 Martina Lavrič
Martina Lavrič Klemen Eler2,3
Klemen Eler2,3 Jožica Gričar
Jožica Gričar- 1Department of Yield and Silviculture, Slovenian Forestry Institute, Ljubljana, Slovenia
- 2Department of Agronomy, Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia
- 3Department of Forest Ecology, Slovenian Forestry Institute, Ljubljana, Slovenia
Intra-annual variations in leaf development, radial growth, including the phloem part, and sap flow have rarely been studied in deciduous trees from drought-prone environments. In order to understand better the chronological order and temporal course of these processes, we monitored leaf phenology, xylem and phloem formation and sap flow in Quercus pubescens from abandoned karst grasslands in Slovenia during the growing season of 2014. We found that the initial earlywood vessel formation started before bud opening at the beginning of April. Buds started to open in the second half of April and full leaf unfolding occurred by the end of May. LAI values increased correspondingly with leaf development. About 28% of xylem and 22% of phloem annual increment were formed by the time of bud break. Initial earlywood vessels were fully lignified and ready for water transport, indicating that they are essential to provide hydraulic conductivity for axial water flow during leaf development. Sap flow became active and increasing contemporarily with leaf development and LAI values. Similar early spring patterns of xylem sap flow and LAI denoted that water transport in oaks broadly followed canopy leaf area development. In the initial 3 weeks of radial growth, phloem growth preceded that of xylem, indicating its priority over xylem at the beginning of the growing season. This may be related to the fact that after bud break, the developing foliage is a very large sink for carbohydrates but, at the same time, represents a small transpirational area. Whether the interdependence of the chronological sequence of the studied processes is fixed in Q. pubescens needs to be confirmed with more data and several years of analyses, although the ‘correct sequence’ of processes is essential for synchronized plant performance and response to environmental stress.
Introduction
Among all European regions, the Mediterranean appears to be the most vulnerable to climate change and subsequent increase in the frequency and intensity of extreme climatic events, such as drought and heat waves, which may create unprecedented climate-caused stress on trees and forest ecosystems (IPCC, 2014). In order to understand better the sensitivity and growth patterns of different tree species in natural adverse environments numerous studies have been carried out on this topic in recent years (e.g., Camarero et al., 2010; Nasr et al., 2011; Martinez del Castillo et al., 2016; Pérez-de-Lis et al., 2016b). Information about tree adaptation strategies and survival in marginal areas is crucial for our understanding of tree functioning and resilience, and is indispensable for future forest management (Tognetti et al., 2007; Hernandez-Santana et al., 2009).
Pubescent oak (Quercus pubescens Willd.), a deciduous ring-porous tree species, typically grows in a Mediterranean-type climate (Damesin and Rambal, 1995; Poyatos et al., 2005, 2007). It has developed various mechanisms and adaptations to survive in drought prone environments. Progressive stomatal closure, with a gradual and large decrease in water potential (Ψ) indicates its conservative water use. It decreases photosynthetic yield substantially during severe drought and shows efficient protection against high irradiance (Damesin and Rambal, 1995). Nevertheless, extreme drought events can lead to a severe decrease of water potential, loss of non-structural carbohydrates and loss of stem hydraulic conductivity, as shown by Nardini et al. (2016) for drought in the year 2012. Tognetti et al. (1998) studied the vulnerability of Q. pubescens to cavitation and estimated a water potential of -2.0 MPa as a threshold value causing 50% loss of water transport capacity (Ψ50). Such Ψ values can also be easily achieved in non-extreme dry summers (Nardini et al., 2016). This suggests that pubescent oak operates close to the point of xylem dysfunction. During summer, it protects against embolism by stomatal regulation which keeps water potentials above that causing hydraulic failure. Yet, stem conductivity can be affected also by frost-induced embolism, which first develops in large diameter vessels. According to Tyree and Cochard (1996) Quercus spp. suffer a high loss due to freezing and survival of the tree depends on the formation of a new ring of sapwood before leaf flush in spring.
In relation to xylem growth, the response of Q. pubescens to prolonged drought is reflected in reduced cambial cell production and, consequently, a lower proportion of latewood, as typical of ring-porous tree species (e.g., Panshin and de Zeeuw, 1980). Earlywood with, on the other hand, is more stable resulting in its smaller proportion in the case of wider xylem rings (Gričar et al., 2014). In addition, decreasing conduit size under drought, leading to a reduction in water-conducting capacity and a lower risk of cavitation, is a stress avoidance strategy of Q. pubescens (Eilmann et al., 2006; Zweifel et al., 2006). However, a reduction in the number of narrow latewood vessels might, in turn, cause its higher susceptibility to drought (Eilmann et al., 2009). In Slovenia, Q. pubescens is one of the prevailing tree species in the Karst region (Ferlan et al., 2016) and is ecologically important since it grows in forests that prevent degradation of vulnerable, shallow and erosion-prone soil (Brus, 2004). Information about its growth and survival strategies in this region therefore deserves more attention.
The radial stem growth of a tree is driven by the water status, which controls the metabolism of the entire tree, and carbon status, as a source of compounds for cambial activity and cell differentiation (Hsiao and Acevedo, 1974; Zweifel et al., 2006; Steppe et al., 2015). Due to their tight coupling, any changes in water-carbon relations (Zweifel et al., 2006) have an impact on xylem and phloem formation processes, affecting their ratios and specific structure (Hinckley and Lassoie, 1981; Gricar et al., 2015; Jyske and Hölttä, 2015; Sass-Klaassen et al., 2016). In this respect, intra-annual xylem and phloem formation analyses are crucial for a better understanding of the mechanisms underlying the environmental response of the xylem and phloem anatomy of different tree species. Information on intra-annual xylem growth of deciduous tree species growing in unfavorable growing conditions, such as Mediterranean areas, is scarce (González-González et al., 2013; Pérez-de-Lis et al., 2016a,b) although populations growing in such areas, together with rear-edge populations, are particularly under threat due to anticipated climate change (IPCC, 2014). Phloem observations in trees from sub-Mediterranean areas are even more rare and have been mainly performed on conifers (Gričar et al., 2016).
Intra-annual radial growth analyses are often coupled to leaf phenological observations to assess their linkage to photosynthesis and water transport (e.g., Michelot et al., 2012; Prislan et al., 2013; Copini et al., 2016; Kitin and Funada, 2016; Sass-Klaassen et al., 2016), since new photoassimilates transported in phloem sap are substrates used for radial growth increment (González-González et al., 2013). The relationship between these two developmental processes is therefore especially relevant in cambial reactivation, because expanding leaves are the main sources of auxin, which has a major role in cambial activity and cell expansion (Lachaud et al., 1999; Aloni, 2013). In Q. pubescens, xylem growth starts before bud burst and almost half of the xylem increment is formed at the time of leaf unfolding (Zweifel et al., 2006). Because of tylose formation in the major part of earlywood vessels of the previous xylem increments (Bréda and Granier, 1996), newly formed earlywood vessels are therefore essential to provide a water supply to the crown in a ring-porous Q. pubescens (Zweifel et al., 2006).
Since studies on leaf phenology, intra-annual radial growth and water relations in trees generally exclude the phloem part, we investigated these key processes (i.e., leaf phenology, xylem and phloem formation and sap flow) jointly in Q. pubescens trees from abandoned karst grasslands in Slovenia in 2014. In particular, three main aims were addressed: (1) to define the main milestones in the formation of xylem and phloem annual increments; (2) to compare the time course and interdependance of leaf development, phloem formation, xylem formation and sap flow measurements, with special attention to the beginning of the growing season and (3) to evaluate the obtained tree-anatomical and eco-physiological measurements in relation to local environmental conditions.
