 Thaura Ghneim-Herrera1*
Thaura Ghneim-Herrera1* Michael G. Selvaraj2*
Michael G. Selvaraj2* Donaldo Meynard3
Donaldo Meynard3 Denis Fabre3Alexandra Peña1
Denis Fabre3Alexandra Peña1 Walid Ben Romdhane4
Walid Ben Romdhane4 Rania Ben Saad5
Rania Ben Saad5 Satoshi Ogawa2,6
Satoshi Ogawa2,6 Maria C. Rebolledo2Manabu Ishitani2
Maria C. Rebolledo2Manabu Ishitani2 Joe Tohme2
Joe Tohme2 Abdullah Al-Doss4
Abdullah Al-Doss4 Emmanuel Guiderdoni3*Afif Hassairi4,7
Emmanuel Guiderdoni3*Afif Hassairi4,7- 1Departamento de Ciencias Biológicas, Universidad Icesi, Cali, Colombia
- 2International Center for Tropical Agriculture, Cali, Colombia
- 3UMR Amélioration Génétique et Adaptation des Plantes Méditerranéennes et Tropicales, Centre de Coopération Internationale en Recherche Agronomique pour le Développement, Montpellier, France
- 4Department of Plant Production, College of Food and Agricultural Sciences, King Saud University, Riyadh, Saudi Arabia
- 5Biotechnology and Plant Improvement Laboratory, Centre of Biotechnology of Sfax, University of Sfax, Sfax, Tunisia
- 6Graduate School of Agricultural and Life Science, Department of Global Agricultural Science, The University of Tokyo, Tokyo, Japan
- 7Centre of Biotechnology of Sfax, Sfax, Tunisia
We evaluated the yields of Oryza sativa L. ‘Nipponbare’ rice lines expressing a gene encoding an A20/AN1 domain stress-associated protein, AlSAP, from the halophyte grass Aeluropus littoralis under the control of different promoters. Three independent field trials were conducted, with drought imposed at the reproductive stage. In all trials, the two transgenic lines, RN5 and RN6, consistently out-performed non-transgenic (NT) and wild-type (WT) controls, providing 50–90% increases in grain yield (GY). Enhancement of tillering and panicle fertility contributed to this improved GY under drought. In contrast with physiological records collected during previous greenhouse dry-down experiments, where drought was imposed at the early tillering stage, we did not observe significant differences in photosynthetic parameters, leaf water potential, or accumulation of antioxidants in flag leaves of AlSAP-lines subjected to drought at flowering. However, AlSAP expression alleviated leaf rolling and leaf drying induced by drought, resulting in increased accumulation of green biomass. Therefore, the observed enhanced performance of the AlSAP-lines subjected to drought at the reproductive stage can be tentatively ascribed to a primed status of the transgenic plants, resulting from a higher accumulation of biomass during vegetative growth, allowing reserve remobilization and maintenance of productive tillering and grain filling. Under irrigated conditions, the overall performance of AlSAP-lines was comparable with, or even significantly better than, the NT and WT controls. Thus, AlSAP expression inflicted no penalty on rice yields under optimal growth conditions. Our results support the use of AlSAP transgenics to reduce rice GY losses under drought conditions.
Introduction
Field drought could severely alter global rice production (740 million tons from over 160 million hectares of paddy in 20131). Twenty percent of Asian rice acreage is drought-prone, and the frequency, extent, and intensity of drought events are amplified by increasing climate instability (Pandey and Bhandari, 2007). Drought mainly occurs in rain-fed and upland rice growing systems; however, the need to save irrigation water in lowland paddy fields may also lead to periods of drought in more favorable environments. Rice is sensitive to drought over its growth cycle; however, drought stress is most detrimental to grain yield (GY) when it occurs at the reproductive stage (from panicle initiation to fertilization). Drought limits transpiration, which is required for panicle cooling and causes spikelet sterility, while impairing proper reserve remobilization and grain filling (Prasad et al., 2008).
Grain yield under drought occuring at flowering stage is the most reliable trait for discrimination of tolerant progeny in breeding programs (Atlin, 2003; Kumar et al., 2008). Long-term efforts to identify quantitative trait loci (QTL) associated with drought-avoidance and -tolerance have identified genomic regions involved in these complex traits (Lanceras et al., 2004; Bernier et al., 2009; Courtois et al., 2009; Khowaja et al., 2009). More recently, genes influencing drought-related traits have been cloned (Uga et al., 2013; Raorane et al., 2015). Undoubtably, the re-sequencing of genetic resources [The 3,000 genomes project (Li et al., 2014)] and target population of environments-phenotyping2 of a broad range of accessions will accelerate the identification of new loci contributing to drought-tolerance in the near-future.
An alternative strategy is the engineering of drought-tolerance using transgenic approaches, which can use various expression systems (constitutive, tissue-specific, or drought-inducible) (Gaudin et al., 2013). While an extensive range of genes from different origins has already been introduced into rice and proven to induce some level of drought-tolerance in laboratory/greenhouse experiments, reports of challenges of transgenic rice with field-drought at the reproductive stage are limited (Gaudin et al., 2013). Transgenic drought-tolerance has primarily been achieved using genes encoding transcription factors (TFs), such as members of the AP2/ERF (Oh et al., 2009; Ambavaram et al., 2014); NAC, NAM, ATAF, and CUC (Hu et al., 2006; Jeong et al., 2010; Redillas et al., 2012); and homeodomain-leucine zipper (Liu et al., 2013) families. Genes involved in abscisic acid (ABA) (Xiao et al., 2009) and proline (You et al., 2012) synthesis, along with those encoding a late embryogenesis abundant protein (Xiao et al., 2007) and a proteinase inhibitor (Huang et al., 2007), have also improved yield under drought in field experiments. In these reports, drought was imposed at the flowering stage, and observed significant gains in yield were explained by reduced sterility, enhanced root systems, and increased tiller/panicle/grain number/size. More importantly, in these examples, no yield penalties were observed under unstressed conditions; indeed, some instances of enhanced yield in the absence of stress were reported (Ambavaram et al., 2014).
Over the last 10 years, there has been increasing interest in a class of zinc-finger proteins, the A20/AN1 domain stress-associated protein (SAP) family. These molecules exhibit structural and functional conservation among plant species (Vij and Tyagi, 2008; Giri et al., 2013). The rice genome harbors 18 SAP genes encoding the A20/AN1 domain, while Arabidopsis contains 14. Interestingly, the majority of the published attempts to constitutively express SAPs in the model plants, tobacco, Arabidopsis, and rice, have resulted in enhanced tolerance to multiple abiotic stress factors, including drought, salinity, cold, heat, oxidative stress, and heavy metals. Similar examples of transgenic protection have also recently been reported in durum wheat (Ben Saad et al., 2011), banana (Sreedharan et al., 2012), and cotton (Hozain et al., 2012).
Stress-associated proteins can exhibit various modes of action. Recent reports of experiments involving Arabidopsis AtSAP5 (Kang et al., 2011, 2013; Choi et al., 2012) and rice OsSAP1/OsSAP11 (Giri et al., 2011) indicate that SAPs can recognize and bind polyubiquitin chains, may exhibit E3-ligase activity, and act through protein–protein interactions involving their A20/AN1 zinc finger domains. SAPs also have a well-established roles in the negative regulation of ABA [AtSAP5 (Kang et al., 2013), OsSAP7 (Sharma et al., 2015)], and gibberellic acid [OsSAP11/OsDOG (Liu et al., 2011)] mediated responses. Other recent evidence indicates that AtSAP12 may have a redox sensor role via cysteine residues outside of the conserved A20/AN1 domain (Stroher et al., 2009). Ectopic accumulation of SAPs can influence the expression of abiotic stress-related genes in Arabidopsis, rice, and cotton; however, a function for these proteins as transcription regulators through direct DNA-binding has been suggested but not demonstrated. A recent transcript profiling report in rice indicated that overexpression of OsSAP1 influenced the transcription of 150 genes, including 43 involved in stress responses (Dansana et al., 2014).
Based on the hypothesis that halophytes could be a valuable source of adaptive genes, we isolated an A20/AN1 SAP encoding gene, AlSAP, from the C4 halophyte grass Aeluropus littoralis (Zouari et al., 2007). AlSAP protein exhibits 79% sequence similarity with rice OsSAP9, is inducible by multiple stress factors (Ben Saad et al., 2010), and, when expressed in transgenic tobacco, durum wheat (Ben Saad et al., 2011), and the japonica rice cv. Nipponbare (Oryza sativa L. ‘Nipponbare’) (Ben Saad et al., 2012), confers enhanced tolerance to cold, drought, salt, and oxidative stress. Under greenhouse conditions, AlSAP transgenic rice plants survived a severe 15-day dry-down experiment (DDE) imposed at the six-leaf, early tillering stage. AlSAP-lines rehydrated at the end of the stress period yielded 50% of the grain produced under unstressed conditions, whereas wild-type Nipponbare (WT-NIP) did not recover from the treatment. Physiological investigations conducted during the DDE revealed that the enhanced performance of the transgenics was likely due to the maintenance of photosynthesis and the integrity of the photosynthetic apparatus under stress (Ben Saad et al., 2012). Importantly, over-expression of AlSAP in rice did not appear to alter its performance under unstressed greenhouse growth conditions. This contrasts with the reports of OsSAP overexpression in rice, where a reduction in GY was observed under unstressed conditions (Kanneganti and Gupta, 2008; Dansana et al., 2014).
As mentioned above, transgenic protection from drought requires testing in the field at several locations and with stress imposed at the most sensitive stage, from panicle initiation to fertilization. Also, potential detrimental or beneficial effects of transgene expression require assessment under stringent yield trials. With this aim, we set up three replicate field trials in Colombia, with drought stress imposed at the flowering stage, either through a naturally occurring dry season or by placement of the field trial under a rainout shelter. Six AlSAP-lines, along with non-transgenic (NT) azygous control lines and WT-NIP were challenged at panicle initiation stage with different soil drought conditions. During these trials, we also monitored the status of plants at the physiological and biochemical levels, to investigate AlSAP function. Parallel, irrigated trials were conducted to determine whether the constitutive expression of AlSAP alters rice yields under unstressed conditions.
Data from the three drought field trials consistently indicated that the two highest-expressing AlSAP lines out-yielded the NT and WT controls under drought field conditions. Consistent and significantly higher yields were also observed among AlSAP-lines in irrigated trials. No significant physiological or biochemical differences were observed between AlSAP-lines and WT and NT controls. Our results suggest that the superior performance of AlSAP-lines under field drought was likely due to a primed status, with beneficial features, including enhanced biomass, accumulated during vegetative growth, that facilitate improved tolerance of water deficit at the flowering stage.
Materials and Methods
Ethical Statement
The authors declare that the transgenic experiments described herein comply with the current biosafety laws of the country in which they were performed.
Rice Transformation
The isolation and cloning of the AlSAP gene from A. littoralis was described previously (Ben Saad et al., 2011, 2012). The AlSAP gene and protein sequences are deposited in the GenBank (DQ885218) and UniProtKB (A1YAQ3) databases. The rice genome contains 12 SAP genes encoding A20 and AN1 domains. The relationship between AlSAP and OsSAP proteins was established by Ben Saad et al. (2010); the SAP gene encoding the protein sharing the highest amino acid identity (79%) is OsSAP9 (Os07g07350) which is the likely ortholog of AlSAP in rice (Supplementary Figure S1).
We evaluated six AlSAP-lines and their NT azygous controls in our field trials (Supplementary Figures S2, S3). RN4, RN2, and RN5 (T5 homozygous lines) expressed AlSAP under control of the 35S CaMV promoter and accumulated AlSAP transcripts at increasing levels (Ben Saad et al., 2012). RN1 and RN3 (T3 homozygous material) expressed AlSAP under the control of the maize Ubiquitin1 regulatory region (Christensen et al., 1992). RN6, also a T3 homozygous material, expressed AlSAP under the rice ACT1 gene regulatory region (Mc Elroy et al., 1990). T3 homozygous RN3, RN1, and RN6 plantlets accumulated AlSAP transcript at moderate, medium, and high levels in leaves, respectively, at the six leaf stage under greenhouse conditions. Each of the three lines harbored a single copy of the transgene (Supplementary Figure S3).
