 Zhenhua Wei
Zhenhua Wei Taisheng Du
Taisheng Du Xiangnan Li
Xiangnan Li Liang Fang
Liang Fang Fulai Liu
Fulai Liu- 1Center for Agricultural Water Research in China, China Agricultural University, Beijing, China
- 2Department of Plant and Environmental Sciences, Faculty of Science, University of Copenhagen, Copenhagen, Denmark
- 3Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, Changchun, China
The interactive effects of CO2 elevation, N fertilization, and reduced irrigation regimes on fruit yield (FY) and quality in tomato (Solanum lycopersicum L.) were investigated in a split-root pot experiment. The plants were grown in two separate climate-controlled greenhouse cells at atmospheric [CO2] of 400 and 800 ppm, respectively. In each cell, the plants were fertilized at either 100 or 200 mg N kg-1 soil and were either irrigated to full water holding capacity [i.e., a volumetric soil water content of 18%; full irrigation (FI)], or using 70% water of FI to the whole pot [deficit irrigation (DI)] or alternately to only half of the pot [partial root-zone irrigation (PRI)]. The yield and fruit quality attributes mainly from sugars (sucrose, fructose, and glucose) and organic acids (OAs; citric acid and malic acid) to various ionic (NH4+, K+, Mg2+, Ca2+, NO3-, SO42-, and PO43-) concentrations in fruit juice were determined. The results indicated that lower N supply reduced fruit number and yield, whereas it enhanced some of the quality attributes of fruit as indicated by greater firmness and higher concentrations of sugars and OAs. Elevated [CO2] (e[CO2]) attenuated the negative influence of reduced irrigation (DI and PRI) on FY. Principal component analysis revealed that the reduced irrigation regimes, especially PRI, in combination with e[CO2] could synergistically improve the comprehensive quality of tomato fruits at high N supply. These findings provide useful knowledge for sustaining tomato FY and quality in a future drier and CO2-enriched environment.
Introduction
As a consequence of anthropogenic activities, the carbon dioxide concentration ([CO2]) in atmosphere has reached 400 ppm at present, and is projected to approximately double by the end of this century (Intergovernmental Panel on Climate Change [IPCC], 2013). The elevated [CO2] (e[CO2]) in the atmosphere contributing to global warming causes changes in precipitation patterns and results in water scarcity in many areas, particularly in arid and semi-arid regions, where agriculture accounts for >70% of freshwater withdrawals (Wang et al., 2010). Moreover, nitrogen (N) fertilization is an essential mineral nutrient, and inadequate N supply could have significant influence on yield and quality of horticultural crops (Sainju et al., 2003), including tomato (Solanum lycopersicum L.), which is one of the most popular fruit crops grown around the world. Therefore, a better understanding of tomato plant responses to e[CO2], N fertilization rate, and water availability is necessary for optimizing tomato fruit yield (FY) and quality in the future climate change scenarios.
Tomato is tasty and nutritional and constitutes an important source of minerals, vitamins, and antioxidants that are essential for human health (Sun et al., 2014). The quality attributes including fruit firmness (FM), minerals, concentrations of total soluble solids (TSS), sugars, and organic acids (OAs), as well as their ratio determine not only the sweetness and sourness but also the overall flavor intensity of tomato fruits (Jones and Scott, 1984). Apart from organic components, minerals are also key quality attributes in tomatoes. Mineral elements commonly found in tomato fruit with significant amounts are N, P, K, Ca, Mg, and S, which are essential for maximizing all aspects of fruit quality (Sainju et al., 2003).
In recent years, it is of great interest to exploit appropriate reduced irrigation management strategies for sustaining crop yield and improving fruit quality in drought prone regions. Alternate partial root-zone irrigation (PRI) and deficit irrigation (DI) are effective irrigation techniques that are being investigated in many regions (Davies et al., 2002; Du et al., 2015). DI is a method that irrigates the entire root zone with an amount of water less than the potential evapotranspiration and the mild stress that develops minimal effects on the yield (Dodd, 2009). PRI is a further refinement of DI and the principle behind PRI is to alternately allow one part of the root system to be irrigated to keep the leaves hydrated while the other part is exposed to soil drying, triggering stronger root-to-shoot abscisic acid (ABA) hormonal signaling, inducing partial stomatal closure (Kang and Zhang, 2004; Liu et al., 2006), better water status in plant (Xu et al., 2009; Sun et al., 2014), greater water, and N use efficiency (Wang et al., 2010). Moreover, accumulated evidence has demonstrated that, compared with the full irrigation (FI), both DI and PRI could save up to 25–50% irrigation water without reduction in yield (Wang et al., 2010; Barrios-Masias and Jackson, 2016; Wei et al., 2016), accompanied with several significantly improved fruit quality attributes, such as firmness, TSS, and sugar to acid ratio (SAR) (Davies et al., 2000; Zegbe et al., 2003, 2004, 2006; Campos et al., 2009; Patanè and Cosentino, 2010; Wang and Frei, 2011; Hou et al., 2017), as well as ionic concentrations in fruit juice of tomatoes (Sun et al., 2014).
In addition to water management, N nutrition is also known to be an important factor affecting plant growth, yield, and quality of fruit crops (Sainju et al., 2003). A low N supply could decrease the development of the foliar canopy, trigger changes in tomato secondary metabolism, and reduce photoassimilates available into fruit, hereby influencing the firmness, TSS, sugar, and acid concentrations (Huett and Dettmann, 1988; Domis et al., 2002; Wang et al., 2007; Bénard et al., 2009).
CO2 plays a crucial role in physiology of plants by affecting the leaf photosynthesis, plant growth, and crop yield. Previous studies have concluded that plant photosynthesis, stomatal aperture, biomass production, yield, and water use efficiency could be modulated by CO2 environment (Ainsworth and Long, 2005; Sanz-Sáez et al., 2010; Pazzagli et al., 2016). For instance, more carbohydrates could be transferred into fruits due to the increased photosynthesis in plants grown under CO2-enriched environment, which could enhance yield and increase the concentrations of starch, sugars, ascorbic acid, and OA (Islam et al., 1996; Bindi et al., 2001; Högy and Fangmeier, 2009; Moretti et al., 2010; Sun et al., 2012). Nevertheless, there is generally a reduction in mineral contents, particularly N concentrations, in cereals grown at e[CO2] (Li et al., 2016); most probably due to restricted root nutrient uptake (i.e., caused by reduced mass flow) and dilution effect (Loladze, 2002; Myers et al., 2014). However, until now the effect of e[CO2] on mineral concentrations in fruit crops has not been examined thoroughly (Loladze, 2014), and the combined effects of e[CO2], N fertilization, and reduced irrigation regimes on fruit quality in tomato remain largely elusive.
In this study, tomato plants were grown in two atmospheric [CO2] (400 and 800 ppm) combined with two N fertilization rates (100 and 200 mg kg-1 soil) and exposed to three different irrigation regimes (FI, DI, and PRI) during flowering to fruiting stages. The yield and quality attributes including firmness and concentrations of TSS, sugars, OAs, and several minerals in fruit juice were determined. It was hypothesized that e[CO2] would ameliorate the negative effects of reduced irrigation on tomato yield; on the other hand, it would cause reduced mineral nutrition, hence decreasing the fruit quality, while PRI could improve plant nutrient uptake and increase of N fertilization might further increase plant yield and sustain fruit quality at e[CO2].
