 Lysiane Brocard1†
Lysiane Brocard1† Françoise Immel2,3†
Françoise Immel2,3† Denis Coulon2,3,4
Denis Coulon2,3,4 Nicolas Esnay2,3Karine Tuphile2,3Stéphanie Pascal2,3Stéphane Claverol5
Nicolas Esnay2,3Karine Tuphile2,3Stéphanie Pascal2,3Stéphane Claverol5 Laëtitia Fouillen2,3
Laëtitia Fouillen2,3 Jean-Jacques Bessoule2,3
Jean-Jacques Bessoule2,3 Claire Bréhélin2,3*
Claire Bréhélin2,3*- 1Plant Imaging Platform, Bordeaux Imaging Center, UMS 3420 Centre National de la Recherche Scientifique, US4 Institut National de la Santé et de la Recherche Médicale, University of Bordeaux, Bordeaux, France
- 2Laboratory of Membrane Biogenesis, Centre National de la Recherche Scientifique, UMR 5200, Villenave d'Ornon, France
- 3Laboratory of Membrane Biogenesis, University of Bordeaux, UMR 5200, Villenave d'Ornon, France
- 4Bordeaux INP, Talence, France
- 5Proteome Platform, Functional Genomic Center of Bordeaux, University of Bordeaux, Bordeaux, France
Lipid droplets (LDs) are cell compartments specialized for oil storage. Although their role and biogenesis are relatively well documented in seeds, little is known about their composition, structure and function in senescing leaves where they also accumulate. Here, we used a label free quantitative mass spectrometry approach to define the LD proteome of aging Arabidopsis leaves. We found that its composition is highly different from that of seed/cotyledon and identified 28 proteins including 9 enzymes of the secondary metabolism pathways involved in plant defense response. With the exception of the TRIGALACTOSYLDIACYLGLYCEROL2 protein, we did not identify enzymes implicated in lipid metabolism, suggesting that growth of leaf LDs does not occur by local lipid synthesis but rather through contact sites with the endoplasmic reticulum (ER) or other membranes. The two most abundant proteins of the leaf LDs are the CALEOSIN3 and the SMALL RUBBER PARTICLE1 (AtSRP1); both proteins have structural functions and participate in plant response to stress. CALEOSIN3 and AtSRP1 are part of larger protein families, yet no other members were enriched in the LD proteome suggesting a specific role of both proteins in aging leaves. We thus examined the function of AtSRP1 at this developmental stage and found that AtSRP1 modulates the expression of CALEOSIN3 in aging leaves. Furthermore, AtSRP1 overexpression induces the accumulation of triacylglycerol with an unusual composition compared to wild-type. We demonstrate that, although AtSRP1 expression is naturally increased in wild type senescing leaves, its overexpression in senescent transgenic lines induces an over-accumulation of LDs organized in clusters at restricted sites of the ER. Conversely, atsrp1 knock-down mutants displayed fewer but larger LDs. Together our results reveal that the abundancy of AtSRP1 regulates the neo-formation of LDs during senescence. Using electron tomography, we further provide evidence that LDs in leaves share tenuous physical continuity as well as numerous contact sites with the ER membrane. Thus, our data suggest that leaf LDs are functionally distinct from seed LDs and that their biogenesis is strictly controlled by AtSRP1 at restricted sites of the ER.
Introduction
In plant cells, neutral lipids constitute a storable source of energy that accumulates in two different compartments: plastoglobules and cytosolic lipid droplets, also known as oil bodies. In contrast to plant-specific plastoglobules, cytosolic lipid droplets (LDs) are described in every organism, from bacteria to human (Yang et al., 2012). LDs are dynamic organelles controlling neutral lipid metabolism and storage. As such, they participate in many critical cellular pathways including lipid transport between organelles (Bartz et al., 2007), protein metabolism (Cermelli et al., 2006; Welte, 2015), or protein degradation (Ohsaki et al., 2006; Welte, 2007; Farese and Walther, 2009; Moldavski et al., 2015). Consequently, dysfunctions in LD biology lead to several metabolic human diseases (Krahmer et al., 2013). In plants, lipid droplets are essential for reproduction by storing oil in seeds, which will be used as source of energy during the post-germinative growth until acquisition of photo-autotrophism. In leaves, LDs accumulate during senescence and transiently store lipids such as steryl esters and triacylglycerol (TAG) derived from the dismantlement of membranes (Chapman et al., 2013; Troncoso-Ponce et al., 2013; Shimada et al., 2015). Yet, the composition, structure and functions of lipid droplets present in vegetative organs, such as leaves or roots (Murphy, 2001; Lersten et al., 2006) remain elusive.
LDs and plastoglobules share a similar structure. They are composed of a central core of neutral lipids, mainly TAGs and lipid-esters (steryl-ester and phytyl ester for LDs and plastoglobules, respectively), covered by a monolayer of polar lipids. While polar lipids found in plastoglobules directly derive from the outer leaflet of the thylakoid membrane, that of LDs derive from the endoplasmic reticulum (ER) membrane (Austin et al., 2006; Jacquier et al., 2011; Kassan et al., 2013; Choudhary et al., 2015). Studies in yeast and animal cells as well as developing seedlings provided current models for LDs biogenesis (reviewed in Chapman et al., 2012; Wilfling et al., 2014). Lipid droplets can arise from previously existing LDs or be formed de novo. During de novo formation, neutral lipids are synthesized at the ER and subsequently accumulate between the two leaflets of one ER membrane forming lens-like structures that gradually bud from the membrane. LDs will eventually detach from the ER, however it is still unclear whether this last step is systematic or not (Jacquier et al., 2011; Hashemi and Goodman, 2015). In plant leaves, the mechanism for LD biogenesis is not documented. Yet, based on what is found in seeds and other organisms, it is assumed that they derive from the ER.
The characterization of the seed LD proteome has brought crucial information toward the understanding of their structure and functions. To date, less than a dozen proteomic studies have been conducted on purified seed lipid droplets of diverse species (Jolivet et al., 2013; Liu et al., 2015), that identified between 8 and 30 proteins. Oleosins are systematically associated with seed lipid droplets; these proteins are involved in the maintenance of the LD structure, notably by preventing their coalescence during seed maturation. Oleosins are major proteins in LDs of reproductive organs such as seeds and stamens, but are absent from vegetative tissues. For example, in Arabidopsis, 16 oleosin isoforms are present, but none of the encoding genes is expressed in leaves (Kim et al., 2002). Thus, other proteins must play structural functions in vegetative lipid droplets. Caleosins are other LD structural proteins described in seeds as well as in vegetative tissues. Their implication in the maintenance of lipid droplet structure has mainly been demonstrated by in vitro assays on artificial lipid droplets (Liu et al., 2009; De Domenico et al., 2011); they are also involved in fatty acid degradation or oxylipin synthesis via their peroxygenase activity (Chapman et al., 2012). Other proteins frequently identified in seed lipid droplets include steroleosins that are involved in sterol and brassinosteroid metabolism. In fact, diverse proteomic approaches of seed LDs identified proteins involved in lipid metabolism, in particular in TAG mobilization during seed germination, as well as in stress response, intracellular transport, and protein biosynthesis (reviewed in Jolivet et al., 2013). In contrast to the seed LDs, the comprehensive protein composition of leaf LDs remains unresolved and only a few proteins have been described to localize at these compartments (Aubert et al., 2010; Shimada and Hara-Nishimura, 2015). Shimada et al. (2014) showed that the caleosin AtCLO3/RD20/PGX3 (At2g33380—thereafter simplified in AtCLO3) interacts with an α-dioxygenase (α-DOX1—At3g01420) in Arabidopsis leaf lipid droplets. This interaction catalyzes the formation of an antifungal phytoalexin, implying a role for leaf lipid droplets in plant defense against fungi. In addition, the TAG lipase Sugar Dependent1 (SDP1), which initiates oil breakdown during germination, localizes to cotyledon lipid droplets during germination (Eastmond, 2006; Thazar-Poulot et al., 2015). However, this localization was described in germinating seedlings, and is therefore associated with seed lipid droplets rather than vegetative lipid droplets from true leaves. During the process of our study, two manuscripts, published concomitantly, demonstrated the localization of a new class of proteins, named either AtSRP for Small Rubber Particle or LDAP for Lipid Droplet Associated Proteins in the LDs of Arabidopsis cotyledon and leaf (Gidda et al., 2016; Kim et al., 2016). These proteins share similarities with the Hevea brasiliensis SRPP (Small rubber particle protein) proteins which participate in the maintenance of the structure of rubber particles (Hillebrand et al., 2012), and were previously identified in LDs of the avocado mesocarp (Horn et al., 2013). Gidda et al. (2016) suggested that, during post-germinative growth, the SRP proteins replace oleosins at the LD surface, which are degraded during germination. Yet, while oleosins are highly abundant proteins in seed LDs, the level of SRP proteins in vegetative LDs remains to be determined.
The abundance of leaf LDs dramatically increases during senescence (Shimada et al., 2015). Yet, apart from their role in lipid storage and their implication in the production of an antifungal phytoalexin (Shimada et al., 2014; Shimada and Hara-Nishimura, 2015), little is known about their composition and functions at this developmental stage. Understanding the structure and biogenesis of LDs in senescing leaves will therefore shed new light onto their role in plant aging. In fact, besides SRP, CLO3, and α-DOX1, it is most likely that other proteins associate to lipid droplets, in particular in senescent leaves. We reasoned that a quantitative and qualitative profile of the LD proteome in this tissue would bring new details into the molecular requirements for LD formation and functions. We thus purified vegetative LDs from Arabidopsis aging leaves and performed a label-free quantitative proteomic approach to determine the proteome of these organelles. Proteins were defined as component of the LD core proteome when they were found enriched in the LD fraction compared to any other cell fraction, whatever their abundances within the LD fraction. We found that the protein composition of leaf LDs is quite different from that of seed LDs. In particular, it is strictly deprived of enzymes involved in lipid metabolism. By determining the abundance of each protein within this LD core proteome, we show that the two most abundant proteins in aging leaf lipid droplets are the previously characterized caleosin AtCLO3, and one protein of the small rubber particle protein family, AtSRP1/LDAP1 (At1g67360). In addition, our proteomics data suggest a new role of aging leaf lipid droplets in several pathways for secondary metabolite biosynthesis and plant resistance to stress, notably herbivory and pathogen resistances. Using laser scanning confocal and electron microscopy, we demonstrate that AtSRP1 is sufficient to induce lipid droplet biogenesis in leaf at restricted sites of the ER, and necessary to regulate LD size and number. We took advantage of this property to establish electron tomography of cryofixed leaf LDs and demonstrate that they share numerous contact sites as well as some continuity with the ER membrane.
Materials and Methods
Plant Material and Culture Conditions
For LD preparation, Arabidopsis thaliana ecotype Columbia (Col-0) were grown at high density on soil in a phytotron in long day conditions (16 h light/8 h dark), 100 μmol of photon.m−2.s−1 at 20–22°C and 65% humidity. For nitrogen starvation assays, 10-day-old plants grown on half strength Murashige and Skoog (MS) medium were transplanted on synthetic minimal medium deprived of nitrogen [0.8% Agarose, 1% sucrose, 1 mM MgSO4, 1 mM KH2PO4, 25 μM Fe-EDTA, 35 μM H3BO3, 7 μM MnCl2, 0.25 μM CuSO4, 0.5 μM ZnSO4, 0.1 μM Na2MoO4, 5 μM NaCl, 5 nM CoCl2, 1 mM CaCl2, and 2.5 mM KCl] (Gaude et al., 2007) and grown for 6–7 additional days. For lipid analysis, seeds were first germinated on half strength MS medium, and then transplanted in individual pots in growth chamber in long day conditions, at 20–22°C and 40% humidity. The atsrp1_1 (SALK_148081) and atsrp1_2 (GK-309G05) T-DNA mutants were obtained from the ABRC seed stock center, and homozygous lines were selected by PCR genotyping (primer sequences are provided in Table S1). For transient transformation, Nicotiana benthamiana leaves or 4-day-old A. thaliana seedlings were infiltrated with Agrobacterium tumefaciens cell cultures as described in Perraki et al. (2014) or Marais et al. (2015) respectively.
Cloning and Transgenic Plants
ORFs clones of AtSRP1 (U17438) and AtULP (U15698) in pENTR vector were obtained from ABRC clone center and verified by sequencing. AtULP clone contained a non-silent mutation (L63 → M63) that was corrected by PCR quick-change site directed mutagenesis. ORFs with or without the stop codon were subsequently transferred by GATEWAY® recombinational cloning technology in diverse destination vectors under the 35S promoter in pGWB661 and pGWB660 (Nakamura et al., 2010), and under the UBQ10 promoter in pUBNYFP or pUBCYFP (Grefen et al., 2010) to obtain TagRFP-AtSRP1, AtSRP1-TagRFP, YFP-AtSRP1, and AtSRP1-YFP constructs respectively. The resulting plasmids were introduced into Arabidopsis plants by an Agrobacterium transfection method (Clough and Bent, 1998). Two or three confirmed lines, numbered #a to c, were used for the different experiments. Sequences of primers used for cloning are given in Table S1.
Preparation of Lipid Droplet Fractions
For lipid droplet purification, c.a. 150 g of 6-week-old A. thaliana leaves were homogenized in cold G-20 buffer (100 mM Tricine (pH7.5), 10 mM KCl, 1 mM EDTA, 1 mM PMSF, plant protease inhibitor cocktail (Sigma), and 20% sucrose) using a Waring blender. The homogenized plant material was filtered through two layers of cheese cloth and one layer of Miracloth (Calbiochem) and split into three equal portions that were processed in parallel to obtain three technical replicates. A scheme representing the purification process is provided in Figure S1. Each portion of the filtrate was centrifuged for 30 min at 10,000 rpm. The floating lipid pad was kept, and the soluble fraction centrifuged at 100,000 g for 30 min. The floating lipid pad was collected and pooled with the previous one. The pellet and the soluble fraction above the floating lipid pad were used for isolation of membrane and soluble proteins respectively. The pooled floating lipid pads were resuspended in 10 mL G-20 buffer using a Potter homogenizer, distributed in 2 centrifuge tubes, covered with 2 mL of G-15 buffer (G-buffer with 15% sucrose) and 5 mL of G-5 buffer (G buffer with 5% sucrose), and centrifuged 50 min at 150,000 g. The two floating lipid pads were pooled, resuspended in 5 mL G-20 buffer, and centrifuged similarly with G-15 and G-5 buffers. Finally, each floating lipid pad was resuspended in 5 mL G-20 buffer with a Potter homogenizer, covered with 7 mL G-0 buffer (without sucrose), and centrifuged 50 min at 150,000 g. The three resulting floating lipid pads from the three starting portions were stored at −80°C. Plastoglobules were purified according to the protocol described by Besagni et al. (2011).
Protein Analysis
Proteins were extracted according to Rensink et al. (1998) and concentrated by chloroforme-methanol precipitation (Wessel and Flugge, 1984). For immunoblot experiments, proteins were separated by SDS-PAGE (Sodium Dodecyl Sulfate—PolyAcrylamide Gel Electrophoresis) and blotted onto nitrocellulose membrane for immunodetection. Blots were probed with an antibody raised against GFP (Roche), or sera raised against plastoglobule, ER, Golgi or thylakoid markers, respectively AtFBN1b (Vidi et al., 2006), BiP (Höfte and Chrispeels, 1992), AtMemb11 (Marais et al., 2015), or P16 (Vallon et al., 1986). Proteins were quantified either according to the Bradford method (Bradford, 1976) or the instructions of the QuantiPro BCA assay kit manual (Sigma).
