 Yan Yang
Yan Yang Bing Zhang1,3†
Bing Zhang1,3† Jörg Romeis
Jörg Romeis Yunhe Li
Yunhe Li- 1State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing, China
- 2Research Division Agroecology and Environment, Agroscope, Zurich, Switzerland
- 3Institute of Tropical Agriculture and Forestry, Hainan University, Haikou, China
Assessing the potential effects of insect-resistant genetically engineered (GE) plants on collembolans is important because these common soil arthropods may be exposed to insecticidal proteins produced in GE plants by ingestion of plant residues, crop pollen, or root exudates. Laboratory studies were conducted to evaluate the potential effects of two Bacillus thuringiensis (Bt)-rice lines expressing Cry1C and Cry2A in pollen and leaves and of their non-Bt conventional isolines on the fitness of the collembolan Folsomia candida and on the activities of its antioxidant-related enzymes, superoxide dismutase and peroxidase, and of its detoxification-related enzymes, glutathione reductase and glutathione S-transferase. Survival, development, reproduction, and the intrinsic rate of increase (rm) were not significantly reduced when F. candida fed on the Bt rice pollen or leaf powder than on the non-Bt rice materials; these parameters, however, were significantly reduced when F. candida fed on non-Bt rice pollen or non-Bt leaf-based diets containing the protease inhibitor E-64 at 75 μg/g. The activities of the antioxidant-related and detoxification-related enzymes in F. candida were not significantly affected when F. candida fed on the Bt rice materials, but were significantly increased when F. candida fed on the non-Bt rice materials containing E-64. The results demonstrate that Cry1C and Cry2A are not toxic to F. candida, and also indicate the absence of unintended effects on the collembolan caused by any change in plant tissue nutritional composition due to foreign gene transformation.
Introduction
Multiple genetically engineered (GE) rice lines producing Cry1 and Cry2 proteins derived from Bacillus thuringiensis (Bt rice) have been developed in China and many of these lines can efficiently control target pests such as Chilo suppressalis and Cnaphalocrocis medinalis (both Lepidoptera: Crambidae) (Li et al., 2016). Planting of Bt rice cultivars thus has great potential for reducing insecticide applications and to thereby benefit the environment and also human and animal health (Li et al., 2016; National Academies of Sciences Engineering Medicine [NASEM], 2016). Nevertheless, the cultivation of GE plants in general and of Bt rice, in particular, remains controversial because of safety concerns, one of which is the potential risk to valuable non-target organisms (Romeis et al., 2008; Li et al., 2017). Assessing the potential effects of Bt rice on non-target organisms is therefore an important part of the environmental risk assessment that must be conducted before the Bt rice can be commercially planted.
Recent studies have investigated the potential effects of Bt rice cultivars producing Cry1 and Cry2 proteins on non-target arthropods under laboratory or field conditions (reviewed by Li et al., 2016). Most of these studies have focused on plant-dwelling arthropods including predators belonging to different orders (Tian et al., 2010; Han et al., 2011, 2014, 2017; Li et al., 2014b, 2015; Wang et al., 2015, 2017; Meng et al., 2016), parasitic Hymenoptera (Han et al., 2015; Tian et al., 2017), and the silkworm Bombyx mori (Lepidoptera) (Yang et al., 2014). In all cases, these studies found that Cry proteins produced by Bt rice plants are very specific to target pest species in the order Lepidoptera and are not toxic to non-target species that do not belong to this order of insects (Li et al., 2016). Among the non-target species tested, only B. mori was adversely affected by consuming large amounts of Bt rice pollen containing Cry1C and Cry2A proteins (Yang et al., 2014). Sensitivity of B. mori to these proteins was expected because this species belongs to the target order Lepidoptera. The negative effects on B. mori, however, were only observed at exposure levels that far exceeded those expected under natural conditions, and the researchers, therefore, concluded that the planting of Bt rice would pose a negligible risk to B. mori (Yang et al., 2014).
Collembolans (springtails) are common arthropods in agricultural soils (Al-Deeb et al., 2003; Bai et al., 2010). In addition to being important consumers of plant residues and soil fungi, collembolans also help create humus (Bitzer et al., 2002). The common soil collembolan Folsomia candida (Collembola: Isotomidae) can be easily maintained in the laboratory and has been widely used as a standard test organism for assessing the non-target effects of insecticides and insecticidal GE plants (Fountain and Hopkin, 2005; Romeis et al., 2013; Yang et al., 2015; Zhang et al., 2017). This species has also been used in non-target risk assessment of Bt rice including rice lines expressing Cry1Ab or Cry1Ab/1Ac (Bai et al., 2010, 2011; Yuan et al., 2011, 2013, 2014). To our knowledge, information regarding the effects of Bt rice lines expressing the cry1C or cry2A genes on F. candida is limited to one study in which the collembolan was fed purified Cry1C and Cry2A proteins at concentrations that were 10 times higher than those in rice tissues. The results showed that F. candida was not sensitive to the Cry proteins (Yang et al., 2015). To date, the effects of feeding F. candida plant tissues from Bt rice lines containing Cry1C or Cry2A proteins (rather than purified proteins) have not been reported.
We therefore assessed the potential effects of ingestion of Bt rice pollen and leaves containing Cry1C or Cry2A proteins on F. candida with the hypothesis that consumption of these Bt rice material will not significantly affect the fitness of F. candida. Because plant tissues were used rather than purified toxins, we expected that the assessment would cover both the direct effects from the Cry proteins as well as possible indirect effects caused by unintended changes in plant composition as a consequence of the genetic transformation.
Materials and Methods
Plant Material
The transgenic Bt rice cultivars T1C-19 and T2A-1 and their corresponding non-transformed near-isoline Minghui 63 (MH63) were used in the experiments. T1C-19 plants express a modified cry1C gene, and T2A-1 plants express a modified cry2A gene; the proteins encoded by both genes target lepidopteran rice pests. The non-Bt rice line MH63 is an elite indica restorer line for cytoplasmic male sterility in China.
The rice lines were simultaneously planted in three adjacent plots at the experimental field station of the Institute of Plant Protection, CAAS, near Langfang City, Hebei Province, China (39.5°N, 116.4°E). Each plot was approximately 0.1 ha, and the plots were separated by a 1-m ridge. The rice seeds were sown in a seeding bed on May 6, 2015. When the seedlings were at the four-leaf stage, they were transplanted in the field (June 14, 2015). The plants were cultivated according to local agricultural practices but without pesticide sprays.
A previous study showed that the Cry protein concentrations in the two transgenic rice lines are higher in the leaves than in the stems or roots and are highest in the leaves at the seedling stage (Wang et al., 2016). Rice leaves were therefore collected from >50 randomly selected seedlings before they were transplanted in the field on June 10, 2015. The collected leaves were immediately frozen in liquid nitrogen, lyophilized, ground to a fine powder, and stored at -20°C until they were used in the experiments.
When rice plants in the plots reached the flowering stage, rice pollen was collected daily from 3 to 13 September 2015 by shaking the rice tassels in a plastic bag. The collected pollen was air dried at room temperature for 48 h and subsequently passed through a fine mesh (0.125 mm) to remove anthers and contaminants. Pollen collected from each rice line was pooled and stored at -20°C until used in the experiments.
Test Insects
The F. candida specimens used in the current study were obtained from the same permanent laboratory colony as described in our previous studies (Yang et al., 2015; Zhang et al., 2017). The collembolans used in the experiments were 12 days old, which followed the OECD guidelines (Organisation for Economic Cooperation and Development [OECD], 2016) and which ensured that the specimens were mature (Snider, 1973). To obtain 12-day-old insects, we placed F. candida adults in Petri dishes with plaster in the bases and allowed the females to oviposit for 48 h before all adults were removed. The eggs hatched after approximately 7 days and the neonates that hatched on a single day were subsequently fed on the baker’s yeast for 11 days and were then used in the experiments.
Rice Pollen Experiment
Our preliminary experiments showed that F. candida survival and development were similar on rice pollen (MH63) and on baker’s yeast, which is a diet that is favored by the collembolan (unpublished data). This indicated that rice pollen is a suitable food for F. candida and can be used in dietary exposure experiments.
