 Yan Ren1
Yan Ren1 Weixiu Hou1
Weixiu Hou1 Caixia Lan2
Caixia Lan2 Bhoja R. Basnet2
Bhoja R. Basnet2 Ravi P. Singh2
Ravi P. Singh2 Wei Zhu3Xiyong Cheng1
Wei Zhu3Xiyong Cheng1 Dangqun Cui1
Dangqun Cui1 Feng Chen1*
Feng Chen1*- 1Collaborative Innovation Center of Henan Grain Crops/National Key Laboratory of Wheat and Maize Crop Science/Agronomy College, Henan Agricultural University, Zhengzhou, China
- 2International Maize and Wheat Improvement Center (CIMMYT), Mexico, Mexico
- 3Shangqiu Academy of Agricultural and Forestry Sciences, Shangqiu, China
CIMMYT wheat (Triticum aestivum L.) lines Francolin#1 and Quaiu#3 displayed effective and stable adult plant resistance (APR) to Chinese Blumeria graminis f. sp. tritici isolates in the field. To elucidate their genetic basis of resistance, two recombinant inbred line (RIL) populations of their crosses with Avocet, the susceptible parent, were phenotyped in Zhengzhou and Shangqiu in the 2014–2015 and 2015–2016 cropping seasons. These populations were also genotyped with SSR (simple sequence repeat markers) and DArT (diversity arrays technology) markers. Two common significant quantitative trait loci (QTL) on wheat chromosomes 1BL and 4BL were detected in both populations by joint and individual inclusive composite interval mapping, explaining 20.3–28.7% and 9.6–15.9% of the phenotypic variance in Avocet × Francolin#1 and 4.8–11.5% and 10.8–18.9% in Avocet × Quaiu#3, respectively. Additional QTL were mapped on chromosomes 1DL and 5BL in Avocet × Francolin#1 and on 2DL and 6BS in Avocet × Quaiu#3. Among these, QPm.heau-1DL is probably a novel APR gene contributing 6.1–8.5% of total phenotypic variance. The QTL on 1BL corresponds to the pleiotropic multi-pathogen resistance gene Yr29/Lr46/Pm39, whereas the QTL on 2DL maps to a similar region where stripe rust resistance gene Yr54 is located. The QTL identified can potentially be used for the improvement of powdery mildew and rust resistance in wheat breeding.
Introduction
Powdery mildew (PM), caused by Blumeria graminis f. sp. tritici (Bgt), is a significant disease of common wheat worldwide, reported to have caused yield losses of up to 34% (Alam et al., 2011). Since the 1980s, PM has been widespread across large areas of China, and is currently a major disease in the Northern, Yellow and Huai wheat growing regions. On average, 7.0 million ha of wheat planting areas were affected annually by PM in China from 2010 to 2015 (National Agro-technical Extension and Service Center (NAESC), 2010, 2011, 2012, 2013, 2014, 2015). Growing resistant cultivars is the most cost effective and environmentally safe approach to manage the disease (Gurung et al., 2009).
Resistance to PM in wheat is of two main types, viz. race specific and race non-specific. The former is usually effective against certain Bgt isolates, but ineffective to others. These types of genes usually exhibit resistance through the whole growth cycle of wheat and cause various degrees of hypersensitive foliar reactions in a gene-for-gene interaction (Flor, 1942; Hsam and Zeller, 2002). Race-specific resistance genes were widely applied in Chinese wheat breeding because of their high level of effectiveness through all growth stages of the crop. However, these genes are known to break down readily due to the selection of pathogen isolates with matching virulence genes. In contrast, race non-specific resistance is often quantitatively inherited. It usually shows resistance at the adult plant stage, hence, it is also called adult plant resistance (APR), and is more durable than the race specific resistance. So far, 82 PM resistance genes have been formally cataloged on 54 loci (McIntosh et al., 2017), but most of them are race specific and are easily overcome by new Bgt isolates (Leath and Murphy, 1985; Limpert et al., 1987; Li et al., 2014). Consequently, breeders have paid more attention to the development of wheat cultivars with APR to PM. So far, only three wheat PM resistance genes, Pm38, Pm39, and Pm46, are known to be race non-specific and confer partial resistance. These three genes also show pleiotropic effects on resistance to stripe rust, leaf rust, and stem rust (William et al., 2003; Spielmeyer et al., 2005; Herrera-Foessel et al., 2011). Moreover, Pm38 and Pm46 have been cloned (Krattinger et al., 2009; Moore et al., 2015). The multi-pathogen resistances conferred by Pm38 and Pm46 encode a putative ABC transporter and a hexose transporter (Krattinger et al., 2009; Risk et al., 2012; Moore et al., 2015), respectively. These pleiotropic APR genes are valuable in wheat breeding for their broad effectiveness and potential durability.
In addition, over 120 PM resistance quantitative trait loci (QTL) have been identified using molecular markers in wheat (Li et al., 2014; Hao et al., 2015). However, some of these QTL might be the same due to their chromosomal location at approximate vicinity. Marone et al. (2013) developed a high-density integrated map for the projection of PM resistance genes, and identified 24 MQTLs containing 2–6 initial QTLs on 15 chromosomes through a QTL meta-analysis. The chromosomes 2B and 7A showed abundant PM resistance loci. Li et al. (2014) constructed an integrated linkage map using mapped QTL with PM and leaf rust resistance based on wheat consensus maps, and found eight resistance QTL clusters on chromosomes 1A (2 QTL clusters), 2A (2 QTL clusters), 3A, 4A, 5A, and 5B, respectively. Overall the A genome contained more PM resistance QTL than the B and D genomes.
Francolin#1 and Quaiu#3 are high yielding spring wheat lines developed by the International Maize and Wheat Improvement Center (CIMMYT), and have been widely used as parents in the CIMMYT wheat breeding program. They do not carry effective seedling resistance genes to PM as they displayed high infection type (IT) against Chinese Bgt isolates E09 and E20 at seedling stage (IT = 3 based on the 0–4 scale). However, the two lines exhibited stable resistance to those isolates at the adult plant stage, indicating the presence of typical APR to PM. QTL analysis for APR to stripe rust and leaf rust in Francolin#1 and Quaiu#3 were previously analyzed by Lan et al. (2014) and Basnet et al. (2014), respectively. However, the genetic basis of resistance to PM in the two parents remains unknown. The objectives of our study were to: (1) investigate the genetic basis of PM resistance in Avocet × Francolin#1 and Avocet × Quaiu#3, and (2) identify and characterize the major APR loci for PM resistance in these populations through joint and individual inclusive composite interval mapping approach.