Experimental Field Design
In Trial 1, six homozygous AlSAP-transformed lines (RN1–6), together with their transformation controls, and non-transformed WT-NIP plants were evaluated in a natural upland rain-fed field from November 2012 to March 2013 at CIAT Santa Rosa station (Villavicencio, Colombia). The transgenic and WT-NIP lines were sown in three replicate 2 m × 1 m plots with a spacing of 25 cm × 10 cm and received regular fertilization. The plants experienced a 42-day rain-free period, which began at panicle initiation and extended for approximately 28 days after attainment of 50% flowering (Supplementary Figure S5).
Trials 2 and 3 were performed in the rainout shelter facilities at CIAT headquarters (Palmira, Colombia); the drought treatment was imposed at panicle initiation stage by suspending irrigation. In Trial 2 (April–July 2013), RN2, RN5, and RN6 were evaluated, together with their NT controls and WT-NIP, while in Trial 3 (June–October 2013) only RN5 and RN6 were assessed, together with their NT controls, and WT-NIP. Lines were sown in a randomized complete block design, with three replicates. Each plot had six rows of 16 plants with a spacing of 20 cm × 10 cm. WT-NIP plants were inserted after every four transgenic line plants for comparison. Drought was imposed for 22 days (Trial 2) and 19-days (Trial 3), after which the plants received full irrigation.
For both Trials 2 and 3, a duplicate set of materials were also sown under flooded conditions in another field, 5 m away from the rainout shelter, to evaluate differences in GY between transgenic and control rice under irrigated conditions.
Harvest was completed for all trials 107–114 days after sowing (DAS).
Soil Moisture Measurement and Calculation of Soil Water Matric Potential
The progress of drought-stress was monitored by measuring changes in soil moisture at four depths, 20, 40, 60, and 80 cm, using Aquapro sensors (Reno, NV); polycarbonate access tubes were installed at regular intervals within each replicate, and readings were taken regularly during the stress period. Humidity data were transformed to soil matric potential (Ψsoil) values using soil water retention curves (SWRC) built for each location. Aquapro soil moisture values (%) were converted to volumetric soil water content using the equation:
where 𝜃vol is the volumetric soil moisture content (%), A is the Aquapro measurement value (Aquapro %), and b is a constant depending on soil type (Tromp-van Meerveld and McDonnell, 2006). SWRC were obtained experimentally using the pressure plate apparatus method (Bittelli and Flury, 2009).
Measurement of Yield and Growth Parameters
Single plant yield and yield components were measured in each plot for all trials. Yield parameters were scored for 15 plants. Unfilled and filled grains were independently counted and weighed. Grain weight was used to determine final yields at 14% moisture content. The following agronomic traits were scored as established in the Standard Evaluation System for Rice (IRRI, 2013): flowering date, plant height (cm), total aerial biomass (g), panicle length (cm), number of tillers, number of panicles, total grain weight (g), and grain fertility (%).
Photosynthetic Performance
The rate of light-saturated photosynthesis (Asat), stomatal conductance (Gs), and intercellular CO2 concentration (Ci) of expanded flag leaves were measured from 9:00 to 12:00 h using an LI-6400 portable photosynthesis open system (Li-Cor Biosciences, Lincoln, NE, United States). Measuring conditions were as follows: photosynthetic photon flux intensity = 1500 μmol/m2/s, leaf temperature = 27.0°C ± 1°C, ambient CO2 concentration = 380 ± 10 μmol/mol, and relative humidity = 60%. The rate of CO2 saturated photosynthesis (Amax) was measured under the conditions described above, except that the ambient CO2 concentration was 1800 μmol/mol.
Leaf Rolling, Leaf Drying, and Drought Recovery Scores
Leaf rolling (LR) was characterized at regular intervals in drought-stressed plants using a scoring scale from 0 (unrolled) to 9 (tight rolling) according to the Standard Evaluation System for Rice (IRRI, 2013). Leaf drying (LD) was evaluated at the peak of stress using a scoring system from 0 (no symptoms) to 9 (dead plant). The drought recovery score (DRR) was recorded 10 days after rewatering using a scoring system from 1 (90–100% recovery) to 9 (0–19% recovery) (IRRI, 2013). All evaluations were performed at noon using whole plants.
Leaf Water (Ψw)
On three different occasions during Trial 2 (before stress, at stress peak, and 10 days after rewatering), Ψw was evaluated (pre-dawn) in drought-stressed plants. Measurements were performed on fully expanded flag leaves, as previously described (Turner, 1981), using a pressure chamber (Soil Moisture Equipment 3005F01) with a measurement limit of -15 MPa.
Oxidative Damage and Antioxidant Capacity in Leaf Tissues
The antioxidant capacity was determined using the ABTS∙+ test system, based on the method of (Du et al., 2009). The level of lipid peroxidation in leaf tissues was measured as MDA content, following a published protocol (Ueda et al., 2013).
Monitoring of AlSAP Transcript Accumulation
Procedures for Southern and northern blot analyses, and RT-PCR were the same as those detailed by Ben Saad et al. (2012). Total DNA and RNA were isolated from leaf tissue of six leaf stage seedlings, and hybridizations performed using P32 labeled Npt2, Hpt, or AlSAP gene probes. RT-PCR analyses were performed using RNA isolated from leaves and primers specific to AlSAP and the housekeeping gene, OsExp (Caldana et al., 2007), used as an internal expression standard (Supplementary Table S1). For samples from field grown plants, inflorescences were collected from the master tiller of three independent plants per line at flowering stage, pooled and stored. RNA isolation and northern blot procedures were performed as indicated above (Ben Saad et al., 2012).
Statistical Analyses
Data for days to flowering, height, total aerial biomass, GY, yield components, leaf MDA, and TEAC-ABTS were subjected to General Linear Model (GLM) analysis, appropriate for a randomized complete block design. Differences between mean values from WT-NIP- and AlSAP-transformed lines were compared using Dunnett’s test. A GLM was also used to test the effect of drought treatment (days after the last irrigation), and repetition on the soil matric potential (Ψsoil) measured at a depth of 40 cm in Trials 2 and 3. All statistical analyses were performed using SAS v9.2 (SAS Institute Inc., Cary, NC, United States). Data (means ± standard errors of raw data) are presented graphically using GraphPad Prism (GraphPad PRISM for Windows v.7.0, GraphPad Software Inc., La Jolla, CA, United States).
Results
Drought Intensity Characterization
Drought intensity and duration varied among trials with harsher conditions in Trial 1 (Santa Rosa station). In this trial, drought progressed rapidly in the first 20 cm of soil, with Ψsoil falling under the permanent wilting point (PWP, -1.6 MPa) 10 days after stress initiation (DASI). Changes in the 40 cm layer proceeded more slowly, with Ψsoil reaching -1.6 MPa by day 22. Thereafter, Ψsoil decreased steadily until the first rains occurred. A reduction in Ψsoil was also observed at 60 cm depth; however, minimum values were approximately -1.0 MPa by the end of the rain-free period (Figure 1A). In the rainout shelter in Trial 2 (Palmira station), Ψsoil in the 20 cm soil layer also fell below the PWP 10 DASI and reached very low values during the drought period (Figure 1B). However, Ψsoil changes at depths of 40 and 60 cm were less pronounced than those in Trial 1. Drought developed in a similar fashion in Trial 3 (Palmira), although the overall stress intensity was less severe than that in Trial 2 (Figure 1C). No significant differences in Ψsoil were detected between replicates in any trials, indicating that drought-stress progressed uniformly among replicates (Supplementary Table S2).
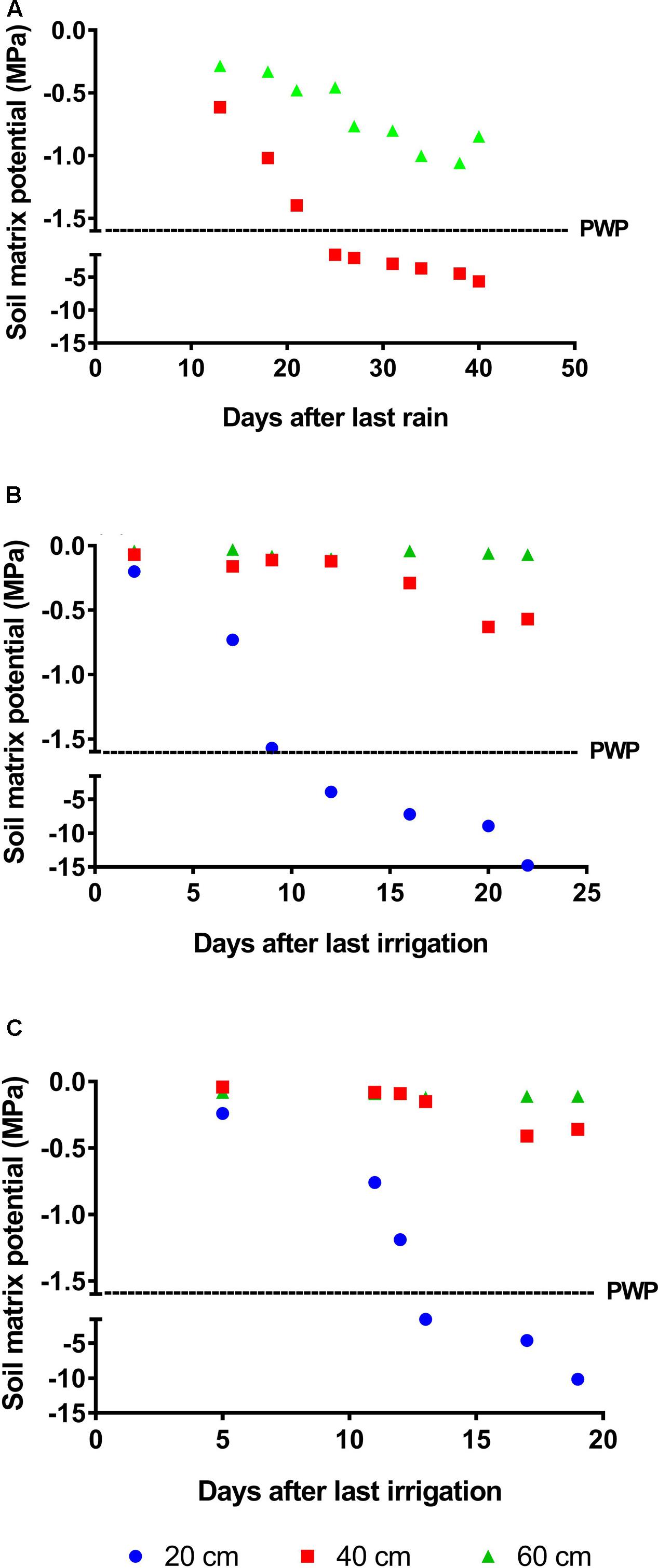
FIGURE 1. Progress of drought-stress measured as changes in ψsoil. (A) Trial 1 was performed under upland conditions in Santa Rosa (Villavicencio, Colombia). In this trial, at a depth of 40 cm the permanent wilting point (PWP, –1.6 MPa) was reached 25 days after the initiation of the rain-free period. Trials 2 and 3 were performed under the rainout facilities at CIAT headquarters (Cali, Colombia). Drought stress was imposed by suspending irrigation and allowing the soil to dry out. (B) In Trial 2, plants were kept under stress for 22 days, and ψsoil at 40 cm reached –0.6 MPa by day 20. (C) For Trial 3, plants were subjected to drought for 19 days; in this trial, relatively higher ψsoil at 40 cm depth (–0.45 MPa) indicated that the plants were submitted to a milder stress relative to the previous trials.