Materials and Methods
Study Site Characteristics
The study site was located in karst grassland at Podgorski Kras (45°32′56.3″N, 13°54′36.1″E, 429.4 m a.s.l.) in Slovenia, which was abandoned ca. 30 years ago from agricultural use after centuries of low intensity sheep and cattle grazing. Abandonment lead to encroachment of various woody plant species, among which pubescent oak dominates, growing in patches or solitary with an average height of 7 m and average age ca. 30–40 years.
The site is influenced by a sub-Mediterranean climate, characterized by fairly harsh winters, and dry and hot summers when drought periods frequently occur. The average annual temperature in the period 1981–2010 was 10.4°C, the average daily maximum 16.5°C and the average daily minimum 5.4°C. The lowest average temperature was recorded in January (1.3°C) and the highest in July (20.1°C). The average total annual precipitation was 1170 mm, with two annual rainfall peaks; in autumn and in late spring. Meteorological data for the region were obtained from the nearby Ilirska Bistrica climate station belonging to the Slovenian Environment Agency (ARSO).
The dominant soil type is rendzic with very uneven soil depth and rocky surface. The soil has mainly a clay texture with a low amount of plant nutrients, 12–15% proportion of organic matter in the top layer and pH value 6.9. Severe weather conditions, shallow soil and frequent wind diminish the impact of high precipitation, so water stress frequently occurs in the growing season (March–October) (Ferlan et al., 2011).
Weather Conditions in 2014
In 2014, daily data of meteorological parameters (air and soil) temperature (T), precipitation (P), vapor pressure deficit (VPD), solar radiation (Rg) and soil water content (SWC) measured at half-hourly intervals were obtained from a micrometeorological tower in the close vicinity of the research plot (Ferlan et al., 2011). The year 2014 was unusually wet, exceeding the long-term average of sum of precipitation by about 50% (Table 1 and Figure 1). A more detailed description of the measured environmental parameters at the study site in 2014 is shown in Table 1.

TABLE 1. Detailed description of the measured environmental parameters at the study site in 2014.
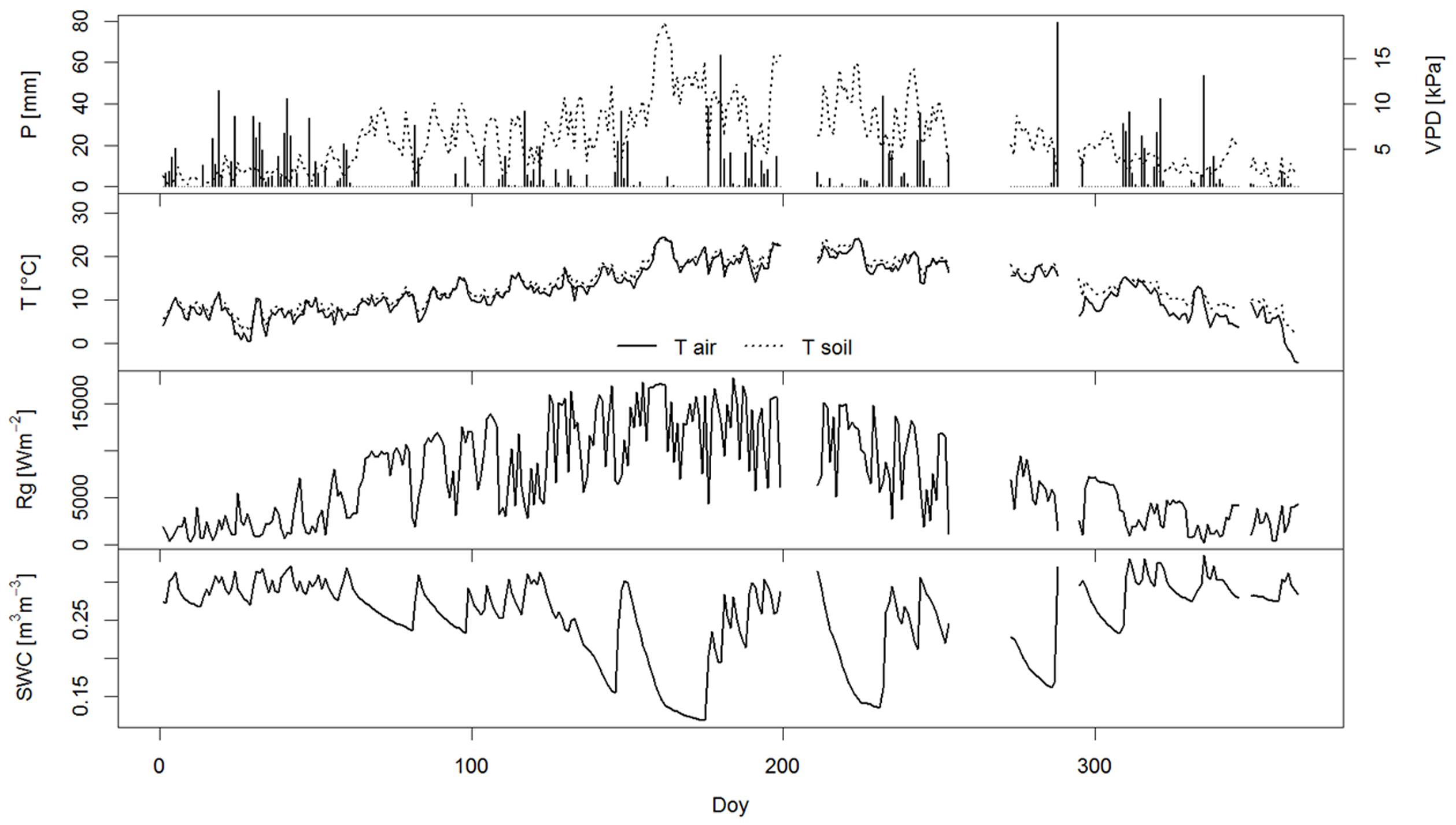
FIGURE 1. Measured meteorological parameters at the study site Podgorski Kras in 2014, including daily sum of precipitation (P), daily sum of vapor pressure deficit (VPD), mean daily air and soil temperature (T), daily sum of sum solar radiation (Rg) and mean daily soil water content (SWC).
Tree Selection and Leaf Phenological Observations
At the beginning of the 2014 growing season, we randomly selected six solitary healthy pubescent oaks (Quercus pubescens Willd.) without any visible injuries on stems and roots. The trees were 35 ± 5 years old and 7 ± 0.5 m high, with diameters at breast height (DBH) of 0.18 ± 0.015 m. The selected oaks were equipped for sap flow measurements and intra-annual microcore sampling was performed in the period March–October of 2014.
Leaf phenological observations and leaf area index (LAI) measurements were carried out on all oaks in the first part of the growing season, i.e., March–July. On the northern side of each tree, one branch was selected for phenological observations. We focused on phenological phase leaf unfolding, which was divided into five subclasses: 1 – buds are fully closed, 2 – buds are open (leaf appearance), 3 – the beginning of leaf unfolding (33–66% leaf development), 4 – leaf unfolding throughout the canopy (66–99% leaf development), 5 – full leaf unfolding. We took weekly images of a selected part in the crown with a digital camera to document leaf development.
Leaf Area Index Measurements
In the period March–July, we measured the LAI, i.e., one-sided leaf surface area per ground surface area (m2m-2) with a LAI-2200 Plant Canopy Analyzer (LI-COR Inc., USA). We performed measurements at weekly intervals following the protocol for measuring LAI of isolated trees (LAI-2200, 2012). At each tree, four “below canopy” readings were taken in N, E, S, and W orientations, placing the sensor equipped with a 270° view restrictor next to the trunk. Data of gap fraction measurements were analyzed with FV2200 software, version 2.0.0 (LI-COR Inc., USA) in which the average LAI values for each tree were calculated.