Materials and Methods
Plant Materials
In the present study, 196 and 195 F5 recombinant inbred lines (RILs) from Avocet-YrA × Francolin#1 and Avocet-YrA × Quaiu#3 crosses, respectively, were used to understand the genetic basis of resistance to powdery mildew. Francolin#1 (Waxwing*2/Vivitsi) and Quaiu#3 (Babax/Lr42//Babax*2/3/Vivitsi) displayed moderate levels of stable resistance to powdery mildew at the adult plant stage. Whereas, Avocet-YrA (hereafter termed Avocet for simplicity), an Australian reselected line that lacks the resistance gene YrA, was susceptible to currently prevalent Chinese Bgt isolates E09 and E20 at all growth stages.
Greenhouse Test
Seedling tests of Francolin#1, Quaiu#3, Avocet and all RILs were conducted in greenhouse using the prevalent Chinese isolates E09 and E20 in 2014 and 2016. About 15 seeds of each entry were sown in 10 × 10 × 10 cm plastic pots where three seeds of susceptible cultivar Jingshuang 16 were also included as controls. Seedlings were uniformly dust inoculated with fresh conidiophores on two-leaf stage. Infection types (IT) were scored 7–10 d after inoculation based on the improved 0–4 scale standard following Wang et al. (2005).
Field Trials
Two RIL populations were hand-sown (~70 plants per line) in a 1.5 m long single row at Zhengzhou and Shangqiu in Henan province, China during the 2014–2015 and 2015–2016 cropping seasons. Every 12th row was planted with the susceptible variety Jingshuang 16 as controls and spreader rows. Inoculations on spreader rows were carried out using two prevalent Chinese isolates E09 and E20 at the early jointing stage at Zhengzhou and Shangqiu. Disease severity was evaluated at the filling stage when the susceptible check (Jingshuang 16) reached 80% severity and the second scoring was done 4–5 days after the first evaluation. Maximum disease severity (MDS) on RILs was used in all statistical analyses and QTL mapping. The field data from the 2014–2015 cropping season at Shangqiu was not used due to poor PM development.
Genetic and Statistical Analyses
To preliminarily estimate the number of PM resistance genes in two RIL populations using Mendelian segregation analysis, the RILs in each population were classified into two phenotypic categories following Lillemo et al. (2006); resistant (R), and susceptible- intermediate (SI). A line was considered R when it had a similar disease score as the resistant parent ± one standard deviation; otherwise, the line was classified into SI. The goodness-of-fit to the expected segregation ratios for two, three, four and five independent genes were tested using Chi squared (χ2) analysis. Moreover, the minimum number of PM resistance genes was also determined using the Wright's method (1968): n = D2/4.27, where D = the phenotype range of the F5lines multiplied by the narrow sense heritability. Narrow sense heritability of PM was calculated using the following formula: h2 = /( + ), where = ( –)/r; in this formula, genetic variance, variance of the F5 lines, error variance, and r = number of replicates.
Map Construction and QTL Mapping
In total, 520 and 450 simple sequence repeat (SSR) markers were screened on the parents of Avocet × Francolin#1 and Avocet × Quaiu#3, respectively. Polymorphic SSR markers were then used to genotype the entire populations. In addition, 141 lines of Avocet × Francolin#1 and 181 lines of Avocet × Quaiu#3 were genotyped with 518 and 361 diversity array technology (DArT) markers, respectively.
Genetic linkage groups were constructed with the statistical software QTL IciMapping V4.1 (http://www.isbreeding.net/). A logarithm of odds (LOD) score of 5.0 was set to make the linkage groups. To finalize the linkage map, marker ordering and ripping were performed using RECORD and COUNT algorithms, respectively. Inclusive composite interval mapping (ICIM) was used to detect the QTL based on MDS in each environment and the means of MDS (MMDS) over all environments. LOD scores for declaring significant QTL were calculated from 1,000 permutations at the P = 0.05 level. A walk speed of 1.0 cM was chosen for all QTL detection. After ICIM analysis, a consensus map was constructed with QTL IciMapping V4.1 using the “CMP” tool. “nnTwoOpt” and “SAD” were used to measure linkage algorithm and criteria. Subsequently, a joint inclusive composite interval-mapping (JICIM) program (Li et al., 2011) with a stepwise regression probability of 0.001 was employed for QTL detection. The LOD threshold was calculated by 1,000 permutations at the P = 0.05 level. In both cases, QTL effects were estimated as the phenotypic variance explained (PVE) and additive effects explained by the QTL.
Results
PM Evaluation in the Field
Francolin#1 and Quaiu#3 were susceptible to Chinese Bgt isolates E09 and E20 at the seedling stage, but showed resistance at the adult plant stage. The RILs from both populations were also susceptible in seedling stage. Final MDS of Francolin#1 and Quaiu#3 were 9.8–22.7 and 26.8–31.7% across three environments, respectively, whereas it ranged from 48.3 to 78.3% for the susceptible parent Avocet over three environments. The frequency distributions of PM severity for RILs exhibited continuous variation in both populations and transgressive segregations for both increased resistance and susceptibility (Figure S1), indicating polygenic inheritance of APR to PM in both parents. Genetic analyses by Mendelian segregation analysis (Table S2) and Wright's method (Table S3) indicated the presence of 3–4 and 2–3 APR genes conferring resistance to powdery mildew in Avocet × Francolin#1 and Avocet × Quaiu#3 populations, respectively.
Pearson's correlation coefficients (r) for PM severities for Avocet × Francolin#1 RILs was between 0.70 and 0.73 across three environments. Highly significant correlation (r = 0.63–0.85) for MDS were also observed in the Avocet × Quaiu#3 population across environments. ANOVA for both populations also showed significant variance (P < 0.0001) among lines, environments and line × environment interactions (Table S1).
Linkage Maps Construction
A total of 647 polymorphic SSR and DArT markers were used to construct the genetic linkage map in Avocet × Francolin#1 population, spanning 779 cM, 1,442 and 450 cM in the sub-genomes A, B, and D, respectively (Table S4). Twenty-four linkage groups were developed, covering all chromosomes of wheat. Meanwhile, 463 polymorphic SSR and DArT markers were used for construction of genetic linkage map in Avocet × Quaiu#3 population, covering a total of 2628.2 cM in all linkage groups (Table S4). Twenty-six linkage groups were developed, representing all chromosomes.
