 Rozaimi Razali1†
Rozaimi Razali1† Salim Bougouffa1†‡
Salim Bougouffa1†‡ Mitchell J. L. Morton2†
Mitchell J. L. Morton2† Damien J. Lightfoot2†‡
Damien J. Lightfoot2†‡ Intikhab Alam1,3
Intikhab Alam1,3 Magbubah Essack1
Magbubah Essack1 Stefan T. Arold1,2
Stefan T. Arold1,2 Allan A. Kamau1,3
Allan A. Kamau1,3 Sandra M. Schmöckel2
Sandra M. Schmöckel2 Yveline Pailles2
Yveline Pailles2 Mohammed Shahid4
Mohammed Shahid4 Craig T. Michell5‡
Craig T. Michell5‡ Salim Al-Babili2
Salim Al-Babili2 Yung Shwen Ho2
Yung Shwen Ho2 Mark Tester2
Mark Tester2 Vladimir B. Bajic1,3*
Vladimir B. Bajic1,3* Sónia Negrão2*‡
Sónia Negrão2*‡- 1Computational Bioscience Research Center, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
- 2Division of Biological and Environmental Sciences and Engineering, The Bioactives Lab, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
- 3Division of Computer, Electrical and Mathematical Science and Engineering Division, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
- 4International Center for Biosaline Agriculture, Dubai, United Arab Emirates
- 5Red Sea Research Center, Biological and Environmental Sciences and Engineering Division, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia
Solanum pimpinellifolium, a wild relative of cultivated tomato, offers a wealth of breeding potential for desirable traits such as tolerance to abiotic and biotic stresses. Here, we report the genome assembly and annotation of S. pimpinellifolium ‘LA0480.’ Moreover, we present phenotypic data from one field experiment that demonstrate a greater salinity tolerance for fruit- and yield-related traits in S. pimpinellifolium compared with cultivated tomato. The ‘LA0480’ genome assembly size (811 Mb) and the number of annotated genes (25,970) are within the range observed for other sequenced tomato species. We developed and utilized the Dragon Eukaryotic Analyses Platform (DEAP) to functionally annotate the ‘LA0480’ protein-coding genes. Additionally, we used DEAP to compare protein function between S. pimpinellifolium and cultivated tomato. Our data suggest enrichment in genes involved in biotic and abiotic stress responses. To understand the genomic basis for these differences in S. pimpinellifolium and S. lycopersicum, we analyzed 15 genes that have previously been shown to mediate salinity tolerance in plants. We show that S. pimpinellifolium has a higher copy number of the inositol-3-phosphate synthase and phosphatase genes, which are both key enzymes in the production of inositol and its derivatives. Moreover, our analysis indicates that changes occurring in the inositol phosphate pathway may contribute to the observed higher salinity tolerance in ‘LA0480.’ Altogether, our work provides essential resources to understand and unlock the genetic and breeding potential of S. pimpinellifolium, and to discover the genomic basis underlying its environmental robustness.
Introduction
The Solanum section Lycopersicon is an economically important clade that consists of 14 species including the cultivated tomato Solanum lycopersicum (formerly Lycopersicon esculentum), which is the most economically important horticultural crop (Peralta et al., 2005; Spooner et al., 2005). This clade also contains Solanum pimpinellifolium, which is the closest wild relative of the cultivated tomato (The Tomato Genome Consortium, 2012; The 100 Tomato Genome Sequencing Consortium et al., 2014). S. pimpinellifolium has a bushy growth type, small red fruits (∼1.5 cm diameter) and is facultatively autogamous (Rick et al., 1978). The distribution of the species includes the dry coastal regions of Peru, Ecuador, and northern Chile (Luckwill, 1943; Warnock, 1991; Peralta et al., 2008), where plants are frequently exposed to brackish groundwater, salt-laden mist and other harsh environmental conditions (Rick et al., 1977; Peralta and Spooner, 2000; Zuriaga et al., 2009; Blanca et al., 2012).