Sap Flow Measurements
At the beginning of the growing season (i.e., first decade of April), the selected oaks were equipped with SFM1 Sap Flow Meters based on the Heat Ratio Method (SFM1 instrument, ICT International, Australia). According to the protocol described in Burgess and Downey (2014), SFM1 were installed on the north side of the tree, 1 ± 0.1 m above the ground, with a 0.5 mm distance between the temperature probes and the heater. Data were stored continuously at 10-min intervals in binary notation and monthly downloaded in Sap Flow Tool software (ICT International, 2014). In the software, data were analyzed using correction factors (e.g., stem diameter, bark thickness, and sapwood depth), which were acquired at the end of the measurement campaign (Pfautsch et al., 2010; Burgess and Downey, 2014). The average sapwood width of pubescent oaks was 2.13 ± 0.6 cm, so only data of the outer temperature thermistor located in the conducting wood were applied. The inner temperature thermistor was already in hardwood, in which water transport is interrupted due to the presence of tyloses in the large earlywood vessels (Cochard and Tyree, 1990). These data were not therefore used for analyses. For determination of the actual sap flux density, zero flow values were calculated for each tree from the data obtained from the period before leaf unfolding, i.e., DOY 1–112 (Do and Rocheteau, 2002; Vandegehuchte and Steppe, 2013). Due to power supply problems, a few gaps in SF data occurred (12% data loss) (Figure 1).
Microcore Collection, Section Preparation and Tissue Analyses
Microcores were collected at weekly intervals from March until August of 2014, using a Trephor tool (Rossi et al., 2006). Additionally, microcores were collected in mid-October, when phloem and xylem rings were fully developed. The samples were collected at 0.7–1.7 m above the ground following a spiral up the stem. In order to avoid wound effects, sampling locations were separated by 5 cm. Each microcore contained phloem (non-collapsed and collapsed), cambium and at least two of the last-formed xylem rings. Immediately after removal from the trees, the samples were fixed in ethanol-formalin acetic acid solution (FAA) (Gričar et al., 2007). After 1 week, the samples were dehydrated in a graded series of ethanol, infiltrated with D-limonene (Bio Clear, Bio Optica, Milano, Italy) and embedded in paraffin blocks (Paraplast plus, ROTH, Karlsruhe, Germany) (Rossi et al., 2006). Transverse sections of 8–12 μm thickness were cut with a Leica RM 2245 rotary microtome (Leica Microsystems, Wetzlar, Germany). The sections were stained with a safranin (Merck, Darmstadt, Germany) (0.04%) and astra blue (Sigma–Aldrich, Steinheim, Germany) (0.15%) water mixture (van der Werf et al., 2007) and mounted in Euparal (Waldeck, Münster, Germany). They were observed under an Olympus BX51 light microscope (Tokyo, Japan) using transmission and polarized light modes. Histometrical analyses (i.e., determination of phenological phases of cambial activity and cell differentiation, and increment measurements) were performed with the Nikon NIS-Elements Basic Research v.2.3 image analysis system (Tokyo, Japan).
On the sections, the number of cell layers in the cambium was counted, as well as the widths of currently formed xylem, and phloem increments along three radial files were measured and then averaged. Additionally, the following phenological phases of cambial activity were assessed: (1) onset of cambial cell production, (2) maximum rate of xylem and phloem cell production, (3) cessation of cambial cell production, (4) cessation of the differentiation process in terminal latewood tracheids and (5) transition from earlywood to latewood and from early to late phloem. Phases were assessed for each tree and were computed in days of the year (DOY).
The phenological phases of cambial activity and cell differentiation listed above were identified and interpreted within the context of the multi-seriate concept that the vascular cambium comprises both the cambial initial cells and the xylem and phloem mother cells (Plomion et al., 2001). The observed xylem and wood formation phases were defined as follows: (1) the onset of cambial cell production was identified as an increased number of thin-walled cambial derivatives (Prislan et al., 2011); (2) the maximum rate of new xylem and phloem cell production was calculated and plotted using a Gompertz function that indicated the maximum growth rate at the inflection point of the curve (Rossi et al., 2003); (3) cessation of cambial cell production was identified as the time at which no new thin-walled cells were observed adjacent to the cambium and the number of cambial cells was comparable to the number before its reactivation in spring; (4) cessation of xylem differentiation was identified by the complete lignification of the terminal latewood cells, as indicated by cell walls stained completely red by the safranin-astra blue procedure; (5) transition from earlywood to latewood was determined when small latewood vessels were no longer arranged in rings, which is characteristic of large earlywood vessels (Gričar, 2010); (6) transition from early to late phloem was identified by the appearance of small, tangentially orientated groups of phloem fibers separating the two parts (Srivastava, 1964; Gričar, 2010).
Data Analysis
Analyses were performed using the statistical software R, version 3.2.3 (R Core Team, 2015). Data were displayed as time-series and analyzed with descriptive statistics. The widths of xylem and phloem increments were fitted to the Gompertz function (Rossi et al., 2003) using the nlme package in R. Model fits were evaluated by computing pseudo-R2 by comparing residual variance of the full model against the residual variance of an intercept-only null model. The first derivative of the Gompertz function was calculated to determine maximum cambial cell production of xylem and phloem rings during the growing season.
Results
Leaf Phenology and LAI Measurements
Leaf phenological observations were performed from March until July, with the main focus on early spring leaf development. Typical of many Quercus sp. are marcescent leaves that are dead and retained through the winter (Sánchez de Dios et al., 2009); consequently, there was still a small proportion of old leaves from the previous year attached to the branches of the pubescent oak in early spring, possibly increasing our LAI values. Buds were fully closed until the second half of April, when they started to open (DOY 110.5 ± 5.5), followed by the appearance of the first leaves. By the end of April, half of the trees’ canopy was covered with leaves. Full leaf unfolding occurred by mid-May (DOY 131.5 ± 5.5). Thus, in 2014 the period from bud opening to full leaf unfolding lasted on average 21 days (Figure 3). LAI values increased correspondingly with leaf development. When the first leaves appeared in mid-April, LAI values were 0.27 ± 0.1 m2m-2 and reached an average maximum of 2.02 m2m-2 at the beginning of June, 3 weeks after full leaf development. Maximum LAI values varied among the trees; for example, in three individuals LAI was 3.45 ± 0.4 m2m-2, while in one oak only 0.86 m2m-2.
Seasonal Dynamics of Xylem and Phloem Formation
In the third week of March, when we started the microcore sampling, cell division in the cambium had just started, as inferred from the increased number of cambial cells (7–8 cell layers) compared to that of the dormant cambium (4–5 cell layers) at the end of the growing season in October. In addition, in some of the trees, initial sieve tubes of early phloem started to expand adjacent to the cambium (16.1 ± 3.8 μm). In the first 3 weeks of radial growth, phloem growth exceeded that of xylem but from 8 April until the cessation of cambial cell production, xylem growth surpassed that of phloem. By the third week of June, earlywood vessels were fully created (Figure 2a). The average duration of the earlywood vessel enlargement period was 46 ± 4 days and of earlywood vessel formation 89 ± 11 days. Latewood started to form in the third week of May (Figure 2b), while the entire earlywood part was fully formed in the second half of June. In 2014, Q. pubescens generally developed one to two rows of earlywood vessels, with the largest ones being mostly located in the first row, at the growth ring boundary (Figure 2b). The transition from early to late phloem occurred at the end of May, as evident from the formation of the groups’ phloem fibers (Figure 2c). The peak of cambial production of xylem cells occurred at the beginning of May (DOY 125.6), when on average 8.7 μm of xylem tissue was formed per day, as calculated from the first derivative of the Gompertz function (Figure 3). At that time, 25% of the annual xylem increment was formed. For phloem, maximum cambial cell production (2.9 μm per day) took place 2 weeks later than in the case of xylem. At that time, 40% of the annual phloem increment was formed. At the height of cambial cell production, the cambium was more than 10 cell layers wide, with the number gradually decreasing thereafter. At the beginning of August, cambial cell production stopped simultaneously on both sides. In 2014, the average width of the completed xylem increment was 1324 ± 440 μm (min = 889 μm; max = 2311 μm) and of phloem 337 ± 69 μm (min = 230 μm; max = 482 μm) (Figure 2d). Earlywood and early phloem parts occupied 36.8 and 40.5%, respectively, of the annual growth increments.
