 Peipei Zhang1Guihong Yin2Yue Zhou3Aiyong Qi1Fengmei Gao4Xianchun Xia4
Peipei Zhang1Guihong Yin2Yue Zhou3Aiyong Qi1Fengmei Gao4Xianchun Xia4 Zhonghu He4
Zhonghu He4 Zaifeng Li1*Daqun Liu1*
Zaifeng Li1*Daqun Liu1*- 1College of Plant Protection, Agricultural University of Hebei, Baoding, China
- 2Zhoukou Academy of Agricultural Sciences, Zhoukou, China
- 3Department of Biochemistry, Baoding University, Baoding, China
- 4Institute of Crop Science, National Wheat Improvement Center – Chinese Academy of Agricultural Sciences, Beijing, China
Wheat leaf rust is an important disease worldwide. Growing resistant cultivars is an effective means to control the disease. In the present study, 244 recombinant inbred lines from Zhou 8425B/Chinese Spring cross were phenotyped for leaf rust severities during the 2011–2012, 2012–2013, 2013–2014, and 2014–2015 cropping seasons at Baoding, Hebei province, and 2012–2013 and 2013–2014 cropping seasons in Zhoukou, Henan province. The population was genotyped using the high-density Illumina iSelect 90K SNP assay and SSR markers. Inclusive composite interval mapping identified eight QTL, designated as QLr.hebau-2AL, QLr.hebau-2BS, QLr.hebau-3A, QLr.hebau-3BS, QLr.hebau-4AL, QLr.hebau-4B, QLr.hebau-5BL, and QLr.hebau-7DS, respectively. QLr.hebau-2BS, QLr.hebau-3A, QLr.hebau-3BS, and QLr.hebau-5BL were derived from Zhou 8425B, whereas the other four were from Chinese Spring. Three stable QTL on chromosomes 2BS, 4B and 7DS explained 7.5–10.6%, 5.5–24.4%, and 11.2–20.9% of the phenotypic variance, respectively. QLr.hebau-2BS in Zhou 8425B might be the same as LrZH22 in Zhoumai 22; QLr.hebau-4B might be the residual resistance of Lr12, and QLr.hebau-7DS is Lr34. QLr.hebau-2AL, QLr.hebau-3BS, QLr.hebau-4AL, and QLr.hebau-5BL are likely to be novel QTL for leaf rust. These QTL and their closely linked SNP and SSR markers can be used for fine mapping, candidate gene discovery, and marker-assisted selection in wheat breeding.
Introduction
Wheat is among the most important food crops (Curtis and Halford, 2014). Leaf rust (LR), caused by Puccinia triticina (Pt), is an important disease on wheat. LR occurred severely in 1969, 1973, 1975, 1979, and 2012 in China and caused serious yield loss (Dong, 2001; Zhou et al., 2013). Although LR can be controlled by fungicides overuse may lead to fungicide residues in the grain, or resistance to fungicides followed by a resurgence of the disease under favorable conditions (Luo, 2009). Therefore, planting resistant cultivars is the most effective, economic and environmentally safe mean of controlling the disease.
There are two kinds of resistance, viz. race specific and race non-specific to wheat LR. Race specific resistance is often controlled by a single gene or simple gene combinations and elicits a HR. This kind of resistance is often not durable as it is readily overcome by new pathotypes that lack the corresponding avirulence factors. In contrast, race non-specific resistance, termed as APR, slow rusting or partial resistance, typically reduces growth and reproduction of the pathogen on adult plants. It is usually controlled by several minor genes, and tends to be more durable compared with race specific resistance.
To date, more than 100 LR genes have been documented, and 76 have been cataloged (McIntosh et al., 2016). Most of these genes are race specific and can be overcome easily by new pathogen pathotypes. Only four known slow rusting resistance genes, viz. Lr34, Lr46, Lr67 and Lr68, have been cataloged at present (Dyck, 1977; Singh et al., 1998; Herrera-Foessel et al., 2011; Herrera-Foessel et al., 2012; Li et al., 2014). Besides the four cataloged slow rusting genes, 80 other APR QTL for LR have been mapped on 16 wheat chromosomes (Li et al., 2014). Although minor gene resistance tends to be more durable than major gene resistance, it also might be overcome by slow evolution in the pathogen populations (McDonald and Linde, 2002). Therefore, it is very important to identify more APR gene in wheat cultivars for controlling wheat rusts in China.
Molecular markers have been widely used in mapping and cloning resistance genes. SSR markers are widely used in linkage mapping due to advantages of co-dominance, stability, high polymorphism, chromosome specificity, and ease of manipulation (Röder et al., 1998). The SNP gene-chip technology developed quickly recently, and it provides a superior way for gene mapping due to its higher accuracy and density than other markers (Yu et al., 2011). A combined use of SSR and SNP markers is rational in that SSR might act as a framework to anchor SNP to the chromosomes. The map provides connections between SNP and SSR markers and can be used for comparative mapping of QTL linked to SSR markers reported before (Li et al., 2015).
The Chinese wheat line Zhou 8425B, developed by the Zhoukou Academy of Agricultural Sciences (ZAAS) in 1984, is still high resistant to wheat rusts and powdery mildew in the field at present. During the past 20 years, about 100 cultivars were derived from this line and had been planted over 33 million ha in China (Yin et al., 2009). In our previous work, a seedling major gene LrZH84 was mapped on chromosome 1BL in Zhou 8425B (Zhao et al., 2008). The aim of present study was to identify APR QTL to LR and their closely linked molecular markers for MAS in wheat breeding.
Materials and Methods
Wheat Materials and P. triticina Pathotypes
A total of 244 F8 RILs derived from Zhou 8425B/Chinese Spring cross were used to map QTL for APR to LR. Zhou 8425B, Chinese Spring, Zhoumai 22, and 36 differential lines were included in the seedling tests with 14 Chinese Pt pathotypes (Table 1). The pathotypes were named following the Pt coding system described by Long and Kolmer (1989).