Due to its exposure to these challenging environmental conditions over evolutionary time, S. pimpinellifolium exhibits a phenotypic robustness that appears to have been lost in cultivated tomato during the domestication process (Miller and Tanksley, 1990; Tanksley and McCouch, 1997; Bai and Lindhout, 2007). Thus, S. pimpinellifolium is regarded as an important source of genes that can confer favorable stress-tolerance to cultivated tomato. For instance, breeding tomatoes with resistance to bacterial speck disease (caused by Pseudomonas syringae) was achieved through the introgression of the resistance gene, Pto, from S. pimpinellifolium into commercial cultivars (Pitblado and Kerr, 1979; Pedley and Martin, 2003; Thapa et al., 2015). Furthermore, horticultural traits of commercial tomato, such as fruit size, have been influenced by the introduction of S. pimpinellifolium alleles (as reviewed by Tanksley, 2004; Azzi et al., 2015), some of which were identified by the molecular mapping of backcross populations developed from S. pimpinellifolium (Tanksley et al., 1996). Additionally, numerous quantitative trait loci (QTLs) have been identified using S. pimpinellifolium, such as those for biotic stress (Salinas et al., 2013; Chen et al., 2014; Víquez-Zamora et al., 2014; Ni et al., 2017), abiotic stress (Villalta et al., 2008; Lin et al., 2010), fruit quality traits (Tanksley et al., 1996; Chen et al., 1999; Xiao et al., 2008; Capel et al., 2016), and other agronomic traits (Doganlar et al., 2002; Cagas et al., 2008; Nakano et al., 2016). Numerous S. pimpinellifolium accessions have been previously characterized as having a high salinity tolerance (ST) and are promising sources of genes and alleles for improvement of ST in cultivated tomato (Bolarin et al., 1991; Cuartero et al., 1992; Foolad and Lin, 1997; Foolad et al., 1998; Cuartero and Fernandez-Munoz, 1999; Foolad, 1999; Foolad and Chen, 1999; Bolarin et al., 2001; Foolad et al., 2001; Zhang et al., 2003; Villalta et al., 2008; Estan et al., 2009; Rao et al., 2013, 2015).
To drive research and to facilitate the discovery of genes that confer favorable traits, the Tomato Genome Consortium published the high-quality genome sequence of S. lycopersicum cv. ‘Heinz 1706,’ as well as a draft sequence of S. pimpinellifolium accession ‘LA1589’ (The Tomato Genome Consortium, 2012). The availability of the cultivated tomato genome has led to several important advances, such as the identification of candidate genes (CG) related to fruit development (Zhong et al., 2013; Liu et al., 2016), the development of single nucleotide polymorphism (SNP) genotyping arrays (Sim et al., 2012a,b; Víquez-Zamora et al., 2014), the design of the CRISPR-cas9 gene-editing system (Brooks et al., 2014), and the identification of loci contributing to improved tomato flavor quality (Tieman et al., 2017). While the draft genome sequence of S. pimpinellifolium ‘LA1589’ has been used in several previous studies (e.g., Kevei et al., 2015), the fragmented nature of the assembly (309,180 contigs), the low sequencing coverage of the genome and the limitations of the available genome annotation constrain the usefulness of this assembly. Additionally, a further three accessions of S. pimpinellifolium (LYC2798, LA1584 and LA1578) were sequenced by the 100 Tomato Genome Project (The 100 Tomato Genome Sequencing Consortium et al., 2014), but genome assemblies and annotations for these accessions have not been performed. Thus, the availability of an improved genome assembly for S. pimpinellifolium is expected to provide increased opportunities for the discovery of new genes unique to wild germplasm within the Lycopersicon clade.
Here we report the results of a field trial that confirms the previously reported high ST of S. pimpinellifolium relative to the commercial tomato, S. lycopersicum ‘Heinz 1706.’ We used the S. pimpinellifolium accession ‘LA0480,’ which ranked in the top 50 accessions in terms of ST out of 200 genotypes in a recent large-scale field experiment (unpublished data). To investigate the genomic basis of this ST, we used Illumina technology to sequence the genome of S. pimpinellifolium ‘LA0480’ to a depth of 197x and produced a genome assembly of 811 Mb, with final scaffold N50 of 75,736 bp (Table 1 and Supplementary Table S4). This assembly is a substantial improvement on the previously reported genome assembly of S. pimpinellifolium accession ‘LA1589.’ We annotated 25,134 protein-coding genes (Table 1 and Supplementary Table S9) within our assembly with Dragon Eukaryotic Analysis Platform (DEAP), which is a new tool for functional genome annotation and comparison and is presented here for the first time. The DEAP (pronounced DEEP) ‘Annotate’ module was used to assign annotation from multiple sources including the Kyoto Encyclopedia of Genes and Genomes (KEGG) database, UniProt and InterProScan. Additionally, the DEAP ‘Compare’ module was used to compare genome annotations of S. pimpinellifolium and S. lycopersicum. The use of multiple comparative genomics approaches led to the identification of genes that may play a role in biotic and abiotic stress tolerance of S. pimpinellifolium ‘LA0480’ and these genes represent promising candidates for future investigation. Additionally, a CG approach led to the identification of genes encoding inositol-3-phosphate synthase (I3PS), a key enzyme involved in salinity response (Nelson et al., 1998), as having a higher copy number in S. pimpinellifolium ‘LA0480’ compared with other, less salt tolerant, tomato species. Our results suggest that I3PS and the inositol pathway may play an important role in ST in ‘LA0480.’
