 Qing Li
Qing Li Jianhua Shao
Jianhua Shao Yueyun Hong
Yueyun Hong- National Key Laboratory of Crop Genetic Improvement, College of Life Science and Technology, Huazhong Agricultural University, Wuhan, China
Wrinkled1 (WRI1) belongs to the APETALA2 transcription factor family; it is unique to plants and is a central regulator of oil synthesis in Arabidopsis. The effects of WRI1 on comprehensive lipid metabolism and plant development were unknown, especially in crop plants. This study found that BnWRI1 in Brassica napus accelerated flowering and enhanced oil accumulation in both seeds and leaves without leading to a visible growth inhibition. BnWRI1 decreased storage carbohydrates and increased soluble sugars to facilitate the carbon flux to lipid anabolism. BnWRI1 is localized to the nucleus and directly binds to the AW-box at proximal upstream regions of genes involved in fatty acid (FA) synthesis and lipid assembly. The overexpression (OE) of BnWRI1 resulted in the up-regulation of genes involved in glycolysis, FA synthesis, lipid assembly, and flowering. Lipid profiling revealed increased galactolipids monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG), and phosphatidylcholine (PC) in the leaves of OE plants, whereas it exhibited a reduced level of the galactolipids DGDG and MGDG and increased levels of PC, phosphatidylethanolamide, and oil [triacylglycerol (TAG)] in the siliques of OE plants during the early seed development stage. These results suggest that BnWRI1 is important for homeostasis among TAG, membrane lipids and sugars, and thus facilitates flowering and oil accumulation in B. napus.
Introduction
Lipids not only serve as storage components of high-density energy, but they also function as essential components of cell membranes and regulators of various cellular processes during growth, development, and stress responses (Wang et al., 2006; Hong et al., 2008, 2009; Phillips et al., 2009; To et al., 2012). Fatty acid (FA) synthesis and lipid assembly involve multiple steps (Li-Beisson et al., 2010). The initial precursors of lipid biosynthesis include acetyl-CoA and glycerol-3-phosphate, which are initially derived from glycolysis and the Calvin–Benson cycle in plants (Kang and Rawsthorne, 1996; Alonso et al., 2007). The acetyl-CoA carboxylase (ACCase) complex is made of three subunits, namely biotin carboxyl carrier protein (BCCP), biotin carboxylase (BC), and carboxyltransferase (CT); this complex is encoded by separated genes in plants and catalyzes acetyl-CoA and CO2 to produce malonyl-CoA, the first committed step in de novo FA synthesis (Slabas and Fawcett, 1992; Ohlrogge and Browse, 1995; Ohlrogge and Jaworski, 1997; Voelker and Kinney, 2001; Sasaki and Nagano, 2004). Malonyl-CoA was then transferred to ACP protein by malonyl-CoA: ACP transferase (MAT) to initiate FA synthesis, and malonyl-CoA provides a two-carbon unit for acyl chain elongation as catalyzed by an FA synthase (FAS) complex (Ohlrogge and Browse, 1995). The synthesized FAs are either retained in the chloroplast for galactolipid synthesis or exported to the endoplasmic reticulum (ER) for membrane phospholipid and storage lipid [triacylglycerol (TAG)] assembly. The final step of TAG assembly as catalyzed by DAG acyltransferase (DGAT), which occurs in the Kennedy pathway, is also regarded as a critical reaction for oil accumulation (Slabas and Fawcett, 1992; Ohlrogge and Browse, 1995; Zou et al., 1999; Voelker and Kinney, 2001). The loss of DGAT1 resulted in reduced seed oil content, whereas DGAT1 overexpression (OE) enhanced the seed oil content in Arabidopsis (Zou et al., 1999; Jako et al., 2001). The OE of maize high-oil DGAT1-2 also promoted oil accumulation in maize seeds (Zheng et al., 2008). Alternatively, phosphatidylcholine (PC) also provides an acyl chain toward DAG for TAG synthesis, as catalyzed by phospholipid:diacylglycerol acyltransferase (PDAT; Dahlqvist et al., 2000; Tarczynski and Shen, 2008; Zhang et al., 2009).
Given the complicated networks that make up the lipid anabolic process, it would be more efficient to boost oil accumulation by enhancing multiple routes in a coordinated fashion including carbon partitioning, FA synthesis, and lipid assembly. Therefore, the identification of the key enzymes or master regulators involved in multiple steps simultaneously becomes an attractive approach for improving oil production (Ohlrogge and Jaworski, 1997; Ruuska et al., 2002; Cahoon et al., 2007; Mu et al., 2008). Transcriptomic profiling revealed that the genes encoding enzymes involved in FA synthesis is co-regulated to the rate of acyl chain synthesis, suggesting that transcriptional regulation plays an important role in the lipid biosynthesis process (Ruuska et al., 2002; Baud and Lepiniec, 2009; Barthole et al., 2012). Recent studies have identified several transcription factors that are capable of governing multiple oil accumulation steps (Cernac and Benning, 2004; Shen et al., 2010; To et al., 2012). Wrinkled1 (WRI1) belongs to the APETALA2 (AP2)-ethylene-responsive element binding protein family of transcription factors, and it acts as a central regulator in seed oil accumulation by modulating numerous genes simultaneously during late glycolysis and FA biosynthesis. A deficiency mutant of Arabidopsis WRI1 (AtWRI1) leads to wrinkled seeds with 80% less seed oil content in Arabidopsis (Focks and Benning, 1998; Cernac and Benning, 2004). The loss of AtWRI1 also leads to impaired seed germination and seedling establishment, whereas AtWRI1 OE enhances oil accumulation in Arabidopsis, which is accompanied by aberrant seedling development (Stone et al., 2001; Kwong et al., 2003; Cernac and Benning, 2004; Cernac et al., 2006; Baud et al., 2007). AtWRI1 binds to the AW-box consensus [CnTnG](n)7[CG] in the proximal promoter of target genes that are involved in the glycolysis and FA synthesis of Arabidopsis (Maeo et al., 2009; To et al., 2012).
The biological significance of WRI1 has been extensively studied in relation to oil accumulation in Arabidopsis. The role of WRI1 in comprehensive lipid regulation in other plant species, particularly in crop plants, remains to be elucidated. A recent study showed that the WRI1 homolog from maize is able to compensate for the impaired oil accumulation and seedling establishment of the Atwri1 mutant in Arabidopsis (Cernac et al., 2006; Pouvreau et al., 2011). The OE of ZmWRI1 in maize increased the levels of FAs and some amino acid residues (Pouvreau et al., 2011), suggesting that the role of WRI1 in oil accumulation is highly conserved between monocot and dicot plants. Arabidopsis that overexpresses AtWRI1 exhibits undesirable agronomic traits, with retarded growth and reduced biomass (Lotan et al., 1998; Stone et al., 2001; Cernac and Benning, 2004; Wang et al., 2007; Mu et al., 2008), whereas ZmWRI1 OE in maize promotes oil accumulation without visible side effects on growth and development (Shen et al., 2010). The results suggest that the role of WRI1 in oil synthesis is conserved but distinguishable in different plant species. The WRI1 in different plant species may exhibit a unique role in addition to its effect on oil accumulation. Furthermore, most WRI1 studies have been focused on oil accumulation, and the effects of WRI1 on membrane phospholipids and galactolipids have remained unknown. In the present study, we characterized BnWRI1 (BnaA09g34250D) from Brassica napus (B. napus). BnWRI1 OE in B. napus resulted in enhanced lipid anabolism by binding to the cis-element CnTnG (n)7CG in the promoter regions of genes involved in FA synthesis and lipid assembly to up-regulate these target genes. BnWRI1 promotes oil accumulation and thylakoid membrane monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG), and PC biosynthesis to regulate homeostasis among membrane lipids, oils, and carbohydrates. Therefore, BnWRI1 OE facilitates flowering, reproduction, and oil production without visible side effects on the growth of B. napus.
Materials and Methods
Plant Materials and Growth Conditions
The Westar cultivar of canola (B. napus L.) was used in this study. Seeds were germinated in either Murashige and Skoog (MS) plates or soil in pots. Three-week-old seedlings were then transferred to pots containing soil. The plants were raised in a growth room under 16 h light (25°C)/8 h dark (20°C), a photosynthetic photon flux density of 200–300 mmol m-2 s-1, and 60% relative humidity, or natural conditions during winter-spring seasons in Wuhan, China. For the field growth test, 3-week-old seedlings were transferred to the field at suitable spacing (33 cm × 50 cm) that were arranged in a one-way randomized block design with 30 plants/lines per block, and three replications.
Gene Cloning, Vector Construction, and B. napus Plant Transformation
To obtain the full-length BnWRI1 cDNA, total RNA was extracted from the leaves of 4-week-old B. napus plants, and it was subjected to reverse transcription to obtain first strand cDNA according to the manufacturer’s instructions (TransGene Biotech, Beijing, China). The full-length BnWRI1 cDNA was amplified by PCR with primers BnWRI1F 5′-GGATCCATGAAGAGACCCTTAACCACT-3′ and BnWRI1R 5′-GAGCTCTCAGACAGAATAGTTCCAAGAA-3′, and then it was ligated into binary vector pBI121, which had been digested by SacI and BamHI. The resulting construct was transformed into B. napus by Agrobacterium GV3101 mediation with hypocotyls used as explants for regeneration. The transgenic shoots were first selected on kanamycin (50 μg/ml), and then the kanamycin resistant shoots were transferred to MS medium containing 1-naphthaleneacetic acid for rooting. The transgenic plants were further confirmed by PCR with a pBI121 vector and BnWRI1 sequence specific primers (Supplemental Table S1).
Subcellular Localization
The full-length cDNA of BnWRI1 was ligated into pCAMBIA1301 vector that had been digested by the restriction enzymes SacI and BamHI. The construct containing BnWRI1-GFP was introduced into Agrobacterium GV3101 and infiltrated into tobacco leaves for 24 h to obtain transient protein expression under the control of the 35S promoter. Subcellular localization was visualized under a confocal laser scanning microscope (Leica, Biberach, Germany) with the exciter filter HFT488 and the transmitting optical filter BP505–530 to observe the green fluorescence. The nuclei were labeled with 4′,6-diamidine-2-phenylindole dihydrochloride (DAPI) staining.
RNA Extraction and Quantitative Real-time PCR
Total RNA was extracted from various tissues at different stages using TransZol reagent (TransGen Biotech, Beijing, China), and it was then treated with RNase-free DNaseI (NEW ENGLAND Biolabs, Ipswitch, MA, USA) to remove any contaminating DNA. The resulting RNA was used for first strand synthesis by reverse transcriptase with an oligo-d (T) 18 primer (TransGen Biotech, Beijing, China) to obtain cDNA according the manufacturer’s protocol. Quantitative real-time PCR was performed with SYBR Green PCR Master Mix (TransGen Biotech, Beijing, China) on a single-color Real-time PCR Detection System (Bio-Rad, Hercules, CA, USA). A BnActin gene was used as the standard control. The quantitative real-time PCR conditions were as follows: 95°C for 1 min; 40 cycles of 95°C for 30 s, 55°C for 30 s, 72°C for 30 s; and 72°C for 10 min for the final extension. The primers used for real-time PCR are listed in Supplemental Table S2.
Lipid Extraction and Analyses
Lipids were extracted from the leaves, developing siliques, and mature seeds. The lipids were separated on a thin layer chromatography (TLC) plate with developing solvent consisting of petroleum ether, ethyl ether, and acetic acid (80:20:1, v/v). The separated lipids were visualized with iodine vapor and the spots were scraped for measurement by GC analysis (Agilent 7890A, Santa Clara, CA, USA) after a methyl ester reaction with methanol and toluene containing 5% H2SO4 at 80°C for 3–4 h. To measure the seed oil contents, oil was extracted from the seeds and tested by GC analysis after the methyl ester reaction as detailed above. The GC running conditions were as follows: the injection port temperature was 180°C, and the oven temperature was set at 180°C for 2 min and was increased by 10°C/min up to 220°C for 5 min. The temperature of the flame ionization detector was 280°C with flow rates of 30, 300, and 25 ml/min for hydrogen, air, and helium, respectively.
