 Roberta Calafiore1†
Roberta Calafiore1† Valentino Ruggieri1†
Valentino Ruggieri1† Assunta Raiola1
Assunta Raiola1 Maria M. Rigano1
Maria M. Rigano1 Adriana Sacco1
Adriana Sacco1 Mohamed I. Hassan2
Mohamed I. Hassan2 Luigi Frusciante1
Luigi Frusciante1 Amalia Barone1*
Amalia Barone1*- 1Department of Agricultural Sciences, University of Naples Federico II, Portici, Italy
- 2Department of Genetics, Faculty of Agriculture, Assiut University, Assiut, Egypt
The tomato is a model species for fleshy fruit development and ripening, as well as for genomics studies of others Solanaceae. Many genetic and genomics resources, including databases for sequencing, transcriptomics and metabolomics data, have been developed and are today available. The purpose of the present work was to uncover new genes and/or alleles that determine ascorbic acid and carotenoids accumulation, by exploiting one Solanum pennellii introgression lines (IL7-3) harboring quantitative trait loci (QTL) that increase the content of these metabolites in the fruit. The higher ascorbic acid and carotenoids content in IL7-3 was confirmed at three fruit developmental stages. The tomato genome reference sequence and the recently released S. pennellii genome sequence were investigated to identify candidate genes (CGs) that might control ascorbic acid and carotenoids accumulation. First of all, a refinement of the wild region borders in the IL7-3 was achieved by analyzing CAPS markers designed in our laboratory. Afterward, six CGs associated to ascorbic acid and one with carotenoids metabolism were identified exploring the annotation and the Gene Ontology terms of genes included in the region. Variants between the sequence of the wild and the cultivated alleles of these genes were investigated for their functional relevance and their potential effects on the protein sequences were predicted. Transcriptional levels of CGs in the introgression region were extracted from RNA-Seq data available for the entire S. pennellii introgression lines collection and verified by Real-Time qPCR. Finally, seven IL7-3 sub-lines were genotyped using 28 species-specific markers and then were evaluated for metabolites content. These analyses evidenced a significant decrease in transcript abundance for one 9-cis-epoxycarotenoid dioxygenase and one L-ascorbate oxidase homolog, whose role in the accumulation of carotenoids and ascorbic acid is discussed. Comprehensively, the reported results demonstrated that combining genetic and genomic resources in tomato, including bioinformatics tools, was a successful strategy to dissect one QTL for the increase of ascorbic acid and carotenoids in tomato fruit.
Introduction
In recent years increasing attention has been given to the nutritional properties of plant antioxidant compounds, since their consumption has demonstrated to be associated with a reduced risk of cancer, inflammation and cardiovascular diseases. A great contribute to these health effects is attributed to secondary metabolites, including ascorbic acid (AsA, vitamin C) and carotenoids (precursors of Vitamin A) (Canene-Adams et al., 2005; Raiola et al., 2014). Besides their critical role in human nutrition, these compounds have major roles in several plant biological processes, such as photoreception and photoprotection, hormone signaling, cell cycle, cell expansion, plant development, responses to biotic and abiotic stresses. The biosynthetic pathway of carotenoids has been extensively studied and most metabolic key-steps that control their accumulation in plants has been identified (Giuliano, 2014). Plants produce AsA through several biosynthetic pathways, including the D-mannose–L-galactose as the main pathway, even though the role of the L-gulose, the D-galacturonate, and the myo-inositol pathways has also been suggested (Valpuesta and Botella, 2004); in addition the recycling pathway can contribute to the regulation of AsA accumulation (Chen et al., 2003). Finally, since AsA doesn’t diffuse through lipid bilayers because of its negatively charged form at physiological pH values, a class of transporters (Nucleobase Ascorbate Transporter, NAT) may be involved in the mechanisms of AsA accumulation (Badejo et al., 2012; Cai et al., 2014). The level of antioxidants in plants is highly influenced by environmental conditions, and this can explain why in recent years many scientific efforts were focused on better understanding the genetic architecture of this complex trait in various plant species (Davey et al., 2006; Stevens et al., 2007; Hayashi et al., 2012; Fantini et al., 2013; Kandianis et al., 2013; Lisko et al., 2014). Indeed, even though the biosynthesis of carotenoids and AsA in plants is well characterized, their gene regulation and their accumulation in fruits still remain elusive.
Humans are unable to synthetize AsA and carotenoids, and their dietary intake mainly derives from fruit and vegetables. Among these, tomato is the second most consumed vegetable in the world, thus being one of main sources of antioxidants. Indeed, tomato consumption reaches 40–45 kg pro capita per year in countries such as Spain, Italy, or USA (source: FAO databases); used as fresh product or processed (paste, juice, sauce and powder), its antioxidant content may protect against cancer, inflammation and cardiovascular diseases (Canene-Adams et al., 2005; Friedman, 2013).
Tomato is also a reference species for genetic and genomic studies in the Solanaceae family, due to its diploid genome with relative small size (950 Mbp), its short generation time, efficient transformation technologies, high synteny with various Solanaceae and numerous genetic and genomics resources already available (Mueller et al., 2005; Barone et al., 2008). Information data on gene function, genetic diversity and evolution in tomato and in other Solanaceae species are available since the year 2012 when the tomato genome was completely sequenced (Tomato Genome Consortium, 2012). Since then, high-throughput datasets and bioinformatics platforms extremely useful for the Solanaceae plant research community were newly generated or implemented. The Sol Genomics Network1 is a clade-oriented database for the Solanaceae family and its close relatives, which hosts genotypic and phenotypic data and analysis tools. The tomato genome resources database (TGRD2) is a resource that allows investigations on genes, quantitative trait loci (QTL), miRNA, transcription factors (TFs), single sequence repeat (SSR) and SNPs. Other specific databases, generated before the release of the tomato genome, are the SolEST, miSolRNA, Tomatoma, KaTomics, Tomato Functional Genomics Database (TFGD) and several others recently reviewed in Suresh et al. (2014).
Some of these resources might be extremely useful to dissect genetic complex traits into quantitative trait loci, especially when combined with the exploitation of genetic resources, such as the introgression lines (IL). These lines contain a defined homozygous segment of wild genome in a cultivated genetic background and, taken all together, represent a genomic library of the wild species (Eshed and Zamir, 1995). IL populations have been obtained from various wild tomato species, such as Solanum pennellii, S. habrochaites, S. pimpinellifolium, S. lycopersicoides, S. chmielewskii, and S. sitiens (Fernie et al., 2006) and they are useful to identify genes involved in QTLs regulation thus helping the detection of favorable wild alleles controlling the trait under study. The S. pennellii IL population is the most exhaustive; it consists of 76 lines with overlapping wild segments in the cultivated genetic background of the variety M82. These ILs have been widely used to map QTLs (Lippman et al., 2007), have been characterized at genomic and transcriptomic level (Chitwood et al., 2013) and, recently, Alseekh et al. (2013, 2015) carried out their high-dense genotyping and detailed metabolic profiling.
In this work we integrated genomic and transcriptomic data to identify candidate genes (CGs) controlling antioxidant metabolite accumulation in the fruit of S. pennellii IL7-3, which has been previously selected in our laboratory since it harbors a positive QTL for AsA and carotenoids content in the fruit (Sacco et al., 2013; Rigano et al., 2014). In addition, in order to restrict the number of CGs, we selected sub-lines of IL7-3 by the aid of species-specific CAPS markers and evaluated their metabolites content. This allowed us to identify one gene that might control carotenoids levels in the fruit. In addition, we could locate the genes controlling AsA content in a restricted part of the introgressed region 7-3, focusing on the role of one gene involved in AsA recycling pathway. These findings can provide valuable tools for improving the nutritional value of tomato and may represent a focus for future investigations.
Materials and Methods
Plant Material
Plant material consisted of one S. pennellii in S. lycopersicum introgression line (IL7-3, accession LA4102) and the cultivated genotype M82 (accession LA3475). The accessions were kindly provided by the Tomato Genetics Resources Centre3. Sub-lines of the region 7-3 (genotypes coded from R200 to R207) were selected from F2 genotypes previously obtained by intercrossing two ILs (IL12-4 × IL7-3; Sacco et al., 2013). The F2 genotypes were selfed for two generations and then screened by species-specific markers in order to select sub-lines carrying different wild regions at the homozygous condition. Additional IL7-3 sub-lines (genotypes coded from R176 to R182) were kindly provided by Dr. Dani Zamir (Hebrew University, Israel). All genotypes were grown in open-field conditions in the years 2014 and 2015 in a randomized complete block design with three replicates per genotype and 10 plants per replicate. Fruits were collected at three developmental stages (MG: mature green, BR: breaker stage, MR: mature red). Seeds and columella were subsequently removed, and fruits were ground in liquid nitrogen and stored at -80°C until analyses.
