 Sadegh Ashkani1,2*
Sadegh Ashkani1,2* Mohd Y. Rafii1*
Mohd Y. Rafii1* Mahmoodreza Shabanimofrad3
Mahmoodreza Shabanimofrad3 Gous Miah1Mahbod Sahebi4Parisa Azizi1Fatah A. Tanweer3,5
Gous Miah1Mahbod Sahebi4Parisa Azizi1Fatah A. Tanweer3,5 Mohd Sayeed Akhtar4,6
Mohd Sayeed Akhtar4,6 Abbas Nasehi7
Abbas Nasehi7- 1Laboratory of Food Crops, Institute of Tropical Agriculture, Universiti Putra Malaysia, Serdang, Malaysia
- 2Department of Agronomy and Plant Breeding, Yadegar –e- Imam Khomeini RAH Shahre-Rey Branch, Islamic Azad University, Tehran, Iran
- 3Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, Serdang, Malaysia
- 4Laboratory of Plantation Crops, Institute of Tropical Agriculture, Universiti Putra Malaysia, Serdang, Malaysia
- 5Department of Plant Breeding and Genetics, Faculty of Crop Production, Sindh Agriculture University Tandojam, Sindh, Pakistan
- 6Department of Botany, Gandhi Faiz-e-Aam College, Shahjahanpur, India
- 7Department of Plant Protection, Faculty of Agriculture, Universiti Putra Malaysia, Serdang, Malaysia
Rice is a staple and most important security food crop consumed by almost half of the world’s population. More rice production is needed due to the rapid population growth in the world. Rice blast caused by the fungus, Magnaporthe oryzae is one of the most destructive diseases of this crop in different part of the world. Breakdown of blast resistance is the major cause of yield instability in several rice growing areas. There is a need to develop strategies providing long-lasting disease resistance against a broad spectrum of pathogens, giving protection for a long time over a broad geographic area, promising for sustainable rice production in the future. So far, molecular breeding approaches involving DNA markers, such as QTL mapping, marker-aided selection, gene pyramiding, allele mining and genetic transformation have been used to develop new resistant rice cultivars. Such techniques now are used as a low-cost, high-throughput alternative to conventional methods allowing rapid introgression of disease resistance genes into susceptible varieties as well as the incorporation of multiple genes into individual lines for more durable blast resistance. The paper briefly reviewed the progress of studies on this aspect to provide the interest information for rice disease resistance breeding. This review includes examples of how advanced molecular method have been used in breeding programs for improving blast resistance. New information and knowledge gained from previous research on the recent strategy and challenges towards improvement of blast disease such as pyramiding disease resistance gene for creating new rice varieties with high resistance against multiple diseases will undoubtedly provide new insights into the rice disease control.
Introduction
Biotic and abiotic stresses cause significant yield losses in food crop production and Improvement in stress tolerance of plant is a major breeding goal. Nowadays, different methods are being used to improve stress tolerance in plants (Figure 1). Diseases are among the most important limiting factors that affect rice production. More than 70 diseases caused by fungi, bacteria, viruses or nematodes have been reported on rice (Zhang et al., 2009). Rice blast (Magnaporthe oryzae) is the most devastating disease of rice because of its wide distribution and its destructiveness under conductive conditions (Skamnioti and Gurr, 2009; Helliwell and Yang, 2013; Helliwell et al., 2013). Among the biotic stresses blast disease is most important. Since there have been many blast disease outbreaks in rice, efforts have been made to develop new cultivars resistant to the blast disease. Earlier studies on the variability of this fungus relied mainly on the phenotypic characters and virulence test using a set of host differentials. These studies were only focused on screening and selection of rice varieties or advanced lines toward selected local blast pathotypes. Most of these phenotypic traits are highly variable as this pathogen is genetically unstable. Such kinds of studies are labor-intensive and time-consuming, require large greenhouse space and often lead to ambiguous results. Furthermore, they are influenced by environmental conditions, inoculation techniques and human errors during scoring (Shivayogi et al., 2002). Now strategic research concentrates on filling the gaps in the existing knowledge of biotic stresses on rice, especially improving molecular genetics of blast disease, with a view to develop an integrated management program for blast resistance. Over the past decades, we have seen the successful use of advanced molecular and genomic tools such as molecular markers, expressed sequence tags (ESTs), microarrays, and genetic transformations to explore the genetic basis of stress tolerance and eventually to develop crop cultivars improved for stress tolerance. The recent developments in DNA marker technology have helped to develop the concepts of QTLs mapping, marker-aided selection (MAS) and genetic transformation to produce plants of superior quality. In addition, molecular markers can be used for assessing genetic diversity, fingerprinting genotypes, separating hybrids from selfed progeny, and other uses. The actual identification of candidate DNA markers linked to resistance genes using fine mapping may well allow rice breeders to efficiently transfer these genes from donor cultivars into new, elite rice cultivars using marker-assisted selection (MAS). There is also a need to connect knowledge about genes and gene function to create new productive varieties that are a necessary element of a sustainable food supply for the future (Ashkani et al., 2015). Here we highlight a set of molecular tools that are currently being used to study the rice blast fungus. The information generated on recent methodology will help breeders to expedite breeding research in rice crops and explore a promising new concept which utilizes such molecular data to breed for durable resistance to rice blast.

FIGURE 1. Method for biotic and abiotic resistance breeding in crops.
Rice And Rice Blast Importance
Rice (Oryza sativa), is the principal food for over half of the population of the world and supplies the main energy resource for almost 50% of the world’s population (Yu et al., 2002). In Asia, where 60% of the earths’ people live 90% of the world’s rice is grown and more than 3 billion Asians obtain 35–75% of their calories from rice and its products (Khush and Jena, 2009). Even though the world’s rice production increased from 257 million tons in 1966 to 600 million tons in 2000, the increase has not kept up with the demand for rice because of the corresponding increase in the human population during this time. It is estimated that rice production must increase by at least 40% in 2030 to meet ever-increasing demands (Khush, 2005). Hence, population increasing at an alarming rate, making the food security the major challenge in future.
Rice serves as an economically important crop and advances in molecular biology have made it a model monocot species among the cereal for genetics studies in breeding programs. Rice in comparison to other grass species has several attributes such as: small genome; extensive genetic resources; genetic transformation potential; synteny with other cereal genomes; comprehensive genetic and physical map of the genome; high density molecular map for gene mapping and map-based gene cloning; complete sequencing of the genome in indica and japonica rice cultivars; development of bacterial artificial chromosome (bac) and yeast artificial chromosome (YAC) libraries and development of the Oryza map alignment project (OMAP); and development of the genetic maps of chloroplast and mitochondrial genomes.
The study of homologies and diversities of markers and genes within and between species, genus or other taxonomic divisions is mentioned to comparative mapping (Paterson et al., 1991). This comparison involves analyzing the conserved area between maps of the order wherein markers occur; the conserved marker order is named ‘synteny.’ Comparative mapping may help in the construction of new linkage maps and the locations predictions of QTLs in varius mapping populations (Young, 1994). Infact, previous linkage maps may show an insight which markers are polymorphic and show an insight of linkage groups and the order of markers in the linkage groups. In the last few years, high-density molecular linkage maps of rice containing approximately 3000 markers have been developed making the marker density in the rice genome, on average, one marker per cM (Causse et al., 1994; Harushima et al., 1998; Lopez-Gerena, 2006).
Rice blast is by far the most important disease that attack rice. The fungus M. oryzae = M. grisae (Cooke) Sacc [anamorph: Pyricularia oryzae], is the causal agent of blast disease (Couch and Kohn, 2002). The fungus colonizes leaves (leaf blast), panicles (panicle blast) and other parts of the rice plants, and causes huge crop loss in rice growing areas. Its most infections occur on the leaves and first symptoms of the disease appear as small brown or grayish dots on the leaves. After 2–3 days the dots develop to almost 1.5 cm long and 0.3–0.5 cm wide diamond-shaped lesions with a gray or white center (Figure 2), causing the infected leaves to die. The yield losses due to blast were reported to be between 30 and 50% in large rice producing areas under favorable environmental conditions (Correa-Victoria José and Zeigler, 1993; Skamnioti and Gurr, 2009). Efforts are underway to develop rice varieties with durable blast resistance. Therefore, continuous studies on blast disease are important in order to overcome this disease problem sustaining rice production in the future.

FIGURE 2. Leaf blast disease symptoms. Lesions are typically spindle-shaped; wide in the center and pointed toward either end. Large lesions usually develop a diamond shape with a grayish center and brown margin.
Management Of Rice Blast And Efficient Ways For Crop Protection
Management of rice blast through the breeding of blast-resistant varieties is the most desirable means of managing blast, especially in developing countries. Rice cultivars with durable blast resistance have been recognized in several production systems. The deployment of rice cultivars with broad-spectrum resistance is practical means of controlling the fungal pathogen (Bonman, 1992; Bonman et al., 1992; Fukuoka et al., 2015). Rice blast control with resistant cultivars is much desired for farmers and consumers, because it can decrease fungicide application, subsequently reducing agrochemical pollution in the rice fields, thus reducing the cost of production. Local and wild varieties are normally used as sources for introgression of a new resistance gene into cultivated rice. Genetic resistance has been, and will continue to be, the major method of disease control of blast. The resistance in newly released rice cultivars to rice blast caused by M. oryzae can be lost due to the high level of instability in the genome of this fungus or due to the frequent breakdown of resistance under field conditions (Bonman, 1992; Bonman et al., 1992; Zeigler et al., 1994). One way to overcome this problem is through pyramiding of multiple R genes, each recognizing a unique set of M. oryzae isolates, into a single cultivar. Molecular markers techniques can be utilize in gene pyramiding in rice breeding programs to produce resistant cultivars and overcome to breakdown of disiease in early stage thereby sustaining rice yields and eventually to map based cloning of the gene. Such techniques as simpler method will save time and minimize costs especially for traits with laborious screening and it is more efficient to use of resources for plant breeding.