Electrophoretic Mobility-shift Assay
The full-length cDNA of Bn WRI1 was amplified by using the forward primer 5′-CCCGGGTATGAAGAGACCCTTAACCAC-3′ coupled with the reverse primer 5′-GGATCCCGACAGAATAGTTCCAAGAA-3′, and then it was ligated to pET28a vectors that were digested by BamHI and SacI. This construct was transformed into Escherichia coli strain Rosetta (DE3), and the BnWRI1 protein was expressed by induction with 0.6 mM isopropyl-β-D-thiogalactopyranoside (IPTG) while the strain was grown in Luria Bertani (LB) medium overnight at 20°C. The cells were harvested and lysed by sonication in a buffer (300 mM NaCl, 20 mM Tris-HCl, pH 8.0, 10 mM imidazole, 5% glycerol, and 50 mM NaH2PO4). The cell lysate was centrifuged at 12,000 r/min for 20 min. The supernatant was incubated with Ni-NTA resin (Shanghai Sangon, http://www.sangon.com) for 3 h at 4°C. The BnWRI1 protein was eluted from the resin after three washes with wash buffer (50 mM NaH2PO4, 300 mM NaCl, and 20 mM imidazole, pH 8). Protein from E. coli cells containing only the pET28a vector was used as a negative control. The DNA sequences that were 300 and 250 bp upstream from the start codon of KASI and GPAT9, respectively, were amplified from Arabidopsis with the KASI forward primer 5′-GAATTCTGTTGAGTTACGAATTGGAG-3′, coupled with the KASI reverse primer 5′-GAGCTCATTGAGAGAGGTATTGAGAG-3′, and the GPAT9 forward primer 5′-GAATTCACATAATATGTCCAAGATCATT-3′ coupled with the GPAT9 reverse primer 5′-GAGCTCCTATTATACTTATACCACAT-3′. The substitutive nucleotide (C→T, T→C, G→A) mutant at the AW-box [CnTnG](n)7[CG] was amplified by using a similar approach. The amplified DNA fragments containing native or mutant AW-box were incubated with purified BnWRI1 protein in binding buffer (20 mM Tris-HCl, pH 8.0, 250 mM NaCl, 2 mM MgCl2, 1% glycerol, 1 mg/ml BSA, 1 mM DTT) for 1 h at 4°C. The resulting mixture was separated on native PAGE (6%) by electrophoresis and was visualized under UV light. The binding activity of BnWRI1 to the AW-box was also determined with biotin labeled DNA probes using a chemiluminescent electrophoretic mobility-shift assays (EMSA) kit (Beyotime, China) according to the manufacturer’s instructions.
Measurements of Protein, Starch, and Soluble Sugar
Proteins were extracted from leaves and seeds by homogenizing in buffer containing 50 mM Tris-HCl, pH 8.0, 250 mM NaCl, 1 mM EDTA, and 1% (w/v) SDS, and incubating the mixture for 2 h at 25°C. The homogenate was centrifuged at 16,000 g for 10 min, the supernatant was diluted 200 times and the protein concentration was measured by Lowry D protein assay (Bio-Rad). Soluble sugars were measured using phenol-sulfuric acid method (DuBois et al., 1956; Chow and Landhausser, 2004). In brief, leaf samples (1 g fresh weight) were homogenized with deionized water and filtered. The extract (50 μl) was mixed with 450 μl of sulfuric acid containing anthrone (2 mg/ml) at 95°C, and then the absorbance at 625 nm was monitored by spectrometer (Infinite M200 PRD, Untersbergstr, Austria). For starch extraction, the remaining sediment was suspended in a solution containing 0.2 N KOH and incubated at 95°C for 1 h, followed by the addition of 1 N acetic acid and incubation for 15 min. After centrifugation at 16,000 g for 5 min, the starch in the supernatant was measured by using a method similar to that of soluble sugars.
Results
Expression Pattern and the Effect of BnWRI1 on Flowering in B. napus
To investigate the temporal and spatial distribution of BnWRI1 mRNA in B. napus, total RNA was extracted from various tissues at different stages and used for analysis by quantitative real-time PCR. During the seedling and bolting stages, the BnWRI1 transcript level was higher in leaves and flower buds than it was in roots and stems (Figure 1A). During the flowering stage, the BnWRI1 expression was higher in flowers than in leaves and stems. The transcript level was rapidly up-regulated in siliques and was highest at 30 days after anthesis (Figure 1A).
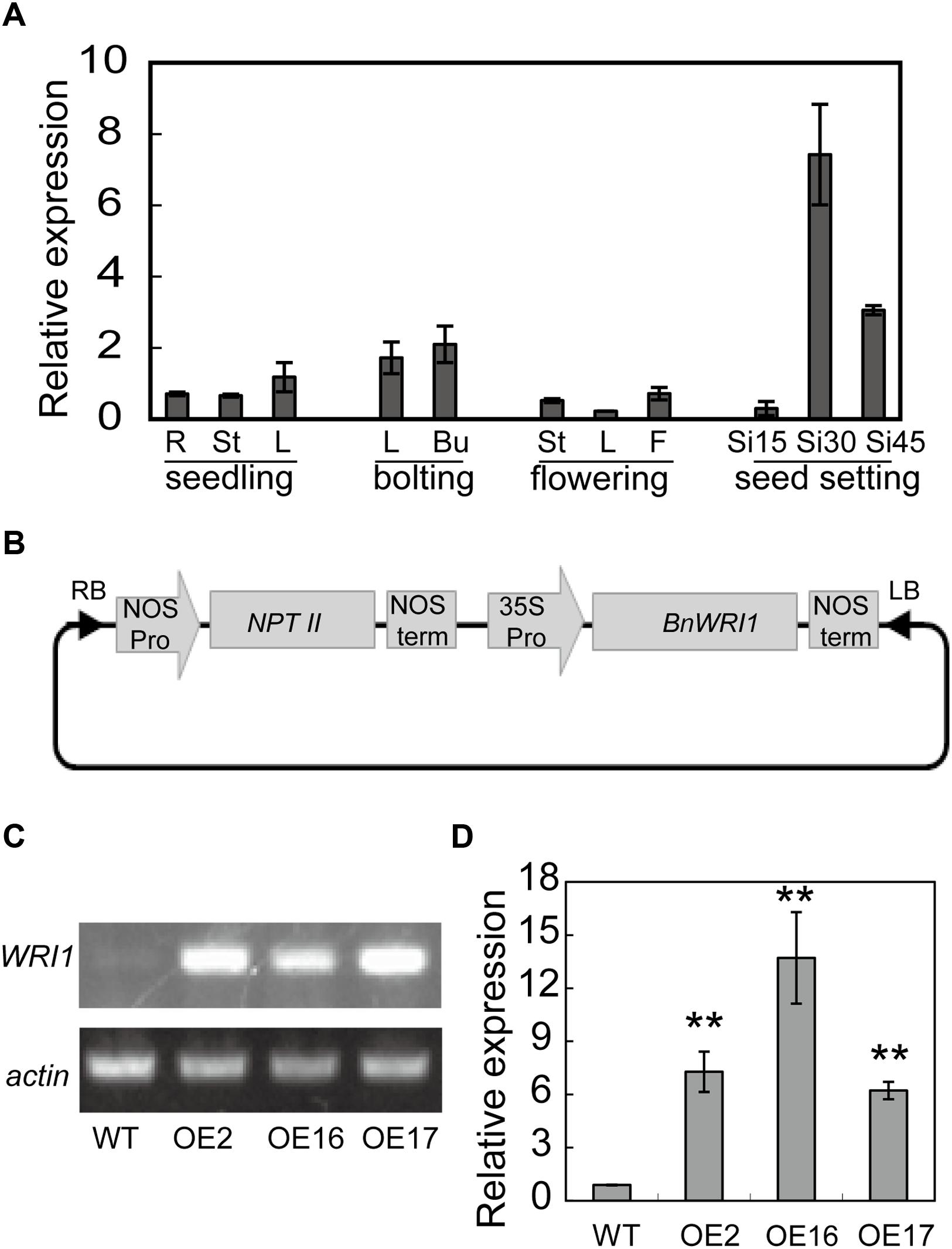
FIGURE 1. BnWRI1 expression pattern and overexpression (OE) of BnWRI1 in Brassica napus. (A) Transcript level of BnWRI1 in different B. napus tissues at seedling, bolting, flowering and seed developing stages. The expression level was quantified by real-time PCR normalized to the expression of BnActin. Values are mean ± SD (n = 3). R, root; St, stem; L, leaf; Bu, flower bud; F, flower; and Si15, Si30, and Si45, siliques at 15, 30, and 45 days, respectively, after anthesis. (B) The construct containing BnWRI1 in the binary vector pBI121. (C,D) The transcript level of BnWRI1 in BnWRI1-OE plants according to semi-quantitative RT-PCR (C) and quantitative real-time PCR (D). Total RNA was extracted from the leaves of 6-week-old plants, and the expression level was detected by using BnWRI1-specific primers. The expression levels were normalized to BnActin. OE2, OE16, and OE16 represent BnWRI1-OE lines. Values are mean ± SD (n = 3). ∗∗Indicates significant difference at P < 0.01 compared with the WT based on Student’s t-test.
To explore the biological function of BnWRI1 in B. napus, full-length BnWRI1 cDNA was cloned by reverse transcription PCR by using mRNA that was extracted from leaves as a template, and the cDNA was ligated into binary vector pBI121. The resulting construct containing BnWRI1 was transformed into B. napus under the control of the 35S promoter (Figure 1B). More than 30 independent transgenic lines were obtained, and the BnWRI1 transcript level in transgenic plants was much higher than that of the wild-type (WT; Figures 1C,D). Three representative, independent BnWRI1 OE lines, OE2, OE16, and OE17, were selected randomly from 30 transgenic lines, and they were used for further characterization. These plants were grown under natural conditions either in the field or in pots, and no visual growth change was observed between OE and WT plants, which showed a similar leaf size, leaf number, and growth rate during the vegetative growth stage (Figure 2A). However, BnWRI1 accelerated flowering; OE plants bolted and flowered 4 to 6 days earlier than WT plants (Figures 2B,C). At 136 days after germination, 60% of the OE plants were flowering, whereas only 23% of the WT plants were flowering (Figure 2D). The earlier flowering in OE plants did not cause changes in the total number of inflorescent branches and biomass at the mature stage compared with the results for the WT plants (Figures 2E,F).
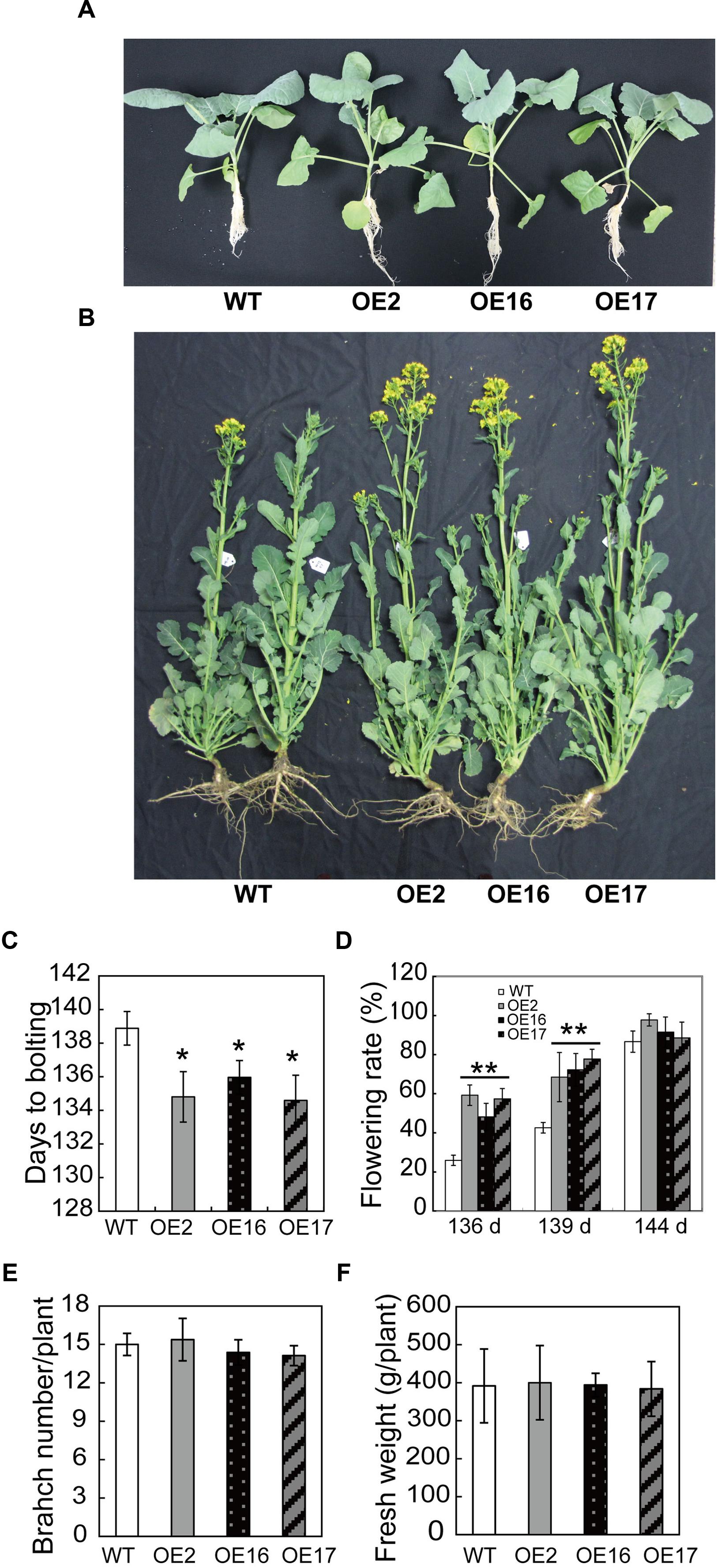
FIGURE 2. Accelerated flowering in BnWRI1-OE plants. (A) BnWRI1-OE and WT plants at the vegetative growth stage. The pictures were taken of plants that were grown in pots at 30 days after germination. (B) Accelerated flowering in BnWRI1-OE plants. The picture was taken of 145-day-old plants grown in the field. (C) Days to bolting based on a flowering rate of 50% (n = 20, r = 3). (D) The flowering rate of OE and WT plants grown under the same conditions. Values are mean ± SD (n = 20, r = 3). (E) Inflorescent branch number of BnWRI1-OE and WT. The data were collected from mature plants (175-day-old) grown in the field (n = 12, r = 3). (F) The fresh weights of aerial part from 175-day-old plants grown in the field. Values are mean ± SD (n = 12, r = 3). ∗,∗∗Indicate significant differences at P < 0.05 and P < 0.01, respectively, compared with the WT based on Student’s t-test.