QTL Analysis in Avocet × Francolin#1 Population
Four QTL for PM resistance were detected in the Avocet × Francolin#1 population (Table 1, Figure 1). The resistance genes mapped on chromosomes 1BL, 1DL, and 5BL were derived from wheat line Francolin#1, whereas the one on chromosome 4BL was from Avocet. The large-effect QTL QPm.heau-1BL, flanked by molecular markers wPt-1770 and wPt-9028, was detected in all three environments. It explained 26.1, 20.3, 21.2, and 28.7% of phenotypic variance in Zhengzhou 2014, Zhengzhou 2015, Shangqiu 2015, and MMDS, respectively (Table 1, Figure 1A). The second consistently detected QTL with larger effect, QPm.heau-4BL, was located on chromosome 4BL between wmc413 and wPt-9067, explaining 9.6, 10.7, 15.1, and 15.9% of phenotypic variance in the three environments and MMDS, respectively (Table 1, Figure 1C). The third QTL QPm.heau-5BL, located between DArT markers wPt-2607 and wPt-1895 on 5BL, explained 10.9, 6.2, and 7.0% of the phenotypic variance in two environments and MMDS, respectively (Table 1, Figure 1D). The fourth QTL, QPm.heau-1DL, mapped in the marker interval of wPt-5721-wPt-1865, was also detected in all three environments and for MMDS, explaining 6.8, 6.1, 8.5, and 8.3% of the phenotypic variance, respectively (Table 1, Figure 1B).
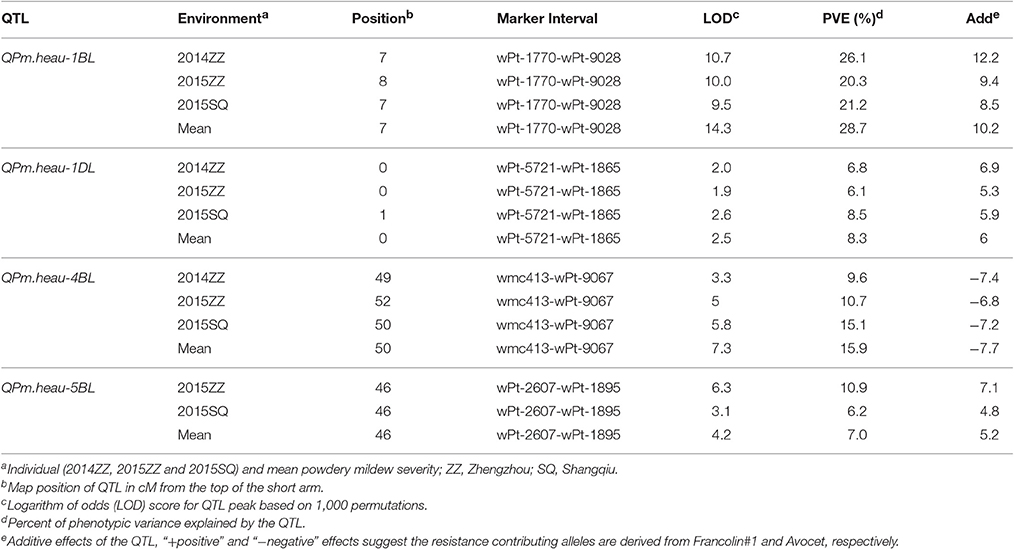
Table 1. Quantitative trait loci (QTL) for adult-plant resistance to powdery mildew detected in Avocet × Francolin#1 RIL population through bi-parental analysis.
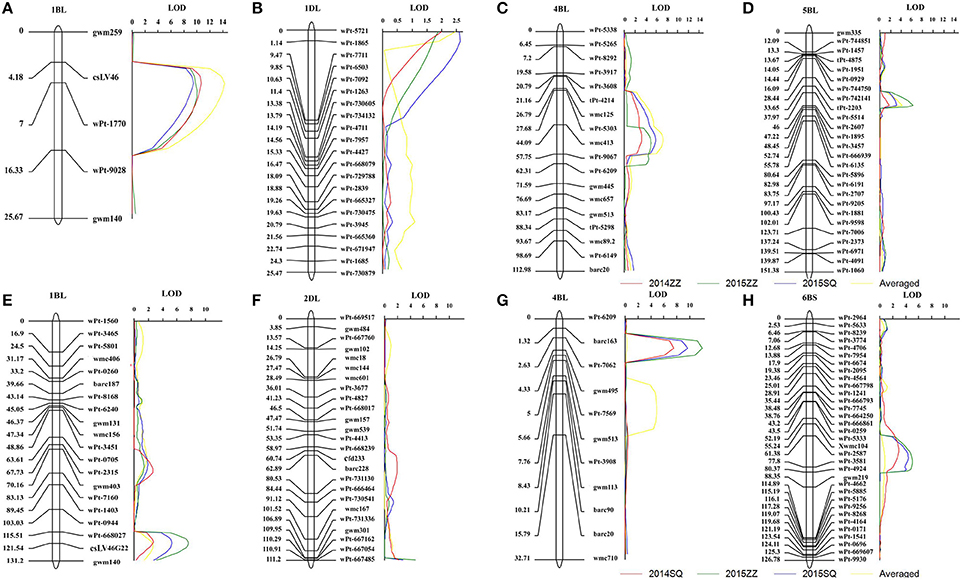
Figure 1. LOD contours for quantitative trait loci (QTL) to powdery mildew on chromosomes 1BL, 1DL, 4BL, and 5BL identified by inclusive composite interval mapping (ICIM) in the Avocet × Francolin#1 population (A–D), and on chromosomes 1BL, 2DL, 4BL, and 6BS in the Avocet × Quaiu#3 population (E–H). The significant LOD thresholds were based on 1,000 permutations. Positions (in cM) of molecular markers along chromosomes are shown on the vertical axes using cumulated genetic distances.
QTL Analysis in Avocet × Quaiu#3 Population
Four QTL for APR to PM were found in the Avocet × Quaiu#3 population (Table 2, Figure 1). The resistance genes on 1BL and 4BL were mapped at proximately chromosomal positions to those identified in the Avocet × Francolin#1 population. The largest effect resistance QTL was located on chromosome 4BL, flanked by SSR markers wPt-7062 and gwm495, explaining from 10.8 to 18.9% of phenotypic variance across three environments as well as MMDS (Table 2, Figure 1G). The second QTL QPm.heau-1BL in the marker interval wPt-668027-csLV46G22 explained 4.8, 11.5, 8.3, and 6% of phenotypic variance in Shangqiu 2014, Zhengzhou 2015, and Shangqiu 2015 and MMDS, respectively (Table 2, Figure 1E). The third QTL, QPm.heau-6BS located in the interval wPt-2587-wPt-3581 on chromosome 6BS, explained from 7.2 to 9.4% of the phenotypic variances in two environments and MMDS (Table 2, Figure 1H). The fourth QTL, QPm.heau-2DL, was between wPt-667054 and wPt-667485, explaining 6.7 and 4.2% of the phenotypic variance in Zhengzhou 2015 and Shangqiu 2015, respectively (Table 2, Figure 1F).