Phenotypic Evaluations
Ascorbic Acid Determination
Ascorbic acid determination was carried out by a colorimetric method (Stevens et al., 2006) with modifications reported by Rigano et al. (2014). Briefly, 500 mg of frozen powder were extracted with 300 μl of ice cold 6% TCA. The mixture was vortexed, incubated for 15 min on ice and centrifuged at 14000 rpm for 20 min at 4°C. Twenty microliters of supernatant were placed in an eppendorf tube with 20 μl of 0.4 M phosphate buffer (pH 7.4) and 10 μl of double distilled (dd) H2O. Then, 80 μl of color reagent solution were prepared by mixing solution A [31% H3PO4, 4.6% (w/v) TCA and 0.6% (w/v) FeCl3] with solution B [4% 2,2′-dipyridil (w/v)]. The mixture was incubated at 37°C for 40 min and measured at 525 nm by a NanoPhotometerTM (Implen). Three separated biological replicates for each sample and three technical assays for each biological repetition were measured. The concentration was expressed in nmol of AsA according to the standard curve, designed over a range of 0–70 nmol; then the values were converted into mg/100 g of fresh weight (FW).
Carotenoids Determination
The extraction of carotenoids was carried out according to the method reported by Zouari et al. (2014) with minor modifications. Briefly, one gram of frozen powder was extracted with a solution of acetone/hexane (40/60, v/v) for 15 min. The mixture was centrifuged at 4000 rpm for 10 min and the absorbance of surnatant was measured at 663, 645, 505, and 453 nm. Total carotenoids were determined by the equation reported by Wellburn (1994). Results were expressed as mg per 100 g FW. All biological replicates per sample were analyzed in triplicate.
Molecular Marker Analysis
In order to define the wild region size of IL sub-lines, polymorphic markers spanning the introgression region 7-3 were searched for by exploring the Sol Genomics Network database4. Some markers were retrieved from the database, others markers instead were designed by searching for polymorphisms between the reference tomato sequence (release SL2.50) and the S. pennellii genome (Bolger et al., 2014) using the Tomato Genome Browser5. The primer pairs used to amplify the genomic region were designed using the Primer3web6. Total genomic DNA was extracted from leaves using the PureLinkTM Genomic DNA Kit (Invitrogen). PCR DNA amplification was carried out in 50 μl reaction volume containing 50 ng DNA, 1X reaction buffer, 0.2 mM each dNTP, 1.0 mM primer and 1.25 U GoTaq polymerase (Promega). Discriminating restriction enzymes were identified using the CAPS Designer tool available at the Sol Genomics Network7. The restriction endonuclease reaction was made in 50 μl of reaction volume containing 20 μl PCR product, 5 μl 10X reaction buffer and 1 μl of the selected restriction enzyme (10 u/ml). Digested fragments were separated by electrophoresis on 2% agarose gel in TAE buffer.
Bioinformatic Identification of Candidate Genes
The search for CGs associated with ascorbic acid and carotenoids metabolism was conducted by exploring the annotations and the Gene Ontology terms of the genes included in the region 7-3 of the tomato chromosome 7 (Alseekh et al., 2013). Due to the preliminary annotation of S. pennelliii genome (Bolger et al., 2014), the genes of the wild parent were computationally re-annotated by Blast2Go program (version 38; Conesa and Götz, 2008), to better characterize the gene set and collect additional information on their function. BlastX algorithm (e-value < 1E-6) and NCBI nr protein database were considered for Blast2Go analysis, while the annotation of all the sequences was performed by using default parameters (e-value < 1E-5). The ‘Augment Annotation by ANNEX’ function was also used to refine annotations (implemented in Blast2Go and described in Zdobnov and Apweiler, 2001).
Variants between S. lycopersicum and S. pennellii for all the CGs were obtained by extracting information from the Tomato Variant Browser (Aflitos et al., 2014). In addition, in order to validate the structural variants, the gene sequences were aligned using the genomic sequence information available for both cultivated S. lycopersicum and wild S. pennellii species. The effects of these variants and the prediction on their functional impact on the protein were analyzed using SnpEff v4.2 (Cingolani et al., 2012). The program performs a simple estimation of putative deleteriousness of the variants, classifying them in four classes (HIGH, MODERATE, LOW, MODIFIER, for detailed information refer to the documentation at http://snpeff.sourceforge.net/SnpEff_manual.html). Variants with high impact cause a stop codon or a frame shift; those with moderate impact are missense variants, whereas those with low impact are synonymous SNPs. The potential effect of these polymorphisms on the protein sequence was also cross-validated with the PROVEAN protein tool (publicly available from the J. Craig Venter Institute at http://provean.jcvi.org/seq_submit.php). According to the author’s guideline, we considered a “deleterious” effect of the variant if the PROVEAN score was equal or below -2.5.
The Tomato Functional Genomic Database (TED9), which reports RNA-seq data from the red fruit of S. pennellii ILs, was exploited to verify the expression of the identified CGs in tomato fruits and to estimate their differential expression in M82 and IL7-3. Finally, the TFs mapping in the introgression region were identified by investigating the 2505 TFs present in the Tomato Genomic Resources Database10 and the 1845 TFs categorized in the Plant Transcription Factor Database11. CGs and TFs with an RPKM value <3 in the RNA-seq database were excluded from further analyses and genes/TFs with Log2 ratio (IL7-3/M82) >1.5 or < -1.5 were considered to be differentially expressed, following the thresholds reported by Ye et al. (2015).
Real-Time PCR Amplification of Candidate Genes
Total RNA was isolated from tomato fruit at the three stages of ripening (MG, BR, MR) by TRIzol® reagent (Invitrogen, Carlsbad, CA, USA) and treated with RNase-free DNase (Invitrogen, Carlsbad, CA, USA; Madison, WI, USA) according to the method reported by the manufacturer (Invitrogen). Total RNA (1 μg) was treated by the Transcriptor High Fidelity cDNA Synthesis Kit (Roche) and cDNA was stored at -20°C until RT-PCR analysis. For each PCR reaction, 1 μL of cDNA diluited 1:10 was mixed with 12.5 μL SYBR Green PCR master mix (Applied Biosystems) and 5 pmol each of forward and reverse primers (Supplementary Table S1) in a final volume of 25 μL. The reaction was carried out by using the 7900HT Fast-Real Time PCR System (Applied Biosystems). The amplification program was carried out according to the following steps: 2 min at 50°C, 10 min at 95°C, 0.15 min at 95°C, and 60°C for 1 min for 40 cycles, and followed by a thermal denaturing step (0.15 min at 95°C, 0.15 min at 60°C, 0.15 min at 95°C) to generate the dissociation curves in order to verify the amplification specificity. All the reactions were run in triplicate for each of the three biological replicates and a housekeeping gene coding for the elongation factor 1-α (Ef 1-α) was used as reference gene. The level of expression relative to the reference gene has been calculated using the formula 2-ΔCT, where ΔCT = (CTRNAtarget – CTreferenceRNA) (Schmittgen et al., 2004). Comparison of RNA expression was based on a comparative CT method (ΔΔCT) and the relative expression has been quantified and expressed according to log2RQ, where RQ was calculated as 2-ΔΔCT, and ΔΔCT = (CTRNAtarget – CTreferenceRNA) – (CTcalibrator – CTreferenceRNA) (Winer et al., 1999; Livak and Schmittgen, 2001). M82 MG, BR, and MR were selected as calibrators for the three analyzed stages of ripening. Quantitative results were expressed as the mean value ± SE. Differences among samples were determined by using Statistical Package for Social Sciences (SPSS) Package 6, version 15.0. Significance was determined by comparing the genotypes for each stage of ripening through a t-Student’s test at a significance level of 0.05.
Results
Phenotypic Evaluation of Parental Lines
In order to confirm the presence of one positive QTL for AsA and carotenoids in the region 7-3, metabolic analyses were performed for two consecutive years on mature red fruits of the cultivated genotype M82 and of IL7-3 grown in open fields (Table 1). In both years, IL7-3 accumulated a significant higher level of AsA and carotenoids in the fruit compared to M82, confirming data previously reported in our laboratory (Di Matteo et al., 2010; Sacco et al., 2013; Rigano et al., 2014). The metabolites content was also estimated in three different ripening stages (mature green – MG, breaker – BR, and mature red – MR) as shown in Figure 1. In M82, the AsA level increased from MG to BR and then decreased from BR to MR; accordingly, in IL7-3 the AsA level increased in the first ripening stages but did not decrease in MR. The total carotenoids content deeply increased in both genotypes from BR to MR as expected, and was higher in IL7-3.