Overexpression of BnWRI1 Enhances Oil Accumulation in Seeds and Leaves without Undesirable Agronomic Traits
To investigate the role of BnWRI1 in oil (TAG) synthesis, the oil content was measured in both the seeds and leaves of OE and WT plants. The oil content of BnWRI1-OE seeds was significantly higher than that of the WT, and it was increased by 31, 38, and 18% in OE2, OE16, and OE17, respectively (Figure 3A). FA profiling revealed that oleic acid (18:1) contributed mostly to oil accumulation in OE plants. Other FA species such as 16:0 and 18:2 also increased in OE seeds (Figure 3B). The OE of BnWRI1 also increased the oil contents of vegetative tissues. The oils in the leaves were primarily composed of 16:0, 18:1, and 18:0 FA species, which account for ∼90% of the total FA species. The total TAG contents of the leaves from OE2, OE16, and OE17 increased by 28, 67, and 63%, respectively, in comparison with the WT plants (Figure 3C). The enhanced TAG in the OE leaves resulted from the increase of 16:0, 18:0, 18:1, and 18:2 FA species (Figure 3D). The relative increase in leaf oil content from BnWRI1 OE was greater than that of the seed oil content.
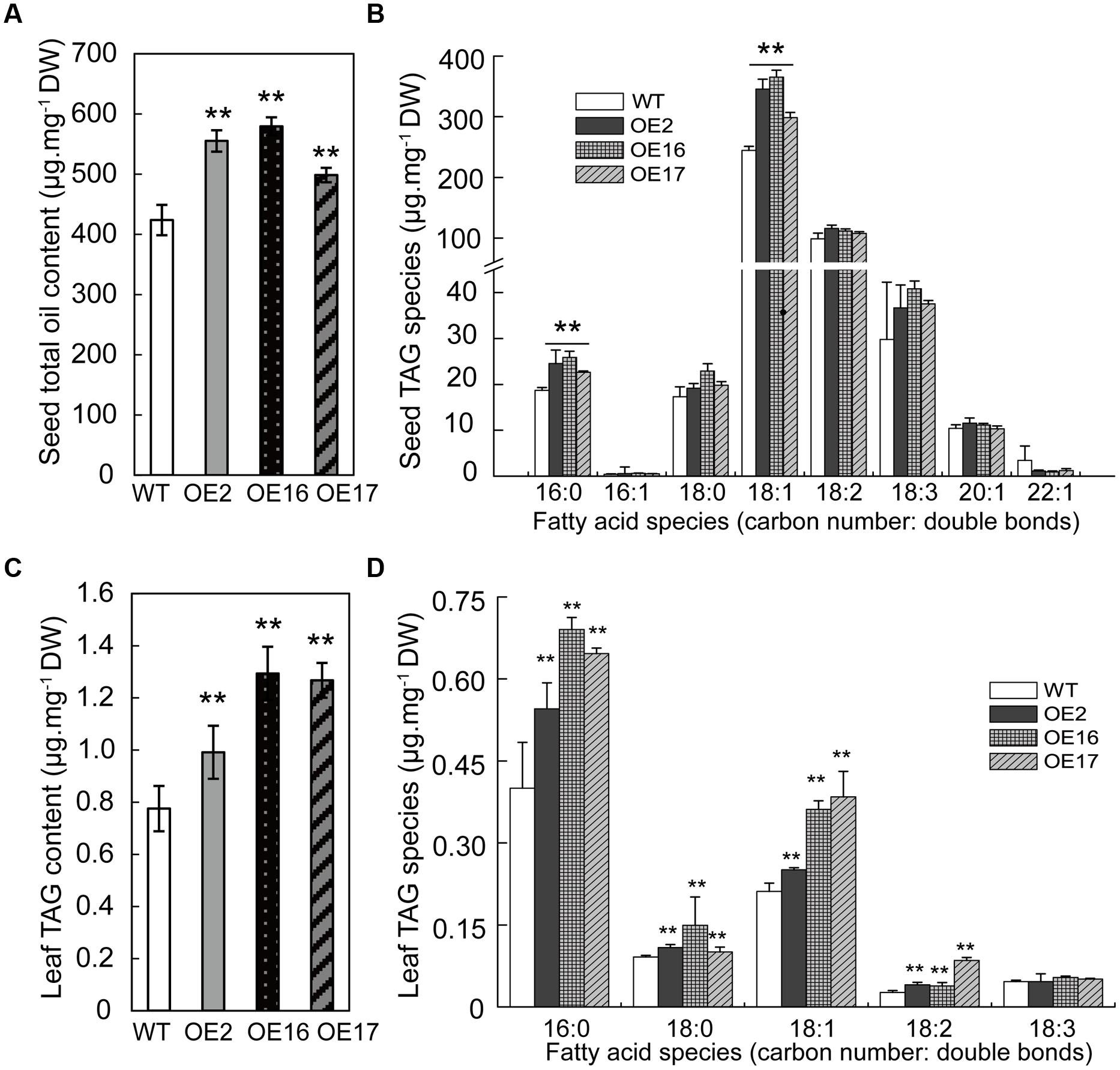
FIGURE 3. Enhanced seed oil accumulation in BnWRI1-OE plants. Total TAG content (A) and fatty acid (FA) composition (B) in the seeds of BnWRI1-OE and WT. Total TAG content (C) and FA composition (D) in the leaves of BnWRI1-OE and WT. Lipids were extracted from the leaves of 8-week-old plants and were separated by using a thin layer chromatography (TLC) plate. TAG spots were scraped and extracted for methylation and GC measurement. Values are mean ± SD (n = 4); ∗∗P < 0.01.
The Effect of BnWRI1 on Total Lipids, Sugar, and Protein Accumulation
Carbohydrates are photosynthesized in green tissues, predominantly in leaves, which are the major source tissue, providing precursors for lipid and protein synthesis. Carbon partitioning among these metabolites is a major factor that influences lipid accumulation. To determine whether enhanced oil accumulation resulted from alterations in carbon partitioning, the contents of the total lipids, soluble sugars, starch, and proteins were measured in the leaves of 3-month-old plants at the flowering stage. The total lipid content of OE leaves was significantly higher than that of WT, and it was increased by 24, 47, and 25% for OE2, OE16, and OE17, respectively (Figure 4A). BnWRI1-OE leaves exhibited increased soluble sugars with reduced starch contents compared with WT plants (Figures 4B,C). However, the total sugars and total protein content in OE leaves were not substantially different from those of WT plants (Figures 4B,C). In comparison with those in the leaves, the contents of soluble sugars, starch, total sugars, and proteins in mature seeds were less altered between the OE and WT (Figures 4D,E). The starch contents of OE2 and OE16 seeds were lower than that of the WT, whereas the soluble sugar, total sugar, and protein contents in OE seeds were not significantly different from that of the WT (Figures 4D,E). These results indicate that enhancing the lipid content by overexpressing BnWRI1 without reducing sugars and proteins, but enhanced leaf sugar moves from storage starch to soluble sugars for lipid accumulation.
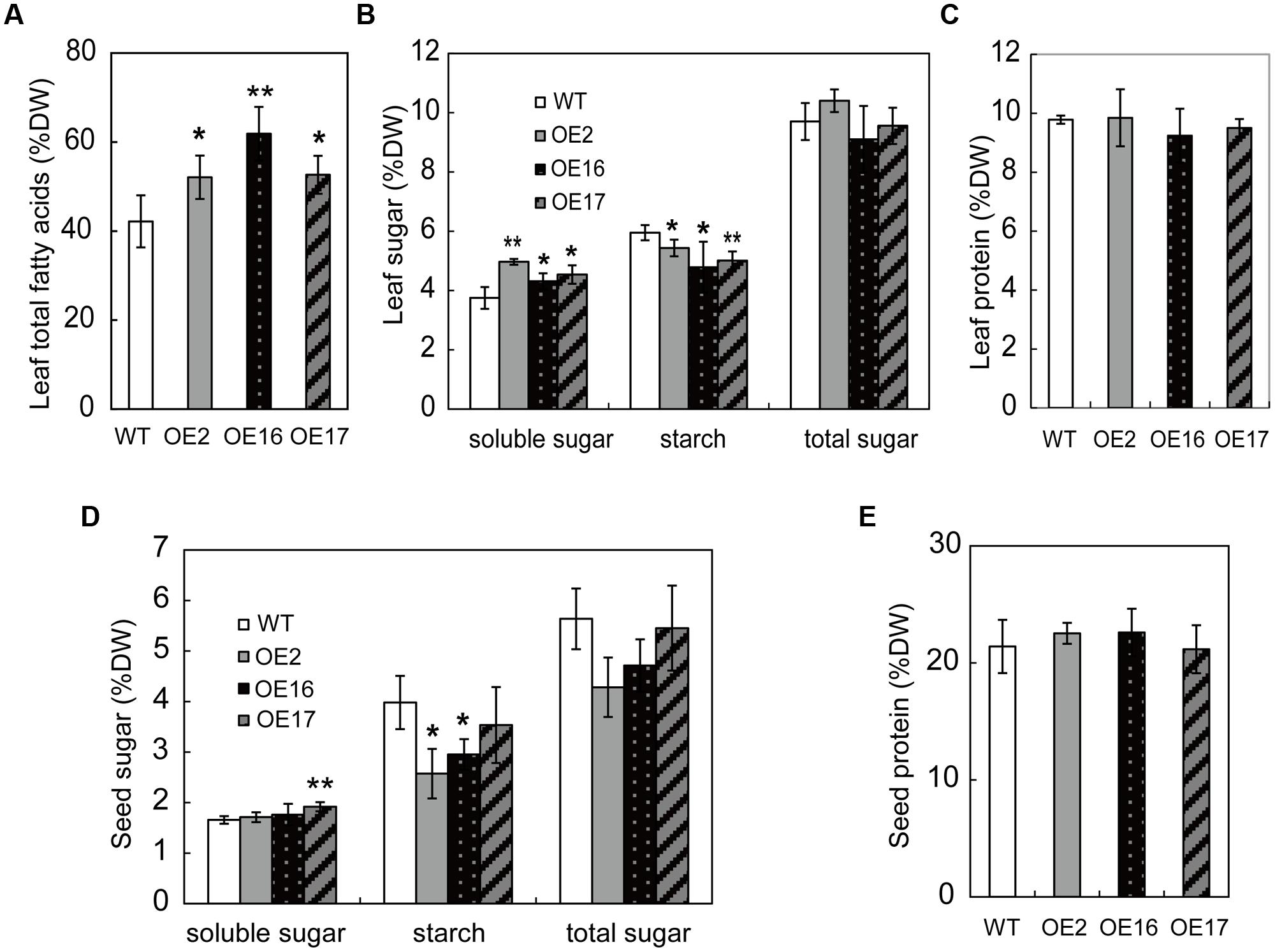
FIGURE 4. The effect of BnWRI1 on partitioning among lipids, sugars, and proteins in B. napus. (A) Total FA contents in leaves from BnWRI1-OE and WT plants. (B,C) The soluble sugars, starch, total sugars, and proteins in the leaves of 2-month-old plants. (D,E) The soluble sugar, starch, total sugar, and protein contents in mature seeds. Values are mean ± SD (n = 3). DW, dry weight; ∗P < 0.05, ∗∗P < 0.01.