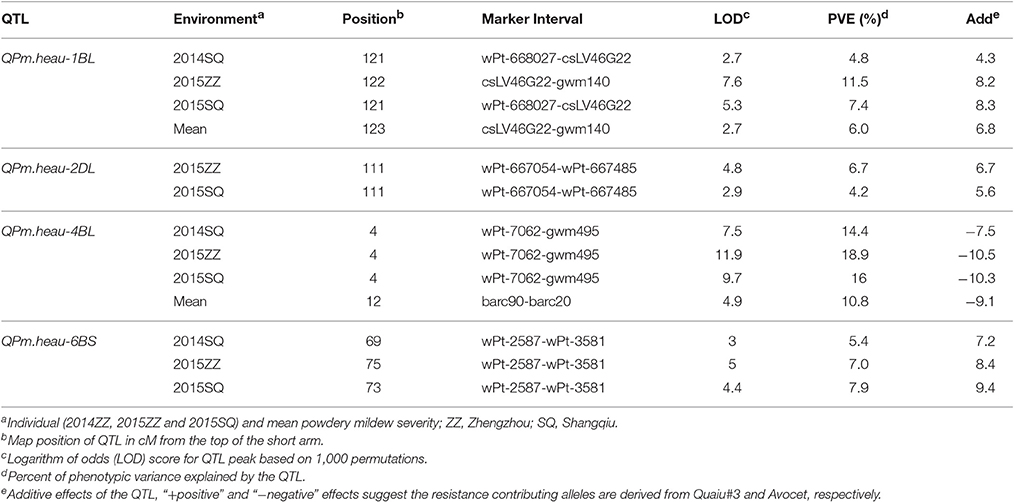
Table 2. Quantitative trait loci (QTL) for adult-plant resistance to powdery mildew detected in Avocet × Quaiu#3 RIL population through bi-parental analysis.
QTL for PM Resistance with JICIM
Joint inclusive composite interval mapping (JICIM) detected five consistent QTL on chromosomes 1BL, 2DL, 4BL, 5BL, and 6BS in two environments as well as MMDS in the two populations (Table 3, Figure 2). The resistance loci on chromosomes 1BL, 4BL, and 5BL were common in both populations, whereas QTL on chromosomes 2DL and 6BS were only detected in the Avocet × Quaiu#3 population. The QTL mapped on the long arm of chromosome 4B was from the susceptible parent Avocet, explaining from 3.9 to 13.1% of phenotypic variances. The second QTL was positioned on 1BL, around the near-diagnostic marker csLV46 and csLV46G22, which explained 5.2–12.9% of phenotypic variances. A third QTL with significant effects in both populations was located on 5BL between DArT markers wPt-4628 and wPt-3503. The QTL on chromosomes 2DL and 6BS were only detected in the Avocet × Quaiu#3 population, explaining 4.4–8.7% and 4.3–6%, respectively, of the phenotypic variances in two environments.
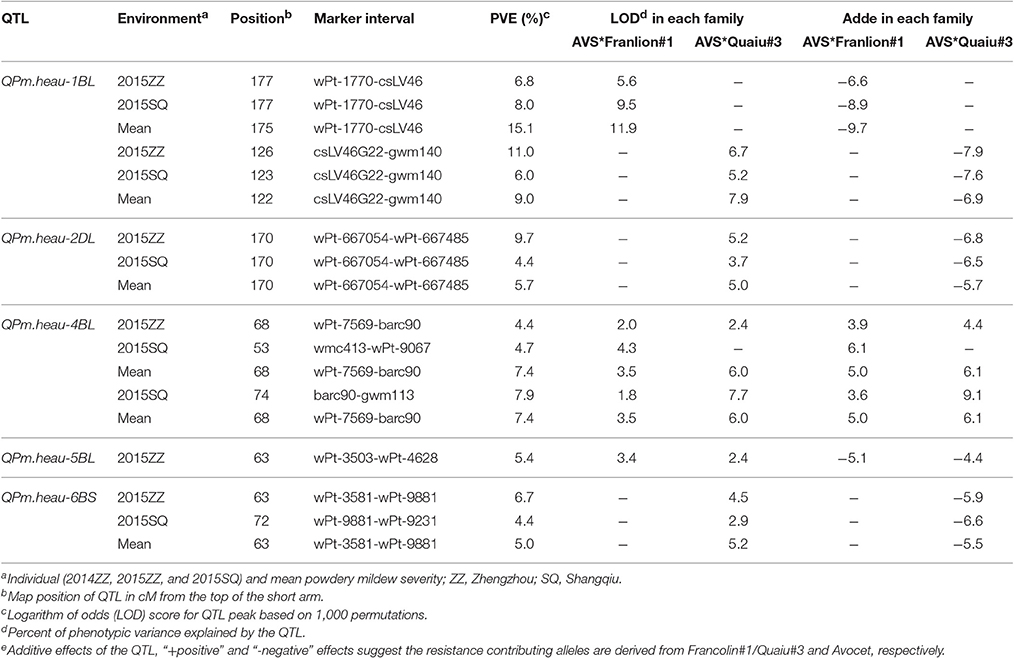
Table 3. Quantitative trait loci (QTL) for adult-plant resistance to powdery mildew detected by joint inclusive composite interval mapping (JICIM) in the two nested association mapping (NAM) populations (Avocet × Francolin#1 and Avocet × Quaiu#3).
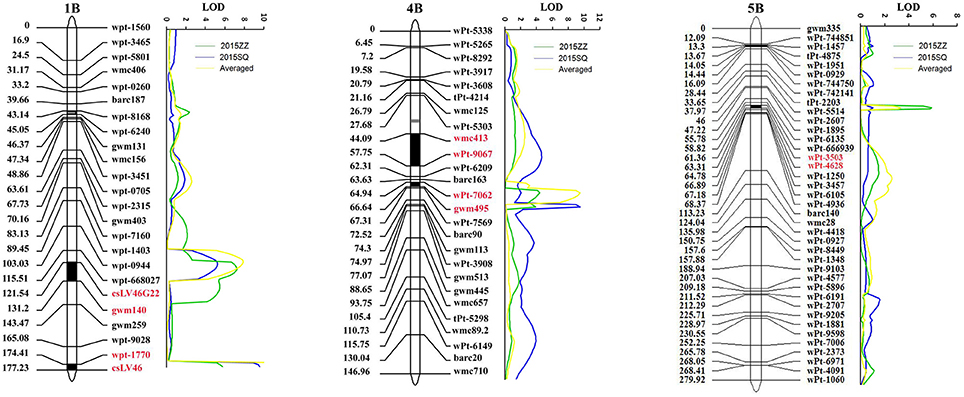
Figure 2. LOD contours for QTL to powdery mildew on chromosomes 1B, 4B, and 5B in the wheat nested association mapping (NAM) population by Joint inclusive composite interval mapping (JICIM).
