 Pei-Li Fu
Pei-Li Fu Jussi Grießinger
Jussi Grießinger Aster Gebrekirstos
Aster Gebrekirstos Ze-Xin Fan
Ze-Xin Fan Achim Bräuning
Achim Bräuning- 1Key Laboratory of Tropical Forest Ecology, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, China
- 2Institute of Geography, University of Erlangen-Nürnberg, Erlangen, Germany
- 3World Agroforestry Centre, Nairobi, Kenya
Stable isotopes in wood cellulose of tree rings provide a high-resolution record of environmental conditions, yet intra-annual analysis of carbon and oxygen isotopes and their associations with physiological responses to seasonal environmental changes are still lacking. We analyzed tree-ring stable carbon (δ13C) and oxygen (δ18O) isotope variations in the earlywood (EW) and latewood (LW) of pines from a secondary forest (Pinus kesiya) and from a natural forest (Pinus armandii) in southwestern China. There was no significant difference between δ13CEW and δ13CLW in P. kesiya, while δ13CEW was significantly higher than δ13CLW in P. armandii. For both P. kesiya and P. armandii, δ13CEW was highly correlated with previous year’s δ13CLW, indicating a strong carbon carry-over effect for both pines. The intrinsic water use efficiency (iWUE) in the earlywood of P. armandii was slightly higher than that of P. kesiya, and iWUE of both pine species showed an increasing trend, but at a considerably higher rate in P. kesiya. Respective δ13CEW and δ13CLW series were not correlated between the two pine species and could be influenced by local environmental factors. δ13CEW of P. kesiya was positively correlated with July to September monthly mean temperature (MMT), whereas δ13CEW of P. armandii was positively correlated with February to May MMT. Respective δ18OEW and δ18OLW in P. kesiya were positively correlated with those in P. armandii, indicating a strong common climatic forcing in δ18O for both pine species. δ18OEW of both pine species was negatively correlated with May relative humidity and δ18OEW in P. armandii was negatively correlated with May precipitation, whereas δ18OLW in both pine species was negatively correlated with precipitation during autumn months, showing a high potential for climate reconstruction. Our results reveal slightly higher iWUE in natural forest pine species than in secondary forest pine species, and separating earlywood and latewood of for δ18O analyses could provide seasonally distinct climate signals in southwestern China.
Introduction
Due to the intense human impact, including large scale forest harvesting and resultant land use changes, the area of natural forests in tropical and subtropical China is remarkably decreasing in favor of secondary forest or plantation (Fang et al., 2001; Piao et al., 2009). The transformation of land use and vegetation types may have strong impacts on the local climate and the regional hydrological cycle (Li et al., 2009). Hence, it is of urgent importance to understand the responses of different vegetation types to climate factors and environmental change, especially in tropical and subtropical areas in southwestern China that are exposed to extreme summer monsoon precipitation events which may trigger devastating flood events.
Stable carbon and oxygen isotopes from tree rings (δ13C, δ18O) are frequently used in environmental research, as they provide a continuous, annually resolved record of environmental conditions and show stronger correlations between tree individuals and environmental variables than radial tree growth (McCarroll and Loader, 2004). Tree-ring δ13C is controlled by the balance between stomatal conductance and photosynthetic rate (Farquhar et al., 1980; Leavitt and Long, 1988). Thus δ13C is frequently correlated with air humidity or precipitation in dry environments (Gebrekirstos et al., 2009, 2011; Kress et al., 2009; Brienen et al., 2011), whereas it is associated with irradiance factors and growing season temperature in humid environments (McCarroll and Loader, 2004). δ13C has been widely used to calculate intrinsic water use efficiency (iWUE) and to estimate differences in water use among different plants (McCarroll and Loader, 2004; Gebrekirstos et al., 2011; Gessler et al., 2014). Recent studies reported an increase of iWUE with elevated CO2 in sub-tropical and tropical regions (Brienen et al., 2011; Xu Y. et al., 2014).
Tree-ring δ18O is mainly controlled by the source water δ18O, leaf water exchange and xylem/phloem water exchange and thus reflects variations in hydroclimate (McCarroll and Loader, 2004). In high mountain regions of southwestern China, δ18O is a good indicator for monsoon moisture (An et al., 2012) or cloudiness (Liu et al., 2014). δ18O in Fokienia hodginsii is a promising proxy of drought variability in Laos (Xu et al., 2011). δ18O in teak (Tectona grandis) is positively correlated with annual precipitation in western and central India, but a negative correlations with annual precipitation was found in southern India (Managave et al., 2011). The combined analysis of δ13C and δ18O is a promising way to investigate tree physiological responses to environmental changes (McCarroll and Loader, 2004). However, there are only few studies that investigated the tree-ring carbon and oxygen isotope variations in subtropical and tropical areas in southwestern China (An et al., 2012; Xu Y. et al., 2014).
The earlywood in tree rings is produced during spring and early summer, while the latewood is produced in late summer and autumn, thus earlywood and latewood may potentially record climatic signals in different seasons (Fritts, 1976/1987). In addition, wood formation may be influenced by carbohydrates synthesized in the previous growing season and remobilized to form earlywood in the following spring (Kagawa et al., 2006). Such a carbon carry-over effect was proven in the tropical conifer Podocarpus falcatus from the Ethiopian highlands (Krepkowski et al., 2013), whereas Kress et al. (2009) showed that Scots pine did not rely on stored carbon reserves from previous years at tree-line sites in European mountain regions. Thus, the contribution of the carry-over effect to wood formation might differ between species and regions. In regions influenced by the Asian summer monsoon climate, An et al. (2012) found that earlywood δ18O was affected by early monsoon season temperature and relative humidity (RH), whereas latewood δ18O was correlated with late monsoon precipitation and RH. Therefore, separate analyses in tree ring earlywood and latewood stable isotopes may shed light on the physiological responses of tree species to seasonal environmental change.
The secondary forest formed by dominant stands of Pinus kesiya Royle ex Gord var. langbianensis is an important type of natural forest replacement and have a wide distribution across southern Yunnan, southwestern China (Li et al., 2015). These secondary forests were established after the destruction the natural evergreen Lithocarpus forest in 1970s (Young and Wang, 1989). Pinus armandii Franchet is a regional natural forest species, which is able to survive in higher elevations than P. kesiya. The two pine species share a very similar wood anatomy and form clearly distinguishable growth ring boundaries, which is not very common among tree species of this subtropical ecosystem. Furthermore, the annual growth rates of the two pine species are rather high, and show a clear color distinction between earlywood and latewood, enabling us to carry out intra-annual stable isotope analysis. We analyzed the tree-ring stable carbon (δ13C) and oxygen (δ18O) isotope variations in the earlywood and latewood of the two pine species. Our aims were (1) to study whether the earlywood and latewood of the two pine species differed in their δ13C and δ18O and if δ13C in earlywood are influenced by a carry-over effect of carbohydrates assimilated in the previous growing season; (2) to investigate if there exist differences in iWUE of pine species from the secondary forest and the natural forest and their response to elevated CO2; (3) to detect the seasonal climatic signals that control tree ring δ13C and δ18O in earlywood and latewood of the studied species. We hypothesized that (1) natural forest pine would have higher iWUE than the secondary forest pine; (2) tree ring δ18O in earlywood and latewood of both pine species might capture seasonal moisture signals (precipitation, relative humidity), that earlywood recorded early growing season moisture signal, while latewood recorded autumn moisture signal.
Materials and Methods
Study Site and Climate
This study was carried out in a subtropical forest ecosystem of the Ailao Mountains, Yunnan Province, southwestern China. The Ailao Mountains form the major climatic border between regions influenced by the southwestern Asian summer monsoon and East Asian summer monsoon (Young and Wang, 1989), and harbors remnants of natural evergreen subtropical forest in higher elevations above 2000 m a.s.l. Local climate data are available from the Ailaoshan Station for Subtropical Forest Ecosystem Studies (ALS) (1982–2012, 24°31′ N, 101°01′ E, 2480 m a.s.l), and a longer data record covering the period 1956–2012 from a weather station in Jingdong County, located 60 km far from ALS in a valley (24°28′ N, 100° 52′ E, 1162.3 m a.s.l.) (Figure 1).