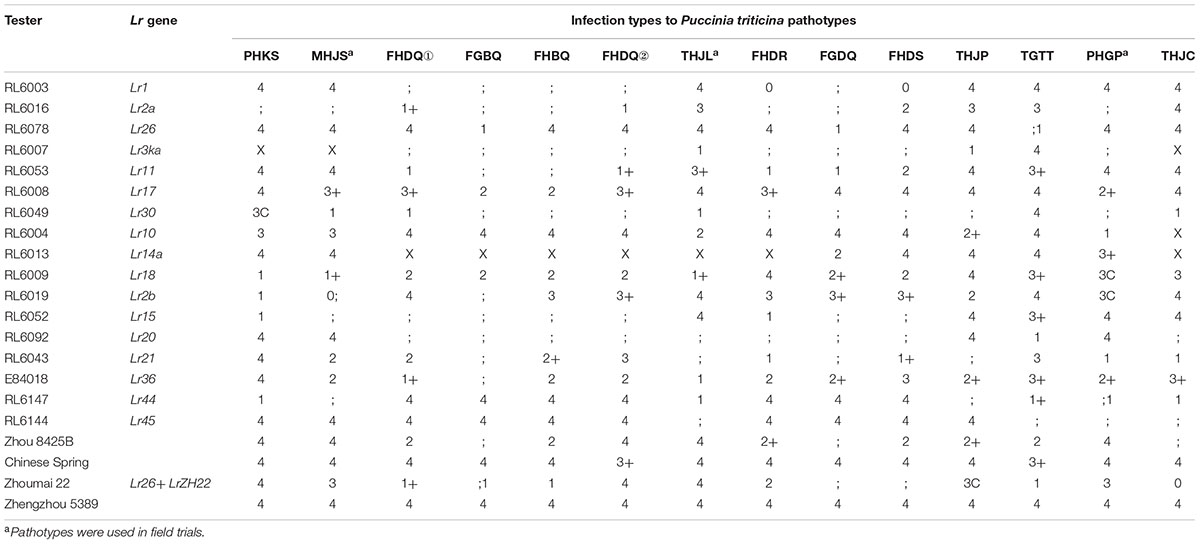
TABLE 1. Seedling infection types on Zhou 8425B, Chinese Spring, Zhoumai 22 and 36 wheat lines with known Lr genes when tested with 14 Chinese Puccinia triticina pathotypes.
Evaluation of Seedling Responses in the Greenhouse
Seedlings were planted in a growth chamber (30 cm × 50 cm). Inoculation of seedlings was carried out by the method provided by Li et al. (2010). ITs were recorded 10–14 days after inoculation based on the 0–4 Stakman scale modified by Roelfs et al. (1992). The gene postulation was conducted using the method reported by Dubin et al. (1989). The Zhou 8425B/Chinese Spring RIL population was also inoculated with the pathotype FHDQ to verify the seedling gene in Zhou 8425B.
to verify the seedling gene in Zhou 8425B.
Leaf Rust Tests in the Field
Zhou 8425B, Chinese Spring and the 244 F8 RILs from the cross Zhou 8425B/Chinese Spring were evaluated for APR for LR in the field at Baoding, Hebei province during the 2011–2012, 2012–2013, 2013–2014, and 2014–2015 cropping seasons and Zhoukou, Henan province during the 2012–2013 and 2013–2014 cropping seasons. The field test designed followed Qi et al. (2016). Spreader rows of susceptible line Zhengzhou 5389 were sown perpendicular and adjacent to the test lines to aid the spread of spores. Equal amounts of Pt pathotypes PHGP, MHJS, and THJL (virulent on seedlings of Zhou 8425B and Chinese Spring) were used for field inoculation. Field inoculation was conducted by the method provided by Zhou et al. (2014). Four weeks after inoculation LR severities were recorded three times at weekly intervals using the modified Cobb scale (Peterson et al., 1948). MDS at Baoding in Hebei in the 2011–2012, 2012–2013, 2013–2014, and 2014–2015 cropping seasons and Zhoukou in Henan in the 2012–2013 and 2013–2014 cropping seasons will hereafter be referred to as 2012BD, 2013BD, 2014BD, 2015BD, 2013ZK, and 2014ZK, respectively; these were used for statistical and QTL analysis.
Statistical Analysis
Analyses of variance (ANOVA) were conducted by IBM SPSS Statistics 19.0 software. The correlation coefficients of phenotypic data between MDS in different environments were calculated by the Microsoft Excel analytical tool. Broad-sense heritabilities (h2) for LR resistance were calculated by the formula: h2 = σg2/(σg2+σge2/e+σε2/re) (Zhou et al., 2014).
Genotyping Using SNP and SSR Markers
Genomic DNA was extracted from 10 non-infected seedling leaves of each line including the parents using the CTAB method (Sharp et al., 1988). Zhou 8425B, Chinese Spring and the 244 RILs were genotyped by the 90K iSelect SNP array in our previous study (Gao et al., 2015). In addition, 26 SSR markers were also used to genotype the whole population for further linkage and QTL mapping in the present study.
QTL Analysis
Linkage maps were constructed using SNP markers combined with SSRs. The software QTL IciMapping 3.1 was used to detect APR QTL (Li et al., 2007). Phenotypic values of all lines in each environment were used for QTL detection. The procedures for QTL detection and digenic interactions analysis between non-allelic QTL were similar with Zhou et al. (2014) and Qi et al. (2016).