TABLE 1. Evaluation of metabolite content (ascorbic acid, and total carotenoids, mean and standard error) in mature red fruit of genotypes M82 and IL7-3 in the years 2014 and 2015.
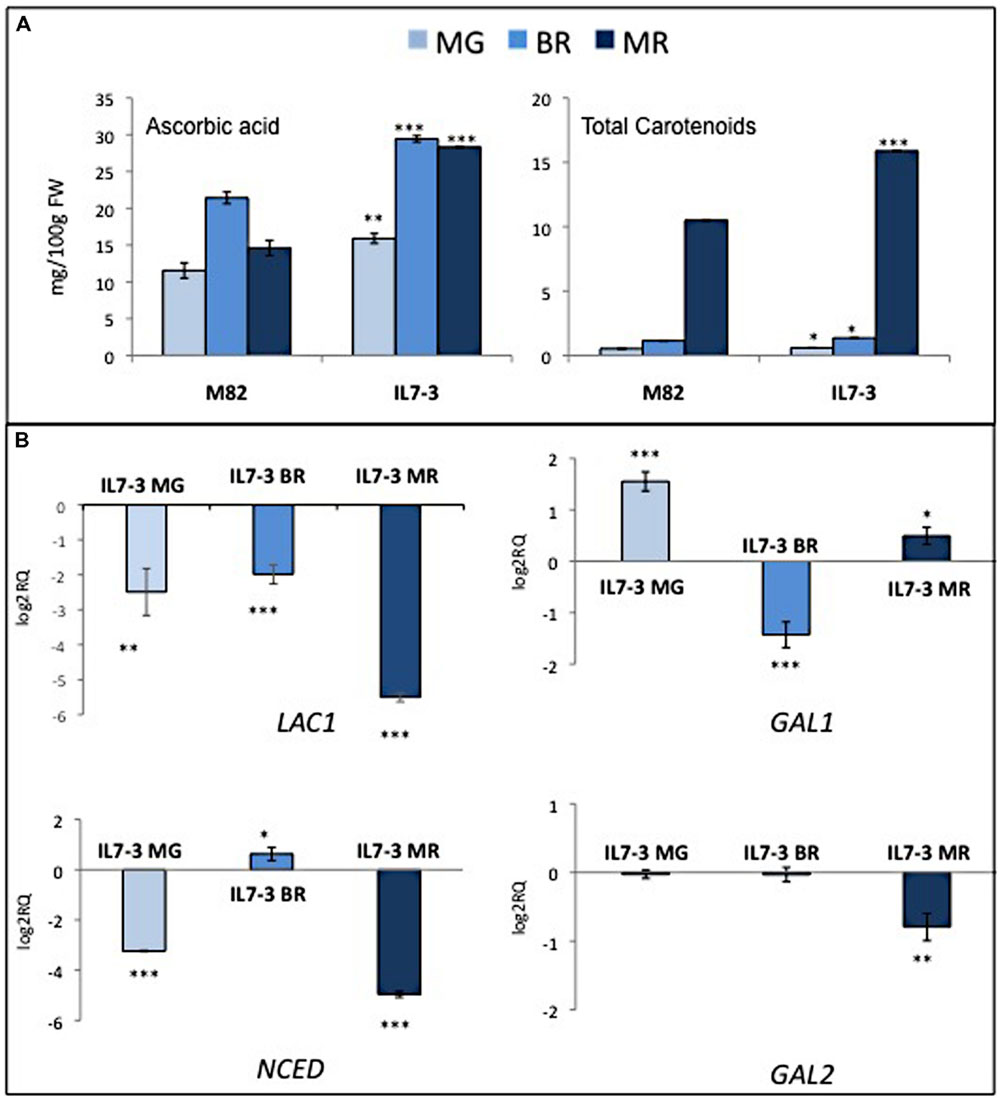
FIGURE 1. Phenotypic and molecular evaluation of the parental genotypes M82 and IL7-3. (A) Metabolite content (AsA and total carotenoids) at three different ripening stages (MG, mature green; BR, breaker; MR, mature red); (B) Expression level of four selected CGs (LAC1: laccase-22/L-ascorbate-oxidase homolog; GAL1 and GAL2: β-1-3-galactosiltrasferase; NCED: 9-cis-epoxycarotenoid dioxygenase) in IL7-3 at different ripening stages. Asterisks indicate statistically significant differences of each ripening stage to the corresponding M82 stage (∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001).
Identification of Candidate Genes (CGs)
In order to identify CGs controlling AsA and carotenoids content in IL7-3, we firstly better defined the introgression region size. At this purpose, we selected species-specific molecular markers at the two region borders, referring to those reported in the Sol Genomics Network database12 and taking into account the information on S. pennellii ILs reported in Chitwood et al. (2013) regarding the chromosomal positions of ILs boundaries. By testing ten markers (from N22 to N28 at the upper border, and from N12 to N30 at the lower border, Table 2) on the parental genotypes M82 and on IL7-3, we ascertained that the wild region stretches from marker N27 (corresponding to Solyc07g048030 at 59,218,716 bp) to marker N17 (corresponding to Solyc07g063330 at 65,816,155 bp), covering about 6.6 Mbp. This region includes 725 genes (Supplementary Table S2), 120 (16.5%) were annotated as unknown proteins, whereas 94 (13.0%) were TFs. Among the remaining 511 annotated genes, we searched for those related to AsA and carotenoids accumulation. Six CGs putatively involved in determining AsA content were detected (Table 3), but none of them belong to the main biosynthetic galactose pathway. The identified genes were: one polygalacturonase (Solyc07g056290, POLYGAL), two beta-1-3-galactosyltransferase (Solyc07g052320 and Solyc07g062590, GAL1 and GAL2, respectively), two laccase-22/L-ascorbate-oxidase homolog (Solyc07g052230 and Solyc07g052240, LAC1 and LAC2, respectively), and one nucleobase-ascorbate transporter (Solyc07g049320, NAT). The investigation of the SolCyc biochemical pathways database13 allowed confirming the involvement of the gene POLYGAL in the galacturonate AsA biosynthetic pathway (enzymatic step EC 3.2.1.15), and of GAL1 and GAL2 in enzymatic reactions (EC 2.1.4-) potentially regulating myo-inositol content, that might feed the glucuronate biosynthetic pathway. LAC1 and LAC2 might enter the recycling pathway of AsA by reducing L-ascorbate into monodehydroascorbate (EC 1.10.3.3), whereas the NAT might have a role in transporting AsA among the different intracellular compartments. In addition, in the introgression region one 9-cis-epoxycarotenoid dioxygenase (Solyc07g056570, NCED) was also mapped that, entering the carotenoids pathway, determines the carotenoids oxidative cleavage with consequent production of apocarotenoids, the direct substrates for abscisic acid (ABA) synthesis. The latter gene was included into the group of those to be further investigated.