The Nipponbare root system normally develops in the first 40 cm of soil (Hanzawa et al., 2013). Thus, we considered changes in Ψsoil at depths of 0–40 cm good indicators of the level of drought-stress experienced by plants in the field.
Yields under Drought
In Trial 1, six homozygous AlSAP-transformed lines (RN1, RN2, RN3, RN4, RN5, and RN6) (generations T3–T5) were evaluated together with their azygous NT controls and WT-NIP. RN2, RN4, and RN5 harbor the AlSAP coding sequence under the control of the CaMV35S promoter (Ben Saad et al., 2012), while RN1, RN3, and RN6 contain one copy of a T-DNA containing the AlSAP coding sequence directed by the maize Ubi-1 promoter or the rice ACT1 promoter (Supplementary Figures S2, S3). All materials reached 50% flowering around 45–47 DAS, indicating that AlSAP accumulation did not affect plant phenology. The accumulation of AlSAP transcripts was monitored by northern blot analysis in inflorescence tissues collected from field grown plants at the initiation of drought stress imposition (Supplementary Figure S4). AlSAP transcript accumulation in these tissues was comparable in RN5 and RN6 and lower in RN2. On the other hand, the level of accumulation in RN1, RN3, and RN4 was very low compared to their respective levels in leaves of young plants at the previous generation. This was particularly surprising for the RN1 line which was identified as a medium expresser. Previous literature report describes high activity of the maize Ubi-1 regulatory regions in reproductive organs of rice (Cornejo et al., 1993). This may tentatively be ascribed to a regulation of the maize ubiquitin promoter under field conditions or to a silencing of the transgene.
In this trial, the plants were subjected to 42 days of drought stress from panicle initiation to 28 days after the plants reached 50% flowering, which caused a marked reduction in Ψsoil. Under these severe stress conditions, RN2, RN5, and RN6 were able to maintain their green biomass (Supplementary Figure S6) and exhibited higher GY than WT-NIP and NT controls, with relative gains of 46, 97, and 115%, respectively, compared with NT controls (Figure 2A). The NT controls tended to be more affected by drought stress than WT-NIP and, although this difference was not significant, it was consistent. However, the RN1 and RN3 NT control lines exhibited significantly lower GY (p < 0.001) than either the WT-NIP or NT controls of the other transformation events following the field drought-stress. This depressive effect could be due to somaclonal variation. We also see previously that these two lines eventually expressed AlSAP at a low level under field conditions. Interestingly, a beneficial effect of AlSAP expression was still observed in these variants; however, this was not sufficient to cause yields significantly higher than those of WT-NIP. The four other lines could be grouped into two classes according to their performance relative to WT-NIP, and their level of accumulation of AlSAP transcripts. The low level of transcript expression in the RN4 line did not result in enhanced yield protection under drought stress at flowering, whereas the medium and high expression levels found in RN2, RN5, and RN6 conferred enhanced tolerance to these lines. Therefore, lines RN1, RN3, and RN4 were not included in subsequent drought trials.
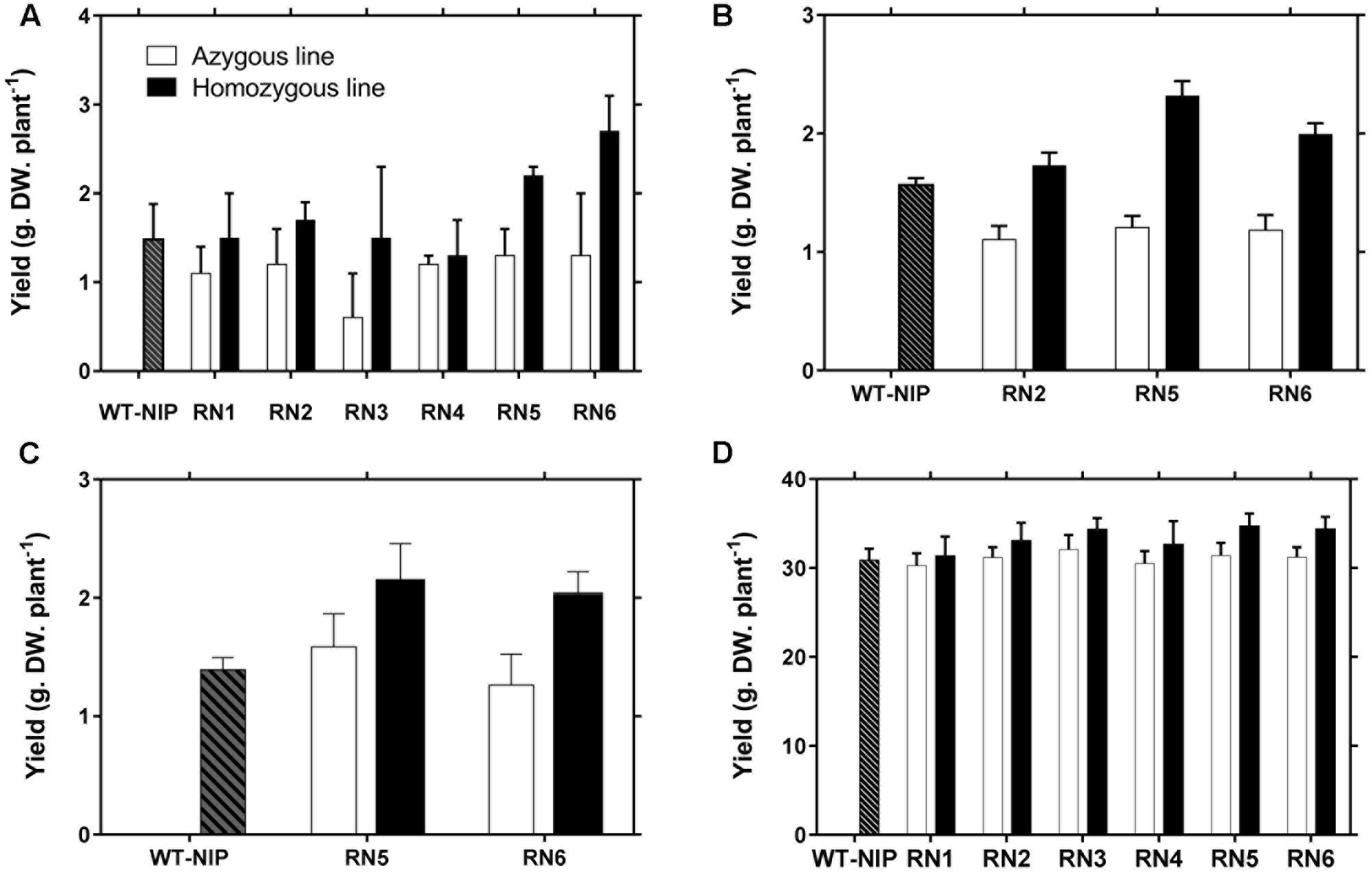
FIGURE 2. Improved grain yield in AlSAP-lines exposed to drought stress. (A) Grain yield (GY) of WT-NIP and AlSAP-lines RN1–6 in Santa Rosa (Trial 1). (B,C) As for the Santa Rosa trial, RN5 and RN6 outperformed WT-NIP and their NT controls under the drought conditions imposed in Trials 2 and 3. (D) Higher GY for AlSAP-lines RN1–6 was also observed in the irrigated control conducted in Trial 2.
Trials 2 and 3 were conducted using a rainout shelter to impose drought-stress. In Trial 2, RN2, RN5, and RN6 consistently achieved higher GYs than their NT controls, exhibiting GY increases of 57, 92, and 69%, respectively (Figure 2B). RN5 and RN6 also exhibited superior GY in Trial 3 (Figure 2C). The higher GYs were associated with significantly higher numbers of panicles (p < 0.001), higher fertility (p < 0.001), and higher accumulation of biomass in AlSAP-lines (p < 0.001) (Table 1). All lines flowered around the same date, 53–55 DAS; thus, they were at very similar physiological stages when subjected to drought (Table 1).
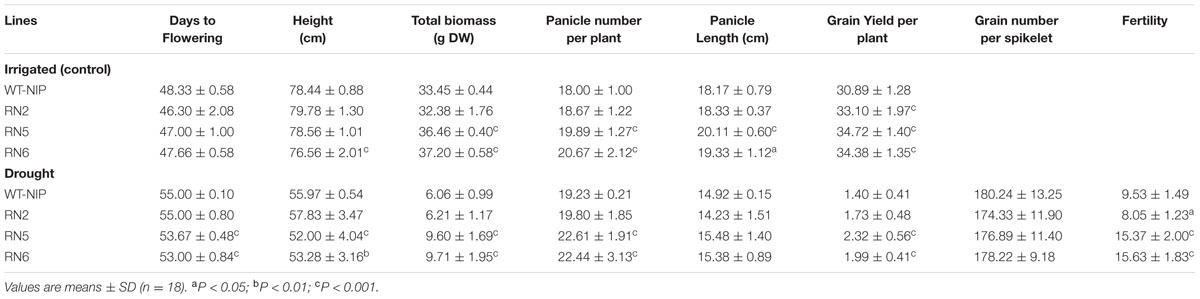
TABLE 1. Days to flowering, height, total aerial biomass, grain yield, and yield components under irrigated and drought stress conditions in the rainout shelter (Trial 2).
In Trial 2, lines RN1–6 were also grown under irrigated conditions, along with their NT controls and WT-NIP to evaluate the effect of AlSAP accumulation in the absence of drought-stress. Interestingly, under irrigated conditions, all NT controls behaved in the same manner as WT-NIP, while the transgenic lines tended to outperform both types of controls (Figure 2D). This indicates that the depressed background represented by the RN1 and RN3 NT controls influence performance only under drought conditions and, could, therefore, affect stress-related traits. Irrigated plots were also established in the third field trial. Under irrigation, the accumulation of AlSAP significantly enhanced GY (p < 0.001). As observed under drought-stress, this effect was related to both higher numbers of panicles (p < 0.001) and accumulation of vegetative biomass (p < 0.001) (Table 1).
Together, these results demonstrate that AlSAP expression increases GY under drought-stress imposed at the reproductive stage, through a mechanism that involves the maintenance of vegetative biomass, a higher number of panicles, and enhanced grain fertility. Furthermore, AlSAP-lines also exhibited higher yields, related to higher panicle numbers and increased vegetative biomass, when grown under irrigation.
Sensitivity to Drought
In Trial 2, we compared the drought sensitivity of AlSAP-lines, their NT controls, and WT-NIP by monitoring the progress of LR, LD, and DRRs (IRRI, 2013). LR appeared earlier and proceeded more rapidly in WT-NIP plants, reaching a score of 5.5 at the peak of stress (22 DASI) (Figure 3A), indicating that leaves were fully cupped. At this time, LR scores for RN5 and RN6 were 3.0 and 3.6 (leaves folding), respectively. At peak stress, the LD score for WT-NIP was 3.2 (drying up to ¼; of the length of the leaf), while RN5 and RN6 scores were 1.6 (slight tip drying) (Figure 3B). DRR was evaluated 10 days after rewatering, RN5 and RN6 showed better recovery (DRR = 1.7) than WT-NIP (DRR = 3.0). These results indicate that the best-performing AlSAP lines were much less sensitive to drought and exhibited improved recovery after rehydration compared with controls (Figure 3C).
