 Enzo R. Bracamonte1
Enzo R. Bracamonte1 Pablo T. Fernández-Moreno2
Pablo T. Fernández-Moreno2 Fernando Bastida3
Fernando Bastida3 María D. Osuna4
María D. Osuna4 Ricardo Alcántara-de la Cruz5*
Ricardo Alcántara-de la Cruz5* Hugo E. Cruz-Hipolito6
Hugo E. Cruz-Hipolito6 Rafael De Prado2
Rafael De Prado2- 1Faculty of Agricultural Sciences, National University of Cordoba (UNC), Cordoba, Argentina
- 2Department of Agricultural Chemistry and Edaphology, University of Cordoba, Cordoba, Spain
- 3Department of Agroforestry Sciences, University of Huelva, Huelva, Spain
- 4Agrarian Research Center “Finca La Orden Valdesequera”, Badajoz, Spain
- 5Departamento de Entomologia/BIOAGRO, Universidade Federal de Viçosa, Viçosa, Brazil
- 6Bayer CropScience, Mexico City, Mexico
The Chloris genus is a C4 photosynthetic species mainly distributed in tropical and subtropical regions. Populations of three Chloris species occurring in citrus orchards from central Cuba, under long history glyphosate-based weed management, were studied for glyphosate-resistant status by characterizing their herbicide resistance/tolerance mechanisms. Morphological and molecular analyses allowed these species to be identified as C. ciliata Sw., Chloris elata Desv., and Chloris barbata Sw. Based on the glyphosate rate that causes 50% mortality of the treated plants, glyphosate resistance (R) was confirmed only in C. elata, The R population was 6.1-fold more resistant compared to the susceptible (S) population. In addition, R plants of C. elata accumulated 4.6-fold less shikimate after glyphosate application than S plants. Meanwhile, populations of C. barbata and C. ciliata with or without glyphosate application histories showed similar LD50 values and shikimic acid accumulation rates, demonstrating that resistance to glyphosate have not evolved in these species. Plants of R and S populations of C. elata differed in 14C-glyphosate absorption and translocation. The R population exhibited 27.3-fold greater 5-enolpyruvyl shikimate-3-phosphate synthase (EPSPS) activity than the S population due to a target site mutation corresponding to a Pro-106-Ser substitution found in the EPSPS gene. These reports show the innate tolerance to glyphosate of C. barbata and C. ciliata, and confirm the resistance of C. elata to this herbicide, showing that both non-target site and target-site mechanisms are involved in its resistance to glyphosate. This is the first case of herbicide resistance in Cuba.
Introduction
The use of herbicides is the most common weed control method (Délye, 2013; Fernández-Moreno et al., 2017b). However, herbicide resistance has caused this method to be quickly undermined. This scenario is the result of evolutionary adaptations in a target weed to herbicide applications (Powles and Yu, 2010; Beckie and Harker, 2017). Glyphosate [(N-phosphonomethyl)-glycine] is one of the most widely used herbicides, although it is also an herbicide with many cases of resistance (37 glyphosate-resistant species; Shaner et al., 2012; Bracamonte et al., 2016; Heap, 2017). This herbicide is systemic, non-selective and is used post-emergence, and it inhibits the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene (EC 2.5.1.19), triggering the catalysis of shikimate-3-phosphate and phosphoenolpyruvate (PEP) to form 5-enolpyruvyl-3-phosphate, an important step in the biosynthesis of aromatic amino acids in plants (Schönbrunn et al., 2001).
The mechanisms conferring glyphosate resistance are grouped into two major groups (Sammons and Gaines, 2014). Target-site resistance (TSR) can involve EPSPS gene mutations or EPSPS gene amplification. TSR was revealed to be caused by point mutations in the EPSPS gene with substitutions at Thr-102 and Pro-106 (Yu et al., 2015; Alcántara-de la Cruz et al., 2016b). Pro-106 substitutions have been found in several weeds (González-Torralva et al., 2012; Alarcón-Reverte et al., 2015; Fernandez et al., 2015), conferring low resistance levels to glyphosate on the order of 2- to 5-fold, while a double mutation (Thr-102 and Pro-106) increases resistance levels. Gene amplification is an adaptation also confers resistance to glyphosate (Gaines et al., 2013). The additional EPSPS produced from the amplified gene copies enables plants to survive higher glyphosate doses (Gaines et al., 2013; Chen et al., 2015; Yu et al., 2015). Non-target-site resistance (NTSR) mechanism results from reduced absorption and/or translocation, increased vacuolar sequestration, and metabolism to non-toxic compounds, causing less glyphosate transport to the EPSPS via the xylem and phloem (Délye, 2013). NTSR has being described as the most common mechanism of resistance to glyphosate and can confer unpredictable resistance (Powles and Yu, 2010). Similar to TSR, NTSR has been found to be a mechanism involved in resistance in many weeds (de Carvalho et al., 2012; Ge et al., 2012; Rojano-Delgado et al., 2012; Vila-Aiub et al., 2012).
The genus Chloris Sw. (Poaceae: Chloridoidae) is a C4 photosynthetic species distributed in tropical and subtropical regions (Molina and Agrasar, 2004). It has also been found in semi-arid areas inhabiting semi-natural grasslands and rural habitats such as roads and barren places (Cerros-Tlatilpa et al., 2015). The genus comprises 50–60 species in both hemispheres (Molina and Agrasar, 2004; Barkworth, 2007). Species of this genus have great economic and ecological importance worldwide because they are a source of forage, resist drought, increase soil fertility, require low inversion, and can be used as plant cover to protect soil from rain-driven erosion (Michael et al., 2012). However, some of them could be considered invasive weed species (Cerros-Tlatilpa et al., 2015).
Studies of herbicide resistant weeds in Cuba are scarce because of lack of knowledge of the issue. This situation is similar to the Dominican Republic, in which studies have already begun to be carried out (Bracamonte et al., 2016). Unfortunately, growers have already started to have many weeds in their citrus groves that are not controlled at the recommended field dose of glyphosate (720 g ae ha−1). Given that weed control strategies in larger commercial fields are absolutely the focus of glyphosate applications at the post-emergence growth stage, scientific confirmation is necessary. In this way, this study could define the species that are evolving toward to glyphosate resistance or even tolerant species to this herbicide.
An accurate assessment of taxonomic identity is a prerequisite to addressing population and individual plant-based functional studies. This is particularly true in the case of highly diverse genera, for which taxonomic and nomenclatural complexities generally arise, as is the case in Chloris (Molina and Agrasar, 2004).
This work aimed to characterize suspicious glyphosate-resistant populations of three different Chloris species in Cuba. Studies were conducted to (1) establish their taxonomical identity based on morphological and molecular analyses, (2) evaluate their resistance/tolerance levels, and (3) determine the mechanisms involved.
Material and Methods
Plant Material and Experimental Conditions
In 2014, our research group (Dr. Rafael De Prado) together with the Weed Science group of the Ministry of Agriculture of Cuba (Dr. Jorge Cueto), prospected for Chloris species in citrus orchards in central Cuba. These fields had been repeatedly treated with glyphosate (5 L ha−1, 36% w/v) continuously for over 10 years, and sometimes received more than one application per year (Cueto, personal communication).