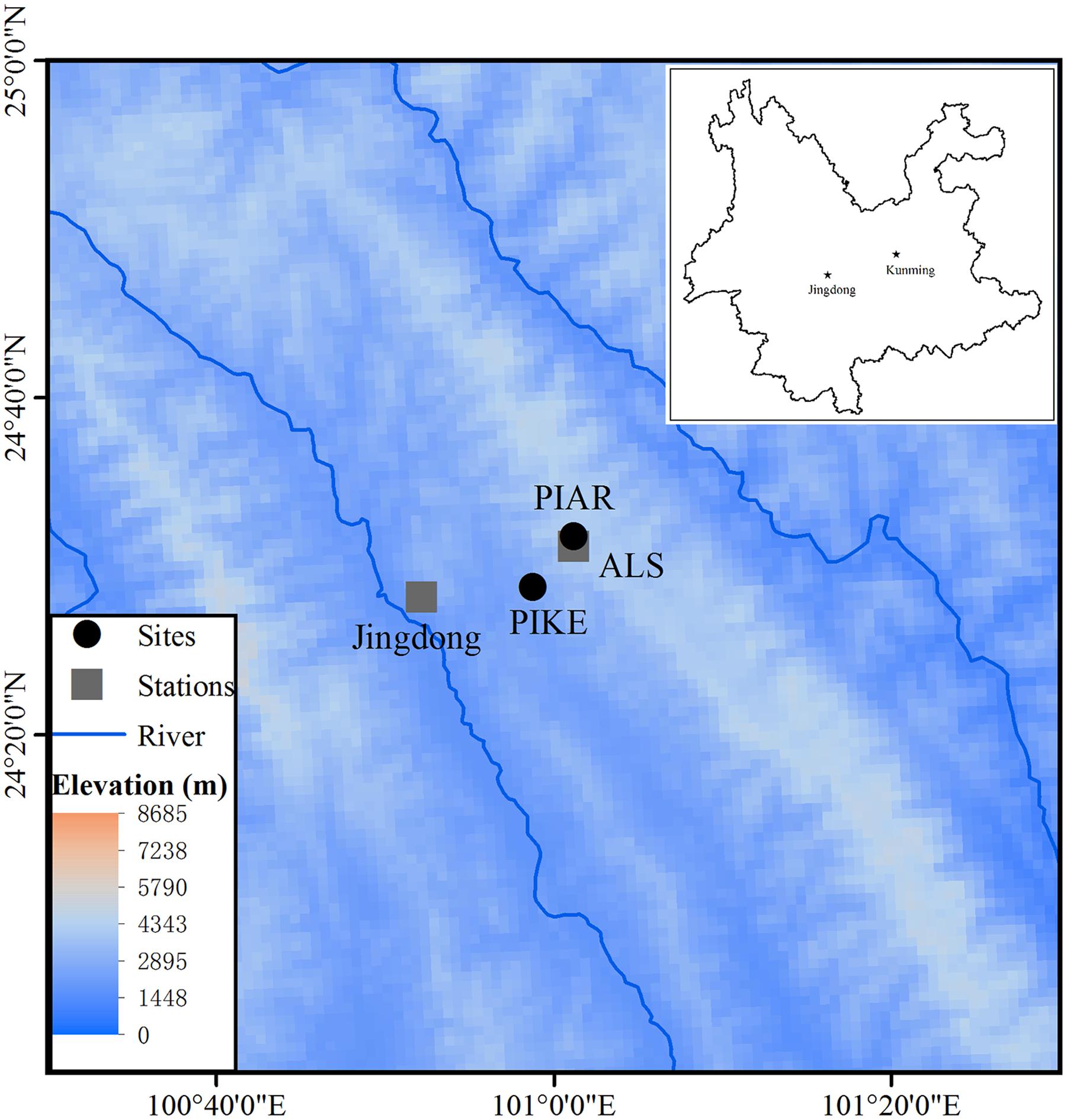
FIGURE 1. Locations of sampling sites of P. kesiya (PIKE) and P. armandii (PIAR) and meteorological stations in the Ailaoshan Station for Subtropical Forest Ecosystem Studies (ALS) and Jingdong County within Yunnan Province, southwest China (in lay map).
The mean annual precipitation at ALS is 1880 mm, of which more than 86% falls during the rainy season from May to October. The mean annual temperature is 11°C, with a minimum of 5.3°C in January and a maximum of 15.3°C in July (Figure 2A). In January, temperature may drop below 0°C. The investigated forest is a subtropical cloud forest with heavy fog during June to August and November to December. Annual mean precipitation at Jingdong County amounts 1118 mm and mean annual temperature is 18.6°C (Figure 2B). Although differing in absolute values, the temporal variations of temperature and precipitation between Jingdong County and ALS are synchronous, therefore the data of both stations are highly correlated (Supplementary Table S1, Figure 2). However, RH during the late summer and autumn months shows poor correlation between the two stations (Supplementary Table S1), since ALS is mainly situated in a dense cloud cover during summer months while Jinglong is located in a river valley. Hence, we used temperature and precipitation data from Jingdong with longer record but RH data from ALS for further correlation analysis.
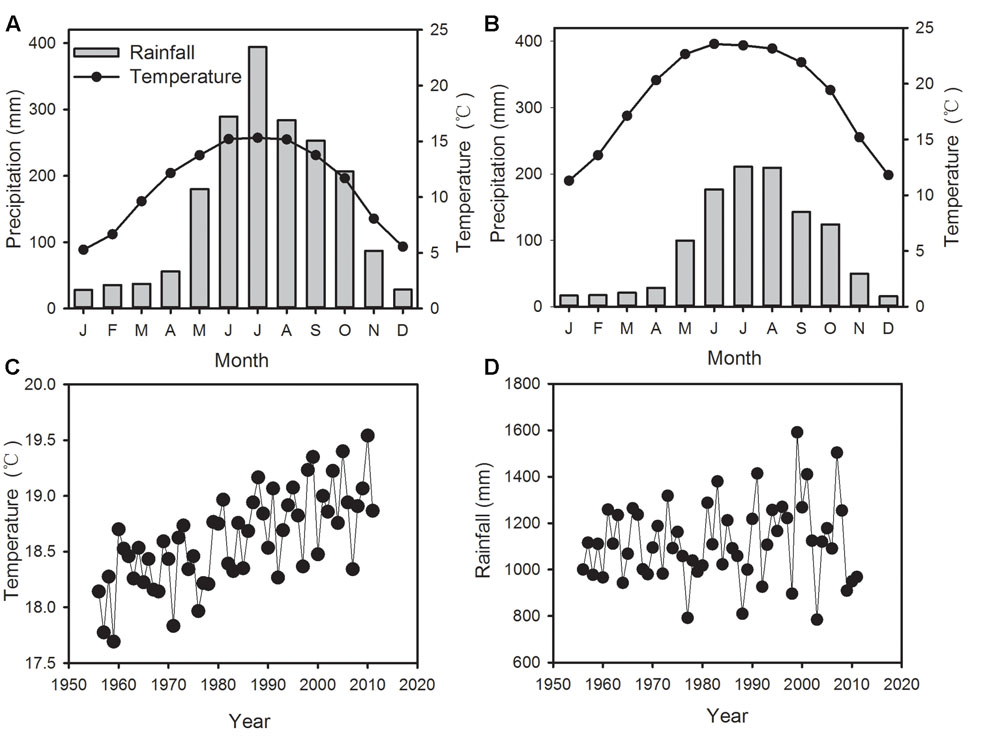
FIGURE 2. Monthly mean temperature and precipitation in Ailaoshan Station for Subtropical Forest Ecosystem Studies (ALS) (A) and in Jingdong County (B), time series of annual mean temperature (C) and precipitation (D) in Jingdong County from 1956–2012.
Study Species and Tree-Ring Sampling
Pinus kesiya var. langbianensis is a geographic variant of P. kesiya, and P. kesiya has a wide distribution in SE Asia, occurring in Myanmar, India, Laos, Vietnam, Thailand, Philippines, and some African countries (Armitage and Burley, 1980). For short, we used ‘P. kesiya’ instead of ‘P. kesiya var. langbianensis’ in the following parts of this paper. P. kesiya is the dominant tree species in the secondary forest of Ailao Mountains and is associated by Schima noronhae at elevations between 700 and 2000 m a.s.l. Pinus armandii can be found in mountain areas and river basins at elevations ranging from 1000 to 3300 m a.s.l. of southwestern China. In our study sites, this species is distributed in remnants of the natural Lithocarpus forests above 2000 m a.s.l. According to the dendrometer data, stem growth of both P. kesiya and P. armandii started in April and ended in October (Supplementary Figure S2).
We collected two increment cores for each tree, and 40 cores (20 trees) of P. kesiya from a secondary pine forest (24°29′ N, 100°59′ E, 1980 m a.s.l.), and 36 cores (18 trees) of P. armandii from a natural forest close to ALS (24°32′ N, 100°1′ E, 2495 m a.s.l.). The stem density in the tree layer of the P. kesiya is lower (1,032 stems ha-1 and canopy cover less than 50%) than those of the natural Lithocarpus forest (21,400 stems ha-1 and canopy cover more than 95%) (Young and Wang, 1989). The sampling site of P. kesiya is located on a north-facing slope with an inclination of ca. 10°, and the soil type is Nitisol. The local farmers continuously harvest the resin of P. kesiya when the tree diameter at breast height (DBH) exceeds 20 cm (Wang et al., 2006). There are no evidences of human managements, such as logging, irrigation and fertilization. The sampling site of P. armandii is a north facing slope with an inclination of ca. 20°, and the soil type is Luvisol.