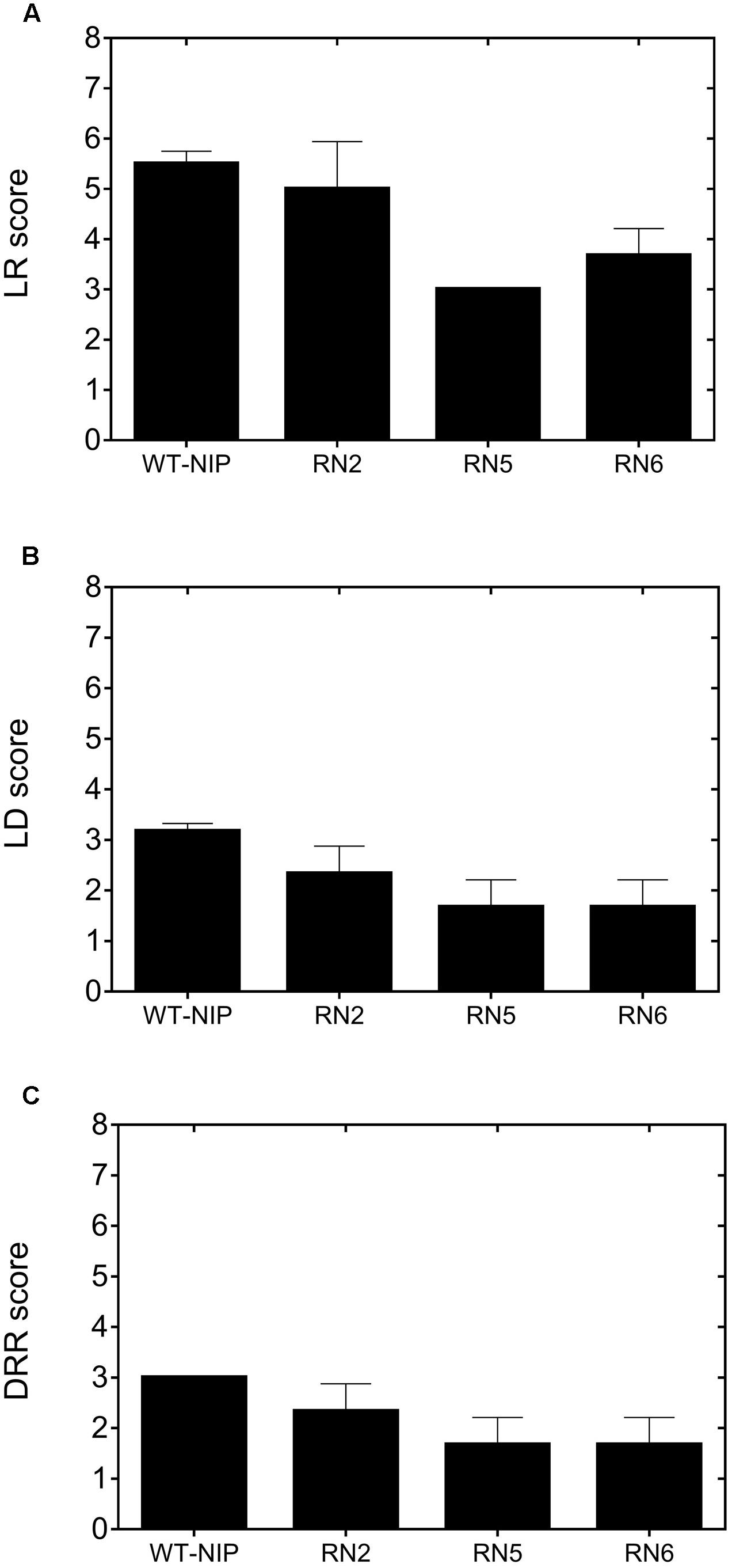
FIGURE 3. Leaf rolling (A, LR), leaf drying (B, LD), and drought recovery scores (C, DRR) exhibited by AlSAP-lines and WT-NIP. AlSAP-lines were less sensitive to drought and exhibited better recovery after rewatering. LR and LD were recorded at the peak of stress, while DRR was evaluated 10 days after rewatering. All measurements were taken at noon.
Leaf Water Potential (ψw)
In rice, the ability to maintain ψw at the flowering and grain filling stages is related to variation in GY under drought-stress (Sibounheuang et al., 2006). Furthermore, there is a negative correlation between ψw and the percentage of spikelet sterility in rice plants subjected to drought at flowering (Jongdee et al., 2002). Therefore, in Trial 3 we evaluated whether the GY differences observed among RN5, RN6, and WT-NIP were related to an improved ability to maintain higher ψw.
No significant differences in ψw were detected before the imposition of drought stress, with ψw values oscillating between -0.15 and -0.50 MPa (Figure 4). After 19 days under drought-stress, ψw decreased significantly in all lines, reaching values of -0.90 to -2.10 MPa. RN6 plants showed slightly, but not significantly, higher ψw than RN5 and WT-NIP. Rewatering of plants resulted in ψw recovery to pre-drought values, with no significant differences among lines (Figure 4).
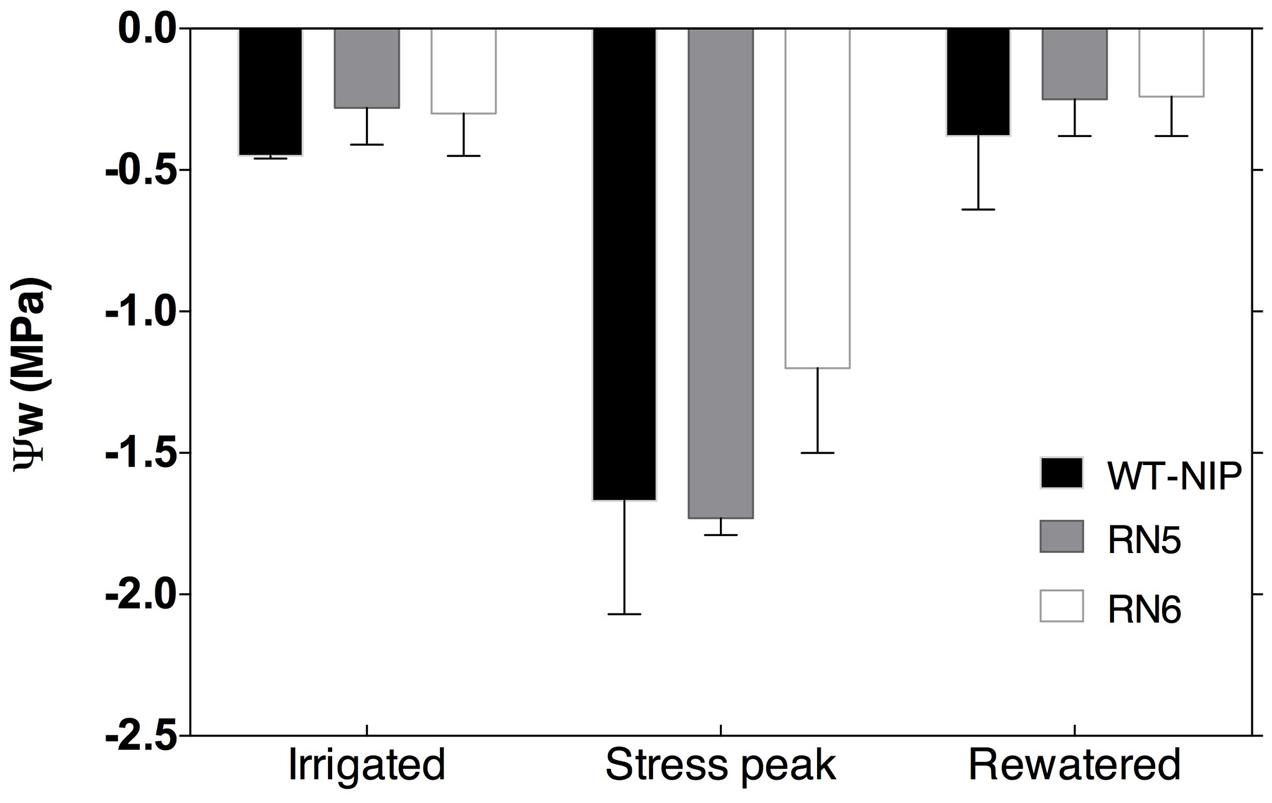
FIGURE 4. Pre-dawn leaf water potential (ψw) in RN5, RN6, and WT–NIP, measured before stress (irrigated), at peak stress, and 10 days after rewatering.
Photosynthetic Performance
Flag leaves of AlSAP-lines and WT-NIP showed a marked reduction in Asat under the stress conditions imposed in Trial 2 (Figure 5A). Plants exhibited Asat rates of approximately 30 μmol/m2/s before the imposition of drought; however, rates were reduced by approximately 70% at the peak of drought stress. The reduction in Asat was matched by a strong diminution in Gs and Ci (Figures 5B,C), indicating that stomatal closure intensely limited photosynthesis.
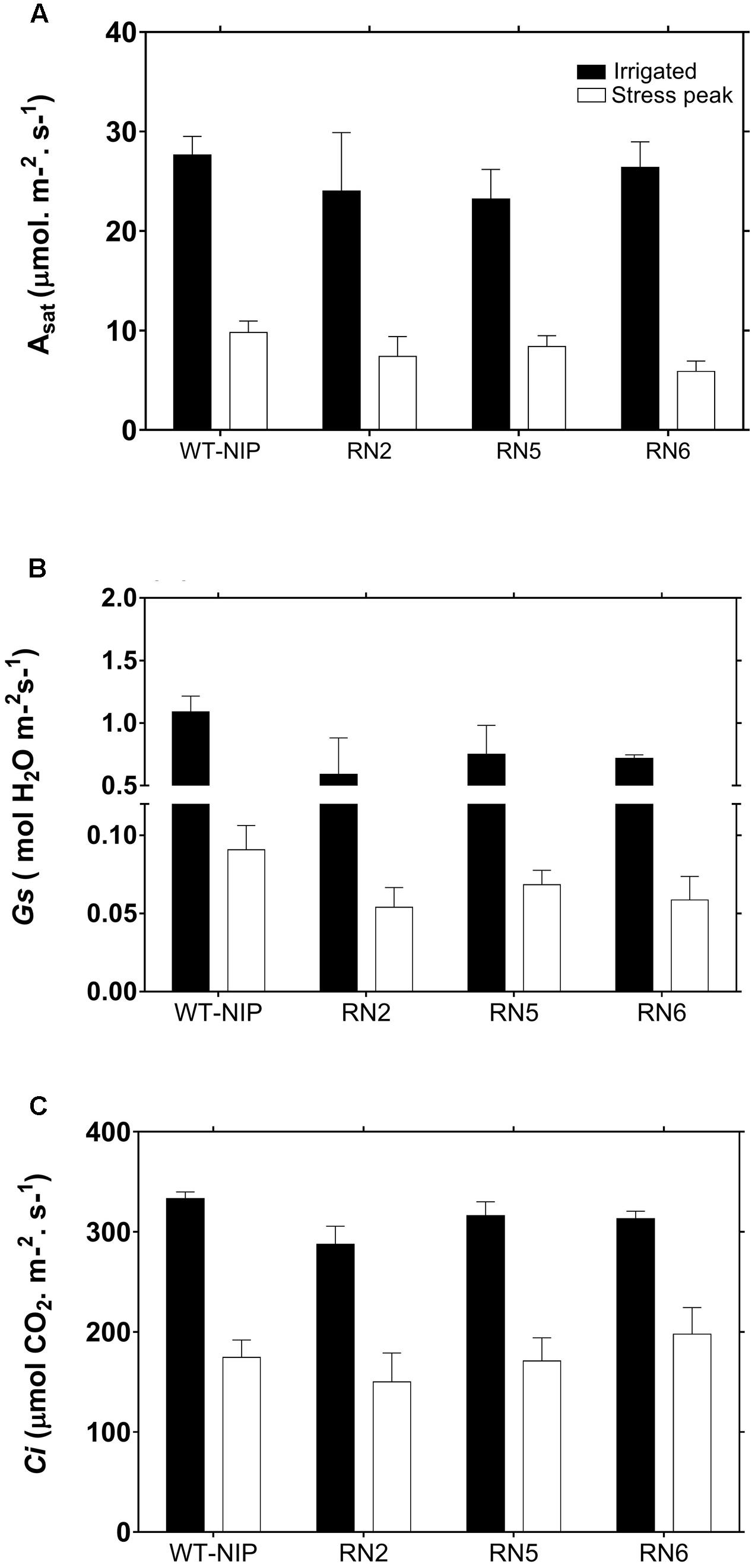
FIGURE 5. Instantaneous photosynthesis rate (A, Asat), stomatal conductance (B, Gs), and internal CO2 concentration (C, Ci) for WT-NIP and AlSAP-lines under irrigated and drought conditions. No significant differences were observed between AlSAP-lines and WT-NIP under the stress conditions imposed in Trial 2. Photosynthetic performance was evaluated before (irrigated) and 19 days after drought-stress was imposed (stress peak). [CO2]ambient = 380 mmol/mol, photosynthetic photon flux intensity = 1500 mmol/m2/s, RH = 60%, LeafTemp = 28°C.