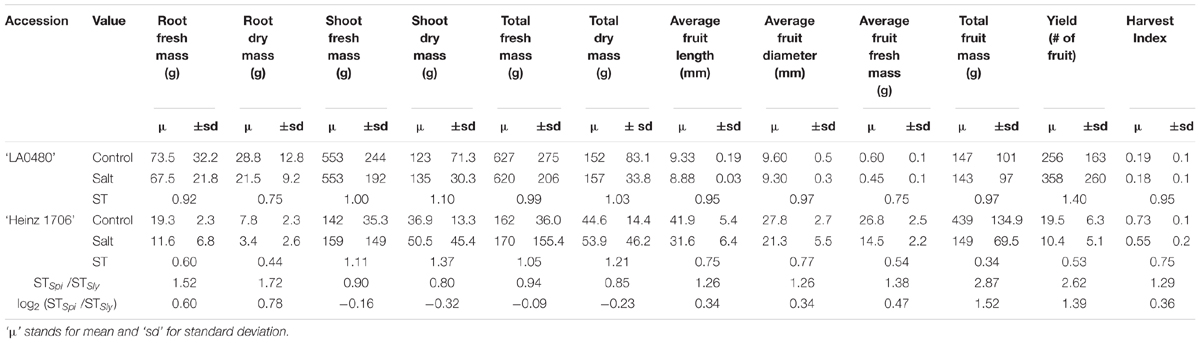
TABLE 1. Summary of field performance of S. pimpinellifolium and S. lycopersicum under control and saline conditions assessing various biomass and yield-related traits and their respective salinity tolerance (ST) index values for both species.
Materials and Methods
Salinity Tolerance Field Trial
A field trial was conducted at the International Center for Biosaline Agriculture (ICBA) in Dubai, United Arab Emirates (N 25° 05.847; E 055° 23.464), between October 2015 and May 2016. The complete experiment included 214 S. pimpinellifolium and 13 commercial accessions, but only ‘LA0480’ and ‘Heinz 1706’ are considered here. We used a randomized block design, with a non-saline and a saline plot, each comprising four blocks for a total of 4 replicates per genotype, per treatment. Plants were planted in rows, with 0.5 m spacing between plants, and 1 m spacing between rows. Plants were grown in a nursery for 6 weeks before being transplanted into the field. Following transplantation, plots were irrigated with non-saline water for the first 5 weeks, after which the irrigation for the saline field was switched to a saline source. Regular water sample analysis over the course of the experiment indicated an average electroconductivity (EC) of 0.7 dS/m-1 and 12.3 dS/m-1, and a NaCl concentration of 0.5 – 10 mM and 70 – 110 mM for the non-saline and saline water sources, respectively. After salt-stress application, the experiment was continued for 17 weeks. Mature fruit were harvested continually throughout the field trial to assess fruit- and yield-related traits and a final destructive harvest was performed to evaluate biomass traits. All measurements were spatially corrected in the R statistical computing environment (v2.12), using custom scripts and the ASReml v3.0-1 (Gilmour et al., 2009) package for R v3.2.0 (R Core Team, 2014).
The Harvest Index (HI) was defined as the fresh fruit yield as a proportion of the total fresh shoot mass (including fruit) (Gianfagna et al., 1997) and calculated with the formula:
The ST was calculated for each trait in each genotype (where Xsalt and Xcontrol are the mean value of variable X under salt stress and control conditions, respectively) using the formula:
‘LA0480’ DNA Library Construction, Sequencing and Assembly
The S. pimpinellifolium accession ‘LA0480’ was sequenced using the HiSeq 2000 Illumina platform at King Abdullah University of Science and Technology (KAUST) (Figure 8). DNA was extracted from whole flowers of a single soil-grown plant ‘LA0480-ref’ using the Qiagen DNeasy Plant Mini Kit (Qiagen, Germany). Two 101 bp paired-end (PE) short-read libraries (139 and 332 bp mean insert length) and five 100 bp mate-pair libraries (2, 6, 8, 10, and >10 kb insert length) were prepared using the NEBNext Ultra DNA Library Prep Kit and the Nextera Mate-pair Library Kit, respectively (New England Biolabs, United Kingdom).