Effect of QPm.heau-1BL and QPm.heau-4BL in Two Populations
Based on the closely linked flanking markers, PM severity distributions were observed among the RILs from the Avocet × Francolin#1 and Avocet × Quaiu#3 crosses in the presence or absence of two consistent loci, QPm.heau-1BL and QPm.heau-4BL. RILs with QPm.heau-4BL displayed mean MDS of 38.5% and 35.6% in the Francolin#1 and Quaiu#3 populations, respectively, whereas the mean severities were 53.9 and 53.3% for RILs lacking QPm.heau-4BL, respectively (Table 4). In Francolin#1 population, a significant MDS reduction of 12.2–16.3% was observed in RILs with QPm.heau-4BL, compared with those without resistance allele across three environments as well as MMDS (Table 4). A significant effect was also found in Quaiu#3 population with a mean PM reduction of 13–21.1% for RILs carrying resistance allele at QPm.heau-4BL locus (Table 4). QPm.heau-1BL also showed a significant effect on PM at the adult plant stage in both populations. A mean MDS reduction of 16.7–24.6 and 10.7–18.9% was observed in Avocet × Francolin#1 and Avocet × Quaiu#3 RILs with QPm.heau-1BL, respectively, compared to RILs without this locus (Table 5).
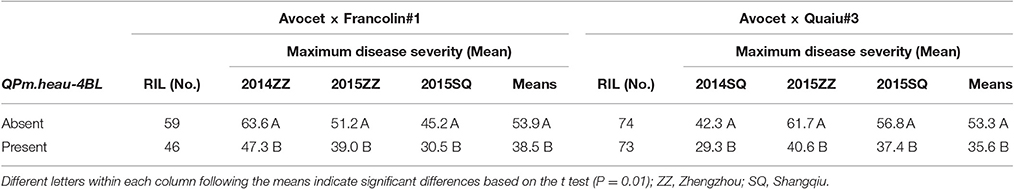
Table 4. t-tests for the comparison of mean maximum disease severity in Avocet × Francolin#1 and Avocet × Quaiu#3 RILs when QPm.heau-4BL was either absent or present.
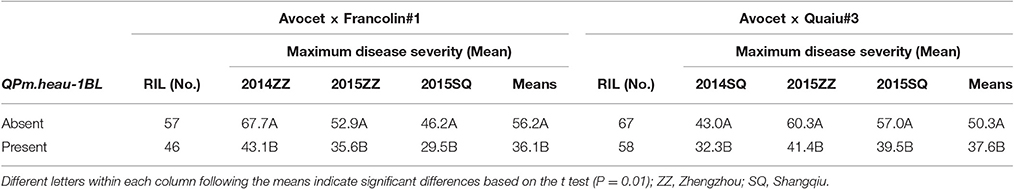
Table 5. t-tests for the comparison of mean maximum disease severity in Avocet × Francolin#1 and Avocet × Quaiu#3 RILs when QPm.heau-1BL was either absent or present.
Discussion
Quantitative and qualitative genetic analyses showed that about three to four PM resistance genes conferred partial resistance in Francolin#1 and Quaiu#3 derived RIL populations, respectively. These results corroborated the QTL mapping results. Avocet contributed a consistent QTL in both mapping populations. Most of the QTL identified through bi-parental QTL analyses were also identified by JICIM with the exception of one minor locus on chromosome 1DL from Francolion#1, indicating JICIM was a powerful method for QTL analysis in nested association mapping populations due to an improvement of linkage maps.
The QTL on chromosome 1BL was common in both populations. It was closely linked to the diagnostic marker csLV46 or csLV46G22 for known PM APR gene Pm39. However, there is only one common marker between the genetic linkage groups on chromosome 1BL for the two populations (Figures 1A,E). Due to the limited number of SSR and DArT markers at present, only three other poorly fit common markers were finally added to the linkage map on 1BL for Avocet × Francolin#1 population, which might lead to the long distance between the markers csLV46 and csLV46G22 on the joint linkage map. PM QTL at the same position on 1BL was reported in Massey (Liu et al., 2001) and its derivative USG3209 (Tucker et al., 2007). The gene Pm39 is known to have pleotropic effects on leaf rust, stripe rust and stem rust (Lillemo et al., 2008; Singh et al., 2013; Kolmer et al., 2015). Previous studies on leaf rust and stripe rust resistance in the two populations have shown that the chromosome 1BL QTL explained 23.0–44.5% and 6.0–43.6% for leaf rust and stripe rust severity variations in Avocet × Francolin#1 and Avocet × Quaiu#3, respectively (Basnet et al., 2014; Lan et al., 2014), indicating that this locus should be the known pleiotropic APR gene Lr46/Yr29/Pm39 based on its effect on multiple diseases. Moreover, our study shows that the genetic background of the host genotype profoundly influences the expression of Pm39; the phenotypic variance attributed to QPm.heau-1BL was 20.3–26.1% in Avocet × Francolin#1 population compared to 4.8–11.5% in the Avocet × Quaiu#3 population.
QPm.heau-1DL, derived from wheat line Francolion#1, was located on the distal region of chromosome 1DL based on the genetic map of USG3209/Jaypee-1D (http://wheat.pw.usda.gov/cgi-bin/GG3/report.cgi?class=marker;query=∗5721∗; name = XwPt5721-1D). The LOD curves indicated that this locus had stable effects on PM resistance across all three environments, and explained 6.1–8.5% of the phenotypic variance. Keller et al. (1999) detected a QTL (QPm.sfr-1D) for PM resistance in Swiss winter wheat Forno, flanked by RFLP markers psr168 and glk558b, explaining 8.1–13.7% of phenotypic variances. Bougot et al. (2006) also identified a QTL for PM resistance on chromosome 1D in the French winter wheat line RE9001. It was closely linked with SSR marker gwm160 with a PVE of 12.6% in a single environment. These QTLs should be different from QPm.heau-1DL based on more than 20 cM genetic distance between them (Somers et al., 2004; He et al., 2011; Marone et al., 2013). Thus, it was supposed that QPm.heau-1DL was a novel QTL for APR to PM.
QPm.heau-2DL, mapped on the distal end of chromosome 2DL, was closely linked to the DArT marker wPt-667054 and co-located with Yr54 for stripe rust resistance in Quaiu#3 (Basnet et al., 2014). Keller et al. (1999) detected a QTL, QPm.sfr-2D, on chromosome 2DL within the marker interval psr932-psr331 explaining 8.2–12.2% of phenotypic variances in the Swiss spelt wheat cultivar Oberkulmer. It seems that the QTL from Oberkulmer is located more proximal on chromosome 2DL than QPm.heau-2DL (Somers et al., 2004; https://wheat.pw.usda.gov/GG3/). Börner et al. (2002) detected a minor QTL for PM resistance from the synthetic wheat W7984 on 2DL and it was flanked by markers glk588 and ksuD23. This QTL was possibly the same as QPm.heau-2DL based on genetic map (http://wheat.pw.usda.gov/GG3/). In addition, two PM QTLs at the distal of chromosome 2DL were also detected to be close to marker gwm301 in the Swedish winter wheat Folke and German spring wheat Naxos (Lu et al., 2012; Lillemo et al., 2012; Windju et al., 2017). Therefore, known stripe rust gene Yr54, which has only a slight effect in seedlings and a moderate effect in adult plants, could be linked to 2DL QTL, or may have pleiotropic effect on powdery mildew resistance. However, fine mapping and/or cloning of Yr54 will be necessary to resolve it.