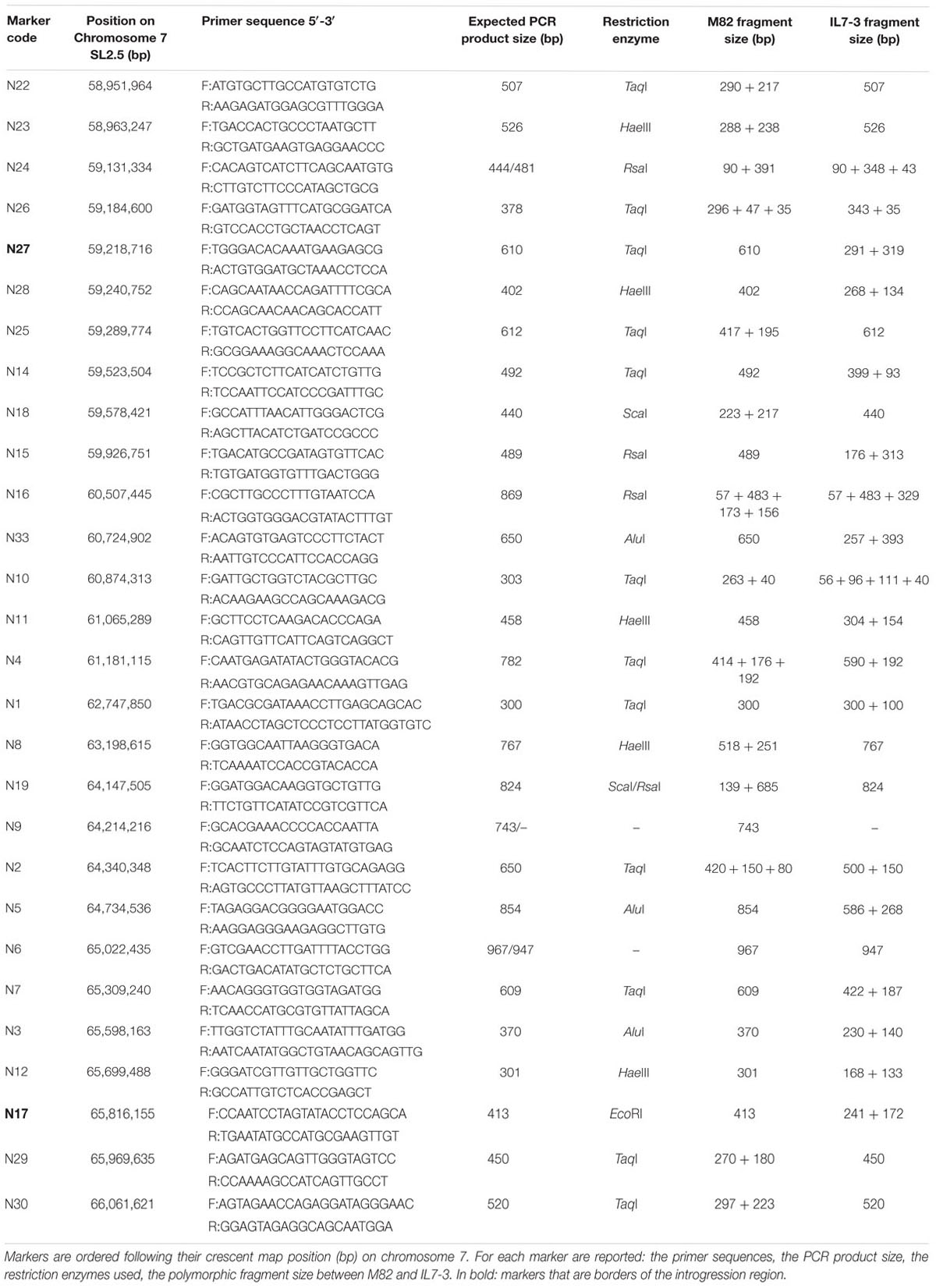
TABLE 2. CAPS molecular markers used to define the whole introgression region 7-3 and the sub-lines obtained from this region.
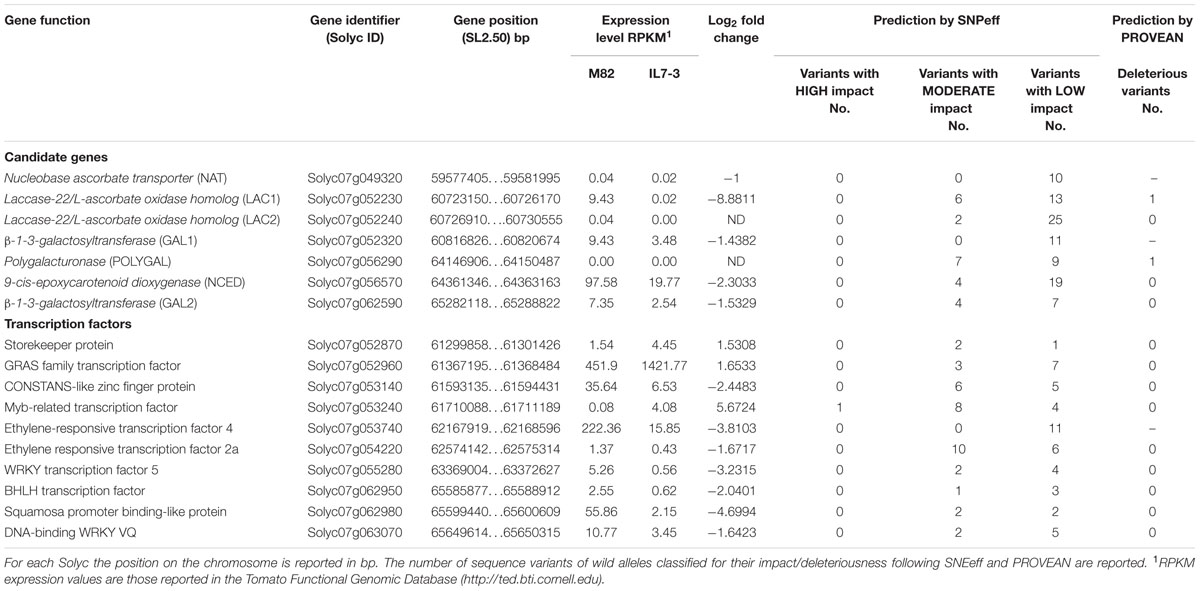
TABLE 3. Candidate genes for ascorbic acid and carotenoids, and transcription factors mapping in the introgressed region 7-3, all selected for their log2 fold change <-1.5 or >1.5.
Sequence Variation and Expression Variability of Selected CGs
In order to better define which CGs determine the different metabolites content between M82 and IL7-3 fruit, differences in their sequence and/or in their expression level were investigated. The impact of polymorphisms between S. lycopersicum and S. pennellii was estimated for the 117 variations identified in CGs mapping in the introgression region (Table 3). No case of high impact polymorphism was detected, and NAT and GAL1 did not even exhibit any variants with moderate impact effect. For the other genes, the number of variants with moderate impact varied from two (LAC2) to seven (POLYGAL), and a deleterious effect at the protein level investigated by PROVEAN was predicted for genes LAC1 and POLYGAL.
The RNA-seq data available for M82 and IL7-3 in the TFGD (Fei et al., 2010) allowed to ascertain that three CGs for AsA were not expressed or expressed at very low levels in the red fruit (Table 3): NAT, LAC2 and POLYGAL. By contrast, GAL1, GAL2, LAC1 and NCED showed a lower expression level in IL7-3 compared to M82. The expression of all CGs was analyzed by qRT-PCR in three developmental stages of M82 and IL7-3 fruits. This analysis allowed confirming the lack of expression of NAT, LAC2 and POLYGAL. The different expression levels of LAC1 and/or GAL2 detected here well correlate with the different trend of AsA accumulation in M82 and IL7-3 in the three developmental fruit stages (Figure 1), even though also the down-regulation of GAL1 at BR stage could be relevant. Finally, the significant lower expression of the gene NCED at the MR stage well correlated with the higher level of total carotenoids observed in IL7-3.
Besides the identified CGs, TFs mapping in the introgression region 7-3 might play a role in increasing antioxidants. Indeed, if differentially expressed or polymorphic between M82 and IL7-3, they could trans-regulate the expression of genes involved in AsA or carotenoids biosynthesis and accumulation mapping in the introgression or in other regions of the genome. Sequence variations with deleterious effects on the protein functionality were found in 27 TFs (Supplementary Table S3), but in most cases (85%) the polymorphic TFs were not expressed in the fruit (Supplementary Table S4). The ten TFs selected for their significant differential expression between IL7-3 and M82 (Supplementary Table S3) did not show any sequence variation that cause deleterious effect as predicted by PROVEAN. Most of them exhibited a lower expression in IL7-3, except for one MYB (Solyc07g053240), one GRAS (Solyc07g052960) and one storekeeper protein (Solyc07g052870), but none corresponded to the TFs identified by Ye et al. (2015) for their correlation with expression of genes involved in high AsA and carotenoids in tomato fruit. The availability of the whole transcriptome of M82 and IL7-3 in the TED database allowed also investigating the expression of all the genes involved in AsA and carotenoids biosynthetic pathways, which were annotated in the tomato genome (Supplementary Table S5). No differentially expressed gene among these was identified. When looking at the whole transcriptome, an unbalance of ascorbate-oxidase activity could be hypothesized in IL7-3 compared to M82. Indeed, besides the gene LAC1 of the introgression 7-3, two other laccases-22/L-ascorbate-oxidase, mapping on chromosomes 2 and 8, showed a decreased expression in IL7-3. They could exert an additive action to that of the wild LAC1 in increasing AsA content in the fruit. Finally, two myo-inositol phosphate synthase (Solyc04g050820 and Solyc05g051850) were over-expressed in IL7-3, and they could affect the myo-inositol pathway, as well as the two GAL1 and GAL2 that map into the introgression.