Materials and Methods
Experimental Setup
The experiment was conducted in a climate-controlled greenhouse at the experimental farm of the Faculty of Science, University of Copenhagen, Taastrup, Denmark, from September 2016 to January 2017. Tomato seeds (Solanum lycopersicum L., cv. Elin) were sown on 26th September 2016. Elin is a common indeterminate tomato cultivar being widely grown in Denmark and Sweden. The seedlings were transplanted into 1.5 L pots filled with peat substance (Sphagnum, 32% organic matter, pH = 5.6–6.4, and EC = 0.45 mscm-1) at the fourth leaf stage. From sowing, half of the plants (24) were grown in a greenhouse cell with ambient CO2 concentration of 400 ppm (a[CO2]), and another half were grown in a cell with elevated CO2 concentration of 800 ppm (e[CO2]). In both greenhouse cells (50 m2 each), the [CO2] was sustained or achieved by emission of pure CO2 from a bottled tank, released in one point, and distributed evenly in the cells through internal ventilation. The [CO2] in the glasshouse cells was monitored every 6 s by a CO2 Transmitter Series GMT220 (Vaisala Group, Helsinki, Finland). The [CO2] concentration was kept almost constant in each cell during the whole treatment period and the data have been shown in Yan et al. (2017). The climatic conditions in the two glasshouse cells were set at: 23/16 ± 2 °C day/night air temperature, 60% relative humidity, 16 h photoperiod, and 500 μmol m-2s-1 photosynthetic active radiation supplied by sunlight plus LED lamps (Philips GreenPower LED Toplighting, Frederikskaj 6, 2450 København SV, Denmark).
Five weeks after sowing, tomato seedlings were transplanted into 10 L pots (17 cm diameter and 50 cm depth) in the greenhouse, filled with 14.5 kg of air-dried soil. All pots used were divided vertically into two equal-sized compartments with plastic sheets such that the water exchange between the two compartments was prevented. A piece of plastic (4 cm × 5 cm) was removed from the middle of the sheet where the tomato seedling was transplanted. Plant spacing was 0.4 m × 0.4 m resulting in six plants per m2. The soil used was classified as sandy loam, with a pH of 6.7, total C 10.3 g kg-1, total N 1.0 g kg-1, NH4+ 0.1 mg kg-1, and NO3- 5.3 mg kg-1. The soil was sieved through 5 mm mesh before filling the pots. The soil had a volumetric soil water content (% vol.) of 18.0% and 5.0% at pot water holding capacity and permanent wilting point, respectively.
Treatments
The experiment was conducted in two greenhouse cells, one with a[CO2] (400 ppm) and the other with e[CO2] (800 ppm). Two N (in the form of NH4NO3) fertilization rates, i.e., 100 (N1) and 200 (N2) mg kg-1 soil, were included and each was assigned to half of the plants in both of the greenhouse cells. The N fertilizer was mixed thoroughly with the soil before filling the pots. At the same time, 50 and 60 mg kg-1 soil P and K, respectively, were also applied as KH2PO4 into the soil to meet the nutrient requirements for plant growth.
The average soil water content in the pot was monitored by a time-domain reflectometer (TRASE; Soil Moisture Equipment Corporation, Santa Barbara, CA, United States) with two probes (35 cm in length) installed in the middle of each soil compartment, namely, four probes in each pot. The tomato plants were well-watered to full pot water holding capacity (i.e., a volumetric soil water content of 18%) to compensate for the evapotranspiration water loss during the previous day in the first 3 weeks after transplanting. The irrigation was done manually at 15:00 h daily. The plants were exposed to three irrigation treatments: (1) FI where both soil compartments were watered daily to a volumetric soil water content of 18%, the irrigation volume (IFI) was calculated as: IFI = 10 × (18% - θmean), where 10 is the soil volume of the whole pots and θmean is the mean soil water content of the two compartments; (2) alternative PRI, where half of the root system was watered to 70% of the FI irrigation volume while the other half was allowed to dry to ca. 6%, and then the irrigation was shifted between the two soil compartments; and (3) DI where the same amount of water for PRI was evenly irrigated to the two soil compartments. No leaching fraction was considered during the irrigation treatment.
The experiment was a complete randomized design with four replicated plants in each irrigation and N treatment at both a[CO2] and e[CO2]. Although the number of replicates for each treatment was limited, the controlled environmental conditions in the greenhouse cells were steady during experimental period, this had facilitated a similar fruit growth and development among the replicated plants for each treatment, and the difference between treatments was obvious. Thus, the data of the FY and quality should be reliable. The water used for the irrigation was tap water with negligible concentrations of nutrients. The irrigation treatments lasted 40 days and each soil compartment of the PRI plants had experienced five drying/wetting cycles.
Measurements
The tomato variety Elin is an indeterminate type. Therefore, after the fourth fruit trusses appearance, the plants were pruned by removing the apex to stop the vegetative growth. At first truss fruits reached red maturity, namely, 64 days after plant transplanting, all the fresh fruits from the four plants of each treatment were harvested and fruit number (FN) and yield (FY) (namely, the fresh weight of all harvested fruits) per plant were recorded. Single fruit weight (SFW) of each plant was calculated as the ratio of FY to FN.
Four ripen fruits per treatment, i.e., one fresh fruit per plant in each treatment at the firm red stage, were chosen for quality measurements. FM was measured on the middle of the pericarp and reading was recorded for each fruit with a FTX Fruit Tester (Wagner Instruments, Greenwich, CT, United States) (Sun et al., 2014). Thereafter, each fruit was cut into small pieces and homogenized thoroughly in a fruit blender. The homogenate was centrifuged at 4,000 × g for 5 min and the supernatant was filtered through a syringe filter (0.22 mm Acetate Cameo; Osmonics, Minnetonka, MN, United States). TSS concentration of the juice was then measured using a digital refractometer with automatic temperature compensation (RFM 90; Struers Ltd., Catcliffe Rotherham, United Kingdom) and was expressed in °Brix.
Fruit juice concentrations of fructose, glucose, sucrose, citric acid, malic acid, NH4+, K+, Mg2+, Ca2+, NO3-, SO42-, and PO43- were analyzed by ion chromatography (Metrohm AG, Herisau, Switzerland). The determination of fructose, glucose, and sucrose was done on a Metrosep Carb 1-150 column using 100 mM sodium hydroxide as eluent, while the measurement of citric and malic acid was done on a Carbohydrate H+ column using 0.5 mM sulfuric acid and 10% acetone as eluent. Total sugar (TS) concentration was calculated as the sum of fructose, glucose, and sucrose, and OA concentration was calculated as the sum of citric and malic acid. The SAR was also calculated. NH4+, K+, Mg2+, and Ca2+ concentrations were determined on a Metrosep C4-100 analytical column (4 mm × 125 mm, 1.7 mM nitric acid/0.7 mM dipicolinic acid eluent), while NO3-, SO42-, and PO43- concentrations were determined on a Metrosep A Supp 4 analytical column (4 mm × 125 mm, 1.8 mM Na2CO3/1.7 mM NaHCO3 eluent). Total cation concentration (TCN) was calculated as the sum of NH4+, K+, Mg2+, and Ca2+ concentration; total anion concentration (TAN) was calculated as the sum of NO3-, SO42-, and PO43- concentration; and total ionic concentration (TIN) was calculated as the sum of TCN and TAN. The above measurements were done for four samples of each treatment.
Statistical Analyses
Three-way analysis of variance was performed for the independent variables: CO2 concentration ([CO2]), N fertilization (N), and irrigation regime (IRRI), as well as for their interactions. Data were analyzed with SPSS version 18.0 (IBM Electronics). LSD’s multiple range test was applied to assess the differences between treatments at a significance level of 5%. Principal component analysis (PCA) approach was used to evaluate the comprehensive fruit quality from FM to TIN (15 parameters in total) as affected by the different treatments. The PCA method has been detailed in Wang et al. (2015).
Results
Soil Water Content
Changes of daily average volumetric soil water content in the 0–35 cm soil profile are illustrated in Figure 1. Irrespective of [CO2] environment, the soil water contents showed similar trends for the same IRRI and N treatment. In FI, soil water contents remained at an average value of ca. 18% during the 40 days after initiation of the irrigation treatment (DAT) under both N1 and N2. In DI, soil water contents declined considerably in the first 8 DAT and remained at an average value of 10 and 12% during the last 32 days under N1 and N2 treatments, respectively. In PRI, soil water contents were dependent on which side of the root system was being irrigated. Soil water contents in the irrigated soil compartment were above 12 and 16% under N1 and N2 treatments, respectively, while soil water contents in the dry side decreased sharply and were about 6% before the shifting of irrigation under both N1 and N2 treatments.
