 Richard W. Zobel
Richard W. Zobel- Plant Science Research Unit, Agricultural Research Service, United States Department of Agriculture, Raleigh, NC, USA
The Arabidopsis root system is frequently considered to have only three classes of root: primary, lateral, and adventitious. Research with other plant species has suggested up to eight different developmental/functional classes of root for a given plant root system. If Arabidopsis has only three classes of root, it may not be an adequate model for eudicot plant root systems. Recent research, however, can be interpreted to suggest that pre-flowering Arabidopsis does have at least five (5) of these classes of root. This then suggests that Arabidopsis root research can be considered an adequate model for dicot plant root systems.
Introduction
Root system architecture (RSA) may be a key to the further improvement of crop productivity (Lynch, 2007). A significant amount of research is being carried out on Arabidopsis roots as a model system (Benfy et al., 2010). This research includes both morphological, anatomical, and physiological/molecular studies. Historically this research has referred to two types of Arabidopsis root, the primary root and its lateral roots. More recent studies of adventitious roots on Arabidopsis have begun to elucidate a more complex rooting system. Zobel (2011) has suggested that both monocot and eudicot root systems are made up of 8 or more genetically and functionally distinct classes of root. Does the Arabidopsis root system architecture fit this model or is it a subset with only 2–3 classes of root?
First, what is Root System Architecture? Two systems for describing RSA have been put forward. Fitter (1987) described a method that classified the architecture of a root system, or parts of a root system, as a “topology.” The topological patterns he demonstrated were shown to coincide well with concepts of efficient root exploration for nutrients (Fitter and Stickland, 1991; Fitter et al., 1991). This topology normally describes roots of very young plants or a single major root with it’s attendant laterals. Topology, as defined in dictionaries, however, is the study of the topography of an object. Zobel (2011) describes the topography of an intact whole plant root system, in terms of the major structures that are included with in that topography. Zobel’s RSA, relative to a given plant, can be further defined (refined) through studies of root angle, root diameter, growth rate, differential function, etc. Fitter’s topology describes visual patterns of lateral root development within that topography and thus is also a refining element that can be explored independent of the overall topography.
Second, what is the significance of Zobel’s (2011) RSA (Table 1). The classification proposed was based on documented differential genetic control of the initiation and development of different roots with in the root system of both monocot and eudicot plants. Combined with this was evidence that some of these classes of root are functionally distinct. For instance, for both soybean (Glycine max L.) and maize (Zea mays L.) the tap root and basal roots can have different abilities to penetrate compacted soil or aluminum toxic soil (Zobel, 2011). These abilities vary with cultivar, such that if a plant breeder desires cultivar of soybean that can penetrate Aluminum toxic soil, that breeder would need to identify a cultivar which has the ability for the tap root to penetrate the toxic soil, and another cultivar that has the ability for the basal roots to penetrate the toxic soil. The breeder would then need to hybridize the two cultivars and select from among the offspring a line which has the ability for both the tap root and basal roots to penetrate the toxic soil. The same requirements hold for the other classes of root. This necessitates that function (and molecular patterns) of different classes of root be studied in isolation of the other classes of root. If Arabidopsis is to be a useful model plant, these different classes of root need to be identified within its RSA so they can be studied with the powerful tools already developed for this species.
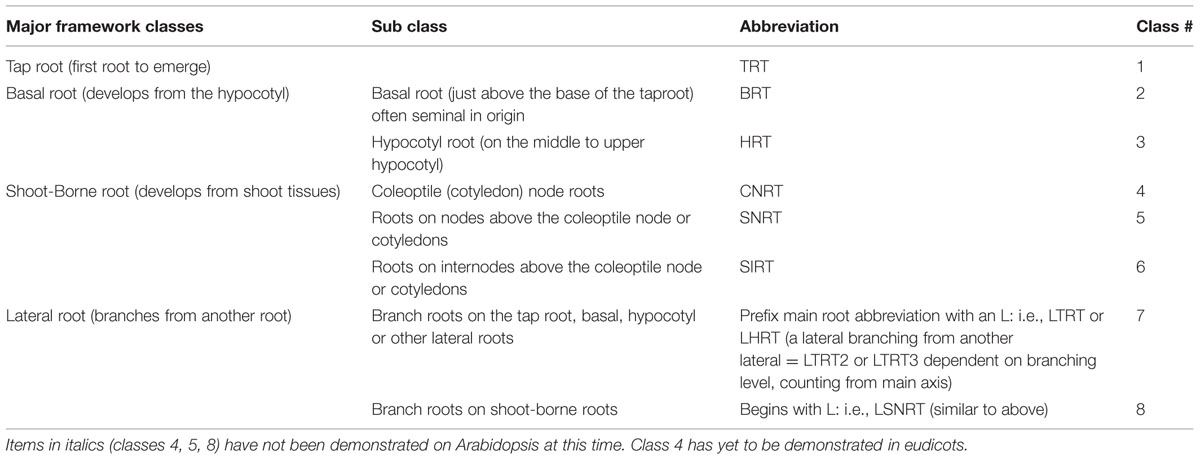
TABLE 1. Table of root nomenclatures, modified from Zobel (2011).