For the experiment, 12-day-old F. candida were randomly selected, individually placed in Petri dishes (diameter 35 mm; height 10 mm; with plaster in the base), and subjected to one of the following dietary treatments: (i) MH63 rice pollen (non-Bt rice pollen; negative control); (ii) T1C-19 rice pollen (Bt pollen containing Cry1C); (iii) T2A-1 rice pollen (Bt pollen containing Cry2A); and (iv) MH63 rice pollen mixed with E-64 [trans-epoxysuccinyl-L-leucylamido (4-guanidino) butane], which served as a positive control. The protease inhibitor E-64 was purchased from Sigma–Aldrich (St. Louis, MO, United States) and was used as a positive control because it is known to be toxic to F. candida (Yang et al., 2015). For preparation of the positive control, stock solutions of E-64 were diluted with distilled water to a defined concentration and then mixed with non-Bt rice pollen (75 μg/g pollen dry weight [DW]). To ensure that the control and pollen treatments were prepared similarly, the same volume of distilled water was mixed with Bt and non-Bt rice pollen. All of the prepared pollen diets were lyophilized and ground into powder 3 days before the initiation of the experiment and were stored at -20°C until used. Each treatment was represented by 50 replicates (one collembolan and dish per replicate). The diets were renewed every 2 days to prevent the degradation of the test compounds. Survival, the number of fecal pellets produced, and the numbers of eggs and offspring produced by each collembolan were recorded twice daily (9:00 a.m. and 9:00 p.m.). Every seven days, the surviving individuals were photographed with a photo-microscope, and body length and head width were measured using a scale in the microscope. The experiment, which was conducted in a climate chamber at 20 ± 1°C with 70 ± 5% RH and a 12-h light/12-h dark cycle, was terminated after 35 days.
To estimate the intrinsic rate of natural increase (rm) of F. candida, individuals in each treatment were randomly assigned to one of three groups (16 or 17 individuals per group), resulting in three replicate groups per treatment. With the observed data, the rm was calculated per group using the equation described in our previous study (Zhang et al., 2017).
Rice Leaf Experiment
Powder from leaves alone is known to be an unsuitable food for F. candida (Yu et al., 1997; Romeis et al., 2003). Our preliminary experiments revealed that rice leaf powder mixed with baker’s yeast at a ratio of 10:1 (w: w) can support the survival, development, and reproduction of F. candida (unpublished data), and this mixture was used as the leaf-based diet in the current study.
The method used for the leaf-feeding experiment was similar to that used for the pollen-feeding experiment. The 12-day-old F. candida larvae were fed the following diets: (i) MH63 leaf powder mixed with baker’s yeast (leaf-based diet; negative control); (ii) T1C-19 leaf-based diet; (iii) T2A-1 leaf-based diet; and (iv) MH63 leaf-based diet containing E-64. Each treatment was represented by 50 replicates (one individual and dish per replicate), and the same life table parameters including the rm were recorded as described for the pollen-feeding experiment.
Uptake of Cry Protein by F. candida during the Feeding Experiments
To estimate the uptake of Cry1C or Cry2A protein by F. candida that fed on diets containing Bt rice pollen or leaf powder as described above, a separate assay was performed in which >30 Petri dishes (diameter 90 mm; height 10 mm), each containing >100 F. candida (12 days old) were provided with the Bt rice diets (pollen or leaf-based) or corresponding non-Bt diets as described above. After 7, 21, and 35 days of feeding, four samples per treatment (with 50–60 individuals per sample) were collected from different Petri dishes, resulting in a total of 72 samples (36 samples for pollen and 36 for leaves). The samples were frozen at -60°C for later ELISA analysis according to the methods described in our previous study (Li et al., 2015).
Stability and Bioactivity of Cry Proteins in the Diets
To evaluate the stability and bioactivity of Cry proteins in pollen or leaf-based diets during the feeding experiments, three 2- to 3-mg (FW) subsamples were collected from fresh diets that had been kept at -20° and from diets that had been exposed to F. candida for 2 days. The concentration and bioactivity of Cry proteins in the diets were analyzed by ELISA and sensitive-insect bioassays, respectively, according to the methods described in our previous study (Li et al., 2015).
Determination of Enzyme Activity
Folsomia candida (12 days old) were placed in Petri dishes (diameter 90 mm; height 10 mm; between 50 and 60 specimens per dish; >50 dishes in total) and exposed to non-Bt or Bt rice pollen or leaf-based diet or non-Bt rice pollen or leaf-based diet containing E-64 for 0, 7, 14, or 21 days as described before. At each sampling date, F. candida samples (200–300 individuals per sample, one sample per diet) were collected and stored at -20°C before the activities of the following enzymes were quantified in each sample: the antioxidant-related enzymes superoxide dismutase (SOD) and peroxidase (POD), and the detoxification-related enzymes glutathione (GR) and glutathione S-transferase (GST). The activities of these enzymes have been widely used as indicators of adverse effects caused by stomach poisons in F. candida and other arthropods (Bai et al., 2011; Yuan et al., 2011; Yang et al., 2015; Zhang et al., 2017). All enzyme activities were measured with enzyme kits from Nanjing Jiancheng Ltd., Co. (Nanjing, China) as described in our previous study (Zhang et al., 2017).
Data Analysis
Dunnett’s tests were used to analyze the difference between the treatments and the negative control for the following parameters: body length, head width, number of fecal pellets, number of eggs, and the intrinsic rate of increase. Hatching rates were analyzed by one-way ANOVA followed by HSD tests. Survival rates were analyzed with the Kaplan–Meier procedure and Logrank test. Cry protein concentrations and enzyme activities in F. candida collected on different days during the feeding assay were analyzed by repeated measures (RM-) ANOVA. In addition, Student’s t-tests were used to compare Cry protein concentrations in the fresh pollen/leaf diets vs. pollen/leaf diets exposed to F. candida for 2 days. Chi-square tests were used to compare the mortalities of the C. suppressalis larvae in the sensitive-insect bioassay. All statistical analyses were conducted using the software package SPSS (version 15.0 for Windows, 2006). Unless noted otherwise, values are presented as means ± SE.
Results
No Effects on Fitness of F. candida by Feeding on Bt Rice Pollen
The survival rates were >90% when F. candida fed on either Bt rice pollen (T1C-19 or T2A-1) or non-Bt rice pollen for 35 days, and there was no significant difference between any Bt pollen treatment and the control pollen treatment (χ2< 0.01, P = 0.99 for T1C-19; χ2 = 0.12, P = 0.73 for T2A-1) (Figure 1A). However, the survival rate was significantly reduced when F. candida fed on non-Bt pollen containing E-64 (χ2 = 17.660, P < 0.001). Similarly, the mean body length and head width of F. candida were not affected by ingestion of Bt rice pollen (P > 0.10 for all sampling dates) (Table 1). In addition, the number of eggs produced per individual and the number of fecal pellets produced per individual were not affected by feeding on Bt rice pollen (Figure 2) (Dunnett’s tests; T1C-19 pollen: P = 0.27 for number of eggs, and P = 1.00 for a number of fecal pellets; T2A-1 pollen: P = 1.00 for number of eggs, and P = 0.93 for a number of fecal pellets). All of these parameters, however, were significantly reduced when F. candida fed on the non-Bt rice pollen containing E-64 (Dunnett’s tests, all P < 0.01). Interestingly, the egg hatching rate of F. candida did not significantly differ among the diets (one-way ANOVA; F3,128 = 1.11, P = 0.35).