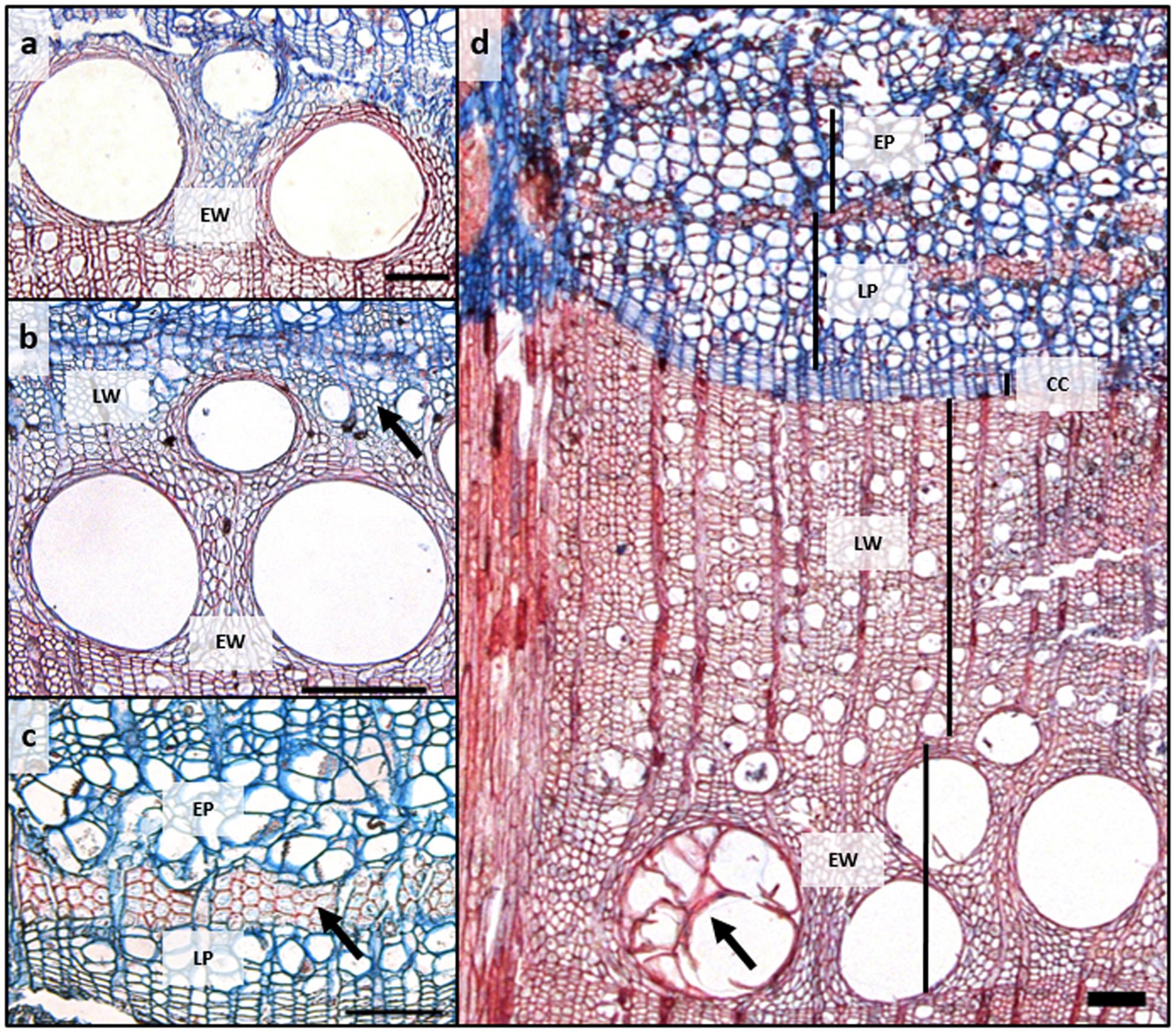
FIGURE 2. (a) Fully developed first ring of earlywood vessels; (b) Transition from early to latewood (black arrow); (c) Transition from early to late phloem (black arrow) and (d) Fully formed xylem and phloem increments in Quercus pubescens in 2014. Black arrow denotes earlywood vessel filled with tyloses. CC, cambium; EW, earlywood; LW, latewood; EP, early phloem; LP, late phloem, Bars = 100 μm.
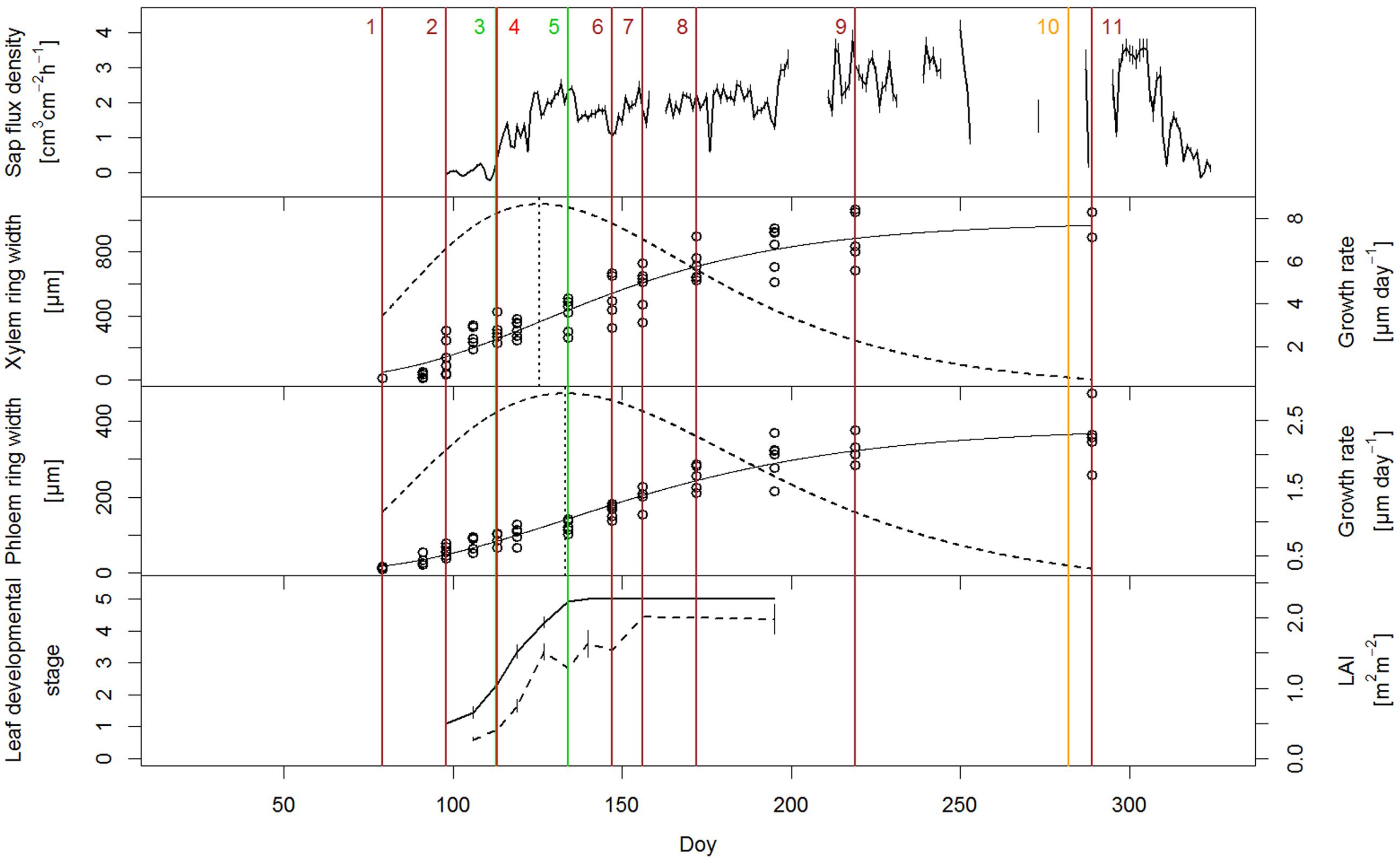
FIGURE 3. Relationship between various measured parameters (sap flow density, seasonal dynamics of xylem and phloem growth ring formation and leaf development) in Quercus pubescens during the growing season of 2014. Xylem and phloem development are presented as cumulative growth (solid line) and as weekly increment growth (dashed line). Leaf development is presented as phenological stages (solid line) and LAI measurements (dashed line). The most important milestones of the analyzed parameters are indicated with numbers: (1) onset of cambial cell production (DOY 79); (2) onset of expansion of initial earlywood vessels (DOY 98); (3) full bud opening (leaf appearance) (DOY 113); (4) complete development of initial earlywood vessels (DOY 113); (5) full leaf unfolding (DOY 134); (6) transition from earlywood to latewood (DOY 147); (7) transition from early phloem to late phloem and reaching peak of LAI (DOY 156); (8) complete development of earlywood (DOY 172); (9) end of cambial cell production (DOY 219); (10) autumn leaf senescence (DOY 282), and (11) cessation of xylem growth ring formation (DOY 289).