Results
Resistance Genes Postulated from Seedling Reactions
The different ITs of 36 wheat differential lines with known Lr genes provided an ability to postulate 17 LR genes, viz. Lr1, Lr2a, Lr26, Lr3ka, Lr11, Lr17, Lr30, Lr10, Lr14a, Lr18, Lr2b, Lr15, Lr20, Lr21, Lr36, Lr44, and Lr45, when inoculated with 14 Pt pathotype (Table 1). Ten genes, viz. Lr9, Lr19, Lr24, Lr28, Lr29, Lr39, Lr42, Lr47, Lr51, and Lr53, showed resistance to all the tested pathotypes. It is not possible to postulate genes Lr2c, Lr3a, Lr3b, Lr13, Lr14b, Lr16, Lr23, Lr33, and LrB because these genes were susceptible to most pathotypes. Zhou 8425B showed a similar response pattern to Zhoumai 22 which carries LrZH22 (Wang et al., 2016). Zhou 8425B is one of the parents of Zhoumai 22, so LrZH22 in Zhoumai 22 was likely derived from Zhou 8425B. In previous report Zhou 8425B also contained Lr26 and LrZH84 (Zhao et al., 2008). In the seedling test Zhou 8425B was resistant to three Pt pathotypes avirulent to Lr26, which further confirmed that Zhou 8425B contained Lr26.
Phenotypic Evaluation for Leaf Rust in the Field
Leaf rust developed well in all environments. As pathotypes used in the field were virulent on seedlings of both parents, low disease severities presumably resulted from slow rusting resistance genes. The MDS of Zhengzhou 5389 ranged from 70 to 100% across environments. Zhou 8425B had mean MDS scores of 10% across six environments, whereas Chinese Spring showed a MDS of 15%. MDS in the RILs ranged from 1 to 100% across all environments exhibiting significant differences among genotypes. The frequency distribution of LR MDS in each environment showed a continuous distribution skewed toward resistance (Supplementary Figure S1), indicating polygenic inheritance. Correlation coefficients for the population ranged from 0.59 to 0.82 across different environments (P < 0.0001) (Table 2). Mean MDS across all environments was 16.7%. Broad-sense heritability of MDS across six environments was 0.77. ANOVA confirmed significant variation among the RILs (Table 3).

TABLE 2. Pearson correlation coefficients (r) for two-way comparisons of leaf rust (LR) severity data from different environments.
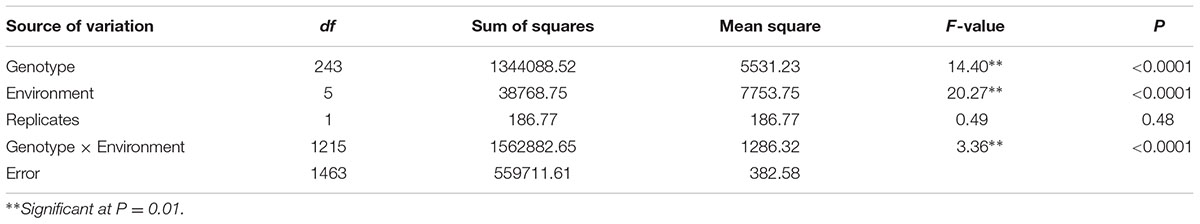
TABLE 3. Analysis of variance of maximum disease severities (MDS) for leaf rust in the population of Zhou 8425B/Chinese Spring.
Linkage Map Construction
A total of 21 linkage groups corresponding to the 21 hexaploid wheat chromosomes were constructed from 5,636 high-quality polymorphic SNP markers (Gao et al., 2015) and 26 SSR markers (Figure 1).

FIGURE 1. Simplified genetic maps of nine chromosomes showing QTL for leaf rust (LR) resistance in the Zhou 8425B/Chinese Spring population. The bar length of each chromosome do not represent the same genetic distance. The epistasis QTLs also list on the figure. 1∗, 2∗, and 3∗ indicate interactions in 2012BD, 2014BD, and 2015BD, respectively, on the map.
Resistance of LrZH84 in the Field
Zhou 8425B was known to carry Lr26 and LrZH84 (Zhao et al., 2008). Lr26 have lost resistance to most of Pt pathotypes in China. LrZH84 was mapped on 1BL, and linked to SSR markers gwm582 and barc8 with genetic distances of 3.9 and 5.2 cM, respectively (Zhao et al., 2008). In the present study, two SSR markers gwm582 and barc8 linked to LrZH84 were used to test the entire RIL population to identify the effect of LrZH84 at adult-plant stage; the result showed that lines with LrZH84 had lost resistance to the mixed virulent pathotypes in the field (data not shown). The APR in Zhou 8425B was conferred by other LR resistance genes.
QTL for LR Resistance
Eight putative LR APR QTL were identified on chromosomes 2AL, 2BS, 3A, 3BS, 4AL, 4B, 5BL, and 7DS (Table 4 and Figure 1) based on the MDS data, and were designated as QLr.hebau-2AL, QLr.hebau-2BS, QLr.hebau-3A, QLr.hebau-3BS, QLr.hebau-4AL, QLr.hebau-4B, QLr.hebau-5BL, and QLr.hebau-7DS, respectively. The QTL on 2BS, 3A, 3BS, and 5BL were derived from Zhou 8425B, whereas those on 2AL, 4AL, 4B, and 7DS were from Chinese Spring. As expected there was no evidence of any contributions from Lr26 or LrZH84.