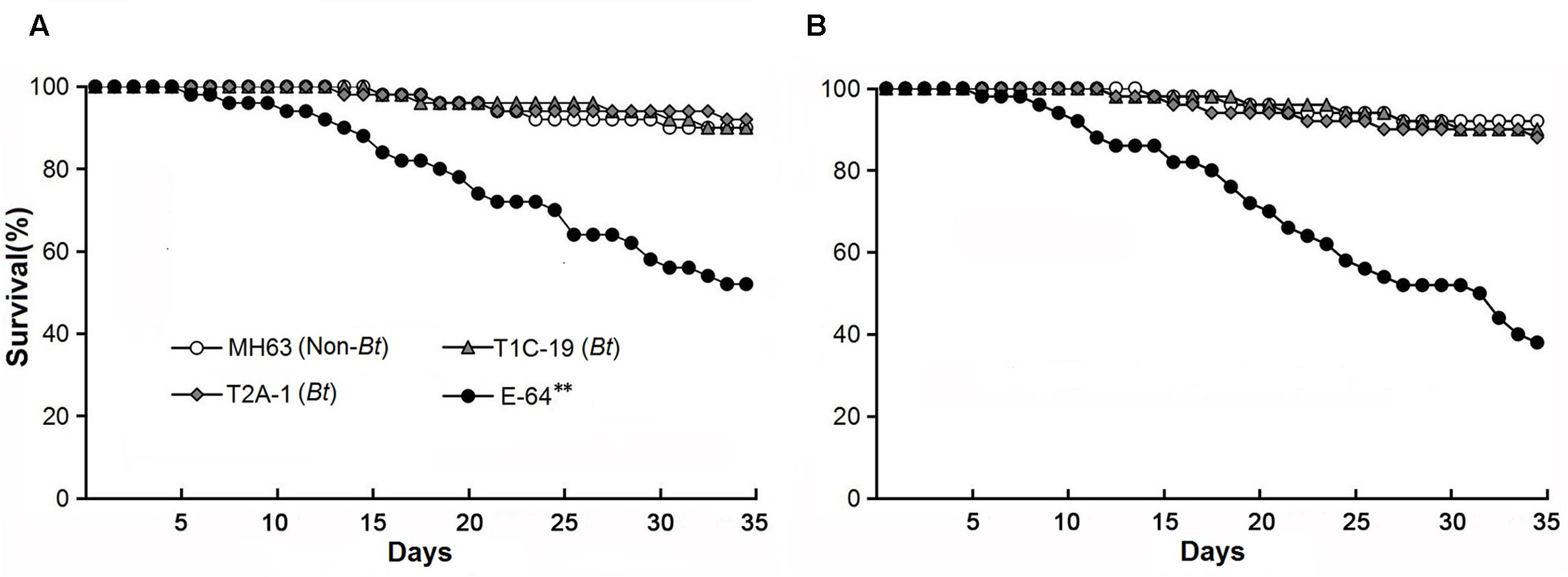
FIGURE 1. Survival of Folsomia candida on a diet consisting of rice pollen (A) or a mixture of rice leaf powder and baker’s yeast (10:1) (B). The pollen and leaf power were obtained from either of two Bt rice lines (T1C-19: Cry1C, and T2A-1: Cry2A) or their non-Bt near isoline MH63 (negative control). For the positive control, the corresponding non-Bt rice tissue was supplemented with E-64. Asterisks indicate a significant difference between the treatment and the negative control (Logrank test, P < 0.01) (n = 50).
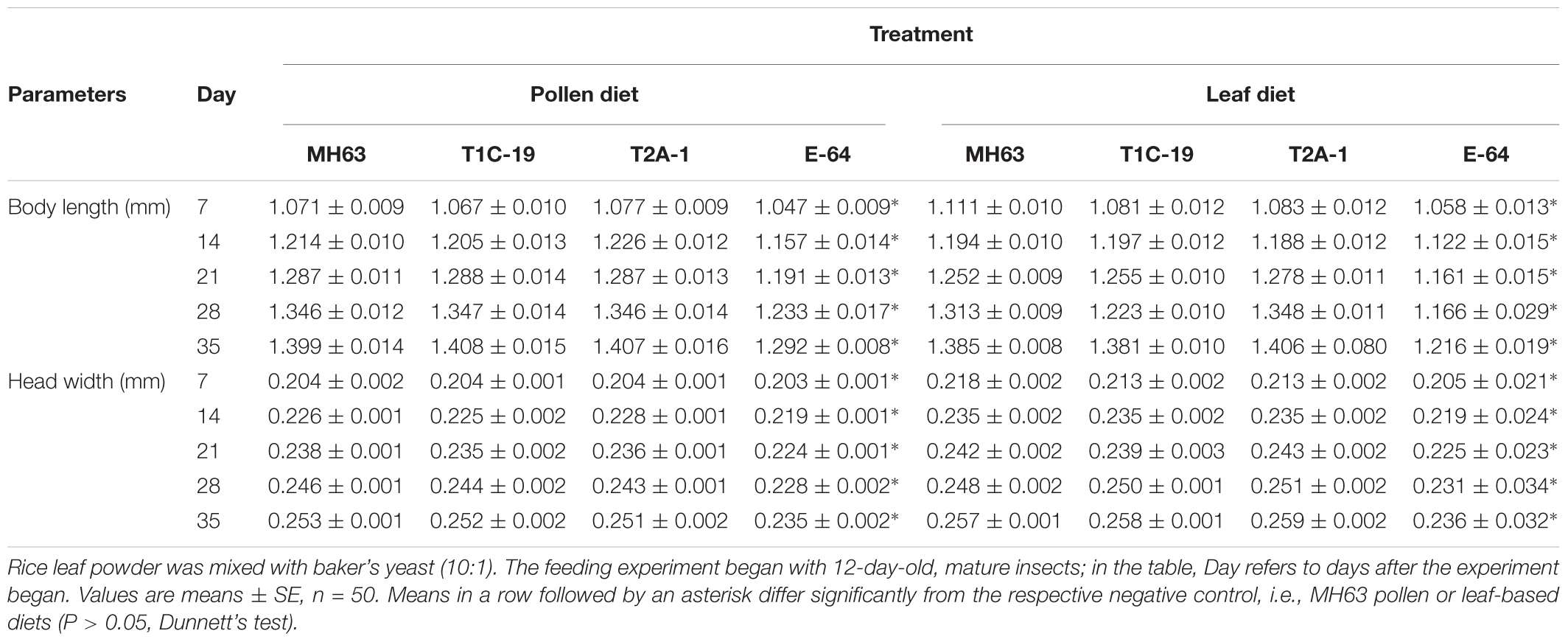
TABLE 1. Body length and head width of Folsomia candida that fed on a diet consisting of non-Bt pollen/leaf (MH63, negative control), Bt pollen/leaf (T1C-19: Cry1C, and T2A-1: Cry2A), or non-Bt pollen/leaf plus E-64 protein (E-64, positive control) for 35 days.
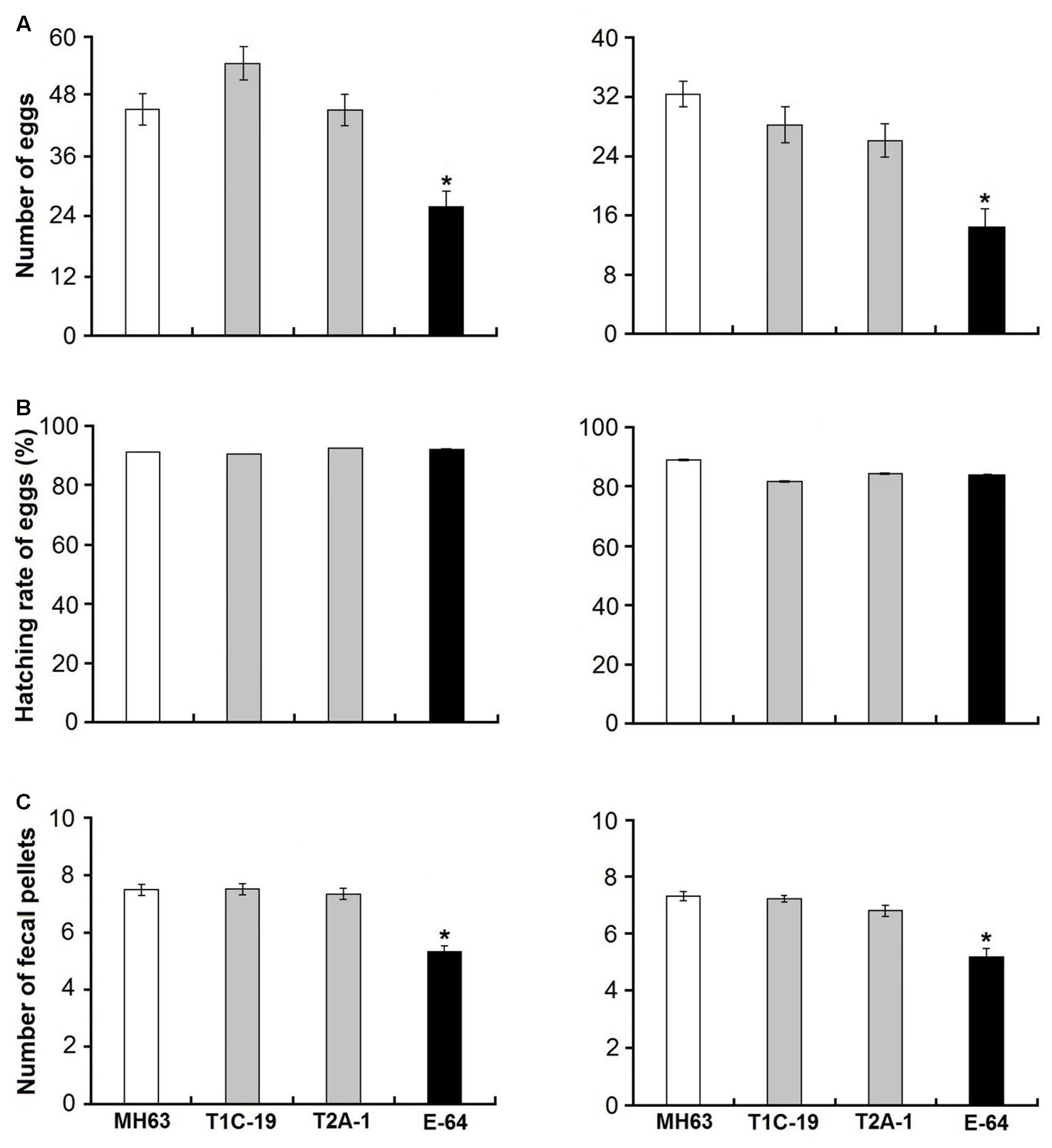
FIGURE 2. Fecundity (numbers of eggs) produced per individual (A), hatching rate of eggs (B), and numbers of fecal pellets produced per individual (C) of F. candida fed on a diet consisting of rice pollen (left) or a mixture of rice leaf powder and baker’s yeast (10:1) (right). The rice pollen and rice leaf powder were obtained from either of two Bt rice lines (T1C-19: Cry1C and T2A-1: Cry2A) or their non-Bt near isoline MH63 (negative control). For the positive control, the corresponding non-Bt rice tissue was supplemented with E-64 (positive control) for 35 days. Values are means ± SE, n = 50. Asterisks indicate a significant difference between the treatment and the negative control (MH63) (Dunnett’s test, P < 0.05).