Molecular Breeding Research To Improve Disease-Resistant Varieties
New rice varieties that combine higher yield potential with excellent grain quality, resistance to biotic and abiotic stresses, and input use efficiency are desperately needed. Diverse strategies for breeding durable resistance have been offered for rice blast. Some of these strategies, such as pyramiding (Bonman, 1992; Bonman et al., 1992), lineage exclusion (Zeigler et al., 1994), multilines (Abe, 2004) and mixtures (Zhu et al., 2000), are based on the use of complete and specific resistance genes. In general, in current agriculture three major strategies are used to improve disease resistance in crops. The first strategy involves improvement of cultural practices. The second approach comprises the improvement of crops through conventional or molecular marker-assisted breeding of disease resistant cultivars. The third strategy is the direct transformation of resistant genes into elite cultivars (Baulcombe, 2004). The use of molecular techniques for example the agrobacterium-mediated transformation allows the stable transfer of a transgene in a different variety or species, as well as to use a different promoter on a resistance-related gene to alter the intensity of gene expression (Baulcombe, 2004). To increase rice production and resistance, conventional rice breeding carried out during the last 50 years, resulted in the release of modern resistant varieties with high quality and yield. Despite, this method has played an important role in rice cultivar development over the past decades there are drawbacks as well. Conventional breeding progress is slow owing to several obstacles, such as: time-consuming and laborious selection process, difficulties in appropriate genotype selection due to the quantitative nature of most agronomic traits, several generations of crossing, selfing, and testing plants for resistance. In addition, traditional breeding is often negatively affected by linkage drag, which resulted in the transferring of loci conferring potentially undesired agronomic traits due to its close linkage with resistance loci. Recent advances in molecular genetics of rice have provided new tools for breeders to develop the rice varieties of the future which is known as molecular breeding. Only few years ago, the status of rice genetics was considered far behind that of other food crops such as maize, wheat, barley, and tomato. However, the last decade has seen a knowledge explosion in this area and rice is now considered a model plant for such research on cereal crops. Rice has been performed as a successful crop in biotechnology approaches leading to crop improvements. A vast reservoir of germplasm (>200,000 accessions) of both domestic and wild rice is available for genetic and breeding research. With the completion of rice genome sequence, many rice research now focused on functional characterization of rice genes, elucidation of the underlying mechanisms involved in major agronomic traits (e.g., high yield, grain quality, abiotic stress tolerance, and disease resistance), and subsequent translation of genomic knowledge into agricultural productivity via molecular breeding and improved cultural practice (Helliwell and Yang, 2013; Helliwell et al., 2013). Genetic studies of blast resistance in rice were established in Japan as early as 1917 (Ballini et al., 2008). To date, there are various molecular and biotechnological approaches to genetically improve rice crop for effective, durable and/or broad-spectrum resistance to major diseases. Currently, many resistance genes (R-genes) and QTLs in rice for blast have been identified and sequenced (Ashkani et al., 2014). These genes contribute to the understanding of the interaction between the disease and the host for breeding proposes. In addition, a wide variety of genes and mechanisms involved in rice defense response (e.g., pathogenesis-related proteins and other defense genes) have been identified and elucidated. Also, many molecular approaches including use of specialized promoters, modification of target protein structures have been studied and proposed to improve the effectiveness of transgenes (Baulcombe, 2004). During the past two decades, some rice research institute has been involved in the rice mutation breeding program to generate new varieties, in support of the crop breeding program. The main purposes of mutation breeding of rice have been improvement of agronomic traits, inducing resistance against diseases and pests, and enhancing the grain quality and grain taste. In the breeding program for rice in collaboration with IRRI (the Philippines) and JIRCAS (Japan) many modern varieties have also been released for commercial use. Traditional rice varieties have been widely used as genetic resources for biotic and abiotic traits of hybridization program. To complement conventional breeding method, molecular and transgenic method represents an increasingly important approach for genetic improvement of disease resistance and reduction of pesticide usage and various molecular strategies including use of specialized promoters, modification of target protein structures have been studied and proposed to improve the effectiveness of transgenes (Helliwell and Yang, 2013; Helliwell et al., 2013). MAS for quick indirect selection of the target gene by using molecular markers closely linked to a target gene as a molecular tag, quantitative trait locus (QTL) analysis and genetic transformation techniques are the most useful tools for rice molecular breeding especially to improve disease-resistant varieties. These techniques have been used to identify new germplasms and elite rice cultivars.
Blast Disease Improvement Due To Molecular Markers Techniques
Molecular markers have played an increasing role in rice breeding for cultivar improvement, screening, selection and germplasm collections (Wang et al., 2007). The new sequencing tools provide valuable informations for the discovery, validation and assessment of genetic markers in populations (Sahebi et al., 2015). For instance, the analysis of next generation sequencing (NGS) data by means of bioinformatics developments allows discovering new genes and regulatory sequences and their positions, and makes available large collections of molecular markers (Perez-de-Castro et al., 2012). The whole genome sequence data substantially enhanced the efficiency of polymorphic marker development for QTL fine mapping and the identification of possible candidate genes (Wan et al., 2006). These performances can be useful as genetic resources for breeding of rice cultivars. The use of molecular markers for rice has been recently reviewed (Temnykh et al., 2001; Xu, 2002; Semagn et al., 2006; Lang et al., 2008; Kumar et al., 2009; Benali et al., 2011). In the case of rice blast (M. oryzae) a large number of the major genes had been identified and was targeted for mapping investigations using a variety of marker systems and approaches (Ashkani et al., 2014). DNA markers including : simple sequence repeats (SSRs), Single-nucleotide polymorphisms (SNPs) and small insertions/deletions (InDels), amplified fragment length polymorphisms (AFLPs), random amplified polymorphic DNAs (RAPDs), cleaved amplified polymorphic sequences (CAPS), and restriction fragment length polymorphisms (RFLPs) have been identified to be linked with blast resistance genes in rice (Ashkani et al., 2014; Tanweer et al., 2015). In recent year, scientists have used these markers for genetic mapping to identify candidate genes and QTLs in many plant species. Several genes of agronomic importance such as those that confer resistance to blast, bacterial leaf blight, brown planthopper, tungro and grassy stunt virus have been transferred from the wild species into the elite breeding lines of rice, including the quantitative trait loci (QTLs) for biotic and abiotic stress resistance (Amante-Bordeos et al., 1992; Brar and Khush, 1997).
DNA markers techniques provide us and rapid tool to select for the existence of multiple blast resistance genes without the need to test the progeny or inexact phenotypic disease screening (Fjellstrom et al., 2004). Through molecular markers tools many useful markers linked to the race-specific blast resistance genes (Pi-genes), has been identified and screened in segregating populations in rice (Fjellstrom et al., 2004; Sharma et al., 2005; Ashkani et al., 2011). PCR-based markers as example SSRs are precise, reliable and cost effective; this marker has been applied for the selection of plants containing blast resistance genes in rice at an early stage (Hittalmani et al., 2002; Ashkani et al., 2012). Microsatellites are SSR markers, and have been used extensively to identify genes and QTLs associated blast resistance in rice. Microsatellites are abundant in plants (McCouch et al., 2001), causing more polymorphism and better repetition over other marker systems. The genetic map covering all 12 rice chromosomes with at least one microsatellite at the distance of 0.5 cM has been developed by International Rice Microsatellite Initiative (IRMI; McCouch et al., 2002). Currently, breeders are focusing on MAS instead of using conventional breeding. Application of MAS reduces the time for phenotypic selection and saves the costs to select a desired trait (Koide et al., 2009). This method is helpful tool, and more accurate approach in introducing novel cultivars and it also help breeders to expedite breeding research in crops by enabling selection based on the genotype rather than on the phenotype. After the discovery of molecular markers linked with gene of interest, selection of specified traits to develop new cultivar could be made at an early level (Zhu et al., 2012). Pyramiding of linked genes into a single line or cultivar is one of the common applications of MAS.
Marker-assisted backcross breeding (MABC) as another technique recently has been given attention in rice breeding for the introgression of blast resistance genes (one or a few genes) into the susceptible or in an adapted or elite varieties. MABC is the process of using markers to select for target loci, minimize the length of the donor segment containing a target locus and accelerate the recovery of the recurrent parent (RP) genome during backcrossing (Charcosset, 1997; Hospital, 2001; Hasan et al., 2015). The main purpose of MABC is to transfer the desired character/or targeted gene along with recovering the recurrent parent characters/or genes. MABC is now playing an important role for the development of blast-resistant cultivars (Sundaram et al., 2009) and is superior to conventional backcrossing in precision and efficiency and time saving. Molecular markers which are tightly linked with important traits are used in MABC. Therefore, molecular markers are the tools that can be used to detect the presence of desire character in backcrossing and greatly increases the efficiency of selection. The methods and potential application of MAS and MABC for the Improvement of rice have been recently reviewed and described (Collard and Mackill, 2008; Hasan et al., 2015). Recently through application of MABC many blast resistance genes have been successfully introgressed into the genetic background and improved the blast resistance. Some successful examples for application of MAS and MABC in rice breeding programs aimed at improving blast resistance in this species are presented in Table 1.
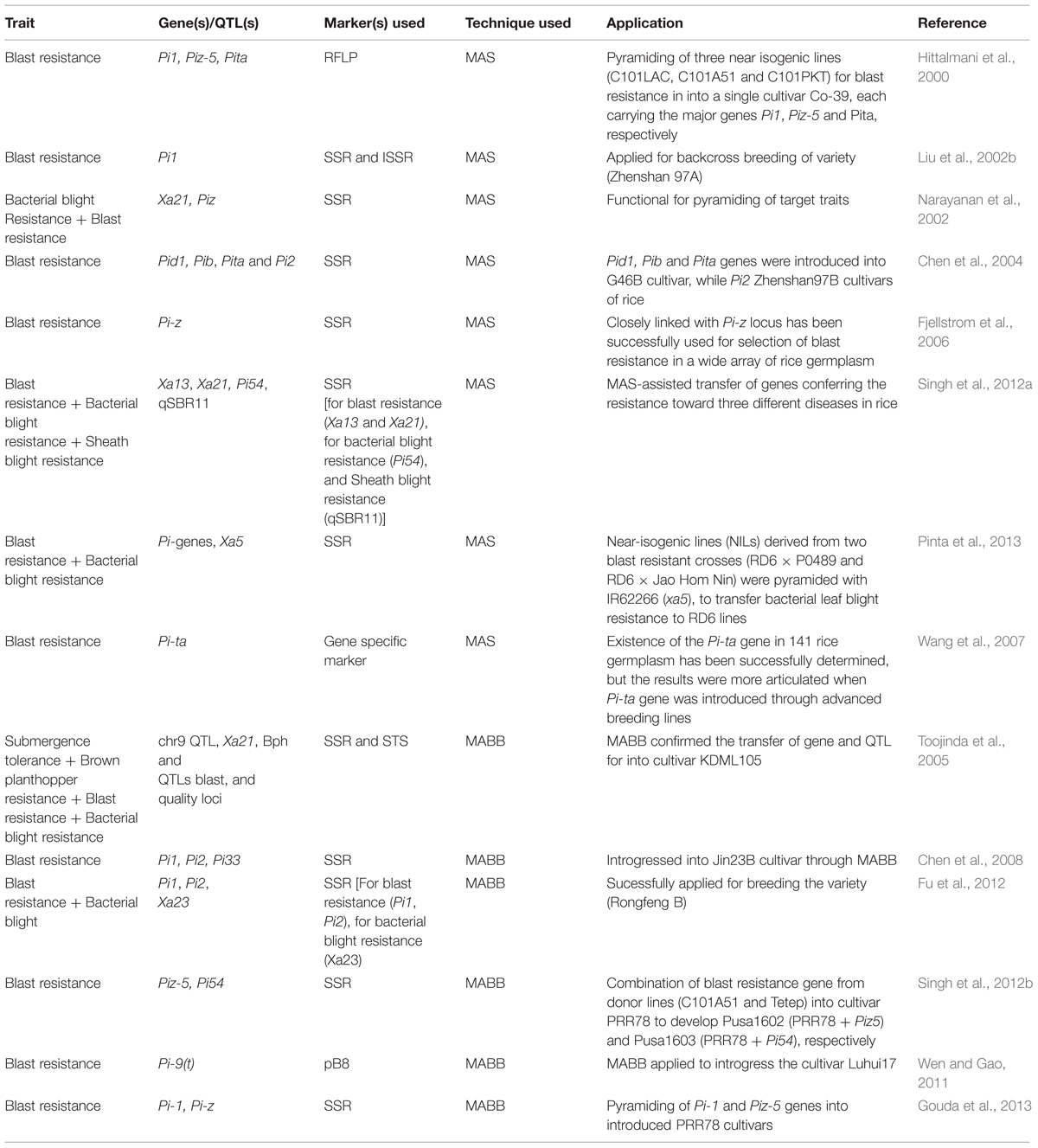
TABLE 1. Examples for application of marker-assisted selection (MAS) and marker-assisted backcrossing (MABB) in rice.