Adapter sequences, low-quality four nucleotide stretches of nucleotides, and low quality leading and trailing bases were removed with Trimmomatic v0.33 (Bolger A.M. et al., 2014) and reads with a final length of less than 36 bp after trimming were discarded (Supplementary Table S1). Processed PE data were de novo assembled into contigs using ABySS (Simpson et al., 2009) with a k-mer length of 77, as determined by k-mer analysis (Supplementary Figure S1). These contigs were scaffolded based on library size information from the PE read libraries (Supplementary Table S2), followed by a second round of scaffolding with mate pair data utilizing the ABySS pipeline. Preliminary quality control was performed by mapping the sequencing reads back to the genome with BWA (BWA MEM) (Supplementary Table S3). GapCloser (Luo et al., 2012) was used to close gaps in the assembled scaffolds (Supplementary Table S4). The completeness of the genome assembly was assessed with BUSCO (Simão et al., 2015).
‘LA0480’ Transcriptome Sequencing and Assembly
RNA was extracted from a single root, young leaf, old leaf, petiole, meristem, flower, and immature fruit tissue sample collected from the mature soil-grown ‘LA0480-ref’ plant. Additionally, a single leaf and root sample from plants (‘LA0480-ref’ progeny) grown hydroponically under control (∼0 mM NaCl and 0 dS/m-1) and salt stress (∼200 mM NaCl and 16 dS/m-1) conditions were collected (Supplementary Material Section 4 and Supplementary Tables S6, S7). RNA was extracted using the ZR Plant RNA MiniPrep Kit (Zymo, Orange County, CA, United States). RNA sequencing libraries were prepared using the NEBNext Ultra Directional RNA Library Prep Kit for Illumina (New England BioLabs, United Kingdom) and sequencing reads were processed with Trimmomatic and assembled into transcripts using Trinity v2.0.6 (Grabherr et al., 2011). Each RNA-seq library was assembled independently to minimize the creation of chimeric transcript isoforms. We removed low quality transcripts using TransRate v1.0.2 (Smith-Unna et al., 2016). BUSCO was used, as previously described, to assess the completeness of the genome annotation (Supplementary Table S8). The final RNA-seq fragment counts are presented in Supplementary Table S11.
‘LA0480’ Repeat Annotation
RepeatModeler v1.0.8 and RepeatMasker v4.0.5 (Tarailo-Graovac and Chen, 2009) were used to identify repetitive elements (RE). A library of de novo repeats was constructed with RepeatModeler and this library was subsequently merged with the RepBase library (v21.02) from RepeatMasker. RepeatMasker was run on the assembled genome (minimum length of 5 kb) using the total repeat library.
‘LA0480’ Gene Structure and Functional Annotation
To identify gene structures, we used the MAKER annotation pipeline v03 (Cantarel et al., 2008) with AUGUSTUS (Stanke et al., 2004) as the base ab initio gene predictor. AUGUSTUS was trained using the existing S. lycopersicum gene model as the basis and the assembled RNA-seq data as hints (Supplementary Material Section 6). Protein-coding genes were predicted using hints from the assembled transcripts as well as from the unassembled raw RNA-seq data and from the aligned proteins from S. lycopersicum, S. pennellii and SwissProt (Bairoch and Apweiler, 2000). tRNA genes were predicted using tRNAscan-SE (Lowe and Eddy, 1997). The predicted genes were assessed and assigned scores using MAKER based on the assembled transcripts and homologous proteins (Supplementary Material Section 6).
Functional annotation was performed using DEAP. KEGG Orthologs (KO) were assigned based on the KEGG database using BLASTp with a BLAST percent identity cut off of 60 and a maximum E-value of 1E-5. Functional domains, protein signatures and their associated Gene Ontology (GO) were assigned using InterProScan (Jones et al., 2014). For versions of the different tools and databases used under DEAP v1.0 refer to http://www.cbrc.kaust.edu.sa/deap/ (Supplementary Material Section 7).
Identification of Orthologous Genes
Orthologous and paralogous protein relationships between the four species were identified using OrthoMCL (Li et al., 2003). Custom Perl scripts were utilized to analyze OrthoMCL outputs for visualization with InteractiVenn (Heberle et al., 2015). Protein datasets for S. pennellii and S. lycopersicum were obtained from the Sol Genomics Network1 (Fernandez-Pozo et al., 2015) while the S. tuberosum protein dataset was obtained from Phytozome2. All sequences were downloaded in February 2017. The proteins corresponding to the primary transcripts were identified with custom scripts.