The rm values were 0.14 ± 0.001, 0.14 ± 0.003, 0.14 ± 0.005, and 0.11 ± 0.003 when F. candida fed on the pollen from non-Bt plants, T1C-19, T2A-1, and non-Bt plants containing E-64, respectively. The mean rm values did not significantly differ between Bt and non-Bt pollen treatments (Dunnett’s tests; P = 0.66 for T1C-19 pollen, P = 0.99 for T2A-1 pollen), but the mean rm value was significantly reduced for F. candida that fed on the non-Bt pollen containing E-64 (P = 0.004).
Concentrations of Cry Proteins in F. candida Feeding on Bt Rice Pollen
As indicated by ELISA measurements, F. candida that fed on the Bt rice pollen diets contained Cry proteins. The concentration of Cry1C detected in F. candida on days 7, 21, and 35 was 38.49 ± 0.23, 41.89 ± 0.76, and 42.54 ± 1.44 ng/g DW, respectively, and the concentration significantly increased over time (RM-ANOVA, F1,3 = 2963.9, P < 0.001). The concentration of Cry2A detected in F. candida on days 7, 21, and 35 was 115.12 ± 6.17, 125.10 ± 0.19, and 252.85 ± 1.74 ng/g DW, respectively, and the concentration significantly increased over time (F1,3 = 79431.2, P < 0.001). No Cry protein was detected in F. candida that fed on the non-Bt rice pollen.
Stability and Bioactivity of Cry Proteins in Pollen Diets
According to ELISA measurements, the concentration of Cry1C protein in the freshly prepared Cry1C rice pollen diet was 1.82 ± 0.02 μg/g of pollen, and the concentration significantly decreased to 1.16 ± 0.02 μg/g of pollen after a 2-day feeding exposure to F. candida (Student’s t-test; t = 29.7, df = 6, P < 0.001). The concentration of Cry2A detected in the freshly prepared Cry2A rice pollen diet was 26.41 ± 0.19 μg/g of pollen, and the concentration significantly decreased to 24.07 ± 0.12 μg/g of pollen after a 2-day feeding exposure to F. candida (Student’s t-test; t = 10.4, df = 6, P < 0.001). No Bt protein was detected in non-Bt rice pollen.
The sensitive-insect bioassay showed that the mortality of C. suppressalis larvae was 3.3 ± 3.3% when the larvae were fed a diet containing the extract from non-Bt rice pollen for 7 days. The mortalities were 86.7 ± 3.3% or 83.3 ± 3.3% when C. suppressalis larvae were fed diets containing the extract from fresh Cry1C pollen (T1C-19) or Cry1C pollen that had been exposed to F. candida for 2 days. The mortalities were 80.0 ± 5.8% or 76.7 ± 3.3% when C. suppressalis larvae were fed diets containing the extract from fresh Cry2A pollen (T2A-1) or Cry2A pollen that had been exposed to F. candida for 2 days. Mortalities were not significantly different for larvae that fed on fresh diet vs. 2-day-old diet (Chi-square test; U = 0.13, df = 1, P = 0.72 for T1C-19 pollen; U = 0.10, df = 1, P = 0.75 for T2A-1 pollen).
No Effects on Fitness of F. candida by Feeding on Bt Rice Leaf Tissue
The survival rates were ≥88% when F. candida fed on the Bt rice (T1C-19 or T2A-1) leaf-based diets for 35 days, and there was no significant difference between any Bt leaf diet treatment and the control treatment (χ2 = 0.11, P = 0.74 for T1C-19; χ2 = 0.53, P = 0.47 for T2A-1) (Figure 1B). However, the survival rate was significantly reduced when F. candida fed on the non-Bt leaf-based diet containing E-64 (χ2 = 32.17, P < 0.001). Similarly, the mean body length and head width were not affected by ingestion of the Bt leaf-based diet (P > 0.10 for all sampling dates) (Table 1). In addition, the number of eggs produced per individual and the number of fecal pellets produced per individual was not affected by feeding on a Bt leaf-based diet (Figure 2) (Dunnett’s tests; T1C-19 leaves: P = 0.37 for number of eggs, and P = 0.98 for a number of fecal pellets; T2A-1 leaves: P = 0.093 for number of eggs, and P = 0.092 for a number of fecal pellets). All of these parameters, however, were significantly reduced when F. candida fed on a non-Bt leaf-based diet containing E-64 (Dunnett’s tests, all P < 0.01). As was the case with the pollen diets, the egg hatching rates of F. candida did not significantly differ among leaf-based diets (one-way ANOVA, F3,147 = 1.73, P = 0.16).
The rm values of F. candida were 0.138 ± 0.004, 0.138 ± 0.001, 0.136 ± 0.003, and 0.117 ± 0.007 for F. candida that fed on a leaf-based diet from non-Bt plants, T1C-19 plants, T2A-1 plants, and non-Bt plants containing E-64, respectively. The mean rm values did not significantly differ between Bt and non-Bt diet treatments (Dunnett’s tests, P = 1.0 for T1C-19 leaves, P = 0.969 for T2A-1 leaves), except that the mean rm value was significantly reduced when F. candida fed on the non-Bt diet containing E-64 (P = 0.017).
Concentrations of Cry Proteins by F. candida Feeding on Bt Rice Leaf Tissue
As indicated by ELISA measurements, F. candida that fed on a Bt rice leaf-based diet contained Cry proteins. The concentration of Cry1C detected in F. candida on days 7, 21, and 35 was 39.19 ± 0.24, 41.99 ± 1.09, and 64.60 ± 5.83 ng/g DW, respectively, when the collembolan fed on a T1C-19 leaf-based diet, and the concentration significantly increased over time (RM-ANOVA, F = 458.1, df = 3, P < 0.001). The concentration of Cry2A detected in F. candida on days 7, 21, and 35 was 147.53 ± 5.33, 140.08 ± 5.84, and 196.53 ± 0.19 ng/g DW, respectively, when the collembolan fed on a T2A-1 leaf-based diet, and the concentration significantly increased over time (F1,3 = 7628957.0, P < 0.001). No Cry protein was detected in F. candida that fed on a leaf-based diet from non-Bt rice plants.
Stability and Bioactivity of Cry Proteins in the Leaf-Based Diets
According to ELISA measurements, the original concentrations of Cry1C and Cry2A in the Bt rice leaf-based diets were 1.55 ± 0.03 and 16.38 ± 0.28 μg/g DW diet, respectively. After a 2-day feeding exposure, the contents had significantly decreased to 1.44 ± 0.01 and 13.64 ± 0.26 μg/g diet for Cry1C and Cry2A, respectively (Student’s t-test; t = 4.1, df = 6, P = 0.006 for Cry1C, and t = 7.1, df = 6, P < 0.001 for Cry2A). No Cry protein was detected in the leaf-based diet made from non-Bt rice plant.