Label-Free Quantitative Data Analysis
Proteins were identified by mass spectrometry and relatively quantified by a label-free approach (for detailed description see Appendix S1). Ten microgram of each fraction were loaded onto a 10% acrylamide SDS-gel. Trypsin digested peptide mixture was analyzed on a Ultimate 3000 nanoLC system (Dionex, Amsterdam, The Netherlands) coupled to a LTQ-Orbitrap XL mass spectrometer (Thermo Fisher Scientific, San Jose, USA). Data were searched by SEQUEST through Proteome Discoverer 1.4 (Thermo Fisher Scientific, San Jose, USA) against the TAIR (The Arabidopsis Information Resource) protein database. Raw LC-MS/MS data were imported in Progenesis QI 2.0 (Nonlinear Dynamics Ltd, Newcastle, UK). Protein abundance was calculated according to two methods: when comparing protein abundance between samples the protein abundance was calculated based on peptide uniqueness, as preconized by the Progenesis tool, while the TOP3 method (sum of the three most abundant peptides), which allows an absolute quantification of the protein (Silva et al., 2006), was chosen to compare abundance of the different proteins in a given sample. If the same set of peptides could be used to identify several different proteins, these proteins were grouped and counted as one protein for quantification. The relative normalized abundance of each protein in a single sample was calculated from its normalized abundance relatively to the sum normalized abundance of all the peptides in the sample. Average relative normalized abundances were calculated based on the triplicates of the enrichment ratios between LD and each of the other fractions calculated for each protein. Quantitative data were considered for proteins quantified by a minimum of 2 peptides. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Vizcaíno et al., 2016) partner repository with the dataset identifier PXD006113.
Lipid Analysis
Lipids were extracted and purified according to Folch et al. (1957). The lipids were then resuspended in chloroform:methanol (2:1, v/v) solution and applied onto a silica-coated chromatography plate (Merck) with lipid standards. Lipids were separated with a single (leaf lipid composition) or double (purified LD analysis) thin layer chromatography (TLC): polar lipids were first separated with a Vitiello-Zanetta solvent mixture (Vitiello and Zanetta, 1978). The neutral lipids were then separated according to Juguelin et al. (1986). The lipids were visualized with primuline under UV light. After fatty acid trans-esterification, lipids were quantified by gas chromatography (Agilent 7890 gas chromatograph equipped with a Carbowax column, Alltech Associates, Deerfield, IL, USA) coupled to a flame ionization detector (GC-FID).
Confocal Microscopy and BODIPY493/503 Coloration
Observations were performed on a confocal microscope Leica TCS SP2. When needed, leaf tissues were stained with a neutral lipid-specific fluorescent dye, BODIPY493/503 (MolecularProbes), at a final concentration of 1 μg/mL for 5 min in the dark. The acquisitions with objective X63 were taken sequentially on each channel with the following acquisition settings: excitation wavelength of 514 nm and emission of 520–550 nm band pass for YFP, excitation wavelength of 543 nm and emission of 600–650 nm band pass for TagRFP, excitation wavelength of 514 nm and emission filter of 520–540 nm band pass for BODIPY493/503, excitation wavelength of 514 nm and emission of 655–690 nm or 695–750 nm band pass for chlorophyll autofluorescence. Stack image deconvolution was achieved with Autoquant software with blind PSF mode and five iterations.
TEM Imagery and Tomography
Leaves were chemically fixed by 2.5% glutaraldehyde in phosphate Buffer 0.1 M pH 7.2 overnight at 4°C and post-fixed by 1% OsO4 for 2 h at room temperature. After dehydration, inclusion in SPURR (Electron Microscopy Science) resin was progressively performed. To finish, the samples were polymerized at 70°C for 16 h. Cryofixation was performed on the first leaves of 6-day-old plants. The samples were high pressure frozen (HPF) with an EM-PACT1 (Leica) device and 20% BSA in half MS medium was used as cryoprotectant. Freeze-substitution steps were achieved in an AFS2 (Leica) system. Samples were first incubated at −90°C into a glutaraldehyde 0.5%, osmium tetroxide 2%, uranyl acetate 0.1% in pure acetone for 48 h. For IEM (Immuno electron microscopy), osmium tetroxyde and glutaraldehyde were omitted. Then a progressive raise of the temperature of 3°C/h was initiated until −50°C is reached, and samples were progressively incubated in HM20 Lowicryl resin (Electron Microscopy Science) and polymerized under UV for 48 h at −50°C followed by 48 h at 20°C. Ultrathin sections of 70–80 nm and 180 nm thickness were made with ultramicrotome Leica EM UC7. IEM was made according to Grison et al. (2015). GFP antibodies (Tp401, Torrey Pines Biolabs) were diluted 1/500 before immuno-gold-labeling with anti-rabbit conjugated with 5 nm colloidal gold (Tebu) diluted 1/30. The transmission electron microscope FEI Tecnai G2 Spirit TWIN 120 kV equipped with a CCD 16 Mpixels Eagle 4k camera was used for acquisition (120 kV). ImageJ software (Schneider et al., 2012) was used for image analysis. For electron tomography, FEI software was used for tilt series acquisition. Alignement and backprojection steps were respectively achieved with IMOD and tomoJ softwares. OSART algorithm with 100 iterations was used.
RNA Extraction and Quantitative RT-PCR
RNAs were extracted with the RNeasy Plant Mini kit (Qiagen) on green parts of plants, treated by DNAse (Turbo DNA-free™ kit, Ambion) and quantified with the NanoDrop 2000 system (Thermo Scientific). Reverse transcriptase reaction was carried out on 143 ng of extracted RNA with SSII Reverse Transcriptase (Invitrogen). For quantitative PCR, three reference expression genes were used: actin (At1g49240), GAPDH (At1g13440) and eIF4A-1 (At3g13920). The sequence of the primers is provided in Table S1. Three wells per condition were quantified and efficiency test was performed on primers. Transcript abundance in samples was determined using a comparative cycle threshold (Ct) method on the CFX96 Touch™ Real-Time PCR Detection System (Bio-Rad). The relative abundance of the three genes of reference in each sample was determined and used to normalize for differences of total RNA level according to the method described by Vandesompele et al. (2002).
Results and Discussion
Purification of Vegetative LDs from Arabidopsis Aging Leaves
LDs have been described to accumulate in senescing leaves (Parker and Murphy, 1981; Shimada et al., 2015), therefore we chose to harvest aging leaves, from 6 to 7 week old Arabidopsis plants as starting material for the isolation of leaf LDs. Arabidopsis of this age, grown at high density, display a mix of green, purple, red and yellow leaves reminiscent of a stress and senescent phenotype (Figure 1A1). The accumulation of LDs was tested by transmission electron microscopy on chemically fixed leaves (Figures 1A2–A4). Parenchymal cells of aging leaves accumulated, per cell section, 1–7 lipid droplets from 400 to 1,600 nm diameter (with an average of c.a. 820 nm) while parenchyma plants of 3 week old green leaves exhibited very little to no LD. In addition to cytosolic LDs, we observed an accumulation of large plastoglobules (ca. 310 nm) in plastids, which was more pronounced in red leaves. This is consistent with previous studies showing that plastoglobules are also known to accumulate in response to a variety of stress as well as during senescence (Nacir and Bréhélin, 2013).
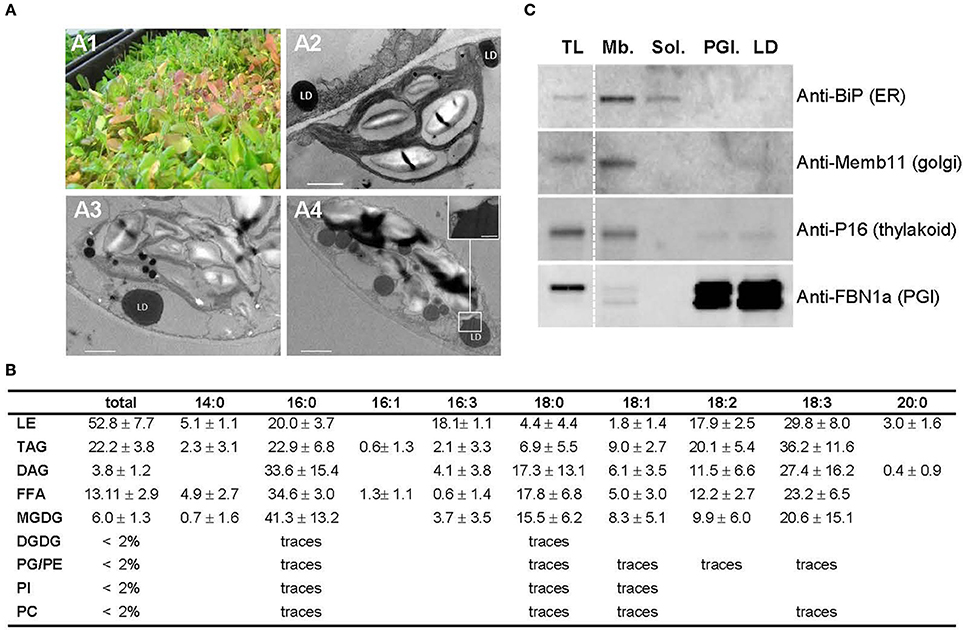
Figure 1. Preparation of aging leaf lipid droplet fraction. (A) Six week old Arabidopsis leaves accumulate lipid droplets. (A1) 6 week old leaves grown under long day conditions; (A2–A4): details of the ultrastructure observed by TEM of a parenchyma cell from green (A2), purple (A3), and red (A4) leaves showing lipid droplet in close proximity to chloroplast. Insert in (A4) shows the plastid—LD contact site. Scale bar: 1 μm, except in insert: 200 nm. LD, lipid droplet. (B) Lipid composition of Arabidopsis leaf lipid droplets (in % of total fatty acids) n = 5. Lipids were extracted from LD fraction, separated by thin layer chromatography, and quantified by GC-FID after transesterification. LE, lipid ester; TAG, triacylglycerol; DAG, Diacylglycerol; FFA, free fatty acid; MGDG, monogalactosyldiacylglycerol; DGDG, digalactosydiacylglycerol; PG, phosphatidylglycerol; PE, phosphatidylethanolamine; PI, phosphatidylinositol; PC, phosphatidylcholine. (C) Immunoblot analysis of cell fractions using marker antibodies. Proteins for total leaf (TL), membranous (Mb.), soluble (Sol.), plastoglobules (PGl.), and lipid droplet (LD) fractions were precipitated and 10 μg from each fraction were loaded on SDS-PAGE. After transfer to nitrocellulose, proteins were probed with sera raised against Binding immunoglobulin protein (BiP) as ER marker, Membrin 11 (Memb11) as Golgi marker, P16 as thylakoid marker, and Fibrillin 1a (FBN1a) as plastoglobule marker. Two lines of the blots were cropped (dashed line) for clarity reason, but brightness and contrast balance were applied to every pixels of the whole blots before cropping.
LDs were purified by successive centrifugations on sucrose gradients (see Figure S1). Because no antibody specific to leaf LDs was available to test the quality of the resulting fraction by immunoblotting, we determined by TLC GC-FID the lipid composition of the final fraction to confirm the presence of lipid droplets. The LD fraction contained, as expected, mainly neutral lipids with 53% of total fatty acids (FAs) in lipid esters, 22% of FAs in TAG and 13% in free fatty acids (FFAs), and less than 5% of FAs in total phospholipids (Figure 1B). We also detected minor quantities of MGDGs in our LD fraction. MGDGs are lipids typically associated with plastid membranes; yet their composition in LDs differed from that of chloroplast MGDGs, in particular in respect to C16:3 (3.7% in LDs compared to 35% in plastid MGDG, Ajjawi et al., 2011), which is an acyl moieties commonly considered as exclusively plastidial (Li-Beisson et al., 2013). Therefore, only 10% of the MGDG present in LD could be explained by a chloroplast contamination, if any, and a major contamination of the LD sample by plastid membranes can be excluded. In addition, the proportion of phospholipids to neutral lipids in our purified fraction was similar to that of seed LDs (Huang, 1992), demonstrating that it was mainly constituted by lipid droplets and contained, if any, only traces of other organelles. To further test the purity of the LD fraction, we performed immunoblot analyses using markers of the most probable contaminants: Golgi vesicles that may float similarly to the LDs, ER membranes that could stay attached to LDs, and thylakoids, the most abundant membranes in leaves. As expected, the LD fraction did not show any signal with Golgi and ER markers (Figure 1C). A faint signal was observed with the thylakoid marker, suggesting a weak contamination by thylakoid membranes. However, the lipid analysis demonstrated that possible contaminating membranes are far less abundant than neutral lipids in the LD fraction. Furthermore, lipid esters in the LD fraction were constituted of 18.1% C16:3 in addition to 20.0% of C16:0, 17.9% C18:2, and 29.8% C18:3 (Figure 1B), while Arabidopsis leaf steryl esters do not contain any C16:3 (Wewer et al., 2011). On the contrary, fatty acid phytyl esters (FAPEs), described to accumulate in plastoglobules are constituted by 50% of C16:3 and neither C18:2 nor C18:3 (Gaude et al., 2007). This suggests a contamination of the LD fraction by FAPEs originating from plastoglobules. We therefore performed immunoblot assays with a serum raised against the fibrillin/plastoglobulin AtFBN1a/AtPGL35 (At4g04020), a plastoglobule marker (Vidi et al., 2007). The LD fraction showed a signal equivalent to the one obtained with purified plastoglobules. It confirmed that our LD fraction contains plastoglobules in addition to cytosolic lipid droplets. This can be easily explained since plastoglobules have a density similar to LDs, and are also purified by floatation on a sucrose gradient (Besagni et al., 2011). During the purification process chloroplasts were inevitably broken, releasing plastoglobules that were then co-purified with LDs. The plastoglobule proteome has already been well described (Vidi et al., 2006; Ytterberg et al., 2006; Lundquist et al., 2012), with a core proteome of 34 proteins (Nacir and Bréhélin, 2013) that can manually be removed from the LD core proteome, therefore we choose to performed the proteomic analysis on this fraction enriched in LDs and plastoglobules, and deprived of any other major contaminants.
Defining the Leaf LD Proteome
In order to establish precisely the leaf LD proteome, we quantified protein abundance by label free quantitative proteomics, and determined the enrichment ratio of each protein in the LD fraction compared to other cell fractions. This should allow eliminating any protein unspecifically detected in LDs (for example contaminant proteins highly abundant in other cell fractions) and identifying with certainty proteins specific to the LD fraction. Arabidopsis leaf LDs were purified from three independent 6 week old cultures. In addition, fractions enriched with soluble or membrane proteins were also prepared from the same starting material. From each independent culture, three technical replicates were analyzed, coming from three gradients performed in parallel (for LD, soluble and membrane fractions) or three different leaves (for total leaf extracts). Proteins within each fraction were thereafter identified and quantified by label free quantitative proteomics. The data obtained are provided in Table S2. Some proteins with a same ontology were grouped under a unique identifier when they share identical peptide sets. To be able to compare the abundance of a protein between the different fractions, we followed a protocol similar to the one previously described by Lundquist et al. (2012) for the identification of the plastoglobule core proteome. First, the relative mass contribution (relative normalized abundance) of each protein within a fraction was calculated from its normalized abundance quantified by the label free method relatively to the sum normalized abundance of all the peptides in the fraction. The averages of relative normalized abundances from triplicates of the same experiment were determined. Second, the enrichment factor between LD and each of the other fractions was obtained for each protein by calculating the ratios of the average of relative normalized abundance in LD to other fractions (see Figures S2, S3 for graphical explanation). Next, these ratios were analyzed according to several criteria to establish a list of proteins enriched in the LD fraction: (1) the LD/total leaf enrichment ratio had to be superior to 6; this threshold was chosen deliberately low (given that the total protein amount in an entire cell is much more than 6 times more abundant than the LD total protein amount of a cell), in order to minimize the risk to exclude major LD proteins that could partition between LDs and other compartments, but sufficient to remove all abundant proteins such as subunits of the photosystem complexes. (2) Proteins with a ratio of LD to soluble proteins and LD to membrane proteins inferior to 1.5 were discarded. This enrichment ratio was chosen as low as possible in order to obtain the most possible exhaustive list of proteins. The resulting data identified plastoglobule proteins in the LD fraction as suspected from our immunoblot analyses, the most abundant being the fibrillins AtFBN4 (At3g23400) and AtFBN1b (At4g04020). Purified plastoglobules highly impoverished in LDs can easily be obtained because plastoglobules are isolated from purified chloroplasts. In order to clearly exclude contaminating plastoglobule proteins from our LD proteome, we purified plastoglobules from the material used for LD purification in one of the three replicates, and the LD/plastoglobule enrichment ratio was determined for each protein. Thirty two of the thirty four plastoglobule proteins defined in Nacir and Bréhélin (2013) were identified both in the plastoglobule and LD fractions (Table S3). In this assay, the known plastoglobule protein with the highest LD/plastoglobule ratio was the kinase ABC1K3 with a ratio of 3.99. Thus proteins with LD/plastoglobule enrichment ratio less than 4 were considered as non-LD proteins but rather plastoglobule ones, and were thus discarded.