To establish whether the reduction in Asat was exclusively due to diffusive limitations, we evaluated the photosynthetic responses of RN5 and WT-NIP under a saturating CO2 concentration (Amax) at regular intervals during the drought period and 10 days after rewatering (Figure 6A). Under irrigated conditions, Amax was approximately 46 μmol/m2/s; however, it reduced progressively during drought reaching values of 21 μmol/m2/s, at the peak of stress. After rewatering, Amax did not fully recover to pre-drought levels. These results indicate that the reduction of photosynthesis also involved biochemical damage. The similar responses exhibited by RN5 and WT-NIP suggest that AlSAP expression did not offer protection to the photosynthetic machinery under the high stress levels imposed in Trial 2.
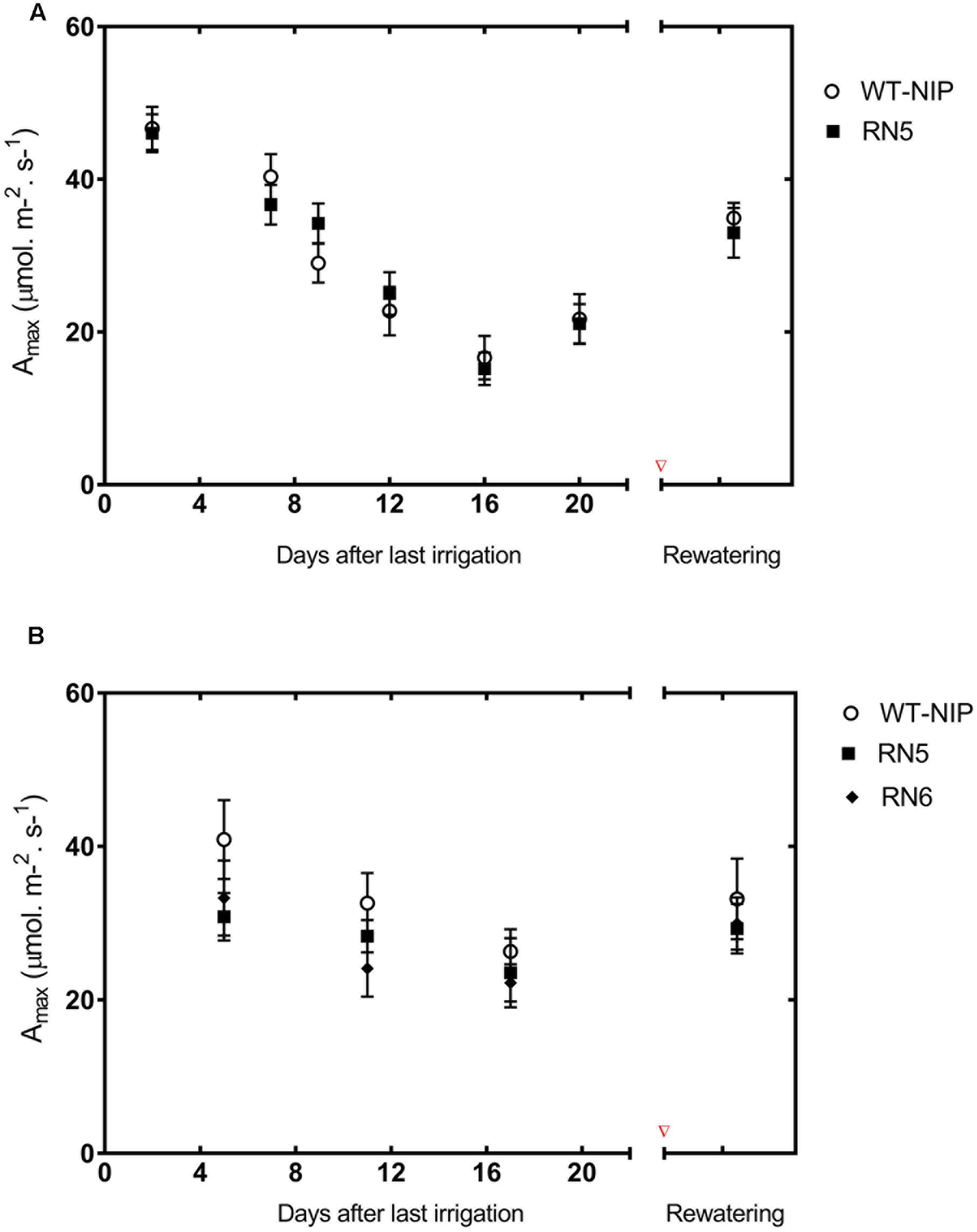
FIGURE 6. Drought-induced changes in CO2 saturated photosynthesis rate (Amax) during Trials 2 (A) and 3 (B). Regular measurements were taken during the drought stress period. Experimental plots were re-irrigated when leaves started to fold, and Amax was measured again 10 days later. [CO2]ambient = 1800 mmol/mol, photosynthetic photon flux intensity = 1500 mmol/m2/s, RH = 60%, LeafTemp = 28°C
We also evaluated Amax under the milder stress conditions imposed in Trial 3. RN6 was also included in this evaluation. Similar to Trial 2, we observed a progressive diminution of Amax in all lines as drought-stress intensified (Figure 6B). However, Amax was reduced to a lesser extent in AlSAP-lines, and recovered almost entirely after rehydration. At the peak of stress, Amax in AlSAP-lines was reduced by 24% compared to the initial rate, while in its NT control Amax diminished by 52%. After reirrigation, AlSAP-lines showed a 95% recovery of Amax; however, the NT control attained only 59%. Thus, it appears that AlSAP may exert a “protective” effect on photosynthesis, specifically at the biochemical level, and that this effect is modulated by the intensity of drought-stress.
Oxidative Stress
Previous work on AlSAP-transformed rice demonstrated that detached leaves of AlSAP-lines maintained high chlorophyll content than controls after exposure to high concentrations of H2O2, indicating tolerance to oxidative stress. Additionally, AlSAP-lines showed enhanced expression of the genes OsSodA and OsAPX1, encoding the enzymes manganese superoxide dismutase and ascorbate peroxidase, respectively, which are involved in oxidative protection (Ben Saad et al., 2012).
Based on these results, in Trial 3 we evaluated whether a superior capacity for removal of reactive-oxygen-species (ROS) was involved in the enhanced drought tolerance observed in AlSAP-lines. We quantified the content of MDA in leaves to monitor the degree of oxidative stress induced by drought (Figure 7). Leaf MDA concentrations increased in response to drought in all lines and continued to rise after rehydration (p < 0.01). RN5 and RN6 exhibited significantly higher MDA content than NT controls before the drought, at the peak of drought-stress, and 10 days after rewatering (p < 0.01). No association was observed between the concentration of MDA and chlorophyll content (data not shown). All plants exhibited decreases in chlorophyll content during the experiment; however, no significant differences were observed among lines (data not shown).
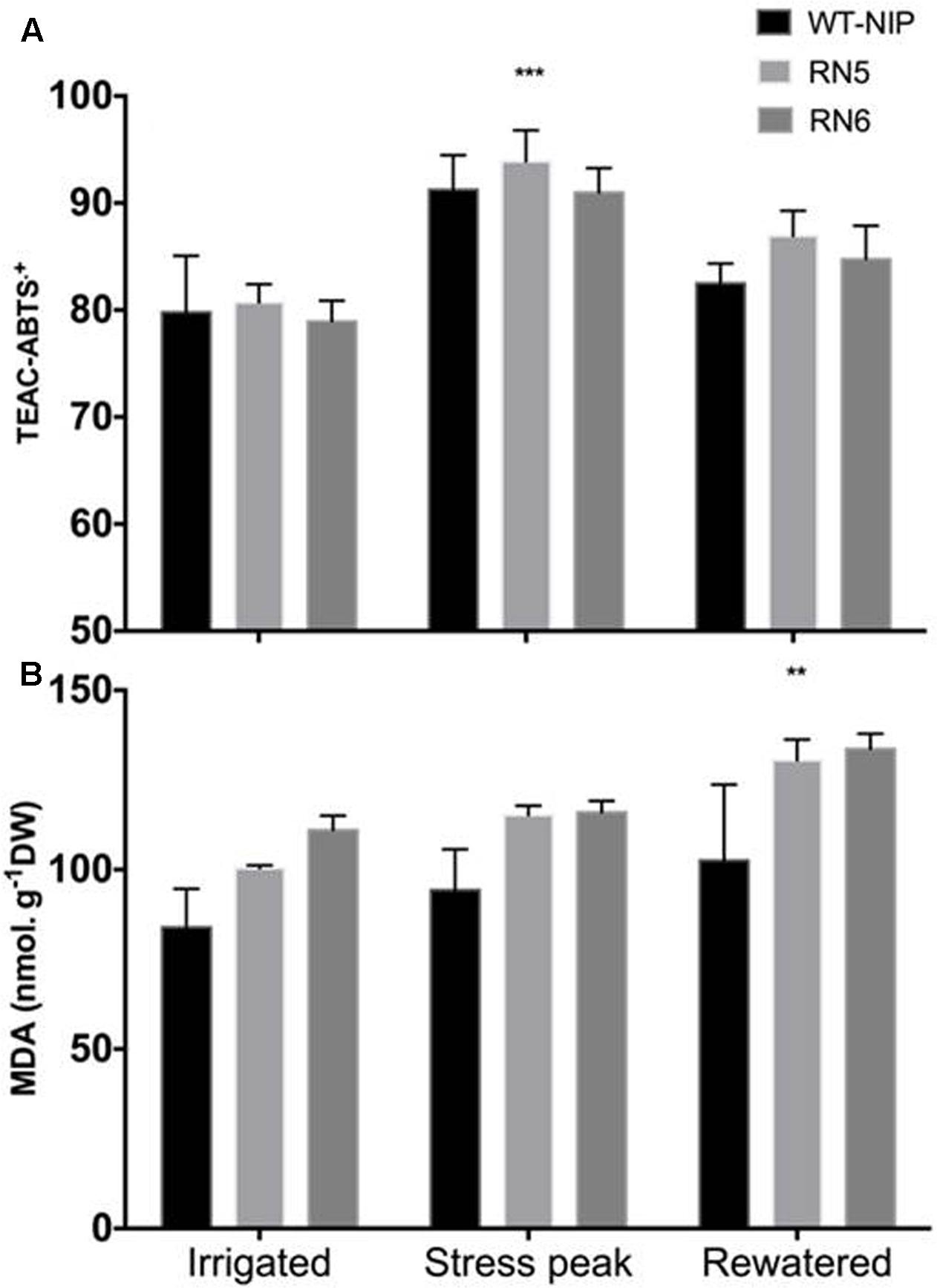
FIGURE 7. Antioxidant capacity determined by radical cation ABTS∙+ and MDA content under irrigated, drought, and rewatered conditions in Trial 2. (A) Drought stress induced a significant increase (p < 0.001) in the antioxidant capacity, measured by the ABTS assay. No significant differences were detected among lines. (B) A significant increase in MDA (p < 0.01) was measured in all lines after rewatering. Significance level ∗∗p < 0.01, ∗∗∗p < 0.001.
We also evaluated the capacity of plants to remove ROS using the ABTS•+-based system, which estimates total antioxidant capacity (Tirzitis and Bartosz, 2010). No significant differences were observed among lines at any time point (Figure 7); however, a marked increase in antioxidants was observed in response to drought. The antioxidant capacity decreased after rewatering, although it remained higher than that observed before stress, possibly due to stress priming (Wang et al., 2014).