A QTL from Avocet was identified in both populations in the current study. This QTL was located between wPt-6209 and barc20 on the long arm of chromosome 4B in the same location as QTL for PM resistance in the winter wheat Forno (Keller et al., 1999), synthetic hexaploid wheat W7984 (Börner et al., 2002), Italian wheat Strampelli (Asad et al., 2013) and the Israeli wheat cultivar Oligoculm (Liang et al., 2006) based on an integrated map developed by Marone et al. (2013). The same QTL contributed by Avocet has shown significant effects on stripe rust and leaf rust in the cross Avocet × Pavon in two seasons (1998–1999 and 1999–2000). Although our two populations were also tested for stripe rust and leaf rust in previous studies (Lan et al., 2014; Basnet et al., 2014), the LOD curves at the same location were far from being significant, which is in agreement with finding by Lillemo et al. (2008). Contribution of the 4BL QTL from Avocet explains the RILs showing transgressive segregation for both increased resistance and susceptibility (Figure S1).
Bougot et al. (2006) identified a QTL, QPm.inra-5BL.2, in the French wheat cultivar Courtot in the marker interval of gwm604-gwm790. However, this QTL was more than 40 cM away from QPm.heau-5BL based on Somers et al. (2004). Keller et al. (1999) reported a QTL for PM severity on chromosome 5BL in the Swiss spelt cultivar Oberkulmer between psr580b and psr143. It was ~30 cM from QPm.inra-5BL.2 (He et al., 2011) and came closer to the terminal end of chromosome, indicating the difference from QPm.heau-5BL. Recently, Asad et al. (2013) identified QPm.caas-5BL.1 and QPm.caas-5BL.2 in the Italian wheat cultivar Strampelli, flanked by gwm335-barc331 and barc331-wmc537, respectively, which fall within the interval of QPm.heau-5BL in the present study. Furthermore, QPm.heau-5BL was also located in the region corresponding to QPm.inra-5BL.1 on chromosome 5B (Bougot et al., 2006; Marone et al., 2013). Lu et al. (2009) detected a significant QTL for stripe rust resistance in a similar chromosomal position corresponding to QPm.caas-5BL.2, but QPm.heau-5BL did not show significant effect on stripe rust (Lan et al., 2014).
Keller et al. (1999) found a QTL within the AFLP marker interval of psr167b-psr964 on the long arm of chromosome 6B in winter wheat Forno, which should differ from QPm.heau-6BS based on the wheat consensus map. Lan et al. (2009) identified a QTL conferring resistance to PM on chromosome 6BS in Chinese winter wheat Bainong 64 on the same position as QPm.heau-6BS. The former explained 10.3–13.2% of total phenotypic variance in PM severity in different environments, which is similar to QPm.heau-6BS in the present study. In addition, a PM QTL at a similar position on chromosome 6BS was also detected in the Swedish winter wheat Folke (Lillemo et al., 2012).
In this study, two consistent QTL, QPm.heau-1BL and QPm.heau-4BL, were detected in two populations based on ICIM and JICIM. Furthermore, we were able to identify another locus (QPm.heau-5BL) in both populations using JICIM. The detection of QPm.heau-5BL is most probably attributed to the higher mapping power of JICIM.
Conclusion
In this study, we detected six QTL conferring APR to PM across environments, including QPm.heau-1BL (Pm39), QPm.heau-1DL, QPm.heau-2DL, QPm.heau-4BL, QPm.heau-5BL, and QPm.heau-6BS. QPm.heau-1DL is likely to be a novel QTL for resistance to PM. Besides Pm39, QPm.heau-2DL might be another co-located/pleotropic resistance gene in wheat, which maps to same position as Yr54. The closely linked molecular markers for each QTL would be of great significance for QTL pyramiding from the parents Francolin#1 and Quaiu#3 in the future. It may be even better to use RILs that have shown significantly higher level of resistance than the parents.
Author Contributions
YR carried out the experiment and wrote the manuscript. WH, WZ, XC, and DC participated in field trials. CL, BB, and RS performed SSR and DArT genotyping and revised the paper. YR and FC designed the study. All authors have read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Thanks to Prof. Kehui Zhan for providing help in field trials. This project was funded by the National Key Research and Development Program (2016YFD0101802), Henan Major Science and Technology Project (161100110500) and Henan Science and Technology Innovation Outstanding Youth Funding (174100510001) of China. CL acknowledges support from the International Foundation for Science (C/5147-1), the Australian Grains Research and Development Corporation (GRDC) and the Australian Cereal Rust Control Program (ACRCP).
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01212/full#supplementary-material
Figure S1. Frequency distribution of Avocet × Francolin#1 (A) and Avocet × Quaiu#3 (B) recombinant inbred lines (RILs) for powdery mildew maximum disease severities (MDS) in three environments. ZZ14, Zhengzhou 2014; ZZ15, Zhengzhou 2015; SQ14, Shangqiu 2014; SQ15, Shangqiu 2015.
Table S1. Analysis of variance for maximum disease severity values for RILs generated from Avocet × Francolin#1 and Avocet × Quaiu#3 crosses.
Table S2. Estimated number of resistance genes conferring adult plant resistance to powdery mildew in 196 Avocet × Francolin#1 and 195 Avocet × Quaiu#3 F5recombinant inbred lines (RILs) based on Mendelian segregation analysis.
Table S3. Range and mean maximum disease severities for parents and Avocet × Francolin#1 and Avocet × Quaiu#3 F5 recombinant inbred lines (RILs), and narrow-sense heritability estimates during 2014–2015 and 2015–2016 cropping seasons at Zhengzhou and Shangqiu, as well as the estimated minimum number of segregating resistance genes using the Wright method.
Table S4. summary of the main features of the wheat linkage maps in Avocet × Francolin#1 and Avocet × Quaiu#3 populations.