Sap Flow
Before bud break, we observed minimum sap flux density, which reached a daily average of 0.012 cm3cm-2h-1. Sap flux density and leaf unfolding showed a linear relationship in the period from milestone (3) to (5), as illustrated in Figure 3, whereas later, in the late stage of leaf development, a decrease of sap-flow was observed due to reduced transpirational demand (rain period).
The average sap flux density in the summer season when the canopy was fully developed but non-senescent (from DOY 134 to 280) was 2.19 cm3cm-2h-1. Fairly high fluctuations were found. These variations could largely be explained by precipitation, soil water availability and fluctuating water vapor pressure deficit. Previous research at the same site has shown that ecosystem evapotranspiration was severely reduced at a SWC lower than 0.145 m3m-3, which indicates a reduction of stomatal conductivity at the stand level (Ferlan et al., 2011). When sap-flow data of Q. pubescens were filtered for days with higher SWC, 46.2 and 37.0% of the variability of sap-flow density could be explained by water vapor deficit and Rg, respectively. The highest mean value of sap flux density was measured at the end of August: 8.65 cm3cm-2h-1. High densities persisted in part of November and then decreased in December, reaching early spring values at the end of the year. The drop in values coincided with the mass autumnal brown coloring of the leaves on trees.
Time Course of Leaf Development, sap Flow and Seasonal Radial Stem Growth in the First Part of the Growing Season
The expansion of the initial earlywood vessels already started at the beginning of April, when buds were still closed (Figure 3). At that time, no sap flow was recorded; its value was below 1 cm3cm-2h-1. By mid-April, when the buds were still closed, about 28% of the xylem and 22% of phloem annual increment were formed. In the last decade of April, buds started to open and the first leaves appeared. At the same time, initial earlywood vessels were fully lignified and thus ready for water transport. Sap flow became active and was contemporarily increasing with leaf development and LAI values. In the meantime, about one third of the xylem and one fourth of the phloem annual increments were created. Full leaf unfolding occurred in mid-May, when about 40% and 33% of xylem and phloem increments, respectively, had formed. In the period of leaf development (DOY 113–134) average sap flux density was 1.60 cm3cm-2h-1. Transition from earlywood to latewood occurred 2 weeks after full leaf development. Transition from early to late phloem, which took place a week later, coincided with the end of LAI increase.
Discussion
Synchronicity of Leaf Development and Increase of LAI Values
The increase in LAI values in the period from mid-April till mid-May corresponded to phenophases of leaf unfolding Furthermore, LAI values continued to rise for about 3 weeks after full leaf development, indicating canopy extension and the development of new shoots (height growth). Average LAI values of our Q. pubescens trees were lower (2.2 m2m-2) than of some other Quercus sp. from the literature, ranging from 3 to 7 m2m-2 (Bréda and Granier, 1996; Jonard et al., 2011; Burner et al., 2014). However, our measurements were in line with LAI values of Quercus robur stands (2.26 m2m-2) (Bequet et al., 2012). Since LAI is an indicator of soil fertility and/or stand productivity (Bréda and Granier, 1996) and is spatially highly variable (Bequet et al., 2012), differences in all values in different studies can be attributed to all these factors, as well as to differences in tree density, because our study was carried out on solitary oaks. Finally, environmental conditions also highly influence leaf development and LAI values (Mooney, 1972).
Seasonal Dynamics of Xylem and Phloem Formation
The main growth period (i.e., leaf expansion and phloem and xylem growth) was April–June, which is very similar to other species (e.g., González-González et al., 2013; Prislan et al., 2013). After that period, carbon is still assimilated, partly used for secondary wall biosynthesis but mostly for below-ground growth (Barbaroux et al., 2003) or storage reserves (Hoch et al., 2003). The transition from early to late phloem occurred after transition from earlywood to latewood. The transition from early to late phloem is associated with changes in photosynthetic allocation. Early in the growing season, a large proportion of the photosynthates is used for respiration, cell division and differentiation, and biosynthesis. As the rate of cell division slows, the principal sink for photosynthate shifts to storage in wood parenchyma, both above and below ground (Taiz and Zeiger, 2006). In addition, the second part of the growing season is important for storage of carbohydrate resources (in parenchyma) (Barbaroux and Bréda, 2002).
The phloem increment represented one fourth of the annual radial increment in 2014 (1.7 ± 0.5 mm). The xylem increment thus predominated, which is typical of productive oaks having a xylem ring wider than 0.75 mm (Gričar et al., 2014). Xylem and phloem ring widths, as well as proportions between xylem and phloem increments, and early and late parts of xylem and phloem of Q. pubescens, are comparable to measurements in Q. robur from lowland forests, suggesting that they might be general patterns for ring-porous oaks (Gričar et al., 2014). Under drought, Q. pubescens mainly reduces the latewood proportion, whereas earlywood remains more or less constant (Eilmann et al., 2009). Large earlywood vessels are most important in terms of water-conductivity, since the bulk water transport in oak occurs in earlywood of the youngest xylem rings (Ellmore and Ewers, 1985). Each spring, therefore, at least one new tangential row of earlywood vessels is formed in order to regenerate its maximum conductivity every spring, which is a successful strategy in summer-dry climates (Eilmann et al., 2009). Small vessels and tracheids in latewood, on the other hand, which are formed in the summer months, are also important for water transport because they represent a safety component of the xylem in ring-porous species. Their role is negligible when conditions are favorable but is critical in the case of cavitation of earlywood vessels (Granier et al., 1994).
Linkage between Leaf Unfolding and Radial Growth
The timing of vessel formation and leaf phenology differs among Quercus sp. (Suzuki et al., 1996; Sass-Klaassen et al., 2011; Takahashi et al., 2013), especially when comparing temperate and sub-Mediterranean oaks (González-González et al., 2013; Pérez-de-Lis et al., 2016b). We confirmed the observations of Zweifel et al. (2006) that a substantial amount of current xylem increment in Q. pubescens is formed at the time of bud burst and full leaf unfolding (28 and 40%, respectively). Since tyloses in large earlywood vessels of the previous growth rings impede water transport in ring-porous Q. pubescens (Cochard and Tyree, 1990), hydraulic conductivity is strongly reduced in early spring (Bréda and Granier, 1996). Initial earlywood vessels formed before bud break are therefore essential in order to provide a water supply to the young transpiring leaves in the crown (Zweifel et al., 2006). This pattern demonstrates that xylem tissue formed before full leaf development relies on carbohydrates stored during the previous growing season, particularly during the summer and autumn of the previous year (Barbaroux and Bréda, 2002).