Mature seeds of three suspicious glyphosate-resistant populations of Clhoris species (treated = T) were harvested separately in in citrus orchards from Arimao and Ceiba, in Cienfuegos Province, from at least 20 plants that had been survived to the last glyphosate treatment. Seeds from a population of each species from nearby locations with no known records of exposure to glyphosate were also collected (non-treated = NT).
The seeds were germinated in containers using a substrate of sand/peat (1:2 v/v), covered with parafilm, and placed in a growth chamber at temperatures of 28/18°C (day/night), with a 16 h photoperiod (850 μmol m−2 s−1) and 80% humidity. Subsequently, the seedlings from each population of the different Chloris species were transplanted individually into pots (1 plant per pot) containing the same substrate and placed in a growth chamber under the conditions described above. Furthermore, 20–30 plants from each population were placed in a greenhouse until flowering and fruiting.
Morphometric Study and Taxonomic Identity
Different taxonomically relevant morphological traits of inflorescences and caryopses were measured in greenhouse-grown plants of each species of the Chloris populations. The examined traits of inflorescences were the number of racemes, raceme length and spikelet density (number of spikelets per cm of raceme). For the spikelets, we examined the length and width of the lower and upper glumes, the number of sterile florets, the length of hairs surrounding the callus, the length and width of the lemma of the fertile floret (fertile lemma), the presence and length of hairs on or adjacent to the keel and on the margins of fertile lemma, the length of the palea of the fertile floret, the lemma length and width of the basal sterile floret, the lemma length of any additional sterile floret, and the presence and length of awns on lemmas. The characteristics of the caryopses were length, width, thickness, shape, and length of the embryo mark. The shape of caryopses was quantified as the variance in their three dimensions, each relative to length (Thompson et al., 1993). This dimensionless shape index varies between 0 for a perfect sphere and 0.22 for a disk- or needle-shaped item. Based on the above morphological characters, the three pairs of study populations were identified to the species level according to Molina and Agrasar (2004), and the nomenclature followed IPNI (2017).
Molecular Characterization of the Chloris Species by AFLP Primer Analysis
Twenty-four accessions from the Chloris spp. were employed as the study material. Genomic DNA was extracted from fresh young leaves of eight individual plants per species (four T and four NT), using the Speedtools Plant DNA Extraction kit (Biotools). The DNA concentration was measured using a NanoDrop ND 1000 spectophotometer. DNA was diluted to a final concentration of 10 ng/μL.
Twelve AFLP primer pairs were used [E36-M48 (E-ACC M-CAC); E36-M60 (E-ACC M-CTC); E37-M49 (E-ACG M-CAG); E38-M50 (E-ACT M-CAT); E40-M61 (E-AGC M-CTG); E35-M49 (E-ACA M-CAG); E36-M49 (E-ACC M-CAG); E35-M61 (E-ACA M-CTG); E40-M62 (E-AGC M-CTT); E32-M60 (E-AAC M-CTC); E33-M50 (E-AAG M-CAT)]. The reaction mix contained 10 ng template DNA, 2.5 U Taq DNA polymerase, 40 pmol primer, 200 μM dNTPs, 2.5 mM MgCl2, and 10 mM Tris-HCl all in a volume of 20 μl. The optimized thermal cycling conditions were 2 min at 94°C, followed by 40 cycles of 94°C for 25 s, 56°C for 25 s, 72°C for 25 s and a final extension at 72°C for 7 min. AFLP fragments were resolved in 25-cm gels (0.25 mm spacer thickness). Electrophoresis and detection were performed on a two-dye, model 4300 LICOR automated DNA Sequencer. Digital AFLP gel images were scored to obtain binary (band presence/absence) data using the SAGA GENERATION 2 software program.
Data clustering was conducted for AFLPs with the NTSYS-pc-2.2 software (Rohlf, 2000) using Jaccard's coefficients to define unweighted pair-group (UPGMA) dendograms. A principal coordinate analysis (PCA) was also performed with the NTSYS-pc program.
Dose-Response Assays
Plants of each Chloris population were sprayed at the 3–4 leaf growth stage. Glyphosate applications were applied with a laboratory chamber (SBS-060 De Vries Manufactering, Hollandale, MN) equipped with 8002 flat fan nozzle delivering 200 L ha−1 at 250 KPa at the height of 50 cm. The following glyphosate (Roundup®, 360 g ae L−1 as isopropylamine salt) rates were used: 0, 62.5, 125, 250, 500, 1,000, 2,000, 3,000, and 4,000 g ae ha−1. The experiment was design using nine replications per rate and was repeated twice. Plants were cut down at the soil surface 21 days after application (DAT).
Shikimic Accumulation Assay
Fifty mg of fresh tissue (4 mm leaf disks) were harvested from the youngest fully expanded leaf at the 3–4 leaf growth stage from 15 plants per population. Shikimic acid accumulation was determined according to Hanson et al. (2009). The glyphosate concentrations used were: 0, 500, and 1,000 μM. Sample absorbance was measured in a Beckman DU-640 spectrophotometer at 380 nm. The test was performed in triplicate on five treated and non-treated plants of each biotype in a completely random design. Results were expressed in mg of shikimic acid g−1 fresh tissue.
Absorption and Translocation
This study was carried out in the two C. elata populations.14C-glyphosate (American Radiolabeled Chemicals, Inc., USA) was added to the commercial herbicide to prepare a solution with a specific activity of 0.834 kBq μL−1. The final glyphosate concentration corresponded to 360 g ae ha−1 in 200 L ha−1. Plants were harvested at 24, 48, 48, 72, and 96 h after 14C-glyphosate treatment (0.834 kBq/plant). Five plants per populations at each time evaluated in a completely random design were handled according to Fernández-Moreno et al. (2017b). Radioactivity was analyzed by liquid scintillation spectrometry (LSS) in a Beckman LS 6500 scintillation counter (Beckman Coulter Inc. Fullerton, USA) during 10 min per sample. Percentage of 14C-glyphosate absorbed was expressed as [kBq in combusted tissue/(kBq in combusted tissue + kBq in leaf washes)] × 100.
Translocation of 14C-glyphosate in plants of the two C. elata populations was visualized using a phosphor imager (Cyclone, Perkin-Elmer, Waltham, MA, USA).
Glyphosate Metabolism
Six plants by each C. elata population at 3–4 leaf growth stage, were treated with 300 g ae ha−1 of glyphosate (as described in the dose-response assays) in a completely randomized design. Untreated plants were used as controls. Leaf tissues were washed with distilled water at 96 HAT, flash-frozen in liquid nitrogen, and stored at −40°C until use. Following the methodology described by Rojano-Delgado et al. (2010), glyphosate and its metabolites [aminomethyl phosphonate (AMPA), glyoxylate, sarcosine, and formaldehyde] were determined by reversed polarity capillary electrophoresis using a 3D Capillary Electrophoresis Agilent G1600A instrument equipped with a diode array detector (DAD, wavelength range 190–600 nm). Standard compounds used (glyphosate, AMPA, sarcosine, formaldehyde, and glyoxylate), were provided by Sigma-Aldrich, Spain. Glyoxylate naturally produced (untreated plants) was subtracted from the average of glyoxylate produced from glyphosate metabolism (treated plants) for each population.