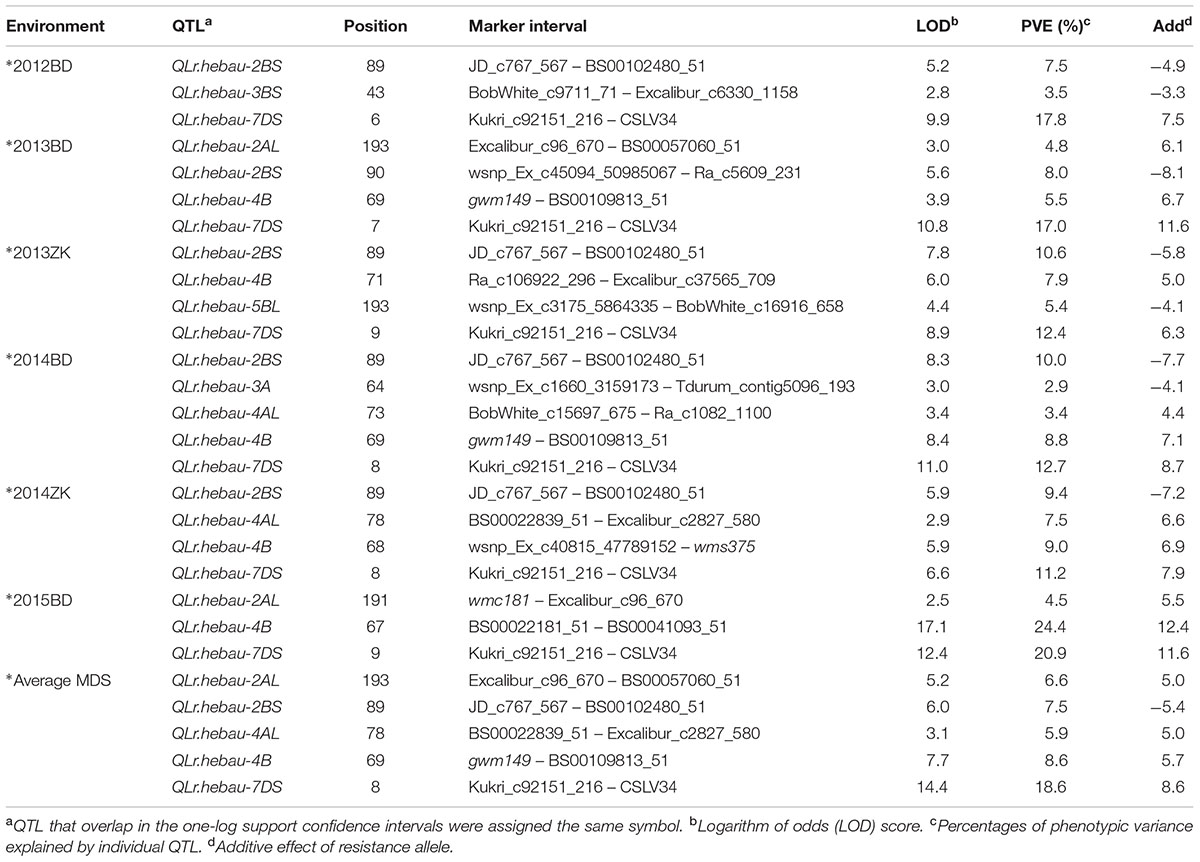
TABLE 4. Quantitative trait locus/loci for MDS to leaf rust by ICIM in the RIL population from Zhou 8425B/Chinese Spring.
QLr.hebau-7DS, in the marker interval Kukri_c92151_216 – csLV34 was stably detected in all environments, explaining 17.8, 17.0, 12.4, 12.7, 11.2, 20.9, and 18.6% of the phenotypic variances in 2012BD, 2013BD, 2013ZK, 2014BD, 2014ZK 2015BD and average MDS, with additive effects from Chinese Spring of 7.5, 11.6, 6.3, 8.7, 7.9, 11.6, and 8.6, respectively.
The second QTL QLr.hebau-2BS identified in 2012BD, 2013BD, 2013ZK, 2014BD, 2014ZK and average MDS accounted for 7.5, 8.0, 10.6, 10.0, 9.4, and 7.5% of the phenotypic variances, respectively. The additive effects from Zhou 8425B were 4.9, 8.1, 5.8, 7.7, 7.2, and 5.4, respectively. To further confirm the relationship between LrZH22 and QLr.hebau-2BS, the pathotype FHDQ avirulent to LrZH22 was used to inoculate the whole population at the seedling stage. The RIL population segregated 112 resistant lines with IT 1–2, 126 susceptible lines with IT 3–4 and 6 segregated lines, indicating single gene LrZH22 conferred seedling resistance to FHDQ (χ2 = 2.09, 2 df, P > 0.25). Then the phenotype data was combined with genotype data for mapping LrZH22 using software Joinmap 4.0. The result showed that the two closest flanking SNP loci were JD_c767_567 and RFL_Contig1483_1765 with genetic distances of 0.3 and 0.6 cM, respectively; this places the LrZH22 locus in the same vicinity as QLr.hebau-2BS. Thus QLr.hebau-2BS should be the same gene in Zhoumai 22. The APR gene effect on 2BS is likely to be residual resistance from LrZH22 (Wang et al., 2016).
QLr.hebau-4B from Chinese Spring in the marker interval BS00022181_51 – Excalibur_c37565_709 identified in 2013BD, 2013ZK, 2014BD, 2014ZK, 2015BD and average MDS explained 5.5, 7.9, 8.8, 9.0, 24.4, and 8.6% of the phenotypic variances, respectively. The additive effects were 6.7, 5.0, 7.1, 6.9, 12.4, and 5.7, respectively.
Two QTL were identified only in two environments and average MDS. QLr.hebau-2AL in the marker interval wmc181 – BS00057060_51 explained 4.8, 4.5, and 6.6% of the phenotypic variance in 2013BD, 2015BD and average MDS with additive effects from Chinese Spring of 6.1, 5.5, and 5.0, respectively. Another QTL QLr.hebau-4AL, located in the region of BobWhite_c15697_675 – Excalibur_c2827_580, explained 3.4, 7.5, and 5.9% of the phenotypic variance in 2014BD, 2014ZK and average MDS, with additive effects from Chinese Spring of 4.4, 6.6, and 5.0, respectively. The effects of the two QTL need to verify using further evidence.