References
Alam, M. A., Xue, F., Wang, C. Y., and Ji, W. Q. (2011). Powdery mildew resistance genes in wheat: identification and genetic analysis. J. Mol. Biol. Res. 1, 20–39. doi: 10.5539/jmbr.v1n1p20
Asad, M. A., Bai, B., Lan, C.-X., Yan, J., Xia, X.-C., Zhang, Y., et al. (2013). QTL mapping for adult plant resistance to powdery mildew in Italian wheat cv. Strampelli. J. Integr. Agric. 12, 756–764. doi: 10.1016/S2095-3119(13)60297-X
Basnet, B. R., Singh, R. P., Ibrahim, A. M. H., Herrera-Foessel, S. A., Huerta-Espino, J., Lan, C. X., et al. (2014). Characterization of Yr54 and other genes associated with adult plant resistance to yellow rust and leaf rust in common wheat Quaiu3. Mol. Breed. 33, 385–399. doi: 10.1007/s11032-013-9957-2
Börner, A., Schumann, E., Fürste, A., Cöster, H., Leithold, B., Röder, M., et al. (2002). Mapping of quantitative trait loci determining agronomic important characters in hexaploid wheat (Triticum aestivum L.). Theor. Appl. Genet. 105, 921–936. doi: 10.1007/s00122-002-0994-1
Bougot, Y., Lemoine, J., Pavoine, M. T., Guyomarc'h, H., Gautier, V., Muranty, H., et al. (2006). A major QTL effect controlling resistance to powdery mildew in winter wheat at the adult plant stage. Plant Breed. 125, 550–556. doi: 10.1111/j.1439-0523.2006.01308.x
Gurung, S., Bonman, J. M., Ali, S., Patel, J., Myrfield, M., Mergoum, M., et al. (2009). New and diverse sources of multiple disease resistance in wheat. Crop Sci. 49, 1655–1666. doi: 10.2135/cropsci2008.10.0633
Hao, Y. F., Parks, R., Cowger, C., Chen, Z. B., Wang, Y. Y., Bland, D., et al. (2015). Molecular characterization of a new powdery mildew resistance gene Pm54 in soft red winter wheat. Theor. Appl. Genet. 128, 465–476. doi: 10.1007/s00122-014-2445-1
He, Z. H., Lan, C. X., Chen, X. M., Zou, Y. C., Zhuang, Q. S., and Xia, X. C. (2011). Progress and perspective in research of adult plant resistance to stripe rust and powdery mildew in wheat. Sci. Agric. Sin. 44, 2193–2215. doi: 10.3864/j.issn.0578-1752.2011.11.001
Herrera-Foessel, S. A., Lagudah, E. S., Huerta-Espino, J., Hayden, M. J., Bariana, H. S., Singh, D., et al. (2011). New slow-rusting leaf rust and stripe rust resistance genes Lr67 and Yr46 in wheat are pleiotropic or closely linked. Theor. Appl. Genet. 122, 239–249. doi: 10.1007/s00122-010-1439-x
Hsam, S. L. K., and Zeller, F. J. (2002). “Breeding for powdery mildew resistance in common wheat (Triticum aestivum L.),” in The Powdery Mildews: A Comprehensive Treatise, eds R. R. Belanger, W. R. Bushnell, A. J. Dik, and T. L. W. Carver (St. Paul, MN: The American Phytopathological Society), 219–238.
Keller, M., Keller, B., Schachermayr, G., Winzeler, M., Schmid, J. E., Stamp, P., et al. (1999). Quantitative trait loci for resistance against powdery mildew in a segregating wheat × spelt population. Theor. Appl. Genet. 98, 903–912. doi: 10.1007/s001220051149
Kolmer, J. A., Lagudah, E. S., Lillemo, M., Lin, M., and Bai, G. (2015). The Lr46 gene conditions partial adult-plant resistance to stripe rust, stem rust, and powdery mildew in Thatcher wheat. Crop Sci. 55, 2557–2565. doi: 10.2135/cropsci2015.02.0082
Krattinger, S. G., Lagudah, E. S., Spielmeyer, W., Singh, R. P., Huerta-Espino, J., McFadden, H., et al. (2009). A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science 323, 1360–1362. doi: 10.1126/science.1166453
Lan, C. X., Liang, S. S., Wang, Z. L., Yan, J., Zhang, Y., Xia, X. C., et al. (2009). Quantitative trait loci mapping for adult plant resistance against powdery mildew in Chinese wheat cultivar Bainong 64. Phytopathology 99, 1121–1126. doi: 10.1094/PHYTO-99-10-1121
Lan, C. X., Rosewarne, G. M., Singh, R. P., Herrera-Foessel, S. A., Huerta-Espino, J., Basnet, B. R., et al. (2014). QTL characterization of resistance to leaf rust and stripe rust in the spring wheat line Francolin#1. Mol. Breed. 34, 789–803. doi: 10.1007/s11032-014-0075-6
Leath, S., and Murphy, J. P. (1985). Virulence genes of the wheat powdery mildew fungus, Erysiphe graminis f. sp. Tritici in North Carolina. Plant Dis. 69:905.