EPSPS Enzyme Activity Assays
Leaf tissue of the C. elata populations (three samples of 5 g each) were ground to fine powder in liquid nitrogen a chilled mortar. The enzyme activity was extracted according to the protocol described by Sammons et al. (2007). The basal EPSPS activity in the extract was measured using a Modified Lowry Kit for Protein Determination (Sigma-Aldrich, Madrid, Spain) in accordance with the manufacturer's instructions. The specific EPSPS activity was determined using the EnzCheck Phosphate Assay Kit (Invitrogen, Carlsbad, CA) following the manufacturer's instructions, to determine the inorganic phosphate release. The glyphosate concentrations used were: 0, 0.1, 1, 10, 100, and 1,000 μM. The EPSPS activity was measured during 10 min at 360 nm in a spectrophotometer (Beckman DU-640) to determine the amount of phosphate (μmol) released μg of total soluble protein (TSP)−1 min−1 and expressed as a percentage with respect to the control (without glyphosate). The experiment was repeated three times for each samples.
EPSPS Gene Sequence
Young tissue (100–200 mg) was collected from 10 plants of each C. elata population and stored at −80°C for RNA extraction. Total RNA was isolated using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. RNA was then treated with TURBO DNase (RNase-Free; Ambion, Warrington, UK) to eliminate any DNA contamination. cDNA synthesis was carried out using 2 μg of total RNA and M-MLV (Moloney Murine Leukemia Virus) Reverse Transcriptase (Invitrogen, Carlsbad, CA, USA) in combination with oligo (dT)12−18 and random nonamers (Amersham Biosciences, Amersham, UK) according to the manufacturer's instructions. To amplify the EPSPS gene, primers previously designed by Perez-Jones et al. (2007) (forward: 5′ AGCTGTAGTCGTTGGCTGTG 3′; reverse: 5′ GCCAAGAAATAGCTCGCACT 3′), and de Carvalho et al. (2012) (forward: 5′ TAGTACAGCCAAAAGGGCAGTC-3′; reverse: 5′ GCCGTTGCTGGAGGAAATTC 3′) were used. These primers expand a 120-bp fragment of the EPSPS gene that contains the mutation site described as conferring resistance to glyphosate. The PCR reactions were carried out using cDNA from 50 ng of total RNA, 1.5 mM MgCl2, 0.2 mM dNTP, 0.2 μM of each primer, 1 × buffer, and 0.625 units of a 100:1 enzyme mixture of non-proofreading (Thermus thermophilus) and proofreading (Pyrococcus furiosus) enzymes (BIOTOOLS, Madrid, Spain) in a final volume of 25 μL. All PCR reactions were completed in duplicate, and the cycling conditions were as follows: 94°C for 3 min, 35 cycles of 94°C for 30 s, 55°C for 30 s and 72°C for 1 min, with a final extension cycle of 72°C for 10 min. An aliquot of the PCR product was loaded onto a 1% agarose gel to confirm the correct band amplification. The remainder of the PCR product was then purified using ExoSAP-IT® for PCR Product Clean-Up (USB, Ohio, USA) as indicated by the manufacturers. Five purified PCR products per population were sequenced (STAB VIDA, Caparica, Portugal).
Statistical Analysis
Dose-response and EPSPS enzyme activity data were subjected to non-linear regression analysis to determine the amount of glyphosate needed to reduce the fresh weight (GR50), increase the mortality (LD50), and inhibit the EPSPS activity (I50) by 50% in each Chloris population using the three-parameter log-logistic function: y = ([(d) / 1+(x/g)b]) where y is, depending on the analysis, the above ground fresh weight, survival, or EPSPS-activity expressed as the percentage of the non-treated control, d is the parameter corresponding to the upper asymptote, b is the slope, g is the GR50, LD50, or I50, and x (independent variable) is the glyphosate rate. Regression analyses were conducted using the drc package in the R program version 3.2.5 (Ritz et al., 2015). Resistance indexes (RI = R/S) were computed as R-to-S GR50, LD50, or I50 ratios.
An analysis of variance (ANOVA) was conducted to test for differences between populations in the different assays. When needed, differences between means were separated using the Tukey HSD test at P < 0.05. Model assumptions of the normal distribution of errors and homogeneous variance were graphically inspected. The ANOVAs were conducted using the Statistix (version 9.0) (Analytical Software, USA) software.
Results
Morphometric Study and Taxonomic Identity
Based on the examined morphological traits, the populations studied were identified as C. ciliata Sw., C. elata Desv., and Chloris barbata Sw. Only the last species listed is an annual species, and it can be easily separated from the other two species by its long-awned lemmas, the glabrous keel of the fertile lemma and the presence of hairs flanking the keel. In addition, caryopses of C. barbata were clearly more elongated in shape than those of the remaining species, as indicated by the higher values of the seed shape index. Distinctive traits of C. ciliata include a low number of racemes in the inflorescences, more than two sterile florets per spikelet, long keel hairs and relatively short awns. Compared to the other two species, racemes of C. elata plants were consistently longer and their fertile lemmas were shorter in length, showing much longer marginal hairs. In addition, the embryo mark in caryopses was shorter in this species (Table 1).
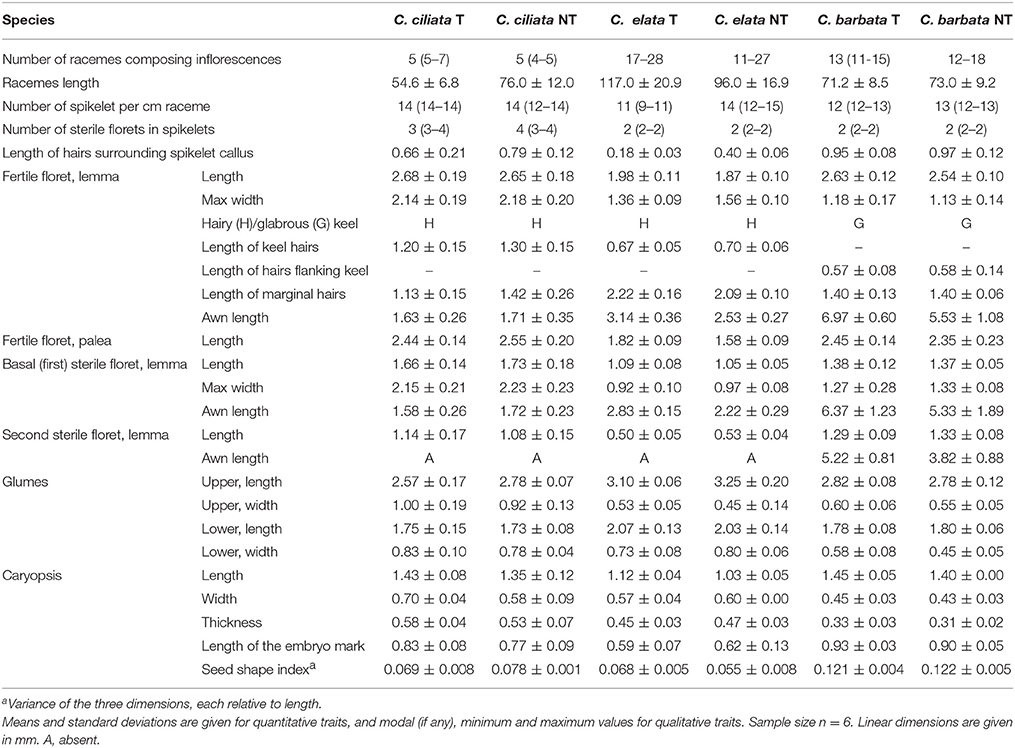
Table 1. Comparison of morphological traits from inflorescences and caryopses for the three pairs of studied Chloris populations, and their taxonomic identity at the species level.