Thirty three proteins or groups of proteins matching all the aforementioned criteria in at least two of the three independent experiments were considered enriched in LDs (Table 1). We further excluded five proteins with a previously characterized plastidial localization and/or function (Table S4). Among the 28 proteins assigned to the core LD proteome (Table 1), three were already known to associate with leaf LDs: Aubert et al. (2010) described the co-localization of the caleosin AtCLO3 with lipid droplets in onion epidermal cells and the accumulation of the corresponding transcripts in leaves, especially in response to drought stress. We also identified the peroxidase α-DOX1 that interacts with AtCLO3 in leaf LDs (Shimada et al., 2014), and AtSRP1, one of the three members of the AtSRP/LDAP family (Gidda et al., 2016; Kim et al., 2016) is also present in our proteome. The identification of these already known LD proteins in the core proteome validated our approach. Twenty three other proteins were classified into seven biological processes according to the MapMan tool provided by the Bio-Analytic Resource for Plant Biology (http://bar.utoronto.ca/), UniProt Gene Ontology (Consortium, 2015) and KEGG BRITE database (Kanehisa et al., 2016): cytoskeleton proteins (3), transport (6), stress response (3), biosynthesis of secondary metabolites (6), biosynthesis of amino acids (2), cell wall modification (1), and peptidases (2), while two proteins remained unclassified: an unknown protein (At5g16550) and a HAD superfamily acid phosphatase.
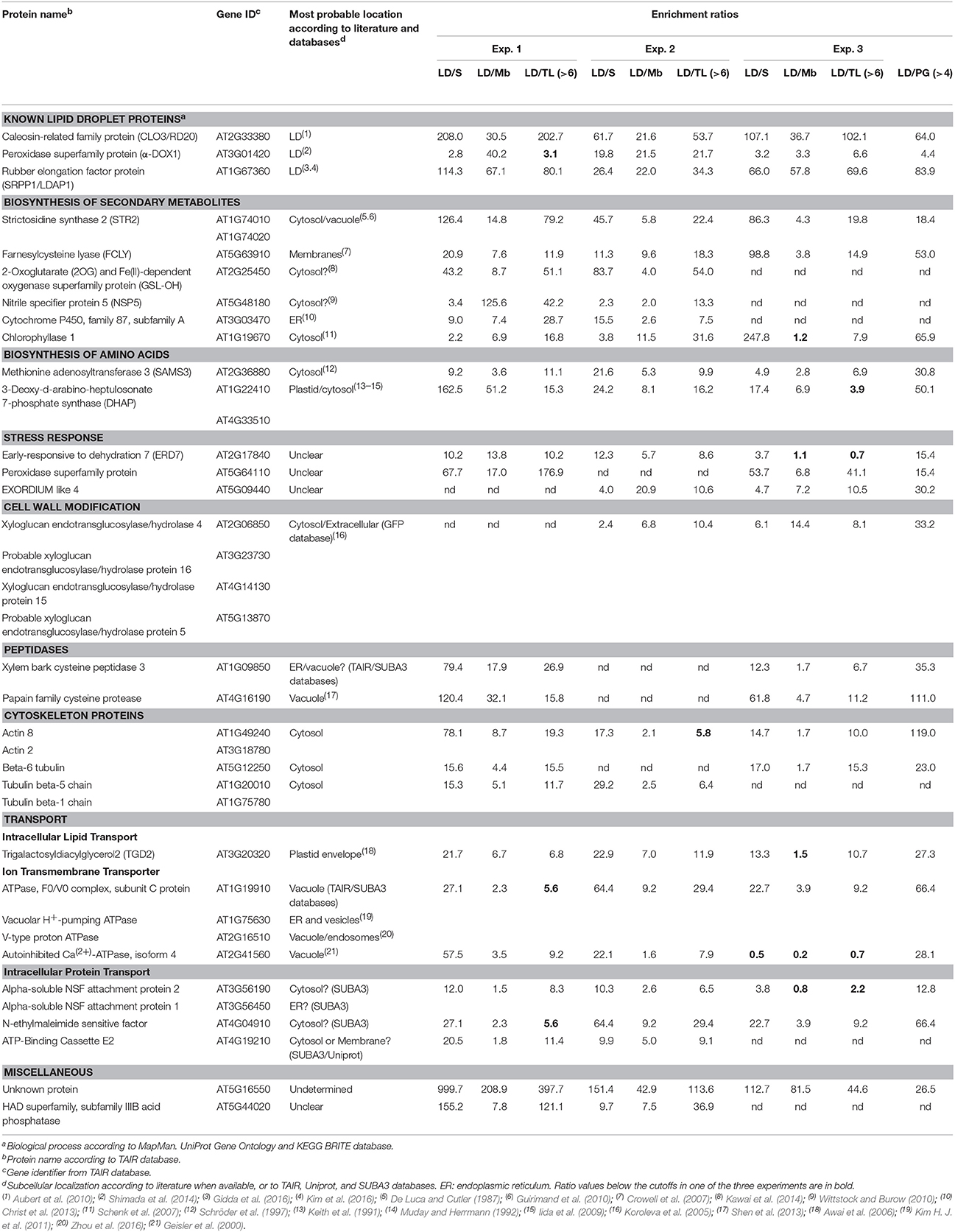
Table 1. Aging leaf LD core proteome determined by label free quantitative comparative proteomics.
To further validate our proteomic approach we tested the localization of another protein highly enriched in LDs according to our proteomics data. We transiently expressed fluorescent fusions of this protein of unknown function, At5g16550, which we choose to call AtULP for Unknown Lipid droplet Protein, either alone in Arabidopsis cotyledons, or in N. benthamiana leaves co-transformed with AtSRP1 fluorescent constructs. The localization of the fluorescent constructs was then observed by confocal laser scanning microscopy. In Arabidopsis cotyledons AtULP, fused to TagRFP at its N-terminal end, co-localized with cytosolic particles stained with the neutral lipid-specific fluorescent dye, BODIPY493/503 (MolecularProbes), suggesting that these structures are lipid rich bodies (Figure 2A). When transiently co-expressed with AtSRP1-YFP constructs in N. benthamiana leaves, the AtULP-TagRFP fluorescence was localized in the same particles as AtSRP1-YFP (Figure 2B), a protein recently shown to be targeted to LDs (Gidda et al., 2016; Kim et al., 2016). Taken together, these results demonstrate the reliability of the LD core proteome identity established here.
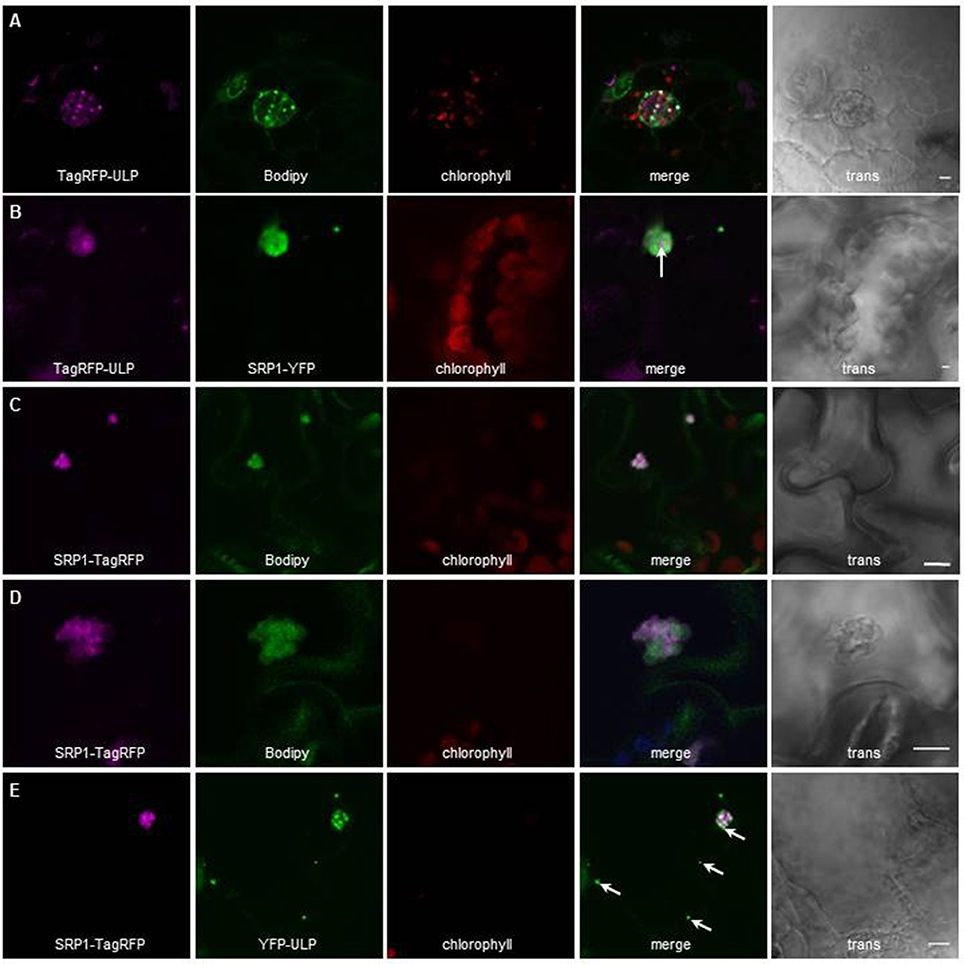
Figure 2. Localization of AtULP and AtSRP1-fluorescent fusions in lipid droplets. Arabidopsis cotyledons (A,C–E), or N. benthamiana leaves (B) expressing TagRFP-AtULP (A,B) AtSRP1-YFP (B), AtSRP1-TagRFP (C–E), or YFP-AtULP (E) were visualized by confocal laser scanning microscopy. Arabidopsis plantlets were co-labeled with neutral lipid specific dye BODIPY493/503(A,C,D). Co-labeling of AtULP or AtSRP1 targeted structures by BODIPY493/503 confirmed that these structures are lipid droplets. Bodipy, chlorophyll, and trans indicate Bodipy fluorescence, chlorophyll autofluorescence and transmission microscopy image respectively. Merge indicates merge of TagRFP, Bodipy and chlorophyll fluorescences (A,C,D) or YFP and TagRFP fluorescence (B,E). Bar: 5 μm. White arrows indicate specific localization of AtULP in cells expressing both AtSRP1 and AtULP transgenes.
Leaf LDs Contain Enzymes of Secondary Metabolite Pathways Involved in Resistance against Pathogens
Among the proteins identified here as core components of the aging leaf LD proteome we found three cytoskeleton proteins, which reflects the dynamism of LD intracellular trafficking (Welte, 2009). Such proteins were previously identified in seed (reviewed in Jolivet et al., 2013) and Chlamydomonas reinhardtii (Tsai et al., 2015) LDs and were recently demonstrated to participate in the recruitment and shape maintenance of LDs in zebrafish (Dutta and Kumar Sinha, 2015). It is highly probable that LDs associate to the cytoskeleton network to transit between organelles in leaves, thus allowing lipid transfer between the ER, plastids and peroxisome. Two proteins of the SNARE (Soluble N-ethylmaleimide-sensitive factor activating protein receptor) system, N-ethylmaleimide sensitive factor (NSF- At4g04910) and alpha soluble NSF attachment protein 2 (At3g56190) were also identified in the core proteome. SNARE proteins are suggested to transit from the plasma membrane or endosomes to LDs in mammalian cells and to be essential for LD growth (Boström et al., 2007). Additionally, the presence of vacuolar ATPase in the core proteome may reflect contacts between LDs and the vacuole. In yeast, LDs can be targeted to vacuoles to be degraded by a process resembling microautophagy (Vevea et al., 2015). Nevertheless, the abundance of these ATPases in leaf LDs is very low, thus their genuine association with LDs remains to be confirmed.
Surprisingly, the LD core proteome contains nine proteins involved in various secondary metabolite pathways associated with plant response to biotic stress (Figure 3). In addition to AtCLO3 and α-DOX1 involved in oxylipin metabolism (Shimada et al., 2014), we identified three proteins which directly participate in mechanisms for plant resistance to herbivores and pathogens: strictosidine synthase (STR) 2-oxoglutarate and Fe(II)-dependent oxygenase (GSL-OH) and nitrile specifier protein5 (NSP5). In addition, we found the farnesylcysteine lyase (FCLY) that contributes to cell detoxification via the recycling of prenylated proteins (Crowell et al., 2007) as well as enzymes of the amino acid metabolism: the 3-deoxy-d-arabino-heptulosonate 7-phosphate (DAHP) synthase that participates in the shikimate pathway, and a methionine adenosyltransferase 3 (SAMS3). The potential functional relevance of these proteins is discussed thereafter.
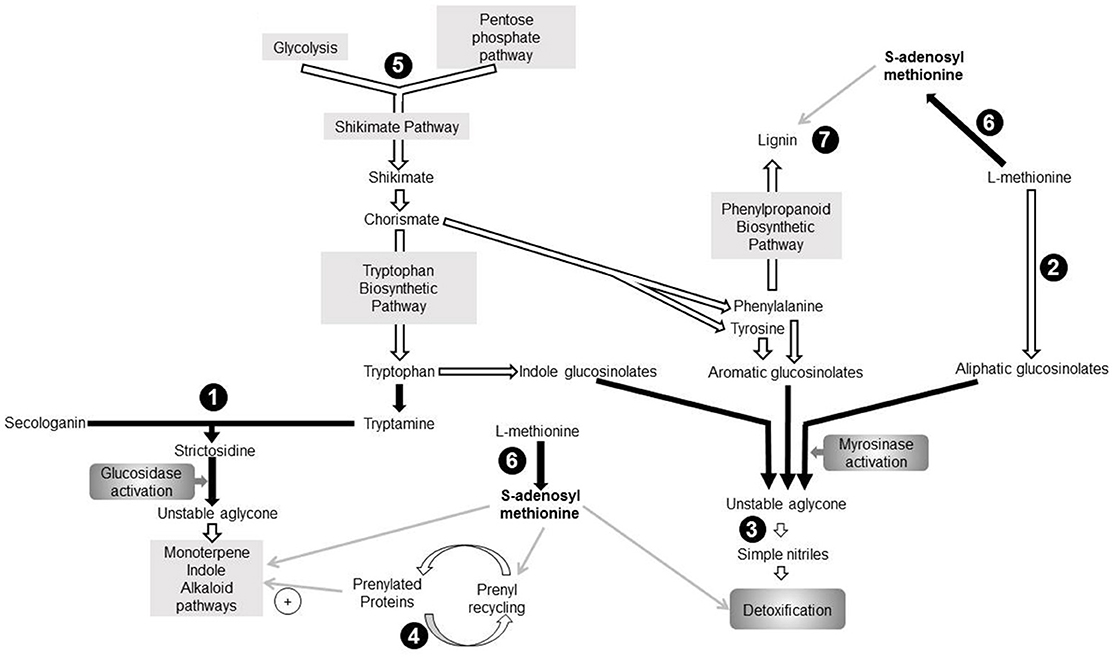
Figure 3. Seven proteins identified in the aging leaf lipid droplet (LD) core proteome participate to secondary metabolism pathways. Enzymes identified in leaf LDs are represented in black circles. Pathways are represented by gray rectangles. Black arrows indicate direct reactions, white arrows represent indirect reactions, and gray arrows depict involvement of one component in a pathway. (1): 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase (DAHP; AT1G22410), (2): Strictosidine synthase (AT1G74010; AT1G74020), (3): Farnesylcysteine lyase (AT5G63910), (4): Methionine adenosyltransferase3 (AT2G36880), (5): 2-oxoglutarate (2OG) and Fe(II)-dependent oxygenase superfamily protein (AT2G25450), (6): Nitrile specifier protein5 (AT5G48180), (7): Peroxidase superfamily protein (Peroxidase70; AT5G64110). DMAPP, Dimethylallyl diphosphate; IPP, Isopentenyl diphosphate; MEP, Methyl-D-erythritol 4-phosphate; MIA, Monoterpenoid indole alkaloid biosynthetic pathway; PP, Diphosphate.