Discussion
GY Is Protected from Field Drought at the Reproductive Stage by AlSAP
In this investigation, we characterized the field performance of rice lines (O. sativa L. ‘Nipponbare’) transformed with the AlSAP gene isolated from the halophyte grass A. littoralis. Our results demonstrate that the constitutive expression of AlSAP did not involve growth, development or phenology trade-offs, and confirm a role for AlSAP in enhancing rice tolerance to drought stress.
AlSAP expression improved the tolerance of rice to drought stress imposed at the reproductive stage, and this effect translated into higher GY. In our three independent field trials, AlSAP-lines achieved higher GYs than NT and WT controls when exposed to drought-stress of different intensities, complementing previous findings by Ben Saad et al. (2012) indicating superior drought tolerance at the vegetative stage. The higher GY corresponded with a significant increase in panicle number and spikelet fertility. No differences were observed in the number of spikelets per panicle, nor in individual grain weight. We disregarded the hypothesis that differences in panicle number and spikelet fertility were due to phenology, as all the lines flowered at around the same time and drought stress began at heading, when the number of panicles was already determined. In fact, we observed no difference in panicle number between plants grown under irrigated and drought conditions for either WT-NIP or AlSAP-lines.
For RN2, RN4, and RN5, the same ranking in stress tolerance (RN5 > RN2 > RN4) correlating with levels of AlSAP transcript accumulation was observed under field drought conditions and previous laboratory and greenhouse experiments (Ben Saad et al., 2012). The RN6 line, harboring the AlSAP coding sequence under the control of the rice ACT1 promoter, performed similarly to RN5. In contrast, somaclonal variation in Ubi1 NT controls, led to inferior performance under drought conditions. This depressed background likely prevented expression of the full benefit of a low level of AlSAP accumulation in the transgenic lines. Variation among plants derived from tissue culture is an extensively reported phenomenon, including in transgenic lines tested for drought-tolerance (Xiao et al., 2007, 2009), and has both genetic (Miyao et al., 2012) and epigenetic (Stroud et al., 2013) origins. These reports stress the requirement for evaluation of homozygous transgenic lines alongside their azygous, sibling progenies.
Drought Protection in AlSAP-Lines Cannot Be Explained by Maintenance of Photosynthesis
Photosynthesis and yield are positively correlated in rice under both irrigated and drought conditions (Centritto et al., 2009; Gu et al., 2012). However, the GY differences observed between AlSAP-lines and WT-NIP under drought-stress were not clearly associated with Asat in flag leaves. This result contrasts with those obtained by Ben Saad et al. (2012), which demonstrated enhanced photosynthetic performance in RN5 under mild and severe drought applied at the vegetative stage, a response engendered by the capacity of this transgenic line to maintain elevated Gs. Similar to that report, we found that stomatal closure limited Asat under drought conditions; however, we did not detect an improved capacity to maintain open stomata in AlSAP-lines. Furthermore, evaluation of Amax during drought and after rehydration indicated that similar non-stomatal limitations operate in AlSAP-lines and WT-NIP under severe stress conditions.
Although we did not observe differences in instantaneous Asat on a single leaf basis, it is still possible that AlSAP-lines may have achieved improved photosynthetic performance. RN5 and RN6 exhibited moderate LR, which can increase photosynthesis and GY in rice (Zhang et al., 2009; Zou et al., 2011). LD was also lower in AlSAP-plants, and they were able to recover their vegetative biomass almost entirely after drought. Additionally, they showed significantly higher above-ground dry matter accumulation under both irrigated and drought conditions, indicating that increased biomass contributed to carbon assimilation and GY. This combination of traits may have led to higher carbon fixation rates, at either the leaf or the whole plant basis, in AlSAP-lines, particularly during the active grain filling phase, which occurred close to the peak of drought-stress.
Delayed LR indicates an ability to sustain turgor, despite drought-stress; for example, through increased water uptake or osmotic adjustment. We did not observe differences in Ψw that could explain the reduction of LR in AlSAP-lines, although Ψw was slightly higher in RN6 at stress peak. However, it is interesting that AlSAP-lines maintained Ψw values similar to those of WT-NIP, despite their elevated biomass accumulation, which would be expected to reduce this characteristic (Pantuwan et al., 2002). Neither was differential regulation of Gs observed in AlSAP-lines; hence, the reduced LR may be associated with improved access to water in the soil. The characterization of the response of AlSAP-seedlings to different abiotic stresses demonstrated their improved ability to maintain root growth and development (Ben Saad et al., 2012). Thus, it would be interesting to evaluate the development of the root system of these plants in the field.
It is important to highlight that AlSAP expression reduced LR but did not affect leaf morphology or plant development (data not shown; Supplementary Figure S2). A similar effect has been reported in rice plants overexpressing the Arabidopsis TFs CBF3/DREB1A (CBF3), ABF3, and SDIR1 (Oh et al., 2005; Zhang et al., 2009), or OsNAC TFs (Hu et al., 2006; Jeong et al., 2010). The molecular mechanisms underlying this effect remain unknown, but may include the interaction of some of these, or similar TFs, with other components in drought-stress signaling pathways, in which SAPs may also participate.
Developmental Stage and Environmental Conditions May Influence the Protective Effect of AlSAP on Photosynthesis
The imposition of stress at different phenological stages may influence plant responses to drought and the mechanisms by which AlSAP contributes to drought tolerance. Ben Saad et al. (2012) performed physiological investigations on the last expanded leaf of AlSAP rice plants at the 5–6 leaf, early tillering stage. In contrast, we evaluated photosynthesis parameters using the flag leaf, which is physiologically different to other leaves, and suffers distinctly under stress conditions. Hormone balance, oxidative damage, water regulation, and particularly drought-induced changes in photosynthesis, proceed distinctly in the flag leaf (Chaves et al., 2003; Biswal and Kohli, 2013). Drought-stress during anthesis reduces the rate of photosynthesis of the flag leaf and increases the remobilization of assimilates from it to sustain grain demand, accelerating its senescence, which has further impacts on photosynthesis (Peng and Ismail, 2004). In contrast, during vegetative growth and especially in young leaves, drought depresses the photosynthesis rate without inducing senescence (Peng and Ismail, 2004). The mechanisms involved in reducing photosynthesis are different from those that enhance remobilization, and do not necessarily involve AlSAP.
Evaluation at distinct physiological stages may also have involved developmentally associated differences in AlSAP transcription levels or expression patterns. We found that a higher accumulation of transcript correlates positively with drought-tolerance in AlSAP-lines. Similar observations were reported for rice, tobacco, and durum wheat (Ben Saad et al., 2010, 2011, 2012). These results may indicate that complete expression of the effects of AlSAP depends to some degree on gene transcript, or protein, accumulation. Alternatively, expression in certain organs/tissues or at specific growth stages may be required for AlSAP to achieve its full potential.
Co-occurrence of multiple abiotic stresses is another factor to consider. In the field, crops encounter a combination of stresses that could require different or antagonistic responses, and consequently may activate varying molecular mechanisms (Mittler, 2006; Mazzucotelli et al., 2008; Salekdeh et al., 2009). Drought and heat stresses are classic examples of this kind of interaction that may elicit antagonistic physiological responses and activate the transcription machinery in a non-additive fashion (Atkinson and Urwin, 2012). In our field trials, plants coped with both drought and high diurnal temperatures. Although, AlSAP appeared to confer tolerance to both heat and drought stresses applied independently to transformed tobacco (Ben Saad et al., 2010), its effect in response to the simultaneous imposition of these factors has not been tested and cannot be predicted from these individual studies (Salekdeh et al., 2009).
It is also possible that the improved yields observed in AlSAP-lines exposed to drought during the reproductive stage is not the result of better performance at the time of stress, but rather signifies a primed status, acquired during vegetative growth, due to the higher accumulation of carbon reserves resulting from enhanced photosynthetic performance at the vegetative stage.
AlSAP as a Candidate for Reduction of Yield Loss under Drought-Stress
Evidence gathered to date demonstrates that AlSAP overexpression in rice is effective in conferring drought tolerance at both the vegetative and reproductive stages. Stress protection during distinct phenological stages was also reported for AlSAP-transformed tobacco and durum wheat. Importantly, AlSAP expression exerted similar effects on growth and yield of durum wheat, tobacco, and rice, under both normal and stressed conditions (Ben Saad et al., 2010, 2011, 2012). These similar responses indicate that the beneficial role of AlSAP in regulating growth and development may be universal in monocotyledonous and dicotyledonous plants.
The gains in GY in AlSAP-transformed rice ranged from 26 to 115%, depending on the line and the severity of the stress exerted in the field. These gains are very high compared with reports for other transgenic rice lines expressing genes conferring drought tolerance and challenged with milder stress intensity, as indicated by the GY loss observed in controls under drought vs. standard conditions (see Table 1 for AlSAP-lines). For example, overexpression of AP37 in rice enhanced drought tolerance at the reproductive stage and resulted in yield gains of 16–57% above controls. However, in this study, the stress intensity was limited, since plants remained irrigated to avoid LR, resulting in a GY loss of only 32% in the WT line (Oh et al., 2009). In another study, rice plants overexpressing OsNAC10 showed enhanced drought tolerance at the reproductive stage, and GY increased by 25–42%; however, again milder stress conditions were applied (Jeong et al., 2010). The expression of HYR, which afforded a 14–39% GY increase under stress, was also evaluated under mild drought stress, since GY in the control line reduced by only 30% (Ambavaram et al., 2014).
A limited number of studies applied drought intensity more comparable to our experimental conditions. Plants overexpressing LOS5 and ZAT10 showed gains ranging from 11 to 36% compared with their controls, which suffered an 82% yield reduction (Xiao et al., 2009). Plants overexpressing AtEDT1/HDG11 exhibited a GY gain of 65% over the control, the yield of which was reduced by 83% when subjected to severe drought (Yu et al., 2013). More remarkably, rice lines expressing OsCPI1 showed a 2.5–3-fold superior GY over the control, the yield of which dropped by 90% under severe drought conditions (Huang et al., 2007).
Grain production penalties, due to panicle sterility and abnormalities, under unstressed conditions have frequently been reported when transgenic TFs are constitutively expressed (Dubouzet et al., 2003; Oh et al., 2009; Park et al., 2013). In our study, the use of three distinct promoters, CaMV35S, maize Ubi-1, and rice ACT1, inducing a range of constitutive gene expression levels, imposed no penalties in either rice growth or GY under irrigated conditions. Conversely, we observed modest, but consistently higher GY in AlSAP rice lines under paddy fields. These results contrast with the effects reported previously from several studies of SAP over-expression. To date, ectopic expression of five (OsSAP1, OsSAP8, OsSAP7, OsSAP9, and OsSAP11) of the 18 rice SAP genes has been investigated in heterologous or homologous systems. Two examples of SAPs (OsSAP1 and OsSAP8) over-expressed in rice under the maize ubiquitin promoter resulted in an enhanced tolerance to abiotic stresses, but detrimental effects on GY under standard growth conditions (Kanneganti and Gupta, 2008; Dansana et al., 2014). A third example (OsSAP7) demonstrated a negative effect of SAP overexpression in Arabidopsis abiotic stress tolerance, due to its role as negative regulator of ABA responses (Sharma et al., 2015).
The fact that these detrimental effects are absent in rice plants expressing the AlSAP gene underlines the potential for the application of AlSAP transgenic lines to reduce yield losses in crop species facing environmental stress.
Conclusion and Prospects
This is the first report of field evaluation of transgenic plants expressing an A20/AN1 stress-associated protein. We consistently observed that AlSAP accumulation enhanced GY under drought, and did not penalize either yield or plant growth under irrigated conditions. Improved GY was associated with higher biomass accumulation, a greater number of productive panicles, and lower panicle sterility, even under severe stress. Milder environments should be tested since it is possible that further GY gains can be attained by AlSAP expression under more favorable conditions. It will also be important to introduce AlSAP in other genetic backgrounds, notably elite upland cultivars with established basal levels of drought-tolerance, to determine whether AlSAP can further enhance their ability to resist drought.