Molecular Characterization of the Genus Chloris
A cluster analysis using UPGMA methods classified the Chloris populations into two major groups (I and II), thus providing complementary information to the morphological analysis. Group I contained all samples of C. ciliata whereas Group II consisted of two subgroups, II-1 including C. elata, and II-2 including C. barbata. It is noting that the cluster analysis did not separate T and NT populations of C. ciliata or C. barbata, while the T and NT populations of C. elata were clearly separated (Figure 1).
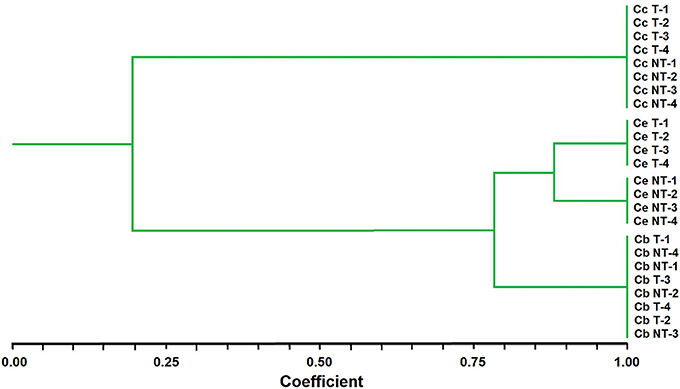
Figure 1. Genetic similarities among Chloris species after UPGMA analysis performed with AFLP marker data. Ce, Chloris elata; Cb, C. barbata; and Cc, C. Ciliata; T, plants with glyphosate history applications; NT, non-treated plants.
Dose-Response Assays
The fresh weight reduction by 50% (GR50) for the NT and T populations of C. elata was achieved at 88.3 and 542.1 g ae ha−1, respectively, i.e., T populations were 6.1-fold more resistance to glyphosate than the NT population. The LD50 values of the T populations exhibited 15-fold resistance than the NT population of C. elata. In contrast, GR50 and LD50 values in both C. barbata and C. ciliata species were not different between T and NT populations (Table 2, Figure 2).
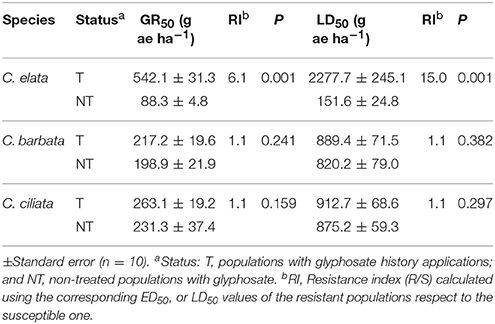
Table 2. Glyphosate rates required for 50 % reduction fresh weight (GR50) and survival (LD50) expressed as percentage of the mean untreated control of Chloris species.
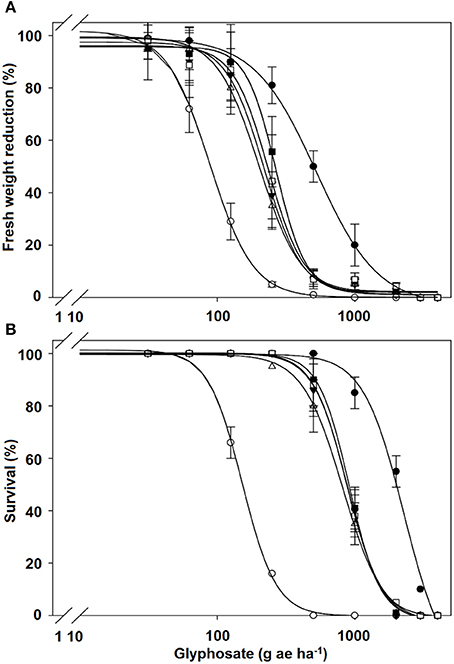
Figure 2. Glyphosate (A) dose-response on fresh weight reduction and (B) % survival expressed as percentage respect to untreated control of the treated (T) (•) and non-treated (NT) (○) populations of Chloris elata; T (▾) and NT (Δ) populations of C. barbata; and T (■) and NT (□) populations of C. ciliata. Vertical bars are ± standard errors (n = 9).
Shikimic Acid Accumulation
No differences were found between the exposure of leaf disks to 500 or 1,000 μM of glyphosate. At 1000 μM, the NT population of C. elata presented 4.6-fold more shikimic than the T population. However, the T populations of C. barbata and C. ciliata showed no differences, accumulating only 1.13 and 1.07-fold more shikimic acid, respectively, than their respective NT populations (Figure 3).
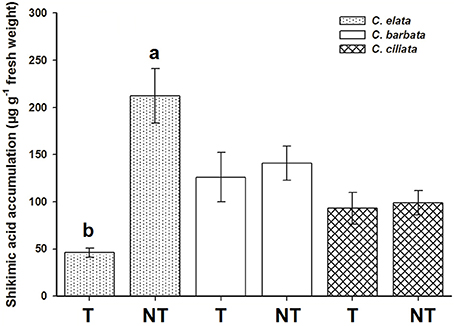
Figure 3. Shikimic acid accumulation in Chloris spp. plants at 1,000 μM glyphosate concentration. T = plants with glyphosate history applications; and NT = non-treated plants. Vertical bars are ± standard errors (n = 5). Different letters shown differences at 95% by the Tukey's test.
It was determined that C. barbata and C. ciliata exhibited natural tolerance to glyphosate, and the T and NT populations of C. elata were renamed resistant (R) and susceptible (S) to glyphosate, respectively. In the following assays, we focused in C. elata.
Absorption and Translocation
Total 14C-glyphosate recovery was 93.2 and 94.3% for the R and S populations of C. elata, respectively (data not shown). 14C-glyphosate absorption increased slowly in the first 72 HAT. At this time, population S had absorbed 30% of glyphosate, while population R had only absorbed 22%. The maximum glyphosate absorption rate was observed between 72 and 96 HAT, which was the two-fold higher in the S population (64%) than in the R population (33%; Figure 4A).
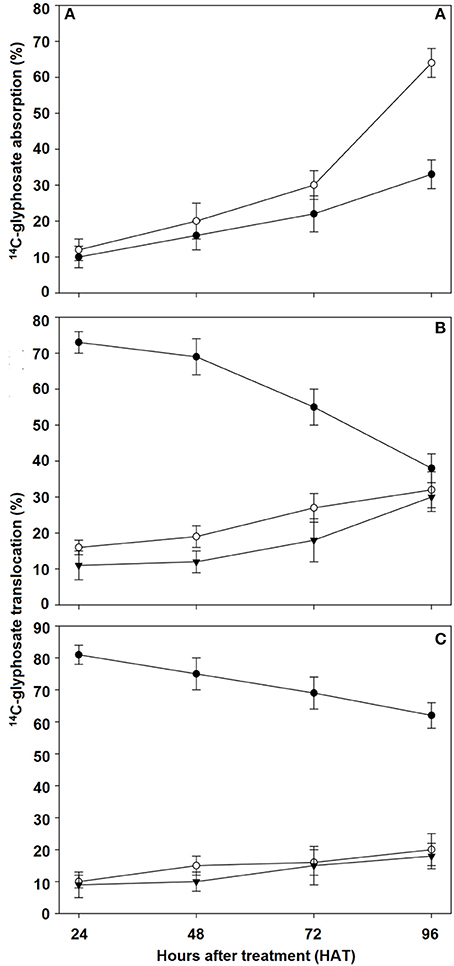
Figure 4. 14C-glyphosate absorption and translocation in susceptible (S) and resistant (R) Chloris elata populations from 24 to 96 h after glyphosate treatment. (A) 14C-glyphosate absorption in S (○) and R (•) plants. 14C-glyphosate translocation in S (B) and R (C) plants. Treated leaf (•), remaining shoot tissue (○), and roots (▾). Vertical bars are ± standard errors (n = 5).