BnWRI1 Binds to the Proximal Upstream Regions of Genes Involved in Lipid Anabolism
To get insight into the molecular mechanism of BnWRI1 in lipid metabolism and carbon partitioning, the subcellular localization of BnWRI1 and its putative target genes involved in lipid metabolism were investigated. The full-length cDNA of BnWRI1 was fused with GFP at the C-terminus and then transiently expressed in the epidermal cells of tobacco leaves by Agrobacterium infiltration. Green fluorescent BnWRI1-GFP was visualized by confocal laser scanning microscopy (Figure 5A), and the resulting image was overlaid with a nucleus marked by 4′,6-diamidine-2-phenylindole dihydrochloride (DAPI) staining, confirming that the introduced BnWRI1 is localized to the nucleus (Figures 5B–D). The proximal upstream regions of Arabidopsis genes such as pyruvate kinase (PKp), BCCP2, KASI, LPAT2, and GPAT9 that are involved in glycolysis, FA synthesis, and lipid assembly contain the AW-box featured with [CnTnG](n)7[CG] (Figure 6A). To test whether BnWRI1 binds to the target DNA fragments containing the AW-box, the BnWRI1 protein was expressed in E. coli and purified for binding assays (Figure 6C). The DNA fragments of 250–300 bp that contained the AW-box [CnTnG](n)7[CG] in the promoter region of two representative genes known as KASI and GPAT9 were amplified and used for EMSA. When BnWRI1 protein was incubated with the DNA fragment amplified from the KASI promoter containing the AW-box [CnTnG](n)7[CG], the target DNA was bound to BnWRI1, as indicated by the shifted bands at the top of gel when it was visualized under UV light (Figure 6D). However, when the DNA fragment with a mutant AW box [TnCnA](n)7[CG] was explored, the binding between the BnWRI1 and the DNA fragment was abolished, as shown by the observation that the DNA band position remaining unchanged in the gel (Figures 6B,D). A similar gel shift was observed when BnWRI1 protein was incubated with the DNA fragment containing an AW box [CnTnG](n)7[CG] from the promoter region of GAPT9 (Figure 6D). The binding was diminished when the consensus was mutated to [CnTnG](n)7[TA] for the GPAT9 promoter (Figures 6B,D). A similar result was found when the DNA probes were labeled with biotin (Figure 6E). These results suggest that BnWRI1 specifically binds to the promoter region containing the AW-box, which is conserved with Arabidopsis WRI1, and the AW-box is essential for the interaction between BnWRI1 and the target gene promoters.
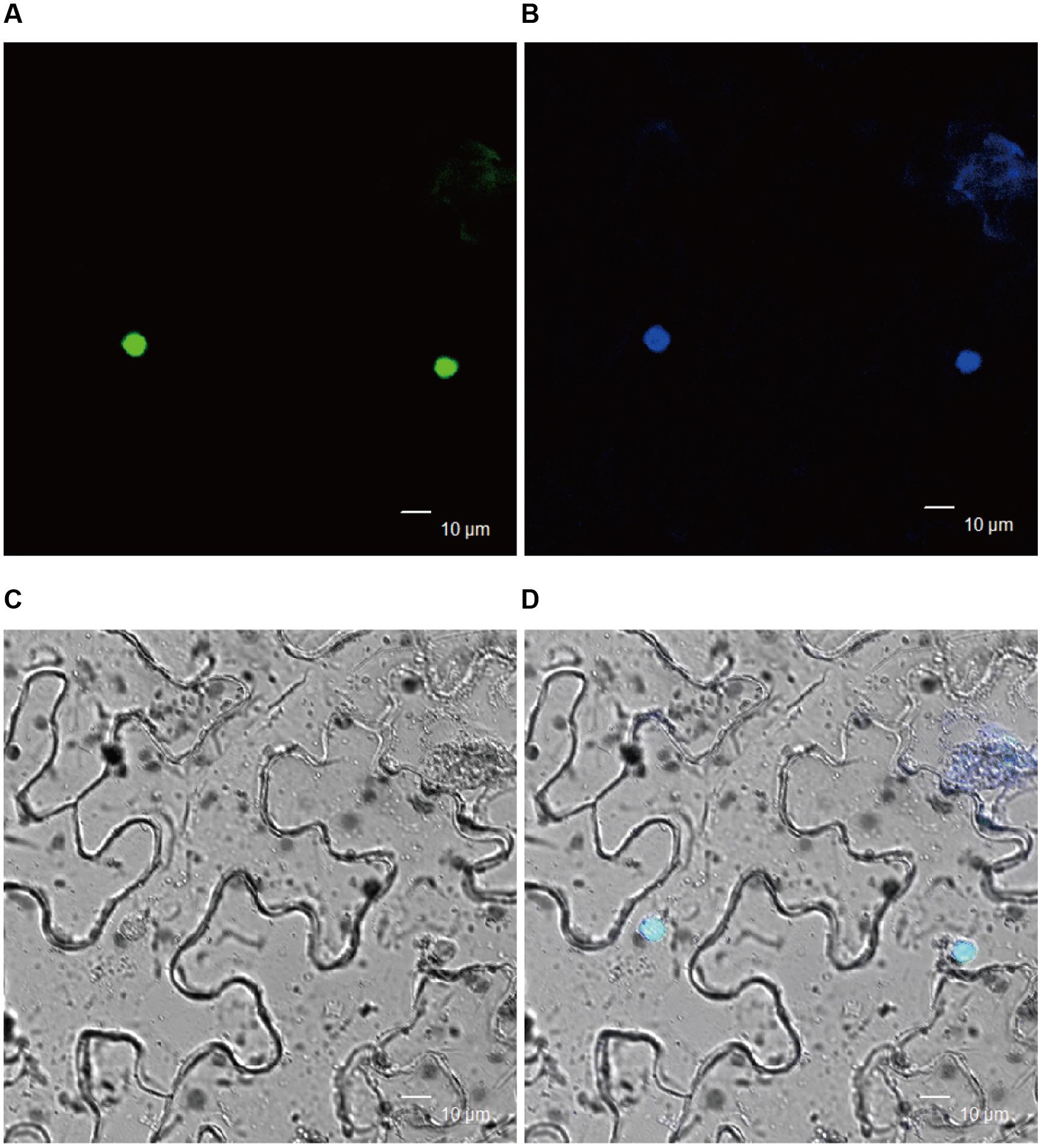
FIGURE 5. Nuclear localization of BnWRI1. (A) Green fluorescence of BnWRI1-GFP. (B) Nucleus stained with DAPI. (C) Bright image. (D) Merged image of the same cells observed in (A). BnWRI1-GFP was transiently expressed in Nicotiana benthamiana epidermal cells and visualized under by confocal laser scanning microscopy. The nuclear localization of BnWRI1 was indicated by green fluorescence that was overlaid with the nucleus indicated by DAPI staining.
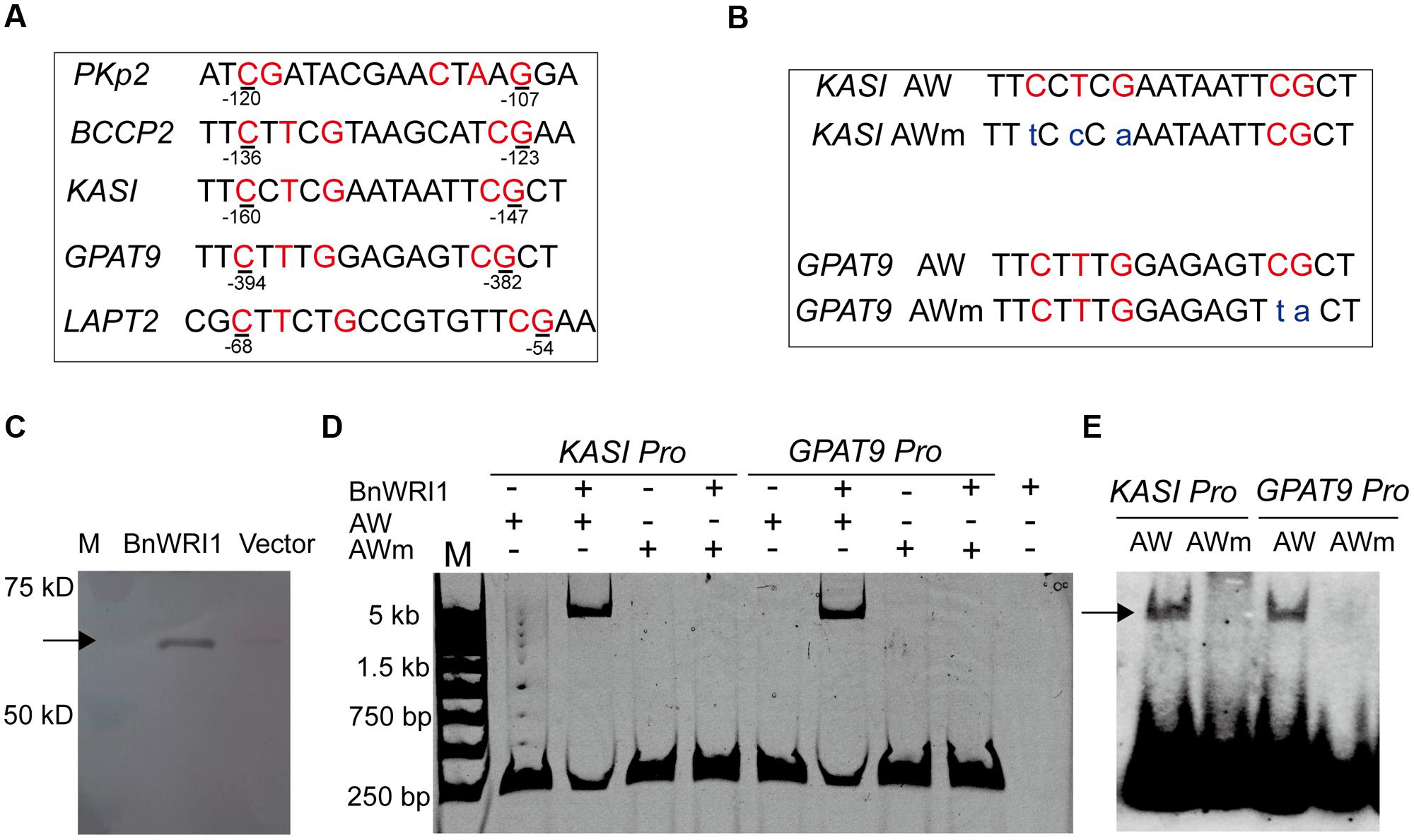
FIGURE 6. BnWRI1 binding to proximal upstream regions of genes involved in lipid anabolism. (A) The AW-box [CnTnG](n)7[CG] in the proximal upstream regions of genes involved in glycolysis, FA synthesis, and lipid assembly. (B) Consensus [CnTnG](n)7[CG] of the AW-box and its mutant AW-box [TnCnA](n)7[CG] of the KASI promoter or [CnTnG](n)7 [TA] of GPAT9 promoter. (C) Recombinant BnWRI1 protein (50 kD) expressed in E. coli cells. (D) BnWRI1 specifically binds to the promoter region of KASI and GPAT9 containing the AW-box as assayed by EMSA. The binding was abolished when the conserved sequence was mutated into the [TnCnA](n)7[CG] of the KASI promoter or the [CnTnG](n)7[TA] of the GPAT9 promoter. The purified BnWRI1 protein (0.4 μg) was incubated with the target DNA fragment for 1 h. The resulting complex was visualized under UV light. The lanes from left to right: M, DNA ladder; KASI promoter only; BnWRI1 (0.4 μg) + KASI promoter; KASI mutant promoter only; BnWRI1 (0.4 μg) + KASI mutant promoter; GPAT9 promoter only; BnWRI1 (0.4 μg) + GPAT9 promoter; GPAT9 mutant promoter only; BnWRI1 (0.4 μg) + GPAT9 mutant promoter; BnWRI1 only. (E) The binding activity of BnWRI1 to the AW-box or its mutant AW-box (mAW) in the promoter regions of KASI and GPAT9 by EMSA using the DNA probes labeled with biotin. The shifted band is indicated by the arrow.