Resistant Genes And Qtls For Blast Disease
Resistance to blast was classified into complete and partial resistance (Wang et al., 1994). Complete resistance is a qualitative character and race specific controlled by a major gene (R genes). Meanwhile, partial resistance is a quantitative character and non-race specific, which is controlled by many genes known as quantitative resistance loci (QRL; Young, 1996). However, if the resistance is highly partial, it can also be controlled by a major gene and is race specific. Qualitative and quantitative blast resistances have been reported in rice germplasm (Ou, 1985). Many qualitative resistance major genes (∼100 genes) for blast resistance have been identified and mapped in the rice genome (Sharma et al., 2012; Ashkani et al., 2014). About 22 R-genes have been successfully cloned and molecularly characterized. In the practice of resistance breeding, using a single R gene which has a broad resistance spectrum is more effective. There have been many reports on introgression of Pi genes related to blast disease into commercial and elite varieties. For example, the Pi-9 gene that exists in the indica rice line 75-1-127 (Liu et al., 2002a), was introgressed from the wild species O. minuta (Amante-Bordeos et al., 1992). The Pi-ta allele was identified in O. rufipogon and O. nivara, or in their hybrids with O. sativa (Jena and Khush, 2000). These R genes function in a gene-for-gene fashion, so the pathogen can adapt by mutating or deleting the corresponding a-virulence gene. Therefore varieties those carrying R genes which confer high levels of resistance typically lose their resistance after a few years (Chen et al., 2003). Quantitative resistance donated by quantitative trait loci (QTL) are long-lasting disease resistance against a wide-range of pathogens, promising for sustainable rice production in the future (Song and Goodman, 2001). QTL mapping is a modern type of study to locate genes controlling a quantitative trait. Since the first publication of a QTL analysis of rice resistant to blast (Wang et al., 1994), several QTLs related to blast resistance have been detected using different type of markers, population and environment and have been published. We have summerized all these events in tabular form (Table 2).
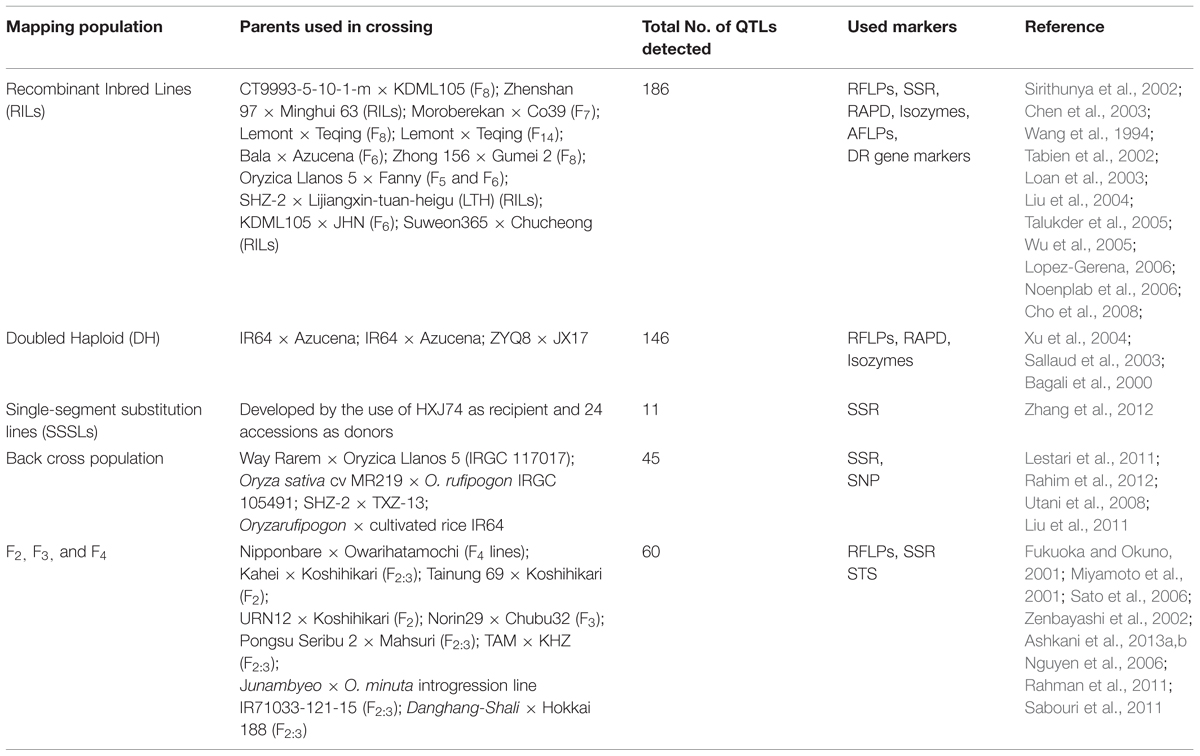
TABLE 2. Quantitative trait loci (QTL), identified for rice blast resistance.
Quantitative trait loci detection approach has been employed to map major or minor genes involved in the resistance (Wang et al., 1994; Fukuoka and Okuno, 2001; Miyamoto et al., 2001; Tabien et al., 2002; Zenbayashi et al., 2002; Chen et al., 2003; Sallaud et al., 2003; Talukder et al., 2004; Wu et al., 2005; Ashkani et al., 2013a,b). Identification of QTLs, associated with blast resistance has been delivered the effective genetics evidences for the molecular marker assisted breeding and cloning of the major genes. In the other word, QTL mapping is useful in identifying multiple loci controlling complete resistance in a highly resistant cultivar as well as in estimating the number, location and effect of genomic region involved in partial blast resistance (Sallaud et al., 2003). Many rice improvement programs now aim to incorporate quantitative or polygenic resistance into rice varieties. Previous studies have verified that genetic linkage maps constructed with various DNA markers are very useful for the analysis and detection of qualitative trait loci (Bao et al., 2000; Price et al., 2000). Molecular linkage maps have helped resolve the effects of minor and major QTLs and estimate the amount phenotypic variation explained at each locus. Molecular linkage maps have led to better understanding of genetic phenomena, such as interloci (epistasis) and intralocus (dominant) interactions (Grandillo and Tanksley, 1996), heterosis (Stuber et al., 1992) and identifying transgressive segregants (Tanksley, 1988, 1993).
Gene Pyramiding For Blast Resistance
Pyramiding is the accumulation of genes into a single line or cultivar. In a gene pyramiding, strategy is to cumulate genes identified in multiple parents into a single genotype (Figure 3). The end product of a gene-pyramiding program is a genotype with all of the target genes. Pyramiding multiple resistance genes provides durable stress resistance expression in crops. Gene Pyramiding technique broadly is used for combining multiple disease or pest resistance genes for specific races of a pathogen or insect to develop durable resistance. It helps in crop improvement program and reduces breeding duration. Different R-genes often confer resistance to different isolates, races or biotypes. Combining their resistance broadens the number of races or isolates that a more than one character in a variety at the same time. Developing elite breeding lines and varieties often requires plant breeders to combine desirable traits from multiple parental lines, particularly in the case of disease resistance. Gene pyramiding can be accelerated by using molecular markers to identify and select plants that contain the desired allele combination in very early stage, resulting in obvious savings of resources including greenhouse or field space, water, and fertilizer. Therefore, marker technology can help existing plant breeding programs and allows researchers to access, transfer and combine genes at a rate and with a precision not previously possible. MAS based gene pyramiding could facilitate in pyramiding of genes effectively into a single genetic background (Joshi and Nayak, 2010). Factors such as the number of genes to be transferred, the distance between the target genes and flanking markers calculated in genetic mapping studies, the number of genotype selected in each breeding generation and the nature of germplasm is critical for successful gene pyramiding program. Gene pyramiding is considered one of the most effective strategies for achieving durable resistance against blast disease in rice (Shinoda et al., 1971; Hittalmani et al., 2000; Koide et al., 2010) and have successfully used for accumulating different blast resistance genes in elite rice cultivars (Table 3).