In both populations, 14C-glyphosate levels in treated leaves of C. eleta declined from 24 to 96 HAT, with the rate of movement out of the treated leaf greater and faster in the S population than in R population. The initial amount quantified of 73% at 24 HAT in the treated leaf decreased to 38% at 96 HAT in S population. Conversely, the 14C-glyphosate was retained mainly in the leaf treated in the R population, dropping from 81 to 62% at 24 and 96 HAT, respectively. An average of 32 and 30% of the glyphosate translocated reached the remaining shoot tissue and roots at 96 HAT in the S population, whereas in R population it was only of 20 and 18%, respectively (Figures 4B,C).
The 14C-glyphosate visualization by phosphor imaging revealed differences in the distribution between the S and R populations of C. elata. There was a difference in the translocation of glyphosate from treated leaves to shoots and roots, and the S population translocated higher amounts of 14 C-glyphosate compared to the R population (Figure 5).
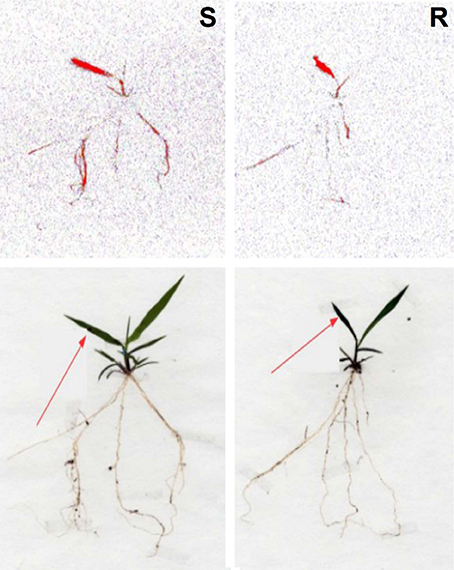
Figure 5. Translocation of 14C-glyphosate in susceptible (S) and resistant (R) Chloris elata plants at 96 h after application. The highest concentration of 14C-glyphosate is highlighted in red. Arrows indicate the treated leaf.
Glyphosate Metabolism
For glyphosate absorbed into the R and S C. elata populations, much of the herbicide was unaltered in plants by 96 HAT. At this time, 89.4 and 91.0% of the applied herbicide remained as glyphosate in plants of the R and S populations, respectively. The levels of AMPA were 7.2 and 6.4%, while glyoxylate levels reached 3.4 and 2.6% in the R and S plants, respectively. For both AMPA and glyoxylate, these differences between the R and S populations were non-significant (P = 0.8741 for AMPA, and P = 0.6318 for glyoxylate).
EPSPS Enzyme Activity Assays
The R population was 27.3-fold more resistant than the S population. The basal enzyme activity showed no differences between populations with 0.0987 to 0.0937 mmol mg−1 TPS−1 min−1 for the R and S populations, respectively (Figure 6).
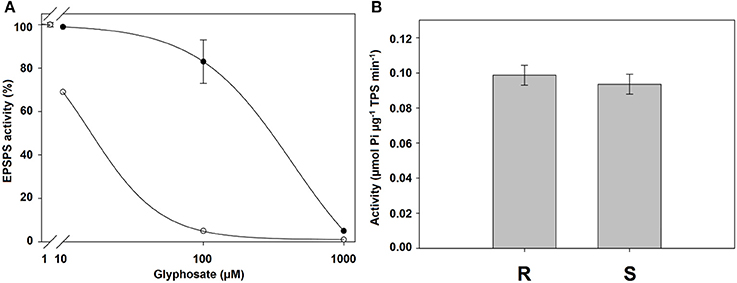
Figure 6. (A) EPSPS enzyme activity expressed as percentage of the untreated control in leaf extracts of plants from glyphosate-susceptible (○) and -resistant (•) Chloris elata plants. (B) Basal EPSPS activity for C. elata populations. Vertical bars are ± standard errors (n = 3).
Sequencing of the EPSPS Gene
A total of 120 bp of the EPSPS gene of the R and S C. elata populations was sequenced. The fragments were aligned and numbered based on a published EPSPS sequence of Leptochloa virgata (GenBank: KX425854) (Alcántara-de la Cruz et al., 2016c). Protein alignment of the predicted EPSPS fragments from R and S populations of C. elata showed 91.6 and 92.5% protein similarity, respectively, to that of L. virgata. The R population of C. elata showed an amino acid substitution at position 106 consisting of a proline to serine change. The substitution resulted in a TCG (serine) codon instead of a CCG (proline) codon (Figure 7).
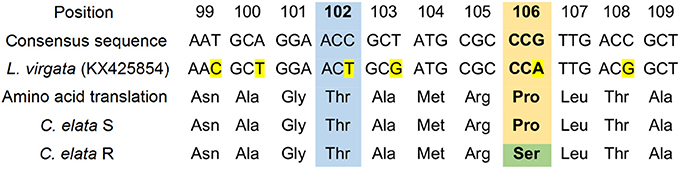
Figure 7. Partial alignment of nucleotides and amino acid sequences of EPSPS genes between glyphosate-susceptible and -resistant Chloris elata populations and Leptochloa virgata (GenBank: KX425854). The yellow color indicates nucleotide changes among species. The green color indicates an amino acid substitution at 106 position from Proline (CCG) to Serine (TCG). Box includes the 102 (blue) and 106 (orange) positions (amino acid number based on the start codon (ATG) of Arabidopsis thaliana, corresponding to point mutations associated for conferring glyphosate resistance.
Discussion
The three Chloris populations from citrus orchards in Cuba were identified as C. barbata, C. ciliata, and C. elata. The number of species of Chloris recognized by different authors in Mexico and in the nearby Caribbean Islands is variable (Barkworth, 2007; Cerros-Tlatilpa et al., 2015). In these regions, the most frequent species are C. ciliata, C. elata, C. barbata, and C. virgata (Cerros-Tlatilpa et al., 2015). In Cuba, 12 species of the Chloris genus have been found (Catasús-Guerra, 2002).
The AFLP-based classification of Chloris populations revealed molecular-based relationships between three basic entities, which closely matched the morphology-based identification of three different species. The selected AFLP markers can be useful candidates in the pursuit of disentangling phylogenetic relationships among Chloris species. However, although these markers allowed to separate the T and NT populations of C. elata, they cannot be used to detect glyphosate resistance, because could produce biased results due to fragment-size homoplasy (Caballero et al., 2008).