Overexpression of BnWRI1 Up-regulates the Transcript Level of Genes in Glycolysis, Fatty Acid Synthesis, Lipid Assembly, and Flowering
To investigate whether BnWRI1 OE up-regulated the transcript level of genes involved in the lipid anabolism process, RNA was extracted from the leaves and analyzed by quantitative real-time PCR. These genes include PKp2 in glycolysis, BCCP2, MAT, KASI, ENR1, and acyl-ACP thioesterase (FATA) in the FA biosynthetic process, and GPAT9, LPAT2, and DGAT1 in lipid assembly. The transcript levels of the tested genes were all significantly higher in OE plants than in the WT. Despite the significant elevation of genes in multiple pathways by BnWRI1, the regulation of BnWRI1 in specific routes differs to variable extents. The transcript level of genes involved in glycolysis and FA synthesis including PKp2, MAT, KASI, ENR1, and FATA was up-regulated the most, and the expression levels in OE plants were more than twofold that of the WT (Figures 7A,B). The BnWRI1 enhanced expression of genes involved in FA synthesis was most prominent among the tested genes (Figure 7B). Moreover, RNA accumulation for the genes involved in lipid assembly was also strongly induced in OE plants. The mRNA level of GPAT9 in OE plants accumulated more than twofold that of the WT (Figure 7C). The LPAT2 and DGAT1 transcript levels were also substantially higher in OE than in WT plants (Figures 7C,D). In addition, the FLOWERING LOCUS T (FT) is a key regulator in the control of flowering time in several plant species, and the FT expression level in OE plants was threefold higher than that of the WT (Figure 7E). The results suggest that BnWRI1 synchronously promotes multiple pathways in transcriptional regulation to enhance the plant reproductive process, seed development, and oil accumulation in B. napus.
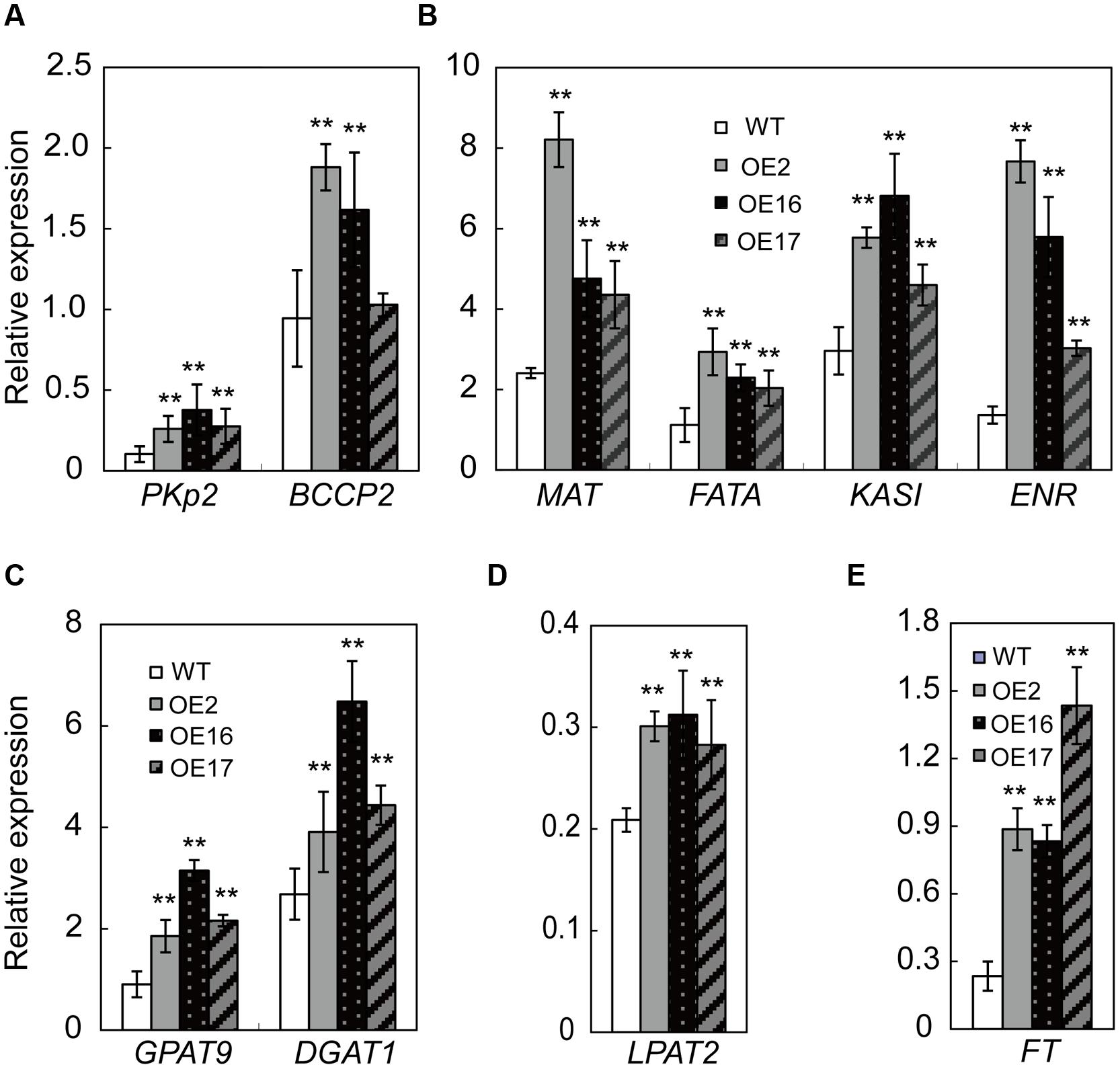
FIGURE 7. Transcript levels of genes involved in lipid anabolism in BnWRI1-OE and WT plants. (A,B) The expression levels of genes involved in glycolysis and FA synthesis. (C,D) The expression levels of genes involved in lipid assembly. (E) The expression level of FT involved in flowering. Total RNA was extracted from the leaves of 7-weeks-old plants and the transcript levels of genes were analyzed by quantitative real-time PCR. PKp2, pyruvate kinase, homolog At5g52920; BCCP2, biotin carboxyl carrier protein, homolog At5g16390; MAT, malonyl-CoA:ACP malonyltransferase, homolog At2g30200; KASI, ketoacyl-ACP synthases, homolog At5g46290; ENR1, enoyl-ACP reductase, homolog At2g05990; FATA, acyl-ACP thioesterase, homolog At3g25110; GPAT9, glycerol-3-P acyltransferase, homolog At5g60620; LPAT2, lysophosphatidic acid acyltransferase, homolog At3g57650; and DGAT1, DAG acyltransferase, homolog At2g19450. The expression level was normalized to that of BnActin. Values are mean ± SD (n = 3); ∗∗P < 0.01.
The Effect of BnWRI1 on the Membrane Lipid Composition
Most studies have focused on the effect of WRI1 on oil accumulation, but the effect of WRI1 on the membrane lipid composition remains elusive. The OE of BnWRI1 in B. napus leads to the involvement of numerous genes in glycolysis, FA biosynthesis, and the lipid assembly process, indicating its significance in lipid anabolism. To investigate the effect of BnWRI1 on various lipid metabolisms further, phospholipids and galactolipids from leaves or siliques during flowering stages were analyzed. The phosphatidylethanol (PE) and phosphatidylglycerol (PG) in leaves remained comparable between OE and WT plants (Figure 8A). However, BnWRI1 OE resulted in a significant increase in MGDG, DGDG, and PC in leaves relative to their levels in WT plants (Figure 8A). The increased DGDG resulted from the increase in 16:0, 18:1, 18:2, and 18:3 FA species, and the elevated MGDG primarily came from an increased 16:0 FA, whereas the enhanced PC level was mostly contributed by increased 16:0, 16:1, and 18:2 FAs (Figures 8B–D).
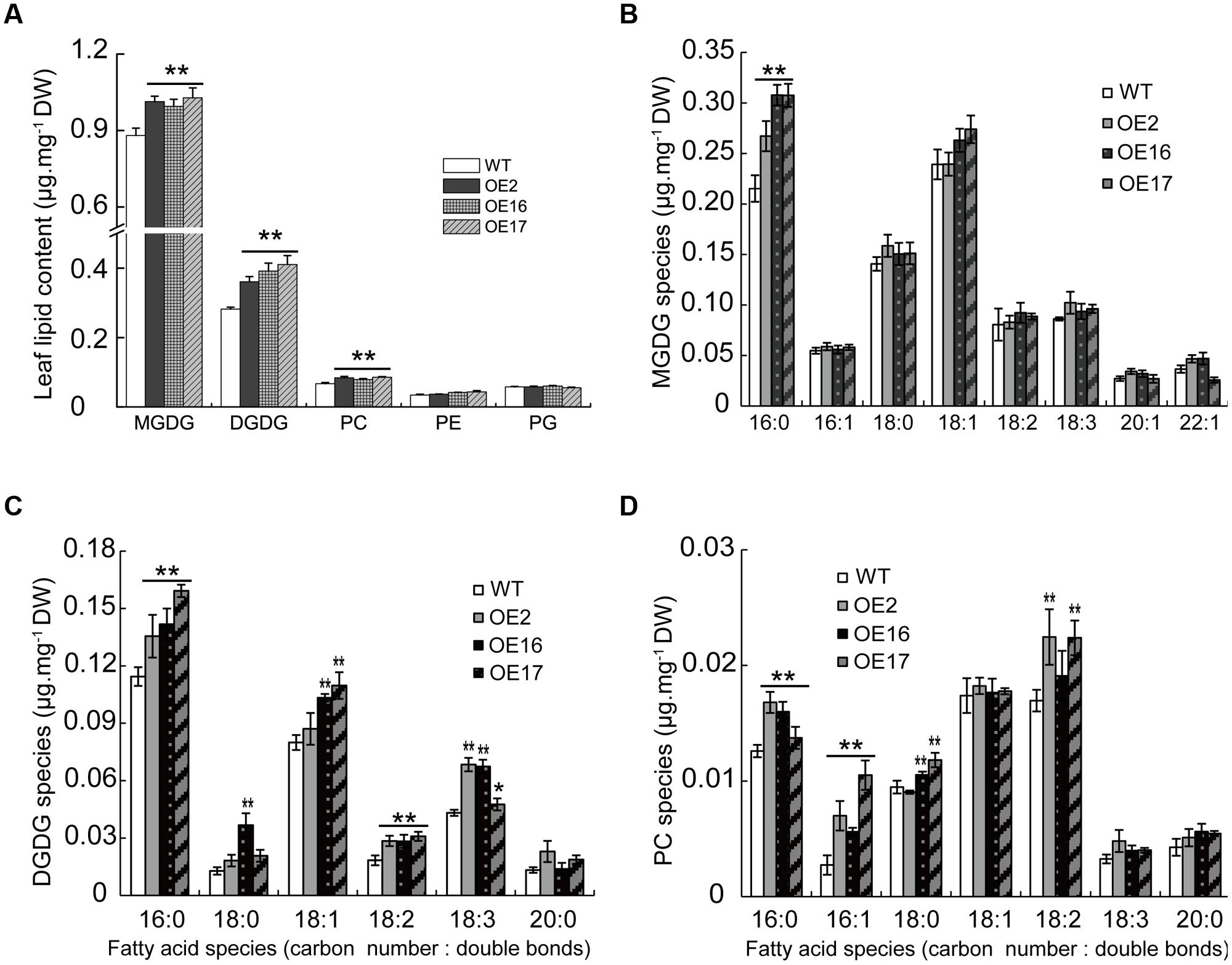
FIGURE 8. Effect of BnWRI1-OE on membrane lipid compositions in leaves. (A) Lipid compositions in the leaves of BnWRI1-OE and WT plants. Fatty acid species of MGDG (B), DGDG (C), and PC (D) in the leaves. Lipids were extracted from 5-month-old plants during the early flowering stage. The extracted lipids were separated by TLC plates and the spot corresponding to a specific lipid was quantitatively analyzed by GC measurement. Values are mean ± SE (n = 4); ∗P < 0.05, ∗∗P < 0.01.