The three tentative QTL, viz. QLr.hebau-3A, QLr.hebau-3BS and QLr.hebau-5BL, were detected only in one environment, and their effects need to be further confirmed. QLr.hebau-3A, in marker interval wsnp_Ex_c1660_3159173 – Tdurum_contig5096_193 explained 2.9% of the phenotypic variance in 2014BD, with an additive effect from Zhou 8425B of 4.1. QLr.hebau-3BS, flanked by SNP markers BobWhite_c9711_71 and Excalibur_c6330_1158, explained 3.5% of the phenotypic variance in 2012BD, with an additive effect from Zhou 8425B of -3.3. QLr.hebau-5BL, in interval wsnp_Ex_c3175_5864335 – BobWhite_c16916_658, accounted for 5.5% of the phenotypic variance in 2013ZK, and the additive effect from Zhou 8425B was 4.1.
The total phenotypic variances explained by all QTL in a simultaneous fit ranged from 28.8 to 49.8% across environments, indicating significant effects of the QTL in reducing LR severity.
Epistasis of QTL for Leaf Rust Resistance
Three epistatic QTL were identified for the LR resistance. The variation explained ranged from 7.5 to 18.4% (Table 5 and Figure 1). The first interaction was detected between the chromosome 1AS and 3B in 2012BD, explaining 7.5% of the phenotypic variance with the additive effect of -5.1. The second interaction was detected between chromosome 2AL and 2BS in 2014BD, explaining the phenotypic variance of 15.1% with the additive effect of -9.5. The QTL on 2BS located at the similar position as QLr.hebau-2BS. The last interaction is detected between chromosome 2AS and 4B in 2015BD, which explained 18.4% of the phenotypic variance with the additive effect of 9.7.
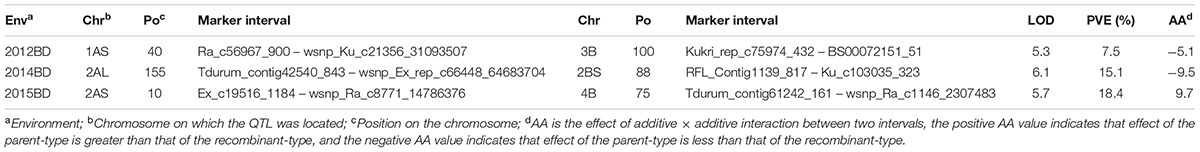
TABLE 5. Epistastic QTL for leaf rust resistance in the Zhou 8425B/Chinese Spring population.
Discussion
The Major QTL
In the present research five major QTL were identified by QTL analysis using the average MDS from different environments.QLr.hebau-2AL.
QLr.hebau-2AL was mapped in the marker interval wmc181 – BS00057060_51. Three known LR QTL, viz. QLr.cimmyt-2AL (Rosewarne et al., 2012), QLr.sfr-2AL (Schnurbusch et al., 2004) and QLr.ubo-2A (Maccaferri et al., 2008), were located on chromosome 2AL. QLr.cimmyt-2AL and QLr.sfr-2AL were mapped at 63 cM on 2A, and QLr.ubo-2A was at the end of 2AL (143 cM) (Li et al., 2014). QLr.hebau-2AL linked to wmc181 was mapped at 103 cM on 2A based on the consensus map of Somers et al. (2004). Therefore, QLr.hebau-2AL appears to be a new QTL.
QLr.hebau-2BS
Six known Lr genes, viz. Lr13, Lr16, Lr23, Lr35, Lr48, and LrZH22 (Li et al., 2014; Wang et al., 2016) are located on chromosome 2BS. QLr.hebau-2BS was closely linked with SSR markers barc55 and wmc474, in the same position as LrZH22 (Wang et al., 2016). In the seedling tests Zhou 8425B was postulated to contain LrZH22, a temperature-sensitive gene on chromosome 2BS in Zhoumai 22 (Wang et al., 2016), and QLr.hebau-2BS is likely to be residual resistance from LrZH22.
QLr.hebau-4AL
QLr.hebau-4AL from Chinese Spring was located between BobWhite_c15697_675 and Excalibur_c2827_580, and also closely linked to SSR marker wmc617. No LR resistance gene was previously reported in this position (Li et al., 2014), hence QLr.hebau-4AL is a new APR QTL.
QLr.hebau-4B
Four known LR resistance genes, viz. Lr12 (Singh and Bowden, 2011), Lr25 (Singh et al., 2012), Lr31, and Lr49 (Bansal et al., 2008) and three APR QTL viz. QLr.sfrs-4B (Messmer et al., 2000), QLr.pbi-4BL (Singh et al., 2009), and QLr.cimmyt-4BL (William et al., 2006) have been mapped on chromosome 4B. In the present study QLr.hebau-4B was detected in Chinese Spring, which carries the race-specific APR gene Lr12. QLr.hebau-4B was flanked by SSR loci Xgwm149 and Xgwm375, which were closely linked to Lr12 with genetic distances of 1.9 and 3.1 cM, respectively. In the field test Chinese Spring was susceptible to inoculated mixture of pathotypes with IT 4, indicating that Lr12 in Chinese Spring has lost resistance to the mixed pathotypes, and the minor effect of QLr.hebau-4B is likely to be the residual effect from the race-specific gene Lr12. However, in another field trial the MDS of RL6011 (Lr12) was 5% with IT 2 and Thatcher (Lr22) was 70% with IT 4 (data not shown); the relationship between QLr.hebau-4B and Lr12 in RL6011 is unclear and needs to be tested in the future.