With few exceptions (e.g., Prislan et al., 2013), the phloem part has been largely overlooked in studies on leaf phenology and intra-annual radial growth, despite its main role in the transport of carbohydrates from photosynthetic and storage tissues (source) to areas of active growth and metabolism (sinks) (Oparka and Robert, 1999). We found that at the time of bud burst and full leaf unfolding, 22 and 33%, respectively, of the early phloem part was formed, mainly consisting of large sieve tubes. In addition, in the initial 3 weeks of radial growth, phloem growth preceded xylem growth, as already previously observed in Q. petraea (Gričar, 2010). Rapid production of phloem indicates its priority over xylem at the beginning of the growing season, which may be related to the fact that, shortly after the beginning of leaf development, the developing foliage is a very large sink for carbohydrates but, at the same time, represents a small transpirational area. It can also be presumed that the hydraulic conductivity and water storage of sapwood developed in preceding years efficiently support the water balance of leaves at this, very early stage of development. On the other hand, one might expect that existing phloem connections might limit carbohydrate allocation at the very start of the season. Sieve tubes function for only one to two growing seasons and late phloem cells may remain functional until new phloem is produced in spring (Zamski and Zimmermann, 1979). Furthermore, overwintering undifferentiated cambial derivatives may differentiate into sieve elements before new elements are formed by the dividing cambium (e.g., Derr and Evert, 1967). Thus, newly formed phloem cells are important to link the storage tissues to developing leaves, which require photosynthates for respiration and biosynthesis in spring (Barbaroux et al., 2003).
Not much is known about the phloem formation patterns of deciduous trees in sub-Mediterranean areas, especially species with marcescent leaves. These leaves are presumed to be dead and retained on a tree through the winter (Sánchez de Dios et al., 2009). Abadía et al. (1996) demonstrated that marcescent leaves from the upper part of the tree crown of Q. subpyrenaica, unlike senescent leaves from the lower part of the crown, are capable of some photosynthetic activity in the last 1–2 months of the growing season (September and October), which significantly increases the possibility of carbon assimilation during the period when high light intensities and mild temperatures may still occur. Whether marcescent behavior of Q. pubescens affects carbohydrate reserves and thus also radial growth in the spring of the following growing season remains to be examined.
Sap Flow With Respect to Leaf Phenology and Environmental Conditions
Early spring patterns of xylem sap flow and LAI were similar, indicating that water transport in the oaks broadly followed canopy leaf area development. A similar relation was found for Quercus prinus by Wullschleger et al. (2001). Sap flow is in general very well correlated with leaf area, with an almost linear relationship (Vertessy et al., 1995). After the full development of foliage, transpiration and sap-flow are mainly governed by climatic/weather (precipitation) variability, causing changes in SWC, vapor pressure deficit and light. A study by Wullschleger et al. (2001) showed that, within the season, 85% of daily sap flow variability in Q. prinus can be explained by radiation, vapor pressure deficit and fractional LAI. Meteorological network data and our measurements of environmental parameters in the field (Figure 1) showed that 2014 was atypically wet. The sum of precipitation and number of rainy days in summer exceeded the 10-year average by 35 and 33%, respectively. Sharp decreases of sap flux density during the season can therefore be attributed to reduced transpirational demand and low light conditions during wet days. During a short dry and hot early summer period, in mid-June SWC decreased below SWCcritical (0.145 m3m-3, Ferlan et al., 2016) and sap flux density reached only medium values (1.89 cm3cm-2h-1), despite the Rg being high. At the same time, (June 21) leaf water potential measurements revealed a Ψmidday value of -2.67 ± 0.13 MPa (Vodnik et al., unpublished data). It can be concluded that the transpiration flow of the pubescent oak was reduced by substantial stomatal closure induced by low water potential and high VPD. This can be supported by the study of Nardini et al. (2016), who report that Q. pubescens leaf conductance to water vapor decreased by 85% when comparing a period with ample of water (Ψmidday = -1.5 MPa) and an extremely dry period (Ψmidday = -3.5 MPa).
Is the Chronologial Sequence of the Studied Patterns (Leaf Development, LAI, Radial Growth and Sap Flow) Fixed in Q. pubescens?
Understanding tree phenology, growth patterns and physiology is crucial to better understanding how trees cope with environmental changes (Granier et al., 1994; Sass-Klaassen et al., 2016). With Q. pubescens, a substantial amount of current xylem increment is formed before leaf unfolding, in order to provide essential hydraulic conductivity for axial water flow during leaf development, which is in line with the findings of Zweifel et al. (2006) for the same species. Comparing intra-annual xylem and phloem formation with other data (Gričar, 2010; Gričar et al., 2014) suggests that these might be general radial growth patterns for ring-porous oaks.
However, based on only 1-year of observation it is too speculative to conclude the interdependence of the chronological sequence of the studied processes (leaf development, LAI, xylem and phloem formation and sap flow) in Q. pubescens. Data of several growing seasons, including years with extreme events typical for this site (e.g., drought, fires) will reveal to what extent the time course of the studied variables is fixed and species-specific and how much it is influenced by environmental factors. After all, the ‘correct sequence’ of development of leaf unfolding, phloem and xylem growth and others, such as root growth, are essential for synchronized plant performance and response to environmental stress.
Author Contributions
Conception or design of the work: DV, MF, and KE; Data collection: ML, DV, and KE; Data analysis and interpretation: all authors; Drafting the article: JG, ML, DV, and KE; Critical revision of the article: all authors; Final approval of the version to be published: all authors.
Funding
This work was supported by the Slovenian Research Agency, young researchers’ program (Martina Lavrič), programs P4-0085 and P4-0107, project J4-7203, and by the FP7 Capacities project EUFORINNO (REGPOT no. 315985).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors gratefully acknowledge the help of Boštjan Zupanc and Gregor Skoberne in both the field and laboratory. We thank Zlatko Rojc for his permission to perform the study on the plot, and Martin Cregeen for language editing.
References
Abadía, A., Gil, E., Morales, F., Montañés, L., Montserrat, G., and Abadía, J. (1996). Marcescence and senescence in a submediterranean oak (Quercus subpyrenaica E.H. del Villar): photosynthetic characteristics and nutrient composition. Plant Cell Environ. 19, 685–694. doi: 10.1111/j.1365-3040.1996.tb00403.x
Aloni, R. (2013). Role of hormones in controlling vascular differentiation and the mechanism of lateral root initiation. Planta 238, 819–830. doi: 10.1007/s00425-013-1927-8
Barbaroux, C., and Bréda, N. (2002). Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiol. 22, 1201–1210. doi: 10.1093/treephys/22.17.1201
Barbaroux, C., Bréda, N., and Dufrêne, E. (2003). Distribution of above-ground and below-ground carbohydrate reserves in adult trees of two contrasting broad-leaved species (Quercus petraea and Fagus sylvatica). New Phytol. 157, 605–615. doi: 10.1046/j.1469-8137.2003.00681.x
Bequet, R., Kint, V., Campioli, M., Vansteenkiste, D., Muys, B., and Ceulemans, R. (2012). Influence of stand, site and meteorological variables on the maximum leaf area index of beech, oak and Scots pine. Eur. J. For. Res. 131, 283–295. doi: 10.1007/s10342-011-0500-x
Bréda, N., and Granier, A. (1996). Intra- and interannual variations of transpiration, leaf area index and radial growth of a sessile oak stand (Quercus petraea). Ann. Sci. For. 53, 521–536. doi: 10.1051/forest:19960232
Burgess, S., and Downey, A. (2014). SFM1 Sap Flow Meter Manual. Version 1.0. Armidale, NSW: ICT International Pty Ltd.