Lipid profiling from developing siliques revealed that the lipid content and composition of siliques were significantly different from those of leaves. The contents of galactolipids such as DGDG and MGDG in siliques, the predominant components of chloroplast membranes, were much lower than the contents in leaves (Figures 8 and 9), suggesting that photosynthesis predominantly occurs in leaves rather than siliques during the flowering and early seed development stages. BnWRI1 OE resulted in increased phospholipids and a corresponding reduction in galactolipids (Figure 9A). OE siliques contain less DGDG with decreased 16:0, 18:2 FAs and increased 20:1 FA in OE siliques (Figure 9C). MGDG was also reduced with decreased 16:0, 22:1 FA and increased 18:3 FA in OE siliques, and the magnitude was smaller than that of DGDG (Figures 9A–C). By comparison, phospholipids PC, PE, and PG in OE siliques were higher than the levels in the WT (Figure 9A). The increased PC and PE in OE siliques were contributed mostly by increased 16:0 FA, whereas elevated PG resulted from increased 16:0 and 18:0 FAs in OE siliques. (Figures 9D–F). The TAG in OE siliques was significantly higher than that of the WT, and increased TAG comes primarily from elevated 16:0 FA. The 18:2 and 18:3 FA contents in both OE2 and OE17 were also higher than those in WT siliques (Figures 9G,H).
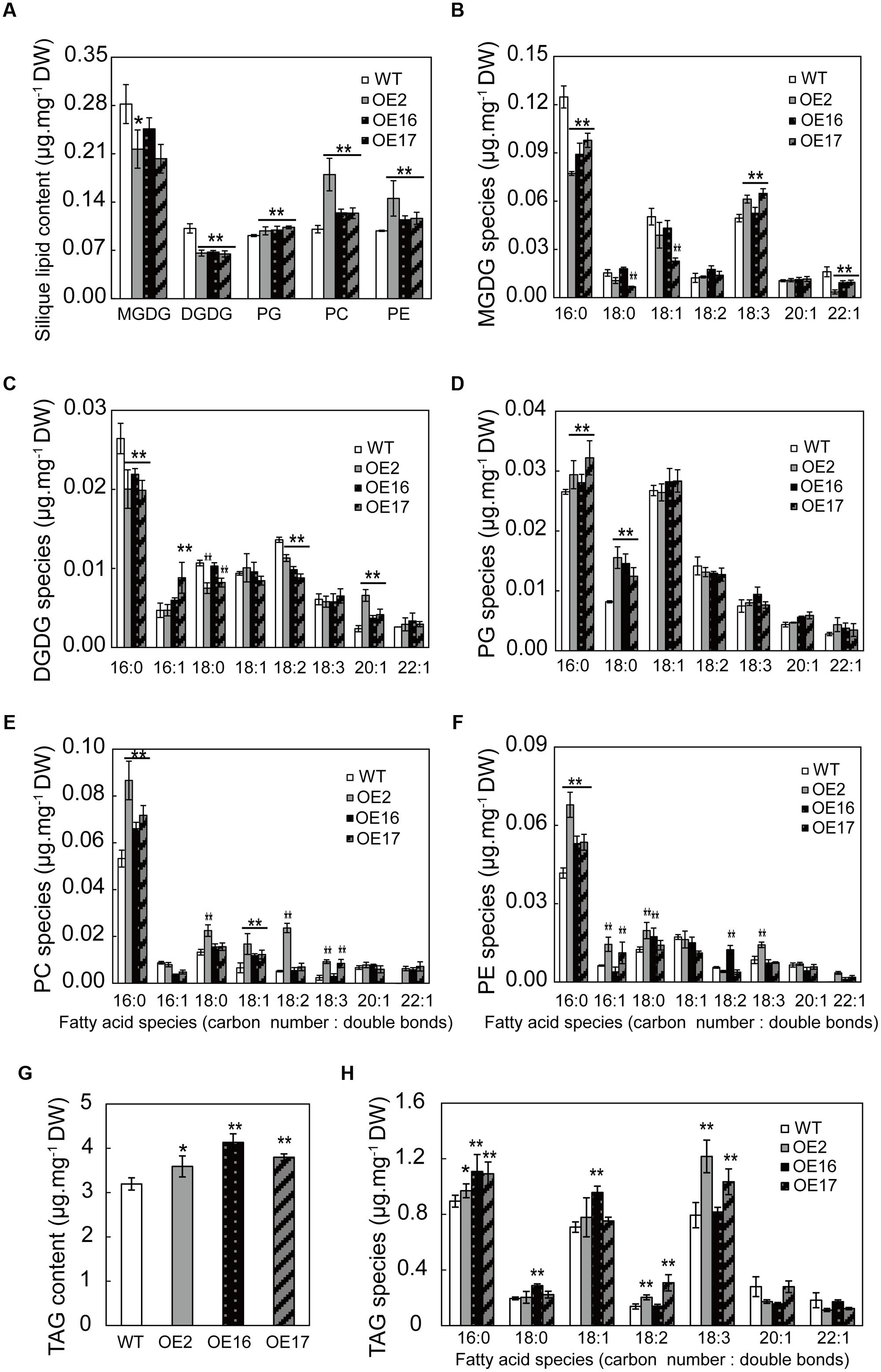
FIGURE 9. Effect of BnWRI1-OE on membrane lipids and TAG levels in siliques. (A) Lipid composition in the siliques of BnWRI1-OE and WT plants. Lipids were extracted from the siliques at 20 days after anthesis (DAA). The extracted lipids were separated by TLC and the spot corresponding to a specific lipid was quantitatively analyzed by GC. (B–F) FA species of MGDG, DGDG, PG, PC, and PE in the siliques of 20 DAA. Oil (TAG) content (G) and FA species of TAG (H) in the siliques of 20 DAA. Values are mean ± SE (n = 4); ∗P < 0.05, ∗∗P < 0.01.
Discussion
Fatty acids are major, basic components of lipid assembly, and their destinations include membrane lipids, storage lipids, lipid messengers, and other derivatives. The FA synthesis enzyme complex consists of multiple subunits including malonyl-CoA: ACP transferase (MAT), acyl-ACP transferase (AT), β-ketoacyl-ACP synthase (KAS), β-ketoacyl-ACP reductase (KAR), hydroxyacyl-ACP dehydrogenase (HAD), and enoyl-ACP reductase (ENR) as encoded by individual genes in plants (Slabas and Fawcett, 1992; Ohlrogge and Browse, 1995; Ohlrogge and Jaworski, 1997). WRI1 is a central regulator that modulates numerous genes and multiple steps in oil synthesis simultaneously, and it is unique to plants (Cernac and Benning, 2004; Shen et al., 2010). However, most studies are focused on WRI1 involvement in oil accumulation in Arabidopsis. The effect of WRI1 on lipid synthesis and its consequence on plant development were unknown, especially in crop plants. This study unraveled the novel roles of BnWRI1 in flowering time control and lipid homeostasis in relation to oil accumulation, membrane phospholipids, galactolipids, and sugars. Enhanced lipid anabolism and accelerated flowering from BnWRI1 OE did not result in the inhibition of protein synthesis and total sugar accumulation. Unlike the results found in Arabidopsis (Cernac and Benning, 2004), BnWRI1 OE in B. napus plants did not cause visible undesirable growth or development traits. These results suggest that the roles of BnWRI1 in B. napus is conserved and yet distinguishable from that of other species such as Arabidopsis and maize (Focks and Benning, 1998; Baud et al., 2007; Pouvreau et al., 2011; To et al., 2012).
A previous study showed that the AP2-type transcription factors play important roles in regulating meristem growth and organ development. AP2-type transcription factors are also involved in ovule development, floral organ growth, and seed size (Jofuku et al., 1994; Elliott et al., 1996; Okamuro et al., 1997; Mizukami and Fischer, 2000; Ohto et al., 2005). WRI1 belongs to the AP2-type transcription factor family, but its role in the flowering time had not been reported. Our result showed that BnWRI1-OE plants flowered 4–6 days earlier than the WT, suggesting that BnWRI1 plays a role in flowering time control. WRI1 from Arabidopsis or maize is not involved in flowering time control (Cernac and Benning, 2004; Pouvreau et al., 2011). Early flowering is an important trait because seeds can be matured within a given time to make field space available for the next crop’s growth. In addition to its role in oil accumulation, a mild earlier flowering by BnWRI1-OE did not lead to a visible inhibition in vegetative growth and biomass, which provides a possibility for oil crop breeding. Gene expression profiling showed that BnWRI1-OE plants enhanced FT expression, suggesting that BnWRI1 modulates flowering in a transcriptional manner. However, the promoter region of FT does not contain a typical AW-box, suggesting that other cis elements in the FT promoter are recognized and regulated by BnWRI1. Alternatively, the early flowering may have resulted from PC elevation as shown by the increased PC level in OE plants. Recent studies showed that PC plays a positive role in Arabidopsis flowering (Nakamura et al., 2014; Wang et al., 2015).
An oil component in the form of TAG is primarily derived from sugars through photosynthesis in plants. In oil seed plants such as B. napus and Arabidopsis, starch was accumulated at the early phase of seed development and converted to TAG and proteins at the later phase of seed maturation (Baud et al., 2002; Ruuska et al., 2002; Hills, 2004). Our study showed that BnWRI1 OE in B. napus resulted in significant oil accumulation in both seeds and leaves, which was accompanied by up-regulated genes involved in glycolysis, FA biosynthesis, and lipid assembly, whereas the AtWRI1 in Arabidopsis was not involved in lipid assembly (Cernac and Benning, 2004; Baud et al., 2007; To et al., 2012). The seed-specific OE of BnWRI1 in B. napus or in Arabidopsis resulted in enhanced seed oil content and seed size (Liu et al., 2010; Wu et al., 2014). The OE of Arabidopsis WRI1 in Camelinasativa also led to increased oil content and seed size (An and Suh, 2015). AtWRI1 binds to the AW-box [CnTnG](n)7[CG] located at the proximal upstream regions of genes involved in glycolysis and FA synthesis (Maeo et al., 2009). BnWRI1 contains two AP2 domains involved in DNA binding. Our results showed that BnWRI1 was localized to the nucleus. An EMSA assay showed that BnWRI1 was capable of binding to the AW-box [CnTnG](n)7[CG] at the proximal promoter region of genes involved in FA synthesis and lipid assembly, and the AW-box is essential for binding because the nucleotide substitutive mutant abolished the binding. Moreover, lipid assembly is also important for oil synthesis as demonstrated by overexpressing the genes encoding yeast glycerol-3-phosphate dehydrogenase and yeast LPAT in B. napus leading to a modest increase in oil content (Zou et al., 1997; Vigeolas et al., 2007). Likewise, an elevated DGAT transcript level in soybeans and maize resulted in enhanced seed oil content (Zheng et al., 2008). The current study showed that BnWRI1 also binds to the promoter region of GPAT9 that is responsible for lipid assembly, which is not found in Arabidopsis and other plant species (Baud et al., 2007; To et al., 2012). These observations suggest that the transcriptional regulation of BnWRI1 and its effect on the lipid anabolic process is more comprehensive in B. napus than in other plant species.
As fossil energy becomes limited, it is attractive to enhance oil accumulation in vegetative tissues to supply renewable biofuel and feed stock for industry and animal foods. The current results showed that BnWRI1-OE in B. napus led to increased oil contents in the leaves without growth retardation, suggesting a potential application in crop plant breeding. However, the increased amount of leaf oil content is moderate at 1.4% of the dry weight, which is similar to that of tobacco leaves that transiently expressed WRI1 from other plant species (Grimberg et al., 2015). A transcriptional analysis showed that WRI1 up-regulated genes in both FA synthesis and degradation, indicating a futile cycle of FA metabolism, which may be responsible for the limitation in leaf oil accumulation (Grimberg et al., 2015). The results suggest that achieving high oil contents in the leaves through single gene manipulation could be challenging. A recent study showed that the co-expression of three genes, WRI1, DGAT and oleosin, led to 15% TAG dry weight in tobacco leaves (Vanhercke et al., 2014).