QLr.hebau-7DS
The known pleiotropic APR gene Lr34/Yr18/Pm38/Sr57 was mapped on chromosome 7DS (Lillemo et al., 2008). In the present study, QLr.hebau-7DS detected from Chinese Spring was on chromosome 7DS, flanked by markers csLV34 (closely linked to Lr34) and Kukri_c92151_216. Chinese Spring contains Lr34 (Piech and Supryn, 1978; Krattinger et al., 2009); QLr.hebau-7DS should be Lr34.
The Tentative QTL
Three QTL can only be detected in few environment(s) and can’t be detected in Average MDS.
QLr.hebau-3A
Three QTL QLr.ubo-3A, QLr.sfrs-3AL, and QLr.fcu-3AL were located on chromosome 3A (Li et al., 2014). QLr.hebau-3A in the marker interval wsnp_Ex_c1660_3159173 – Tdurum_contig5096_193 was linked to SSR markers wmc651 and wmc264 with genetic distances of 4.6 and 13.6 cM, respectively. Based on the Somers map QLr.hebau-3A was located at a similar position to QLr.ubo-3A and QLr.sfrs-3AL but with different genetic background. The relationships among the three genes were unclear and need to be verified in future work.
QLr.hebau-3BS
QLr.hebau-3BS from Zhou 8425B was located in marker interval BobWhite_c9711_71 – Excalibur_c6330_1158; it was linked to SSR markers barc147 and gwm493, at genetic distances of 7.4 and 6.9 cM, respectively, at the position 10 cM on the Somers map. Two QTL, QLr.sfrs-3B (Messmer et al., 2000) and QLr.fcu-3BL (Chu et al., 2009), have been located on chromosome 3B. QLr.fcu-3BL was mapped on chromosome 3BL and QLr.sfrs-3B was mapped at the position 61 cM on 3B, which is near the centromere based on the Somers map. Therefore, QLr.hebau-3BS is likely to be new.
QLr.hebau-5BL
Three known LR QTL QLr.hbau-5BL (Zhang et al., 2009), QLr.sfrs-5BL (Messmer et al., 2000), and QLr.fcu-5BL (Chu et al., 2009) were located at a similar position of 140 cM on chromosome 5B (Li et al., 2014), about was 34.1 cM from QLr.hebau-5BL in the present study. Therefore QLr.hebau-5BL is likely to be new.
QTL and Their Phenotypic Effects
The total phenotypic variances explained by all additive QTL ranged from 28.8 to 49.8% across environments. The effect of APR QTL can be easily influenced by environment (Zhang et al., 2009). For example, QLr.hebau-4B explained 5.5–24.4% of the phenotypic variance across different environments, indicating the QTL was not very stable among different environments. Although the phenotypic variance explained by the epistatic QTL was changed across different environments, the interaction between QTL also played an important role in the genetic control of quantitative traits (Roncallo et al., 2012). In this study, different epistatic QTL were detected in 2012BD, 2014BD and 2015BD, and explained 7.5–18.4% of the phenotypic variance. Some interactions didn’t involve the detected QTL, but their effects are just as important as some of the individual QTL in terms of LOD, PVE, and AA effect. These suggested that the epistasis as a genetic factor played an important role in the LR resistance. The additive and epistatic effects of QTL together contributed to the LR resistance.
Conclusion
In this study, 8 QTL were detected in the Zhou 8425B/Chinese Spring cross. Among these, QLr.hebau-2BS might be the same as LrZH22 in Zhoumai 22 (Wang et al., 2016). QLr.hebau-4B is likely to be the residual resistance of Lr12. QLr.hebau-7DS is Lr34. QLr.hebau-2AL, QLr.hebau-3BS, QLr.hebau-4AL, and QLr.hebau-5BL might be new APR QTL. The genes and closely linked markers can permit accurate selection in marker-assisted wheat breeding for durable resistance to LR.
Ethics Statement
We declare that these experiments comply with the ethical standards in China.
Author Contributions
PZ prepared phenotypic data in the field and drafted the initial manuscript. GY, YZ, and AQ prepared some phenotypic data and prepared some tables and figures. FG prepared genotypic data. XX and ZH played a major role in planning this study and drafted some sections of the manuscript. ZL and DL led the design and coordination of this study.
Funding
This study was supported by National Natural Science Foundation of China (31361140367, 31571662, and 31601299), Natural Science Foundation of Hebei Province (C2014204113 and C2016104013), Natural Science Research Project of Provincial Higher Education in Hebei province (QN2016316), and Graduate Student Innovation project of Hebei Province (1099009).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
We would like to thank Prof. R. A. McIntosh (Plant Breeding Institute, University of Sydney) for the critical review of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00793/full#supplementary-material
Abbreviation
APR, adult plant resistance; HR, hypersensitive response; IT, infection type; LOD, logarithm of odds; LR, leaf rust; MAS, marker-assisted selection; MDS, maximum disease severity; Pt, Puccinia triticina; PVE, phenotypic variance explained; QTL, quantitative trait locus/loci; RIL, recombinant inbred line; SNP, single nucleotide polymorphism; SSR, simple sequence repeat.