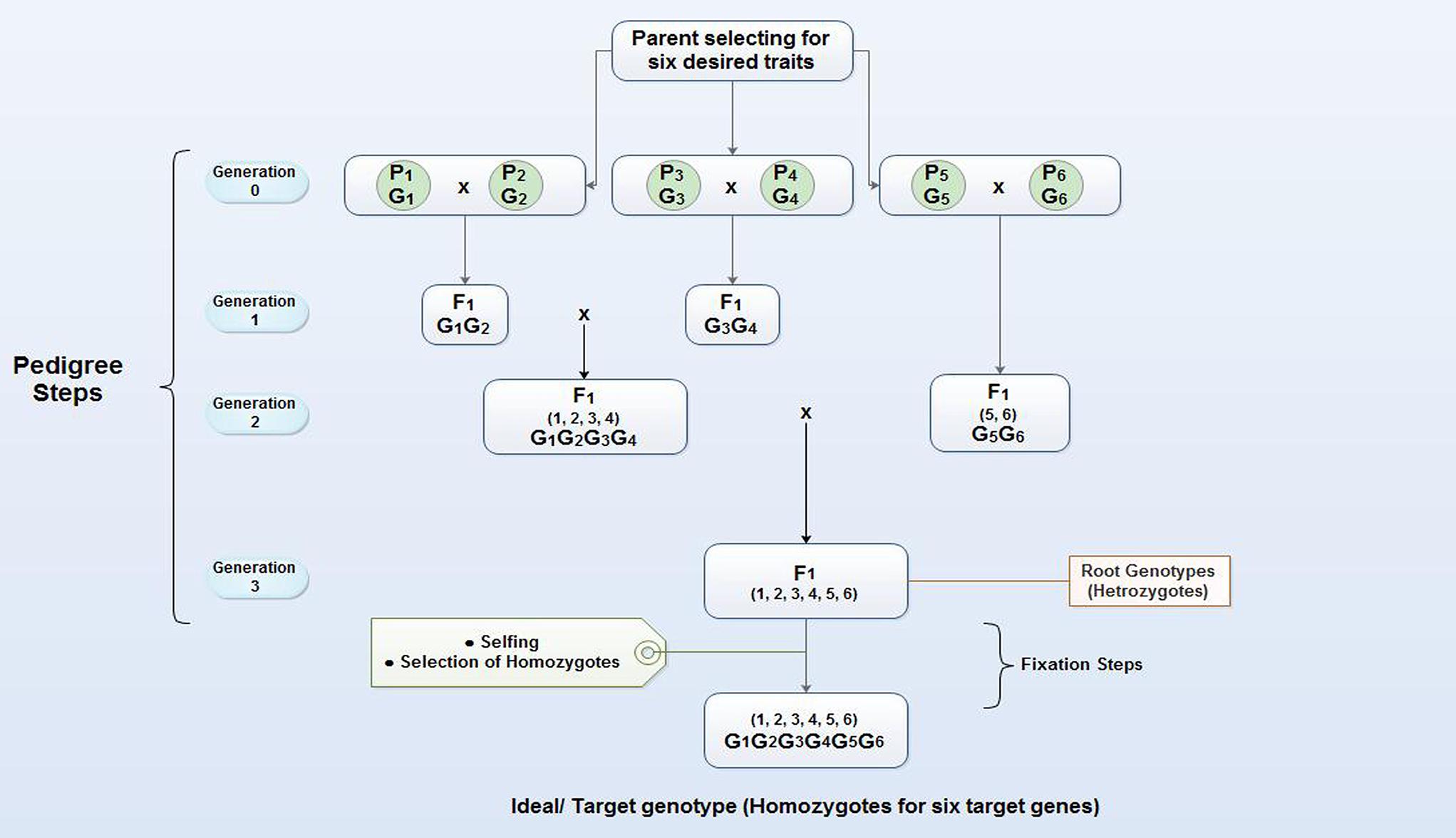
FIGURE 3. Gene pyramiding scheme for cumulating six desired genes (G1–G6) which are present in 6 different parents or lines (P1–P6). The gene pyramiding consists of two steps, pedigree, which aims at cumulating of all target genes in a single genotype (Root genotype) by crossing and selection; the second step is fixation which aims at fixing the target genes into a homozygous state (Ideal/target genotype).
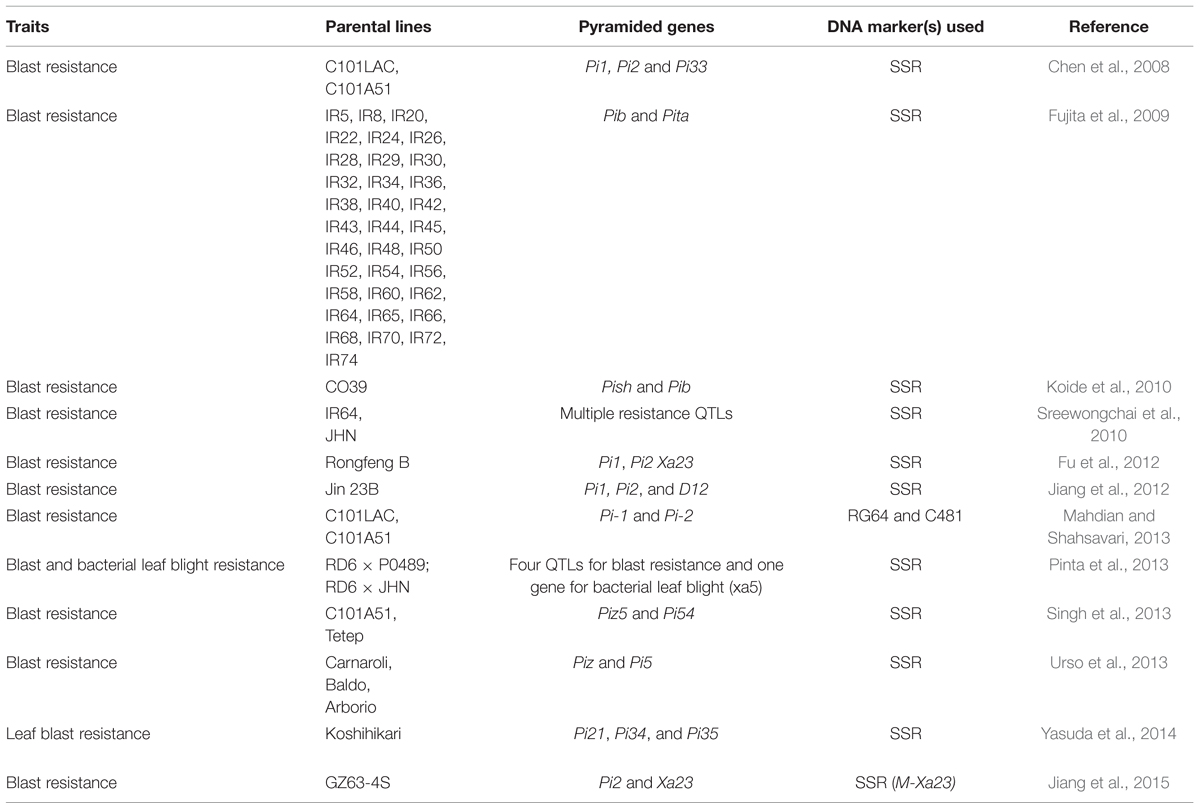
TABLE 3. Example of gene pyramiding for blast resistance trait in rice.
Allele Mining And Blast Resistance Genes
Allele mining is the commonly used approach to identify novel alleles or allelic variants of a gene/or candidate genes of interest, based on the available information about the genes, from a wide range of germplasm. This technique possesses good potential to be used in molecular plant breeding of crop improvement programs. The success of allele mining mainly depends on the type of genetic materials used for screening and should be as diverse as possible and availability of genome and gene sequence information of a particular crop species. For efficient allele mining, wild relatives and local landraces are used because they are reservoirs of useful alleles hidden in their phenotype (Tanksley et al., 1996). The current availability of complete rice genome sequences in addition to several bioinformatic tools have made it possible to mine allelic diversity throughout rice germplasm. EcoTilling and sequence based allele mining are the two widely used approaches in allele mining. Compared to EcoTilling, sequence based allele mining strategy is reported to be simpler and cost effective approach (Ramkumar et al., 2010; Ashkani et al., 2015). Allele mining possesses wide range of applications within crop improvement among them are, allele identification, allelic variation characterization, haplotypes identification, analysis of haplotypes diversity among different haplotypes of the same gene or among the related haplotypes, evolutionary relationship, similarity analysis and development of molecular markers to differentiate a particular allele from other alleles. To date allele mining strategy has been well demonstrated by many researchers. Allele mining of genes from wild and cultivated rice species aims to detect superior alleles for blast resistance (Kumari et al., 2013). So far, mining approaches have been used to identify novel and superior alleles of many major blast resistance genes from different cultivated rice varieties and wild species (Table 4). Through allele mining techniques functional marker to differentiate the resistance and susceptible alleles of Pi54 has been developed (Ramkumar et al., 2011). Costanzo and Jia (2010) analyzed the sequence level similarity for Pikm alleles, derived from 15 different rice cultivars. M. oryza has also been differentiated from M. grisea by using allele mining (Couch and Kohn, 2002).
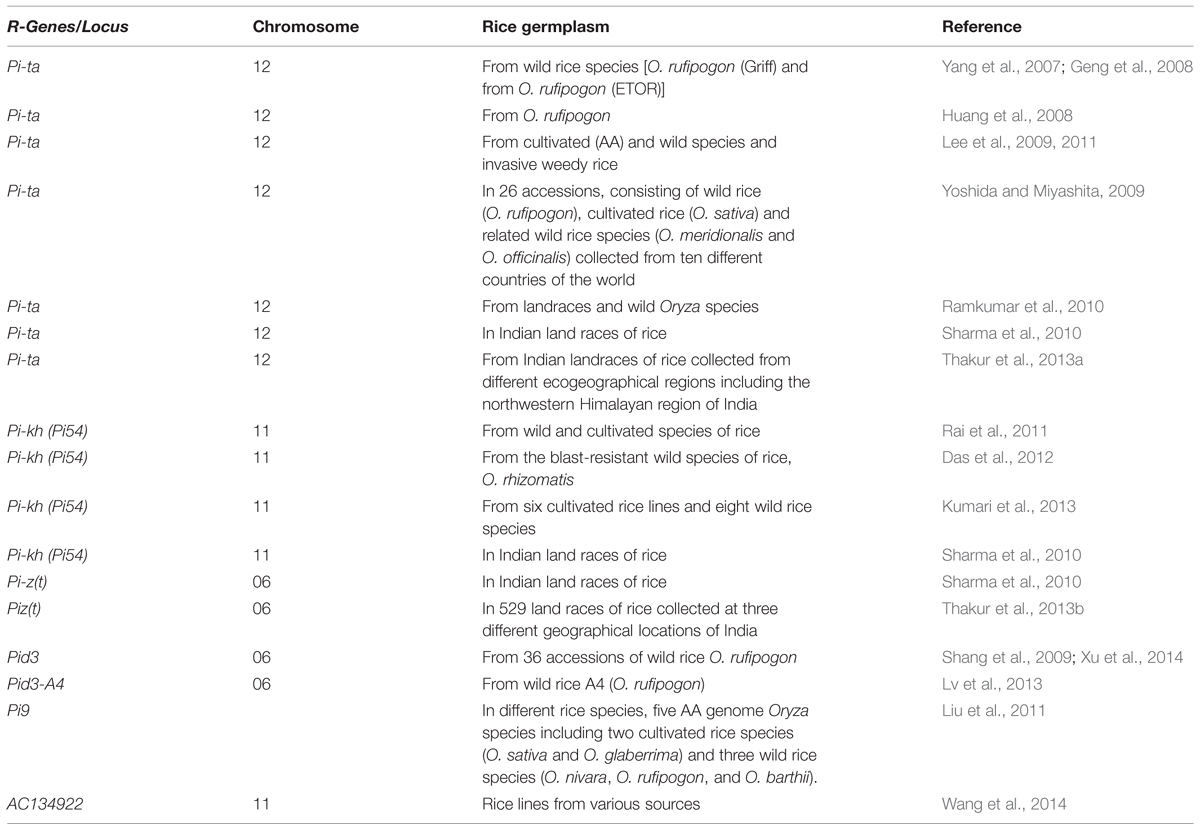
TABLE 4. Summary of allele mining report for blast resistance genes.