STR participates to the “nuclear time bomb” for defense against herbivory (Guirimand et al., 2010) and catalyzes the synthesis of strictosidine (St-Pierre et al., 1999), thereafter activated in an unstable aglycone (Figure 3). This aglycone is a precursor of monoterpene indole alkaloid (MIA) biosynthesis pathway (Rischer et al., 2006; Barleben et al., 2007). MIAs are critical for plant defense against biotic stress, and possess a wide range of pharmaceutical properties and applications. The subcellular localization of STR is controversial, and its exact localization in A. thaliana remains elusive. It is predicted to be extracellular by the SUBAtool (Hooper et al., 2014), but subcellular fractionation of Catharanthus roseus leaves showed that strictosidine synthase was cytoplasmic (De Luca and Cutler, 1987) whereas fluorescent microscopy pointed to a vacuolar localization (Guirimand et al., 2010). The enrichment of STR in the proteome of LDs suggests that strictosidine synthesis occurs at these organelles, the LD thus providing storage for the molecules before their subsequent activation.
The “mustard oil bomb” or glucosinolate-myrosinase defense system implies either aliphatic, aromatic or indole glucosinolates. The GSL-OH protein catalyzes one of the last steps of aliphatic glucosinolate biosynthesis (Figure 3), and the expression of the corresponding gene correlates with the accumulation of aliphatic glucosinolates in leaves (Hansen et al., 2008). Furthermore, the accumulation of 2-hydroxybut-3-enyl glucosinolate synthesized by this GSL-OH in leaves provides an increased resistance to insect herbivory. All plant GSL-OH are predicted to be cytosolic (Kawai et al., 2014). Upon biotic or abiotic stress, glucosinolates are hydrolysed into an unstable aglycone which is converted in different active products such as isothyocianates, thiocyanates or simple nitriles. NSP5 was shown to convert unstable aglycones into simple nitriles in vitro (Kissen and Bones, 2009; Kong et al., 2012). A. thaliana nitrile specifier proteins are predicted to be cytosolic (Wittstock and Burow, 2010). Here we point to a LD colocalization of an enzyme catalyzing the synthesis of glucosinolate (GSL-OH) with NSP5. This could represent a security system favoring the synthesis of simple nitrile rather than breakdown compounds with a higher toxicity. An FCLY activity was detected in membrane fraction of whole A. thaliana extract (Crowell et al., 2007), suggesting that it is not specifically localized in LDs but may partition between both compartments. Interestingly, a prenyl cysteine lyase has been identified in human lipoproteins (Banfi et al., 2009), suggesting that the hydrophobic environment procured by lipoproteins or LDs may provide adequate conditions for lipidated cystein recycling. Prenylated proteins are involved in response to biotic and abiotic stress (Galichet and Gruissem, 2003; Crowell and Huizinga, 2009), thus indirectly linking FCLY and plant stress response. Furthermore, some prenylated proteins also regulate the MIA biosynthetic pathway in C. roseus (Courdavault et al., 2009).
Our work also reveals DAHP as part of the LD proteome of aging leaves, a protein with a potential role in plant response to stress as both A. thaliana isoforms respond to wounding and/or pathogen attacks (Keith et al., 1991). DAHP synthases catalyze the first stage of shikimate biosynthesis which yields the three aromatic amino acids incorporated into diverse secondary metabolites, such as alkaloids, glucosinolates, or phenylpropanoid, a precursor of lignin (Maeda and Dudareva, 2012). In addition to the well-established plastidial shikimate pathway, some plants also possess cytosolic isoforms of the enzymes (Maeda and Dudareva, 2012). For example, Muday and Herrmann (1992) identified two DAHP synthase isoenzymes in Solanum tuberosum with cytosolic or plastidial localizations. Even though both A. thaliana DAHP synthases displayed a chloroplast transit peptide, it was suggested that an alternative transcription start site of transcript of DAHP synthase At4g33510 exists, possibly generating a truncated variant without N-terminal chloroplast transit peptide (Keith et al., 1991; Iida et al., 2009) that could target LDs.
SAMS3, another LD-enriched protein found in our study, catalyzes the production of S-adenosyl-L-methionine (SAM) which participates in multiple pathways (Roje, 2006), including some described hereinabove (Figure 3). (i) SAM is involved in the MIA biosynthetic pathway (Fahn et al., 1985). (ii) SAM participates in the detoxification of glucosinolate sub-products. As mentioned before, the glucosinolate catabolism generates a variety of toxic breakdown products involved in plant defense against biotic stress. In the presence of SAM, thiol methyltransferases catalyze the methylation of these compounds into less toxic products associated with airborne plant-insect signals and plant-pathogen interactions (Attieh et al., 2000, 2002). (iii) SAM is involved in the recycling of prenylated proteins: when isoprenylated proteins transit throught the ER, the prenylcysteine is methylated by an isoprenylcysteine methyltransferase with SAM as methyl donor. (iv) SAMS3 is involved in lignin biosynthesis during which SAM is a substrate for multiple methylations (Campbell and Sederoff, 1996; Shen et al., 2002). Lignin itself is synthesized from products of the shikimate pathway (Maeda and Dudareva, 2012) and deposited at the cell wall in response to stress (Le Gall et al., 2015). In C. roseus, the 3 SAMS isoenzymes localize in the cytoplasm and their microenvironment seems to participate in their specialization (Schröder et al., 1997). In this context, we propose that the presence of SAMS3 in LDs of aging leaves procure close proximity with other afore-mentioned enzymes thus, enabling the corresponding reactions. In other words, in senescing tissue, the LD would provide a platform where the concentration of enzymes of the secondary pathways will allow a fast and coordinated response to a variety of biotic stress. Our model thus reinforce the idea proposed by Shimada et al. (2014, 2015) for LDs and phytoalexins: during senescence, leaf LDs participate to plant defense by accumulating a mixture of compounds with antimicrobial properties or enzymes involved in the synthesis of such compounds.
Leaf LDs Are Deprived of Lipid Metabolism Enzymes
In contrast to the LD proteomes of seeds (Jolivet et al., 2013), or microalgae (Goold et al., 2015), we did not identified enzymes involved in sterol or TAG metabolism in leaf LDs. This suggests that (i) the breakdown of TAG or steryl ester does not occur at LDs in aging leaves, on the contrary to what is described in germinating seeds (Eastmond, 2006); (ii) Leaf LDs do not grow by local lipid synthesis. Instead, LD growth would occur by direct transfer of neutral lipids from the ER or by LD fusion via, for example, the SNARE system. In leaves of AtSRP1 overexpressing lines, we observed LDs in contact with or in close proximity to, the ER membranes (see below). This suggests that leaf LDs are not isolated from the ER, but rather share a connection that would allow for a rapid transfer of molecules, such as lipids. TGD2 was the only enzyme involved in lipid metabolism identified in the core proteome of LDs. Located at the inner membrane of the plastid envelope, TGD2 is part of a transporter complex involved in lipid transfer from the ER to the plastid (Awai et al., 2006). Its presence in LDs is unlikely originating from contamination by chloroplast membranes, since we did not find any other abundant proteins from this compartment. Furthermore, TGD2 was also described in the LD proteome of Chlamydomonas reinhardtii, where it was proposed to originate from contact sites established between LDs and plastid envelope (Tsai et al., 2015). Interestingly we often observed similar contacts by TEM in aging leaves (Figure 1A4), supporting a genuine association of TGD2 with leaf LDs. However, additional studies are required to further substantiate and understand how this inner envelope membrane protein could be associated with cytosolic LDs.
Aging Leaf LDs Are Mainly Composed of the Caleosin AtCLO3 and the Small Rubber Particle Protein AtSRP1
In order to estimate the abundance of each protein constituting the leaf LD proteome, a TOP-3 quantitative analysis was carried out on the nine replicates of the LD fraction (Table 2). This reliable method was previously used in diverse label free quantitative proteomic approaches (Ahrné et al., 2013; Nikolovski et al., 2014). It is based on the correlation of the sum intensity of the three most intense peptides of a protein with its abundance in the sample (Silva et al., 2006). Results from this analysis showed that the two most abundant proteins of aging leaf LDs are the caleosin AtCLO3 and the small rubber particle protein AtSRP1, together constituting more than 70% of the LD protein mass. Seed LDs are described to contain several oleosins and caleosins to maintain the LD structure. However, in leaf LDs only one isoform of caleosin, AtCLO3, was detected. AtCLO3 is believed to play structural role in LDs (Chapman et al., 2012). In addition AtCLO3 has a peroxigenase activity and is involved in oxylipin biosynthesis together with α-DOX1 (Blée et al., 2014; Shimada et al., 2014). Interestingly, we found that AtCLO3 is over a 100 times more abundant than α-DOX1 in the leaf LD proteome. This suggests that only a few AtCLO3 molecules are engaged in a complex with α-DOX1, enabling the major part of AtCLO3 to play a role toward structure maintenance or other functions. AtCLO4, another caleosin, was previously reported to localize in leaf LDs (Kim Y. Y. et al., 2011), yet we did not find it enriched in our LD proteome. This may reflect that AtCLO4 is not specifically enriched in LDs but partitions with other compartments as suggested by the localization pattern of AtCLO4-GFP fusion (Kim Y. Y. et al., 2011). Alternatively, endogenous AtCLO4 may not be present in aging leaves, since it was not detected in total leaf extracts. In addition to AtCLO3, we found that AtSRP1 is highly enriched in the LD proteome of aging leaves. AtSRP1 is part of the small rubber particle family, constituted of 3 members, which was recently characterized to localize in vegetative LDs (Gidda et al., 2016; Kim et al., 2016). Interestingly, we only detected AtSRP1 in our proteome, suggesting that this protein has an unsuspected specific role in aging leaves compared to AtSRP2 and AtSRP3.
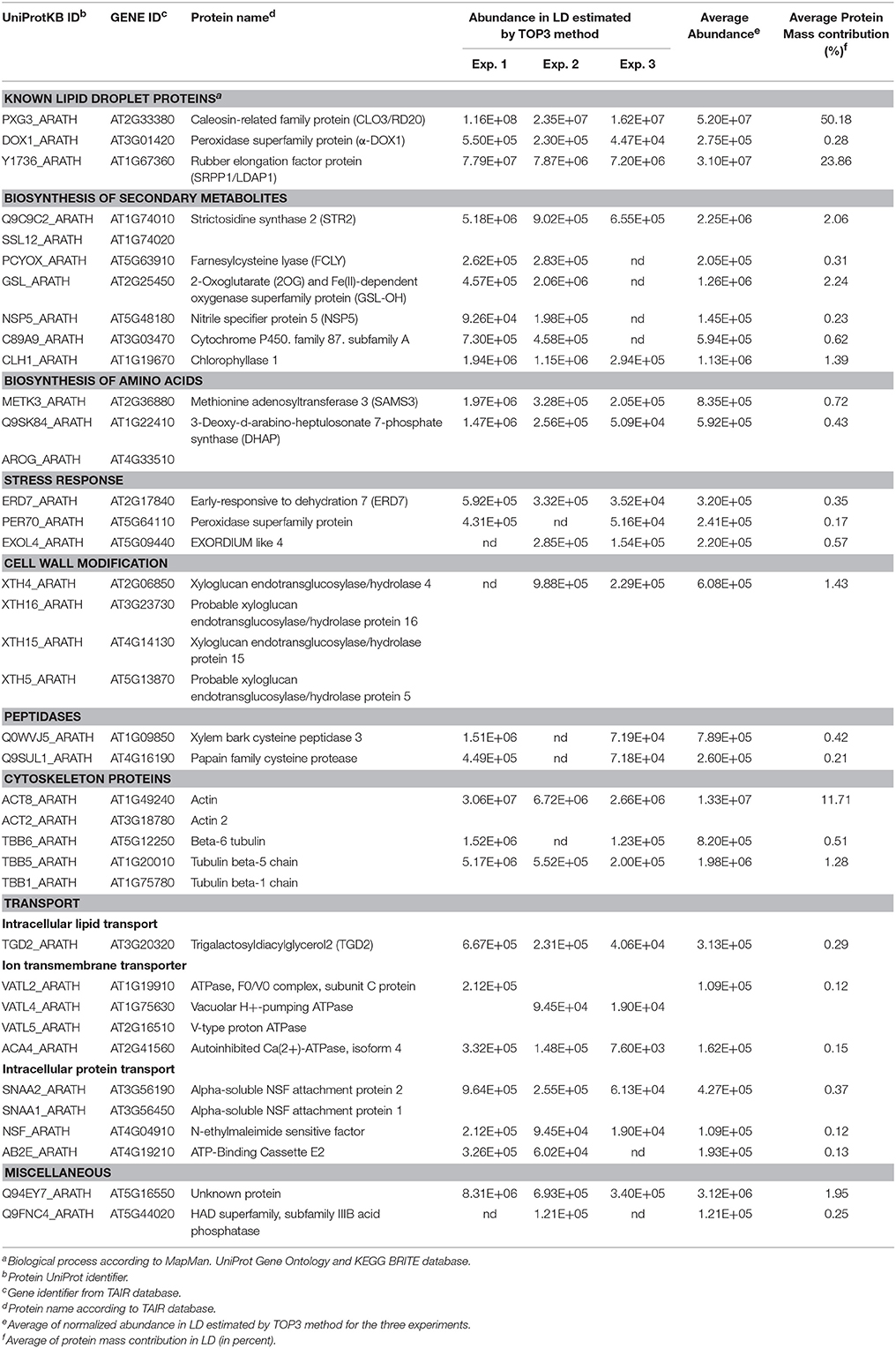
Table 2. Protein abundance in aging leaf LD determined by TOP3 method.
The high content of AtSRP1 in aging leaf LDs prompted us to explore its function in these compartments. We thus generated Arabidopsis lines overexpressing AtSRP1 fused to fluorescent tags. The overexpression of the transgenes in leaves was confirmed by quantitative RT-PCR (Figure S4). We also obtained two T-DNA insertional mutants of AtSRP1 from the ABRC. Basal transcript levels of AtSRP1 were detected by quantitative RT-PCR in 4 and 6 week old T-DNA mutants (Figure 4A), demonstrating that they are knock-down rather than knock-out lines. Because AtCLO3 is the most abundant protein and the unique caleosin found in leaf LDs, we compared the expression levels of AtSRP1 and AtCLO3, in knock-down as well as in four independent AtSRP1 over-expressing lines. In 4 and 6 week old leaves, grown in the same conditions used for the proteomic approach, the expression of AtCLO3 strictly follows that of AtSRP1 (Figure 5A, Figure S5). AtCLO3 is up-regulated in AtSRP1 over-expressing lines, and down-regulated in AtSRP1 knock-down lines. This indicates that AtSRP1 is critical for the regulation of AtCLO3. Both caleosin and SRP proteins have been demonstrated to participate in stabilizing artificial oil bodies or inhibiting their coalescence (Chen et al., 2004; Jiang et al., 2009; Berthelot et al., 2012, 2014). Their high content in LDs of aging leaves, as well as the co-regulation of AtCLO3 and AtSRP1 expression, suggest that they both play fundamental role in stabilizing leaf LD structure and that the maintenance of a strict stoichiometry between both proteins may be determinant to preserve the LD structure.