The sensitive-insect bioassay showed that the mortality of C. suppressalis larvae was 6.7 ± 3.3% when the larvae fed on a diet containing the extract from the non-Bt leaf-based diet for 7 days. The mortality was 90.0 ± 5.8% or 73.3 ± 3.3% when C. suppressalis larvae fed on diets containing the extract from a fresh Cry1C leaf-based diet (T1C-19) or from a Cry1C leaf-based diet that had been exposed to F. candida for 2 days. The mortality was 90.0 ± 5.8% or 80.0 ± 5.8% when C. suppressalis larvae were fed diets containing the extract from the fresh Cry2A leaf-based diet (T2A-1) or a Cry2A leaf-based diet that had been exposed to F. candida for 2 days. Mortalities were not significantly different for larvae that fed on fresh vs. 2-day-old diet (Chi-square test; U = 2.78, df = 1, P = 0.10 for Cry1C diet; U = 1.18, df = 1, P = 0.28 for Cry2A diet).
No Effects on Enzyme Activities in F. candida by Feeding Bt Rice Pollen or Leaf Tissue
The activity of the four enzymes did not significantly differ in F. candida that fed on diets containing Bt pollen vs. non-Bt pollen (Figure 3) or Bt leaf powder vs. non-Bt leaf powder (Figure 4) (RM-ANOVA, all P > 0.05). In contrast, the activities of the four enzymes were significantly higher (P ≤ 0.003) in F. candida that fed on the non-Bt rice pollen containing E-64 rather than on non-Bt rice pollen without E-64 on all test days for POD, on days 14 and 21 for SOD, on days 7 and 21 for GR, and on day 21 for GST (Figure 3). The enzyme activities were also significantly higher (P ≤ 0.009) in F. candida that fed on the non-Bt rice leaf-based diet containing E-64 rather than on the same diet without E-64 on all test days for SOD, on days 7 and 21 for POD, and on day 21 for GR and GST (Figure 4).
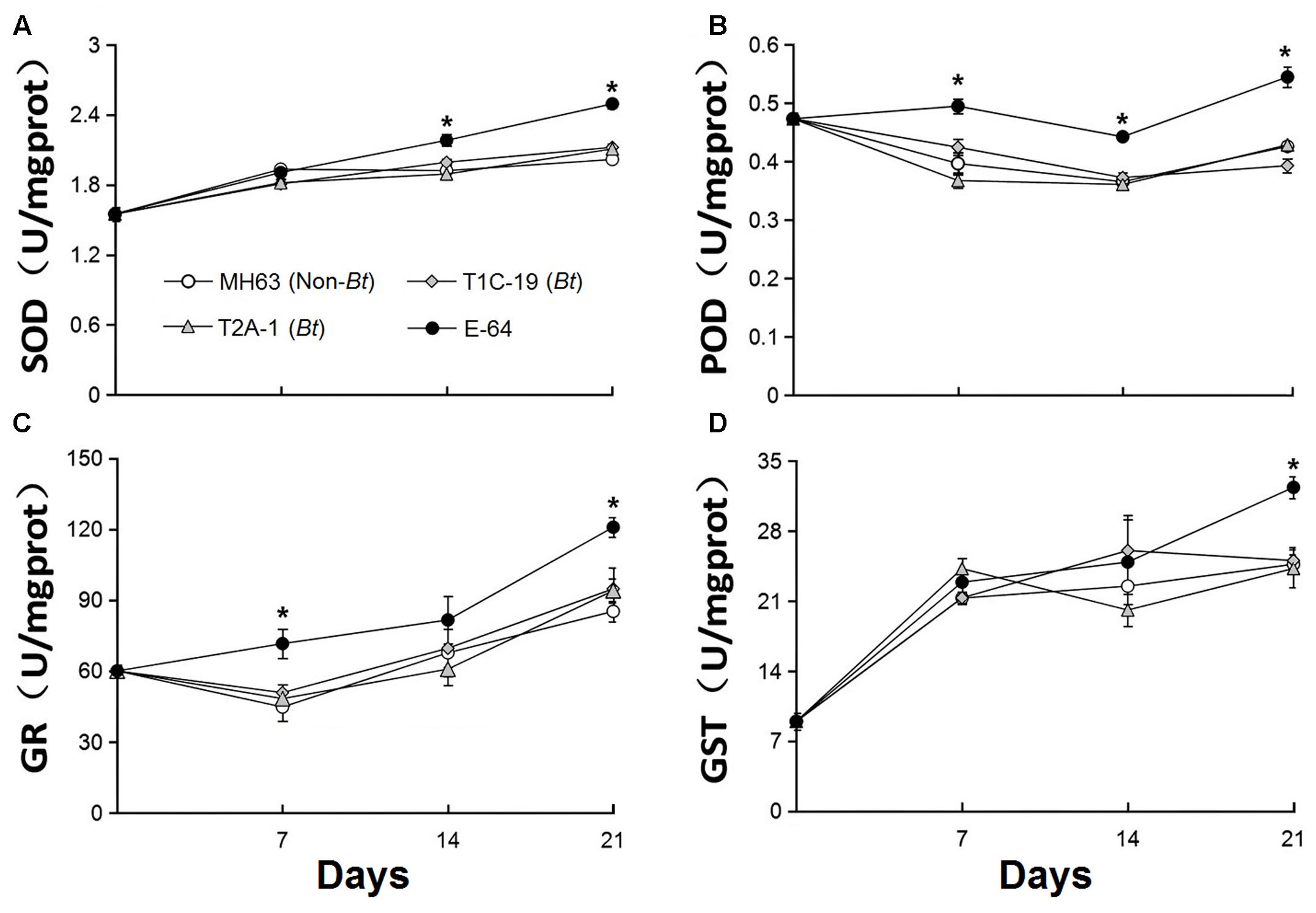
FIGURE 3. Enzyme activities (A: peroxidase, POD; B: superoxide dismutase, SOD; C: glutathione, GR; and D: glutathione S-transferase, GST) in F. candida that fed on a diet consisting of non-Bt rice pollen (MH63, negative control), Bt rice pollen containing Cry1C or Cry2A (T1C-19 or T2A-1), or non-Bt rice pollen containing E-64 protein (positive control) for 35 days. Values are means ± SE, n = 4. Asterisks indicate a significant difference between the treatment and the negative control (MH63) (P < 0.05).
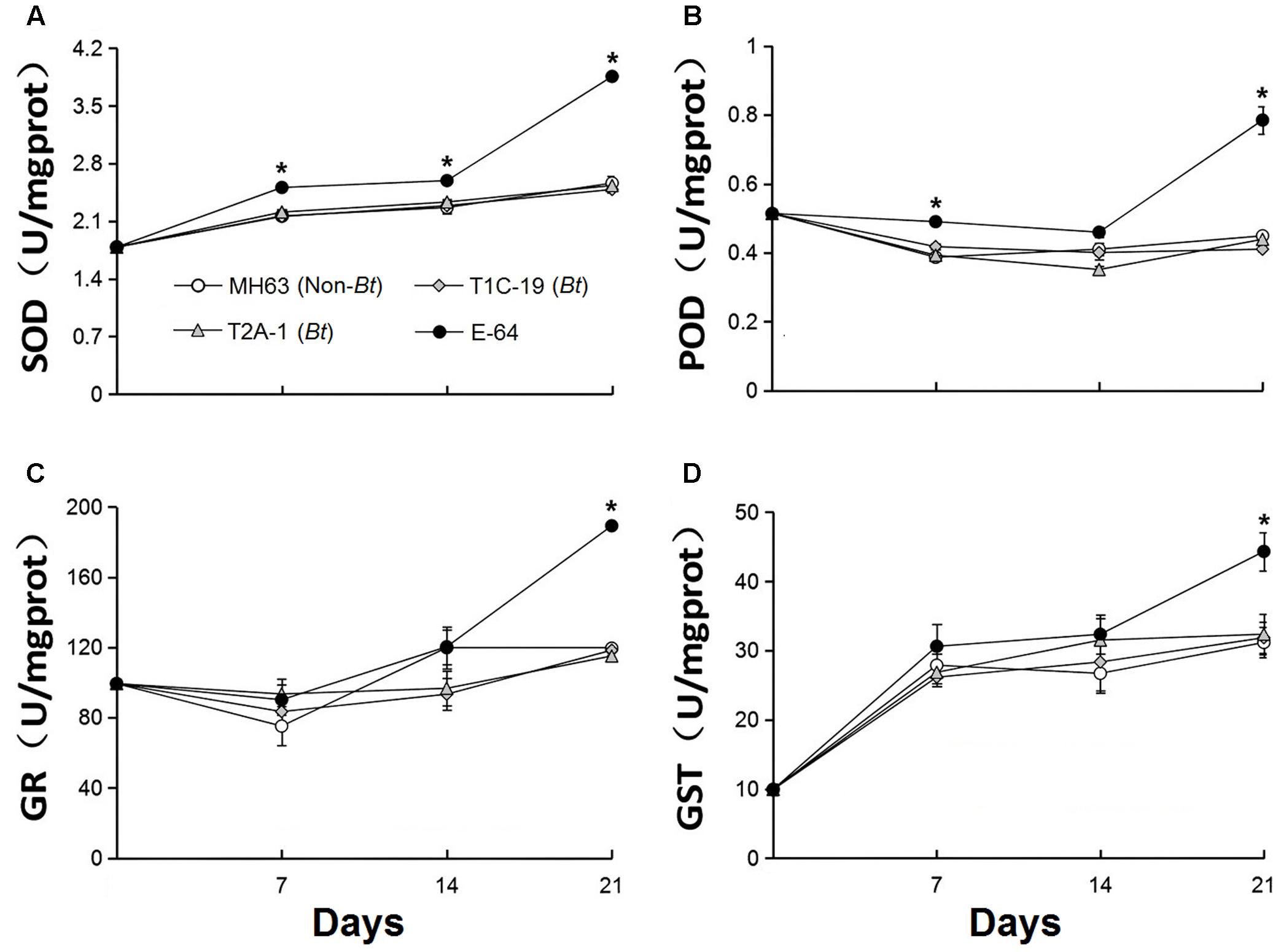
FIGURE 4. Enzyme activities (A: POD, B: SOD, C: GR, and D: GST) in F. candida that fed on non-Bt rice leaves (MH63, negative control), Bt rice leaves containing Cry1C or Cry2A (T1C-19 or T2A-1), or non-Bt leaves containing E-64 protein (E-64, positive control) for 35 days. Rice leaf powder was mixed with baker’s yeast (10:1). Values are means ± SE, n = 4. Asterisks indicate a significant difference between the treatment and the negative control (MH63) (P < 0.05).