CNV-seq and SNP Analyses
CNVs were investigated using CNV-seq v0.2.7 (Xie and Tammi, 2009). Genomic raw reads from S. pimpinellifolium and S. lycopersicum (SRR404081) were aligned to the S. lycopersicum reference genome (NCBI assembly accession GCF_000188115.3) using BWA v0.7.10 (Li and Durbin, 2009) and alignment files were post-processed using SAMtools v1.3.1 (Li et al., 2009). Following this, short read data from S. pimpinellifolium and S. lycopersicum were mapped to the S. lycopersicum genome using the following settings: p ≤ 0.001, log2 threshold ≥±1, window size = 276, minimum window of 4 and using a genome-size of 813 Mb. The circular plot was generated using CIRCOS v0.69.3 (Krzywinski et al., 2009). We also produced high and low CNVs graphs for all 12 chromosomes using R (R Core Team, 2014) (Supplementary Figure S7). The complete dataset regarding the CNV analysis is present in Data Sheet 2.
For SNP analysis, the short-read sequence data from S. pimpinellifolium were mapped to the S. lycopersicum reference genome as described above. SNPs were called using the mpileup command of SAMtools (v1.3.1) and custom Perl scripts were used to filter SNPs for a depth of at least 8 and a SNP allele frequency greater than 75%. SNPs were binned into 1 Mb bins, and plotted together with the CNV data using CIRCOS.
KO Enrichment Analysis
KO enrichment analysis for the S. pimpinellifolium and S. lycopersicum genomes was performed using DEAP Compare (Supplementary Material Section 7). Only KO terms that were assigned based on BLAST percentage identity of at least 60% and above were considered (E-value ≤ 1E-5). For each observed KOi, we compared the ratio KOi / KOtotal-observed in each species using Fisher’s exact test (confidence interval 0.95). An enrichment is defined where the P-value is significant (P < 0.05). We corrected for multiple testing using the Benjamini–Hochberg method.
Identification of Salt Tolerance Candidate Genes and Orthologs
The salt tolerance CG list was adapted from Roy et al. (2014) (Table 3 and Supplementary Table S17) and verified against ‘Dragon Explorer of Osmoprotection associated Pathways’ – DEOP (Bougouffa et al., 2014). For CGs with supporting literature in S. pimpinellifolium, protein sequences were compared using BLASTp and multiple sequence alignment (MSA) tools such as MUSCLE (Edgar, 2004). We also performed BLASTp searches (identity thresholds of usually > 90%) and used OrthoMCL orthogroups to verify the orthology. For CGs with no supporting literature in S. pimpinellifolium, we investigated CG orthologs in S. lycopersicum using a combination of approaches: (1) BLASTp against S. lycopersicum total proteins; (2) orthogroup identification using OrthoDB; (3) inspection and comparison of functional domains; and (4) MSA and visual assessment of the alignment. Alignments for the CGs are presented in Supplementary Figures S10–S24. The workflow is summarized in the Supplementary Figure S9 and Supplementary Material Section 13.
Phylogenetic Analysis
The online tool Phylogeny.fr (Dereeper et al., 2008) was used for the phylogenetic analysis of Solanaceae species, with A. thaliana set as the outgroup. Multiple sequence alignment of the I3PS genes from these species was performed using ClustalOmega with two combined guide-trees and HMM iterations (Sievers et al., 2011). Details for DNA sequences can be found in Supplementary Table S18. The construction of the phylogenetic tree was estimated using the maximum likelihood method (PhyML), and the Generalized Time Reversible substitution model (GTR) (bootstrap value = 100). The tree was drawn with TreeDyn (Chevenet et al., 2006).
Structural Analysis of I3PS Proteins
SwissModel (Arnold et al., 2006) was used to produce homology models based on the ∼55% identical structure of the yeast MIP 1-L-myo-inositol-1-phosphate synthase [PDB id 1jki (Stein and Geiger, 2002); QMEAN values are between -2.0 and -2.3 for SpiI3PSa and SpiI3PSb alleles, and -3.2 for SpiI3PSc]. Models were manually inspected, and mutations evaluated, using the Pymol program3.
Myo-Inositol Content Determination
The selected tissues, old leaf (youngest fully expanded leaf at the time of salt imposition) and young leaf (youngest fully expanded leaf at the time of harvest) were harvested from seedlings grown following the “Hydroponics 2” protocol (Supplementary Material Section 4) 7 days after salt stress application. Three samples were collected and measured per genotype, tissue type and treatment. Frozen leaf samples were ground, freeze-dried and 20 mg of tissue was mixed with water. After centrifugation, myo-inositol content in supernatant was measured using K-INOSL assay kit according to the manufacturer’s instructions (Megazyme International Ireland, Bray, Wicklow, Ireland).