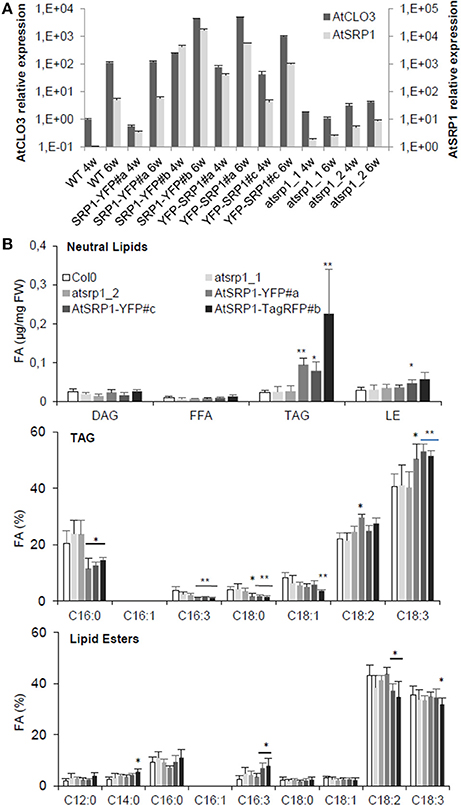
Figure 4. Impact of AtSRP1 misexpression on AtCLO3 expression and neutral lipid metabolism. (A) Relative quantification of AtCLO3 and AtSRP1 expression in wild-type (WT), AtSRP1 overexpressors (SRP1-YFP and YFP-SRP1), and atsrp1 knock-down (atsrp1_1 and atsrp1_2) lines. Relative expression levels of AtSRP1 were determined by quantitative RT-PCR in 4 (4w) and 6 (6w) week old plants, and normalized to three reference genes: actin, GAPDH, and eIF4A-1. Relative expression quantities are represented related to wild type level at 4 weeks, which was set to one. Error bars represent standard deviation of technical replicates. Similar results were obtained when repeated with different plants (see Figure S8). (B) Neutral lipid analysis of AtSRP1 knock-down or overexpressing leaves. Lipids from 6 week-old leaves were quantified by GC-FID after transesterification. Neutral lipid quantification (in μg of fatty acid (FA)/mg fresh weight) is represented in top panel, triacylglycerol (TAG) fatty acid composition (in % of total FA) in middle, and lipid ester fatty acid composition (in % of total FA) in down panel. Values represent average and SD from six biological replicates. Statistically significant differences from the wild type are indicated by *p < 0.05 or **p < 0.01 as determined by Wilcoxon's-test.
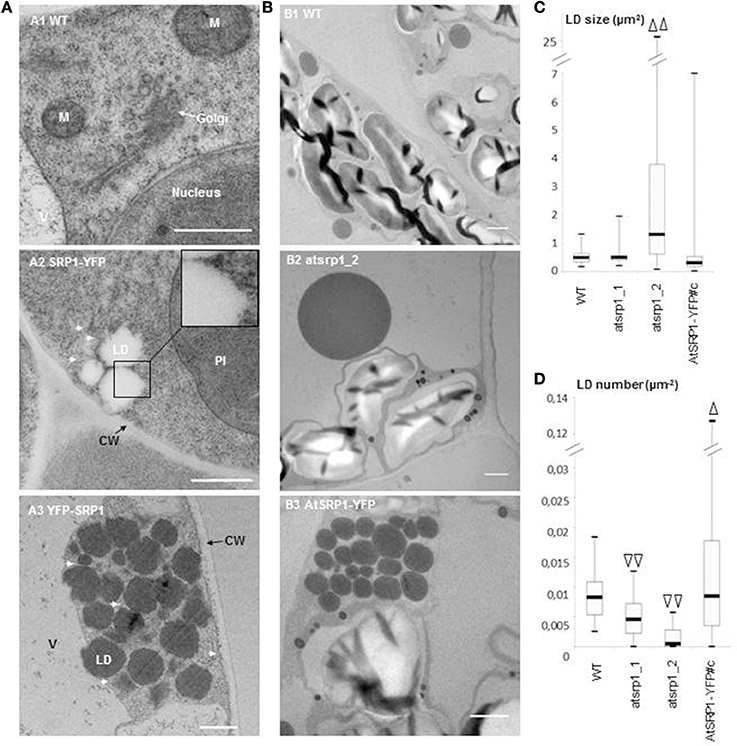
Figure 5. Ultrastructure study of AtSRP1 knock-down and overexpressing leaves. (A) Ultrastructure of lipid droplet (LD) clusters in AtSRP1 overexpressing leaves. Cryo-fixed and cryo-substituted first leaves of 6 day old wild type (A1) or pUB10::AtSRP1-YFP (A2) transgenic plantlets, and chemically fixed leaves of 20 day old pUB10::YFP-AtSRP1 plantlets (A3) were observed by TEM. M, mitochondria; V, vacuole; CW, cell wall. For cosmetic reason, only one LD is indicated among a cluster. White arrows show endoplasmic reticulum membranes in proximity to LDs. Bar = 500 nm. (B–D) Ultrastructure of AtSRP1 misexpressing leaves under nitrogen starvation. Fifteen day-old plantlets were incubated on medium deprived of nitrogen for 5 days. The ultrastructure of the fourth leaf of wild type (B1), atsrp1 knock down (B2), and AtSRP1-YFP overexpressing (B3) plants were observed by TEM after chemical fixation (B). For each line, lipid droplet (LD) area (C) and LD number per cell area (D) were determined thanks to the ImageJ software. Arrowheads show significant differences compared to the wild-type, as determined by Student's test [n > 67 in (C) and n > 36 in (D), Δp < 0.05, and ΔΔp < 0.01]. Measurements of LD area obtained for each individual plant are presented as box-plots in Figure S9. Bar in B = 1 μm. WT: wild-type Columbia 0, atsrp1_1 and atsrp1_2: AtSRP1 knock-down lines, AtSRP1-YFP: AtSRP1 overexpressing lines under pUB10 promoter, with YFP fused at the C-term of AtSRP1.
Different Vegetative LDs Co-exist in Cotyledons and Mature Leaves
Next, we assayed the localization of fluorescent AtSRP1 constructs using confocal laser scanning microscopy. In agreement with Gidda et al. (2016) and Kim et al. (2016), AtSRP1-TagRFP co-localized with cytosolic mobile particles stained with the neutral lipid-specific fluorescent dye BODIPY493/503, suggesting that AtSRP1 localizes to lipid rich bodies (Figure 2C). A similar localization was observed with AtSRP1 constructs fused to YFP or TagRFP either at the C- or N-terminal of the protein. We further observed larger BODIPY-stained AtSRP1 structures present in cotyledon (Figure 2D and Figure S6) and noticed that (i) these large lipid rich bodies are in fact lipid droplet aggregates, and that (ii) the TagRFP fluorescence seemed restricted to the periphery of the lipid droplet. These aggregates of LDs were not observed in wild-type plants, implying that the overexpression of AtSRP1 induces the neo-formation of LD clusters in standard growth conditions.
Similarly to AtSRP1, AtULP, an uncharacterized LD-enriched protein (Figure 2), was visualized in clusters of LDs in agro-infiltrated leaves of N. benthamiana. When co-expressed in the same cell, both targeted the same LD clusters, but not exactly at the same area of the clusters, suggesting that AtULP and AtSRP1 are not systematically targeted to the same LDs (Figure 2B). This difference was even more striking when Arabidopsis cotyledons overexpressing AtSRP1-TagRFP were transiently agro-infiltrated with YFP-AtULP (Figure 2E): YFP-AtULP fluorescence was observed in LD clusters tagged with AtSRP1-TagRFP, but also in some isolated LDs of the same cell untagged by AtSRP1-TagRFP. This suggests that diverse populations of LDs, with differential protein compositions, could co-exist in cotyledons and leaves. In cotyledons, these diverse populations could correspond to a concomitant mix of “seed” LDs derived from the embryo with neo-formed “vegetative” LDs. Nevertheless, their observation in mature leaves of N. benthamiana strongly supports our upraising idea that various specialized LDs coexist in vegetative tissues.
AtSRP1 Overexpression in Leaves Induces the Formation of New LDs Organized in Clusters in Contact or in Continuity with the ER
To gain insight into the LD ultrastructure, we performed transmission electron microscopy (TEM) after cryo-fixation of AtSRP1 overexpressing Arabidopsis leaves. This analysis showed an accumulation of round-shaped cytosolic aggregates, which were never observed in wild-type leaves (Figure 5A2). To confirm the nature of these clusters, immunogold labeling with an antibody specific to the GFP/YFP protein was performed on cryo-fixed roots of a YFP-AtSRP1 overexpressing line. Labeling was restricted to these cytosolic aggregates (Figure S7), while no such specific labeling was observed in control experiments where anti-GFP/YFP antibody was omitted. Therefore, our data indicate that the YFP-AtSRP1 protein is specifically associated with these aggregates. These structures were not surrounded by a typical bilayer membrane but a monolayer (insert Figure 5A2), suggesting that they are not vesicles or small vacuoles. Similar clusters homogeneously stained by osmium tetroxide were observed in chemically fixed leaves of YFP-AtSRP1 overexpressing lines (Figure 5A3). The colocalization of AtSRP1-TagRFP protein with the neutral lipid specific dye BODIPY493/503 (Figure 2), together with the homogeneous staining of the aggregates by osmium tetroxide when traditional chemical fixation was used, confirmed that these structures are filled with neutral lipids. Moreover, these data confirm that the overexpression of AtSRP1 is responsible for the formation of such LD clusters.
LD clusters were often observed in the close vicinity of the ER membranes (Figures 5A2,A3). In order to obtain a better vision of the position of LDs in the ER network, N. benthamiana leaves were transiently co-transformed with pUB10::AtSRP1-YFP and a fluorescent marker of the ER, HDEL-mCherry (Nelson et al., 2007). As shown in Figure 6A, the transient expression of AtSRP1 in tobacco leaves was sufficient to induce the formation of LD clusters similar to that observed in stably transformed Arabidopsis. These LD clusters were mobile, likely following cytosolic or ER streaming (Ueda et al., 2010). YFP tagged LD clusters appeared not only in contact to mCherry fluorescent ER membranes, but even trapped in the ER network, showing a multitude of contact sites between LDs and ER membranes (Figure 6A and Movie S1). Unfortunately, the confocal microscopy resolution is not sufficient to resolve the AtSRP1-ER localization and determine if AtSRP1 is exclusively present in LD periphery or also at ER-LD biogenesis sites. However, results of our proteomic approach, as well as of our IEM experiment (Figure S7) clearly argue for a preferential, if not exclusive, localization of AtSRP1 at LDs. Chapman et al. (2012) proposed that in non-seed tissues LDs originate from small LDs formed all along the ER, “pinched off form the ER, and then fused to form larger droplets.” However, the cluster organization of LDs in AtSRP1 overexpressing lines rather suggests that LDs are formed at restricted, specialized areas of the ER.
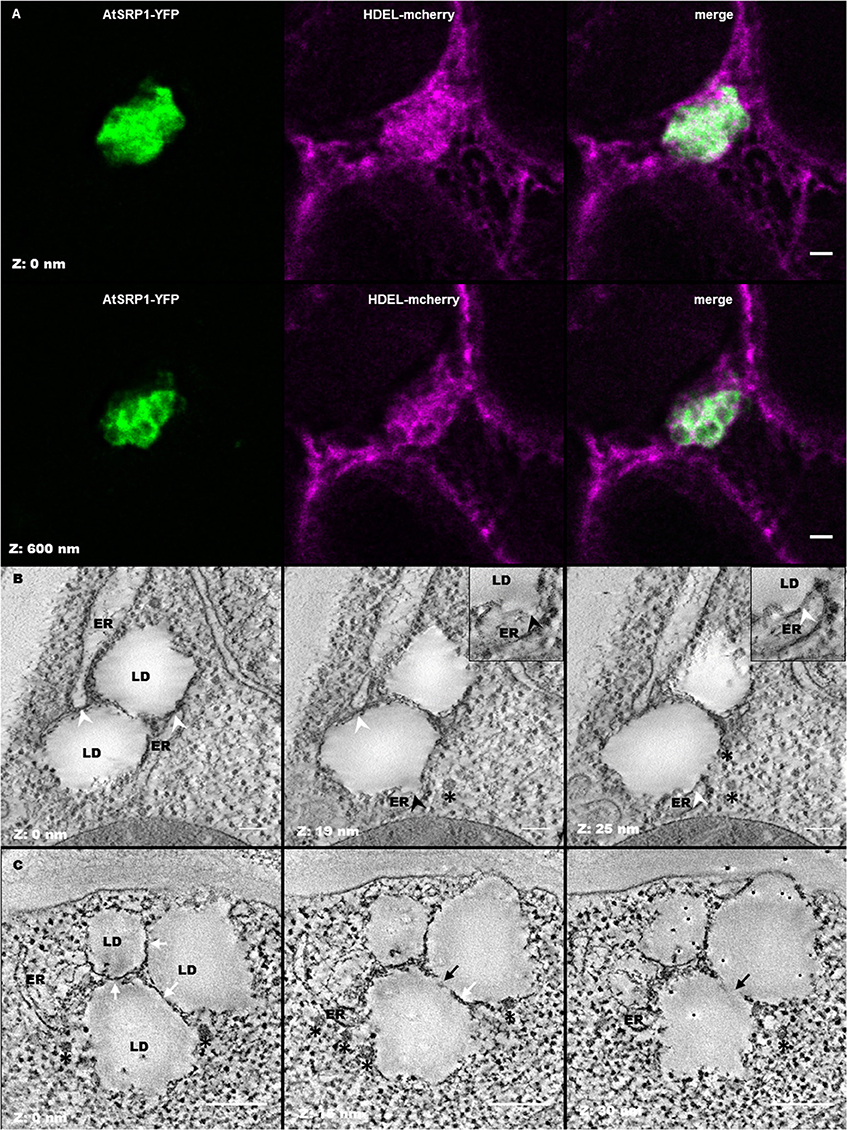
Figure 6. Leaf lipid droplets (LDs) are in close proximity with the endoplasmic reticulum (ER). (A) Fluorescent co-labeling of leaf LDs and the ER. N. benthamiana leaves were transiently co-transformed with AtSRP1-YFP and HDEL-mCherry constructs labeling LDs and ER membranes respectively. Deconvolution after confocal imaging of epidermal cells demonstrates multiple contact sites between LDs and the ER network, and AtSRP1-YFP localization at the LD periphery. Merge indicates overlap of YFP (green) and mCherry (magenta) fluorescences. Bar: 2 μm. (B,C) Electron tomography of leaf LDs in AtSRP1 overexpressing Arabidopsis plantlets with single (B) and dual (C) axis acquisition. White arrowheads indicate LD/ER contacts. Black arrowhead indicates the ER mono-leaflet at the LD budding point. White arrows indicate apposition of two LDs, black arrows indicate direct continuity between two LDs. Black asterisks are positioned below vesicle-like structures. Black points with white halo in last panel are gold fiducials used for alignment steps during tomogram. Bar: 200 nm.
As emphasized by Lersten et al. (2006) as well as Xu and Shanklin (2016), the biology and structure of LDs in leaf cells still remains to be examined. While one could argue that cotyledon and leaf LDs are similar organelles and that LDs have already been described in chemically fixed cotyledons (Frey-Wyssling et al., 1963; Wanner and Theimer, 1978; Wanner et al., 1981), the results of our proteomic analysis demonstrated that leaf LDs have a protein composition totally different from that of seed/cotyledon LDs, suggesting a divergence in function and possibly in structure. The ability to cryo-fix leaves was crucial to fix instantaneously the sample in a near to native state and optimize ultrastructure conservation. Aging leaves, accumulating LDs, are difficult to cryo-fix but young leaves only contain minor quantity of LDs. We thus took advantage of the induction of LD formation in young leaves of our AtSRP1 overexpressing plants to deeply decipher the leaf LD ultrastructure using an electron tomography of LDs in leaves of 6 day old plantlets (Figure 6B and Movie S2). Our analysis showed several ER membranes closely opposed to each LD, in a conformation excluding ribosomes from these sites. Given that the exclusion of ribosomes from the ER membrane is, with the proximity of membranes, “one defining feature for membrane contact sites” (Pérez-Sancho et al., 2016; Phillips and Voeltz, 2016), this suggests that in leaf, LDs and ER share domains reminiscent of membrane contact sites, as illustrated for example in Caenorhabditis elegans (Xu et al., 2012). In addition, a direct continuity between the ER membrane and the LD was observed in some sections (Black arrows in Figure 6B), with the inner leaflet of the ER membrane still separating the LD from the ER lumen. This site, representing a specialized ER domain for LD formation, is highly restricted, with a thickness of only a few tens of nm. We propose that this connection, together with multiple contact sites allow the transit of metabolites between the two compartments, in particular the LD accumulation of TAGs synthesized at the ER membrane, as previously demonstrated in yeast and mammalian cells (Jacquier et al., 2011; Kassan et al., 2013; Choudhary et al., 2015). Given the numerous contact sites observed between LDs and the ER in AtSRP1 overexpressing leaves, one can wonder why no ER protein originating from ER remnants were identified in the LD fraction. It probably reflects that ER-LD connections in wild-type plants are highly labile and dynamic, which is indispensable to allow LD intracellular motion (Welte, 2009). LDs of a same cluster seemed also appressed to each other rather than simply side by side. By electron tomography of a restricted volume, interconnections between some LDs could be observed (Figure 6C), suggesting either that newly formed LDs could bud from pre-existing one or that when a LD reaches its optimal size, the formation of a new one is induced at the exact same site of the ER. The first LD does not detach from the following one, leading to the formation of these clusters of interconnected LDs. Yet, the existence of such an interconnection between two LDs in wild-type leaf remains to be proven. Finally, multiple COP-like vesicles were observed by electron tomography in the vicinity of the LD clusters (black asterisks in Figures 6B,C). The identity of these vesicles remains elusive, however, they could participate in the LD biogenesis, similarly to the model proposed by Kalantari et al. (2010) for animal cells where “lipid droplets form in close association with ER membrane seeded from a COPI vesicle.”