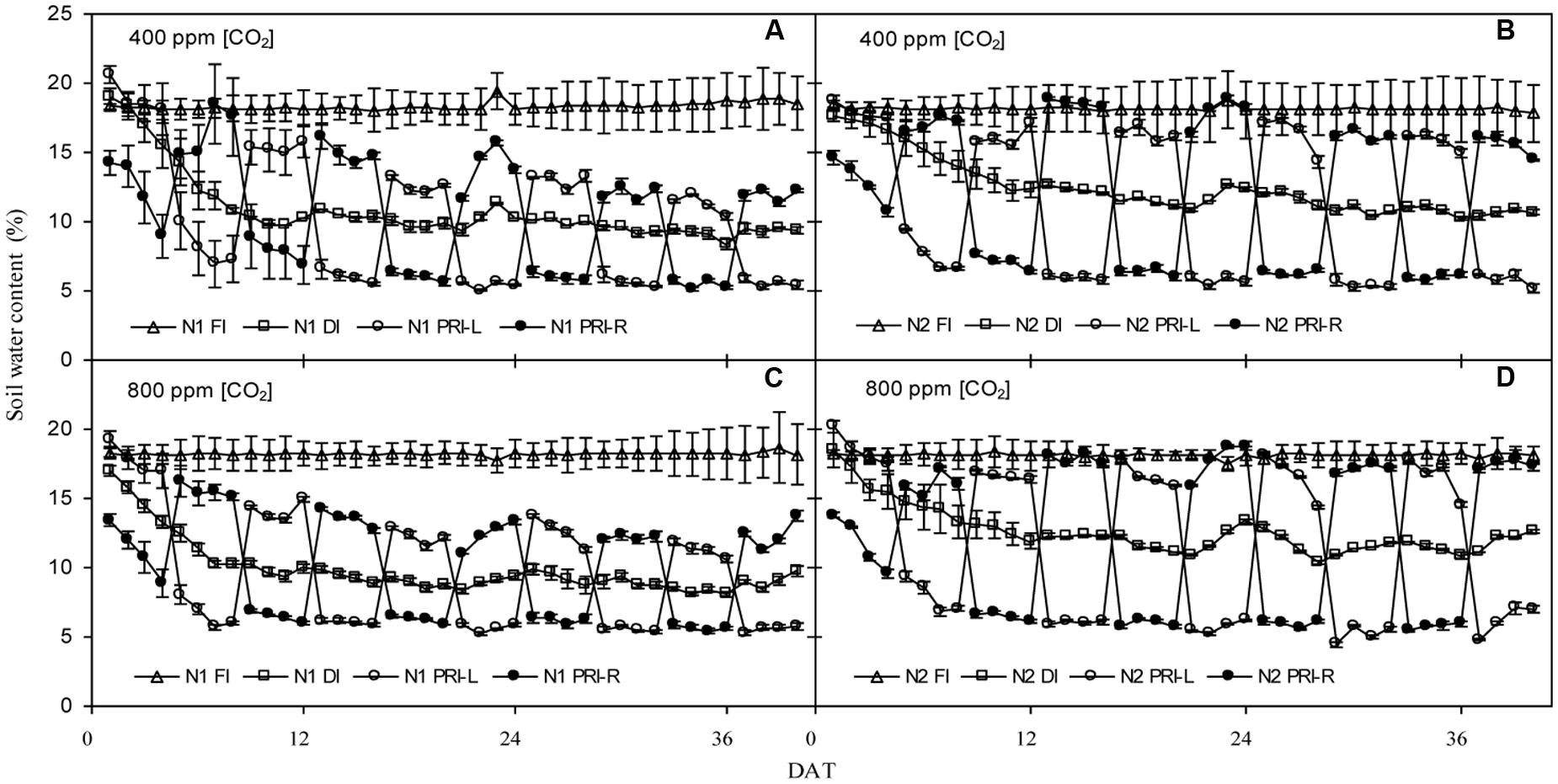
FIGURE 1. Daily average volumetric soil water content (%) in the pots of tomatoes exposed to three irrigation regimes (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRI). PRI-L and PRI-R represent the left and the right soil compartment of the PRI pots, respectively, combined with two N fertilization rates (N1 and N2) under atmospheric 400 ppm [CO2] (A,B) and with two N fertilization rates (N1 and N2) under atmospheric 800 ppm [CO2] (C,D). Error bars indicate standard error of the mean (n = 4).
Fruit Number, Single Fruit Weight, and Fresh Fruit Yield
Fruit number was only significantly affected by N (Table 1), and was higher in plants grown under N2 than N1 treatment. SFW was not affected by any of the [CO2], N, and IRRI treatments (Table 1). Fresh FY was affected significantly by [CO2], N, and IRRI (Table 1). N2 had higher FY than N1 plants. Under N2 treatment, DI and PRI plants grown in e[CO2] had greater FY than those grown in a[CO2] environment. Across N treatments, the FI plants had similar FY to DI and PRI plants at e[CO2].
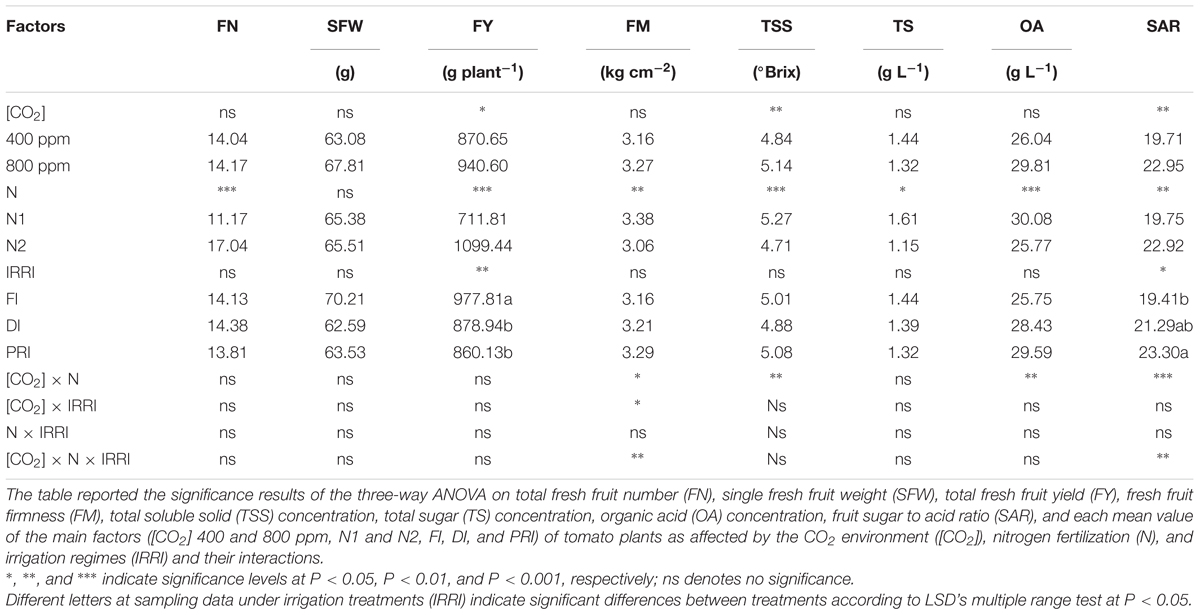
TABLE 1. Output of three-way analysis of variance (ANOVA) and mean value on measured yield and some fruit quality parameters of tomato plants as affected by the atmospheric [CO2] (400 and 800 ppm), N fertilization rates (N1 and N2), and irrigation regimes (full irrigation, FI; deficit irrigation, DI; and alternative partial root-zone irrigation, PRI).