To characterize the glyphosate susceptibility in Chloris species, it is important to consider the innate tolerance to this herbicide that has been observed in some species of the genus. Depending on the species studied, the LD50 (% survival plant) values can vary between 515 and 703 g ae ha−1 (Ngo et al., 2017a,b). These values are lower than those we have found for C. barbata and C. ciliata, including the populations never exposed to this herbicide, demonstrating an innate tolerance to glyphosate in these two species. Similar results were described for Chloris polydactyla from Brazil, where even some accessions with no history of applications presented lower susceptibility to glyphosate than those accessions with a history of glyphosate applications (Barroso et al., 2014). Innate tolerance has been well studied in grass weeds (Fernández-Moreno et al., 2016), and leguminous species (Cruz-Hipolito et al., 2009, 2011; Rojano-Delgado et al., 2012; Alcántara-de la Cruz et al., 2016a; Mao et al., 2016). The mechanism proposed is a lack of 14C-glyphosate absorption and/or translocation in tolerant plants compared to susceptible ones.
C. elata has a different profile than C. barbata and C. ciliata, and its GR50 and LD50 values demonstrate a clear quantitative difference between those plants harvested from T fields compared to those plants from NT fields. The lower shikimic accumulation by the T C. elata population (4.9 times) compared to the NT population showed the greatest resistance level of this species, similar to other Chloris species, such as C. elata (5.4) from Brazil (Brunharo et al., 2016) and C. virgata (2.0–9.7) and Chloris truncata (2.4–8.7) from Australia (Ngo et al., 2017a,b). When glyphosate is applied via foliar application, the EPSPS enzyme is inhibited, and there is a rapid accumulation of shikimate (Shaner et al., 2005). The amount of glyphosate in the NT population of C. elata determined the inhibition of EPSPS and rapid shikimate accumulation demonstrating the high susceptibility (S) of this population. Therefore, the T population of C. elata was characterized as resistant (R) to glyphosate, and the populations T and NT of C. barbata and C. ciliata are tolerant to this herbicide. These results are reflected in those obtained in dose response assays. For this reason, we continued to study the glyphosate resistance mechanisms only in the case of C. elata.
To date, few cases of reduced glyphosate absorption and/or translocation have been studied as a mechanism of resistance in the genus Chloris, and the results are contradictory. 14C-glyphosate studies on C. virgata and C. truncata do not show significant differences in the absorption and subsequent translocation of the herbicide, and the resistance was determined by mechanisms within the target site (Ngo et al., 2017a,b). However, another study on C. elata shows that lower glyphosate absorption and translocation in the R population are the only mechanisms involved in its glyphosate resistance (Brunharo et al., 2016). In our case, the R C. elata population collected in Cuba shows a resistance mechanism similar to that previously found for the species. Thus, 14C-absorption and translocation are higher in the S population than in the R population. These results suggest that less absorption and translocation contributed to the resistance to glyphosate in R C. elata plants.
Glyphosate metabolism has thus far not been identified as a major mechanism of resistance in plants, but is likely the result of plants not succumbing to glyphosate because of the expression of another resistance mechanism (Sammons and Gaines, 2014; Fernández-Moreno et al., 2017a). Only in a few cases has metabolism been demonstrated to be a secondary mechanism in glyphosate resistance, because in these cases, other major mechanisms are involved (Bracamonte et al., 2016). Our research substantiates that of other studies, with <90% of the absorbed glyphosate remaining unaltered in R and S plants of C. elata. It is likely that the ability of grass weeds to metabolize glyphosate is diminished once EPSPS is inhibited (González-Torralva et al., 2012; Fernandez et al., 2015). Considering the small extent of glyphosate metabolism, the significance of this result is unlikely biologically meaningful in the resistance to glyphosate in C. elata.
The I50 values were significantly different between the C. elata populations. The R population exhibited a high resistance level compared to the S population. The results with high I50 values and low shikimic acid values, as has already been explained and demonstrated in other studies, are associated with alterations in the gene encoding the herbicide target protein (Sammons and Gaines, 2014; Yu et al., 2015). Then, the TSR mechanism could be involved in the resistance to this species. Similar results have been shown in other weed species, including L. virgata (Alcántara-de la Cruz et al., 2016c), Lolium multiflorum (Salas et al., 2015), and L. rigidum (Fernandez et al., 2015). In these cases, higher I50 values, as well as the higher basal activity of EPSPS, were found in the resistant populations compared to the susceptible populations. It was thought that an overexpression of EPSPS played a role as a resistance mechanism (Ngo et al., 2017a). However, there were no significant differences in the basal activity of EPSPS between R and S populations of C. elata, precluding the involvement of such a mechanism.
The EPSPS sequence alignment showed only a mutation point at position Pro-106-Ser in the R C. elata population. Four substitutions in this genomic EPSPS position (Pro-106-Ser, Pro-106-Thr, Pro-106-Ala, and Pro-106-Leu), have been reported in mono- and dicotyledonous weeds, endowing resistance to glyphosate (Sammons and Gaines, 2014). A mutation to a different amino acid at this point causes a structural change in the target site, shifting the other amino acids toward the inhibitor by reducing the available space (Healy-Fried et al., 2007). These explain the resistance of the R population of C. elata at a molecular level. Some grassweed species which have shown a mutation at Pro-106 position are: C. virgata (Ngo et al., 2017a), Echonoclhoa colona (Alarcón-Reverte et al., 2015), L. virgata (Alcántara-de la Cruz et al., 2016c), L. rigidum (Fernandez et al., 2015), and Poa annua (Cross et al., 2015), among others.
Conclusions
Morphological- and molecular-based analysis allowed the identification of the three Chloris species collected in citrus orchards from central Cuba. C. barbata and C. ciliata were characterized as being innately tolerant to glyphosate, and C. elata was identified as resistant to this herbicide. The last species had non-target site (reduced absorption and translocation) and target site (Pro-106-Ser mutation) resistance mechanisms to glyphosate.
These results confirm the first case of herbicide resistance in Cuba and strongly suggest that species of the Chloris genus can be either resistant or tolerant to glyphosate, supporting the previous reports of both glyphosate statuses in this genus.
Author Contributions
EB and RDP: Idea and designed the experiments; EB, PF-M, MO, FB, HC-H, and RA-dlC: Performed the research. PF-M, FB, MO, RA-dlC, and RDP; Analyzed the results. All authors contributed to write and approve the manuscript.
Funding
This work was funded by the AGL2016-78944-R (MICINN, Spain), GR15112 (Research Group AGA001 - Junta de Extremadura, Spain) and 242088 (CONACYT, Mexico) projects.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to Maria Cecilia Rodriguez-Garzón and Rafael A. Roldán-Gómez for their technical help, and the Dr. Jorge Cueto by the field prospections.