Tree-ring widths were measured with traditional technique under the stereomicroscope linked to a LINTAB digital positioning table with a resolution of 0.001 mm (LIBTABTM 6, Rinntech, Germany). Tree-ring measurements were cross-dated to the calendar year of their formation by growth pattern matching, and statistical tests using the software package TSAP-Win (Rinn, 2003). A visual dating was applied for P. kesiya, since the trees are mostly younger than 30 years. Tree ring-width chronology of P. armandii spanned from 1906 to 2010, with an average growth rate of 3.34 mm per year (Supplementary Figure S1). The mean sensitivity (0.426) and high inter-series correlation (r = 0.66) indicate the reliable quality of the ring-width chronologies.
We selected cores from each of six P. kesiya individuals and each of five P. armandii individuals for further analysis of δ13C and δ18O. The average tree ages for selected trees is 32 ± 4 years. for P. kesiya, and 45 ± 5 years. for P. armandii, respectively. To better disentangle seasonal influences on the δ13C and δ18O variations in the tree-rings, earlywood (EW) and latewood (LW) of the annuals rings were cut separately with a razor blade, according to their bright and dark color.
Cellulose Extraction
Alpha-cellulose was extracted by using the Multiple Samples Isolation Systems according to Wieloch et al. (2011) and Qin et al. (2015). Resin, fatty acids, etheric oils, and hemicellulose were extracted with a solution of 5% NaOH for 2 h at 60°C for two times. Then lignin was extracted with 7% NaClO2 solution at 60°C, an 8 h × 5 times process was applied in order to achieve an adequate chemical treatment. The remaining hemicelluloses were extracted with 17% NaOH for 2 h at room temperature. A washing procedure with boiled water was interposed for three times across different steps. Finally, samples were treated once with 1% HCl and rinsed with boiling de-ionized water for three times and transferred from the filter funnels into Eppendorf tubes with 1 ml de-ionized water. The samples were then homogenized by using an ultrasound unit (Hielscher Ultrasonics, c.f. Laumer et al., 2009). After freeze drying for 72 h in a lyophilisation unit, the dried α-cellulose samples were processed to the mass spectrometry. δ13C and δ18O were measured with an Elemental Analyzer coupled to a Delta V Advantage IRMS (Thermo Fisher) while laboratory standards were periodically interposed to test analytical replication. The δ13C and δ18O values were referred to International Standards (VPDB, VSMOW) and their analytical error lie within typically reported analytical precisions δ13C = ±0.15′, δ18O = ±0.3′
Calculation of Intrinsic Water Use Efficiency (iWUE)
The measured carbon isotope values were corrected for anthropogenically induced trends of a rising atmospheric CO2 (McCarroll and Loader, 2004). The δ13C and CO2 concentration (ca) of ambient air during 1956–2004 were obtained from McCarroll and Loader (2004) and those data for 2005–2012 were provided by Prof. Danny McCarroll (University of Swansea, UK).
According to Farquhar et al. (1982), isotope discrimination (Δ) is defined as follows:
Where δ13Cplant and δ13Cair refer to the δ13C values of the α-cellulose and of atmospheric CO2, respectively. Δ was also showed to be closely correlated with ci/ca by Farquhar et al. (1982):
Where a represents isotopic discrimination occurring during diffusion of CO2 from the atmosphere into the intercellular spaces of leaves with a = 4.4‰; b represents the isotopic fractionation through discrimination that occurs during enzymatic carboxylation with b = 27‰. By combining Equations (1) and (2), the intercellular CO2 (ci) can be calculated. iWUE was calculated according to Linares and Camarero (2012):
Statistical Analysis
The carbon and oxygen isotope series of different individuals were analyzed using the numerical mix method to produce a mean isotope chronology via arithmetic average (Liu et al., 2012). To remove the impact of extreme values of individual series on the mean chorology, the individual carbon and oxygen isotope series were standardized by subtracting the long-term mean and dividing them by their standard deviation. The average δ13C and δ18O chronology for the six individuals of P. kesiya and five individuals of P. armandii were used to investigate the correlations between mean δ13C and δ18O chronologies in earlywood and latewood and climate data. The mean Pearson’s correlation coefficients among carbon and oxygen isotope series of tree individuals within each species (Rbar) were calculated to examine the common climate signal carried by individual series. The expressed population signal (EPS; Wigley et al., 1984) was also calculated to estimate the internal coherence of the individual tree-ring time series. Trees may use carbohydrates assimilated during the previous growing season and stored in wood parenchyma to form earlywood in the following year (carry-over effects). Hence, it is possible that climate not only affects plant growth in the same year, but also in the subsequent year. Thus, a period of 18-months from previous year’s July to current December was used for the calculation of correlation coefficients between monthly means of temperature, precipitation and RH and stable isotope variations to determine the climatic factors influencing stable isotope fractionation, and to test if stable isotope ratios in a tree ring were also influenced by previous year’s climate. Differences between EW and LW of δ13C and δ18O in the two species were tested for significance by using Independent samples t-test. We used the fraction model of Waterhouse et al. (2002) to reconstruct the earlywood and latewood growth period source water δ18O during the period 1986–2003, and compared these results with the weighted mean δ18O of precipitation at the nearest Global Network of Isotopes in Precipitation (GNIP) station, in Kunming (World Metrological Organization station #5677800; 25°02′ N, 102°43′ E, 1895 m a.s.l.).
Results
Characteristics of the Stable Carbon and Oxygen Isotope Chronologies
Both δ13CEW and δ13CLW of secondary forest pine P. kesiya showed similar mean values (Figure 3A, P = 0.989, Independent Samples t-test), while δ13CEW was higher than δ13CLW in natural forest pine P. armandii (Table 1; Figure 3B, P < 0.001). There was no significant difference between δ18OEW and δ18OLW in P. kesiya (Figure 3C, P = 0.224), while δ18OEW of P. armandii was higher than that of δ18OLW (Table 1; Figure 3D, P < 0.001). For δ13C, the inter-series correlations are higher in earlywood than latewood of both pine species. In contrast, δ18O showed higher inter-series correlations in latewood than earlywood for both pine species. There were significant correlations between δ13CEW and previous year’s δ13CLW in both pine species (Table 1). δ18OEW was positively correlated with previous year’s δ18OLW in P. armandii, while there were no significant correlations between δ18OEW and previous year’s δ18OLW in P. kesiya (Table 1). δ13CEW of P. kesiya vs. δ13CEW of P. armandii, and δ13CLW of P. kesiya vs. δ13CLW of P. armandii were not correlated between two pine species, whereas both δ18OEW and δ18OLW in P. kesiya were positively and significantly correlated with those of P. armandii, respectively (Table 2).
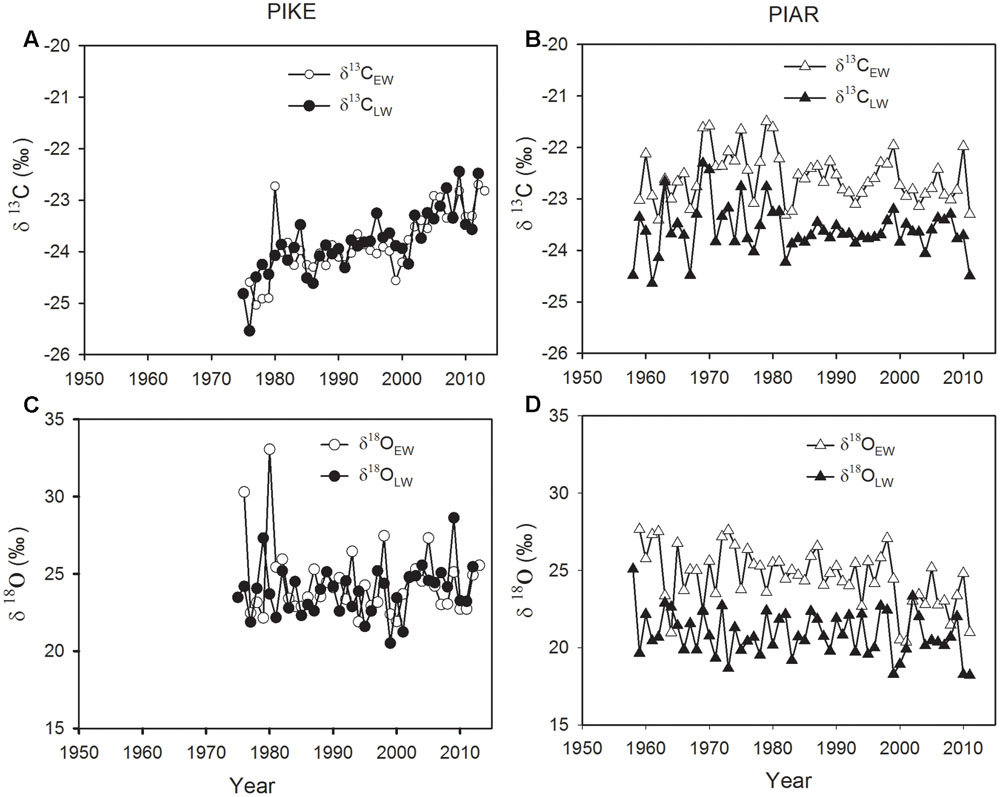
FIGURE 3. Carbon isotope variations in earlywood (δ13CEW, open circle) and latewood (δ13CLW, closed circle) of secondary forest pine P. kesiya (PIKE, A) and in earlywood (δ13CEW open triangle) and latewood (δ13CLW, closed triangle) of P. armandii (PIAR, B), and oxygen isotope variations in earlywood (δ18OEW, open circle) and latewood (δ18OLW, closed circle) of P. kesiya (C) and in earlywood (δ18OEW, open triangle) and latewood (δ18OEW, closed triangle) of forest pine P. armandii (D).