Burner, D. M., Brauer, D. K., Snider, J. L., Harrington, C. A., and Moore, P. A. (2014). Phenological responses of juvenile pecan and white oak on an upland site. Agroforest Syst. 88, 141–155. doi: 10.1007/s10457-013-9662-5
Camarero, J. J., Olano, J. M., and Parras, A. (2010). Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 185, 471–480. doi: 10.1111/j.1469-8137.2009.03073.x
Cochard, H., and Tyree, M. T. (1990). Xylem dysfunction in Quercus: vessel sizes, tyloses, cavitation and seasonal changes in embolism. Tree Physiol. 6, 393–407. doi: 10.1093/treephys/6.4.393
Copini, P., Den Ouden, J., Robert, E. M. R., Tardif, J. C., Loesberg, W. A., Goudzwaard, L., et al. (2016). Flood-ring formation and root development in response to experimental flooding of young Quercus robur trees. Front. Plant Sci. 7:775. doi: 10.3389/fpls.2016.00775
Damesin, C., and Rambal, S. (1995). Field study of leaf photosynthetic performance by a Mediterranean deciduous oak tree (Quercus pubescens) during a severe summer drought. New Phytol. 131, 159–167. doi: 10.1111/j.1469-8137.1995.tb05717.x
Derr, W. F., and Evert, R. F. (1967). The cambium and seasonal development of the phloem in Robinia pseudoacacia. Am. J. Bot. 54, 147–153. doi: 10.2307/2440792
Do, F., and Rocheteau, A. (2002). Influence of natural temperature gradients on measurements of xylem sap flow with thermal dissipation probes. 2. Advantages and calibration of a noncontinuous heating system. Tree Physiol. 22, 649–654. doi: 10.1093/treephys/22.9.649
Eilmann, B., Weber, P., Rigling, A., and Eckstein, D. (2006). Growth reactions of Pinus sylvestris L. and Quercus pubescens Willd. to drought years at a xeric site in Valais, Switzerland. Dendrochronologia 23, 121–132. doi: 10.1016/j.dendro.2005.10.002
Eilmann, B., Zweifel, R., Buchmann, N., Fonti, P., and Rigling, A. (2009). Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 29, 1011–1020. doi: 10.1093/treephys/tpp035
Ellmore, G. S., and Ewers, F. W. (1985). Hydraulic conductivity in trunk xylem of elm, Ulmus americana. IAWA Bull. 6, 303–307. doi: 10.3389/fpls.2014.00253
Ferlan, M., Alberti, G., Eler, K., Batič, F., Peressotti, A., Miglietta, F., et al. (2011). Comparing carbon fluxes between different stages of secondary succession of a karst grassland. Agric. Ecosyst. Environ. 140, 199–207. doi: 10.1016/j.agee.2010.12.003
Ferlan, M., Eler, K., Simončič, P., Batič, F., and Vodnik, D. (2016). Carbon and water flux patterns of a drought-prone mid-succession ecosystem developed on abandoned karst grassland. Agric. Ecosyst. Environ. 220, 152–163. doi: 10.1016/j.agee.2016.01.020
González-González, B. D., García-González, I., and Vázquez-Ruiz, R. A. (2013). Comparative cambial dynamics and phenology of Quercus robur L. and Q. pyrenaica Willd. in an Atlantic forest of the northwestern Iberian Peninsula. Trees 27, 1571–1585. doi: 10.1007/s00468-013-0905-x
Granier, A., Anfodillo, T., Sabatti, M., Cochard, H., Dreyer, E., Tomasi, M., et al. (1994). Axial and radial water flow in the trunks of oak trees: a quantitative and qualitative analysis. Tree Physiol. 14, 1383–1396. doi: 10.1093/treephys/14.12.1383
Gričar, J. (2010). Xylem and phloem formation in sessile oak from Slovenia in 2007. Wood Res. 55, 15–22.
Gričar, J., Jagodic, Š.,Šefc, B., Trajković, J., and Eler, K. (2014). Can the structure of dormant cambium and the widths of phloem and xylem increments be used as indicators for tree vitality? Eur. J. For. Res. 133, 551–562. doi: 10.1007/s10342-014-0784-8
Gricar, J., Prislan, P., De Luis, M., Gryc, V., Hacurova, J., Vavrcik, H., et al. (2015). Plasticity in variation of xylem and phloem cell characteristics of Norway spruce under different local conditions. Front. Plant Sci. 6:730. doi: 10.3389/fpls.2015.00730
Gričar, J., Prislan, P., De Luis, M., Novak, K., Longares, L. A., Martinez Del Castillo, E., et al. (2016). Lack of annual periodicity in cambial production of phloem in trees from Mediterranean areas. IAWA J. 37, 332–348. doi: 10.1163/22941932-20160138
Gričar, J., Zupančič, M., Čufar, K., and Oven, P. (2007). Regular cambial activity and xylem and phloem formation in locally heated and cooled stem portions of Norway spruce. Wood Sci. Technol. 41, 463–475. doi: 10.1007/s00226-006-0109-2
Hernandez-Santana, V., Martinez-Vilalta, J., Martinez-Fernandez, J., and Williams, M. (2009). Evaluating the effect of drier and warmer conditions on water use by Quercus pyrenaica. For. Ecol. Manage. 258, 1719–1730. doi: 10.1016/j.foreco.2009.07.038
Hinckley, T. M., and Lassoie, J. P. (1981). Radial growth in conifers and deciduous trees: a comparison. Mitt. Forstl. Bundes Versanst. Wien 142, 17–56.
Hoch, G., Richter, A., and Körner, C. (2003). Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 26, 1067–1081. doi: 10.1046/j.0016-8025.2003.01032.x
Hsiao, T. C., and Acevedo, E. (1974). Plant responses to water deficits, water-use efficiency, and drought resistance. Agric. Meteorol. 14, 59–84. doi: 10.1016/0002-1571(74)90011-9
ICT International (2014). Available at: http://au.ictinternational.com/products/plants/sap-flow/ [accessed March 3, 2014].
IPCC (2014). Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds Core Writing Team, R. K. Pachauri, and L. A. Meyer. Geneva: IPCC.
Jonard, F., André, F., Ponette, Q., Vincke, C., and Jonard, M. (2011). Sap flux density and stomatal conductance of European beech and common oak trees in pure and mixed stands during the summer drought of 2003. J. Hydrol. 409, 371–381. doi: 10.1016/j.jhydrol.2011.08.032
Jyske, T., and Hölttä, T. (2015). Comparison of phloem and xylem hydraulic architecture in Picea abies stems. New Phytol. 205, 102–115. doi: 10.1111/nph.12973
Kitin, P., and Funada, R. (2016). Earlywood vessels in ring-porous trees become functional for water transport after bud burst and before the maturation of the current-year leaves. IAWA J. 37, 315–331. doi: 10.1163/22941932-20160136
Lachaud, S., Catesson, A. M., and Bonnemain, J. L. (1999). Structure and functions of the vascular cambium. C. R. Acad. Sci. III 322, 633–650. doi: 10.1016/S0764-4469(99)80103-6
Martinez del Castillo, E., Longares, L. A., Gričar, J., Prislan, P., Gil-Pelegrín, E., Čufar, K., et al. (2016). Living on the edge: contrasted wood-formation dynamics in Fagus sylvatica and Pinus sylvestris under Mediterranean conditions. Front. Plant Sci. 7:370. doi: 10.3389/fpls.2016.00370
Michelot, A., Simard, S., Rathgeber, C., Dufrêne, E., and Damesin, C. (2012). Comparing the intra-annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 32, 1033–1045. doi: 10.1093/treephys/tps052
Mooney, H. A. (1972). The carbon balance of plants. Annu. Rev. Ecol. Syst. 3, 315–346. doi: 10.1146/annurev.es.03.110172.001531
Nardini, A., Casolo, V., Dal Borgo, A., Savi, T., Stenni, B., Bertoncin, P., et al. (2016). Rooting depth, water relations and non-structural carbohydrate dynamics in three woody angiosperms differentially affected by an extreme summer drought. Plant Cell Environ. 39, 618–627. doi: 10.1111/pce.12646
Nasr, Z., Woo, S. Y., Zineddine, M., Khaldi, A., et al. (2011). Sap flow estimates of Quercus suber according to climatic conditions in north Tunisia. Afr. J. Agric. Res. 6, 4705–4710.