Measurement of Shoot Ion Concentration
Young and old leaves were collected from plants grown in parallel to those prepared for myo-inositol quantitation to assess the concentration of Na and K in leaf tissues. The fresh and dry mass of each sample (total of three replicates) was measured to determine the tissue water content. Dried leaf samples were digested overnight in 1% (v/v) nitric acid (HNO3) at 70°C. The concentrations of Na and K were determined in three biological replicates using a flame photometer (model 420; Sherwood Scientific Ltd., Cambridge, United Kingdom).
Results and Discussion
S. pimpinellifolium ‘LA0480’ Shows a Higher Salinity Tolerance Than Cultivated Tomato
To assess the ST of S. pimpinellifolium ‘LA0480’ under field conditions, we phenotyped S. lycopersicum ‘Heinz 1706’ and S. pimpinellifolium ‘LA0480’ under both control (non-saline) and saline conditions. From this field trial, we collected physiological measurements for both species specifically focusing on yield-related traits in the field that are the most relevant to breeders for downstream applications. We observed that the majority of traits were affected by salt stress in both genotypes (Table 1 and Figure 1), and that there were also clear differences in ST index between genotypes across different traits, with statistically significant differences between genotypes determined by ANOVA with Tukey pairwise comparison (Supplementary Figure S8). Strikingly, ‘LA0480’ ST values across all fruit- and yield-related traits were ∼1.25 to 2.5 times greater than in ‘Heinz 1706’ (Table 1). The high ST for yield (total fruit fresh mass) in ‘LA0480’ relative to ‘Heinz 1706’ cannot be attributed to differences in fruit dimensions (fruit length and fruit diameter) or individual fruit mass (average fruit fresh mass) but appears to be the result of a marked increase in fruit number in response to salt in ‘LA0480,’ whereas ‘Heinz 1706’ showed substantial reductions in all these traits under stress. That is, under salt stress, S. pimpinellifolium produced an increased quantity of fruit of similar size but reduced mass compared to control conditions; while S. lycopersicum produced fewer and smaller fruit under salt stress relative to control conditions. Interestingly, ST indices for shoot and total fresh and dry mass are 10–20% higher in ‘Heinz 1706’ than in ‘LA0480.’ While this difference is modest, it is informative that this did not translate to enhanced yield maintenance under stress compared with control conditions. This observation highlights the importance of studying agronomically important traits directly, rather than relying solely on expedient proxies such as biomass measurements at the immature stage, which is in line with the findings of Rao et al. (2013). Higher ST for root traits in ‘LA0480’ than in ‘Heinz 1706’ provides an interesting correlation with high fruit- and yield-related ST, but further studies are required to understand this potential relationship.