Recent reports of transgenic protection of rice from drought-stress associated beneficial transgene effects with stronger root systems (Jeong et al., 2010, 2013; Redillas et al., 2012; Ambavaram et al., 2014). The influence of AlSAP on root development and hydraulic conductance require further assessment in specifically designed experiments. Also, a combination of transgenes with different modes of action, and that improve distinct traits contributing to yield under drought could be attempted.
We note apparent discrepancies in the effects of AlSAP in protection of photosynthesis observed in our greenhouse and field experiments. Several hypotheses to explain these findings have been presented and require further exploration. To provide additional clues regarding the physiological and molecular mechanisms operating in AlSAP crops, a better understanding of AlSAP gene function is required; in particular, investigations to identify proteins interacting with AlSAP in rice and decipher its regulation network will be beneficial. Such studies will provide opportunities to consolidate the protection afforded by AlSAP through stress-inducible/developmentally regulated expression strategies and combined expression with other genes (e.g., NAC genes) affording abiotic stress tolerance through different modes of action.
Author Contributions
TG-H and EG conceptualized and wrote the manuscript. TG-H, AP, SO, MR, and DM performed the experiments. MS, DF, WB, RB, MI, JT, AAD, and AH, revised the paper critically. All authors revised and approved the final manuscript.
Funding
This work was funded by the joint FIRST program of Agropolis Foundation, France, and the Cariplo Foundation, Italy, under the “Rice connections” project to EG, DM, DF, MI, MS, and JT, and supported by the National Program for Sciences, Technology and Innovation (NPSTI, Project N° 11-Bio1828-02 2012-2014) in the Kingdom of Saudi Arabia to EG, WB, AAD, and AH and the Visiting Professor program, King Saud University Saudi Arabia, to EG.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We appreciate the technical assistance kindly supplied by Milton Valencia, Maria Recio, Carlos Erasso, and Myriam C. Duque from CIAT, and by Luis Ramos, Stefania Belalcázar, and Alejandra Londoño from Icesi University.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.00994/full#supplementary-material
Footnotes
References
Ambavaram, M. M., Basu, S., Krishnan, A., Ramegowda, V., Batlang, U., Rahman, L., et al. (2014). Coordinated regulation of photosynthesis in rice increases yield and tolerance to environmental stress. Nat. Commun. 5:5302. doi: 10.1038/ncomms6302
Atkinson, N. J., and Urwin, P. E. (2012). The interaction of plant biotic and abiotic stresses: from genes to the field. J. Exp. Bot. 63, 3523–3543. doi: 10.1093/jxb/ers100
Atlin, G. (2003). “Improving drought tolerance by selecting for yield,” in Breeding Rice for Drought Prone Environments, eds K. S. Fischer, R. Lafitte, S. Fukai, G. Atlin, and B. Hardy (Los Baños: International Rice Research Institute), 14–23.
Ben Saad, R., Ben-Ramdhan, W., Zouari, N., Azaza, J., Mieulet, D., Guiderdoni, E., et al. (2011). Marker-free transgenic durum wheat cv. Karim expressing the AlSAP gene exhibits a high level of tolerance to salinity and dehydration stresses. Mol. Breed. 30, 521–533. doi: 10.1007/s11032-011-9641-3
Ben Saad, R., Fabre, D., Mieulet, D., Meynard, D., Dingkuhn, M., Al-Doss, A., et al. (2012). Expression of the Aeluropus littoralis AlSAP gene in rice confers broad tolerance to abiotic stresses through maintenance of photosynthesis. Plant Cell Environ. 35, 626–643. doi: 10.1111/j.1365-3040.2011.02441.x
Ben Saad, R., Zouari, N., Ben Ramdhan, W., Azaza, J., Meynard, D., Guiderdoni, E., et al. (2010). Improved drought and salt stress tolerance in transgenic tobacco overexpressing a novel A20/AN1 zinc-finger “AlSAP” gene isolated from the halophyte grass Aeluropus littoralis. Plant Mol. Biol. 72, 171–190. doi: 10.1007/s11103-009-9560-4
Bernier, J., Serraj, R., Kumar, A., Venuprasad, R., Impa, S., Gowda, V. R. P., et al. (2009). The large-effect drought resistance QTL qtl12.1 increases water uptake in upland rice. Field Crop. Res. 110, 139–146. doi: 10.1016/j.fcr.2008.07.010
Biswal, A. K., and Kohli, A. (2013). Cereal flag leaf adaptations for grain yield under drought: knowledge status and gaps. Mol. Breed. 31, 749–766. doi: 10.1007/s11032-013-9847-7
Bittelli, M., and Flury, M. (2009). Errors in water retention curves determined with pressure plates. Soil Sci. Soc. Am. J. 73, 1453–1460. doi: 10.2136/sssaj2008.0082
Caldana, C., Scheible, W. R., Mueller-Roeber, B., and Ruzicic, S. (2007). A quantitative RT-PCR platform for high-throughput expression profiling of 2500 rice transcription factors. Plant Methods 3:7. doi: 10.1186/1746-4811-3-7
Centritto, M., Lauteri, M., Monteverdi, M. C., and Serraj, R. (2009). Leaf gas exchange, carbon isotope discrimination, and grain yield in contrasting rice genotypes subjected to water deficits during the reproductive stage. J. Exp. Bot. 60, 2325–2339. doi: 10.1093/Jxb/Erp123
Chaves, M. M., Maroco, J., and Pereira, J. S. (2003). Understanding plant responses to drought; from genes to the whole plant. Funct. Plant Biol. 30, 239–264. doi: 10.1071/FP02076
Choi, H., Han, S., Shin, D., and Lee, S. (2012). Polyubiquitin recognition by AtSAP5, an A20-type zinc finger containing protein from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 419, 436–440. doi: 10.1016/j.bbrc.2012.02.044
Christensen, A. H., Sharrock, R. A., and Quail, P. H. (1992). Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing, and promoter activity following transfer to protoplasts by electroporation. Plant Mol. Biol. 18, 675–689. doi: 10.1007/BF00020010
Cornejo, M.-J., Luth, D., Blankenship, K., Anderson, O., and Blechl, A. (1993). Activity of a maize ubiquitin promoter in transgenic rice. Plant Mol. Biol. 23, 567–581. doi: 10.1007/BF00019304
Courtois, B., Ahmadi, N., Khowaja, F., Price, A. H., Rami, J.-F., Frouin, J., et al. (2009). Rice root genetic architecture: meta-analysis from a drought QTL database. Rice 2, 115–128. doi: 10.1007/s12284-009-9028-9
Dansana, P. K., Kothari, K. S., Vij, S., and Tyagi, A. K. (2014). OsiSAP1 overexpression improves water-deficit stress tolerance in transgenic rice by affecting expression of endogenous stress-related genes. Plant Cell Rep. 33, 1425–1440. doi: 10.1007/s00299-014-1626-3
Du, Y. M., Tian, J., Liao, H., Bai, C. J., Yan, X. L., and Liu, G. D. (2009). Aluminium tolerance and high phosphorus efficiency helps Stylosanthes better adapt to low-P acid soils. Ann. Bot. 103, 1239–1247. doi: 10.1093/aob/mcp074
Dubouzet, J. G., Sakuma, Y., Ito, Y., Kasuga, M., Dubouzet, E. G., Miura, S., et al. (2003). OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. Plant J. 33, 751–763. doi: 10.1046/j.1365-313X.2003.01661.x
Gaudin, A. C., Henry, A., Sparks, A. H., and Slamet-Loedin, I. H. (2013). Taking transgenic rice drought screening to the field. J. Exp. Bot. 64, 109–117. doi: 10.1093/jxb/ers313
Giri, J., Dansana, P. K., Kothari, K. S., Sharma, G., Vij, S., and Tyagi, A. K. (2013). SAPs as novel regulators of abiotic stress response in plants. Bioessays 35, 639–648. doi: 10.1002/bies.201200181
Giri, J., Vij, S., Dansana, P. K., and Tyagi, A. K. (2011). Rice A20/AN1 zinc-finger containing stress-associated proteins (SAP1/11) and a receptor-like cytoplasmic kinase (OsRLCK253) interact via A20 zinc-finger and confer abiotic stress tolerance in transgenic Arabidopsis plants. New Phytol. 191, 721–732. doi: 10.1111/j.1469-8137.2011.03740.x
Gu, J., Yin, X., Stomph, T.-J., Wang, H., and Struik, P. C. (2012). Physiological basis of genetic variation in leaf photosynthesis among rice (Oryza sativa L.) introgression lines under drought and well-watered conditions. J. Exp. Bot. 63, 5137–5153. doi: 10.1093/jxb/ers170
Hanzawa, E., Sasaki, K., Nagai, S., Obara, M., Fukuta, Y., Uga, Y., et al. (2013). Isolation of a novel mutant gene for soil-surface rooting in rice (Oryza sativa L.). Rice 6:30. doi: 10.1186/1939-8433-6-30
Hozain, M., Abdelmageed, H., Lee, J., Kang, M., Fokar, M., Allen, R. D., et al. (2012). Expression of AtSAP5 in cotton up-regulates putative stress-responsive genes and improves the tolerance to rapidly developing water deficit and moderate heat stress. J. Plant Physiol. 169, 1261–1270. doi: 10.1016/j.jplph.2012.04.007
Hu, H., Dai, M., Yao, J., Xiao, B., Li, X., Zhang, Q., et al. (2006). Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc. Natl. Acad. Sci. U.S.A. 103, 12987–12992. doi: 10.1073/pnas.0604882103
Huang, Y., Xiao, B., and Xiong, L. (2007). Characterization of a stress responsive proteinase inhibitor gene with positive effect in improving drought resistance in rice. Planta 226, 73–85. doi: 10.1007/s00425-006-0469-8
Jeong, J. S., Kim, Y. S., Baek, K. H., Jung, H., Ha, S. H., Do Choi, Y., et al. (2010). Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 153, 185–197. doi: 10.1104/pp.110.154773
Jeong, J. S., Kim, Y. S., Choi, Y. D., and Kim, J. K. (2013). OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield. FEBS J. 280, 516–517. doi: 10.1111/pbi.12011
Jongdee, B., Fukai, S., and Cooper, M. (2002). Leaf water potential and osmotic adjustment as physiological traits to improve drought tolerance in rice. Field Crop. Res. 76, 153–163. doi: 10.1016/S0378-4290(02)00036-9
Kang, M., Abdelmageed, H., Lee, S., Reichert, A., Mysore, K. S., and Allen, R. D. (2013). AtMBP-1, an alternative translation product of LOS2, affects abscisic acid responses and is modulated by the E3 ubiquitin ligase AtSAP5. Plant J. 76, 481–493. doi: 10.1111/tpj.12312
Kang, M., Fokar, M., Abdelmageed, H., and Allen, R. D. (2011). Arabidopsis SAP5 functions as a positive regulator of stress responses and exhibits E3 ubiquitin ligase activity. Plant Mol. Biol. 75, 451–466. doi: 10.1007/s11103-011-9748-2
Kanneganti, V., and Gupta, A. K. (2008). Overexpression of OsiSAP8, a member of stress associated protein (SAP) gene family of rice confers tolerance to salt, drought and cold stress in transgenic tobacco and rice. Plant Mol. Biol. 66, 445–462. doi: 10.1007/s11103-007-9284-2
Khowaja, F. S., Norton, G. J., Courtois, B., and Price, A. H. (2009). Improved resolution in the position of drought-related QTLs in a single mapping population of rice by meta-analysis. BMC Genomics 10:276. doi: 10.1186/1471-2164-10-276
Kumar, A., Bernier, J., Verulkar, S., Lafitte, H. R., and Atlin, G. N. (2008). Breeding for drought tolerance: direct selection for yield, response to selection and use of drought-tolerant donors in upland and lowland adapted populations. Field Crop. Res. 107, 221–231. doi: 10.1016/j.fcr.2008.02.007
Lanceras, J. C., Pantuwan, G., Jongdee, B., and Toojinda, T. (2004). Quantitative trait loci associated with drought tolerance at reproductive stage in rice. Plant Phys. Biochem. 135, 384–399. doi: 10.1104/pp.103.035527
Li, J.-Y., Wang, J., and Zeigler, R. S. (2014). The 3,000 rice genomes project: new opportunities and challenges for future rice research. Gigascience 3:8. doi: 10.1186/2047-217X-3-8
Liu, Q.-L., Xu, K.-D., Zhong, M., Pan, Y.-Z., Jiang, B.-B., Liu, G.-L., et al. (2013). Overexpression of a novel chrysanthemum Cys2/His2-type zinc finger protein gene DgZFP3 confers drought tolerance in tobacco. Biotechnol. Lett. 35, 1953–1959. doi: 10.1007/s10529-013-1289-0
Liu, Y., Xu, Y., Xiao, J., Ma, Q., Li, D., Xue, Z., et al. (2011). OsDOG, a gibberellin-induced A20/AN1 zinc-finger protein, negatively regulates gibberellin-mediated cell elongation in rice. J. Plant Physiol. 168, 1098–1105. doi: 10.1016/j.jplph.2010.12.013
Mazzucotelli, E., Mastrangelo, A. M., Crosatti, C., Guerra, D., Stanca, A. M., and Cattivelli, L. (2008). Abiotic stress response in plants: when post-transcriptional and post-translational regulations control transcription. Plant Sci. 174, 420–431. doi: 10.1016/j.plantsci.2008.02.005
Mc Elroy, D., Zhang, W., Cao, J., and Wu, R. (1990). lsolation of an efficient actin promoter for use in rice transformation. Plant Cell 2, 163–171. doi: 10.1105/tpc.2.2.163
Mittler, R. (2006). Abiotic stress, the field environment and stress combination. Trends Plant Sci. 11, 15–19. doi: 10.1016/j.tplants.2005.11.002
Miyao, A., Nakagome, M., Ohnuma, T., Yamagata, H., Kanamori, H., Katayose, Y., et al. (2012). Molecular spectrum of somaclonal variation in regenerated rice revealed by whole-genome sequencing. Plant Cell Physiol. 53, 256–264. doi: 10.1093/pcp/pcr172
Oh, S. J., Kim, Y. S., Kwon, C. W., Park, H. K., Jeong, J. S., and Kim, J. K. (2009). Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. Biochem. 150, 1368–1379. doi: 10.1104/pp.109.137554
Oh, S. J., Song, S. I., Kim, Y. S., Jang, H. J., Kim, S. Y., Kim, M., et al. (2005). Arabidopsis CBF3/DREB1A and ABF3 in transgenic rice increased tolerance to abiotic stress without stunting growth. Plant Physiol. 138, 341–351. doi: 10.1104/pp.104.059147
Pandey, S., and Bhandari, H. (2007). “Drought: an overview,” in Economic Costs of Drought and Rice Farmers’ Coping Mechanisms: A Cross-Country Comparative Analysis, eds S. Pandey, H. Bhandari, and B. Hardy (Los Baños: International Rice Research Institute), 203.