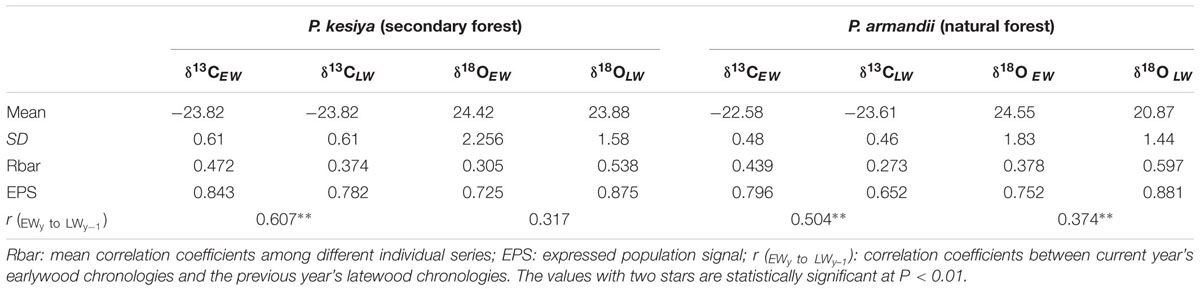
TABLE 1. Statistics of tree-ring carbon and oxygen isotope chronologies for Pinus kesiya (1975–2012) and Pinus armandii (1958–2011).
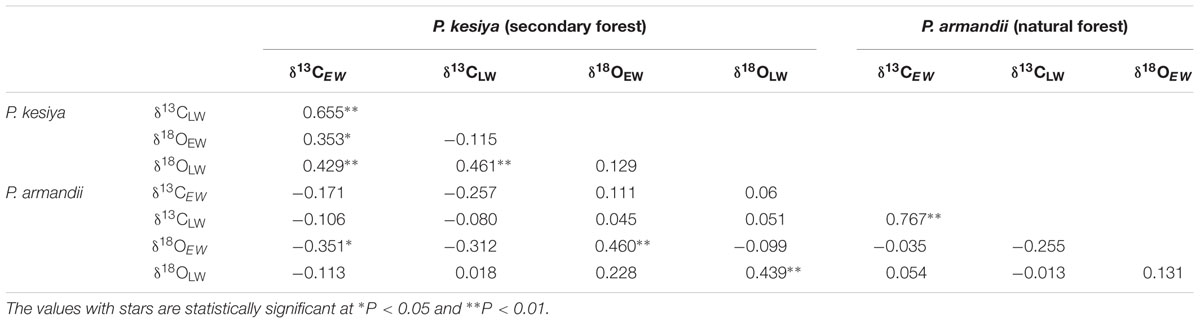
TABLE 2. Correlation coefficients between the mean carbon and oxygen isotope chronologies of secondary forest pine Pinus kesiya and natural forest pine Pinus armandii.
Despite of removing the impact of atmospheric δ13C on the tree ring δ13C, both δ13CEW and δ13CLW of P. kesiya showed an increasing trend (Figure 3A). In contrast, both δ13CEW and δ13CLW of P. armandii showed an increasing trend before 1980 and then remain stable after 1980 with reduced inter-annual variability (Figure 3B). The δ18OEW and δ18OLW in P. kesiya showed no significant trends (Figure 3C), however, δ18OEW in P. armandii displayed slightly decreasing trends (Figure 3D).
Intrinsic Water Use Efficiency
iWUE in earlywood and latewood of both pine species showed increasing trends from the 1960s to 2010s (Figures 4A,B). There was no significant difference in iWUE between earlywood and latewood in P. kesiya (P = 0.811, Figure 4A), while the iWUE in earlywood was consistently higher than that of latewood in P. armandii (Figure 4B, P < 0.001). The earlywood of P. armandii had higher iWUE than that of P. kesiya (P < 0.001). The mean intercellular CO2 concentration (ci) tended to increase with time in both pine species (Figure 4C), and ci in earlywood was consistently lower than that of latewood in P. armandii (Figure 4D, P < 0.001).
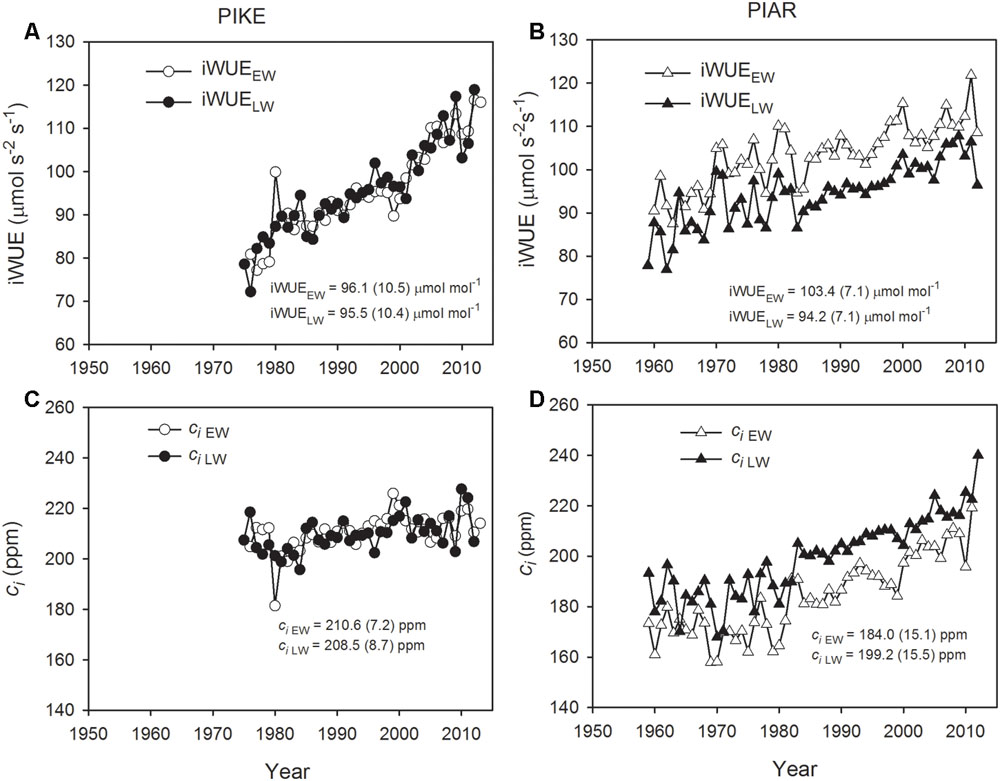
FIGURE 4. Intrinsic water use efficiency (iWUE) for earlywood (iWUEEW, open circle) and latewood of secondary forest pine P. kesiya (PIKE) (iWUELW, closed circle) (A), and for earlywood (iWUEEW, open triangle) and latewood of natural forest pine P. armandii (PIAR) (iWUELW, closed triangle) (B), intercellular CO2 concentration (ci) of earlywood (ciEW, open circle) and latewood of secondary forest pine P. kesiya (ciLW, closed circle) (C), and for earlywood (ci EW, open triangle) and latewood of natural forest pine Pinus armandii (ci LW, closed triangle) (D). Mean values and standard deviations of iWUE and ci series are indicated inside the panels.