AtSRP1 Regulates the Biogenesis and the Size of LDs in Induced Senescent Leaves
We next evaluated the role of AtSRP1 on the biogenesis and ultrastructure of LDs under conditions inducing their formation. In these experiments, plantlets were submitted to nitrogen starvation in order to induce precocious senescence (Balazadeh et al., 2014). As expected, nitrogen starvation induced a scattered LD formation in wild type plants (Figure 5B) with an average of 0.009 ± 0.004 μm−2 LD per cell area (corresponding roughly to 3.3 LDs per cell section) while LDs were scarcely observable under control conditions. The average area of a single LD was 0.51 ± 0.20 μm2, with no LD larger than 1.3 μm2 (Figure 5C, Figure S9). Moreover, 15% of wild type cells contained clusters of LD, with clusters composed of a maximum of 4 LDs. Under nitrogen starvation conditions, the number of LDs per cell area in AtSRP1 overexpressing line was significantly increased compared to the wild type (Figure 5D), with an average of 0.017 ± 0.003 μm−2 LDs per cell area. In addition, LDs were distributed in 50% of the cases in clusters of 2–35 LDs. The LD size in AtSRP1 overexpressing line was heterogeneous, ranging from 0.026 to more than 6 μm2; yet this was not statistically significantly different from the wild type (Figure 5C, Figure S9). The effect of nitrogen starvation was also tested on the two atsrp1 knock-down mutants where it also induced LD formation but significantly (p < 0.01) less than in the wild type. Furthermore, the LD size was highly heterogeneous and significantly bigger in one knock-down line compared to the wild-type (Figures 5B,D and Figure S9), with some very large LDs (up to 25 μm2). It seems that when knock-down atsrp1 lines are submitted to conditions that should induce LD formation, no or few new LDs are generated, and molecules usually sequestered in newly formed LDs accumulate in pre-existing LDs thus leading to abnormally large LDs. Altogether, these results indicate that SRP1 is a key element in (1) preventing the coalescence of LDs, (2) regulating their size, and (3) inducing their neo-formation in senescent leaves.
AtSRP1 Induces Accumulation of TAG Enriched in C18:3 in Aging Leaves
The induced neo-formation of LDs in AtSRP1 overexpressing leaves, suggests that AtSRP1 is able to mobilize components required for the formation of LDs, including structural proteins and neutral lipids. Gidda et al. (2016) described an increase of neutral lipid content in AtSRP1 overexpressing plantlets grown for 15 days on Murashige and Skoog plates, with an enrichment of C18:2 and C18:3, and no perturbation of the lipid composition of knock-down lines. In order to better evaluate the role of AtSRP1 under conditions that naturally induce AtSRP1 expression and neutral lipid accumulation in LDs, we analyzed the neutral lipid composition of 6-week old senescent AtSRP1 overexpressing or knock-down leaves (Figure 4B). The lipid composition of knock-down lines was not perturbed, but AtSRP1 overexpressing lines accumulated more neutral lipids in leaves than the wild type (Figure 4B). Even though lipid esters are the most abundant neutral lipids in LDs of aging leaves (Figure 1B), the overexpression of AtSRP1 specifically led to an increase of the TAG and not of the lipid ester content. In the three independent overexpressing lines, TAGs were more than three times more abundant than in the wild-type. In addition, overexpression of AtSRP1 modified the TAG fatty acid composition with a significant enrichment in C18:3 acyl moieties, mainly at the expense of C16:0. By contrast, the fatty acid composition of lipid esters was only slightly modified and the fatty acid composition of galactolipids and phospholipids were similar in the wild type and in overexpressing lines (Figure S8). These data show that the overexpression of AtSRP1 does not impact fatty acid metabolic pathways. Further, our results suggest that the significant enrichment in C18:3 acyl moieties of TAG does not originate from an imbalance in the C18:3 homeostasis but may rather originate from recycling of thylakoid galactolipids (that are rich in C18:3 and relatively poor in C16:0). The capacity of the cell to form new LDs seems to limit the recycling of acyl groups into TAGs. When AtSRP1 induces LD biogenesis, TAG synthesis is no more limited and TAG leaf content increases. An increase in TAG content was similarly described in LEC2 and/or SEIPIN overexpressing plants (Cai et al., 2015). LEC2 is a seed specific transcription factor that promotes the accumulation of TAG by stimulating the expression of seed-specific genes. AtSEIPINs are human lipodystrophy orthologs, which localize in discrete ER subdomains and promote LD biogenesis. In LEC2 and AtSEIPIN overexpressing plants, the TAG fatty acid composition is modified showing an increase of C16:0 and a decrease of C18:3 contents. C16:0 acyls are enriched in the ER, suggesting that both AtSEIPIN and LEC2 promote the accumulation of ER-derived acyl groups into TAGs. In contrast, AtSRP1 seems to induce a different mechanism which would rather promote the recycling of plastid lipids for TAG synthesis. However, additional evidences are needed to support this hypothesis. It is worth noting that, in aging leaves, LDs were predominantly observed in close proximity to plastids (more than 85% of the observed LDs were next to the plastid envelope). This further suggests a possible transfer of metabolites from the plastids to the LDs during senescence, as already suggested for plastids and ER (reviewed in Mehrshahi et al., 2014), or for plastid and LDs in C. reinhardtii (Goodson et al., 2011).
Conclusion
Altogether, our fine microscopic analyses demonstrated that AtSRP1 plays a dual structural role by inducing LD formation and controlling LD size. Indeed, when LD formation is induced in atsrp1 knock-down lines, striking large LDs are generated. Conversely, in AtSRP1 overexpressing lines LD biogenesis is exacerbated, leading to the formation of numerous LDs that stay aggregated in clusters but do not coalesce in one large LD. These LDs stay in close proximity with the ER, being wrapped within its network, and, as illustrated by tomography, can share direct connections with the ER lumen as well as with other LDs. A tight regulation of LD size and distribution is most probably crucial for the optimal functioning of the LDs. Our proteomics data show that leaf LDs are abundantly associated with proteins of the cytoskeleton, highlighting the importance of intracellular LD motion for their functions (Welte, 2009). Large LDs or aggregation of LDs in clusters most probably impair fast cell dynamic movements as well as LD exchanges with other organelles. In addition, we show that AtSRP1 overexpression enhances TAG synthesis in a different way to that described in cotyledons with SEIPIN proteins and suggest a possible involvement of plastid lipid metabolism in LD biogenesis of senescent leaves. Because SEIPINs co-localized with LDs and the ER, and influenced the structure of the ER, Cai et al. (2015) hypothesized that “SEIPINs participate in the organization of ER subdomains that are devoted to (…) LD formation.” It is possible that once SEIPINs have reorganized the ER in specialized subdomains dedicated to LD formation, AtSRP1 is recruited to induce the formation of new LDs at these subdomains. Furthermore, AtSRP1 could maintain a correct radius curvature of the monolayer surrounding the LD, therefore controlling its size. Our tomographic study demonstrates physical connections between LDs of a same cluster and between LDs and the ER in AtSRP1 overexpressing leaves. These connections may also exist in wild type leaves. However, given the involvement of LDs in intracellular movement and exchange, it is improbable that these connections permanently exist in the wild type. The fission of organelles, for example of coat protein (COP)II vesicles from the ER, or of the Golgi vesicles, necessitates the intervention of specialized proteins (Brown et al., 2008; Fujimoto and Tsutsumi, 2014). In the case of LDs, it is unclear if they systematically dissociate from the ER membrane and if this detachment is spontaneous or supported by specific proteins. The separation of LDs from the ER (and from other LDs) seems perturbed by the overexpression of AtSRP1, therefore indicating that this dissociation is not spontaneous. The unusual accumulation of AtSRP1 at the LD surface may impair the membrane layer fusion process necessary for LD detachment, or the proteins responsible for the LD closing and detachment process may not be enough represented in the overexpressing lines to separate all the neo-formed LDs from the ER.
Results from proteomic study presented here show that AtCLO3 and AtSRP1 are the only structural proteins present in the proteome of leaf LDs. What are the respective roles of AtSRP1 and AtCLO3, the two most abundant proteins in leaf LDs, in the maintenance of LD ultrastructure? How is SRP1 recruited to the ER? Do SEIPIN proteins interact with AtSRP1 and when? Are there any additional partners needed for the dissociation of the LDs from the ER? What is the exact chronology of the diverse events responsible for the LD biogenesis in leaves? In this purpose, the availability of AtSRP1, AtCLO3 and SEIPINs overexpressing lines, as well as the ability to cryofix plant tissues and to perform a correlative light electron microscopy approach, represent valuable tools to better understand events of the LD biogenesis in non-seed tissue.
Accession Numbers
Sequence data from this article can be found in the Arabidopsis Genome Initiative databases under the following accession numbers: AtSRP1: At1g67360; AtCLO3: At2g33380; AtULP: At5g16550; actin: At1g49240; GAPDH: At1g13440; eIF4A-1: At3g13920.
Author Contributions
LB, FI, DC, NE, and CB performed most of the experiments; LB, LF, SC, and CB supervised the experiments; KT, SP, SC, and NE provided technical assistance to LB, FI, and CB; LB, FI, JB, and CB designed the experiments and analyzed the data; CB conceived and supervised the project. FI and CB wrote the article with contributions of all the authors.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Authors would like to acknowledge L. Combes-Soia and A-M. Lomenech for technical help in lipidomics and proteomics respectively, Dr. C. Garcion for VBA programming skills, Dr. P. Legros for lending his confocal microscope, Dr. M. Bonneu for his expertise in proteomic data analysis, Dr. M Blatt and Dr. T. Nakagawa for sharing publish material, and members of the laboratory of membrane biogenesis for their supportive encouragements, especially Drs. A. Bernard, J. Joubes, S. Mongrand and E. M. Bayer for critical reading of the manuscript, and A. Chevin for fund management. The lipid analyses were performed at the Metabolome Facility of Bordeaux-MetaboHUB (ANR-11-INBS-0010). The imagery experiments were achieved on the Bordeaux Imaging center, a member of the France Bioimaging Infrastructure. This work was supported by Idex Bordeaux and CNRS (PEPS 2013-0260). Fundings for the TEM equipment were obtained thanks to the Region Aquitaine via the Convention N°2011 13 04 007 PFM.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00894/full#supplementary-material
References
Ahrné, E., Molzahn, L., Glatter, T., and Schmidt, A. (2013). Critical assessment of proteome-wide label-free absolute abundance estimation strategies. Proteomics 13, 2567–2578. doi: 10.1002/pmic.201300135
Ajjawi, I., Coku, A., Froehlich, J. E., Yang, Y., Osteryoung, K. W., Benning, C., et al. (2011). A J-Like Protein Influences Fatty Acid Composition of Chloroplast Lipids in Arabidopsis. PLoS ONE 6:e25368. doi: 10.1371/journal.pone.0025368
Attieh, J., Djiana, R., Koonjul, P., Etienne, C., Sparace, S. A., and Saini, H. S. (2002). Cloning and functional expression of two plant thiol methyltransferases: a new class of enzymes involved in the biosynthesis of sulfur volatiles. Plant Mol. Biol. 50, 511–521. doi: 10.1023/A:1019865829534
Attieh, J., Kleppinger-Sparace, K. F., Nunes, C., Sparace, S. A., and Saini, H. S. (2000). Evidence implicating a novel thiol methyltransferase in the detoxification of glucosinolate hydrolysis products in Brassica oleracea L. Plant Cell Environ. 23, 165–174. doi: 10.1046/j.1365-3040.2000.00541.x
Aubert, Y., Vile, D., Pervent, M., Aldon, D., Ranty, B., Simonneau, T., et al. (2010). RD20, a Stress-Inducible Caleosin, Participates in Stomatal Control, Transpiration and Drought Tolerance in Arabidopsis thaliana. Plant Cell Physiol. 51, 1975–1987. doi: 10.1093/pcp/pcq155
Austin, J. R., Frost, E., Vidi, P. A., Kessler, F., and Staehelin, L. A. (2006). Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 18, 1693–1703. doi: 10.1105/tpc.105.039859
Awai, K., Xu, C., Tamot, B., and Benning, C. (2006). A phosphatidic acid-binding protein of the chloroplast inner envelope membrane involved in lipid trafficking. Proc. Natl. Acad. Sci. U.S.A. 103, 10817–10822. doi: 10.1073/pnas.0602754103
Balazadeh, S., Schildhauer, J., Araújo, W. L., Munné-Bosch, S., Fernie, A. R., Proost, S., et al. (2014). Reversal of senescence by N resupply to N-starved Arabidopsis thaliana: transcriptomic and metabolomic consequences. J. Exp. Bot. 65, 3975–3992. doi: 10.1093/jxb/eru119
Banfi, C., Brioschi, M., Barcella, S., Wait, R., Begum, S., Galli, S., et al. (2009). Proteomic analysis of human low-density lipoprotein reveals the presence of prenylcysteine lyase, a hydrogen peroxide-generating enzyme. Proteomics 9, 1344–1352. doi: 10.1002/pmic.200800566
Barleben, L., Panjikar, S., Ruppert, M., Koepke, J., and Stöckigt, J. (2007). Molecular architecture of strictosidine glucosidase: the gateway to the biosynthesis of the monoterpenoid indole alkaloid family. Plant Cell 19, 2886–2897. doi: 10.1105/tpc.106.045682