Conclusion
Disease management extremely needed to sustain the world for food consumption. Rice blast caused by M. oryzae is the most severe fungal disease, which limits the rice production and causing the yield loss of 157 million tons of rice per annum in the worldwide (Kaundal et al., 2006). Development of resistant varieties with durable resistance by incorporating new genes into the improved germplasm has been proved to be economical, environmentally friendly and effective to control the rice blast disease (Skamnioti and Gurr, 2009). The availability of different molecular tools allows characterization of genes of interest and identification of plants carrying the target genes and might well serves to improve the efficiency of conventional breeding. Due to molecular dissection it is now possible to identify blast resistance genes and QTLs or combined effects of multiple loci with major and minor effects. The marker developed from these genes or QTLs can be used in marker assisted selection for selection of resistance without confounding the effects of environmental factors. DNA markers that co-segregate with the gene are a powerful method for use in crop protection and can be routinely employed in various aspects of plant genome analysis such as genetics and plant breeding. Information provided on genetics of blast resistance of local traditional variety is very useful for rice resistance breeding program in every country. Recent molecular breeding strategy such as gene pyramiding and allele mining holds greater prospects to attain durable resistance against biotic and abiotic stresses in crops. Identification of novel and superior resistance alleles of the blast resistance genes is an important task in the rice breeding program. The novel alleles are very useful in breeding programs and can be utilized gainfully to develop productive and superior plants.
Future Perspectives And Consideration
The major difficulty in controlling rice blast is the durability of genetic resistance. Enhancing the host plant resistance is being considered as the best approach to handle the rice blast disease. Rice cultivars containing only a single R gene to a specific pathogen race often become susceptible over time due to the emergence of new virulent races. Understanding of genetic identity of contemporary M. oryzae is important for accurate deployment of rice cultivars with different R genes. Stacking R genes with overlapped resistance spectra can lead to long lasting resistance. For combinations of different blast resistance genes in host plant in the rice blast breeding programs, superior alleles of the targeted genes should be considered. During the evolution and artificial selection processes, a significant portion of beneficial alleles have been left behind in the landraces and wild species (McCouch et al., 2007), which can be used for the development of better rice varieties. Effective blast management also requires international cooperation. The knowledge gained by collaborative effort ought to lead to more effective methods to reduce crop loss due to blast disease worldwide. Although considerable progress has been made toward understanding the nature of disease resistance genes, defense responses, and the signal transduction leading to activation of defense responses in rice, the whole story is still far from clear. Studies of the molecular biology of disease resistance will be helpful in improving rice varieties for high production for increasing population. The completion of the rice genome project and availability of structural genomic data for the public can undoubtedly accelerate research on the molecular biology of rice disease resistance. Development of new molecular techniques, methodologies such as functional geonomics and DNA microarrays for global analysis of gene expression should urge rice breeders to integrate these techniques to conventional breeding. Rice research should more focus on identifying more durably resistant genes, tagging of these genes with molecular markers and pyramiding these genes or QTLs through molecular MAS. Monogenic resistance to blast is less stable but varieties with pyramided monogenes or QTLs are durably resistant. Molecular breeding strategy can help in the introduction of durably blast-resistant rice cultivars thereby sustaining rice yields. Candidate gene identification through rice functional genomics has great potential for developing more durably resistant varieties.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank our university colleagues for their helpful suggestions, insightful comments and discussions on this valuable review. This work was supported by the Long-term Research Grants Scheme (LRGS), Rice Food Security Project from the Ministry of Education, Malaysia.
References
Abe, S. (2004). Breeding of a blast resistant multiline variety of rice. Sasanishiki BL. Japan Agric. Res. Q. 38, 149–154. doi: 10.6090/jarq.38.149
Amante-Bordeos, A., Sitch, L., Nelson, R., Dalmacio, R., Oliva, N., Aswidinnoor, H., et al. (1992). Transfer of bacterial blight and blast resistance from the tetraploid wild rice Oryza minuta to cultivated rice. Oryza sativa. Theor. Appl. Genet. 84, 345–354. doi: 10.1007/BF00229493
Ashkani, S., Rafii, M. Y., Rahim, H. A., and Latif, M. A. (2013a). Genetic dissection of rice blast resistance by QTL mapping approach using an F3 population. Mol. Biol. Rep. 40, 2503–2515. doi: 10.1007/s11033-012-2331-3
Ashkani, S., Rafii, M. Y., Rahim, H. A., and Latif, M. A. (2013b). Mapping of the quantitative trait locus (QTL) conferring partial resistance to rice leaf blast disease. Biotechnol. Lett. 35, 799–810. doi: 10.1007/s10529-012-1130-1
Ashkani, S., Rafii, M. Y., Rusli, I., Sariah, M., Abdullah, S. N. A., Rahim, H. A., et al. (2012). SSRs for marker-assisted selection for blast resistance in rice (Oryza sativa L.). Plant Mol. Biol. Rep. 30, 79–86. doi: 10.1007/s11105-011-0315-4
Ashkani, S., Rafii, M. Y., Sariah, M., Siti, N. A. A., Rusli, I., Rahim, H. A., et al. (2011). Analysis of simple sequence repeat markers linked with blast disease resistance genes in a segregating population of rice (Oryza sativa). Genet. Mol. Res. 10, 1345–1355. doi: 10.4238/vol10-3gmr1331
Ashkani, S., Rafii, M. Y., Shabanimofrad, M., Ghasemzadeh, A., Ravanfar, S., and Latif, M. A. (2014). Molecular progress on the mapping and cloning of functional genes for blast disease in rice (Oryza sativa L.): current status and future considerations. Crit. Rev. Biotechnol. 14, 1–15.
Ashkani, S., Rafii, M. Y., Shabanimofrad, M., Azadi, A., Ghasemzadeh, A., Azizi, P., et al. (2015). Allele mining strategies: principles and utilisation for blast resistance genes in rice (Oryza sativa L.). Curr. Issues Mol. Biol. 17, 57–74.
Bagali, P. G., Hittalmani, S., Shashidhar, S. Y., and Shashidhar, H. (2000). “Identification of DNA markers linked to partial resistance for blast disease in rice across four locations,” in Advances in Rice Blast Research, eds D. Tharreau, M. H. Lebrun, N. J. Talbot, and J. L. Notteghem (Berlin: Springer), 34–42.
Ballini, E., Morel, J.-B., Droc, G., Price, A., Courtois, B., Notteghem, J.-L., et al. (2008). A genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol. Plant Microbe Inter. 21, 859–868. doi: 10.1094/MPMI-21-7-0859
Bao, J., Zheng, X., Xia, Y., He, P., Shu, Q., Lu, X., et al. (2000). QTL mapping for the paste viscosity characteristics in rice (Oryza sativa L.). Theor. Appl. Genet. 100, 280–284. doi: 10.1139/G10-070
Benali, S., Mohamed, B., Eddine, H. J., and Neema, C. (2011). Advances of molecular markers application in plant pathology research. Eur. J. Sci. Res. 50, 110–123.
Bonman, J. (1992). Durable resistance to rice blast disease-environmental influences. Euphytica 63, 115–123. doi: 10.1007/BF00023917
Bonman, J., Khush, G., and Nelson, R. (1992). Breeding rice for resistance to pests. Annu. Rev. Phytopathol. 30, 507–528. doi: 10.1146/annurev.py.30.090192.002451
Brar, D., and Khush, G. (1997). Alien introgression in rice. Plant Mol. Biol. 35, 35–47. doi: 10.1023/A:1005825519998
Causse, M. A., Fulton, T. M., Cho, Y. G., Ahn, S. N., Chunwongse, J., Wu, K., et al. (1994). Saturated molecular map of the rice genome based on an interspecific backcross population. Genetics 138, 1251–1274.
Charcosset, A. (1997). Marker-assisted introgression of quantitative trait loci. Genetics 147, 1469–1485.
Chen, H., Chen, Z., Ni, S., Zuo, S., Pan, X., and Zhu, X. (2008). Pyramiding three genes with resistance to blast by marker assisted selection to improve rice blast resistance of Jin23B. Chinese J. Rice Sci. 22, 23–27.
Chen, H., Wang, S., Xing, Y., Xu, C., Hayes, P. M., and Zhang, Q. (2003). Comparative analyses of genomic locations and race specificities of loci for quantitative resistance to Pyricularia grisea in rice and barley. Proc. Natl. Acad. Sci. U.S.A 100, 2544–2549. doi: 10.1073/pnas.0437898100
Chen, X. W., Li, S. G., Ma, Y. Q., Li, H. Y., Zhou, K. D., and Zhu, L. H. (2004). Marker-assisted selection and pyramiding for three blast resistance genes, Pi-d (t) 1, Pi-b, Pi-ta2, in rice. Chinese J. Biotechnol. 20, 708–714.
Cho, Y.-C., Kwon, S.-W., Suh, J.-P., Kim, J.-J., Lee, J.-H., Roh, J.-H., et al. (2008). QTLs identification and confirmation of field resistance to leaf blast in temperate japonica rice (Oryza sativa L.). J. Crop Sci. Biotechnol. 11, 269–276.