Correlations of Stable Isotope Chronologies With Climate Factors
The δ13CEW of P. kesiya was positively correlated with monthly mean temperatures (MMT) of the previous year’s July and August, and current July to September (1975–2012), while δ13CLW of P. kesiya was positively correlated with MMT of previous July and December, and current January and July (Figure 5A). We found no significant correlations between δ13CEW and δ13CLW of P. kesiya and precipitation. The δ13CEW of P. armandii was positively correlated with MMT from February to May (1958–2011), and negatively correlated with monthly precipitation of current February (Figure 5B).
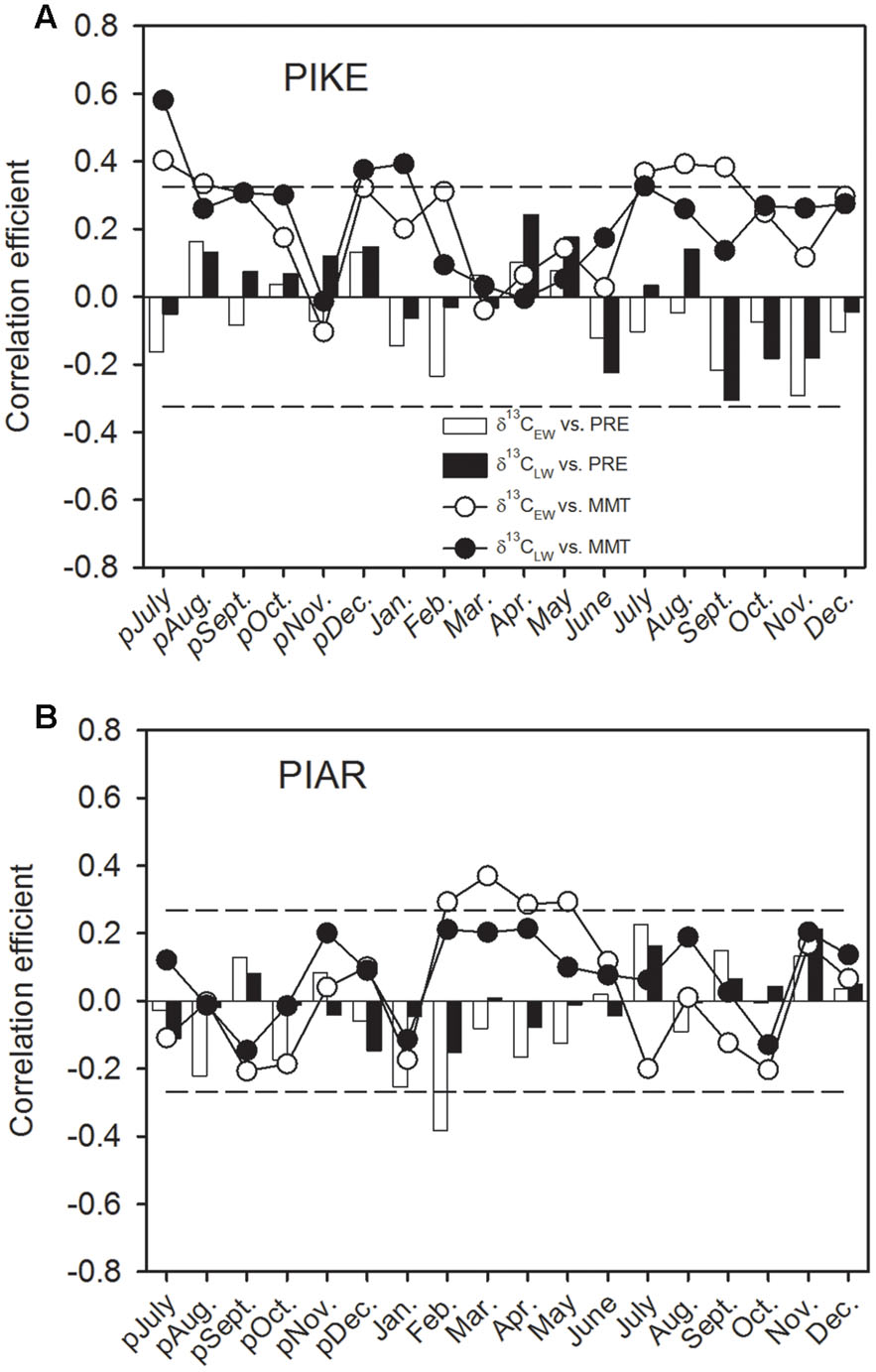
FIGURE 5. Correlations of carbon isotope in earlywood (δ13CEW) and latewood (δ13CLW) of secondary forest pine P. kesiya (PIKE) (1975-2012) (A), and natural forest pine P. armandii (PIAR) (1958-2011) (B) with monthly mean temperature (MMT, open circles for δ13CEW and closed circles for δ13CLW) and monthly precipitation (PRE, open bars for δ13CEW and closed bars for δ13CLW). The dashed lines indicated the 95% confidence intervals.
The δ18OEW of P. kesiya showed significantly negative correlation with previous October MMT (1975–2012), while δ18OLW of P. kesiya correlated negatively with current September, October, and November precipitation (Figure 6A). The δ18OEW of P. armandii was negatively correlated with MMT of previous year’s July to December, and current August and September, and was negatively correlated with previous year’s July and current May precipitation (1958–2011). The δ18OLW of P. armandii was negatively correlated with current September and October precipitation (Figure 6B).
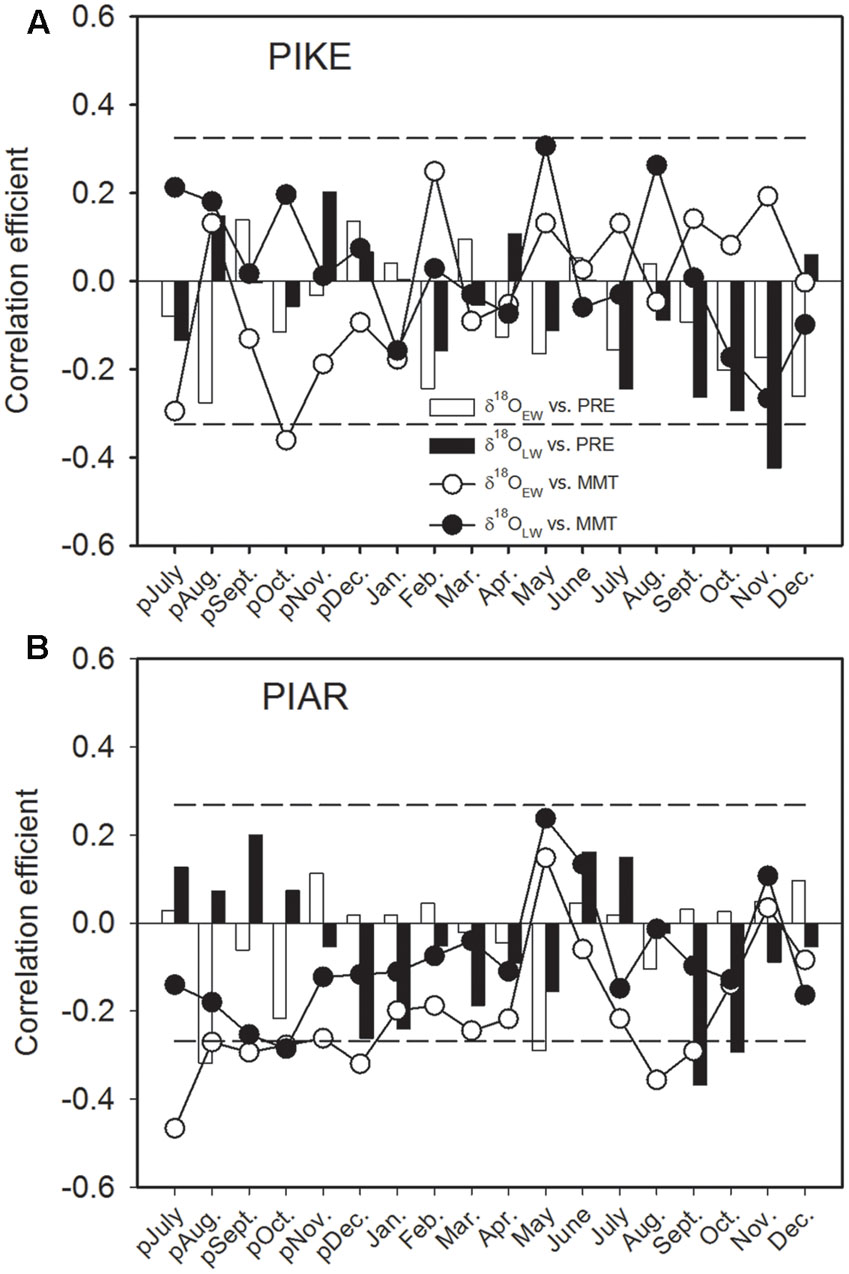
FIGURE 6. Correlations of oxygen isotope in earlywood (δ18OEW) and latewood (δ18OLW) of secondary forest pine P. kesiya (PIKE) (1975-2012) (A), and natural forest pine P. armandii (PIAR) (1958–2011) (B) with monthly mean temperature (MMT, open circles for δ18OEW and closed circles for δ18OLW) and monthly precipitation (PRE, open bars for δ18OEW and closed bars for δ18OLW). The dashed lines indicated the 95% confidence intervals.