Selection and Phenotyping of IL7-3 Sub-lines
In order to reduce the number of CGs potentially responsible for the higher antioxidant content in the IL7-3 red ripe fruit, introgression sub-lines of the wild region were selected through the analysis of 28 polymorphic markers mapping within this region. Comprehensively, we identified seven distinct sub-lines showing a reduced wild region compared to IL7-3 and carrying different combinations of wild alleles for four CGs (Figure 2). Only two sub-lines, R182 (from marker N27 to N14) and R181 (from marker N7 to N17), had no wild alleles for the CGs, even though they carried different introgressed wild regions. Four sub-lines carried two wild CGs for AsA (LAC1 and GAL1) but differed for the presence (R201, R202) or absence (R176, R178) of the wild allele for NCED gene. Finally, one sub-line (R179) carried wild alleles for GAL2 and NCED genes. All the sub-lines were grown in open field and were evaluated for AsA and carotenoids content in the fruit (Table 4). The carotenoids level of the three sub-lines carrying the wild allele for NCED (R179, R201, R202) was higher than in M82; sub-lines carrying the cultivated allele showed a level of carotenoids comparable to M82. As for AsA, it was evident that two sub-lines (R179 and R181) exhibited levels of AsA comparable to that of the control genotype M82, whereas all the others showed AsA content significantly higher than M82 and similar to IL7-3. Therefore, considering the different combinations of wild CGs carried by the sub-lines, it is possible to exclude the role of GAL2 wild allele in increasing AsA content in IL7-3. By contrast, the wild alleles of both LAC1 and GAL1 might be involved in increasing AsA in IL7-3 and in a group of sub-lines, even though the qRT-PCR results evidenced that the action of GAL1 occurs earlier at BR. The expression level of the selected CGs NCED and LAC1 was verified in the sub-lines (Figure 3) to better ascertain their role in affecting carotenoids and AsA, respectively. Surprisingly, a higher expression of NCED compared to M82 and a concurrent higher level of AsA were observed in R182, a sub-line with a reduced introgression size of 200 kb, where no wild allele for any CGs and no differentially expressed or polymorphic TFs was retained. A total of 24 genes map in this region, including two unknown genes.

FIGURE 2. Size and genomic identity of the seven sub-lines selected from the region 7-3. For each sub-line (coded R), the boundary edges are indicated by the CAPS markers (coded N) that separated the wild genomic region (in green) from the cultivated one (in red). The position of the four CGs (LAC1, GAL1, NCED, GAL2) in the region 7-3 is also reported.

TABLE 4. Phenotyping and genotyping of the seven S. pennellii introgression sub-lines: for each sub-line the higher content of ascorbic acid (AsA) and total carotenoids respect to M82 is reported, together with the presence of wild alleles for four candidate genes (CG), and the number of genes classified as unknown and transcription factors (TF), which map in the introgressed region.
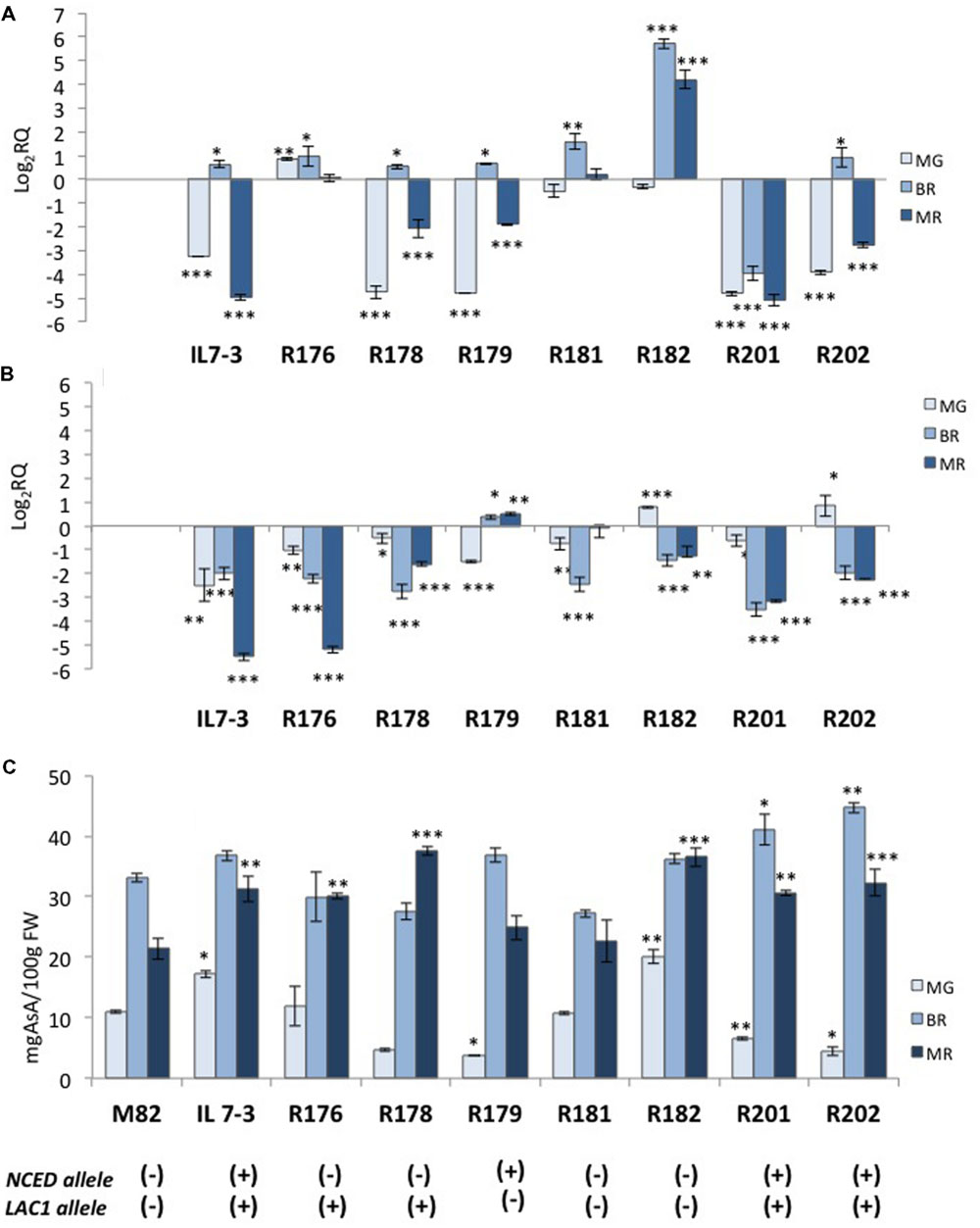
FIGURE 3. Gene expression of two CGs (NCED and LAC1) and ascorbic acid (AsA) content in the fruit of the seven sub-lines of the region 7-3. The expression level of NCED (A) and LAC1 (B) in the parental IL7-3 and its sub-lines is reported in comparison to that observed in the parental M82 at three different ripening stages, together with the AsA content (C) in the fruit evaluated at the same stages for all genotypes. Asterisks indicate statistically significant differences of each ripening stage to the corresponding M82 stage (∗P < 0.05, ∗∗P < 0.01, ∗∗∗P < 0.001). The presence of the wild (+) or cultivated (-) alleles of NCED and LAC1 genes for each genotype is reported at the bottom.
Discussion
The exploitation of wild Solanum species has driven the improvement of tomato varieties for several traits by using traditional and innovative breeding approaches (Bai and Lindhout, 2007). The wild species are precious sources of new alleles for improving specific traits, most of which are quantitatively inherited and therefore highly influenced by environmental conditions and by multiple interactions among a consistent number of genes. In this view, the production of ILs from tomato wild species helped to dissect many complex traits into major QTLs (Lippman et al., 2007), which might be then transferred into improved varieties. This genetic effort is boosted by the availability of genomic tools and resources, which may have a deep impact on the success of this breeding strategy.
The S. pennellii IL7-3 has been selected in our laboratory for its higher antioxidant properties compared to the cultivated variety M82 (Sacco et al., 2013; Rigano et al., 2014). It exhibited stable performances in different years and we could therefore assume that these properties depend on a strong genetic basis. By integrating data coming from many genomics resources publicly available, we exploited this genetic resource with the aim of identifying CGs and their wild alleles, which may contribute to increase AsA and total carotenoids in the fruit. One CG was identified that might affect carotenoids accumulation: the NCED gene, which controls a key-enzyme in ABA biosynthesis (Zhang et al., 2009). The lower expression of this gene in IL7-3 might reduce the metabolic flux toward ABA production, pushing the upstream metabolic pathway and thus feeding carotenoids accumulation, as proposed by Sun et al. (2012), who observed an increase of carotenoids level and a reduction of ABA when the gene SlNCED1 was silenced in tomato fruit. Also in our case, a concurrent increase of carotenoids and decrease of ABA was observed in IL7-3 compared to M82 (data not shown), thus supporting the role attributed to the wild allele of NCED. In addition, even though no deleterious impact on proteins was detected by PROVEAN when comparing the wild and cultivated alleles of NCED, the alteration of amino acid sequences may result in enzymes with modified activities (Yuan et al., 2015).