Collard, B. C., and Mackill, D. J. (2008). Marker-assisted selection: an approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. B 363, 557–572. doi: 10.1098/rstb.2007.2170
Correa-Victoria José, F., and Zeigler, R. S. (1993). Pathogenic variability in Pyricularia grisea at a rice blast hot spot breeding site in eastern Colombia. Plant Dis. 77, 1029–1035. doi: 10.1094/PD-77-1029
Costanzo, S., and Jia, Y. (2010). Sequence variation at the rice blast resistance gene Pi-km locus: implications for the development of allele specific markers. Plant Sci. 178, 523–530. doi: 10.1016/j.plantsci.2010.02.014
Couch, B. C., and Kohn, L. M. (2002). A multilocus gene genealogy concordant with host preference indicates segregation of a new species, Magnaporthe oryzae, from M. grisea. Mycologia 94, 683–693. doi: 10.2307/3761719
Das, A., Soubam, D., Singh, P., Thakur, S., Singh, N., and Sharma, T. (2012). A novel blast resistance gene, Pi54rh cloned from wild species of rice, Oryza rhizomatis confers broad spectrum resistance to Magnaporthe oryzae. Funct. Integr. Genom. 12, 215–228. doi: 10.1007/s10142-012-0284-1
Fjellstrom, R., Conaway-Bormans, C. A., Mcclung, A. M., Marchetti, M. A., Shank, A. R., and Park, W. D. (2004). Development of DNA markers suitable for marker assisted selection of three genes conferring Resistance to multiple pathotypes. Crop Sci. 44, 1790–1798. doi: 10.2135/cropsci2004.1790
Fjellstrom, R., Mcclung, A. M., and Shank, A. R. (2006). SSR markers closely linked to the Pi-z locus are useful for selection of blast resistance in a broad array of rice germplasm. Mol. Breed. 17, 149–157. doi: 10.1007/s11032-005-4735-4
Fu, C., Wu, T., Liu, W., Wang, F., Li, J., Zhu, X., et al. (2012). Genetic improvement of resistance to blast and bacterial blight of the elite maintainer line Rongfeng B in hybrid rice (Oryza sativa L.) by using marker-assisted selection. Afr. J. Biotechnol. 11, 13104–13114. doi: 10.5897/AJB12.1465
Fujita, D., Ebron, L. A., Kobayashi, N., and Fukuta, Y. (2009). “DNA marker analysis of blast resistance genes Pib and Pita in IRRI-bred rice varieties comparision with gene estimation by a differential system,” in Advances in Genetics, Genomics and Control of Rice Blast Disease, eds G. L. Wang and B. Valent (Berlin: Springer), 315–324.
Fukuoka, S., and Okuno, K. (2001). QTL analysis and mapping of pi21, a recessive gene for field resistance to rice blast in Japanese upland rice. Theor. Appl. Genet. 103, 185–190. doi: 10.1007/s001220100611
Fukuoka, S., Saka, N., Mizukami, Y., Koga, H., Yamanouchi, U., Yoshioka, Y., et al. (2015). Gene pyramiding enhances durable blast disease resistance in rice. Sci. Rep. 5:7773. doi: 10.1038/srep07773
Geng, X. S., Yang, M. Z., Huang, X. Q., Cheng, Z. Q., Fu, J., Sun, T., et al. (2008). Cloning and analyzing of rice blast resistance gene Pi-ta+ allele from Jinghong erect type of common wild rice (Oryza rufipogon Griff) in Yunnan. Yi Chuan 30, 109–114. doi: 10.3724/SP.J.1005.2008.00109
Gouda, P. K., Saikumar, S., Varma, C. M., Nagesh, K., Thippeswamy, S., Shenoy, V., et al. (2013). Marker-assisted breeding of Pi-1 and Piz-5 genes imparting resistance to rice blast in PRR78, restorer line of Pusa RH-10 Basmati rice hybrid. Plant Breed. 132, 61–69. doi: 10.1111/pbr.12017
Grandillo, S., and Tanksley, S. (1996). QTL analysis of horticultural traits differentiating the cultivated tomato from the closely related species Lycopersicon pimpinellifolium. Theor. Appl. Genet. 92, 935–951. doi: 10.1007/BF00224033
Harushima, Y., Yano, M., Shomura, A., Sato, M., Shimano, T., Kuboki, Y., et al. (1998). A high-density rice genetic linkage map with 2275 markers using a single F2 population. Genetics 148, 479–494.
Hasan, M. M., Rafii, M. Y., Ismail, M. R., Mahmood, M., Rahim, H. A., Alam, M. A., et al. (2015). Marker-assisted backcrossing: a useful method for rice improvement. Biotechnol. Biotechnol. Equip. 29, 237–254. doi: 10.1080/13102818.2014.995920
Helliwell, E. E., Wang, Q., and Yang, Y. (2013). Transgenic rice with inducible ethylene production exhibits broad-spectrum disease resistance to the fungal pathogens Magnaporthe oryzae and Rhizoctonia solani. Plant Biotechnol. J. 11, 33–42. doi: 10.1111/pbi.12004
Helliwell, E. E., and Yang, Y. (2013). Molecular strategies to improve rice disease resistance. Methods Mol. Biol. 956, 285–309. doi: 10.1007/978-1-62703-194-3_21
Hittalmani, S., Parco, A., Mew, T., Zeigler, R., and Huang, N. (2000). Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor. Appl. Genet. 100, 1121–1128. doi: 10.1007/s001220051395
Hittalmani, S., Shashidhar, H., Bagali, P. G., Huang, N., Sidhu, J., Singh, V., et al. (2002). Molecular mapping of quantitative trait loci for plant growth, yield and yield related traits across three diverse locations in a doubled haploid rice population. Euphytica 125, 207–214. doi: 10.1023/A:1015890125247
Hospital, F. (2001). Size of donor chromosome segments around introgressed loci and reduction of linkage drag in marker-assisted backcross programs. Genetics 158, 1363.
Huang, C.-L., Hwang, S.-Y., Chiang, Y.-C., and Lin, T.-P. (2008). Molecular evolution of the Pi-ta gene resistant to rice blast in wild rice (Oryza rufipogon). Genetics 179, 1527–1538. doi: 10.1534/genetics.108.089805
Jena, K. K., and Khush, G. S. (2000). “Exploitation of species in rice improvement-opportunities, achievements and future challenges,” in Rice Breeding and Genetic Research Priorities and Challenges, ed. J. S. Nanda (Enfield: Science Publication), 269–284.
Jiang, H., Feng, Y., Bao, L., Li, X., Gao, G., Zhang, Q., et al. (2012). Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding. Mol. Breed. 30, 1679–1688. doi: 10.1007/s11032-012-9751-6
Jiang, J., Yang, D., Ali, J., and Mou, T. (2015). Molecular marker-assisted pyramiding of broad-spectrum disease resistance genes, Pi2 and Xa23, into GZ63-4S, an elite thermo-sensitive genic male-sterile line in rice. Mol. Breed. 35, 1–12. doi: 10.1007/s11032-015-0282-9
Joshi, R. K., and Nayak, S. (2010). Gene pyramiding-A broad spectrum technique for developing durable stress resistance in crops. Biotechnol. Mol. Biol. Rev. 5, 51–60.
Kaundal, R., Kapoor, A. S., and Raghava, G. P. (2006). Machine learning techniques in disease forecasting: a case study on rice blast prediction. BMC Bioinformatics 7:485. doi: 10.1186/1471-2105-7-485
Khush, G. S. (2005). What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 59, 1–6. doi: 10.1007/s11103-005-2159-5
Khush, G. S., and Jena, K. K. (2009). “Current status and future prospects for research on blast resistance in rice (Oryza sativa L.),” in Advances in Genetics, Genomics and Control of Rice Blast Disease, eds G. L. Wang and B. Valent (Berlin: Springer), 1–10.
Koide, Y., Kawasaki, A., Telebancoâaaa-Yanoria, M., Hairmansis, A., Nguyet, N., Bigirimana, J., et al. (2010). Development of pyramided lines with two resistance genes, Pish and Pib, for blast disease (Magnaporthe oryzae B. Couch) in rice (Oryza sativa L.). Plant Breed. 129, 670–675. doi: 10.1111/j.1439-0523.2010.01781.x
Koide, Y., Kobayashi, N., Xu, D., and Fukuta, Y. (2009). Resistance genes and selection DNA markers for blast disease in rice (Oryza sativa L.). Japan Agric. Res. Q. 43, 255–280. doi: 10.1016/j.crvi.2014.11.003
Kumar, P., Gupta, V., Misra, A., Modi, D., and Pandey, B. (2009). Potential of molecular markers in plant biotechnology. Plant Omics J. 2, 141–162.
Kumari, A., Das, A., Devanna, B., Thakur, S., Singh, P., Singh, N., et al. (2013). Mining of rice blast resistance gene Pi54 shows effect of single nucleotide polymorphisms on phenotypic expression of the alleles. Eur. J. Plant Pathol. 137, 55–65. doi: 10.1007/s10658-013-0216-5
Lang, N., Buu, B. C., and Ismail, A. (2008). Molecular mapping and marker-assisted selection for major-gene traits in rice (Oryza sativa L.). Omonrice 16, 50–56.
Lee, S., Costanzo, S., Jia, Y., Olsen, K. M., and Caicedo, A. L. (2009). Evolutionary dynamics of the genomic region around the blast resistance gene Pi-ta in AA genome Oryza species. Genetics 183, 1315–1325. doi: 10.1534/genetics.109.108266
Lee, S., Jia, Y., Jia, M., Gealy, D. R., Olsen, K. M., and Caicedo, A. L. (2011). Molecular evolution of the rice blast resistance gene Pi-ta in invasive weedy rice in the USA. PLoS ONE 6:e26260. doi: 10.1371/journal.pone.0026260
Lestari, P., Trijatmiko, K. R., Warsun, A., Ona, I., Cruz, C. V., and Bustamam, M. (2011). Mapping quantitative trait loci conferring blast resistance in upland indica rice (Oryza sativa L.). J. Crop Sci. Biotechnol. 14, 57–63. doi: 10.1007/s12892-010-0030-y
Liu, B., Zhang, S., Zhu, X., Yang, Q., Wu, S., Mei, M., et al. (2004). Candidate defense genes as predictors of quantitative blast resistance in rice. Mol. Plant Microbe Inter. 17, 1146–1152. doi: 10.1094/MPMI.2004.17.10.1146
Liu, G., Lu, G., Zeng, L., and Wang, G.-L. (2002a). Two broad-spectrum blast resistance genes, Pi9 (t) and Pi2 (t), are physically linked on rice chromosome 6. Mol. Genet. Genomics 267, 472–480. doi: 10.1007/s00438-002-0677-2
Liu, S., Li, X., Wang, C., Li, X., and He, Y. (2002b). Improvement of resistance to rice blast in Zhenshan 97 by molecular marker-aided selection. Acta Bot. Sinica 45, 1346–1350.
Liu, Y., Zhu, X. Y., Zhang, S., Bernardo, M., Edwards, J., Galbraith, D. W., et al. (2011). Dissecting quantitative resistance against blast disease using heterogeneous inbred family lines in rice. Theor. Appl. Genet. 122, 341–353. doi: 10.1007/s00122-010-1450-2
Loan, L., Du, P., and Li, Z. (2003). Identification of genes conferring resistance to some Philippine and Vietnamese races of blast. Omonrice 11, 49–62.