The δ13CEW of P. kesiya was negatively correlated with November relative humidity (RH) (Figure 7A), while while δ13CEW of P. armandii was negatively correlated with previous year’s October and current June RH, and δ13CLW of P. armandii was negatively correlated with previous year’s September and current April RH (Figure 7B). The δ18OEW of both pine species were negatively correlated with May relative humidity (RH), while δ18OEW of P. armandii was positively with July RH (Figures 7C,D). δ18OLW of P. kesiya was negatively correlated with previous year’s July and November RH, and δ18OLW of P. armandii was negatively correlated with previous year’s July and August RH (Figures 7C,D).
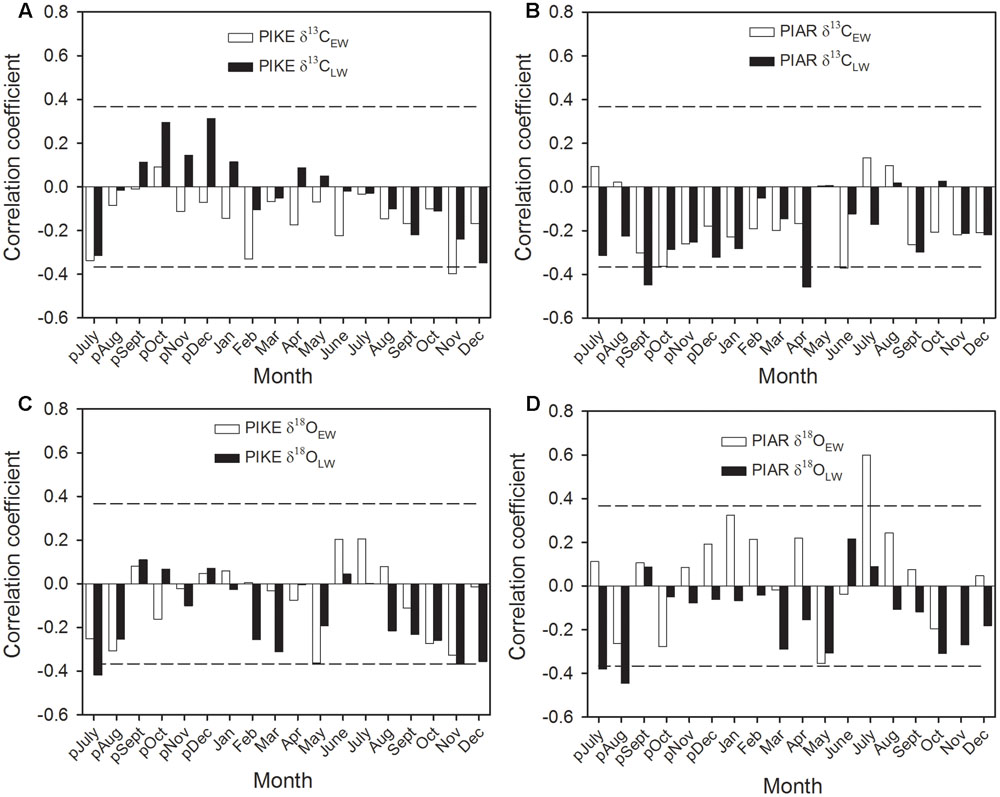
FIGURE 7. Correlations of carbon isotope in earlywood (δ13CEW, open bar) and latewood (δ13CLW, closed bar) of secondary forest pine P. kesiya (PIKE) (A), and natural forest pine P. armandii (PIAR) (B), oxygen isotope in earlywood (δ18OEW) and latewood (δ18OLW) of P. kesiya (PIKE) (C), and δ18OEW and δ18OLW of P. armandii (D) and relative humidity (RH) from Ailaoshan Station for Subtropical Forest Ecosystem Studies (ALS) (1982–2012). The dashed lines indicated the 95% confidence intervals.
Discussion
Intra-annual Variations of Carbon and Oxygen Isotopes
We found that δ13CEW are closely related with previous year’s δ13CLW in both secondary forest pine P. kesiya and natural forest pine P. armandii, indicating that there was a significant carbon carry-over effect, which is consistent with Krepkowski et al. (2013). However Kress et al. (2009) showed no carry-over effect for two tree-line conifers due to the limitation of carbohydrates caused by short growing season lengths. Because of the existence of carry over effect in both pine species in the present study, it is suggested to analysis earlywood and latewood carbon isotope separately (Kagawa et al., 2006). The carbon isotopes (δ13C) in both earlywood and latewood showed low correlations between P. kesiya and P. armandii (Table 2), indicating that the carbon isotope chronologies reflect mainly species-specific environmental signal.
Since carbon isotopes in tree rings were reported to be possibly affected by age effect, the innermost ca. 30 rings are usually excluded in the carbon isotope study (Schleser et al., 1999). The average age of P. kesiya at our site is 30 years., so the carbon isotope series of P. kesiya might potentially be affected by age effects. However, the canopy at our study site had a crown density of ca. 20% only, so air may ventilate freely between stems. Hence, there is no dense canopy that may possibly cause CO2 enrichment of 13C depleted air from soil respiration inside the stand, which is the most responsible factor for age effects in tree-ring 13C (Schleser et al., 1999; Dorado Liñán et al., 2012). Although we cannot completely exclude any possible age effects on carbon isotope ratios in P. kesiya, we consider this ‘age effects’ is negligible in our case.
The seasonal amplitude of δ18O is considerably higher in P. armandii (3.68‰) than P. kesiya (0.56‰) (Table 1; Figures 2C,D), but these amplitudes are lower than those found from high-altitude fir (Abies forrestii) in the nearby Yulong Snow Mountains, southwestern China (ca. 8.76‰, An et al., 2012) or from Fokienia hodginsii in subtropical lowland China (ca. 6.0‰; Xu et al., 2016). The seasonal amplitude of δ18O is related to rooting depth, growing season length and sampling resolution, and thus can be species- or site-dependent (Xu C. et al., 2014). Intraspecific δ18OEW was uncorrelated with δ18OLW for both pine species (Table 2), indicating that oxygen isotope variations in the earlywood and latewood represent different climate signals. Moreover, respective δ18OEW and δ18OLW of natural forest pine are positively correlated with those of secondary forest pine (Table 2), pointing to a strong common seasonal climatic forcing imprinted on oxygen isotope. Our results suggest that separating EW and LW δ18O may provide seasonally distinct climate information.
Species-Specific Differences in Water Use Efficiency
Our results showed that pine species from natural forest (P. armandii) had a slightly higher iWUE compared to pine species from a secondary forest (P. kesiya) (Figures 4A,B), indicating that the former could have a more conservative water use than the secondary forest pine species, which is consistent with our first hypothesis. Conservative water use strategy associated with higher iWUE and lower growth rates were also reported from tree species in East Africa (Gebrekirstos et al., 2011). We found that the earlywood of P. armandii had higher iWUE than that of latewood (Figure 4B), which could be due to the lower water status and reduced stomatal conductance in the early growing season (Qi et al., 2012). However, we found no significant difference between iWUE in earlywood and latewood in P. kesiya. One reason may be related to the regular resin harvesting in secondary forest pine which may reduce carbohydrates within the tree in the early growing season (Wu M. et al., 2015).
With the increase of atmospheric CO2, the intercellular CO2 (ci) and iWUE in both species have been increasing, which is consistent with previous studies (Xu Y. et al., 2014; Wu G. et al., 2015). The increasing rate of iWUE in secondary forest species (0.82 μmol mol-1 per ppm) is twice as high as that of forest pine (0.38 μmol mol-1 per ppm), and the increase rate of iWUE of P. kesiya is also considerably higher than those found from other coniferous species (Xu Y. et al., 2014).
Relationships between Intra-annual Stable Isotopes and Climate Factors
We found that inter-series correlations of earlywood δ13C were higher than those of latewood δ13C for both pine species, whereas δ18O showed higher inter-series correlations in latewood than earlywood for both pine species (Table 2). Our results indicated that both δ13C in earlywood and δ18O in latewood of the two pine species carried a stronger common signal that most likely related to climatic factors. Earlywood δ13C of both pine species mainly stored a temperature signal (Figure 5), while latewood δ18O recorded precipitation signal (Figure 6).