Pantuwan, G., Fukai, S., Cooper, M., Rajatasereekul, S., and O’Toole, J. C. (2002). Yield response of rice (Oryza sativa L.) genotypes to drought under rainfed lowland: 3. Plant factors contributing to drought resistance. Field Crop Res. 73, 181–200. doi: 10.1016/S0378-4290(01)00194-0
Park, S.-H., Jeong, J. S., Redillas, M. C. F. R., Jung, H., Bang, S. W., Kim, Y. S., et al. (2013). Transgenic overexpression of UIP1, an interactor of the 3’ untranslated region of the Rubisco small subunit mRNA, increases rice tolerance to drought. Plant Biotechnol. Rep. 7, 83–90. doi: 10.1007/s11816-012-0239-y
Peng, S., and Ismail, A. M. (2004). “Physiological basis of yield and environmental adaptation in rice,” in Physiology and Biotechnology Integration for Plant Breeding, eds A. Blum and H. T. Nguyen (New York, NY: Marcel Dekker, INC), 72–118.
Prasad, P. V. V., Staggenborg, S. A., and Ristic, Z. (2008). “Impacts of drought and/or heat stress on physiological, developmental, growth, and yield processes of crop plants,” in Response of Crops to Limited Water: Understanding and Modeling Water Stress Effects on Plant Growth Processes, eds L. R. Ahuja, V. R. Reddy, S. A. Saseendran, and Q. Yu (Madison, WI: American Society of Agronomy), 301–355.
Raorane, M. L., Pabuayon, I. M., Varadarajan, A. R., Mutte, S. K., Kumar, A., Treumann, A., et al. (2015). Proteomic insights into the role of the large-effect QTL qDTY12.1 for rice yield under drought. Mol. Breed. 35:139. doi: 10.1007/s11032-015-0321-6
Redillas, M. C. F. R., Jeong, J. S., Kim, Y. S., Jung, H., Bang, S. W., Choi, Y. D., et al. (2012). The overexpression of OsNAC9 alters the root architecture of rice plants enhancing drought resistance and grain yield under field conditions. Plant Biotechnol. J. 10, 792–805. doi: 10.1111/j.1467-7652.2012.00697.x
Salekdeh, G. H., Reynolds, M., Bennett, J., and Boyer, J. (2009). Conceptual framework for drought phenotyping during molecular breeding. Trends Plant. Sci. 14, 488–496. doi: 10.1016/j.tplants.2009.07.007
Sharma, G., Giri, J., and Tyagi, A. K. (2015). Rice OsiSAP7 negatively regulates ABA stress signalling and imparts sensitivity to water-deficit stress in Arabidopsis. Plant Sci. 237, 80–92. doi: 10.1016/j.plantsci.2015.05.011
Sibounheuang, V., Basnayake, J., and Fukai, S. (2006). Genotypic consistency in the expression of leaf water potential in rice (Oryza sativa L.). Field Crop. Res. 97, 142–154. doi: 10.1016/j.fcr.2005.09.006
Sreedharan, S., Shekhawat, U. K., and Ganapathi, T. R. (2012). MusaSAP1, a A20/AN1 zinc finger gene from banana functions as a positive regulator in different stress responses. Plant Mol. Biol. 80, 503–517. doi: 10.1007/s11103-012-9964-4
Stroher, E., Wang, X. J., Roloff, N., Klein, P., Husemann, A., and Dietz, K. J. (2009). Redox-dependent regulation of the stress-induced zinc-finger protein SAP12 in Arabidopsis thaliana. Mol. Plant 2, 357–367. doi: 10.1093/mp/ssn084
Stroud, H., Ding, B., Simon, S. A., Feng, S., Bellizzi, M., Pellegrini, M., et al. (2013). Plants regenerated from tissue culture contain stable epigenome changes in rice. Elife 19:e00354. doi: 10.7554/eLife.00354
Tirzitis, G., and Bartosz, G. (2010). Determination of antiradical and antioxidant activity: basic principles and new insights. Acta Biochim. Pol. 57, 139–142.
Tromp-van Meerveld, H. J., and McDonnell, J. J. (2006). On the interrelations between topography, soil depth, soil moisture, transpiration rates and species distribution at the hillslope scale. Adv. Water Resources 29, 293–310. doi: 10.1016/j.advwatres.2005.02.016
Turner, N. C. (1981). Techniques and experimental approaches for the measurement of plant water status. Plant Soil 58, 339–366. doi: 10.1007/BF02180062
Ueda, Y., Uehara, N., Sasaki, H., Kobayashi, K., and Yamakawa, T. (2013). Impacts of acute ozone stress on superoxide dismutase (SOD) expression and reactive oxygen species (ROS) formation in rice leaves. Plant Physiol. Biochem. 70, 396–402. doi: 10.1016/j.plaphy.2013.06.009
Uga, Y., Sugimoto, K., Ogawa, S., Rane, J., Ishitani, M., Hara, N., et al. (2013). Control of root system architecture by DEEPER ROOTING 1 increases rice yield under drought conditions. Nat. Genet. 45, 1097–1102. doi: 10.1038/ng.2725
Vij, S., and Tyagi, A. K. (2008). A20/AN1 zinc-finger domain-containing proteins in plants and animals represent common elements in stress response. Funct. Integr. Genomics 8, 301–307. doi: 10.1007/s10142-008-0078-7
Wang, X., Vignjevic, M., Jiang, D., Jacobsen, S., and Wollenweber, B. (2014). Improved tolerance to drought stress after anthesis due to priming before anthesis in wheat (Triticum aestivum L.) var. Vinjett. J. Exp. Bot. 65, 6441–6456. doi: 10.1093/jxb/eru362
Xiao, B., Huang, Y., Tang, N., and Xiong, L. (2007). Over-expression of a LEA gene in rice improves drought resistance under the field conditions. Theor. Appl. Genet. 115, 35–46. doi: 10.1007/s00122-007-0538-9
Xiao, B. Z., Chen, X., Xiang, C. B., Tang, N., Zhang, Q. F., and Xiong, L. Z. (2009). Evaluation of seven function-known candidate genes for their effects on improving drought resistance of transgenic rice under field conditions. Mol. Plant 2, 73–83. doi: 10.1093/mp/ssn068
You, J., Hu, H., and Xiong, L. (2012). Ornithine δ-aminotransferase gene OsOAT confers drought and oxidative stress tolerance in rice. Plant Sci. 197, 59–69. doi: 10.1016/j.plantsci.2012.09.002
Yu, L., Chen, X., Wang, Z., Wang, S., Wang, Y., Zhu, Q., et al. (2013). Arabidopsis enhanced drought tolerance1/HOMEODOMAIN GLABROUS11 confers drought tolerance in transgenic rice without yield penalty. Plant Physiol. 162, 1378–1391. doi: 10.1104/pp.113.217596
Zhang, G. H., Xu, Q., Zhu, X. D., Qian, Q., and Xue, H. W. (2009). SHALLOT-LIKE1 is a KANADI transcription factor that modulates rice leaf rolling by regulating leaf abaxial cell development. Plant Cell 21, 719–735. doi: 10.1105/tpc.108.061457
Zou, L. P., Sun, X. H., Zhang, Z. G., Liu, P., Wu, J. X., Tian, C. J., et al. (2011). Leaf rolling controlled by the homeodomain leucine zipper class IV gene Roc5 in rice. Plant Physiol. 156, 1589–1602. doi: 10.1104/pp.111.176016
Keywords: AlSAP, A20/AN1 stress-associated protein, drought, reproductive stage, rice, yield
Citation: Ghneim-Herrera T, Selvaraj MG, Meynard D, Fabre D, Peña A, Ben Romdhane W, Ben Saad R, Ogawa S, Rebolledo MC, Ishitani M, Tohme J, Al-Doss A, Guiderdoni E and Hassairi A (2017) Expression of the Aeluropus littoralis AlSAP Gene Enhances Rice Yield under Field Drought at the Reproductive Stage. Front. Plant Sci. 8:994. doi: 10.3389/fpls.2017.00994
Received: 17 January 2017; Accepted: 26 May 2017;
Published: 12 June 2017.
Edited by:
Henry T. Nguyen, University of Missouri, United StatesReviewed by:
Dong-Keun Lee, Seoul National University, South KoreaRohit Joshi, Jawaharlal Nehru University, India
Copyright © 2017 Ghneim-Herrera, Selvaraj, Meynard, Fabre, Peña, Ben Romdhane, Ben Saad, Ogawa, Rebolledo, Ishitani, Tohme, Al-Doss, Guiderdoni and Hassairi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thaura Ghneim-Herrera, dGdobmVpbUBpY2VzaS5lZHUuY28= Michael G. Selvaraj, bS5zZWx2YXJhakBjZ2lhci5vcmc= Emmanuel S. Guiderdoni, Z3VpZGVyZG9uaUBjaXJhZC5mcg==