Lopez-Gerena, J. (2006). Mapping QTL Controlling Durable Resistance to Rice Blast in the Cultivar Oryzica llanos 5, Ph.D. thesis, Kansas State University, Manhattan, KS.
Lv, Q., Xu, X., Shang, J., Jiang, G., Pang, Z., Zhou, Z., et al. (2013). Functional analysis of Pid3-A4, an ortholog of rice blast resistance gene Pid3 revealed by allele mining in common wild rice. Phytopathology 103, 594–599. doi: 10.1094/PHYTO-10-12-0260-R
Mahdian, S., and Shahsavari, A. (2013). Pyramiding of blast resistance genes Pi-1 and Pi-2 in tarom mahalli rice cultivar. Seed Plant Improv. J. 29, 391–395.
McCouch, S., Temnykh, S., Lukashova, A., Coburn, J., Declerck, G., Cartinhour, S., et al. (2001). “Microsatellite markers in rice: abundance, diversity, and applications,” in Rice genetics IV, eds G. S. Khush, D. S. Brar, and B. Hardy (Willingford: CAB International), 117–135.
McCouch, S. R., Teytelman, L., Xu, Y., Lobos, K. B., Clare, K., Walton, M., et al. (2002). Development and mapping of 2240 new SSR markers for rice (Oryza sativa L.). DNA Res. 9, 199–207. doi: 10.1093/dnares/9.6.199
McCouch, S. R., Sweeney, M., Li, J., Jiang, H., Thomson, M., Septiningsih, E., et al. (2007). Through the genetic bottleneck: O. rufipogon as a source of trait-enhancing alleles for O. sativa. Euphytica 154, 317–339. doi: 10.1007/s10681-006-9210-8
Miyamoto, M., Yano, M., and Hirasawa, H. (2001). Mapping of quantitative trait loci conferring blast field resistance in the japanese upland rice variety kahei. Breed. Sci. 51, 257–261. doi: 10.1270/jsbbs.51.257
Narayanan, N., Baisakh, N., Cruz, V., Gnanamanickam, S., Datta, K., and Datta, S. (2002). Molecular breeding for the development of blast and bacterial blight resistance in rice cv. IR50. Crop Sci. 42, 2072–2079. doi: 10.2135/cropsci2002.2072
Nguyen, T., Koizumi, S., La, T., Zenbayashi, K., Ashizawa, T., Yasuda, N., et al. (2006). Pi35 (t), a new gene conferring partial resistance to leaf blast in the rice cultivar Hokkai 188. Theor. Appl. Genet. 113, 697–704. doi: 10.1007/s00122-006-0337-8
Noenplab, A., Vanavichit, A., Toojinda, T., Sirithunya, P., Tragoonrung, S., Sriprakhon, S., et al. (2006). QTL Mapping for leaf and neck blast resistance in Khao Dawk Mali105 and Jao Hom Nin recombinant inbred lines. Sci. Asia 32, 133–142. doi: 10.2306/scienceasia1513-1874.2006.32.133
Paterson, A. H., Tanksley, S. D., and Sorrells, M. E. (1991). DNA markers in plant improvement. Adv. Agron. 44, 39–90. doi: 10.1016/S0065-2113(08)60578-7
Perez-de-Castro, A. M., Vilanova, S., Canizares, J., Pascual, L., Blanca, J. M., Diez, M. J., et al. (2012). Application of genomic tools in plant breeding. Curr. Genomics 13, 179. doi: 10.2174/138920212800543084
Pinta, W., Toojinda, T., Thummabenjapone, P., and Sanitchon, J. (2013). Pyramiding of blast and bacterial leaf blight resistance genes into rice cultivar RD6 using marker assisted selection. Afr. J. Biotechnol. 12, 4432–4438. doi: 10.5897/AJB12.2028
Price, A., Steele, K., Moore, B., Barraclough, P., and Clark, L. (2000). A combined RFLP and AFLP linkage map of upland rice (Oryza sativa L.) used to identify QTLs for root-penetration ability. Theor. Appl. Genet. 100, 49–56. doi: 10.1007/s001220050007
Rahim, H., Bhuiyan, M., Lim, L., Sabu, K., Saad, A., Azhar, M., et al. (2012). Identification of quantitative trait loci for blast resistance in BC. Genet. Mol. Res. 11, 3277–3289. doi: 10.4238/2012.September.12.11
Rahman, L., Khanam, S., and Jaehwan, R. (2011). Mapping of QTLs involved in resistence to rice blast (Magnaporthe grisea) using Oryza minuta introgression lines. Czech. J. Genet. Plant Breed. 47, 85–94.
Rai, A. K., Kumar, S. P., Gupta, S. K., Gautam, N., Singh, N. K., and Sharma, T. R. (2011). Functional complementation of rice blast resistance gene Pi-kh (Pi54) conferring resistance to diverse strains of Magnaporthe oryzae. J. Plant Biochem. Biotechnol. 20, 55–65. doi: 10.1007/s13562-010-0026-1
Ramkumar, G., Biswal, A. K., Mohan, K. M., Sakthivel, K., Sivaranjani, A. K. P., Neeraja, C. M., et al. (2010). Rani NS, Viraktamath, B.C., and, Madhav, M.S. (2010b). Identifying novel alleles of rice blast resistance genes Pikh and Pita through allele mining. Int. Rice Res. Notes 35, 1–6.
Ramkumar, G., Srinivasarao, K., Mohan, K. M., Sudarshan, I., Sivaranjani, A., Gopalakrishna, K., et al. (2011). Development and validation of functional marker targeting an InDel in the major rice blast disease resistance gene Pi54 (Pikh). Mol. Breed. 27, 129–135. doi: 10.1007/s11032-010-9538-6
Sabouri, H., Sabouri, A., Jafarzadeh, M. R., and Mollashahi, M. (2011). Detection of QTLs controlling field blast resistance in rice (Oryza sative L.). Plant Omics J. 4, 1–5.
Sahebi, M., Hanafi, M. M., Azizi, P., Hakim, A., Ashkani, S., and Abiri, R. (2015). Suppression subtractive hybridization versus next-generation sequencing in plant genetic engineering: challenges and perspectives. Mol. Biotechnol. 57, 880–903. doi: 10.1007/s12033-015-9884-z
Sallaud, C., Lorieux, M., Roumen, E., Tharreau, D., Berruyer, R., Svestasrani, P., et al. (2003). Identification of five new blast resistance genes in the highly blast-resistant rice variety IR64 using a QTL mapping strategy. Theor. Appl. Genet. 106, 794–803.
Sato, H., Takeuchi, Y., Hirabayashi, H., Nemoto, H., Hirayama, M., Kato, H., et al. (2006). Mapping QTLs for field resistance to rice blast in the Japanese upland rice variety Norin 12. Breed. Sci. 56, 415–418. doi: 10.1270/jsbbs.56.415
Semagn, K., Bjørnstad,Å., and Ndjiondjop, M. (2006). An overview of molecular marker methods for plants. Afr. J. Biotechnol. 5, 2540–2568.
Shang, J., Tao, Y., Chen, X., Zou, Y., Lei, C., Wang, J., et al. (2009). Identification of a new rice blast resistance gene, Pid3, by genomewide comparison of paired nucleotide-binding site-leucine-rich repeat genes and their pseudogene alleles between the two sequenced rice genomes. Genetics 182, 1303–1311. doi: 10.1534/genetics.109.102871
Sharma, T. R., Madhav, M. S., Singh, B. K., Shanker, P., Jana, T., Dalal, V., et al. (2005). High-resolution mapping, cloning and molecular characterization of the Pi-k h gene of rice, which confers resistance to Magnaporthe grisea. Mol. Genet. Genom. 274, 569–578. doi: 10.1007/s00438-005-0035-2
Sharma, T., Rai, A., Gupta, S., Vijayan, J., Devanna, B., and Ray, S. (2012). Rice blast management through host-plant resistance: retrospect and prospects. Agric. Res. 1, 37–52. doi: 10.1007/s40003-011-0003-5
Sharma, T. R., Rai, A. K., Gupta, S. K., and Singh, N. K. (2010). Broad-spectrum blast resistance gene Pi-kh cloned from rice line Tetep designated Pi54. J. Plant Biochem. Biotechnol. 191, 87–89. doi: 10.1007/BF03323441
Shinoda, H., Toriyama, K., Yunoki, T., Ezuka, A., and Sakurai, Y. (1971). Studies on the varietal resistance of rice to blast. 6. Linkage relationship of blast resistance genes. Bull. Chugoku Agric. Exp. Stn. Set. A 20, 1–25.
Shivayogi, S., Vaishali, M., Shashidhar, H., and Kumar, K. G. (2002). Genetic analysis of rice blast fungus of southern Karnataka using DNA markers and reaction of popular rice genotypes. Curr. Sci. 82, 732–735.
Singh, A., Singh, V. K., Singh, S., Pandian, R., Ellur, R. K., Singh, D., et al. (2012a). Molecular breeding for the development of multiple disease resistance in Basmati rice. AoB Plants 2012:pls029. doi: 10.1093/aobpla/pls029
Singh, V. K., Singh, A., Singh, S., Ellur, R. K., Choudhary, V., Sarkel, S., et al. (2012b). Incorporation of blast resistance into “PRR78”, an elite Basmati rice restorer line, through marker assisted backcross breeding. Field Crops Res. 128, 8–16. doi: 10.1016/j.fcr.2011.12.003
Singh, V. K., Singh, A., Singh, S., Ellur, R. K., Singh, D., Gopala Krishnan, S., et al. (2013). Marker-assisted simultaneous but stepwise backcross breeding for pyramiding blast resistance genes Piz5 and Pi54 into an elite Basmati rice restorer line ‘PRR78’. Plant Breed. 132, 486–495.
Sirithunya, P., Tragoonrung, S., Vanavichit, A., Pa-in, N., Vongsaprom, C., and Toojinda, T. (2002). Quantitative trait loci associated with leaf and neck blast resistance in recombinant inbred line population of rice (Oryza sativa). DNA Res. 9, 79–88. doi: 10.1093/dnares/9.3.79
Skamnioti, P., and Gurr, S. J. (2009). Against the grain: safeguarding rice from rice blast disease. Trends Biotechnol. 27, 141–150. doi: 10.1016/j.tibtech.2008.12.002
Song, F., and Goodman, R. M. (2001). Molecular biology of disease resistance in rice. Physiol. Mol. Plant Pathol. 59, 1–11. doi: 10.1006/pmpp.2001.0353
Sreewongchai, T., Toojinda, T., Thanintorn, N., Kosawang, C., Vanavichit, A., Tharreau, D., et al. (2010). Development of elite indica rice lines with wide spectrum of resistance to Thai blast isolates by pyramiding multiple resistance QTLs. Plant Breed. 129, 176–180. doi: 10.1111/j.1439-0523.2009.01669.x
Stuber, C. W., Lincoln, S. E., Wolff, D., Helentjaris, T., and Lander, E. (1992). Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics 132, 823–839.