Fruit Firmness, Total Soluble Solids, Total Sugars, Organic Acids, and Sugar to Acid Ratio
Fruit firmness was significantly affected by N, [CO2] × N, and [CO2] × IRRI, as well as [CO2] × N × IRRI (Figure 2A and Table 1). Plants grown with reduced irrigation possessed higher FM than those exposed to FI under N2 + e[CO2] treatment. TSS concentration was affected by [CO2] and N as well as [CO2] × N (Figure 2B and Table 1), being that plants grown at e[CO2] had the same and greater TSS than those grown at a[CO2] under N1 and N2 treatment, respectively. TS was only affected by N treatment (Table 1), and was greater under N1 than N2. Reduced irrigation did not decrease TS as compared with FI at e[CO2]. OA was significantly affected by N and [CO2] × N (Figure 2C and Table 1), and the N2 plants had significantly lower OA than N1 plants. SAR was significantly affected by [CO2], N, and IRRI as well as [CO2] × N and [CO2] × N × IRRI (Figure 2D and Table 1). Regardless of IRRI treatments, plants grown under e[CO2] + N1 had the highest SAR than plants exposed to other treatments. Most interestingly, DI and PRI plants possessed greater SAR than FI at e[CO2] + N2 treatment.
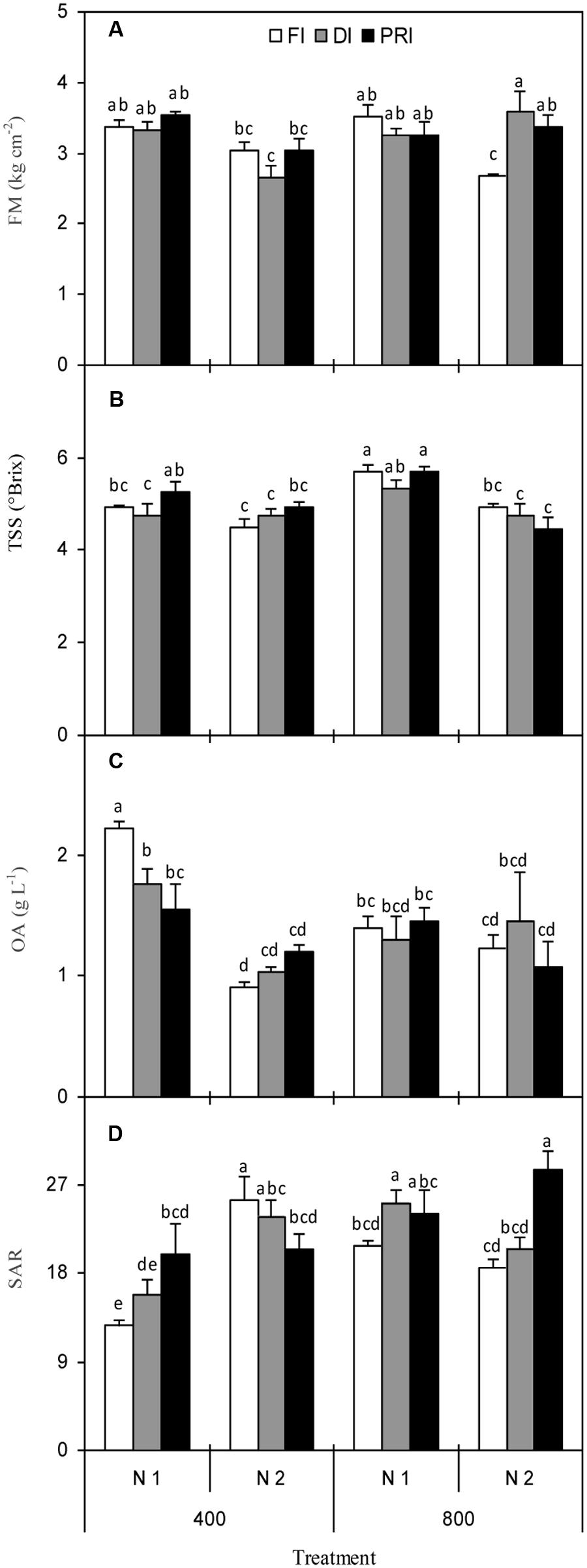
FIGURE 2. Fresh fruit firmness (FM) (A), total soluble solid (TSS) concentration (B), organic acid (OA) concentration (C), and sugar to acid ratio (SAR) (D) of tomato plants as affected by the atmospheric [CO2] (400 and 800 ppm), N fertilization rates (N1 and N2), and irrigation regimes (FI, DI, and alternative PRI). Error bars indicate standard error of the mean (n = 4). Different letters at each sampling data indicate significant differences between treatments according to LSD’s multiple range test at P < 0.05. Statistical comparisons among the treatments are presented in Table 1.
Cation Concentrations in Fruit Juice
NH4+ concentration in fruit juice was affected by N and [CO2] × N (Figure 3A and Table 1). Plants grown under N2 had significantly higher NH4+ than those grown under N1 treatment, especially under e[CO2] environment. K+ concentration was affected by [CO2] and [CO2] × N (Figure 3B and Table 2), and e[CO2] plants had significantly greater K+ than a[CO2] plants. Mg2+ concentration was remarkably affected by [CO2], [CO2] × N, and [CO2] × IRRI (Figure 3C and Table 2). Disregarding N treatment, DI and PRI plants grown under e[CO2] treatment possessed higher Mg2+ than those grown under a[CO2] treatment. Ca2+ concentration in fruit juice was only significantly affected by N treatment (Table 2), and was greater for N2 than N1 plants.
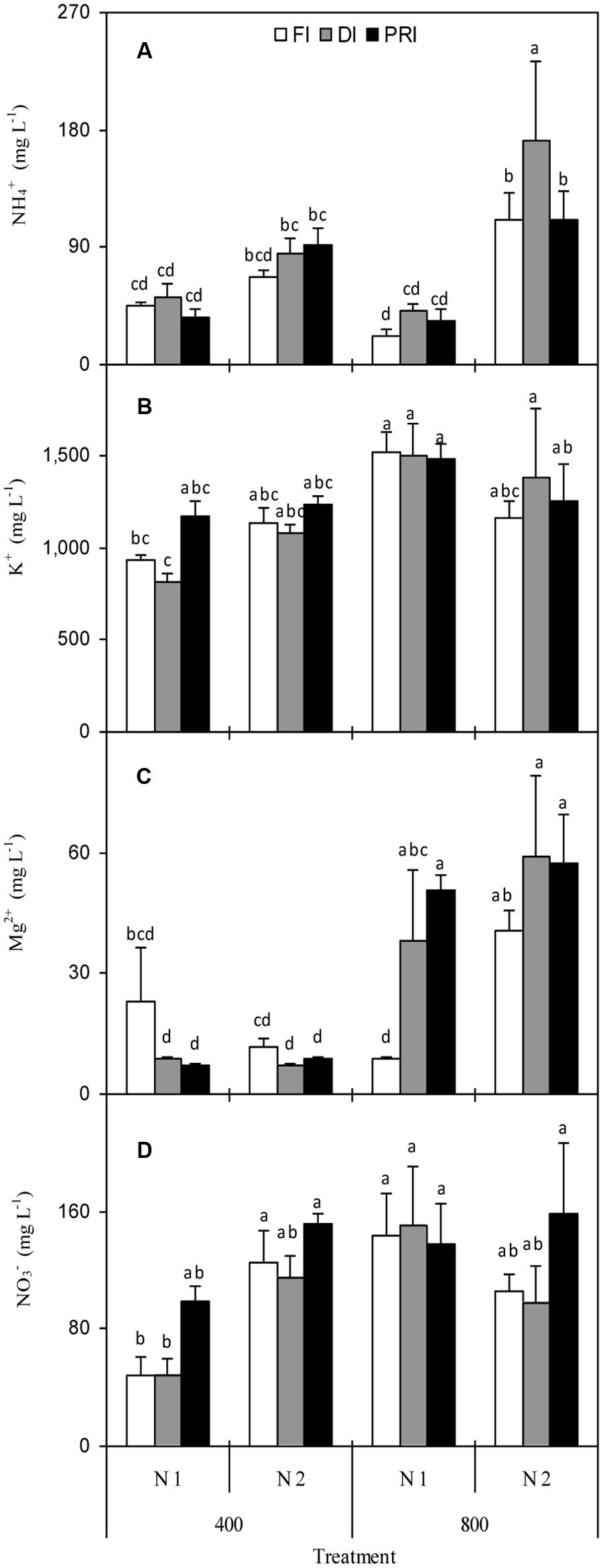
FIGURE 3. NH4+ (A), K+ (B), Mg2+ (C), and NO3- (D) concentration in fruit juice of tomato plants as affected by the atmospheric [CO2] (400 and 800 ppm), N fertilization rates (N1 and N2), and irrigation regimes (FI, DI, and alternative PRI). Error bars indicate standard error of the mean (n = 4). Different letters at each sampling data indicate significant differences between treatments according to LSD’s multiple range test at P < 0.05. Statistical comparisons among the treatments are presented in Table 2.