Oparka, K. J., and Robert, T. (1999). Sieve elements and companion cells – traffic control centers of the phloem. Plant Cell 11, 739–750. doi: 10.1105/tpc.11.4.739
Panshin, A. J., and de Zeeuw, C. (1980). Textbook of Wood Technology, 4th Edn. New York, NY: McGraw-Hill. doi: 10.5194/bg-13-5499-2016
Pérez-de-Lis, G., García-González, I., Rozas, V., and Olano, J. M. (2016a). Feedbacks between earlywood anatomy and non-structural carbohydrates affect spring phenology and wood production in ring-porous oaks. Biogeosciences 13, 5499–5510. doi: 10.1111/nph.13610
Pérez-de-Lis, G., Rossi, S., Vázquez-Ruiz, R. A., Rozas, V., and García-González, I. (2016b). Do changes in spring phenology affect earlywood vessels? Perspective from the xylogenesis monitoring of two sympatric ring-porous oaks. New Phytol. 209, 521–530. doi: 10.1016/j.foreco.2010.01.006
Pfautsch, S., Bleby, T. M., Rennenberg, H., and Adams, M. A. (2010). Sap flow measurements reveal influence of temperature and stand structure on water use of Eucalyptus regnans forests. For. Ecol. Manage. 259, 1190–1199. doi: 10.1104/pp.010816
Plomion, C., Leprovost, G., and Stokes, A. (2001). Wood formation in trees. Plant Physiol. 127, 1513–1523. doi: 10.1093/treephys/27.4.537
Poyatos, R., Čermák, J., and Llorens, P. (2007). Variation in the radial patterns of sap flux density in pubescent oak (Quercus pubescens) and its implications for tree and stand transpiration measurements. Tree Physiol. 27, 537–548. doi: 10.5194/hess-9-493-2005
Poyatos, R., Llorens, P., and Gallart, F. (2005). Transpiration of montane Pinus sylvestris L. and Quercus pubescens Willd. forest stands measured with sap flow sensors in NE Spain. Hydrol. Earth Syst. Sci. 9, 493–505. doi: 10.1016/j.agrformet.2013.06.001
Prislan, P., Gričar, J., De Luis, M., Smith, K. T., and Čufar, K. (2013). Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Meteorol. 180, 142–151.
Prislan, P., Schmitt, U., Koch, G., Gričar, J., and Čufarl, K. (2011). Seasonal ultrastructural changes in the cambial zone of beech (Fagus sylvatica) grown at two different altitudes. IAWA J. 32, 443–459. doi: 10.1163/22941932-90000070
R Core Team (2015). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rossi, S., Anfodillo, T., and Menardi, R. (2006). Trephor: a new tool for sampling microcores from tree stems. IAWA J. 27, 89–97. doi: 10.1163/22941932-90000139
Rossi, S., Deslauriers, A., and Morin, H. (2003). Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia 21, 33–39. doi: 10.1078/1125-7865-00034
Sánchez de Dios, R., Benito-Garzón, M., and Sainz-Ollero, H. (2009). Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant Ecol. 204, 189–205. doi: 10.1007/s11258-009-9584-5
Sass-Klaassen, U., Fonti, P., Cherubini, P., Gričar, J., Robert, E. M. R., Steppe, K., et al. (2016). A tree-centered approach to assess impacts of extreme climatic events on forests. Front. Plant Sci. 7:1069. doi: 10.3389/fpls.2016.01069
Sass-Klaassen, U., Sabajo, C. R., and Den Ouden, J. (2011). Vessel formation in relation to leaf phenology in pedunculate oak and European ash. Dendrochronologia 29, 171–175. doi: 10.1016/j.dendro.2011.01.002
Srivastava, L. M. (1964). Anatomy, Chemistry, and Physiology of Bark. New York, NY: Academic Press. doi: 10.1016/j.tplants.2015.03.015
Steppe, K., Sterck, F., and Deslauriers, A. (2015). Diel growth dynamics in tree stems: linking anatomy and ecophysiology. Trends Plant Sci. 20, 335–343. doi: 10.1016/j.tplants.2015.03.015
Suzuki, M., Yoda, K., and Suzuki, H. (1996). Phenological comparison on the onset of vessel formation between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest. IAWA J. 17, 431–444. doi: 10.1163/22941932-90000641
Takahashi, S., Okada, N., and Nobuchi, T. (2013). Relationship between the timing of vessel formation and leaf phenology in ten ring-porous and diffuse-porous deciduous tree species. Ecol. Res. 28, 615–624. doi: 10.1007/s11284-013-1053-x
Tognetti, R., Cherubini, P., Marchi, S., and Raschi, A. (2007). Leaf traits and tree rings suggest different water-use and carbon assimilation strategies by two co-occurring Quercus species in a Mediterranean mixed-forest stand in Tuscany, Italy. Tree Physiol. 27, 1741–1751. doi: 10.1093/treephys/27.12.1741
Tognetti, R., Longobucco, A., and Raschi, A. (1998). Vulnerability of xylem to embolism in relation to plant hydraulic resistance in Quercus pubescens and Quercus ilex co-occurring in a Mediterranean coppice stand in central Italy. New Phytol. 139, 437–447. doi: 10.1046/j.1469-8137.1998.00207.x
Tyree, M. T., and Cochard, H. (1996). Summer and winter embolism in oak: impact on water relations. Ann. Sci. For. 53, 173–180. doi: 10.1051/forest:19960201
van der Werf, G. W., Sass-Klaassen, U., and Mohren, G. M. J. (2007). The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 25, 103–112. doi: 10.1016/j.dendro.2007.03.004
Vandegehuchte, M. W., and Steppe, K. (2013). Sap-flux density measurement methods: working principles and applicability. Funct. Plant Biol. 40, 213–223. doi: 10.1071/FP12233
Vertessy, R. A., Benyon, R. G., O’Sullivan, S. K., and Gribben, P. R. (1995). Relationships between stem diameter, sapwood area, leaf area and transpiration in a young mountain ash forest. Tree Physiol. 15, 559–567. doi: 10.1093/treephys/15.9.559
Wullschleger, S. D., Hanson, P. J., and Todd, D. E. (2001). Transpiration from a multi-species deciduous forest as estimated by xylem sap flow techniques. For. Ecol. Manage. 143, 205–213. doi: 10.1016/S0378-1127(00)00518-1
Zamski, E., and Zimmermann, M. H. (1979). Sieve tube longevity in white ash (Fraxinus americana) studied with a new histochemical test for the identification of sugar. Can. J. Bot. 57, 650–656. doi: 10.1139/b79-083
Keywords: pubescent oak, cambium, radial growth, cell differentiation, leaf development, sub-Mediterranean
Citation: Lavrič M, Eler K, Ferlan M, Vodnik D and Gričar J (2017) Chronological Sequence of Leaf Phenology, Xylem and Phloem Formation and Sap Flow of Quercus pubescens from Abandoned Karst Grasslands. Front. Plant Sci. 8:314. doi: 10.3389/fpls.2017.00314
Received: 03 January 2017; Accepted: 20 February 2017;
Published: 06 March 2017.
Edited by:
Boris Rewald, University of Natural Resources and Life Sciences, Vienna, AustriaReviewed by:
Cristina Nabais, University of Coimbra, PortugalUwe Hacke, University of Alberta, Canada
Copyright © 2017 Lavrič, Eler, Ferlan, Vodnik and Gričar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jožica Gričar, am96aWNhLmdyaWNhckBnb3pkaXMuc2k=