Sundaram, R. M., Vishnupriya, M., Laha, G. S., Rani, N. S., Rao, P. S., Balachandran, S. M., et al. (2009). Introduction of bacterial blight resistance into Triguna, a high yielding, mid-early duration rice variety. Biotechnol. J. 4, 400–407. doi: 10.1002/biot.200800310
Tabien, R., Li, Z., Paterson, A., Marchetti, M., Stansel, J., and Pinson, S. (2002). Mapping QTLs for field resistance to the rice blast pathogen and evaluating their individual and combined utility in improved varieties. Theor. Appl. Genet. 105, 313–324. doi: 10.1007/s00122-002-0940-2
Talukder, Z. I., Mcdonald, A. J. S., and Price, A. H. (2005). Loci controlling partial resistance to rice blast do not show marked QTL × environment interaction when plant nitrogen status alters disease severity. New Phytol. 168, 455–464. doi: 10.1111/j.1469-8137.2005.01507.x
Talukder, Z. I., Tharreau, D., and Price, A. H. (2004). Quantitative trait loci analysis suggests that partial resistance to rice blast is mostly determined by race-specific interactions. New Phytol. 162, 197–209. doi: 10.1111/j.1469-8137.2004.01010.x
Tanksley, S. (1993). QTL analysis of transgressive segregation in an interspecific tomato cross. Genetics 134, 585–596.
Tanksley, S. D. (1988). Resolution of quantitative traits into Mendelian factors by using a complete linkage map of restriction fragment length polymorphisms. Nature 335, 721–726. doi: 10.1038/335721a0
Tanksley, S., Grandillo, S., Fulton, T., Zamir, D., Eshed, Y., Petiard, V., et al. (1996). Advanced backcross QTL analysis in a cross between an elite processing line of tomato and its wild relative L. pimpinellifolium. Theor. Appl. Genet. 92, 213–224. doi: 10.1007/BF00223378
Tanweer, F. A., Rafii, M. Y., Sijam, K., Rahim, H. A., Ahmed, F., and Latif, M. A. (2015). Current advance methods for the identification of blast resistance genes in rice. C. R. Biol. 338, 321–334. doi: 10.1016/j.crvi.2015.03.001
Temnykh, S., Declerck, G., Lukashova, A., Lipovich, L., Cartinhour, S., and Mccouch, S. (2001). Computational and experimental analysis of microsatellites in rice (Oryza sativa L.): frequency, length variation, transposon associations, and genetic marker potential. Genome Res. 11, 1441–1452. doi: 10.1101/gr.184001
Thakur, S., Gupta, Y., Singh, P., Rathour, R., Variar, M., Prashanthi, S., et al. (2013a). Molecular diversity in rice blast resistance gene Pi-ta makes it highly effective against dynamic population of Magnaporthe oryzae. Funct. Integr. Genom. 13, 309–322. doi: 10.1007/s10142-013-0325-4
Thakur, S., Singh, P., Rathour, R., Variar, M., Prashanthi, S., Singh, A., et al. (2013b). Positive selection pressure on rice blast resistance allele Piz-t makes it divergent in Indian land races. J. Plant Inter. 8, 34–44.
Toojinda, T., Tragoonrung, S., Vanavichit, A., Siangliw, J. L., Pa-in, N., Jantaboon, J., et al. (2005). Molecular breeding for rainfed lowland rice in the Mekong region. Plant Prod. Sci. 8, 330–333. doi: 10.1626/pps.8.330
Urso, S., Orasen, G., Perrini, R., Tacconi, G., Delfanti, S., Biselli, C., et al. (2013). “Pyramiding of Pi resistance genes to increase blast resistance in Italian rice varieties using marker-assisted selection approaches,” in Proceedings of the 57th Italian Society of Agricultural Genetics Annual Congress, 16th-19th September, Foggia.
Utani, D. W., Moeljopawiro, S., Aswidinnoor, H., Setiawan, A., and Hanarida, I. (2008). Blast resistance genes in wild rice Oryza rufipogon and rice cultivar IR64 Indonesian. J. Agric. 1, 71–76.
Wan, X. Y., Wan, J. M., Jiang, L., Wang, J. K., Zhai, H. Q., Weng, J. F., et al. (2006). QTL analysis for rice grain length and fine mapping of an identified QTL with stable and major effects. Theor. Appl. Genet. 112, 1258–1270. doi: 10.1007/s00122-006-0227-0
Wang, D., Guo, C., Huang, J., Yang, S., Tian, D., and Zhang, X. (2014). Allele-mining of rice blast resistance genes at AC134922 locus. Biochem. Biophys. Res. Commun. 446, 1085–1090. doi: 10.1016/j.bbrc.2014.03.056
Wang, G.-L., Mackill, D. J., Bonman, J. M., Mccouch, S. R., Champoux, M. C., and Nelson, R. J. (1994). RFLP mapping of genes conferring complete and partial resistance to blast in a durably resistant rice cultivar. Genetics 136, 1421–1434.
Wang, Z., Jia, Y., Rutger, J., and Xia, Y. (2007). Rapid survey for presence of a blast resistance gene Pi-ta in rice cultivars using the dominant DNA markers derived from portions of the Pi-ta gene. Plant Breed. 126, 36–42. doi: 10.1111/j.1439-0523.2007.01304.x
Wen, S., and Gao, B. (2011). Introgressing blast resistant gene Pi-9 (t) into elite rice restorer Luhui17 by marker-assisted selection. Rice Genom. Genet. 2, doi: 10.5376/ rgg.2011.02.0004
Wu, J.-L., Fan, Y.-Y., Li, D.-B., Zheng, K.-L., Leung, H., and Zhuang, J.-Y. (2005). Genetic control of rice blast resistance in the durably resistant cultivar Gumei 2 against multiple isolates. Theor. Appl. Genet. 111, 50–56. doi: 10.1007/s00122-005-1971-2
Xu, J., Wang, J., Ling, Z., and Zhu, L. (2004). Analysis of rice blast resistance genes by QTL mapping. Chinese Sci. Bull. 49, 337–342. doi: 10.1007/BF02900315
Xu, X., Lv, Q., Shang, J., Pang, Z., Zhou, Z., Wang, J., et al. (2014). Excavation of Pid3 orthologs with differential resistance spectra to Magnaporthe oryzae in rice resource. PLoS ONE 9:e93275. doi: 10.1371/journal.pone.0093275
Xu, Y. (2002). “Global View of QTL: rice as a model,” in Quantitative Genetics, Genomics, and Plant Breeding, ed. M. S. Kang (Willingford: CAB International), 109–134.
Yang, M.-Z., Cheng, Z.-Q., Chen, S.-N., Qian, J., Xu, L.-L., and Huang, X.-Q. (2007). A rice blast-resistance genetic resource from wild rice in Yunnan. China. J. Plant Physiol. Mol. Biol. 33, 589–595.
Yasuda, N., Mitsunaga, T., Hayashi, K., Koizumi, S., and Fujita, Y. (2014). Effects of pyramiding quantitative resistance genes pi21, Pi34, and Pi35 on rice leaf blast disease. Plant Dis. 99, 904–909. doi: 10.1094/PDIS-02-14-0214-RE
Yoshida, K., and Miyashita, N. T. (2009). DNA polymorphism in the blast disease resistance gene Pita of the wild rice Oryza rufipogon and its related species. Genes Genet. Syst. 84, 121–136. doi: 10.1266/ggs.84.121
Young, N. D. (1994). “Constructing a plant genetic linkage map with DNA markers,” in DNA-Based Markers in Plants, eds I. K. V. Ronald and L. Phillips (Dordrecht: Kluwer), 39–57.
Young, N. D. (1996). QTL mapping and quantitative disease resistance in plants. Annu. Rev. Phytopathol. 34, 479–501. doi: 10.1146/annurev.phyto.34.1.479
Yu, J., Hu, S., Wang, J., Wong, G. K.-S., Li, S., Liu, B., et al. (2002). A draft sequence of the rice genome (Oryza sativa L. ssp. indica). Science 296, 79–92. doi: 10.1126/science.1068037
Zeigler, R. S., Leong, S. A., and Teng, P. S. (1994). Rice Blast Disease. Wallingford: CAB International.
Zenbayashi, K., Ashizawa, T., Tani, T., and Koizumi, S. (2002). Mapping of the QTL (quantitative trait locus) conferring partial resistance to leaf blast in rice cultivar Chubu 32. Theor. Appl. Genet. 104, 547–552. doi: 10.1007/s00122-001-0779-y
Zhang, H., Li, G., Li, W., and Song, F. (2009). Transgenic strategies for improving rice disease resistance. Afri. J. Biotechnol. 8, 1750–1757. doi: 10.1007/978-1-62703-194-3_21
Zhang, Y., Yang, J., Shan, Z., Chen, S., Qiao, W., Zhu, X., et al. (2012). Substitution mapping of QTLs for blast resistance with SSSLs in rice (Oryza sativa L.). Euphytica 184, 141–150. doi: 10.1007/s10681-011-0601-0
Zhu, X., Chen, S., Yang, J., Zhou, S., Zeng, L., Han, J., et al. (2012). The identification of Pi50 (t), a new member of the rice blast resistance Pi2/Pi9 multigene family. Theor. Appl. Genet. 124, 1295–1304. doi: 10.1007/s00122-012-1787-9
Keywords: rice blast disease, molecular breeding, DNA markers, QTL mapping, marker-aided selection, gene pyramiding
Citation: Ashkani S, Rafii MY, Shabanimofrad M, Miah G, Sahebi M, Azizi P, Tanweer FA, Akhtar MS and Nasehi A (2015) Molecular Breeding Strategy and Challenges Towards Improvement of Blast Disease Resistance in Rice Crop. Front. Plant Sci. 6:886. doi: 10.3389/fpls.2015.00886
Received: 19 June 2015; Accepted: 06 October 2015;
Published: 16 November 2015.
Edited by:
Tiegang Lu, Chinese Academy of Agricultural Sciences, ChinaCopyright © 2015 Ashkani, Rafii, Shabanimofrad, Miah, Sahebi, Azizi, Tanweer, Akhtar and Nasehi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sadegh Ashkani, YXNoa2FuaS5zYWRlZ2hAdXBtLmVkdS5teQ==; Mohd Y. Rafii, bXJhZmlpQHVwbS5lZHUubXk=