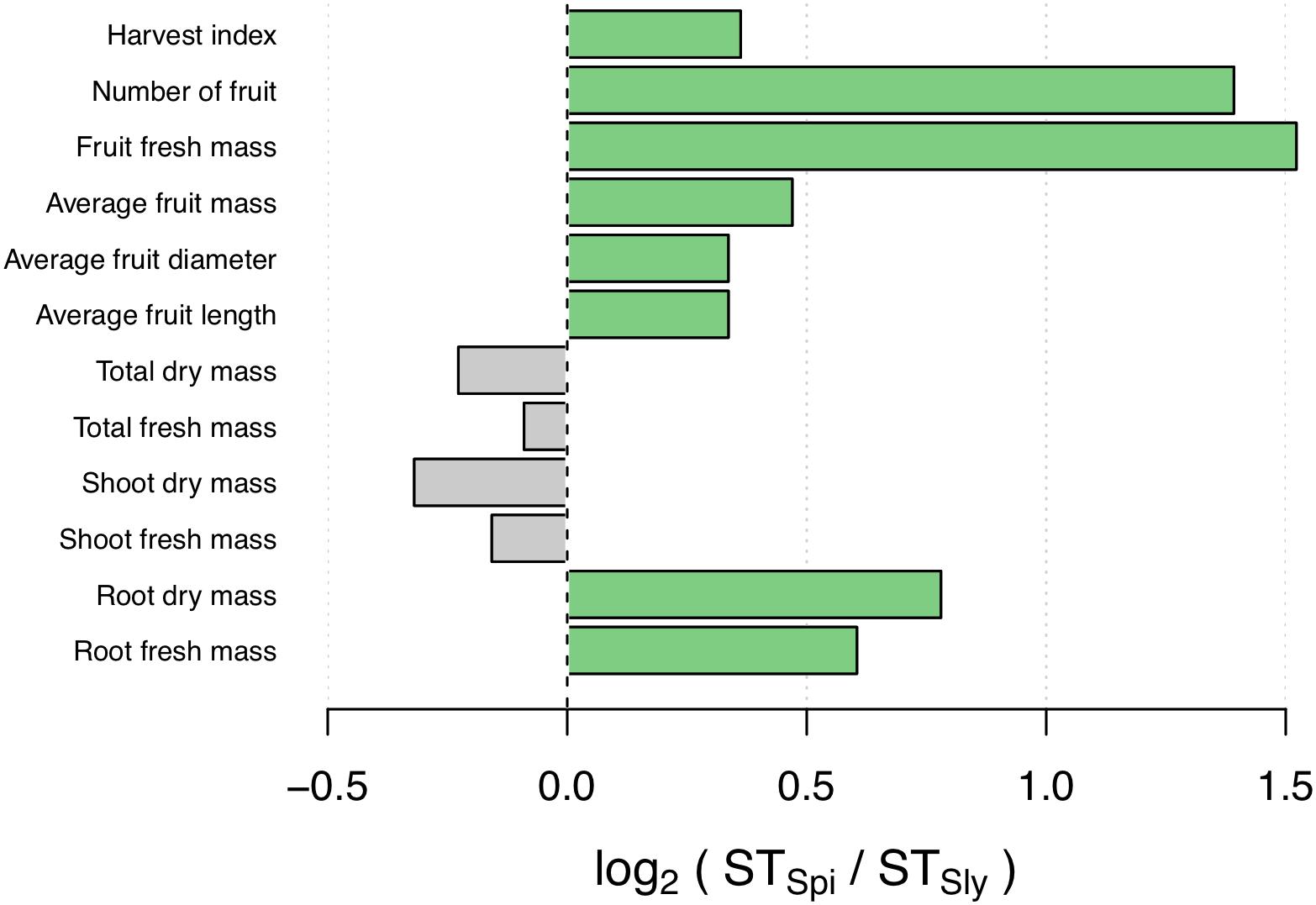
FIGURE 1. Comparison of S. pimpinellifolium and S. lycopersicum salinity tolerance (ST) indices across various traits measured in the field (log2 ratio). Traits for which the ST index is higher in S. pimpinellifolium and S. lycopersicum are in green and gray, respectively.
Altogether, our field results confirm previous reports of high ST in S. pimpinellifolium (Bolarin et al., 1991; Villalta et al., 2008; Rao et al., 2013) and show, specifically, that ‘LA0480’ is more salt tolerant than ‘Heinz 1706’ in fruit and yield-related traits. These findings underline compelling physiological differences between the two accessions that merit further investigation and open possibilities to improve ST in cultivated tomato. To establish the foundation for future research, we present the genome of S. pimpinellifolium accession ‘LA0480’ and investigate the genomic basis for its high ST.
Assembly and Annotation of the S. pimpinellifolium Reference Accession ‘LA0480’ Genome
The genome of S. pimpinellifolium ‘LA0480’ was sequenced using the Illumina HiSeq 2000 sequencing platform. We generated two paired-end libraries (insert sizes: 139 and 332 bp) and five mate-pair libraries (insert sizes: 2, 6, 8, 10, and >10 kb) (Supplementary Table S1), resulting in ∼108 and ∼52 Gb of data, respectively, producing an estimated genome coverage of ∼197x. The initial 160 Gb of raw data were processed to remove low quality sequences generating over 138 Gb of high quality data that were then assembled, scaffolded (Supplementary Table S2) and gap-closed into 163,297 final scaffolds with an N50 of 75,736 bp and a total size of 811.3 Mb (Table 2 and Supplementary Table S4). The assembled genome size is within the expected range compared to closely related species such as S. lycopersicum (900 Mb) (The Tomato Genome Consortium, 2012) and S. pennellii (942 Mb – 1.2 Gb, (Bolger A. et al., 2014). To assess the completeness of our genome assembly for all scaffolds above 1 kb, we used the Benchmarking Universal Single-Copy Orthologs (BUSCO) database (Simão et al., 2015). Of the 1,440 complete plant-specific single copy orthologs in the BUSCO database, we identified 1,375 (95.5%) orthologs in our assembly, denoting a high quality and nearly complete genome assembly (Supplementary Table S5).