Bartz, R., Li, W.-H., Venables, B., Zehmer, J. K., Roth, M. R., Welti, R., et al. (2007). Lipidomics reveals that adiposomes store ether lipids and mediate phospholipid traffic,. J. Lipid Res. 48, 837–847. doi: 10.1194/jlr.M600413-JLR200
Berthelot, K., Lecomte, S., Estevez, Y., Coulary-Salin, B., Bentaleb, A., Cullin, C., et al. (2012). Rubber Elongation Factor (REF), a major allergen component in Hevea brasiliensis latex has amyloid properties. PLoS ONE 7:e48065. doi: 10.1371/journal.pone.0048065
Berthelot, K., Lecomte, S., Estevez, Y., Coulary-Salin, B., and Peruch, F. (2014). Homologous Hevea brasiliensis REF (Hevb1) and SRPP (Hevb3) present different auto-assembling. Biochim. Biophys. Acta 1844, 473–485. doi: 10.1016/j.bbapap.2013.10.017
Besagni, C., Piller, L. E., and Bréhélin, C. (2011). Preparation of plastoglobules from Arabidopsis plastids for proteomic analysis and other studies. Methods Mol. Biol. 775, 223–239. doi: 10.1007/978-1-61779-237-3_12
Blée, E., Boachon, B., Burcklen, M., Guédard, M. L., Hanano, A., Heintz, D., et al. (2014). The reductase activity of the Arabidopsis caleosin RESPONSIVE TO DESSICATION20 mediates gibberellin-dependent flowering time, abscisic acid sensitivity, and tolerance to oxidative stress. Plant Physiol. 166, 109–124. doi: 10.1104/pp.114.245316
Boström, P., Andersson, L., Rutberg, M., Perman, J., Lidberg, U., Johansson, B. R., et al. (2007). SNARE proteins mediate fusion between cytosolic lipid droplets and are implicated in insulin sensitivity. Nat. Cell Biol. 9, 1286–1293. doi: 10.1038/ncb1648
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Brown, W. J., Plutner, H., Drecktrah, D., Judson, B. L., and Balch, W. E. (2008). The lysophospholipid acyltransferase antagonist CI-976 inhibits a late step in COPII vesicle budding. Traffic Cph Den 9, 786–797. doi: 10.1111/j.1600-0854.2008.00711.x
Cai, Y., Goodman, J. M., Pyc, M., Mullen, R. T., Dyer, J. M., and Chapman, K. D. (2015). Arabidopsis SEIPIN proteins modulate triacylglycerol accumulation and influence lipid droplet proliferation. Plant Cell 27, 2616–2636. doi: 10.1105/tpc.15.00588
Campbell, M. M., and Sederoff, R. R. (1996). Variation in lignin content and composition (mechanisms of control and implications for the genetic improvement of plants). Plant Physiol. 110, 3–13. doi: 10.1104/pp.110.1.3
Cermelli, S., Guo, Y., Gross, S. P., and Welte, M. A. (2006). The lipid-droplet proteome reveals that droplets are a protein-storage depot. Curr. Biol. 16, 1783–1795. doi: 10.1016/j.cub.2006.07.062
Chapman, K. D., Dyer, J. M., and Mullen, R. T. (2012). Biogenesis and functions of lipid droplets in plants: thematic review series: lipid droplet synthesis and metabolism: from yeast to man. J. Lipid Res. 53, 215–226. doi: 10.1194/jlr.R021436
Chapman, K. D., Dyer, J. M., and Mullen, R. T. (2013). Commentary: why don't plant leaves get fat? Plant Sci. 207, 128–134. doi: 10.1016/j.plantsci.2013.03.003
Chen, M. C. M., Chyan, C.-L., Lee, T. T. T., Huang, S.-H., and Tzen, J. T. C. (2004). Constitution of stable artificial oil bodies with triacylglycerol, phospholipid, and caleosin. J. Agric. Food Chem. 52, 3982–3987. doi: 10.1021/jf035533g
Choudhary, V., Ojha, N., Golden, A., and Prinz, W. A. (2015). A conserved family of proteins facilitates nascent lipid droplet budding from the ER. J. Cell Biol. 211, 261–271. doi: 10.1083/jcb.201505067
Christ, B., Süssenbacher, I., Moser, S., Bichsel, N., Egert, A., Müller, T., et al. (2013). Cytochrome P450 CYP89A9 is involved in the formation of major chlorophyll catabolites during leaf senescence in Arabidopsis. Plant Cell 25, 1868–1880. doi: 10.1105/tpc.113.112151
Clough, S. J., and Bent, A. F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Consortium, T. U. (2015). UniProt: a hub for protein information. Nucleic Acids Res. 43, D204–D212. doi: 10.1093/nar/gku989
Courdavault, V., Burlat, V., St-Pierre, B., and Giglioli-Guivarc'h, N. (2009). Proteins prenylated by type I protein geranylgeranyltransferase act positively on the jasmonate signalling pathway triggering the biosynthesis of monoterpene indole alkaloids in Catharanthus roseus. Plant Cell Rep. 28, 83–93. doi: 10.1007/s00299-008-0610-1
Crowell, D. N., and Huizinga, D. H. (2009). Protein isoprenylation: the fat of the matter. Trends Plant Sci. 14, 163–170. doi: 10.1016/j.tplants.2008.12.001
Crowell, D. N., Huizinga, D. H., Deem, A. K., Trobaugh, C., Denton, R., and Sen, S. E. (2007). Arabidopsis thaliana plants possess a specific farnesylcysteine lyase that is involved in detoxification and recycling of farnesylcysteine. Plant J. 50, 839–847. doi: 10.1111/j.1365-313X.2007.03091.x
De Domenico, S., Bonsegna, S., Lenucci, M. S., Poltronieri, P., Di Sansebastiano, G. P., and Santino, A. (2011). Localization of seed oil body proteins in tobacco protoplasts reveals specific mechanisms of protein targeting to leaf lipid droplets. J. Integr. Plant Biol. 53, 858–868. doi: 10.1111/j.1744-7909.2011.01077.x
De Luca, V., and Cutler, A. J. (1987). Subcellular localization of enzymes involved in indole alkaloid biosynthesis in Catharanthus roseus. Plant Physiol. 85, 1099–1102. doi: 10.1104/pp.85.4.1099
Dutta, A., and Kumar Sinha, D. (2015). Turnover of the actomyosin complex in zebrafish embryos directs geometric remodelling and the recruitment of lipid droplets. Sci. Rep. 5:13915. doi: 10.1038/srep13915
Eastmond, P. J. (2006). SUGAR-DEPENDENT1 encodes a patatin domain triacylglycerol lipase that initiates storage oil breakdown in germinating Arabidopsis seeds. Plant Cell 18, 665–675. doi: 10.1105/tpc.105.040543
Fahn, W., Laußermair, E., Deus-Neumann, B., and Stöckigt, J. (1985). Late enzymes of vindoline biosynthesis : S-Adenosyl-L-methionine: 11-O-demethyl-17-O-deacetylvindoline 11-O-methyltransferase and unspecific acetylesterase. Plant Cell Rep. 4, 337–340. doi: 10.1007/BF00269893
Farese, R. V. J., and Walther, T. C. (2009). Lipid droplets finally get a little R-E-S-P-E-C-T. Cell 139, 855–860. doi: 10.1016/j.cell.2009.11.005
Folch, J., Lees, M., and Stanley, G. H. S. (1957). A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 226, 497–509.
Frey-Wyssling, A., Grieshaber, E., and Mühlethaler, K. (1963). Origin of spherosomes in plant cells. J. Ultrastruct. Res. 8, 506–516. doi: 10.1016/S0022-5320(63)80052-0
Fujimoto, M., and Tsutsumi, N. (2014). Dynamin-related proteins in plant post-Golgi traffic. Front. Plant Sci. 5:408. doi: 10.3389/fpls.2014.00408
Galichet, A., and Gruissem, W. (2003). Protein farnesylation in plants–conserved mechanisms but different targets. Curr. Opin. Plant Biol. 6, 530–535. doi: 10.1016/j.pbi.2003.09.005
Gaude, N., Bréhélin, C., Tischendorf, G., Kessler, F., and Dormann, P. (2007). Nitrogen deficiency in Arabidopsis affects galactolipid composition and gene expression and results in accumulation of fatty acid phytyl esters. Plant J. 49, 729–739. doi: 10.1111/j.1365-313X.2006.02992.x
Geisler, M., Frangne, N., Gomès, E., Martinoia, E., and Palmgren, M. G. (2000). The ACA4 Gene of Arabidopsis encodes a vacuolar membrane calcium pump that improves salt tolerance in yeast. Plant Physiol. 124, 1814–1827. doi: 10.1104/pp.124.4.1814
Gidda, S. K., Park, S., Pyc, M., Yurchenko, O., Cai, Y., Wu, P., et al. (2016). Lipid droplet-associated proteins (LDAPs) are required for the dynamic regulation of neutral lipid compartmentation in plant cells. Plant Physiol. 170, 2052–2071. doi: 10.1104/pp.15.01977
Goodson, C., Roth, R., Wang, Z. T., and Goodenough, U. (2011). Structural correlates of cytoplasmic and chloroplast lipid body synthesis in Chlamydomonas reinhardtii and stimulation of lipid body production with acetate boost. Eukaryotic Cell 10, 1592–1606. doi: 10.1128/EC.05242-11
Goold, H., Beisson, F., Peltier, G., and Li-Beisson, Y. (2015). Microalgal lipid droplets: composition, diversity, biogenesis and functions. Plant Cell Rep. 34, 545–555. doi: 10.1007/s00299-014-1711-7
Grefen, C., Donald, N., Hashimoto, K., Kudla, J., Schumacher, K., and Blatt, M. R. (2010). A ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies. Plant J. 64, 355–365. doi: 10.1111/j.1365-313X.2010.04322.x
Grison, M. S., Brocard, L., Fouillen, L., Nicolas, W., Wewer, V., Dörmann, P., et al. (2015). Specific membrane lipid composition is important for plasmodesmata function in Arabidopsis. Plant Cell 27, 1228–1250. doi: 10.1105/tpc.114.135731
Guirimand, G., Courdavault, V., Lanoue, A., Mahroug, S., Guihur, A., Blanc, N., et al. (2010). Strictosidine activation in Apocynaceae: towards a “nuclear time bomb”? BMC Plant Biol. 10:182. doi: 10.1186/1471-2229-10-182
Hansen, B. G., Kerwin, R. E., Ober, J. A., Lambrix, V. M., Mitchell-Olds, T., Gershenzon, J., et al. (2008). A novel 2-oxoacid-dependent dioxygenase involved in the formation of the goiterogenic 2-hydroxybut-3-enyl glucosinolate and generalist insect resistance in Arabidopsis. Plant Physiol. 148, 2096–2108. doi: 10.1104/pp.108.129981
Hashemi, H. F., and Goodman, J. M. (2015). The life cycle of lipid droplets. Curr. Opin. Cell Biol. 33, 119–124. doi: 10.1016/j.ceb.2015.02.002
Hillebrand, A., Post, J. J., Wurbs, D., Wahler, D., Lenders, M., Krzyzanek, V., et al. (2012). Down-regulation of small rubber particle protein expression affects integrity of rubber particles and rubber content in Taraxacum brevicorniculatum. PLoS ONE 7:e41874. doi: 10.1371/journal.pone.0041874
Höfte, H., and Chrispeels, M. J. (1992). Protein sorting to the vacuolar membrane. Plant Cell 4, 995–1004. doi: 10.1105/tpc.4.8.995
Hooper, C. M., Tanz, S. K., Castleden, I. R., Vacher, M. A., Small, I. D., and Millar, A. H. (2014). SUBAcon: a consensus algorithm for unifying the subcellular localization data of the Arabidopsis proteome. Bioinforma Oxf. Engl. 30, 3356–3364. doi: 10.1093/bioinformatics/btu550
Horn, P. J., James, C. N., Gidda, S. K., Kilaru, A., Dyer, J. M., Mullen, R. T., et al. (2013). Identification of a new class of lipid droplet-associated proteins in plants. Plant Physiol. 162, 1926–1936. doi: 10.1104/pp.113.222455
Huang, A. H. C. (1992). Oil bodies and oleosins in seeds. Annu. Rev. Plant Physiol. Plant Mol. Biol. 43, 177–200. doi: 10.1146/annurev.pp.43.060192.001141
Iida, K., Fukami-Kobayashi, K., Toyoda, A., Sakaki, Y., Kobayashi, M., Seki, M., et al. (2009). Analysis of multiple occurrences of alternative splicing events in Arabidopsis thaliana using novel sequenced full-length cDNAs. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 16, 155–164. doi: 10.1093/dnares/dsp009
Jacquier, N., Choudhary, V., Mari, M., Toulmay, A., Reggiori, F., and Schneiter, R. (2011). Lipid droplets are functionally connected to the endoplasmic reticulum in Saccharomyces cerevisiae. J. Cell Sci. 124, 2424–2437. doi: 10.1242/jcs.076836
Jiang, P.-L., Chen, J. C. F., Chiu, S.-T., and Tzen, J. T. C. (2009). Stable oil bodies sheltered by a unique caleosin in cycad megagametophytes. Plant Physiol. Biochem. PPB 47, 1009–1016. doi: 10.1016/j.plaphy.2009.07.004
Jolivet, P., Acevedo, F., Boulard, C., d'Andréa, S., Faure, J.-D., Kohli, A., et al. (2013). Crop seed oil bodies: from challenges in protein identification to an emerging picture of the oil body proteome. Proteomics 13, 1836–1849. doi: 10.1002/pmic.201200431
Juguelin, H., Heape, A., Boiron, F., and Cassagne, C. (1986). A quantitative developmental study of neutral lipids during myelinogenesis in the peripheral nervous system of normal and trembler mice. Brain Res. 390, 249–252. doi: 10.1016/S0006-8993(86)80233-5
Kalantari, F., Bergeron, J. J. M., and Nilsson, T. (2010). Biogenesis of lipid droplets – how cells get fatter. Mol. Membr. Biol. 27, 462–468. doi: 10.3109/09687688.2010.538936
Kanehisa, M., Sato, Y., Kawashima, M., Furumichi, M., and Tanabe, M. (2016). KEGG as a reference resource for gene and protein annotation. Nucleic Acids Res. 44, D457–D462. doi: 10.1093/nar/gkv1070
Kassan, A., Herms, A., Fernández-Vidal, A., Bosch, M., Schieber, N. L., Reddy, B. J. N., et al. (2013). Acyl-CoA synthetase 3 promotes lipid droplet biogenesis in ER microdomains. J. Cell Biol. 203, 985–1001. doi: 10.1083/jcb.201305142
Kawai, Y., Ono, E., and Mizutani, M. (2014). Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. Cell Mol. Biol. 78, 328–343. doi: 10.1111/tpj.12479
Keith, B., Dong, X. N., Ausubel, F. M., and Fink, G. R. (1991). Differential induction of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase genes in Arabidopsis thaliana by wounding and pathogenic attack. Proc. Natl. Acad. Sci. U.S.A. 88, 8821–8825. doi: 10.1073/pnas.88.19.8821
Kim, E. Y., Park, K. Y., Seo, Y. S., and Kim, W. T. (2016). Arabidopsis small rubber particle protein homolog SRPs play dual roles as positive factors for tissue growth and development and in drought stress responses. Plant Physiol. 170, 2494–2510. doi: 10.1104/pp.16.00165
Kim, H. J., Ok, S. H., Bahn, S. C., Jang, J., Oh, S. A., Park, S. K., et al. (2011). Endoplasmic Reticulum– and golgi-localized phospholipase A2 plays critical roles in arabidopsis pollen development and germination. Plant Cell 23, 94–110. doi: 10.1105/tpc.110.074799
Kim, H. U., Hsieh, K., Ratnayake, C., and Huang, A. H. C. (2002). A novel group of oleosins is present inside the pollen of Arabidopsis. J. Biol. Chem. 277, 22677–22684. doi: 10.1074/jbc.M109298200
Kim, Y. Y., Jung, K. W., Yoo, K. S., Jeung, J. U., and Shin, J. S. (2011). A stress-responsive caleosin-like protein, AtCLO4, acts as a negative regulator of ABA responses in Arabidopsis. Plant Cell Physiol. 52, 874–884. doi: 10.1093/pcp/pcr039
Kissen, R., and Bones, A. M. (2009). Nitrile-specifier proteins involved in glucosinolate hydrolysis in Arabidopsis thaliana. J. Biol. Chem. 284, 12057–12070. doi: 10.1074/jbc.M807500200
Kong, X. Y., Kissen, R., and Bones, A. M. (2012). Characterization of recombinant nitrile-specifier proteins (NSPs) of Arabidopsis thaliana: dependency on Fe(II) ions and the effect of glucosinolate substrate and reaction conditions. Phytochemistry 84, 7–17. doi: 10.1016/j.phytochem.2012.08.004