Carbon allocation by WRI1 occurred in Arabidopsis and cotton plants. The constitutive expression of AtWRI1 in Arabidopsis led to a significant increase in the oil content, which was accompanied by defective seed germination and plant growth (Cernac and Benning, 2004). Suppressing WRI1 in cotton led to increased fiber length and reduced seed oil content (Qu et al., 2012). Our results showed that the total sugars and proteins remained constant between OE and WT, but OE plants displayed reduced carbohydrate storage with a corresponding increase in soluble sugars in the leaves and seeds. In addition to oil accumulation, BnWRI1 OE also increased phospholipids and galactolipids significantly in leaves. Higher levels in BnWRI1-OE leaves of the galactolipids MGDG and DGDG, the important components of chloroplast thylakoid membranes that are essential for photosynthesis, may be responsible for enhanced source capacity. A recent study showed that the seed-specific expression of BnWRI1 promoted the expression of the photosynthesis gene (Wu et al., 2014). The results suggest that the enhanced lipid anabolic process in leaves by BnWRI1 is not a result of a reduction in other organic components but a consequence of enhanced source/sink strength by enhanced galactolipids and carbon flux to lipid accumulation. This finding may explain, at least in part, how BnWRI1-OE enhances oil accumulation and flowering without growth inhibition in B. napus. However, the lipid metabolism regulated by BnWRI1 in siliques is quite different from that of leaves. A lipid analysis of the siliques revealed reduced galactolipids MGDG and DGDG, and corresponding increases in the phospholipids PC, PE, PG, and storage lipid TAG in OE siliques during flowering and early seed developing stages. These results indicate that enhanced PC and PE may facilitate lipid mobilization to oil accumulation in seeds because TAG can be directly derived from PC by PDAT activation (Dahlqvist et al., 2000; Fan et al., 2013, 2014). Our data support the facts that BnWRI1 regulates homeostasis among sugars, membrane lipids, and storage lipids, and its consequences for enhancing reproduction, seed development and oil accumulation. Our results shed light on the roles of BnWRI1 in lipid regulatory networks and its application in oil crop plant breeding.
In summary, the results from this study identified several novel roles of BnWRI1 in B. napus that have not been characterized before. First, the OE of BnWRI1 led to accelerated plant flowering 4–6 days earlier without reduced vegetative growth, which is a good agronomic trait to open up field space for the next crop’s planting. Second, BnWRI1 enhanced oil accumulation in both seeds and leaves without visible side effects on growth. BnWRI1 decreased storage carbohydrates and increased soluble sugars to facilitate carbon flux to lipid anabolism. Third, BnWRI1 is localized in the nucleus and binds to the AW-box at the proximal promoter region of genes involved in FA synthesis as well as in the lipid assembly process that is not found in Arabidopsis. The OE of BnWRI1 led to the up-regulated transcription of genes in the lipid anabolic pathway, and of FT in flowering control. Finally, BnWRI1 not only enhances oil accumulation, but it also affects membrane lipid metabolism and turnover in both leaves and developing siliques. BnWRI1 OE caused increased galactolipid MGDG, DGDG, and phospholipid PC in leaves, but it led to reduced DGDG and MGDG and increased PC, PE, and TAG in siliques during the early seed development stage. Enhanced galactolipid synthesis by BnWRI1 is beneficial for photosynthesis to enhance the source/sink capacity, and thus facilitates the sugar flux to oil accumulation without the expense of sugar and protein in both leaves and seeds. Therefore, BnWRI1 plays a positive role in homeostasis among sugars, membrane lipids, and oil accumulation by modulating the coordination of multiple metabolism pathways, thus enhancing flowering and oil accumulation without growth inhibition. Based on the results from this study, we proposed a work model for BnWRI1 in lipid anabolic process, plant growth and development (Figure 10). The early flowering led by BnWRI1 may be caused by its transcriptional regulation through binding to the promoter region of FT or other flowering-related genes. It would be interesting to identify new targets of BnWRI1 that are involved in flowering control in future studies. In addition, BnWRI1 OE led to an elevated PC level in both leaves and siliques. PC functions not only as an essential component of the membrane structure, but it also plays important roles in flowering regulation and providing acyl chains for TAG accumulation (Dahlqvist et al., 2000; Zhang et al., 2009; Nakamura et al., 2014; Wang et al., 2015). It would be interesting to explore how PC synthesis is regulated by BnWRI1 in future work.
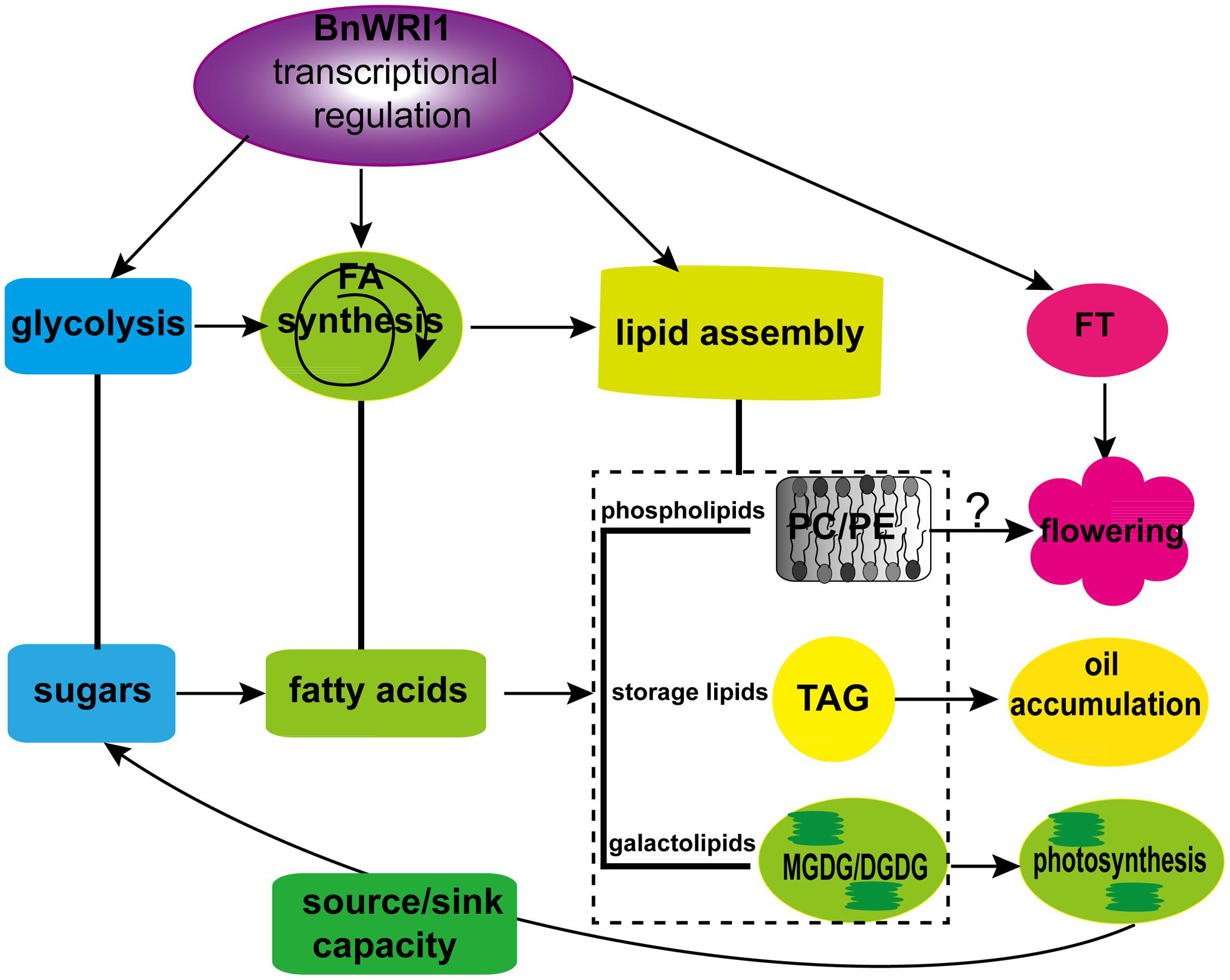
FIGURE 10. A proposed model for BnWRI1 in regulating lipid anabolism and flowering in B. napus. BnWRI1 up-regulates transcript levels of genes involved in glycolysis, FA synthesis, and lipid assembly to enhance accumulation of oil (TAG), galactolipids, and phospholipids. The increased MGDG and DGDG by BnWRI1 promote source/sink capacity, thus BnWRI1 plays a positive role in homeostasis among sugars, TAG, and membrane lipids in B. napus. Moreover, BnWRI1 also enhances FT expression, and PC level to accelerate flowering.
Author Contributions
YH, QL, and JS designed the study. QL and JS performed most of the experiments; ST helped cloning and transformation; QS performed EMSA; TW analyzed some of metabolites; WC helped isolation and identification of transgenic plants.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Professor Xuemin Wang for the critical reading for this manuscript, and Dr. Shaoping Lu for assistance in canola transformation in this study. This work was supported by grants from the Chinese National Key Basic Research Project (2015CB150204), and the National Science Foundation of China (31271514).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2015.01015
References
Alonso, A. P., Goffman, F. D., Ohlrogge, J. B., and Shachar-Hill, Y. (2007). Carbon conversion efficiency and central metabolic fluxes in developing sunflower (Helianthus annuus L.) embryos. Plant J. 52, 296–308. doi: 10.1111/j.1365-313X.2007.03235.x
An, D., and Suh, M. C. (2015). Overexpression of Arabidopsis WRI1 enhanced seed mass and storage oil content in Camelina sativa. Plant Biotechnol. Rep. 9, 137–148. doi: 10.1007/s11816-015-0351-x
Barthole, G., Lepiniec, L., Rogowsky, P. M., and Baud, S. (2012). Controlling lipid accumulation in cereal grains. Plant Sci. 18, 33–39. doi: 10.1016/j.plantsci.2011.09.002
Baud, S., Boutin, J. P., Miquel, M., Lepiniec, L., and Rochat, C. (2002). An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol. Biochem. 40, 151–160. doi: 10.1016/S0981-9428(01)01350-X
Baud, S., and Lepiniec, L. (2009). Regulation of de novo fatty acid synthesis in maturing oilseeds of Arabidopsis. Plant Physiol. Biochem. 47, 448–455. doi: 10.1016/j.plaphy.2008.12.006
Baud, S., Mendoza, M. S., To, A., Harscoët, E., Lepiniec, L., and Dubreucq, B. (2007). WRINKLED1 specifies the regulatory action of LEAFY COTYLEDON2 towards fatty acid metabolism during seed maturation in Arabidopsis. Plant J. 50, 825–838. doi: 10.1111/j.1365-313X.2007.03092.x
Cahoon, E. B., Shockey, J. M., Dietrich, C. R., Gidda, S. K., Mullen, R. T., and Dyer, J. M. (2007). Engineering oilseeds for sustainable production of industrial and nutritional feedstocks: solving bottlenecks in fatty acid flux. Curr. Opin. Plant Biol. 10, 236–244. doi: 10.1016/j.pbi.2007.04.005
Cernac, A., Andre, C., Hoffmann-Benning, S., and Benning, C. (2006). WRI1 is required for seed germination and seedling establishment. Plant Physiol. 141, 745–757. doi: 10.1104/pp.106.079574
Cernac, A., and Benning, C. (2004). WRINKLED1 encodes an AP2/EREB domain protein involved in the control of storage compound biosynthesis in Arabidopsis. Plant J. 40, 575–585. doi: 10.1111/j.1365-313X.2004.02235.x
Chow, P., and Landhausser, S. (2004). A method for routine measurements of total sugar and starch content in woody plant tissues. Tree Physiol. 24, 1129–1136. doi: 10.1093/treephys/24.10.1129
Dahlqvist, A., Stahl, U., Lenman, M., Banas, A., Le, M., Sandager, L., et al. (2000). Phospholipid: diacylglycerol acyltransferase: an enzyme that catalyzes the acyl-CoA-independent formation of triacylglycerol in yeast and plants. Proc. Natl. Acad. Sci. U.S.A. 97, 6487–6492. doi: 10.1073/pnas.120067297
DuBois, M., Gilles, K., Hamilton, J., Rebers, P., and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356. doi: 10.1021/ac60111a017
Elliott, R. C., Betzner, A. S., Huttner, E., Oakes, M. P., Tucker, W. Q., Gerentes, D., et al. (1996). AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8, 155–168. doi: 10.1105/tpc.8.2.155
Fan, J., Yan, C., Roston, R., Shanklin, J., and Xu, C. (2014). Arabidopsis lipins, PDAT1 acyltransferase, and SDP1 triacylglycerol lipase synergistically direct fatty acids toward β-oxidation, thereby maintaining membrane lipid homeostasis. Plant Cell 26, 4119–4134. doi: 10.1105/tpc.114.130377
Fan, J., Yan, C., Zhang, X., and Xu, C. (2013). Dual role for phospholipid: diacylglycerol acyltransferase: enhancing fatty acid synthesis and diverting fatty acids from membrane lipids to triacylglycerol in Arabidopsis leaves. Plant Cell 25, 3506–3518. doi: 10.1105/tpc.113.117358