Li, H. H., Bradbury, P., Ersoz, E., Buckler, E. S., and Wang, J. K. (2011). Joint QTL linkage mapping for multiple-cross mating design sharing one common parent. PLoS ONE 6:e17573. doi: 10.1371/journal.pone.0017573
Li, Z. F., Lan, C. X., He, Z. H., Singh, R. P., Rosewarne, G. M., Chen, X. M., et al. (2014). Overview and application of QTL for adult plant resistance to leaf rust and powdery mildew in wheat. Crop Sci. 54, 1907–1925. doi: 10.2135/cropsci2014.02.0162
Liang, S. S., Suenaga, K., He, Z. H., Wang, Z. L., Liu, H. Y., Wang, D. S., et al. (2006). Quantitative trait loci mapping for adult-plant resistance to powdery mildew in bread wheat. Phytopathology 96, 784–789. doi: 10.1094/PHYTO-96-0784
Lillemo, M., Asalf, B., Singh, R. P., Huerta-Espino, J., Chen, X. M., He, Z. H., et al. (2008). The adult plant rust resistance loci Lr34/Yr18 and Lr46/Yr29 are important determinants of partial resistance to powdery mildew in bread wheat line Saar. Theor. Appl. Genet. 116, 1155–1166. doi: 10.1007/s00122-008-0743-1
Lillemo, M., Bjørnstad, A., and Skinnes, H. (2012). Molecular mapping of partial resistance to powdery mildew in winter wheat cultivar Folke. Euphytica 185, 47–59. doi: 10.1007/s10681-011-0620-x
Lillemo, M., Skinnes, H., Singh, R. P., and Ginkel, M. V. (2006). Genetic analysis of partial resistance to powdery mildew in bread wheat line Saar. Plant Dis. 90, 225–228. doi: 10.1094/PD-90-0225
Limpert, E., Felsenstein, F. G., and Andrivon, D. (1987). Analysis of virulence in populations of wheat powdery mildew in Europe. Phytopathology 120, 1–8. doi: 10.1111/j.1439-0434.1987.tb04408.x
Liu, S. X., Griffey, C. A., and Maroof, M. A. S. (2001). Identification of molecular markers associated with adult plant resistance to powdery mildew in common wheat cultivar Massey. Crop Sci. 41, 1268–1275. doi: 10.2135/cropsci2001.4141268x
Lu, Q. X., Bjørnstad, Å., Ren, Y., Asad, M. A., Xia, X. C., Chen, X. M., et al. (2012). Partial resistance to powdery mildew in German spring wheat ¡®Naxos' is based on multiple genes with stable effects in diverse environments. Theor. Appl. Genet. 125, 297–309. doi: 10.1007/s00122-012-1834-6
Lu, Y. M., Lan, C. X., Liang, S. S., Zhou, X. C., Liu, D., Zhou, G., et al. (2009). QTL mapping for adult plant resistance to stripe rust in Italian common wheat cultivars Libellula and Strampelli. Theor. Appl. Genet. 119, 1349–1359. doi: 10.1007/s00122-009-1139-6
Marone, D., Russo, M. A., Laidò, G., Vita, P. D., Papa, R., Blanco, A., et al. (2013). Genetic basis of qualitative and quantitative resistance to powdery mildew in wheat: from consensus regions to candidate genes. BMC genomics 14:562. doi: 10.1186/1471-2164-14-562
McIntosh, R. A., Dubcovsky, J., Rogers, W. J., Morris, C., and Xia, X. C. (2017). Catalogue of Gene Symbols for Wheat: 2017 Supplement. Available online at: http://shigen.nig.ac.jp/wheat/komugi/genes/macgene/supplement2017.pdf
Moore, J. W., Herrera-Foessel, S., Lan, C. X., Schnippenkoetter, W., Ayliffe, M., Huerta-Espino, J., et al. (2015). A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat. Genet. 47, 1494–1498. doi: 10.1038/ng.3439
National Agro-technical Extension and Service Center (NAESC) (2010). Epidemic characteristics of wheat stripe rust in China in 2010 and its control strategies. China Plant Prot. 30, 32–35. doi: 10.3969/j.issn.1672-6820.2010.03.013
National Agro-technical Extension and Service Center (NAESC) (2011). Epidemic characteristics of wheat stripe rust in China in 2011 and its control strategies. China Plant Prot. 31, 28–32. doi: 10.3969/j.issn.1672-6820.2011.02.010
National Agro-technical Extension and Service Center (NAESC) (2012). Epidemic characteristics of wheat stripe rust in China in 2012 and its control strategies. China Plant Prot. 2, 43–46. doi: 10.3969/j.issn.1672-6820.2012.02.014
National Agro-technical Extension and Service Center (NAESC) (2013). Epidemic characteristics of wheat stripe rust in China in 2013 and its control strategies. China Plant Prot. 2, 35–39. doi: 10.3969/j.issn.1672-6820.2013.02.009
National Agro-technical Extension and Service Center (NAESC) (2014). Epidemic characteristics of wheat stripe rust in China in 2014 and its control strategies. China Plant Prot 2, 47–52. doi: 10.3969/j.issn.1672-6820.2014.02.013
National Agro-technical Extension and Service Center (NAESC) (2015). Epidemic characteristics of wheat stripe rust in China in 2015 and its control strategies. China Plant Prot 2, 43–48. doi: 10.3969/j.issn.1672-6820.2015.02.012
Risk, J. M., Selter, L. L., Krattinger, S. G., Viccars, L. A., Richardson, T. M., Buesing, G., et al. (2012). Functional variability of the Lr34 durable resistance gene in transgenic wheat. Plant Biotechnol. J. 10, 477–487. doi: 10.1111/j.1467-7652.2012.00683.x
Singh, R. P., Herrera-Foessel, S. A., Huerta-Espino, J., Lan, C. X., Basnet, B. R., Bhavani, S., et al. (2013). “Pleiotropic gene Lr46/Yr29/Pm39/Ltn2 confers slow rusting, adult plant resistance to wheat stem rust fungus,” in Proceedings Borlaug Global Rust Initiative, 2013 Technical Workshop (New Delhi), 17.1.
Somers, D. J., Isaac, P., and Edwards, K. (2004). A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 109, 1105–1114. doi: 10.1007/s00122-004-1740-7
Spielmeyer, W., McIntosh, R. A., Kolmer, J., and Lagudah, E. S. (2005). Powdery mildew resistance and Lr34/Yr18 genes for durable resistance to leaf and stripe rust cosegregate at a locus on the short arm of chromosome 7D of wheat. Theor. Appl. Genet. 111, 731–735. doi: 10.1007/s00122-005-2058-9
Tucker, D. M., Griffey, C. A., Liu, S., Brown-Guedira, G., Marshall, D. S., and Maroof, M. A. S. (2007). Confirmation of three quantitative trait loci conferring adult plant resistance to powdery mildew in two winter wheat populations. Euphytica 155, 1–13. doi: 10.1007/s10681-006-9295-0
Wang, Z. L., Li, L. H., He, Z. H., Duan, X. Y., Zhou, Y. L., Chen, X. M., Lillemo, M., and Singh, R. P. (2005). Seedling and adult-plant resistance to powdery mildew in Chinese bread wheat cultivars and lines. Plant Dis. 89, 457–463. doi: 10.1094/PD-89-0457
William, M., Singh, R. P., Huerta-Espino, J., Ortiz Islas, S., and Hoisington, D. (2003). Molecular marker mapping of leaf rust resistance gene Lr46 and its association with stripe rust resistance gene Yr29 in wheat. Phytopathology 93, 153–159. doi: 10.1094/PHYTO.2003.93.2.153
Keywords: Triticum aestivum L., powdery mildew, molecular marker, QTL, JICIM
Citation: Ren Y, Hou W, Lan C, Basnet BR, Singh RP, Zhu W, Cheng X, Cui D and Chen F (2017) QTL Analysis and Nested Association Mapping for Adult Plant Resistance to Powdery Mildew in Two Bread Wheat Populations. Front. Plant Sci. 8:1212. doi: 10.3389/fpls.2017.01212
Received: 09 May 2017; Accepted: 27 June 2017;
Published: 27 July 2017.
Edited by:
Giampiero Valé, Research Center for Cereal and Industrial Crops (CREA CER)-Vercelli, ItalyReviewed by:
Xianchun Xia, Institute of Crop Science, Chinese Academy of Agricultural Sciences, ChinaDaniela Marone, Cereal Research Center (CREA CER)-Foggia, Italy
Morten Lillemo, Norwegian University of Life Sciences, Norway
Copyright © 2017 Ren, Hou, Lan, Basnet, Singh, Zhu, Cheng, Cui and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng Chen, Y2hmMDA4OEAxNjMuY29t