Discussion
Pollen grains contain multiple organic and inorganic nutrients, such as sugars, starch, amino acids, proteins, lipids, vitamins, and minerals, and can serve as a food source for many arthropods (Li et al., 2010). Plant pollen has therefore been commonly used in dietary exposure assays with bees, lacewings, and ladybird beetles; such assays are essential components of non-target risk assessment of insect-resistant GE crops (Li et al., 2008, 2015; Wang et al., 2012, 2017; Meissle et al., 2014; Zhang et al., 2014). The current study shows that F. candida can survive, develop, and reproduce using rice pollen as a sole food source. The results are consistent with our previous study, in which F. candida survived well on only maize pollen (Zhang et al., 2017). In contrast, previous studies found that potato, cotton, and wheat leaves alone are not suitable foods for F. candida (Yu et al., 1997; Romeis et al., 2003; Bakonyi et al., 2006). Similarly, we found that the fitness of F. candida was significantly reduced when the collembolan fed on rice leaf powder alone (Yang et al., unpublished data). Based on our preliminary results, we developed a rice leaf-based diet in which baker’s yeast was mixed with lyophilized leaf powder at a ratio of 1:10. The results indicate that when fed this diet formula, F. candida survival rates ≥88%, which meets the standard for such dietary exposure assays (Romeis et al., 2011).
Dietary exposure assays require an appropriate positive control to confirm that the assay is sensitive, i.e., to confirm that the assay can detect the toxic effects of a test compound (Li et al., 2014a). In the current study, E-64 was used as a positive control because it is readily accepted by F. candida and is known to be toxic to the collembolan (Zhang et al., 2017). Our feeding experiments showed that ingestion of Bt rice pollen or leaf powder from T1C-19 or T2A-1 rice plants did not reduce the survival, development, reproduction, or the intrinsic rate of natural increase (rm) of F. candida. We found, however, that all of these life table parameters, except for the egg hatching rate, were significantly reduced by the consumption of the pollen or leaf-based diet containing E-64. This result demonstrates that the dietary exposure assays developed in our study were able to detect negative effects, and that they are therefore valid for assessing the effects of Bt rice pollen or leaf powder on F. candida. The results from the feeding bioassays thus indicate that consumption of Bt rice pollen or leaf powder has no adverse effects on F. candida individuals or populations. That E-64 did not reduce the egg hatching rate of F. candida that fed on the compound indicates that egg hatching is not a sensitive life-table parameter for assessing chemical toxicity to F. candida.
In addition to the life-table parameters mentioned above, the activities of two antioxidant-related enzymes, SOD and POD, and two detoxification-related enzymes, GR and GST, were measured, because they are known to be involved in the detoxification of reactive oxygen species (ROS) (Felton and Summers, 1995; Hayes and Strange, 2000). ROS might be induced when insects ingest toxic substances, and high levels of ROS may seriously damage the insects (Felton and Summers, 1995). It follows that an increase in the activity of these enzymes in insects may represent a response to the ingestion of a toxic, ROS-inducing compound. For these reasons, the activities of SOD, POD, GR, and GST have been widely used as indicators of the toxicity of Bt proteins and other insecticidal compounds (Bai et al., 2011; Yuan et al., 2011; Yang et al., 2015; Zhang et al., 2017). In the current study, the activities of SOD, POD, GR, and GST in F. candida were not affected by feeding on Bt rice pollen or leaf-based diets containing Cry1C or Cry2A protein. These results are consistent with previous studies. For example, the activities of SOD and POD in F. candida were not affected when the collembolan fed on the yeast mixed with Cry1Ab and Cry1Ac proteins (Yuan et al., 2011). Bai et al. (2011) found that SOD activity was not significantly altered in F. candida after ingestion of Cry1Ab-containing rice tissue for 35 days (Bai et al., 2011). Yang et al. (2015) showed that ingestion of pure Cry1C or Cry2A protein did not affect the activities of six enzymes in F. candida including antioxidant enzymes (SOD and POD), detoxification enzymes (GR and CES), and the proteases (T-Pro and TPS). More recently, Zhang et al. (2017) reported that SOD and POD were not influenced in F. candida that fed on the Bt corn pollen containing Cry1Ab/2Aj protein. In both Yang et al. (2015) and Zhang et al. (2017), the activities of these enzymes were significantly increased when F. candida ingested diets containing E-64, indicating that the assays used were able to detect toxic dietary effects. The lack of effects of consumption of Bt rice materials on life-table parameters further indicates that F. candida is not affected by Cry1C and Cry2A.
To quantify the exposure of F. candida to Cry protein in the feeding experiments, we measured the stability of the Cry proteins in the diets and the uptake of the proteins by F. candida. The results showed that the concentrations of Cry1C and Cry2A proteins in both pollen and leaf-based diets declined significantly during the feeding period, but that >60% of the Cry proteins was still detectable after a 2-day feeding exposure. The ingestion of Cry2A and Cry1C proteins by F. candida in the experiments was also confirmed by ELISA. In general, the contents of Cry proteins in F. candida increased over time with continually feeding on Bt pollen or leaf-based diets, which may be due to increased food consumption with F. candida growth. Furthermore, the bioactivity of the Cry proteins in the pollen or leaf-based diets was confirmed in a bioassay with Bt protein-sensitive C. suppressalis larvae. These results demonstrate that F. candida larvae ingested bioactive Cry1C and Cry2A protein in our feeding experiments. Given that collembolans are soil organisms with a broad range of food (Ponge, 1991), the F. candida in our study, which were exclusively fed Bt rice material, were exposed to Cry proteins at levels much higher than would occur under field conditions. That no lethal or sublethal effects were detected under our worst-case exposure conditions demonstrates that F. candida is not sensitive to Cry1C or Cry2A proteins in Bt rice pollen and leaves. Our results also provide evidence that the genetic engineering of the rice plants has not resulted in any unintended or unexpected changes in rice that affect F. candida (Gong and Wang, 2013; Ladics et al., 2015; Schnell et al., 2015; Devos et al., 2016).