Focks, N., and Benning, C. (1998). wrinkled1: a novel, low-seed-oil mutant of Arabidopsis with a deficiency in the seed-specific regulation of carbohydrate metabolism. Plant Physiol. 118, 91–101. doi: 10.1104/pp.118.1.91
Grimberg,Å, Carlsson, A. S., Marttila, S., Bhalerao, R., and Hofvander, P. (2015). Transcriptional transitions in Nicotiana benthamiana leaves upon induction of oil synthesis by WRINKLED1 homologs from diverse species and tissues. BMC Plant Biol. 15:192. doi: 10.1186/s12870-015-0579-1
Hills, M. J. (2004). Control of storage-product synthesis in seeds. Curr. Opin. Plant Biol. 7, 302–308. doi: 10.1016/j.pbi.2004.03.003
Hong, Y., Devaiah, S. P., Bahn, S. C., Thamasandra, B. N., Li, M., Welti, R., et al. (2009). Phospholipase D and phosphatidic acid enhance Arabidopsis nitrogen signaling and growth. Plant J. 58, 376–387. doi: 10.1111/j.1365-313X.2009.03788.x
Hong, Y., Pan, X., Welti, R., and Wang, X. (2008). Phospholipase D(3 is involved in the hyperosmotic response in Arabidopsis. Plant Cell 20, 803–816. doi: 10.1105/tpc.107.056390
Jako, C., Kumar, A., Wei, Y., Zou, J., Barton, D. L., Giblin, E. M., et al. (2001). Seed-specific over-expression of an Arabidopsis cDNA encoding a diacylglycerol acyltransferase enhances seed oil content and seed weight. Plant Physiol. 126, 861–874. doi: 10.1104/pp.126.2.861
Jofuku, K. D., den Boer, B. G., Van Montagu, M., and Okamuro, J. K. (1994). Control of Arabidopsis flower and seed development by the homeotic gene APETALA2. Plant Cell 6, 1211–1225. doi: 10.1105/tpc.6.9.1211
Kang, F., and Rawsthorne, S. (1996). Metabolism of glucose-6- phosphate and utilization of multiple metabolites for fatty acid synthesis by plastids from developing oilseed rape embryos. Planta 199, 321–327. doi: 10.1007/BF00196575
Kwong, R. W., Bui, A. Q., Lee, H., Kwong, L. W., Fischer, R. L., Goldberg, R. B., et al. (2003). LEAFY COTYLEDON1-LIKE defines a class of regulators essential for embryo development. Plant Cell 15, 5–18. doi: 10.1105/tpc.006973
Li-Beisson, Y., Shorrosh, B., Beisson, F., Andersson, M. X., Arondel, V., Bates, P. D., et al. (2010). Acyl-lipid metabolism. Arabidopsis Book 8:e0133. doi: 10.1199/tab.0133
Liu, J., Hua, W., Zhan, G., Wei, F., Wang, X., Liu, G., et al. (2010). Increasing seed mass and oil content in transgenic Arabidopsis by the overexpression of wri1-like gene from Brassica napus. Plant Physiol. Biochem. 48, 9–15. doi: 10.1016/j.plaphy.2009.09.007
Lotan, T., Ohto, M., Yee, K. M., West, M. A., Lo, R., Kwong, R. W., et al. (1998). Arabidopsis LEAFY COTYLEDON1 is sufficient to induce embryo development in vegetative cells. Cell 93, 1195–1205. doi: 10.1016/S0092-8674(00)81463-4
Maeo, K., Tokuda, T., Ayame, A., Mitsui, N., Kawai, T., Tsukagoshi, H., et al. (2009). An AP2-type transcription factor, WRINKLED1, of Arabidopsis thaliana binds to the AW-box sequence conserved among proximal upstream regions of genes involved in fatty acid synthesis. Plant J. 60, 476–487. doi: 10.1111/j.1365-313X.2009.03967.x
Mizukami, Y., and Fischer, R. L. (2000). Plant organ size control: AINTEGUMENTA regulates growth and cell numbers during organogenesis. Proc. Natl. Acad. Sci. U.S.A. 97, 942–947. doi: 10.1073/pnas.97.2.942
Mu, J., Tan, H., Zheng, Q., Fu, F., Liang, Y., Zhang, J., et al. (2008). LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis. Plant Physiol. 148, 1042–1054. doi: 10.1104/pp.108.126342
Nakamura, Y., Andrés, F., Kanehara, K., Liu, Y. C., Dörmann, P., and Coupland, G. (2014). Arabidopsis florigen FT binds to diurnally oscillating phospholipids that accelerate flowering. Nat. Commun. 5, 3553. doi: 10.1038/ncomms4553
Ohlrogge, J., and Browse, J. (1995). Lipid biosynthesis. Plant Cell 7, 957–970. doi: 10.1105/tpc.7.7.957
Ohlrogge, J. B., and Jaworski, J. G. (1997). Regulation of fatty acid synthesis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 109–136. doi: 10.1146/annurev.arplant.48.1.109
Ohto, M. A., Fischer, R. L., Goldberg, R. B., Nakamura, K., and Harada, J. J. (2005). Control of seed mass by APETALA2. Proc. Natl. Acad. Sci. U.S.A. 102, 3123–3128. doi: 10.1073/pnas.0409858102
Okamuro, J. K., Szeto, W., Lotys-Prass, C., and Jofuku, K. D. (1997). Photo and hormonal control of meristem identity in the Arabidopsis flower mutants apetala2 and apetala1. Plant Cell 9, 37–47. doi: 10.1105/tpc.9.1.37
Phillips, R., Ursell, T., Wiggins, P., and Sens, P. (2009). Emerging roles for lipids in shaping membrane-protein function. Nature 459, 379–385. doi: 10.1038/nature08147
Pouvreau, B., Baud, S., Vernoud, V., Morin, V., Py, C., Gendrot, G., et al. (2011). Duplicate maize Wrinkled1 transcription factors activate target genes involved in seed oil biosynthesis. Plant Physiol. 156, 674–686. doi: 10.1104/pp.111.173641
Qu, J., Ye, J., Geng, Y. F., Sun, Y. W., Gao, S. Q., Zhang, B. P., et al. (2012). Dissecting functions of KATANIN and WRINKLED1 in cotton fiber development by virus-induced gene silencing. Plant Physiol. 160, 738–748. doi: 10.1104/pp.112.198564
Ruuska, S., Girke, T., Benning, C., and Ohlrogge, J. (2002). Contrapuntal networks of gene expression during Arabidopsis seed filling. Plant Cell 14, 1191–1206. doi: 10.1105/tpc.000877
Sasaki, Y., and Nagano, Y. (2004). Plant acetyl-CoA carboxylase: structure, biosynthesis, regulation, and gene manipulation for plant breeding. Biosci. Biotechnol. Biochem. 68, 1175–1184. doi: 10.1271/bbb.68.1175
Shen, B., Allen, W. B., Zheng, P., Li, C., Glassman, K., Ranch, J., et al. (2010). Expression of ZmLEC1 and ZmWRI1 increases seed oil production in maize. Plant Physiol. 153, 980–987. doi: 10.1104/pp.110.157537
Slabas, A. R., and Fawcett, T. (1992). The biochemistry and molecular biology of plant lipid biosynthesis. Plant Mol. Biol. 19, 169–191. doi: 10.1007/BF00015613
Stone, S. L., Kwong, L. W., Yee, K. M., Pelletier, J., Lepiniec, L., Fischer, R., et al. (2001). LEAFY COTYLEDON2 encodes a B3 domain transcription factor that induces embryo development. Proc. Natl. Acad. Sci. U.S.A. 98, 11806–11811. doi: 10.1073/pnas.201413498
Tarczynski, M. C., and Shen, B. (2008). A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat. Genet. 40, 367–372. doi: 10.1038/ng.85
To, A., Joubès, J., Barthole, G., Lécureuil, A., Scagnelli, A., Jasinski, S., et al. (2012). WRINKLED transcription factors orchestrate tissue-specific regulation of fatty acid biosynthesis in Arabidopsis. Plant Cell 24, 5007–5023. doi: 10.1105/tpc.112.106120
Vanhercke, T., El Tahchy, A., Liu, Q., Zhou, X. R., Shrestha, P., Divi, U. K., et al. (2014). Metabolic engineering of biomass for high energy density: oilseed-like triacylglycerol yields from plant leaves. Plant Biotechnol. J. 12, 231–239. doi: 10.1111/pbi.12131
Vigeolas, H., Waldeck, P., Zank, T., and Geigenberger, P. (2007). Increasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific promoter. Plant Biotechnol. J. 5, 431–441. doi: 10.1111/j.1467-7652.2007.00252.x
Voelker, T., and Kinney, A. J. (2001). Variations in the biosynthesis of seed-storage lipids. Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 335–361. doi: 10.1146/annurev.arplant.52.1.335
Wang, H. W., Zhang, B., Hao, Y. J., Huang, J., Tian, A. G., Liao, Y., et al. (2007). The soybean Dof-type transcription factor genes, GmDof4 and GmDof11, enhance lipid content in the seeds of transgenic Arabidopsis plants. Plant J. 52, 716–729. doi: 10.1111/j.1365-313X.2007.03268.x
Wang, X., Devaiah, S. P., Zhang, W., and Welti, R. (2006). Signaling functions of phosphatidic acid. Prog. Lipid Res. 45, 250–278. doi: 10.1016/j.plipres.2006.01.005
Wang, Z., Zhou, Z., Liu, Y., Liu, T., Li, Q., Ji, Y., et al. (2015). Functional evolution of phosphatidylethanolamine binding proteins in soybean and Arabidopsis. Plant Cell 27, 323–336. doi: 10.1105/tpc.114.135103
Wu, X. L., Liu, Z. H., Hu, Z. H., and Huang, R. Z. (2014). BnWRI1 coordinates fatty acid biosynthesis and photosynthesis pathways during oil accumulation in rapeseed. J. Integr. Plant Biol. 56, 582–593. doi: 10.1111/jipb.12158
Zhang, M., Fan, J., Taylor, D. C., and Ohlrogge, J. B. (2009). DGAT1 and PDAT1 acyltransferases have overlapping functions in Arabidopsis triacylglycerol biosynthesis and are essential for normal pollen and seed development. Plant Cell 21, 3885–3901. doi: 10.1105/tpc.109.071795
Zheng, P., Allen, W. B., Roesler, K., Williams, M. E., Zhang, S., Li, J., et al. (2008). A phenylalanine in DGAT is a key determinant of oil content and composition in maize. Nat. Genet. 40, 367–372. doi: 10.1038/ng.85
Zou, J., Katavic, V., Giblin, E. M., Barton, D. L., MacKenzie, S. L., Keller, W. A., et al. (1997). Modification of seed oil content and acyl composition in the Brassicaceae by expression of a yeast sn-2 acyltransferase gene. Plant Cell 9, 909–923. doi: 10.1105/tpc.9.6.909
Keywords: Wrinkled1 (WRI1), oil accumulation, flowering, lipid homeostasis, transcriptional regulation, Brassica napus
Citation: Li Q, Shao J, Tang S, Shen Q, Wang T, Chen W and Hong Y (2015) Wrinkled1 Accelerates Flowering and Regulates Lipid Homeostasis between Oil Accumulation and Membrane Lipid Anabolism in Brassica napus. Front. Plant Sci. 6:1015. doi: 10.3389/fpls.2015.01015
Received: 03 September 2015; Accepted: 02 November 2015;
Published: 19 November 2015.
Edited by:
Chandrashekhar Pralhad Joshi, Michigan Technological University, USAReviewed by:
Biswapriya Biswavas Misra, University of Florida, USAUpinder S. Gill, The Samuel Roberts Noble Foundation, USA
Copyright © 2015 Li, Shao, Tang, Shen, Wang, Chen and Hong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yueyun Hong, aG9uZ3l5QG1haWwuaHphdS5lZHUuY24=