References
Alarcón-Reverte, R., García, R., Watson, S. B., Abdallah, I., Sabaté, S., Hernández, M. J., et al. (2015). Concerted action of target-site mutations and high EPSPS activity in glyphosate-resistant junglerice (Echinochloa colona) from California. Pest Manag. Sci. 71, 996–1007. doi: 10.1002/ps.3878
Alcántara-de la Cruz, R., Barro, F., Domínguez-Valenzuela, J. A., and De Prado, R. (2016a). Physiological, morphological and biochemical studies of glyphosate tolerance in Mexican cologania (Cologania broussonetii (Balb.) DC.). Plant Physiol. Biochem. 98, 72–80. doi: 10.1016/j.plaphy.2015.11.009
Alcántara-de la Cruz, R., Fernández-Moreno, P. T., Ozuna, C. V., Rojano-Delgado, A. M., Cruz-Hipolito, H. E., Domínguez-Valenzuela, J. A., et al. (2016b). Target and non-target site mechanisms developed by glyphosate-resistant hairy beggarticks (Bidens pilosa L.). Front. Plant Sci. 7:1492. doi: 10.3389/fpls.2016.01492
Alcántara-de la Cruz, R., Rojano-Delgado, A. M., Giménez, M. J., Cruz-Hipolito, H. E., Domínguez-Valenzuela, J. A., Barro, F., et al. (2016c). First resistance mechanisms characterization in glyphosate-resistant Leptochloa virgata. Front. Plant Sci. 7:1742. doi: 10.3389/fpls.2016.01742
Barkworth, M. E. (2007). “Chloris Sw,” in Manual of Grasses for North America, eds M. E.Barkworth, L. K. Anderton, K. M. Capels, S. Long, and M. B. Piep (Logan, UT: Utah State University Press) 185–253.
Barroso, A. A. M., Albrecht, A. J. P., Reis, F. C., Placido, H. F., Toledo, R. E., Albrecht, L. P., et al. (2014). Different glyphosate susceptibility in Chloris polydactyla accessions. Weed Technol. 28, 587–591 doi: 10.1614/WT-D-14-00039.1
Beckie, H. J., and Harker, K. N. (2017). Our top 10 herbicide-resistant weed management practices. Pest Manag. Sci. 73, 1045–1052. doi: 10.1002/ps.4543
Bracamonte, E., Fernández-Moreno, P. T., Barro, F., and De Prado, R. (2016). Glyphosate-resistant Parthenium hysterophorus in the Caribbean islands: non target site resistance and target site resistance in relation to resistance levels. Front. Plant Sci. 7:1845. doi: 10.3389/fpls.2016.01845
Brunharo, C. A., Patterson, E. L., Carrijo, D. R., Melo, M. S., Nicolai, M., Gaines, T. A., et al. (2016). Confirmation and mechanism of glyphosate resistance in tall windmill grass (Chloris elata) from Brazil. Pest Manag. Sci. 72, 1758–1764. doi: 10.1002/ps.4205
Caballero, A., Quesada, H., and Alvarez, E. R. (2008). Impact of amplified fragment length polymorphism size homoplasy on the estimation of population genetic diversity and the detection of selective loci. Genetics 179, 539–554. doi: 10.1534/genetics.107.083246
Cerros-Tlatilpa, R., Siqueiros-Delgado, M. L., and Skendzic, E. M. (2015). El género Chloris Sw. (Poaceae: Chloridoideae) en México. Acta Bot. Mex. 112, 95–147. doi: 10.21829/abm112.2015.1091
Chen, J., Huang, H., Zhang, C., Wei, S., Huang, Z., Chen, J., et al. (2015). Mutations and amplification of EPSPS gene confer resistance to glyphosate in goosegrass (Eleusine indica). Planta 242, 859–868. doi: 10.1007/s00425-015-2324-2
Cross, R., McCarty, L. B., Tharayil, N., McElroy, J. S., Chen, S., McCullough, P. E., et al. (2015). A Pro106 to Ala substitution is associated with resistance to glyphosate in annual bluegrass (Poa annua). Weed Sci. 63, 613–622. doi: 10.1614/WS-D-15-00033.1
Cruz-Hipolito, H. E., Osuna, M. D., Heredia, A., Ruiz-Santaella, J. P., and De Prado, R. (2009). Nontarget mechanisms involved in glyphosate tolerance found in Canavalia ensiformis plants. J. Agr. Food Chem. 57, 4844–4848. doi: 10.1021/jf9003253
Cruz-Hipolito, H. E., Rojano-Delgado, A., Domínguez-Valenzuela, J. A., Heredia, A., Luque De Castro, M. D., and De Prado, R. (2011). Glyphosate tolerance by Clitoria ternatea and Neonotonia wightii plants involves differential absorption and translocation of the herbicide. Plant Soil 347, 221–230. doi: 10.1007/s11104-011-0840-9
de Carvalho, L. B., Alves, P., González-Torralva, F., Cruz-Hipolito, H. E., Rojano- Delgado, A. M., De Prado, R., et al. (2012). Pool of resistance mechanisms to glyphosate in Digitaria insularis. J. Agric. Food Chem. 60, 615–622. doi: 10.1021/jf204089d
Délye, C. (2013). Unravelling the genetic bases of non-target-site-based resistance (NTSR) to herbicides: a major challenge for weed science in the forthcoming decade. Pest Manag. Sci. 69, 176–187. doi: 10.1002/ps.3318
Fernández-Moreno, P. T., Alcantara-de la Cruz, R., Cruz-Hipólito, H. E., Rojano-Delgado, A. M., Travlos, I., and De Prado, R. (2016). Non-target site tolerance mechanisms describe tolerance to glyphosate in Avena sterilis. Front. Plant Sci. 7:1220. doi: 10.3389/fpls.2016.01220
Fernández-Moreno, P. T., Alcántara-de la Cruz, R., Smeda, R. J., and De Prado, R. (2017a). Differential resistance mechanisms to glyphosate result in fitness cost for Lolium perenne and L. multiflorum. Front. Plant Sci. 8:1796. doi: 10.3389/fpls.2017.01796
Fernández-Moreno, P. T., Bastida, F., and De Prado, R. (2017b). Evidence, Mechanism and alternative chemical seedbank-level control of glyphosate resistance of a rigid ryegrass (Lolium rigidum) biotype from Southern Spain. Front. Plant Sci. 8:450. doi: 10.3389/fpls.2017.00450
Fernandez, P., Gauvrit, C., Barro, F., Menendez, J., and De Prado, R. (2015). First case of glyphosate resistance in France. Agron. Sustain. Dev. 35, 1469–1476. doi: 10.1007/s13593-015-0322-1
Gaines, T. A., Wright, A. A., Molin, W. T., Lorentz, L., Riggins, C. W., Tranel, P. J., et al. (2013). Identification of genetic elements associated with EPSPS gene amplification. PLoS ONE 8:e65819. doi: 10.1371/journal.pone.0065819
Ge, X., d'Avignon, D. A., Ackerman, J. J. H., Collavo, A., Sattin, M., Ostrander, E. L., et al. (2012). Vacuolar glyphosate-sequestration correlates with glyphosate resistance in ryegrass (Lolium spp.) from Australia, South America and Europe: a 31P-NMR investigation. J. Agric. Food Chem. 60, 1243–1250. doi: 10.1021/jf203472s
González-Torralva, F., Gil-Humanes, J., Barro, F., Brants, I., and De Prado, R. (2012). Target site mutation and reduced translocation are present in a glyphosate-resistant Lolium multiflorum Lam. biotype from Spain. Plant Physiol. Biochem. 58, 16–22. doi: 10.1016/j.plaphy.2012.06.001
Hanson, B. D., Shrestha, A., Shaner, D. L. (2009). Distribution of glyphosate-resistant horseweed (Conyza canadensis) and relationship to cropping systems in the Central Valley of California. Weed Sci. 57, 48–53. doi: 10.1614/WS-08-103.1
Healy-Fried, M. L., Funke, T., Priestman, M. A., Han, H., and Schonbrunn, E. (2007). Structural basis of glyphosate tolerance resulting from mutations of Pro(101) in Escherichia coli 5-enolpyruvylshikimate-3-phosphate synthase. J. Biol. Chem. 282, 32949–32955. doi: 10.1074/jbc.M705624200
Heap, I. (2017). International Survey of Herbicide Resistant Weeds. Available online at: http://www.weedscience.org/ (Accessed May14, 2017).