As a surrogate collembolan species, F. candida has been commonly used in non-target risk assessment of insecticidal GE plants including cotton, potato, wheat, maize, and rice (Yu et al., 1997; Romeis et al., 2003; Bitzer et al., 2005; Clark and Coats, 2006; Bai et al., 2011; Bakonyi et al., 2011; Yuan et al., 2011, 2013; Zhang et al., 2017). Most studies have reported that ingestion of Bt proteins or Bt protein-containing plant tissues did not have any adverse effects. Two exceptions are the studies by Bakonyi et al. (2006, 2011), in which F. candida produced significantly fewer fecal pellets after consuming powder from Bt (Cry1Ab) maize leaves rather than powder from non-Bt leaves. The reasons for this effect, however, were not elucidated. In summary, the available data with F. candida suggest that the currently used Bt Cry1, Cry2, and Cry3 proteins are not toxic to collembolans.
To our knowledge, the current report is the first to assess the potential effects of Bt rice pollen or leaves containing Cry1C or Cry2A proteins on F. candida. The results from our toxicological and biochemical experiments confirmed that Cry1C and Cry2A are not toxic to F. candida. The results also indicated the absence of unintended effects on the collembolan caused by any change in plant tissue nutritional composition due to foreign gene transformation. We therefore conclude that the planting of the Bt rice lines will pose a negligible risk to F. candida.
Author Contributions
YL designed the study. BZ and YY performed all of the experiments. YY, YL, BZ, JR, and YP analyzed the data and wrote the manuscript. XZ and YP provided the experimental materials. All authors have read and approved the manuscript for publication.
Funding
This study was supported by the National GMO New Variety Breeding Program of PRC (2016ZX08011-001).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge Bruce Jaffee (http://jaffeerevises.com) for critical comments and language editing on an earlier version of the manuscript.
References
Al-Deeb, M. A., Wilde, G. E., Blair, J. M., and Todd, T. C. (2003). Effect of Bt corn for corn rootworm control on nontarget soil microarthropods and nematodes. Environ. Entomol. 32, 859–865. doi: 10.1603/0046-225X-32.4.859
Bai, Y., Yan, R., Ke, X., Ye, G., Huang, F., Luo, Y., et al. (2011). Effects of transgenic Bt rice on growth, reproduction, and superoxide dismutase activity of Folsomia candida (Collembola: Isotomidae) in laboratory studies. J. Econ. Entomol. 104, 1892–1899. doi: 10.1603/EC11095
Bai, Y., Yan, R., Ye, G., Huang, F., and Cheng, J. (2010). Effects of transgenic rice expressing Bacillus thuringiensis Cry1Ab protein on ground-dwelling collembolan community in postharvest seasons. Environ. Entomol. 39, 243–251. doi: 10.1603/EN09149
Bakonyi, G., Dolezsai, A., Mátrai, N., and Székács, A. (2011). Effects of consumption of Bt-maize (MON 810) on the collembolan Folsomia candida, over multiple generations: a laboratory study. Insects 2, 243–252. doi: 10.3390/insects2020243
Bakonyi, G., Szira, F., Kiss, I., Villányi, I., Seres, A., and Szekacs, A. (2006). Preference tests with Collembolas on isogenic and Bt-maize. Eur. J. Soil Biol. 42, S132–S135. doi: 10.1016/j.ejsobi.2006.06.005
Bitzer, R. J., Buckelew, L. D., and Pedigo, L. P. (2002). Effects of transgenic herbicide-resistant soybean varieties and systems on surface-active springtails (Entognatha: Collembola). Environ. Entomol. 31, 449–461. doi: 10.1603/0046-225X-31.3.449
Bitzer, R. J., Rice, M. E., Pilcher, C. D., Pilcher, C. L., and Lam, W. K. F. (2005). Biodiversity and community structure of epedaphic and euedaphic springtails (Collembola) in transgenic rootworm Bt corn. Environ. Entomol. 34, 1346–1376. doi: 10.1093/ee/34.5.1346
Clark, B. W., and Coats, J. R. (2006). Subacute effects of CrylAb Bt corn litter on the earthworm Eisenia fetida and the springtail Folsomia candida. Environ. Entomol. 35, 1121–1129. doi: 10.1603/0046-225X-35.4.1121
Devos, Y., Álvarez-Alfageme, F., Gennaro, A., and Mestdagh, S. (2016). Assessment of unanticipated unintended effects of genetically modified plants on non-target organisms: a controversy worthy of pursuit? J. Appl. Entomol. 140, 1–10. doi: 10.1111/jen.12248
Felton, G. W., and Summers, C. B. (1995). Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 29, 187–197. doi: 10.1002/arch.940290208
Fountain, M. T., and Hopkin, S. P. (2005). Folsomia candida (Collembola): a “standart” soil arthropod. Annu. Rev. Entomol. 50, 201–222. doi: 10.1146/annurev.ento.50.071803.130331
Gong, C., and Wang, T. (2013). Proteomic evaluation of genetically modified crops: current status and challenges. Front. Plant Sci. 4:41. doi: 10.3389/fpls.2013.00041
Han, Y., Ma, F., Nawaz, M., Wang, Y., Cai, W., Zhao, J., et al. (2017). The tiered-evaluation of the effects of transgenic cry1c rice on Cyrtorhinus lividipennis, a main predator of Nilaparvata lugens. Sci. Rep. 7:42572. doi: 10.1038/srep42572
Han, Y., Meng, J., Chen, J., Cai, W., Wang, Y., Zhao, J., et al. (2014). Bt rice expressing Cry2Aa does not harm Cyrtorhinus lividipennis, a main predator of the nontarget herbivore Nilapavarta lugens. PLOS ONE 9:e112315. doi: 10.1371/journal.pone.0112315
Han, Y., Wang, H., Chen, J., Cai, W., and Hua, H. (2015). No impact of transgenic cry2Aa rice on Anagrus nilaparvatae, an egg parasitoid of Nilaparvata lugens, in laboratory tests. Biol. Control 82, 46–51. doi: 10.1016/j.biocontrol.2014.12.005
Han, Y., Xu, X. L., Yuan, B., Wang, H., Liu, F., Wang, M., et al. (2011). The influence of transgenic cry1Ab/cry1Ac, cry1C and cry2A rice on non-target planthoppers and their main predators under field conditions. Agric. Sci. China 10, 1739–1747. doi: 10.1007/s11427-011-4234-2
Hayes, J. D., and Strange, R. C. (2000). Glutathione S-transferase polymorphisms and their biological consequences. Pharmacology 61, 154–166. doi: 10.1159/000028396
Ladics, G. S., Bartholomaeus, A., Bregitzer, P., Doerrer, N. G., Gray, A., Holzhauser, T., et al. (2015). Genetic basis and detection of unintended effects in genetically modified crop plants. Transgenic Res. 24, 587–603. doi: 10.1007/s11248-015-9867-7
Li, Y., Chen, X., Hu, L., Romeis, J., and Peng, Y. (2014a). Bt rice producing Cry1C protein does not have direct detrimental effects on the green lacewing Chrysoperla sinica (Tjeder). Environ. Toxicol. Chem. 33, 1391–1397. doi: 10.1002/etc.2567
Li, Y., Hallerman, E. M., Liu, Q., Wu, K., and Peng, Y. (2016). The development and status of Bt rice in China. Plant Biotechnol. J. 14, 839–848. doi: 10.1111/pbi.12464
Li, Y., Hu, L., Romeis, J., Wang, Y., Han, L., Chen, X., et al. (2014b). Use of an artificial diet system to study the toxicity of gut-active insecticidal compounds on larvae of the green lacewing Chrysoperla sinica. Biol. Control 69, 45–51.
Li, Y., Meissle, M., and Romeis, J. (2008). Consumption of Bt maize pollen expressing Cry1Ab or Cry3Bb1 does not harm adult green lacewings, Chrysoperla carnea (Neuroptera: Chrysopidae). PLOS ONE 3:e2909. doi: 10.1371/journal.pone.0002909
Li, Y., Meissle, M., and Romeis, J. (2010). Use of maize pollen by adult Chrysoperla carnea (Neuroptera: Chrysopidae) and fate of Cry proteins in Bt-transgenic varieties. J. Insect Physiol. 56, 157–164. doi: 10.1016/j.jinsphys.2009.09.011
Li, Y., Zhang, Q., Liu, Q., Meissle, M., Yang, Y., Wang, Y., et al. (2017). Bt rice in China—focusing the nontarget risk assessment. Plant Biotechnol. J. 15, 1340–1345. doi: 10.1111/pbi.12720
Li, Y., Zhang, X., Chen, X., Romeis, J., Yin, X., and Peng, Y. (2015). Consumption of Bt rice pollen containing Cry1C or Cry2A does not pose a risk to Propylea japonica (Thunberg) (Coleoptera: Coccinellidae). Sci. Rep. 5:7679. doi: 10.1038/srep07679
Meissle, M., Zünd, J., Waldburger, M., and Romeis, J. (2014). Development of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) on pollen from Bt-transgenic and conventional maize. Sci. Rep. 4:5900. doi: 10.1038/srep05900
Meng, J., Mabubu, J. I., Han, Y., He, Y., Zhao, J., Hua, H., et al. (2016). No impact of transgenic cry1C rice on the rove beetle Paederus fuscipes, a generalist predator of brown planthopper Nilaparvata lugens. Sci. Rep. 6:30303. doi: 10.1038/srep30303
National Academies of Sciences Engineering Medicine [NASEM] (2016). Genetically Engineered Crops: Experiences and Prospects. Washington, DC: National Academies Press.