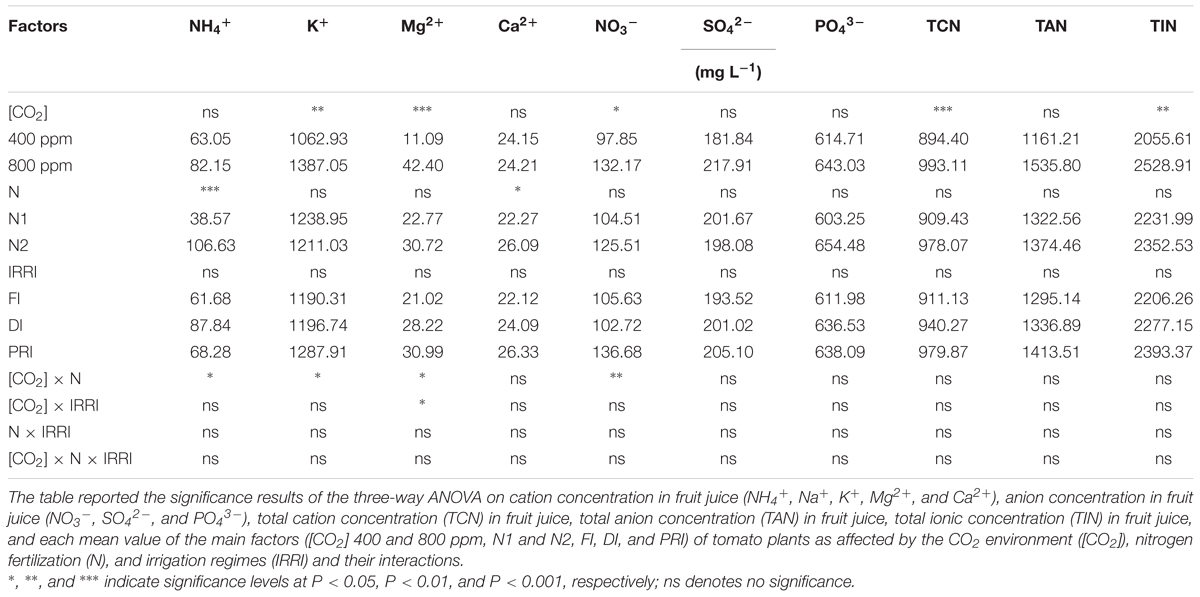
TABLE 2. Output of three-way ANOVA and mean value on fruit quality parameters of overall ion concentration in tomato plants as affected by the atmospheric [CO2] (400 and 800 ppm), N fertilization rates (N1 and N2), and irrigation regimes (FI, DI, and alternative PRI).
Anion Concentrations in Fruit Juice
NO3- concentration in fruit juice was affected by [CO2] and [CO2] × N (Figure 3D and Table 2). NO3- for a[CO2] plants was significantly lower than those for e[CO2] plants. SO42- and PO43- concentrations, however, were unaffected by any of the [CO2], N, and IRRI treatments (Table 2).
Total Cation, Anion, and Ionic Concentrations in Fruit Juice
Total cation concentration in fruit juice was only significantly affected by [CO2] treatment (Table 2); e[CO2] plants had significantly greater TCN than a[CO2] plants. TAN was unaffected by any of the [CO2], N, and IRRI treatments (Table 2). TIN was only remarkably affected by [CO2] treatment (Table 2), and was greater in plants grown at e[CO2] than those grown under a[CO2] environment.
PCA Evaluation of Comprehensive Fruit Quality
The PCA revealed that PRI and DI plants grown under N2 and e[CO2] environment had the highest Pi∗ (0.65 and 0.59) values, leading to the first and second rank of comprehensive fruit quality, respectively, among all treatments (Figure 4).
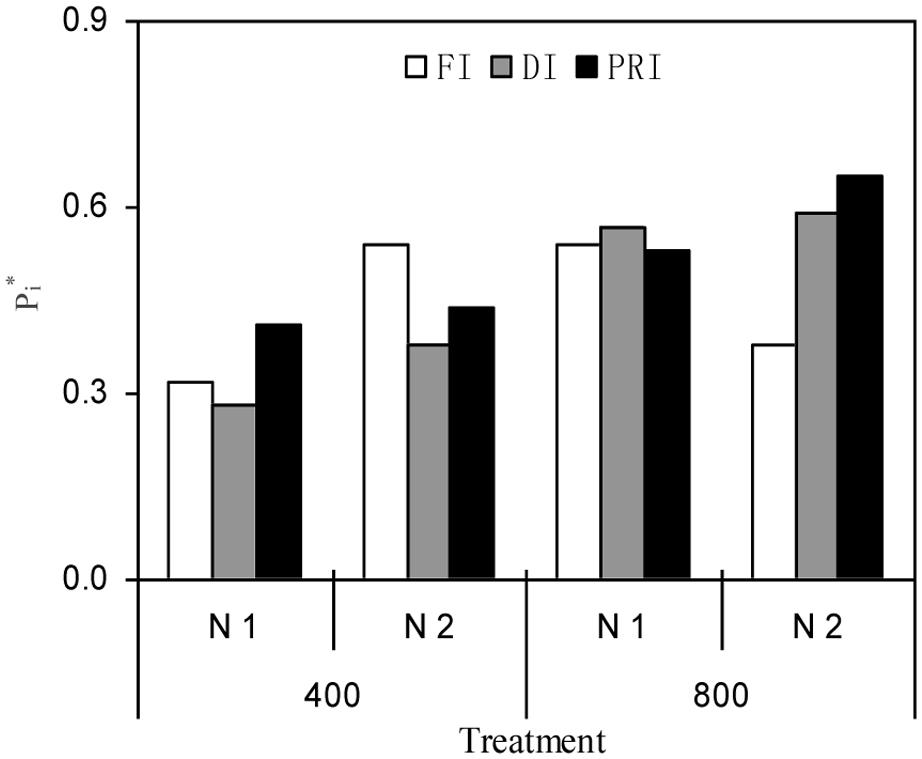
FIGURE 4. PCA evaluation of comprehensive tomato quality attributes (15 parameters) among all treatments (the atmospheric [CO2] (400 and 800 ppm), N fertilization rates (N1 and N2), and irrigation regimes (FI, DI, and alternative PRI). Pi∗ value indicates the closeness between principal components in each treatment.