δ13CEW of P. kesiya was positively correlated with MMT of previous year’s July and current July to September (1975–2012), however, due to the impact of resin collection on P. kesiya, correlations between δ13C and climate should be interpreted with care. In contrast, δ13CEW of P. armandii was positively correlated with MMT from February to May (1958–2011). The spring season is characterized by increasing temperature as well as relatively dry conditions before the onset of the rainy summer monsoon season (Figure 1). Stomatal conductance and intercellular CO2 concentration would be reduced due to lower soil water status in the spring season, and there would be a weaker carboxylation discrimination against δ13C and resulting in positive correlations between δ13CEW of P. armandii and MMT in the early growing season. In contrast to earlywood, δ13CLW of P. armandii was not significantly correlated to any climate parameter, which was consistent with the fact that the inter-series correlations of latewood δ13C were lower than those from δ13CEW.
The present study showed that δ18O in both P. armandii and P. kesiya shared common moisture signals, that δ18OEW of both pine species was negatively correlated with May RH, and δ18OLW of both pine species was negatively correlated with the autumn (September to November) precipitation, which is consistent with our second hypothesis. As can be derived from dendrometer measurements nearby from our study site, cambium growth of P. kesiya last until October (Supplementary Figure S2). However, the biomass accumulation and cell wall thickening of latewood cells may even continue until November, which needs to be corroborated by wood anatomical studies. The δ18O in the earlywood and latewood of P. armandii represent pre-monsoon precipitation and late monsoon precipitation signals, respectively (Figure 6). Moreover, we found that the δ18O in the earlywood and latewood of both P. kesiya and P. armandii were negatively with the pre-monsoon (May) RH and autumn RH, respectively (Figures 7C,D), which were also convinced by other studies (An et al., 2012; Xu et al., 2016; Zeng et al., 2016). This suggests that a contrasting seasonal moisture signal carried by δ18O of earlywood and latewood. In the subtropical area of southwestern China, early vegetation period (April, May) is characterized by sunny conditions with little rainfall and low humidity, while ample monsoon precipitation and high humidity occurs during summer (Figure 1).
The correlations coefficients between reconstructed source water δ18O and the weighted mean δ18O of precipitation in Kunming were higher in earlywood growth period (EWG) than that of latewood period (LWG) for both pine species (Supplementary Figure S3), which were consistent with An et al. (2012), and the correlation coefficients was higher in EWG of P. armandii (r = 0.35) than that of P. kesiya (r = 0.25). However, the correlations between reconstructed δ18O and weighted mean of precipitation were much lower than other studies (Xu C. et al., 2014), which could be due to the long distance between Kunming station and our study sites as well as short time period of the available data (1986–2003).
Conclusion
We found a strong carry-over effect for δ13C in both secondary forest pine species P. kesiya and natural forest pine species P. armandii. In P. kesiya, there was no significant difference between earlywood and latewood for either δ13C or δ18O, however, both δ13C and δ18O in the earywood were significantly higher than those of latewood in P. armandii. Our results showed that P. armandii had slightly higher iWUE in than that of P. kesiya. Water use efficiency is increasing in both pine species, however, with a higher rate in P. kesiya. δ13C variations in earlywood and latewood differed among the studied species and represented a site-specific or species-specific climate signals. Due to the impact of resin collection on P. kesiya, care should be taken to interpret the correlations relationships between tree-ring δ13C and climate factors. In contrast to δ13C, δ18O variations showed strong coherence between the study species. δ18OEW of both pine species was negatively correlated with May RH, whereas δ18OLW of both species had a strong significant autumn precipitation signal and thus has the potential to reconstruct autumn precipitation in the study area. The contrasting climatic signals in δ18OEW and δ18OLW imply that separate analyses of earlywood and latewood are recommendable to derive clear climatic signals in δ18O series from conifer species growing in the mid-mountain zone of the summer monsoon region of southwestern China.
Author Contributions
AB, AG, Z-XF, and P-LF designed the experiment. P-LF, JG, and AG performed the experiment. P-LF and Z-XF analyzed the data. All authors contributed to the writing of the manuscript.
Funding
This work was supported by the Scientific Fund of Yunnan Province, China (2013FB081), National Natural Science Foundation of China (31300333, 31370496, U1502231), the Robert-Bosch Foundation (Stuttgart, Germany), the “Light of West China” Program and Outstanding Young Research Fund of Chinese Academy of Sciences (CAS).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Ms Roswitha Hoefner-Stich and Melanie Viehauser from the Institute of Geography, University of Erlangen-Nuremberg for their assistance in cellulose extraction and isotope analysis. We thank Shankar Panthi and the two reviewers for their valuable comments and suggestions on the manuscript. Meteorological data were provided by Ailaoshan Station for Subtropical Forest Ecosystem Studies and the National Meteorological Information Center of China.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.02050/full#supplementary-material
References
An, W., Liu, X., Leavitt, S. W., Ren, J., Sun, W., Wang, W., et al. (2012). Specific climatic signals recorded in earlywood and latewood δ18O of tree rings in southwestern China. Tellus B 64:18703. doi: 10.3402/tellusb.v64i0.18703
Armitage, F. B., and Burley, J. (1980). Pinus Kesiya Royle ex Gordon (syn. P. khasya Royle; P. insularis Endlicher). Oxford: Tropical forestry, Commonwealth Forestry Institute.
Brienen, R. J. W., Wanek, W., and Hietz, P. (2011). Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species. Trees 25, 103–113. doi: 10.1007/s00468-010-0474-1
Dorado Liñán, I., Gutierrez, E., Heinrich, I., Andreu-Hayles, L., Muntan, E., Campelo, F., et al. (2012). Age effects and climate response in trees: a multi-proxy tree-ring test in old-growth life stages. Eur. J. For. Res. 131, 933–944. doi: 10.1007/s10342-011-0566-5
Fang, J., Chen, A., Peng, C., Zhao, S., and Ci, L. (2001). Changes in forest biomass carbon storage in China between 1949 and 1998. Science 292, 2320–2322. doi: 10.1126/science.1058629
Farquhar, G. D., Oleary, M. H., and Berry, J. A. (1982). On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Aust. J. Plant Physiol. 9, 121–137. doi: 10.1111/j.1469-8137.2008.02518.x
Farquhar, G. D., von Caemmerer, S., and Berry, J. A. (1980). A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149, 78–90. doi: 10.1007/BF00386231
Fritts, H. C. (1976/1987). “Tree Rings and Climate,” inMethods of Dendrochronology, Vol. II and III, eds L. Kairiukstis, Z. Bednarz, and E. Feliksik (Warsaw: International Institute for Applied Systems Analysis and the Polish Academy of Sciences), 567.