Koroleva, O. A., Tomlinson, M. L., Leader, D., Shaw, P., and Doonan, J. H. (2005). High-throughput protein localization in Arabidopsis using Agrobacterium-mediated transient expression of GFP-ORF fusions. Plant J. 41, 162–174. doi: 10.1111/j.1365-313X.2004.02281.x
Krahmer, N., Farese, R. V., and Walther, T. C. (2013). Balancing the fat: lipid droplets and human disease: lipid droplets in disease. EMBO Mol. Med. 5, 973–983. doi: 10.1002/emmm.201100671
Le Gall, H., Philippe, F., Domon, J.-M., Gillet, F., Pelloux, J., and Rayon, C. (2015). Cell wall metabolism in response to abiotic stress. Plants 4, 112–166. doi: 10.3390/plants4010112
Lersten, N. R., Czlapinski, A. R., Curtis, J. D., Freckmann, R., and Horner, H. T. (2006). Oil bodies in leaf mesophyll cells of angiosperms: overview and a selected survey. Am. J. Bot. 93, 1731–1739. doi: 10.3732/ajb.93.12.1731
Li-Beisson, Y., Shorrosh, B., Beisson, F., Andersson, M. X., Arondel, V., Bates, P. D., et al. (2013). Acyl-lipid metabolism. Arab. Book 11:e0161. doi: 10.1199/tab.0161
Liu, H., Wang, C., Chen, F., and Shen, S. (2015). Proteomic analysis of oil bodies in mature Jatropha curcas seeds with different lipid content. J. Proteomics 113, 403–414. doi: 10.1016/j.jprot.2014.10.013
Liu, T., Chyan, C., Li, F., and Tzen, J. T. C. (2009). Stability of artificial oil bodies constituted with recombinant caleosins. J. Agric. Food Chem. 57, 2308–2313. doi: 10.1021/jf803566w
Lundquist, P. K., Poliakov, A., Bhuiyan, N. H., Zybailov, B., Sun, Q., and van Wijk, K. J. (2012). The functional network of the Arabidopsis plastoglobule proteome based on quantitative proteomics and genome-wide coexpression analysis. Plant Physiol. 158, 1172–1192. doi: 10.1104/pp.111.193144
Maeda, H., and Dudareva, N. (2012). The shikimate pathway and aromatic amino Acid biosynthesis in plants. Annu. Rev. Plant Biol. 63, 73–105. doi: 10.1146/annurev-arplant-042811-105439
Marais, C., Wattelet-Boyer, V., Bouyssou, G., Hocquellet, A., Dupuy, J.-W., Batailler, B., et al. (2015). The Qb-SNARE Memb11 interacts specifically with Arf1 in the Golgi apparatus of Arabidopsis thaliana. J. Exp. Bot. 66, 6665–6678. doi: 10.1093/jxb/erv373
Mehrshahi, P., Johnny, C., and DellaPenna, D. (2014). Redefining the metabolic continuity of chloroplasts and ER. Trends Plant Sci. 19, 501–507. doi: 10.1016/j.tplants.2014.02.013
Moldavski, O., Amen, T., Levin-Zaidman, S., Eisenstein, M., Rogachev, I., Brandis, A., et al. (2015). Lipid droplets are essential for efficient clearance of cytosolic inclusion bodies. Dev. Cell 33, 603–610. doi: 10.1016/j.devcel.2015.04.015
Muday, G. K., and Herrmann, K. M. (1992). Wounding induces one of two isoenzymes of 3-deoxy-d-arabino-heptulosonate 7-phosphate synthase in Solanum tuberosum L. Plant Physiol. 98, 496–500. doi: 10.1104/pp.98.2.496
Murphy, D. J. (2001). The biogenesis and functions of lipid bodies in animals, plants and microorganisms. Prog. Lipid Res. 40, 325–438. doi: 10.1016/S0163-7827(01)00013-3
Nacir, H., and Bréhélin, C. (2013). When proteomics reveals unsuspected roles: the plastoglobule example. Front. Plant Sci. 4:114. doi: 10.3389/fpls.2013.00114
Nakamura, S., Mano, S., Tanaka, Y., Ohnishi, M., Nakamori, C., Araki, M., et al. (2010). Gateway binary vectors with the bialaphos resistance gene, bar, as a selection marker for plant transformation. Biosci. Biotechnol. Biochem. 74, 1315–1319. doi: 10.1271/bbb.100184
Nelson, B. K., Cai, X., and Nebenführ, A. (2007). A multicolored set of in vivo organelle markers for co-localization studies in Arabidopsis and other plants. Plant J. 51, 1126–1136. doi: 10.1111/j.1365-313X.2007.03212.x
Nikolovski, N., Shliaha, P. V., Gatto, L., Dupree, P., and Lilley, K. S. (2014). Label-free protein quantification for plant golgi protein localization and abundance1[W]. Plant Physiol. 166, 1033–1043. doi: 10.1104/pp.114.245589
Ohsaki, Y., Cheng, J., Fujita, A., Tokumoto, T., and Fujimoto, T. (2006). Cytoplasmic lipid droplets are sites of convergence of proteasomal and autophagic degradation of apolipoprotein, B. Mol. Biol. Cell 17, 2674–2683. doi: 10.1091/mbc.E05-07-0659
Parker, M. L., and Murphy, G. J. P. (1981). Oleosomes in flag leaves of wheat; their distribution, composition and fate during senesence and rust-infection. Planta 152, 36–43. doi: 10.1007/BF00384982
Pérez-Sancho, J., Tilsner, J., Samuels, A. L., Botella, M. A., Bayer, E. M., and Rosado, A. (2016). Stitching organelles: organization and function of specialized membrane contact sites in plants. Trends Cell Biol. 26, 705–717. doi: 10.1016/j.tcb.2016.05.007
Perraki, A., Binaghi, M., Mecchia, M. A., Gronnier, J., German-Retana, S., Mongrand, S., et al. (2014). StRemorin1.3 hampers Potato virus X TGBp1 ability to increase plasmodesmata permeability, but does not interfere with its silencing suppressor activity. FEBS Lett. 588, 1699–1705. doi: 10.1016/j.febslet.2014.03.014
Phillips, M. J., and Voeltz, G. K. (2016). Structure and function of ER membrane contact sites with other organelles. Nat. Rev. Mol. Cell Biol. 17, 69–82. doi: 10.1038/nrm.2015.8
Rensink, W. A., Pilon, M., and Weisbeek, P. (1998). Domains of a transit sequence required for iimport in Arabidopsis chloroplasts. Plant Physiol 118, 691–699. doi: 10.1104/pp.118.2.691
Rischer, H., Orešič, M., Seppänen-Laakso, T., Katajamaa, M., Lammertyn, F., Ardiles-Diaz, W., et al. (2006). Gene-to-metabolite networks for terpenoid indole alkaloid biosynthesis in Catharanthus roseus cells. Proc. Natl. Acad. Sci. U.S.A. 103, 5614–5619. doi: 10.1073/pnas.0601027103
Roje, S. (2006). S-Adenosyl-l-methionine: beyond the universal methyl group donor. Phytochemistry 67, 1686–1698. doi: 10.1016/j.phytochem.2006.04.019
Schenk, N., Schelbert, S., Kanwischer, M., Goldschmidt, E. E., Dörmann, P., and Hörtensteiner, S. (2007). The chlorophyllases AtCLH1 and AtCLH2 are not essential for senescence-related chlorophyll breakdown in Arabidopsis thaliana. FEBS Lett. 581, 5517–5525. doi: 10.1016/j.febslet.2007.10.060
Schneider, C. A., Rasband, W. S., and Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675. doi: 10.1038/nmeth.2089
Schröder, G., Eichel, J., Breinig, S., and Schröder, J. (1997). Three differentially expressed S-adenosylmethionine synthetases from Catharanthus roseus: molecular and functional characterization. Plant Mol. Biol. 33, 211–222. doi: 10.1023/A:1005711720930
Shen, B., Li, C., and Tarczynski, M. C. (2002). High free-methionine and decreased lignin content result from a mutation in the Arabidopsis S-adenosyl-L-methionine synthetase 3 gene. Plant J. 29, 371–380. doi: 10.1046/j.1365-313X.2002.01221.x
Shen, J., Suen, P. K., Wang, X., Lin, Y., Lo, S. W., Rojo, E., et al. (2013). An in vivo expression system for the identification of cargo proteins of vacuolar sorting receptors in Arabidopsis culture cells. Plant J. 75, 1003–1017. doi: 10.1111/tpj.12257
Shimada, T. L., and Hara-Nishimura, I. (2015). Leaf oil bodies are subcellular factories producing antifungal oxylipins. Curr. Opin. Plant Biol. 25, 145–150. doi: 10.1016/j.pbi.2015.05.019
Shimada, T. L., Takano, Y., and Hara-Nishimura, I. (2015). Oil body-mediated defense against fungi: from tissues to ecology. Plant Signal. Behav. 10:e989036. doi: 10.4161/15592324.2014.989036
Shimada, T. L., Takano, Y., Shimada, T., Fujiwara, M., Fukao, Y., Mori, M., et al. (2014). Leaf oil body functions as a subcellular factory for the production of a phytoalexin in Arabidopsis. Plant Physiol. 164, 105–118. doi: 10.1104/pp.113.230185
Silva, J. C., Gorenstein, M. V., Li, G.-Z., Vissers, J. P. C., and Geromanos, S. J. (2006). Absolute quantification of proteins by LCMSE: a virtue of parallel MS acquisition. Mol. Cell Proteomics MCP 5, 144–156. doi: 10.1074/mcp.M500230-MCP200
St-Pierre, B., Vazquez-Flota, F. A., and Luca, V. D. (1999). Multicellular compartmentation of Catharanthus roseus alkaloid biosynthesis predicts intercellular translocation of a pathway intermediate. Plant Cell 11, 887–900. doi: 10.1105/tpc.11.5.887
Thazar-Poulot, N., Miquel, M., Fobis-Loisy, I., and Gaude, T. (2015). Peroxisome extensions deliver the Arabidopsis SDP1 lipase to oil bodies. Proc. Natl. Acad. Sci. U.S.A. 112, 4158–4163. doi: 10.1073/pnas.1403322112
Troncoso-Ponce, M. A., Cao, X., Yang, Z., and Ohlrogge, J. B. (2013). Lipid turnover during senescence. Plant Sci. 205–206, 13–19. doi: 10.1016/j.plantsci.2013.01.004
Tsai, C.-H., Zienkiewicz, K., Amstutz, C. L., Brink, B. G., Warakanont, J., Roston, R., et al. (2015). Dynamics of protein and polar lipid recruitment during lipid droplet assembly in Chlamydomonas reinhardtii. Plant J. 83, 650–660. doi: 10.1111/tpj.12917
Ueda, H., Yokota, E., Kutsuna, N., Shimada, T., Tamura, K., Shimmen, T., et al. (2010). Myosin-dependent endoplasmic reticulum motility and F-actin organization in plant cells. Proc. Natl. Acad. Sci. U.S.A. 107, 6894–6899. doi: 10.1073/pnas.0911482107
Vallon, O., Wollman, F. A., and Olive, J. (1986). Lateral distribution of the main protein complexes of the photosynthetic apparatus in Chlamydomonas reinhardtii and in spinach: an immunocytochemical study using intact thylakoid membranes and a Ps-II enriched membrane preparation. Photobiochem. Photobiophys. 12, 203–220.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., et al. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3:RESEARCH0034. doi: 10.1186/gb-2002-3-7-research0034
Vevea, J. D., Garcia, E. J., Chan, R. B., Zhou, B., Schultz, M., Di Paolo, G., et al. (2015). Role for lipid droplet biogenesis and microlipophagy in adaptation to lipid imbalance in yeast. Dev. Cell 35, 584–599. doi: 10.1016/j.devcel.2015.11.010
Vidi, P. A., Kanwischer, M., Baginsky, S., Austin, J. R., Csucs, G., Dormann, P., et al. (2006). Tocopherol cyclase (VTE1) localization and vitamin E accumulation in chloroplast plastoglobule lipoprotein particles. J. Biol. Chem. 281, 11225–11234. doi: 10.1074/jbc.M511939200
Vidi, P. A., Kessler, F., and Bréhélin, C. (2007). Plastoglobules: a new address for targeting recombinant proteins in the chloroplast. BMC Biotechnol. 7:4. doi: 10.1186/1472-6750-7-4
Vitiello, F., and Zanetta, J. P. (1978). Thin-layer chromatography of phospholipids. J. Chromatogr. 166, 637–640. doi: 10.1016/S0021-9673(00)95654-1
Vizcaíno, J. A., Csordas, A., del-Toro, N., Dianes, J. A., Griss, J., Lavidas, I., et al. (2016). 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 44, D447–D456. doi: 10.1093/nar/gkv1145
Wanner, G., Formanek, H., and Theimer, R. R. (1981). The ontogeny of lipid bodies (spherosomes) in plant cells. Planta 151, 109–123. doi: 10.1007/BF00387812
Wanner, G., and Theimer, R. R. (1978). Membranous appendices of spherosomes (oleosomes). Planta 140, 163–169. doi: 10.1007/BF00384916
Welte, M. A. (2007). Proteins under new management: lipid droplets deliver. Trends Cell Biol. 17, 363–369. doi: 10.1016/j.tcb.2007.06.004
Welte, M. A. (2009). Fat on the move: intracellular motion of lipid droplets. Biochem. Soc. Trans. 37, 991–996. doi: 10.1042/BST0370991
Welte, M. A. (2015). As the fat flies: the dynamic lipid droplets of Drosophila embryos. Biochim Biophys Acta BBA Mol Cell Biol Lipids 1851, 1156–1185. doi: 10.1016/j.bbalip.2015.04.002
Wessel, D., and Flugge, U. I. (1984). A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 138, 141–143. doi: 10.1016/0003-2697(84)90782-6
Wewer, V., Dombrink, I., Dorp vom, K., and Dörmann, P. (2011). Quantification of sterol lipids in plants by quadrupole time-of-flight mass spectrometry. J. Lipid Res. 52, 1039–1054. doi: 10.1194/jlr.D013987
Wilfling, F., Haas, J. T., Walther, T. C., and Farese, R. V. Jr. (2014). Lipid droplet biogenesis. Curr. Opin. Cell Biol. 29, 39–45. doi: 10.1016/j.ceb.2014.03.008
Wittstock, U., and Burow, M. (2010). Glucosinolate breakdown in Arabidopsis: mechanism, regulation and biological significance. Arab. Book Am. Soc. Plant Biol. 8:e0134. doi: 10.1199/tab.0134
Xu, C., and Shanklin, J. (2016). Triacylglycerol metabolism, function, and accumulation in plant vegetative tissues. Annu. Rev. Plant Biol. 67, 179–206. doi: 10.1146/annurev-arplant-043015-111641
Xu, N., Zhang, S. O., Cole, R. A., McKinney, S. A., Guo, F., Haas, J. T., et al. (2012). The FATP1–DGAT2 complex facilitates lipid droplet expansion at the ER–lipid droplet interface. J. Cell Biol. 198, 895–911. doi: 10.1083/jcb.201201139
Yang, L., Ding, Y., Chen, Y., Zhang, S., Huo, C., Wang, Y., et al. (2012). The proteomics of lipid droplets: structure, dynamics, and functions of the organelle conserved from bacteria to humans. J. Lipid Res. 53, 1245–1253. doi: 10.1194/jlr.R024117
Ytterberg, A. J., Peltier, J. B., and van Wijk, K. J. (2006). Protein profiling of plastoglobules in chloroplasts and chromoplasts. A surprising site for differential accumulation of metabolic enzymes. Plant Physiol. 140, 984–997. doi: 10.1104/pp.105.076083
Keywords: lipid droplet, leaf senescence, label free proteomics, Small rubber particle protein1, electron tomography, ER contact site, ultrastructure, secondary metabolism
Citation: Brocard L, Immel F, Coulon D, Esnay N, Tuphile K, Pascal S, Claverol S, Fouillen L, Bessoule J-J and Bréhélin C (2017) Proteomic Analysis of Lipid Droplets from Arabidopsis Aging Leaves Brings New Insight into Their Biogenesis and Functions. Front. Plant Sci. 8:894. doi: 10.3389/fpls.2017.00894
Received: 09 January 2017; Accepted: 12 May 2017;
Published: 29 May 2017.
Edited by:
Wei Wang, Henan Agricultural University, ChinaReviewed by:
Bing Bai, Umeå Plant Science Centre, SwedenTong Zhang, University of Florida, United States
Copyright © 2017 Brocard, Immel, Coulon, Esnay, Tuphile, Pascal, Claverol, Fouillen, Bessoule and Bréhélin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claire Bréhélin, Y2xhaXJlLmJyZWhlbGluQHUtYm9yZGVhdXguZnI=
†These authors have contributed equally to this work.