Discussion
It is well known that a low N supply could reduce the vegetative growth, number of fruit sets, and yield of fruits (Huett and Dettmann, 1988). In good agreement with this, here, N1 plants had significantly lower FN and FY than N2 plants, regardless of [CO2] and IRRI treatments (Table 1). Nonetheless, SFW was similar among treatments (Table 1). Thus, the reduced FY for lower N fertilization was attributed mainly to the decreased FN. It has been suggested that plants grown in limited N possess lower photosynthetic capacity, and reduced photoassimilates from source leaves to be translocated into the fruits, hereby decreasing fruit set and FY (Bénard et al., 2009). Likewise, plants grown under e[CO2] possessed greater FY than those grown under a[CO2] treatment. This is consistent with the findings by Domis et al. (2002); Ainsworth and Long (2005), and Pazzagli et al. (2016) that e[CO2] environment could enhance the yield or biomass production of crop. The reduction in oxygenase activity of Rubisco at e[CO2] plants could lead to a decrease of photorespiration rate and in turn to an increase on net photosynthesis (Ainsworth and Long, 2005). Therefore, sustaining higher rates of photosynthesis by either sufficient N supply and/or in combination with e[CO2] could substantially enhance FY in tomato plant (Moretti et al., 2010). On the other hand, for the same N and irrigation treatment, there was no difference in soil water content between the e[CO2] and a[CO2] treatments during the treatment period (Figure 1), indicating that e[CO2] plants consumed similar amount of water compared with a[CO2] plants (Wei et al., 2018). Thus, an improvement of water use efficiency for fruit production could be expected (data not shown).
Previous studies have frequently observed that both DI and PRI could consume 25–50% less water without significant reduction in FY or biomass compared to FI (Kang and Zhang, 2004; Zegbe et al., 2004, 2006; Wang et al., 2010; Du et al., 2015; Barrios-Masias and Jackson, 2016; Wei et al., 2016), although several reports revealed a significant reduction of tomato FY in PRI compared with FI (Kirda et al., 2004; Zegbe et al., 2004). In addition, studies have shown that PRI was superior to DI in improving FY (Dodd, 2009) and fruit quality (Sun et al., 2013, 2014) in tomatoes. In the present study, it is notable that the FY of plant grown under reduced irrigation (DI and PRI) was largely sustained when compared to FI treatment at e[CO2], whereas this response was not seen in a[CO2] environment where significant reduction of FY due to reduced irrigation was observed (Table 1). Hereby, this result clearly demonstrated that plants grown in the e[CO2] environment could attenuate the negative influence of water stress on yield, which was in good agreement with earlier findings (Zhou et al., 2014; Mamatha et al., 2015; Pazzagli et al., 2016).
This study found that DI and PRI plants possessed greater FM than FI with N2 under e[CO2] environment (Figure 2A and Table 1), implying that better tomato fruit transport and storage quality was achieved when exposed to reduced irrigation associated with e[CO2] treatment. Also, plants grown at N1 generally possessed greater concentrations of TSS, TS, and OA than those grown at N2 (Figures 2B,C and Table 1). As it is well known, N level has a strong effect on the amounts of primary and secondary metabolites in fruits (Bénard et al., 2009). Simonne et al. (2007) reported that increased OA content was associated with decreased N supply; however, other researchers found the opposite results (Huett and Dettmann, 1988; Wang et al., 2007). Such discrepancy could be due to the different cultivars and experimental conditions as well as the source–sink balance in the plants.
The concentrations of TS, OA, and SAR are the primary biochemical fruit quality attributes that contribute to the flavor in tomatoes (Domis et al., 2002). Here, under N1 fertilization, the e[CO2] plants had higher TSS and TS and lower OA than a[CO2] plants (Figures 2B,C and Table 1). Also, reduced irrigation, especially PRI plants tended to increase TS, decrease OA, and contributed to significantly greater SAR in relation to the FI under N1 + a[CO2] and N2 + e[CO2] treatments (Figures 2C,D and Table 1). It is widely accepted that reduced irrigation could maintain or enhance various aspects of tomato fruit quality (Zegbe et al., 2003; Campos et al., 2009; Xu et al., 2009; Patanè and Cosentino, 2010; Sun et al., 2014). The higher TS and lower OA together with higher SAR in fruit juice under reduced irrigation regimes indicate an improvement of fruit sweetness (Wang and Frei, 2011; Hou et al., 2017). As for the increased TSS and TS concentrations in tomato fruit, it is probably ascribed to the modulation of underlying physiological mechanisms. First, the reduction of side shoots and reproductive growth in reduced irrigation-treated plants result in a relative improvement of the sink activity in tomato fruit; thus, the carbohydrate previously partitioned toward the side shoots is redirected toward into the fruit, accompanied by the increased availability of assimilates for the remaining fruits (Davies et al., 2000; Patanè and Cosentino, 2010). Second, moderate water stress could induce a greater starch accumulation in the fruits during the earlier stage of fruit development, along with the higher conversion of starch to sugars during fruit maturation (Zegbe et al., 2003). Third, the activities of carbohydrate-metabolizing enzymes, primarily invertase plays an important role in modulating sugar concentration in tomato fruits and its activity could be modulated by ABA (Ruan et al., 2010; Sun et al., 2014). Reduced irrigation, especially PRI had the ability to induce higher ABA concentration in the tomato plants (Wang et al., 2010; Sun et al., 2013). Thereby, this hormone may stimulate invertase activity and trigger expected greater sugars in the fruit (Ruan et al., 2010).
The beneficial effects of CO2 enrichment on photosynthetic rate and fruit quality of crops including tomato have been extensively reported by numerous studies (Islam et al., 1996; Bindi et al., 2001; Högy and Fangmeier, 2009; Moretti et al., 2010; Sun et al., 2012). Increases in rubisco content and activity result in enhanced leaf photosynthesis, which probably lead to accumulated concentration of carbon-based compounds in response to e[CO2] environment due to the source–sink balance hypothesis (Peñuelas and Estiarte, 1998), such as TSS, diverse sugars, and acids. Furthermore, in this study, there was no decrease in water consumption at e[CO2], which could facilitate the transport of assimilates into fruits and anticipate to induce greater increase in the flavor components of fruits.
The quantity and ingredient of minerals mainly macronutrient in the fruit have a prominent effect on the nutritional quality of tomatoes (Domis et al., 2002). Here, it was found that high N supply resulted in greater NH4+ and NO3- concentrations in fruit juice under PRI (Figures 3A,D and Table 2), confirming the fact that N deficiency could lead to the inadequate N uptake in the plant. Moreover, regardless of N fertilization, e[CO2] plants combined with reduced irrigation commonly showed no decrease in concentrations of NH4+, Ca2+, SO42-, PO43- and total anion (TAN) or increase in concentrations of K+, Mg2+, NO3-, total cation (TCN) and total ion (TIN) in fruit juice (Figure 3 and Table 2). It is well demonstrated that soil water dynamics under reduced irrigation, especially PRI induced drying and wetting cycles in the soil can have a predominantly positive influence on the bioavailability of mineral nutrients and their movement from the bulk soil to the roots of plant (Wang et al., 2010), and may enhance the root acquisition of mineral elements thereby increasing the nutritional status of the plants (Wang et al., 2012), including the ionic concentrations in the tomato fruits (Sun et al., 2013, 2014). Furthermore, due to an enhanced xylem connection to the fruits, the proportion of the ions in the plants allocated to the fruit could also be enhanced by the reduced irrigation treatments, particularly PRI (Davies et al., 2000).
It has been reported that across diverse tissues and experimental conditions, there is a nearly 8% reduction in the overall mineral content of plants grown under e[CO2] environment (Loladze, 2014), which is mainly attributed to the decreased mass flux for nutrients transport from bulk soil to root surface, limited root nutrient acquisition capacity, and dilution by a greater plant biomass (Loladze, 2002; Myers et al., 2014). In the present study, the e[CO2] plants showed similar water consumption with the a[CO2] plants, implying the equivalent mass flow at e[CO2] compared to a[CO2]. Additionally, studies have indicated that the proportion of assimilates allocated to roots is enhanced at e[CO2], resulting in a larger root system which could enhance the ability to absorb more minerals from the surrounding soil and improve the ionic contents in fruits (Idso and Idso, 2001; Wullschleger et al., 2002). Therefore, the expected negative impacts of drought stress and e[CO2] on tomato ions uptake were not evident in this study, and concentration of some minerals in fruit juice could be enhanced under e[CO2] combined with PRI strategy.