Gebrekirstos, A., van Noordwijk, M., Neufeldt, H., and Mitlöhner, R. (2011). Relationships of stable carbon isotopes, plant water potential and growth: an approach to assess water use efficiency and growth strategies of dry land agroforestry species. Trees Struct. Funct. 25, 95–102. doi: 10.1007/s00468-010-0467-0
Gebrekirstos, A., Worbes, M., Teketay, D., Fetene, M., and Mitlohner, R. (2009). Stable carbon isotope ratios in tree rings of co-occurring species from semi-arid tropics in Africa: patterns, and climatic signals. Glob. Planet. Change 66, 253–260. doi: 10.1016/j.gloplacha.2009.01.002
Gessler, A., Ferrio, J. P., Hommel, R., Treydte, K., Werner, R. A., and Monson, R. K. (2014). Stable isotopes in tree rings: towards a mechanistic understanding of isotope fractionation and mixing processes from the leaves to the wood. Tree Physiol. 34, 796–818. doi: 10.1111/j.1365-3040.2006.01533.x
Kagawa, A., Sugimoto, A., and Maximov, T. C. (2006). (CO2)-C-13 pulse-labelling of photoassimilates reveals carbon allocation within and between tree rings. Plant Cell Environ. 29, 1571–1584. doi: 10.1111/j.1365-3040.2006.01533.x
Krepkowski, J., Gebrekirstos, A., Shibistova, O., and Brauning, A. (2013). Stable carbon isotope labeling reveals different carry-over effects between functional types of tropical trees in an Ethiopian mountain forest. New Phytol. 199, 431–440. doi: 10.1111/nph.12266
Kress, A., Young, G. H. F., Saurer, M., Loader, N. J., Siegwolf, R. T. W., and McCarroll, D. (2009). Stable isotope coherence in the earlywood and latewood of tree-line conifers. Chem. Geol. 268, 52–57. doi: 10.1016/j.chemgeo.2009.07.008
Laumer, W., Andreu, L., Helle, G., Schleser, G., Wieloch, T., and Wissel, H. (2009). A novel approach for the homogenization of cellulose to use micro-amounts for stable isotope analyses. Rapid Commun. Mass Spectrom. 23, 1934–1940. doi: 10.1002/rcm.4105
Leavitt, S. W., and Long, A. (1988). Stable carbon isotope chronologies from trees in the southwestern United States. Glob. Biogeochem. Cycle 2, 189–198. doi: 10.1371/journal.pone.0102545
Li, S. F., Su, J. R., Liu, W. D., Lang, X. D., Huang, X. B., Jia, C., et al. (2015). Changes in biomass carbon and soil organic carbon stocks following the conversion from a secondary coniferous forest to a pine secondary forest. PLoS ONE 10:e0135946. doi: 10.1371/journal.pone.0135946
Li, Z., Liu, W.-Z., Zhang, X.-Z., and Zheng, F.-L. (2009). Impacts of land use change and climate variability on hydrology in an agricultural catchment on the Loess Plateau of China. J. Hydrol. 377, 35–42. doi: 10.1016/j.jhydrol.2009.08.007
Linares, J. C., and Camarero, J. J. (2012). From pattern to process: linking intrinsic water-use efficiency to drought-induced forest decline. Glob. Change Biol. 18, 1000–1015. doi: 10.1111/j.1365-2486.2011.02566.x
Liu, X., Xu, G., Grießinger, J., An, W., Wang, W., Zeng, X., et al. (2014). A shift in cloud cover over the southeastern Tibetan Plateau since 1600: evidence from regional tree-ring δ18O and its linkages to tropical oceans. Quat. Sci. Rev. 88, 55–68. doi: 10.1016/j.quascirev.2014.01.009
Liu, Y., Wang, R. Y., Leavitt, S. W., Song, H. M., Linderholm, H. W., Li, Q., et al. (2012). Individual and pooled tree-ring stable-carbon isotope series in Chinese pine from the Nan Wutai region, China: common signal and climate relationships. Chem. Geol. 33, 17–26. doi: 10.1016/j.chemgeo.2012.08.008
Managave, S. R., Sheshshayee, M. S., Ramesh, R., Borgaonkar, H. P., Shah, S. K., and Bhattacharyya, A. (2011). Response of cellulose oxygen isotope values of teak trees in differing monsoon environments to monsoon rainfall. Dendrochronologia 29, 89–97. doi: 10.1016/j.dendro.2010.05.002
McCarroll, D., and Loader, N. J. (2004). Stable isotopes in tree rings. Quat. Sci. Rev. 23, 771–801. doi: 10.1016/j.quascirev.2003.06.017
Piao, S. L., Fang, J. Y., Ciais, P., Peylin, P., Huang, Y., Sitch, S., et al. (2009). The carbon balance of terrestrial ecosystems in China. Nature 458, 1009–1013. doi: 10.1038/nature07944
Qi, J. H., Zhang, Y. J., Zhang, Y. P., Liu, Y. H., Yang, Q. Y., Song, L., et al. (2012). Water-holding capacity of an evergreen broadleaf forest in Ailao Mountain and its functions in mitigating the effects of Southwest China drought. Acta Ecol. Sin. 32, 1692–1702. doi: 10.5846/stxb201103030259
Qin, C., Yang, B., Brauning, A., Griessginger, J., and Wernicke, J. (2015). Drought signals in tree-ring stable oxygen isotope series of Qilian juniper from the arid northeastern Tibetan Plateau. Glob. Planet. Change 125, 48–59. doi: 10.1016/j.gloplacha.2014.12.002
Rinn, F. (2003). TSAP-Win, Software for Tree-Ring Measurement, Analysis and Presentation. Heidelberg: Rinntech.
Schleser, G. H., Helle, G., Lucke, A., and Vos, H. (1999). Isotope signals as climate proxies: the role of transfer functions in the study of terrestrial archives. Quat. Sci. Rev. 18, 927–943. doi: 10.1016/S0277-3791(99)00006-2
Wang, Z., Calderon, M. M., and Carandang, M. G. (2006). Effects of resin tapping on optimal rotation age of pine secondary forest. J. For. Econ. 11, 245–260. doi: 10.1016/j.jfe.2005.10.001
Waterhouse, J. S., Switsur, V. R., Barker, A. C., Carter, A. H. C., and Robertson, I. (2002). Oxygen and hydrogen isotope ratios in tree rings: How well do models predict observed values? Earth Planet. Sci. Lett. 201, 421–430. doi: 10.1016/S0012-821X(02)00724-0
Wieloch, T., Helle, G., Heinrich, I., Voigt, M., and Schyma, P. (2011). A novel device for batch-wise isolation of α-cellulose from small-amount wholewood samples. Dendrochronologia 29, 115–117. doi: 10.1016/j.dendro.2010.08.008
Wigley, T. M. L., Briffa, K. R., and Jones, P. D. (1984). On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Climatol. 23, 201–213. doi: 10.1175/1520-04501984023<0201:OTAVOC>2.0.CO;2
Wu, G., Liu, X., Chen, T., Xu, G., Wang, W., Zeng, X., et al. (2015). Elevation-dependent variations of tree growth and intrinsic water-use efficiency in Schrenk spruce (Picea schrenkiana) in the western Tianshan Mountains, China. Front. Plant Sci. 6:309. doi: 10.3389/fpls.2015.00309
Wu, M., Pu, A., Zhou, R., Pan, Y., and Ye, J. (2015). The effect of resin tapping on tree growth of Pinus kesiya var. langbianensis. J. West China For. Sci. 44, 47–50.
Xu, C., Sano, M., and Nakatsuka, T. (2011). Tree ring cellulose δ18O of Fokienia hodginsii in northern Laos: a promising proxy to reconstruct ENSO? J. Geophys. Res. Atmos. 116:D24109. doi: 10.1029/2011JD016694
Xu, C., Sano, M., Yoshimura, K., and Nakatsuka, T. (2014). Oxygen isotopes as a valuable tool for measuring annual growth in tropical trees that lack distinct annual rings. Geochem. J. 48, 371–378. doi: 10.2343/geochemj.2.0312
Xu, C., Zheng, H., Nakatsuka, T., Sano, M., Li, Z., and Ge, J. (2016). Inter-and intra-annual tree-ring cellulose oxygen isotope variability in response to precipitation in Southeast China. Trees. 30, 785–794. doi: 10.1007/s00468-015-1320-2
Xu, Y., Li, W. J., Shao, X. M., Xu, Z. H., and Nugroho, P. (2014). Long-term trends in intrinsic water-use efficiency and growth of subtropical Pinus tabuliformis Carr. and Pinus taiwanensis Hayata in central China. J. Soils Sediments 14, 917–927. doi: 10.1007/s11368-014-0878-4
Young, S. S., and Wang, Z.-J. (1989). Comparison of secondary and natural forests in the Ailao Shan region of Yunnan, China. For. Ecol. Manage. 28, 281–300. doi: 10.1016/0378-1127(89)90008-X
Zeng, X. M., Liu, X. H., Evans, M. N., An, W. L., Xu, G. B., and Wu, G. J. (2016). Seasonal incursion of Indian Monsoon humidity and precipitation into the southeastern Qinghai–Tibetan Plateau inferred from tree ring δ18O values with intra-seasonal resolution. Earth Planet. Sci. Lett. 443, 9–19. doi: 10.1016/j.epsl.2016.03.011
Keywords: stable carbon isotope, stable oxygen isotopes, intrinsic water use efficiency, subtropical pine species, Asian summer monsoon, intra-annual resolution
Citation: Fu P-L, Grießinger J, Gebrekirstos A, Fan Z-X and Bräuning A (2017) Earlywood and Latewood Stable Carbon and Oxygen Isotope Variations in Two Pine Species in Southwestern China during the Recent Decades. Front. Plant Sci. 7:2050. doi: 10.3389/fpls.2016.02050
Received: 14 September 2016; Accepted: 21 December 2016;
Published: 10 January 2017.
Edited by:
Jian-Guo Huang, University of Chinese Academy of Sciences, ChinaReviewed by:
Bao Yang, Cold and Arid Regions Environmental and Engineering Research Institute (CAS), ChinaXiaohong Liu, Cold and Arid Regions Environmental and Engineering Research Institute (CAS), China
Copyright © 2017 Fu, Grießinger, Gebrekirstos, Fan and Bräuning. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ze-Xin Fan, ZmFuemV4aW5AeHRiZy5vcmcuY24= Achim Bräuning, YWNoaW0uYnJhZXVuaW5nQGZhdS5kZQ==