Understanding the genetic control of the higher AsA content in IL7-3 fruit was more complicated. Out of six CGs identified in the introgressed region, which might be involved in the synthesis and accumulation of AsA, only three were expressed in the fruit. Among these, the gene LAC1 was expressed only in traces in IL7-3 fruits respect to M82, as retrieved from available RNAseq data in the TED database, and confirmed in our work using three primer pairs for Real Time PCR targeting different regions of this gene. In addition, a deleterious missense variant (-6.337 PROVEAN score) has been detected in LAC1 when comparing the sequences of S. lycopersicum and S. pennellii. This caused a substitution of a glycine in glutamic acid at position 194 (G194E), which affects the cupredoxin domain. The deleterious alteration of the protein in that position might be crucial for its correct functionality. The two GAL genes mapping in the introgression exhibited both a slight lower expression in IL7-3, and might contribute to enhance AsA content in the fruit. Finally, the wild allele of NCED might be indirectly related to the increased level of AsA, since an intricated relationship between ABA biosynthesis and AsA metabolism has been already hypothesized in different species, including Arabidopsis, strawberry, tomato, and Ocimum (Ghassemian et al., 2008; Nair et al., 2009; Lima-Silva et al., 2012; Dongdong et al., 2015). However, in some cases the correlation occurred between ABA content and ascorbate oxidase expression level without any modifications of NCED expression (Lopez-Carbonell et al., 2006; Fotopoulos et al., 2008). Therefore, the AsA increase in IL7-3 might be also attributed to variations of ABA content determined by the reduced activity of NCED, LAC1 or both the enzymes.
Since the introgressed region 7-3 has a large size (6.6 Mbp), we analyzed sub-lines of the region, which allowed focusing on a restricted number of CGs. The selected sub-lines carried only two or three CGs, or even no CG at all. The phenotypic characterization of the sub-lines led us to draw some conclusions on the potential role of the CGs. Firstly, the carotenoids content well-reflected the presence/absence of the wild allele for NCED, even though its specific action should be further investigated, since the lower expression of NCED is correlated to a higher carotenoids content only in three out of four sub-lines (R179, R201, and R202). Therefore, approaches of gene replacement between the wild and the cultivated allele will be undertaken to verify that the presence of the wild allele may be effectively correlated to higher levels of carotenoids content. Secondly, the wild allele of NCED is not essential for determining the higher AsA content in IL7-3, even though it can contribute to increase it, as discussed above. Thirdly, the down-regulation of GAL2 is not correlated to the higher AsA content in IL7-3 and in sub-lines.
Unfortunately, it was not possible to disrupt the linkage between the two CGs LAC1 and GAL1, and then clearly identify if only one or both of them control the AsA increase. However, it is expected that more genes mapping in one QTL contribute to affect one phenotype, and therefore in our case it may be assumed that both LAC1 and GAL1 might have a determinant role in increasing AsA, and this could confirm the existence of polygenes in the QTL under study. In particular, the down-regulation of GAL1 at BR stage would reduce the metabolic flux toward the myo-inositol biosynthetic pathway. Moreover, it is worth saying that, when the whole transcriptome of IL7-3 was analyzed in comparison to that of M82, two genes annotated as inositol-3-phosphate synthase were over-expressed in IL7-3, with a potential contrasting action on the myo-inositol pathway respect to GAL1. Since the involvement of the latter pathway in the AsA biosynthesis in plants is still controversial (Endres and Tenhaken, 2009; Torabinejad et al., 2009; Gallie, 2013), we did not focus further attention on GAL1.
The role of LAC1 is supported by its down-regulation in IL7-3 MR fruit and in the sub-lines that exhibited a high level of AsA (R176, R178, R201, R202). Indeed, the ascorbate oxidase is an apoplastic enzyme that catalyzes the reversible oxidation of ascorbate to dehydroascorbate, through the formation of monodehydroascorbate, with the concomitant reduction of molecular oxygen to water. Transgenic plants over-expressing or under-expressing this gene have shed light on its role in regulating the apoplastic ascorbate pool (Pignocchi et al., 2003; Sanmartin et al., 2003), and therefore the ascorbate redox state, thus also influencing the perception of environmental stresses (Yamamoto et al., 2005; Fotopoulos et al., 2006, 2008; Garchery et al., 2013). In our case, the down-regulation of the wild laccase-22/L-ascorbate oxidase LAC1 might have an effect comparable to that described in RNAi lines with reduced ascorbate oxidase activity (Zhang et al., 2011), which exhibited high ascorbic acid accumulation in tomato fruit. This effect could explain the significantly higher Asa level observed in IL7-3 and in the sub-lines with respect to the control M82. In the future, the potential correlation between the down-regulation of LAC1 in our tomato genotypes and their improved response to agents imposing oxidative stress will be also investigated.
Finally, in the sub-line R182 the increased level of AsA detected in the red ripe fruits should be further investigated, but it confirms that the IL7-3 AsA QTL consists of more than one CG. Among the 24 genes mapping in the small introgressed region of R182, two annotated as unknown were detected, whereas no TFs was differentially expressed in tomato fruit. In the future, the functional role of all these 24 genes in modulating AsA in tomato fruit will be further investigated by using reverse genetic approaches to clearly define their specific role in controlling AsA content in the tomato mature fruit.
It is also worth saying that the existence in the 7-3 region of several genes annotated as unknown proteins paves the way to other hypotheses. Indeed, besides the differential expression and the structural variants affecting enzyme activity, additional transcriptional and translational interations may occur and contribute to influence it. Recently, regulators of AsA biosynthesis in plants have been described (reviewed in Zhang, 2012). In Arabidopsis it has been demonstrated that light regulates AsA synthesis through the interaction between a photomorphogenetic factor and the enzyme GDP-mannose pyrophosphorylase (Wang et al., 2013), as well as that a feedback regulation of Asa biosynthesis occurs following the interaction between ascorbate and an Open Reading Fame (ORF) in the long 5′ UTR (untranslated region) of the GDP-L-galactose phosphorylase gene (Laing et al., 2015). Now, it will be crucial to demonstrate if similar mechanisms or others may operate in tomato fruit and, therefore, influence AsA concentration in IL7-3.
Conclusion
Results reported in the present work clearly demonstrated that exploiting the genetic and genomic resources nowadays available for tomato, including bioinformatics tools, was a successful strategy to dissect one positive QTL for the increase of AsA and carotenoids in the mature fruit. In particular, two CGs for improving these metabolites were detected in the wild region 7-3 introgressed from the species S. pennellii. These were one L-ascorbate oxidase (LAC1) and one 9-cis-epoxycarotenoid dioxygenase (NCED), whose wild alleles, exhibiting polymorphisms and/or differential transcript levels, might increase AsA and total carotenoids content. The first CG favors the accumulation of reduced ascorbate controlling the redox state of ascorbate in the apoplast. The action of the second CG still needs to be elucidated, even though the presence of the wild allele for NCED was correlated to higher carotenoids content. Finally, the latter gene might also indirectly contribute to increase AsA content, as revealed by the sub-line R182, which showed a high expression of the cultivated allele for NCED combined with high AsA content. A group of 24 genes mapping in the wild introgression of the sub-line R182 will be further investigated in the future to better understand their role in the architecture of the QTL that positively influences the level of antioxidants in the investigated region of the chromosome 7.
Author Contributions
RC and VR contributed to bioinformatic and experimental analyses carried out to identify CGs and the introgression sub-lines, and to draft the manuscript; AR contributed to metabolic and transcriptomic analyses; MR contributed to metabolic analysis and critically revised the manuscript; AS contributed to the bioinformatic analysis and drafted the manuscript; MH contributed to molecular marker analysis and to grow materials; LF contributed to the conception of the experiment and critically revised the manuscript; AB contributed to the experiment design, to data analysis and interpretation, to draft the manuscript.
Funding
This research was supported by the Italian Ministry of University and Research (MIUR) [grant MIUR-PON02-GenoPOMpro].