It is well known that PCA approach is a popular tool for statistical analysis and plays an important role in featuring extraction and dimensionality reduction, further realizing the accurate integrated appraisement on the variance of data source (Wang et al., 2015). In this study, the PCA assessment of comprehensive fruit quality attributes including 15 different parameters indicated that plants grown under sufficient N supply together with e[CO2] condition, the comprehensive fruit quality of deficit irrigation-treated plants, particularly for PRI, was superior to those under other treatments (Figure 4). Therefore, a greater fruit quality could be achieved under PRI associated with e[CO2] environment.
Conclusion
Conclusively, low N supply decreased tomato FN and yield but increased fruit quality across irrigation and [CO2] treatments. Plants grown in e[CO2] environment could eliminate the negative impact of reduced irrigation on FY. Moreover, at adequate N fertilization rate, an improvement of comprehensive fruit quality could be achieved by growing in e[CO2] environment combined with reduced irrigation regimes, particularly with the PRI strategy. These findings are of great relevance for formulating agronomical practices to sustain tomato yield and quality in a future drier and CO2-enriched environment.
Author Contributions
ZW: done the experiments and finished the first manuscript. TD and FL: supervised the work. XL and LF: helped to do the experiments.
Funding
This work was partially supported by the National Natural Science Foundation of China (Grant Nos. 51725904, 51439006, and 51621061) and the National Key Research Program (Grant No. 2016YFC0400207). Zhenhua Wei appreciates the Chinese Scholarship Council (CSC) for supporting his study at the Faculty of Science, University of Copenhagen, Denmark.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Technical assistance by Rene Hvidberg Petersen and Lene Korsholm Jørgensen is gratefully acknowledged.
References
Ainsworth, E. A., and Long, S. P. (2005). What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 165, 351–372. doi: 10.1111/j.1469-8137.2004.01224.x
Barrios-Masias, F. H., and Jackson, L. E. (2016). Increasing the effective use of water in processing tomatoes through alternate furrow irrigation without a yield decrease. Agric. Water Manag. 177, 107–117. doi: 10.1016/j.agwat.2016.07.006
Bénard, C., Gautier, H., Bourgaud, F., Grasselly, D., Navez, B., Caris-Veyrat, C., et al. (2009). Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem. 57, 4112–4123. doi: 10.1021/jf8036374
Bindi, M., Fibbi, L., and Miglietta, F. (2001). Free Air CO2 Enrichment (FACE) of grapevine (Vitis vinifera L.): II. Growth and quality of grape and wine in response to elevated CO2 concentrations. Eur. J. Agron. 14, 145–155. doi: 10.1016/S1161-0301(00)00093-9
Campos, H., Trejo, C., Peña-Valdivia, C. B., Ramírez-Ayala, C., and Sánchez-García, P. (2009). Effect of partial rootzone drying on growth, gas exchange, and yield of tomato (Solanum lycopersicum L.). Sci. Hortic. 120, 493–499. doi: 10.1016/j.scienta.2008.12.014
Davies, W. J., Bacon, M. A., Thompson, D. S., Sobeih, W., and Rodrígues, L. G. (2000). Regulation of leaf and fruit growth in plants growing in drying soil: exploitation of the plants’ chemical signalling system and hydraulic architecture to increase the efficiency of water use in agriculture. J. Exp. Bot. 51, 1617–1626. doi: 10.1093/jexbot/51.350.1617
Davies, W. J., Wilkinson, S., and Loveys, B. (2002). Stomatal control by chemical signalling and the exploitation of this mechanism to increase water use efficiency in agriculture. New Phytol. 153, 449–460. doi: 10.1046/j.0028-646X.2001.00345.x
Dodd, I. C. (2009). Rhizosphere manipulations to maximize ‘crop per drop’ during deficit irrigation. J. Exp. Bot. 60, 2454–2459. doi: 10.1093/jxb/erp192
Domis, M., Papadopoulos, A. P., and Gosselin, A. (2002). Greenhouse tomato fruit quality. Hortic. Rev. 26, 239–319. doi: 10.1002/9780470650806.ch5
Du, T. S., Kang, S. Z., Zhang, J. H., and Davies, W. J. (2015). Deficit irrigation and sustainable water-resource strategies in agriculture for China’s food security. J. Exp. Bot. 66, 2253–2269. doi: 10.1093/jxb/erv034
Högy, P., and Fangmeier, A. (2009). Atmospheric CO2 enrichment affects potatoes: 2. Tuber quality traits. Eur. J. Agron. 30, 85–94. doi: 10.1016/j.eja.2008.07.006
Hou, M. M., Jin, Q., Lu, X. Y., Li, J. Y., Zhong, H. Z., and Gao, Y. (2017). Growth, water use, and nitrate-15N uptake of greenhouse tomato as influenced by different irrigation patterns, 15N labeled depths, and transplant times. Front. Plant Sci. 8:666. doi: 10.3389/fpls.2017.00666
Huett, D. O., and Dettmann, E. B. (1988). Effect of nitrogen on growth, fruit quality and nutrient uptake of tomatoes grown in sand culture. Aust. J. Exp. Agric. 28, 391–399. doi: 10.1071/EA9880391
Idso, S. B., and Idso, K. E. (2001). Effects of atmospheric CO2 enrichment on plant constituents related to animal and human health. Environ. Exp. Bot. 45, 179–199. doi: 10.1016/S0098-8472(00)00091-5
Intergovernmental Panel on Climate Change [IPCC] (2013). Climate Change 2014-Impacts, Adaptation and Vulnerability: Regional Aspects. Cambridge: Cambridge University Press.
Islam, M. S., Matsui, T., and Yoshida, Y. (1996). Effect of carbon dioxide enrichment on physico-chemical and enzymatic changes in tomato fruits at various stages of maturity. Sci. Hortic. 65, 137–149. doi: 10.1016/0304-4238(95)00867-5
Jones, R. A., and Scott, S. J. (1984). Genetic potential to improve tomato flavour in commercial F1 hybrids. J. Am. Soc. Hortic. Sci. 109, 318–321.