IPNI (2017). The International Plant Names Index. Plant Name Query. Available online at: http://www.ipni.org (Accessed May 18, 2017).
Mao, C., Xie, H., Chen, S., Valverde, B. E., and Qiang, S. (2016). Multiple mechanism confers natural tolerance of three lilyturf species to glyphosate. Planta 243, 321–335. doi: 10.1007/s00425-015-2408-z
Michael, P. J., Yeoh, P. B., and Scott, J. K. (2012). Potential distribution of the Australian native Chloris truncata based on modelling both the successful and failed global introductions. PLoS ONE 7:e42140. doi: 10.1371/journal.pone.0042140
Molina, A. M., and Agrasar, Z. E. R., (2004). Revisión taxonómica de las especies del género Chloris (Poaceae: Chloridoideae) en Sudamérica. Candollea 59, 347–428.
Ngo, T. D., Krishnan, M., Boutsalis, P., Gill, G., and Preston, C. (2017a). Target-site mutations conferring resistance to glyphosate in feathertop Rhodes grass (Chloris virgata) populations in Australia. Pest Manag. Sci. doi: 10.1002/ps.4512. [Epub ahead of print].
Ngo, T. D., Malone, J. M., Boutsalis, P., Gill, G., and Preston, C. (2017b). EPSPS gene amplification conferring resistance to glyphosate in windmill grass (Chloris truncata) in Australia. Pest Manag. Sci. doi: 10.1002/ps.4573. [Epub ahead of print].
Perez-Jones, A., Park, K. W., Polge, N., Colquhoun, J., and Mallory-Smith, C. A. (2007). Investigating the mechanisms of glyphosate resistance in Lolium multiflorum. Planta 226, 395–404. doi: 10.1007/s00425-007-0490-6
Powles, S. B., and Yu, Q. (2010). Evolution in action: plants resistant to herbicides. Annu. Rev. Plant Biol. 61, 317–347. doi: 10.1146/annurev-arplant-042809-112119
Ritz, C., Baty, F., Streibig, J. C., and Gerhard, D. (2015). Dose-response analysis using R. PLoS ONE 10:e0146021. doi: 10.1371/journal.pone.0146021
Rohlf, F. J. (2000). Numerical Taxonomy and Multivariate Analysis System. Setauket, NY: Applied Biostatistics.
Rojano-Delgado, A. M., Cruz-Hipólito, H. E., De Prado, R., Luque de Castro, M. D., and Franco, A. (2012). Limited absorption, translocation and enhanced metabolic degradation contribute to glyphosate tolerance in Mucuna pruriens var. utilis plants. Phytochemistry 73, 34–41. doi: 10.1016/j.phytochem.2011.09.007
Rojano-Delgado, A. M., Priego-Capote, F., Luque de Castro, M. D., and De Prado, R. (2010). Screening and confirmatory analysis of glyoxylate: a biomarker of plants resistance against herbicides. Talanta 82, 1757–1762. doi: 10.1016/j.talanta.2010.07.068
Salas, R. A., Scott, R. C., Dayan, F. E., and Burgos, N. R. (2015). EPSPS Gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) populations from Arkansas (United States). J. Agric. Food Chem. 63, 5885–5893. doi: 10.1021/acs.jafc.5b00018
Sammons, R. D., and Gaines, T. A. (2014). Glyphosate resistance: state of knowledge. Pest Manag. Sci. 70, 1367–1377. doi: 10.1002/ps.3743
Sammons, R. D., Meyer, J., Hall, E., Ostrander, E., and Schrader, S. (2007). A Simple Continuous Assay for EPSP Synthase from Plant Tissue. Available online at: http://www.cottoninc.com/fiber/AgriculturalDisciplines/Weed-Management/Managing-Glyphosate-Resistant-Palmer-Amaranth/Research-Programs/Monsanto/11a-Industry-Sammons-NCWSS07-poster.pdf (Accessed May 16, 2017).
Schönbrunn, E., Eschenburg, S., Shuttleworth, W. A., Schloss, J. V., Amrheini, N., Evans, J. N. S., et al. (2001). Interaction of the herbicide glyphosate with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic detail. Proc. Natl. Acad. Sci. U.S.A. 48, 1376–1380. doi: 10.1073/pnas.98.4.1376
Shaner, D. L., Lindenmeyer, R. B., and Ostlie, M. H. (2012). What have the mechanisms of resistance to glyphosate taught us? Pest Manag. Sci. 68, 3–9. doi: 10.1002/ps.2261
Shaner, D. L., Nadler-Hassar, T., Henry, W. B., and Koger, C. H. (2005). A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 53, 769–774. doi: 10.1614/WS-05-009R.1
Thompson, K., Band, S. R., and Hodgson, J. G. (1993). Seed size and shape predict persistence in soil. Funct. Ecol. 7, 236–241. doi: 10.2307/2389893
Vila-Aiub, M. M., Balbi, M. C., Distéfano, A. J., Fernández, L., Hopp, E., Yu, Q., et al. (2012). Glyphosate resistance in perennial Sorghum halepense (Johnsongrass), endowed by reduced glyphosate translocation and leaf absorption. Pest Manag. Sci. 68, 430–436. doi: 10.1002/ps.2286
Yu, Q., Jalaludin, A., Han, H., Chen, M., Sammons, R. D., and Powles, S. B. (2015). Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant Physiol. 167, 1440–1447. doi: 10.1104/pp.15.00146
Keywords: 5-enolpyruvyl shikimate-3-phosphate synthase, glyphosate translocation, herbicide resistance mechanisms, Pro-106 mutation, tall windmill grass
Citation: Bracamonte ER, Fernández-Moreno PT, Bastida F, Osuna MD, Alcántara-de la Cruz R, Cruz-Hipolito HE and De Prado R (2017) Identifying Chloris Species from Cuban Citrus Orchards and Determining Their Glyphosate-Resistance Status. Front. Plant Sci. 8:1977. doi: 10.3389/fpls.2017.01977
Received: 12 June 2017; Accepted: 02 November 2017;
Published: 15 November 2017.
Edited by:
Urs Feller, University of Bern, SwitzerlandReviewed by:
Nimesha Fernando, Federation University, AustraliaFernando José Cebola Lidon, Faculdade de Ciências e Tecnologia da Universidade Nova de Lisboa, Portugal
Khawar Jabran, Düzce University, Turkey
Ping Si, University of Western Australia, Australia
Copyright © 2017 Bracamonte, Fernández-Moreno, Bastida, Osuna, Alcántara-de la Cruz, Cruz-Hipolito and De Prado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ricardo Alcántara-de la Cruz, cmljYXJkby5sYUB1ZnYuYnI=