References
Bansal, U. K., Hayden, M. J., Venkata, B. P., Khanna, R., Saini, R. G., and Bariana, H. S. (2008). Genetic mapping of adult plant leaf rust resistance genes Lr48 and Lr49 in common wheat. Theor. Appl. Genet. 117, 307–312. doi: 10.1007/s00122-008-0775-6
Chu, C. G., Friesen, T. L., Xu, S. S., Faris, J. D., and Kolmer, J. A. (2009). Identification of novel QTLs for seedling and adult plant leaf rust resistance in a wheat doubled haploid population. Theor. Appl. Genet. 119, 263–269. doi: 10.1007/s00122-009-1035-0
Curtis, T., and Halford, N. G. (2014). Food security: the challenge of increasing wheat yield and the importance of not compromising food safety. Ann. Appl. Biol. 164, 354–372. doi: 10.1111/aab.12108
Dubin, H. J., Johnson, R., and Stubbs, R. W. (1989). Postulated genes for resistance to stripe rust in selected CIMMYT and related wheats. Plant Dis. 73, 472–475. doi: 10.1094/PD-73-0472
Dyck, P. L. (1977). Genetics of leaf rust reaction in three introductions of common wheat. Can. J. Genet. Cytol. 19, 711–716. doi: 10.1139/g77-077
Gao, F. M., Wen, W. E., Liu, J. D., Rasheed, A., Yin, G. H., Xia, X. C., et al. (2015). Genome-wide linkage mapping of QTL for yield components, plant height and yield-related physiological traits in the Chinese wheat cross Zhou 8425B/Chinese Spring. Front. Plant Sci. 6:1099. doi: 10.3389/fpls.2015.01099
Herrera-Foessel, S. A., Lagudah, E. S., Huerta-Espino, J., Hayden, M. J., Bariana, H. S., Singh, D., et al. (2011). New slow-rusting leaf rust and stripe rust resistance genes Lr67 and Yr46 in wheat are pleiotropic or closely linked. Theor. Appl. Genet. 122, 239–249. doi: 10.1007/s00122-010-1439-x
Herrera-Foessel, S. A., Singh, R. P., Huerta-Espino, J., Rosewarne, G. M., Periyannan, S. K., Viccars, L., et al. (2012). Lr68: a new gene conferring slow rusting resistance to leaf rust in wheat. Theor. Appl. Genet. 124, 1475–1486. doi: 10.1007/s00122-012-1802-1
Krattinger, S. G., Lagudah, E. S., Spielmeyer, W., Sing, R. P., Huerta-Espino, J., McFadden, H., et al. (2009). A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323, 1360–1363. doi: 10.1126/science.1166453
Li, C. L., Bai, G. H., Chao, S. A. M., and Wang, Z. H. (2015). A high-density SNP and SSR consensus map reveals segregation distortion regions in wheat. Biomed Res. Int. 2015:830618. doi: 10.1155/2015/830618
Li, H. H., Ye, G. Y., and Wang, J. K. (2007). A modified algorithm for the improvement of composite interval mapping. Genetics 175, 361–374. doi: 10.1534/genetics.106.066811
Li, Z. F., Lan, C. X., He, Z. H., Singh, R. P., Rosewarne, G. M., Chen, X. M., et al. (2014). Overview and application of QTL for adult plant resistance to leaf rust and powdery mildew in wheat. Crop Sci. 54, 1907–1925. doi: 10.2135/cropsci2014.02.0162
Li, Z. F., Xia, X. C., He, Z. H., Li, X., Zhang, L. J., Wang, H. Y., et al. (2010). Seedling and slow rusting resistance to leaf rust in Chinese wheat cultivars. Plant Dis. 94, 45–53. doi: 10.1094/PDIS-94-1-0045
Lillemo, M., Asalf, B., Singh, R. P., Huerta-Espino, J., Chen, X. M., He, Z. H., et al. (2008). The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theor. Appl. Genet. 116, 1155–1166. doi: 10.1007/s00122-008-0743-1
Long, D. L., and Kolmer, J. A. (1989). A North American system of nomenclature for Puccinia recondita f. sp. tritici. Phytopathology 79, 525–529.
Maccaferri, M., Mantovani, P., Tuberosa, R., DeAmbrogio, E., Giuliani, S., Demontis, A., et al. (2008). A major QTL for durable leaf rust resistance widely exploited in durum wheat breeding programs maps on the distal region of chromosome arm 7BL. Theor. Appl. Genet. 117, 1225–1240. doi: 10.1007/s00122-008-0857-5
McDonald, B. A., and Linde, C. (2002). Pathogen population genetics, evolutionary potential, and durable resistance. Annu. Rev. Phytopathol. 40, 349–379. doi: 10.1146/annurev.phyto.40.120501.101443
McIntosh, R. A., Dubcovsky, J., Rogers, W. J., Morris, C., Appels, R., and Xia, X. C. (2016). Catalogue of Gene Symbols for Wheat: 2015-2016 Supplement. Available at: http://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2015.pdf
Messmer, M. M., Seyfarth, R., Keller, M., Schachermayr, G., Winzeler, M., Zanetti, S., et al. (2000). Genetic analysis of durable leaf rust resistance in winter wheat. Theor. Appl. Genet. 100, 419–431. doi: 10.1007/s001220050055
Peterson, R. F., Campbell, A. B., and Hannah, A. E. (1948). A diagrammatic scale for rust intensity on leaves and stems of cereals. Can. J. Res. 26, 496–500. doi: 10.1139/cjr48c-033
Piech, J., and Supryn, S. (1978). Location of adult-plant leaf rust resistance on chromosome 7D in Chinese Spring wheat. Cereal Res. Commun. 6, 367–375.
Qi, A. Y., Zhang, P. P., Zhou, Y., Yao, Z. J., Li, Z. F., and Liu, D. Q. (2016). Mapping of QTL conferring leaf rust resistance in Chinese wheat lines W014204 and Fuyu 3 at adult plant stage. J. Integr. Agric. 15, 18–28. doi: 10.1016/S2095-3119(14)60974-6
Röder, M. S., Korzun, V., Wendehake, K., Plaschke, J., Tixier, M. H., Leroy, P., et al. (1998). A microsatellite map of wheat. Genetics 149, 2007–2023.
Roelfs, A. P., Singh, R. P., and Saari, E. E. (1992). Rust Diseases of Wheat: Concepts and Methods of Disease Management. Texcoco: CIMMYT.