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.00397
Footnotes
- ^https://solgenomics.net/
- ^http://59.163.192.91/tomato2/
- ^http://tgrc.ucdavis.edu/
- ^https://solgenomics.net/
- ^http://www.tomatogenome.net/VariantBrowser
- ^http://primer3.ut.ee/
- ^http://solgenomics.net/tools/caps_designer/caps_input.pl
- ^https://www.blast2go.com/
- ^http://ted.bti.cornell.edu/
- ^59.163.192.91/tomato2/tfs.html
- ^planttfdb.cbi.pku.edu.cn
- ^solgenomics.net
- ^solcyc.solgenomics.net/
References
Aflitos, S. A., Schijlen, E., de Jong, H., de Ridder, D., Smit, S., Finkers, R., et al. (2014). Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 80, 136–148. doi: 10.1111/tpj.12616
Alseekh, S., Ofner, I., Pleban, T., Tripodi, P., Di Dato, F., Cammareri, M., et al. (2013). Resolution by recombination: breaking up Solanum pennellii introgressions. Trends Plant Sci. 8, 536–538. doi: 10.1016/j.tplants.2013.08.003
Alseekh, S., Tohge, T., Wendenberg, R., Scossa, F., Omranian, N., Li, J., et al. (2015). Identification and mode of inheritance of quantitative trait loci for secondary metabolite abundance in tomato. Plant Cell 27, 485–512. doi: 10.1105/tpc.114.132266
Badejo, A. A., Wada, K., Gao, Y., Maruta, T., Sawa, Y., Shigeoka, S., et al. (2012). Translocation and the alternative D-galacturonate pathway contribute to increasing the ascorbate level in ripening tomato fruits together with the D-mannose/L-galactose pathway. J. Exp. Bot. 63, 229–239. doi: 10.1093/jxb/err275
Bai, Y., and Lindhout, P. (2007). Domestication and breeding of tomatoes: what have we gained and what can we gain in the future? Ann. Bot. 100, 1085–1094. doi: 10.1093/aob/mcm150
Barone, A., Chiusano, M. L., Ercolano, M. R., Giuliano, G., Grandillo, S., and Frusciante, L. (2008). Structural and functional genomics of tomato. Int. J. Plant Genomics 2008:820274. doi: 10.1155/2008/820274
Bolger, A., Scossa, F., Bolger, M. E., Lanz, C., Maumus, F., and Tohge, T. (2014). The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat. Genet. 46, 1034–1038. doi: 10.1038/ng.3046
Cai, X., Ye, J., Hu, T., Zhang, Y., and Ye, Z. (2014). Genome-wide classification and expression analysis of nucleobase–ascorbate transporter (NAT) gene family in tomato. Plant Growth Regul. 73, 19–30. doi: 10.1007/s10725-013-9864-x
Canene-Adams, K., Campbell, J. K., Zaripheh, S., Jeffery, E. H., and Erdman, J. W. Jr. (2005). The tomato as a functional food. J. Nutr. 135, 1226–1230.
Chen, Z., Young, T. E., Ling, J., Chang, S. C., and Gallie, D. R. (2003). Increasing vitamin C content of plants through enhanced ascorbate recycling. Proc. Natl. Acad. Sci. U.S.A. 100, 3525–3530. doi: 10.1073/pnas.0635176100
Chitwood, D. H., Kumar, R., Headland, L. R., Ranjan, A., Covington, M. F., Ichihashi, Y., et al. (2013). A quantitative genetic basis for leaf morphology in a set of precisely defined tomato introgression lines. Plant Cell 25, 2465–2481. doi: 10.1105/tpc.113.112391
Cingolani, P., Platts, A., Wang le, L., Coon, M., Nguyen, T., Wang, L., et al. (2012). A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 6, 80–92. doi: 10.4161/fly.19695
Conesa, A., and Götz, S. (2008). Blast2GO: a comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genomics 2008:619832. doi: 10.1155/2008/619832
Davey, M. W., Kenis, K., and Keulemans, J. (2006). Genetic control of fruit vitamin C contents. Plant Physiol. 142, 343–351. doi: 10.1104/pp.106.083279
Di Matteo, A., Sacco, A., Anacleria, M., Pezzetti, M., Delledonne, M., Ferrarini, A., et al. (2010). The ascorbic acid content of tomato fruits is associated with the expression of genes involved in pectin degradation. BMC Plant Biol. 10:163. doi: 10.1186/1471-2229-10-163
Dongdong, L., Li, L., Luo, Z., Mou, W., Mao, L., and Yieng, T. (2015). Comparative transcriptomic analysis reveals the influence of abscissic acid on the metabolism of pigments, ascorbic acid and folic acid during strawberry fruit ripening. PLoS ONE 10:e0130037. doi: 10.1371/journal.pone.0130037
Endres, S., and Tenhaken, R. (2009). Myoinositol oxygenase controls the level of myoinositol in Arabidopsis, but does not increase ascorbic acid. Plant Physiol. 149, 1042–1049. doi: 10.1104/pp.108.130948
Eshed, Y., and Zamir, D. (1995). An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics 141, 1147–1162.
Fantini, E., Falcone, G., Frusciante, S., Giliberto, L., and Giuliano, G. (2013). Dissection of tomato lycopene biosynthesis through virus-induced gene silencing. Plant Physiol. 163, 986–998. doi: 10.1104/pp.113.224733
Fei, Z., Joung, J. G., Tang, X., Zheng, Y., Huang, M., Lee, J. M., et al. (2010). Tomato functional genomics database: a comprehensive resource and analysis package for tomato functional genomics. Nucleic Acids Res. 39, D1156–D1163. doi: 10.1093/nar/gkq991
Fernie, A. R., Tadmore, Y., and Zamir, D. (2006). Natural genetic variation for improving crop quality. Curr. Opin. Plant Biol. 9, 196–202. doi: 10.1016/j.pbi.2006.01.010
Fotopoulos, V., De Tullio, M. C., Barnes, J., and Kanellis, A. K. (2008). Altered stomatal dynamics in ascorbate oxidase over-expressing tobacco plants suggest a role for dehydroascorbate signalling. J. Exp. Bot. 59, 729–737. doi: 10.1093/jxb/erm359
Fotopoulos, V., Sanmartin, M., and Kanellis, A. K. (2006). Effect of ascorbate oxidase over-expression on ascorbate recycling gene expression in response to agents imposing oxidative stress. J. Exp. Bot. 57, 3933–3943. doi: 10.1093/jxb/erl147
Friedman, M. (2013). Anticarcinogenic, cardioprotective, and other health benefits of tomato compounds lycopene, α-tomatine, and tomatidine in pure form and in fresh and processed tomatoes. J. Agr. Food Chem. 61, 9534–9550. doi: 10.1021/jf402654e
Gallie, D. R. (2013). L-Ascorbic acid: a multifunctional molecule supporting plant growth and development. Scientifica 2013:795964. doi: 10.1155/2013/795964
Garchery, C., Gest, N., Do, P. T., Alhagdow, M., Baldet, P., Menard, G., et al. (2013). A diminution in ascorbate oxidase activity affect carbon allocation and improves yield in tomato under water deficit. Plant Cell Environ. 36, 159–175. doi: 10.111/j.1365-3040.2012.02564.x
Ghassemian, M., Lutes, J., Chang, H. S., Lange, I., Chen, W., Zhu, T., et al. (2008). Abscisic acid-induced modulation and redox control pathways in Arabidopsis thaliana. Phytochemistry 69, 2899–2911. doi: 10.1016/j.phytochem.2008.09.020
Giuliano, G. (2014). Plant carotenoids: genomics meets multi-gene engineering. Curr. Opin. Plant Biol. 19, 111–117. doi: 10.1016/j.pbi.2014.05.006
Hayashi, E., You, Y., Lewis, R., Calderon, M. C., Wan, G., and Still, D. W. (2012). Mapping QTL, epistasis and genotype x environment interaction of antioxidant activity, chlorophyll content and head formation in domesticated lettuce (Lactuca sativa). Theor. Appl. Genet. 124, 1487–1502. doi: 10.1007/s00122-012-1803-0
Kandianis, C. B., Stevens, R., Liu, W., Palacios, N., Mintgomery, K., Pixley, K., et al. (2013). Genetic architecture controlling variation in grain carotenoid composition and concentrations in two maize populations. Theor. Appl. Genet. 126, 2879–2895. doi: 10.1007/s00122-013-2179-5
Laing, W. A., Martinez-Sanchez, M., Wright, M. A., Bulley, S. M., Brewster, D., Dare, A. P., et al. (2015). An upstream Open Reading Frame is essential for feedback regulation of ascorbate biosynthesis in Arabidopsis. Plant Cell 27, 772–786. doi: 10.1105/tcp.114.133777
Lima-Silva, V., Rosado, A., Amorim-Silva, V., Muñoz-Mérida, A., Pons, C., Bombarely, A., et al. (2012). Genetic and genome-wide transcriptomic analyses identify co-regulation of oxidative response and hormone transcript abundance with vitamin C content in tomato fruit. BMC Genomics 13:187. doi: 10.1186/1471-2164-13-187
Lippman, Z. B., Semel, Y., and Zamir, D. (2007). An integrated view of quantitative trait variation using tomato interspecific introgression lines. Curr. Opin. Genet. Dev. 17, 545–552. doi: 10.1016/j.gde.2007.07.007
Lisko, K. A., Aboobucker, S. I., Torres, R., and Lorence, A. (2014). “Engineering elevated vitamin C content in plants to improve their nutritional content, growth, and tolerance to abiotic stress,” in Phytochemicals-Biosynthesis, Function and Application, Recent Advances in Phytochemistry 44, ed. R. Jetter (Berlin: Springer International Publishing Switzerland), 109–128.