Organisation for Economic Cooperation and Development [OECD] (2016). Test No. 232: Collembolan Reproduction Test in Soil. Paris: OECD Publishing, doi: 10.1787/9789264264601-en
Ponge, J. F. (1991). Food resources and diets of soil animals in a small area of Scots pine litter. Geoderma 49, 33–62. doi: 10.1016/0016-7061(91)90090-G
Romeis, J., Bartsch, D., Bigler, F., Candolfi, M. P., Gielkens, M. M., Hartley, S. E., et al. (2008). Assessment of risk of insect-resistant transgenic crops to nontarget arthropods. Nat. Biotechnol. 26, 203–208. doi: 10.1038/nbt1381
Romeis, J., Battini, M., and Bigler, F. (2003). Transgenic wheat with enhanced fungal resistance causes no effects on Folsomia candida (Collembola: Isotomidae). Pedobiologia 47, 141–147. doi: 10.1078/0031-4056-00177
Romeis, J., Hellmich, R. L., Candolfi, M. P., Carstens, K., Schrijver, A. D., Gatehouse, A. M. R., et al. (2011). Recommendations for the design of laboratory studies on non-target arthropods for risk assessment of genetically engineered plants. Transgenic Res. 20, 1–22. doi: 10.1007/s11248-010-9446-x
Romeis, J., Raybould, A., Bigler, F., Candolfi, M. P., Hellmich, R. L., Huesing, J. E., et al. (2013). Deriving criteria to select arthropod species for laboratory tests to assess the ecological risks from cultivating arthropod-resistant genetically engineered crops. Chemosphere 90, 901–909. doi: 10.1016/j.chemosphere.2012.09.035
Schnell, J., Steele, M., Bean, J., Neuspiel, M., Girard, C., Dormann, N., et al. (2015). A comparative analysis of insertional effects in genetically engineered plants: considerations for pre-market assessments. Transgenic Res. 24, 1–17. doi: 10.1007/s11248-014-9843-7
Snider, R. (1973). Laboratory observations on the biology of Folsomia candida (Willem) (Collembola: Isotomidae). Rev. Ecol. Biol. Sol. 10, 103–124.
Tian, J., Romeis, J., Liu, K., Zhang, F., Zheng, X., Xu, H., et al. (2017). Assessing the effects of Cry1C rice and Cry2A rice to Pseudogonatopus flavifemur, a parasitoid of rice planthoppers. Sci. Rep. 7:7838. doi: 10.1038/s41598-017-08173-w
Tian, J. C., Liu, Z. C., Chen, M., Chen, Y., Chen, X. X., Peng, Y. F., et al. (2010). Laboratory and field assessments of prey-mediated effects of transgenic Bt rice on Ummeliata insecticeps (Araneida: Linyphiidae). Environ. Entomol. 39, 1369–1377. doi: 10.1603/EN10003
Wang, Y., Dai, P., Chen, X., Romeis, J., Shi, J., Peng, Y., et al. (2017). Ingestion of Bt rice pollen does not reduce the survival or hypopharyngeal gland development of Apis mellifera adults. Environ. Toxicol. Chem. 36, 1243–1248. doi: 10.1002/etc.3647
Wang, Y., Ke, K., Li, Y., Han, L., Liu, Y., Hua, H., et al. (2016). Comparison of three transgenic Bt rice lines for insecticidal protein expression and resistance against a target pest, Chilo suppressalis (Lepidoptera: Crambidae). Insect Sci. 23, 78–87. doi: 10.1111/1744-7917.12178
Wang, Y., Li, Y., Huang, Z., Chen, X., Romeis, J., Dai, P., et al. (2015). Toxicological, biochemical, and histopathological analyses demonstrating that Cry1C and Cry2A are not toxic to larvae of the honeybee, Apis mellifera. J. Agric. Food Chem. 63, 6126–6132. doi: 10.1021/acs.jafc.5b01662
Wang, Y., Li, Y., Romeis, J., Chen, X., Zhang, J., Chen, H., et al. (2012). Consumption of Bt rice pollen expressing Cry2Aa does not cause adverse effects on adult Chrysoperla sinica Tjeder (Neuroptera: Chrysopidae). Biol. Control 61, 246–251. doi: 10.1016/j.biocontrol.2012.02.011
Yang, Y., Chen, X., Cheng, L., Cao, F., Romeis, J., Li, Y., et al. (2015). Toxicological and biochemical analyses demonstrate no toxic effect of Cry1C and Cry2A to Folsomia candida. Sci. Rep. 5:15619. doi: 10.1038/srep15619
Yang, Y., Liu, Y., Cao, F., Chen, X., Cheng, L., Romeis, J., et al. (2014). Consumption of Bt rice pollen containing Cry1C or Cry2A protein poses a low to negligible risk to the silkworm Bombyx mori (Lepidoptera: Bombyxidae). PLOS ONE 9:e102302. doi: 10.1371/journal.pone.0102302
Yu, L., Berry, R. E., and Croft, B. A. (1997). Effects of Bacillus thuringiensis toxins in transgenic cotton and potato on Folsomia candida (collembola: Isotomidae) and Oppianitens (Acari: Orbatidae). J. Econ. Entomol. 90, 113–118. doi: 10.1093/jee/90.1.113
Yuan, Y., Ke, X., Chen, F., Krogh, P. H., and Ge, F. (2011). Decrease in catalase activity of Folsomia candida fed a Bt rice diet. Environ. Pollut. 159, 3714–3720. doi: 10.1016/j.envpol.2011.07.015
Yuan, Y., Krogh, P. H., Bai, X., Roelofs, D., Chen, F., Zhu-Salzman, K., et al. (2014). Microarray detection and qPCR screening of potential biomarkers of Folsomia candida (Collembola: Isotomidae) exposed to Bt proteins (Cry1Ab and Cry1Ac). Environ. Pollut. 184, 170–178. doi: 10.1016/j.envpol.2013.08.014
Yuan, Y., Xiao, N., Krogh, P. H., Chen, F., and Ge, F. (2013). Laboratory assessment of the impacts of transgenic Bt rice on the ecological fitness of the soil non-target arthropod, Folsomia candida (Collembola: Isotomidae). Transgenic Res. 22, 791–803. doi: 10.1007/s11248-013-9687-6
Zhang, B., Yang, Y., Zhou, X., Shen, P., Peng, Y., and Li, Y. (2017). A laboratory assessment of the potential effect of Cry1Ab/Cry2Aj-containing Bt maize pollen on Folsomia candida by toxicological and biochemical analyses. Environ. Pollut. 222, 94–100. doi: 10.1016/j.envpol.2016.12.079
Keywords: Bt rice pollen, Bt rice leaf, environmental risk assessment, ELISA, non-target effects, enzyme activity
Citation: Yang Y, Zhang B, Zhou X, Romeis J, Peng Y and Li Y (2018) Toxicological and Biochemical Analyses Demonstrate the Absence of Lethal or Sublethal Effects of cry1C- or cry2A-Expressing Bt Rice on the Collembolan Folsomia candida. Front. Plant Sci. 9:131. doi: 10.3389/fpls.2018.00131
Received: 06 August 2017; Accepted: 23 January 2018;
Published: 06 February 2018.
Edited by:
James Lloyd, Stellenbosch University, South AfricaReviewed by:
Abdul Rasheed War, World Vegetable Center South Asia, IndiaKi-Hong Jung, Kyung Hee University, South Korea
Mark Laing, University of KwaZulu-Natal, South Africa
Monica Garcia-Alonso, Estel Consult Ltd., United Kingdom
Copyright © 2018 Yang, Zhang, Zhou, Romeis, Peng and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yunhe Li, bGl5dW5oZUBjYWFzLmNu
† These authors have contributed equally to this work.