Roncallo, P. F., Cervigni, G. L., Jensen, C., Miranda, R., Carrera, A. D., Helguera, M., et al. (2012). QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 185, 77–92. doi: 10.1007/s10681-012-0628-x
Rosewarne, G. M., Singh, R. P., Huerta-Espino, J., Herrera-Foessel, S. A., Forrest, K. L., Hayden, M. J., et al. (2012). Analysis of leaf and stripe rust severities reveals pathotype changes and multiple minor QTLs associated with resistance in an Avocet × Pastor wheat population. Theor. Appl. Genet. 124, 1283–1294. doi: 10.1007/s00122-012-1786-x
Schnurbusch, T., Paillard, S., Schori, A., Messmer, M., Schachermayr, G., Winzeler, M., et al. (2004). Dissection of quantitative and durable leaf rust resistance in Swiss winter wheat reveals a major resistance QTL in the Lr34 chromosome region. Theor. Appl. Genet. 108, 477–484. doi: 10.1007/s00122-003-1444-4
Sharp, P. J., Kreis, M., Shewry, P. R., and Gale, M. D. (1988). Location of β-amylase sequence in wheat and its relatives. Theor. Appl. Genet. 75, 286–290.
Singh, A., Pallavi, J. K., Gupta, P., and Prabhu, K. V. (2012). Identification of microsatellite markers linked to leaf rust resistance gene Lr25 in wheat. J. Appl. Genet. 53, 19–25. doi: 10.1007/s13353-011-0070-0
Singh, D., Simmonds, J., Park, R. F., Bariana, H. S., and Snape, J. W. (2009). Inheritance and QTL mapping of leaf rust resistance in the European winter wheat cultivar ‘Beaver’. Euphytica 169, 253–261. doi: 10.1007/s10681-009-9959-7
Singh, R. P., Mujeeb-Kazi, A., and Huerta-Espino, J. (1998). Lr46: a gene conferring slow-rusting resistance to leaf rust in wheat. Phytopathology 88, 890–894. doi: 10.1094/PHYTO.1998.88.9.890
Singh, S., and Bowden, R. L. (2011). Molecular mapping of adult-plant race-specific leaf rust resistance gene Lr12 in bread wheat. Mol. Breed. 28, 137–142. doi: 10.1007/s11032-010-9467-4
Somers, D. J., Isaac, P., and Edwards, K. (2004). A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 109, 1105–1114. doi: 10.1007/s00122-004-1740-7
Wang, C. F., Yin, G. H., Xia, X. C., He, Z. H., Zhang, P. P., Yao, Z. J., et al. (2016). Molecular mapping of a new temperature-sensitive gene LrZH22 for leaf rust resistance in Chinese wheat cultivar Zhoumai 22. Mol. Breed. 36:18. doi: 10.1007/s11032-016-0437-3
William, H. M., Singh, R. P., Huerta-Espino, J., Palacios, G., and Suenaga, K. (2006). Characterization of genetic loci conferring adult plant resistance to leaf rust and stripe rust in spring wheat. Genome 49, 977–990. doi: 10.1139/G06-052
Yin, G. H., Wang, J. W., Wen, W. E., He, Z. H., Li, Z. F., Wang, H., et al. (2009). Mapping of wheat stripe rust resistance gene YrZH84 with RGAP markers and its application. Acta Agron. Sin. 35, 1274–1281. doi: 10.3724/SP.J.1006.2009.01274
Yu, H. H., Xie, W. B., Wang, J., Xing, Y. Z., Xu, C. G., Li, X. H., et al. (2011). Gains in QTL detection using an ultra-high density SNP map based on population sequencing relative to traditional RFLP/SSR markers. PLoS ONE 6:e17595. doi: 10.1371/journal.pone.0017595
Zhang, L. J., Li, Z. F., Lillemo, M., Xia, X. C., Liu, D. Q., Yang, W. X., et al. (2009). QTL mapping for adult-plant resistance to leaf rust in CIMMYT wheat cultivar Saar. Sci. Agric. Sin. 42, 388–397.
Zhao, X. L., Zheng, T. C., Xia, X. C., He, Z. H., Liu, D. Q., Yang, W. X., et al. (2008). Molecular mapping of leaf rust resistance gene LrZH84 in Chinese wheat line Zhou 8425B. Theor. Appl. Genet. 117, 1069–1075. doi: 10.1007/s00122-008-0845-9
Zhou, H. X., Xia, X. C., He, Z. H., Li, X., Wang, C. F., Li, Z. F., et al. (2013). Molecular mapping of leaf rust resistance gene LrNJ97 in Chinese wheat line Neijiang 977671. Theor. Appl. Genet. 126, 2141–2147. doi: 10.1007/s00122-013-2124-7
Keywords: APR, Puccinia triticina, single nucleotide polymorphism, simple sequence repeat marker, Triticum aestivum
Citation: Zhang P, Yin G, Zhou Y, Qi A, Gao F, Xia X, He Z, Li Z and Liu D (2017) QTL Mapping of Adult-Plant Resistance to Leaf Rust in the Wheat Cross Zhou 8425B/Chinese Spring Using High-Density SNP Markers. Front. Plant Sci. 8:793. doi: 10.3389/fpls.2017.00793
Received: 15 February 2017; Accepted: 27 April 2017;
Published: 16 May 2017.
Edited by:
Chengdao Li, Murdoch University, AustraliaReviewed by:
Daniela Marone, Centre of Cereal Research (CREA-CER), ItalyMeixue Zhou, University of Tasmania, Australia
Copyright © 2017 Zhang, Yin, Zhou, Qi, Gao, Xia, He, Li and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zaifeng Li, bHpmNzU1MUBhbGl5dW4uY29t Daqun Liu, bGRxQGhlYmF1LmVkdS5jbg==