Discussion
Although Arabidopsis is generally considered “the model plant” for eudicots, it’s root system topography (RSA) has been generally accepted to have only two classes of root: tap root (primary root), and lateral roots. This conclusion was probably based on the early research with Arabidopsis roots being restricted to seedling plants, and the classical definition of root system architecture (Esau, 1965). Falasca and Altamura (2003) demonstrated that a third possible class of root, “adventitious,” was distinct from lateral roots, thus raising the number of root classes for Arabidopsis to three. Unfortunately such a conclusion, when presented with Zobel’s (2011) conclusion, suggests that Arabidopsis may not be a fully representative model for eudicot root systems.
More recent research with “adventitious” roots of somewhat older Arabidopsis plants provides some clarity to this situation. The term “adventitious” has two different competing definitions, the strict definition and the common usage definition. The strict definition is that adventitious roots do not arise from pericycle cells. The more general definition is: any root that is not a lateral branch of another root. Because neither of these definitions allows the reader to easily ascertain the actual site of origin of the roots in question, in 1996 the International Society for Root Research (ISRR), declared the term “adventitious” obsolete, and replaced it with term “shoot-borne” (Zobel and Waisel, 2010). Falasca and Altamura (2003) demonstrated that roots arising from the hypocotyl (i.e., non-root tissue) were derived from a pericycle in a manner identical with that of lateral roots of the tap root. Therefore, hypocotyl-borne roots of Arabidopsis, are not adventitious (sensu stricto). Furthermore, Verstraeten et al. (2014) have demonstrated that these hypocotyl-borne roots are distinct from lateral roots in terms of their molecular based initiation patterns. Similar roots from the hypocotyl of beans (Phaseolus vulgaris L.) have been shown to be important for adaptation to low phosphorus conditions (Lynch and Brown, 2001). Welander et al. (2014) took the study of adventitious roots in Arabidopsis one step further and demonstrated molecular differentiation between hypocotyl-borne roots and shoot-borne roots. To add to this, a visual assessment of Celenza et al. (1995 - Figures 9A,C), suggests that the Arabidopsis mutant alf1-1 is a proliferate hypocotyl-borne root mutant. Thus it would appear that Arabidopsis has both hypocotyl roots (HRT) and shoot-borne roots (SIRT) from Zobel’s (2011) topography (Table 1).
Images of wild type Arabidopsis, often show one or more large roots originating at the hypocotyl/taproot junction (i.e., basal; base of the root and base of the shoot; cf Figures 1D,E of King et al., 1995; Figure 3B “WT” in Pacurar et al., 2014; and Figure 1 in Verstraeten et al., 2014; for example). Li and Gao (2011) discuss stimulation of root initiation at the junction of the tap root with the hypocotyl of Arabidopsis by a coumarin derivative (4-nethyllumbelliferone). Combining this with the visual observations, it can be suggested that Arabidopsis has a distinct class of roots originating at the base of the shoot/root. Both the ISRR (Zobel and Waisel, 2010) root classification system and Zobel’s (2011) topography, classify roots from this position as basal roots (BRT). It has been demonstrated in bean that the angle of BRT outgrowth from the axis of the shoot can be critical for efficient phosphorus uptake (Lynch and Brown, 2001).
The topography of the Arabidopsis root system appears to fit with the typical topography of eudicot root systems as presented by Zobel (2011). The root classes of Arabidopsis, using the terms of Zobel (2011, Table 1), thus are: tap root (TRT), basal root (BRT), hypocotyl root (HRT), internodal shoot-borne root (SIRT), and lateral root (LRT). This commonality with other eudicots, strengthens the argument for Arabidopsis root systems being model dicot root systems. Continued molecular and developmental research with these different root classes should help to solidify or adjust the proposed classification of Zobel (2011) for the functional topography of eudicot root systems.