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lopez-Carbonell, M., Munné-Bosch, S., and Alegre, L. (2006). The ascorbate-deficient vtc-1 Arabidopsis mutant shows altered ABA accumulation in leaves and chloroplast. J. Plant Growth Regulat. 25, 137–144. doi: 10.1007/s00344-005-0119-6
Mueller, L. A., Solow, T., Taylor, N., Skwarecki, B., Buels, R., Binns, J., et al. (2005). The SOL Genomics Network. A comparative resource for Solanaceae biology and beyond. Plant Physiol. 138, 1310–1317. doi: 10.1104/pp.105.060707
Nair, V. D., Cheruth, A. J., Gopi, R., Gomathinayagam, M., and Panneerselvam, R. (2009). Antioxidant potential of Ocimum sanctum under growth regulator treatments. Eur. Asian J. Biosci. 3, 1–9. doi: 10.5053/ejobios.2009.3.0.1
Pignocchi, C., Fletcher, J. M., Wilkinson, J. E., Barnes, J. D., and Foyer, C. H. (2003). The function of ascorbate oxidase in tobacco. Plant Physiol. 132, 1631–1641. doi: 10.1104/pp.103.022798
Raiola, A., Rigano, M. M., Calafiore, R., Frusciante, L., and Barone, A. (2014). Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediat. Inflammat. 2014, 1–16. doi: 10.1155/2014/139873
Rigano, M. M., Raiola, A., Tenore, G. C., Monti, D. M., Del Giudice, R, Frusciante, L., et al. (2014). Quantitative trait loci pyramiding can improve the nutritional potential of tomato (Solanum lycopersicum) fruits. J. Agric. Food. Chem. 62, 11519–11527. doi: 10.1021/jf502573n
Sacco, A., Di Matteo, A., Lombardi, N., Trotta, N., Punzo, B., Mari, A., et al. (2013). Quantitative trait loci pyramiding for fruit quality traits in tomato. Mol. Breed 31, 217–222. doi: 10.1007/s11032-012-9763-2
Sanmartin, M., Drogoudi, P. A., Lyons, T., Pateraki, I., Barnes, J., and Kanellis, A. K. (2003). Over-expression of ascorbate oxidase in the apoplast of transgenic tobacco results in altered ascorbate and glutathione redox states and increased sensitivity to ozone. Planta 216, 918–928.
Schmittgen, T. D., Jiang, J., Liu, Q., and Yang, L. (2004). A high-throughput method to monitor the expression of microRNA precursors. Nucleic Acids Res. 32, 43. doi: 10.1093/nar/gnh040
Stevens, R., Buret, M., Duffe, P., Garchery, C., Baldet, P., Rothan, C., et al. (2007). Candidate genes and Quantitative Trait Loci affecting fruit ascorbic acid content in three tomato populations. Plant Physiol. 143, 1943–1953. doi: 10.1104/pp.106.091413
Stevens, R., Buret, M., Garchery, C., Carretero, Y., and Causse, M. (2006). Technique for rapid small-scale analysis of vitamin C levels in fruit and application to a tomato mutant collection. J. Agric. Food Chem. 54, 6159–6165. doi: 10.1021/jf061241e
Sun, L., Yuan, B., Zhang, M., Wang, L., Cui, M., Wang, Q., et al. (2012). Fruit-specific RNAi-mediated suppression of SlNCED1 increases both lycopene and β-carotene contents in tomato fruit. J. Exp. Bot. 63, 3097–3108. doi: 10.1093/jxb/ers026
Suresh, B. V., Roy, R., Sahu, K., Misra, G., and Chattopadhyay, D. (2014). Tomato Genomic Resources Database: an integrated repository of useful tomato genomic information for basic and applied research. PLoS ONE 9:86387. doi: 10.1371/journal.pone.0086387
Tomato Genome Consortium (2012). The tomato genome sequence provides insight into fleshy tomato. Nature 485, 635–641. doi: 10.1038/nature11119
Torabinejad, J., Donahue, J. L., Gunesekera, B. N., Allen-Daniels, M. J., and Gillaspy, G. E. (2009). VTC4 is a bifunctional enzyme that affects myoinositol and ascorbate biosynthesis in plants. Plant Physiol. 150, 951–961. doi: 10.1104/pp.108.135129
Valpuesta, V., and Botella, M. A. (2004). Biosynthesis of L-ascorbic acid in plants: new pathways for an old antioxidant. Trends Plant Sci. 9, 573–577. doi: 10.1016/j.tplants.2004.10.002
Wang, J., Yu, Y., Zhang, Z., Quan, R., Zhang, H., Ma, L., et al. (2013). Arabidopsis CSN5B interacts with VTC1 and modulates ascorbic acid synthesis. Plant Cell 25, 626–636. doi: 10.1105/tpc.112.106880
Wellburn, A. R. (1994). The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J. Plant Physiol. 144, 307–313. doi: 10.1016/S0176-1617(11)81192-2
Winer, J., Jung, C. K., Shackel, I., and Williams, P. M. (1999). Development and validation of real-time quantitative reverse transcriptase-polymerase chain reaction for monitoring gene expression in cardiac myocytes in vitro. Anal. Biochem. 270, 41–49. doi: 10.1006/abio.1999.4085
Yamamoto, A., Bhuiyan, M. N., Waditee, R., Tanaka, Y., Esaka, M., Oba, K., et al. (2005). Suppressed expression of the apoplastic ascorbate oxidase gene increases salt tolerance in tobacco and Arabidopsis plants. J. Exp. Bot. 56, 1785–1796. doi: 10.1093/jxb/eri167
Ye, J., Hu, T., Yang, C., Li, H., Yang, M., Ijaz, R., et al. (2015). Transcriptome profiling of tomato fruit development reveals transcription factors associated with ascorbic acid, carotenoid and flavonoid biosynthesis. PLoS ONE 10:e0130885. doi: 10.1371/journal.pone.0130885
Yuan, H., Zhang, J., Nageswaran, D., and Li, L. (2015). Carotenoid metabolism and regulation in horticultural crops. Hort. Res. 2, 15036. doi: 10.1038/hortres.2015.36
Zdobnov, E. M., and Apweiler, R. (2001). InterProScan-an integration platform for the signature-recognition methods in InterPro. Bioinformatics 17, 847–848. doi: 10.1093/bioinformatics/17.9.847
Zhang, M., Yuan, B., and Leng, P. L. (2009). The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 60, 1579–1588. doi: 10.1093/jxb/erp026
Zhang, Y. (2012). Ascorbic Acid in Plants: Biosynthesis, Regulation and Enhancement. Berlin: Springer Science & Business Media.
Zhang, Y., Li, H., Shu, W., Zhang, C., Zhang, W., and Ye, Z. (2011). Suppressed expression of ascorbate oxidase gene promotes ascorbic acid accumulation in tomato fruit. Plant Mol. Biol. Rep. 29, 638–645. doi: 10.1007/s11105-010-0271-4
Keywords: ascorbic acid, total carotenoids, Solanum pennellii, wild alleles, introgression sub-lines, L-ascorbate oxidase, 9-cis-epoxycarotenoid dioxygenase
Citation: Calafiore R, Ruggieri V, Raiola A, Rigano MM, Sacco A, Hassan MI, Frusciante L and Barone A (2016) Exploiting Genomics Resources to Identify Candidate Genes Underlying Antioxidants Content in Tomato Fruit. Front. Plant Sci. 7:397. doi: 10.3389/fpls.2016.00397
Received: 14 January 2016; Accepted: 14 March 2016;
Published: 08 April 2016.
Edited by:
Antonio Granell, Consejo Superior de Investigaciones Científicas, SpainReviewed by:
Angelos K. Kanellis, Aristotle University of Thessaloniki, GreeceLorenzo Zacarias, Consejo Superior de Investigaciones Científicas, Spain
Copyright © 2016 Calafiore, Ruggieri, Raiola, Rigano, Sacco, Hassan, Frusciante and Barone. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amalia Barone, YW1iYXJvbmVAdW5pbmEuaXQ=
†These authors have contributed equally this work.