Kang, S. Z., and Zhang, J. H. (2004). Controlled alternate partial root zone irrigation: its physiological consequences and impact on water use efficiency. J. Exp. Bot. 55, 2437–2446. doi: 10.1093/jxb/erh249
Kirda, C., Cetin, M., Dasgan, Y., Topcu, S., Kaman, H., Ekici, B., et al. (2004). Yield response of greenhouse grown tomato to partial root drying and conventional deficit irrigation. Agric. Water Manag. 69, 191–201. doi: 10.1016/j.agwat.2004.04.008
Li, X. N., Jiang, D., and Liu, F. L. (2016). Dynamics of amino acid carbon and nitrogen and relationship with grain protein in wheat under elevated CO2 and soil warming. Environ. Exp. Bot. 132, 121–129. doi: 10.1016/j.envexpbot.2016.08.013
Liu, F. L., Shahnazari, A., Andersen, M. N., Jacobsen, S. E., and Jensen, C. R. (2006). Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J. Exp. Bot. 57, 3727–3735. doi: 10.1093/jxb/erl131
Loladze, I. (2002). Rising atmospheric CO2 and human nutrition: toward globally imbalanced plant stoichiometry? Trends Ecol. Evol. 17, 457–461. doi: 10.1016/S0169-5347(02)02587-9
Loladze, I. (2014). Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. Elife 3:e02245. doi: 10.7554/eLife.02245
Mamatha, H., Srinivasa Rao, N. K., and Vijayalakshmi, T. (2015). Physiological responses of tomato (Lycopersicon esculentum Mill) cv. Arka Ashish to elevated atmospheric CO2 under water limiting conditions. Indian J. Agric. Res. 49, 299–307. doi: 10.5958/0976-058X.2015.00055.4
Moretti, C. L., Mattos, L. M., Calbo, A. G., and Sargent, S. A. (2010). Climate changes and potential impacts on postharvest quality of fruit and vegetable crops: a review. Food Res. Int. 43, 1824–1832. doi: 10.1016/j.foodres.2009.10.013
Myers, S. S., Zanobetti, A., Kloog, I., Huybers, P., Leakey, A. D. B., Bloom, A. J., et al. (2014). Increasing CO2 threatens human nutrition. Nature 510, 139–142. doi: 10.1038/nature13179
Patanè, C., and Cosentino, S. L. (2010). Effects of soil water deficit on yield and quality of processing tomato under a Mediterranean climate. Agric. Water Manag. 97, 131–138. doi: 10.1016/j.agwat.2009.08.021
Pazzagli, P. T., Weiner, J., and Liu, F. L. (2016). Effects of CO2 elevation and irrigation regimes on leaf gas exchange, plant water relations, and water use efficiency of two tomato cultivars. Agric. Water Manag. 169, 26–33. doi: 10.1016/j.agwat.2016.02.015
Peñuelas, J., and Estiarte, M. (1998). Can elevated CO2 affect secondary metabolism and ecosystem function? Trends Ecol. Evol. 13, 20–24. doi: 10.1016/S0169-5347(97)01235-4
Ruan, Y. L., Jin, Y., Yang, Y. J., Li, G. J., and Boyer, J. S. (2010). Sugar input, metabolism, and signaling mediated by invertase: roles in development, yield potential, and response to drought and heat. Mol. Plant 3, 942–955. doi: 10.1093/mp/ssq044
Sainju, U. M., Dris, R., and Singh, B. (2003). Mineral nutrition of tomato. Food Agric. Environ. 1, 176–184.
Sanz-Sáez,Á., Erice, G., Aranjuelo, I., Nogués, S., Irigoyen, J. J., and Sánchez-Díaz, M. (2010). Photosynthetic down-regulation under elevated CO2 exposure can be prevented by nitrogen supply in nodulated alfalfa. J. Plant Physiol. 167, 1558–1565. doi: 10.1016/j.jplph.2010.06.015
Simonne, A. H., Fuzere, J. M., Simonne, E., Hochmuth, R. C., and Marshall, M. R. (2007). Effects of nitrogen rates on chemical composition of yellow grape tomato grown in a subtropical climate. J. Plant Nutr. 30, 927–935. doi: 10.1080/15226510701375465
Sun, P., Mantri, N., Lou, H. Q., Hu, Y., Sun, D., Zhu, Y. Q., et al. (2012). Effects of elevated CO2 and temperature on yield and fruit quality of strawberry (Fragaria × ananassa Duch.) at two levels of nitrogen application. PLoS One 7:e41000. doi: 10.1371/journal.pone.0041000
Sun, Y. Q., Feng, H., and Liu, F. L. (2013). Comparative effect of partial root-zone drying and deficit irrigation on incidence of blossom-end rot in tomato under varied calcium rates. J. Exp. Bot. 64, 2107–2116. doi: 10.1093/jxb/ert067
Sun, Y. Q., Holm, P. E., and Liu, F. L. (2014). Alternate partial root-zone drying irrigation improves fruit quality in tomatoes. Hortic. Sci. 41, 185–191.
Wang, C. X., Gu, F., Chen, J. L., Yang, H., Jiang, J. J., Du, T. S., et al. (2015). Assessing the response of yield and comprehensive fruit quality of tomato grown in greenhouse to deficit irrigation and nitrogen application strategies. Agric. Water Manag. 161, 9–19. doi: 10.1016/j.agwat.2015.07.010
Wang, Y. S., Liu, F. L., Andersen, M. N., and Jensen, C. R. (2010). Improved plant nitrogen nutrition contributes to higher water use efficiency in tomatoes under alternate partial root-zone irrigation. Funct. Plant Biol. 37, 175–182. doi: 10.1071/FP09181
Wang, Y. S., Liu, F. L., and Jensen, C. R. (2012). Comparative effects of deficit irrigation and alternate partial root-zone irrigation on xylem pH, ABA and ionic concentrations in tomatoes. J. Exp. Bot. 63, 1907–1917. doi: 10.1093/jxb/err370
Wang, Y. T., Huang, S. W., Liu, R. L., and Jin, J. Y. (2007). Effects of nitrogen application on flavor compounds of cherry tomato fruits. J. Plant Nutr. Soil Sci. 170, 461–468. doi: 10.1002/jpln.200700011
Wang, Y. X., and Frei, M. (2011). Stressed food-The impact of abiotic environmental stresses on crop quality. Agric. Ecosyst. Environ. 141, 271–286. doi: 10.1016/j.agee.2011.03.017
Wei, Z. H., Du, T. S., Li, X. N., Fang, L., and Liu, F. L. (2018). Interactive effects of CO2 concentration elevation and nitrogen fertilization on water and nitrogen use efficiency of tomato grown under reduced irrigation regimes. Agric. Water Manag. 202, 174–182.
Wei, Z. H., Du, T. S., Zhang, J., Xu, S. J., Cambre, P. J., and Davies, W. J. (2016). Carbon isotope discrimination shows a higher water use efficiency under alternate partial root-zone irrigation of field-grown tomato. Agric. Water Manag. 165, 33–43. doi: 10.1016/j.agwat.2015.11.009
Wullschleger, S. D., Tschaplinski, T. J., and Norby, R. J. (2002). Plant water relations at elevated CO2-implications for water-limited environments. Plant Cell Environ. 25, 319–331. doi: 10.1046/j.1365-3040.2002.00796.x
Xu, H. L., Qin, F. F., Du, F. L., Xu, Q. C., Wang, R., Shah, R. P., et al. (2009). Application of xerophyticphysiology in plant production-partial root drying improves tomato crops. J. Food Agric. Environ. 7, 981–988.
Yan, F., Li, X. N., and Liu, F. L. (2017). ABA signaling and stomatal control in tomato plants exposure to progressive soil drying under ambient and elevated atmospheric CO2 concentration. Environ. Exp. Bot. 139, 99–104. doi: 10.1016/j.envexpbot.2017.04.008
Zegbe, J. A., Behboudian, M. H., and Clothier, B. E. (2004). Partial rootzone drying is a feasible option for irrigating processing tomatoes. Agric. Water Manag. 68, 195–206. doi: 10.1016/j.agwat.2004.04.002
Zegbe, J. A., Behboudian, M. H., and Clothier, B. E. (2006). Responses of ‘Petopride’ processing tomato to partial rootzone drying at different phonological stages. Irrig. Sci. 24, 203–210. doi: 10.1007/s00271-006-0043-y
Keywords: CO2, nitrogen, reduced irrigation, yield, fruit quality, tomato
Citation: Wei Z, Du T, Li X, Fang L and Liu F (2018) Interactive Effects of Elevated CO2 and N Fertilization on Yield and Quality of Tomato Grown Under Reduced Irrigation Regimes. Front. Plant Sci. 9:328. doi: 10.3389/fpls.2018.00328
Received: 01 November 2017; Accepted: 28 February 2018;
Published: 27 March 2018.
Edited by:
Stefania De Pascale, University of Naples Federico II, ItalyReviewed by:
María Serrano, Universidad Miguel Hernández de Elche, SpainPaolo Sambo, Università degli Studi di Padova, Italy
Copyright © 2018 Wei, Du, Li, Fang and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Taisheng Du, ZHV0YWlzaGVuZ0BjYXUuZWR1LmNu Fulai Liu, ZmxAcGxlbi5rdS5kaw==