Author Contributions
The author confirms being the sole contributor of this work and approved it for publication.
Funding
Base funds received from U.S. Department of Agriculture.
Conflict of Interest Statement
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Benfy, P. N., Bennett, M., and Schiefelbein, J. (2010). Getting to the root of plant biology: impact of the Arabidopsis genome sequence on root research. Plant J. 61, 992–1000. doi: 10.1111/j.1365-313X.2010.04129.x
Celenza, J. L., Grisafi, P. L., and Fink, G. R. (1995). A pathway for lateral root formation in Arabidopsis thaliana. Genes Dev. 9, 2131–2142. doi: 10.1101/gad.9.17.2131
Falasca, G., and Altamura, M. M. (2003). Histological analysis of adventitious rooting in Arabidopsis thaliana (L.) Heynh seedlings. Plant Biosyst. 137, 265–273. doi: 10.1080/11263500312331351511
Fitter, A. H. (1987). An architectural approach to the comparative ecology of plwnt root systems. New Phytol. 106, 61–77. doi: 10.1111/j.1469-8137.1987.tb04683.x
Fitter, A. H., and Stickland, T. R. (1991). Architectural analysis of plant root sustems 2. Influence of nutrient supply on architecture in contrasting plant species. New Phytol. 118, 383–389. doi: 10.1111/j.1469-8137.1991.tb00019.x
Fitter, A. H., Stickland, T. R., Harvey, M. L., and Wilson, G. W. (1991). Architectural analysis of plant root systems 1. Architectural correlates of exploitation efficiency. New Phytol. 118, 375–382. doi: 10.1111/j.1469-8137.1991.tb00018.x
King, J. J., Stimart, D. P., Fisher, R. H., and Bleeker, A. B. (1995). A mutation altering auxin homeostasis and plant morphology in Arabidopsis. Plant Cell 7, 2023–2037. doi: 10.1105/tpc.7.12.2023
Li, X., and Gao, M. J. (2011). Modulation of root branching by a coumarin derivative. Plant Signal. Behav. 6, 1654–1655. doi: 10.4161/psb.6.11.17768
Lynch, J. P. (2007). Roots of the second green revolution. Aust. J. Bot. 55, 493–512. doi: 10.1071/BT06118
Lynch, J. P., and Brown, K. M. (2001). Topsoil foraging – an architectural adaptation of plants to low phosphorus availability. Plant Soil 237, 225–237. doi: 10.1023/A:1013324727040
Pacurar, D. I., Pacurar, M. L., Bussell, J. D., Schwambach, J., Pop, T. I., Kowalczyk, M., et al. (2014). Identification of new adventitious rooting mutants suppressors of the Arabidopsis thaliana superroot2 mutation. J. Expt. Bot. 65, 1605–1618. doi: 10.1093/jxb/eru026
Verstraeten, I., Schotte, S., and Geelen, D. (2014). Hypocotyl adventitious root organogenesis differs from lateral root development. Front. Plant Sci. 5:495. doi: 10.3389/fpls.2014.00495
Welander, M., Geier, T., Smolka, A., Ahlman, A., Fan, J., and Zhu, L. H. (2014). Origin, timing, and gene expression profile of adventitious rooting in Arabidopsis hypocotyls and stems. Am. J. Bot. 101, 255–266. doi: 10.3732/ajb.1300258
Zobel, R. W. (2011). A developmental genetic basis for defining root classes. Crop Sci. 51, 1410–1413. doi: 10.2135/cropsci2010.11.0652
Keywords: Arabidopsis, roots, root classes, model plants, root system architecture (RSA)
Citation: Zobel RW (2016) Arabidopsis: An Adequate Model for Dicot Root Systems? Front. Plant Sci. 7:58. doi: 10.3389/fpls.2016.00058
Received: 20 November 2015; Accepted: 14 January 2016;
Published: 05 February 2016.
Edited by:
Anna Maria Mastrangelo, CRA-Centro di Ricerca per la Cerealicoltura, ItalyReviewed by:
Frantisek Baluska, University of Bonn, GermanyBoris Rewald, University of Natural Resources and Life Sciences, Vienna (BOKU), Austria
Copyright © 2016 Zobel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Richard W. Zobel, cmljaC56b2JlbEBhcnMudXNkYS5nb3Y=