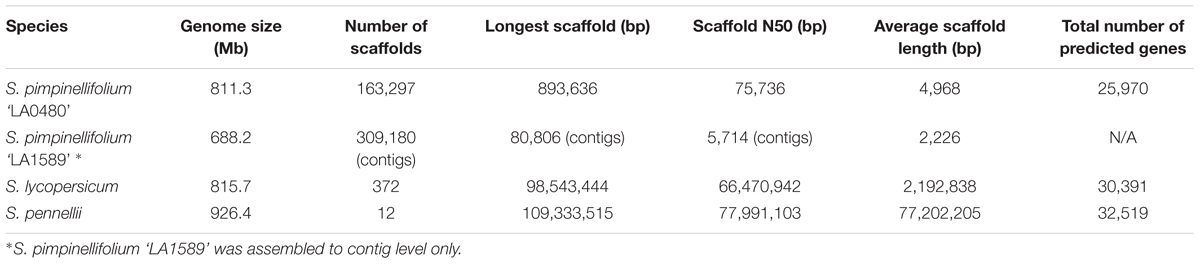
TABLE 2. Genome assembly and annotation statistics for S. pimpinellifolium ‘LA0480’ in comparison to S. pimpinellifolium ‘LA1589’ (The Tomato Genome Consortium, 2012), S. lycopersicum (The Tomato Genome Consortium, 2012), and S. pennellii (Bolger A. et al., 2014).
Analysis of the S. pimpinellifolium genome indicated that 59.5% of the assembled genome consisted of repetitive elements, with Long Terminal Repeats (LTR) retrotransposons of the Gypsy-type being the most abundant, comprising 37.7% of the assembled genome (Supplementary Table S14 and Supplementary Figure S6). This result is consistent with the repeat content of the genomes of both S. lycopersicum- 37.9% (The Tomato Genome Consortium, 2012) and S. pennellii- 40.1% (Bolger A. et al., 2014). The S. pimpinellifolium assembly presented here represents a substantial improvement over the previously published S. pimpinellifolium draft genome, which contained 309,180 contigs and had an estimated genome size of 739 Mb (The Tomato Genome Consortium, 2012). We used a combination of ab initio prediction and transcript evidence supported by RNA-seq data from multiple tissues and conditions to annotate a total of 25,970 genes (25,134 protein-coding genes producing 25,744 mRNAs of which 610 are isoforms) (Table 2 and Supplementary Table S9), with 21,016 genes (80.9%) assigned an annotation edit distance (AED) score of less than, or equal to, 0.3, indicating that they are well supported. A BUSCO completeness score of 91.9% for the genome annotation (Supplementary Table S8) was obtained, which is lower than the BUSCO results that we obtained for S. lycopersicum (99.3%) and S. pennellii (98.9%). This result is expected as the S. lycopersicum and S. pennellii genomes are more complete, as evidenced by their chromosome-level assemblies.
To investigate functional features of the protein-coding genes of S. pimpinellifolium and to compare with the protein-coding genes from closely related species (S. lycopersicum, S. pennellii and S. tuberosum), we developed DEAP, http://www.cbrc.kaust.edu.sa/deap/ (Supplementary Figures S2–S4), which is an extension of Dragon Metagenomic Analyses Platform (DMAP4). The longest protein isoform of each gene was submitted to DEAP Annotate v1.0 for functional annotation (Supplementary Table S10). Additionally, the longest protein isoform of each gene from the S. lycopersicum (NCBI annotation release 102, November 2016), S. pennellii (NCBI annotation release 100, December 2015) and S. tuberosum (NCBI annotation release 101, January 2016) genomes were annotated in the same manner and used for comparison. In addition, we analyzed the protein domains of S. pimpinellifolium, S. pennellii, and S. lycopersicum using InterProScan (Supplementary Table S12), and we observed that most abundant PFAM families shared between the S. pimpinellifolium and S. lycopersicum genomes are the protein kinase domains and the pentatricopeptide repeat family (PRR) (Supplementary Figure S5).
Comparative Genomics Within the Solanaceae
To investigate the gene space of the S. pimpinellifolium genome, we undertook a comparative genomics approach to compare S. pimpinellifolium to three other related species: a second wild tomato (S. pennellii); cultivated tomato (S. lycopersicum); and the more distantly related cultivated potato (S. tuberosum) (Figure 2). OrthoMCL analysis revealed 14,126 clusters of orthologs (containing 78,973 proteins) that are common to all four species analyzed and may represent the core set of genes in Solanum. A total of 715 clusters (2,438 proteins) were identified as being specific to the three members of the Lycopersicon clade, while 4,028 proteins were determined to be specific to S. pimpinellifolium, including 682 protein-coding genes with paralogs (Figure 2) and 3,346 proteins with no identified homologs (Supplementary Table S13).
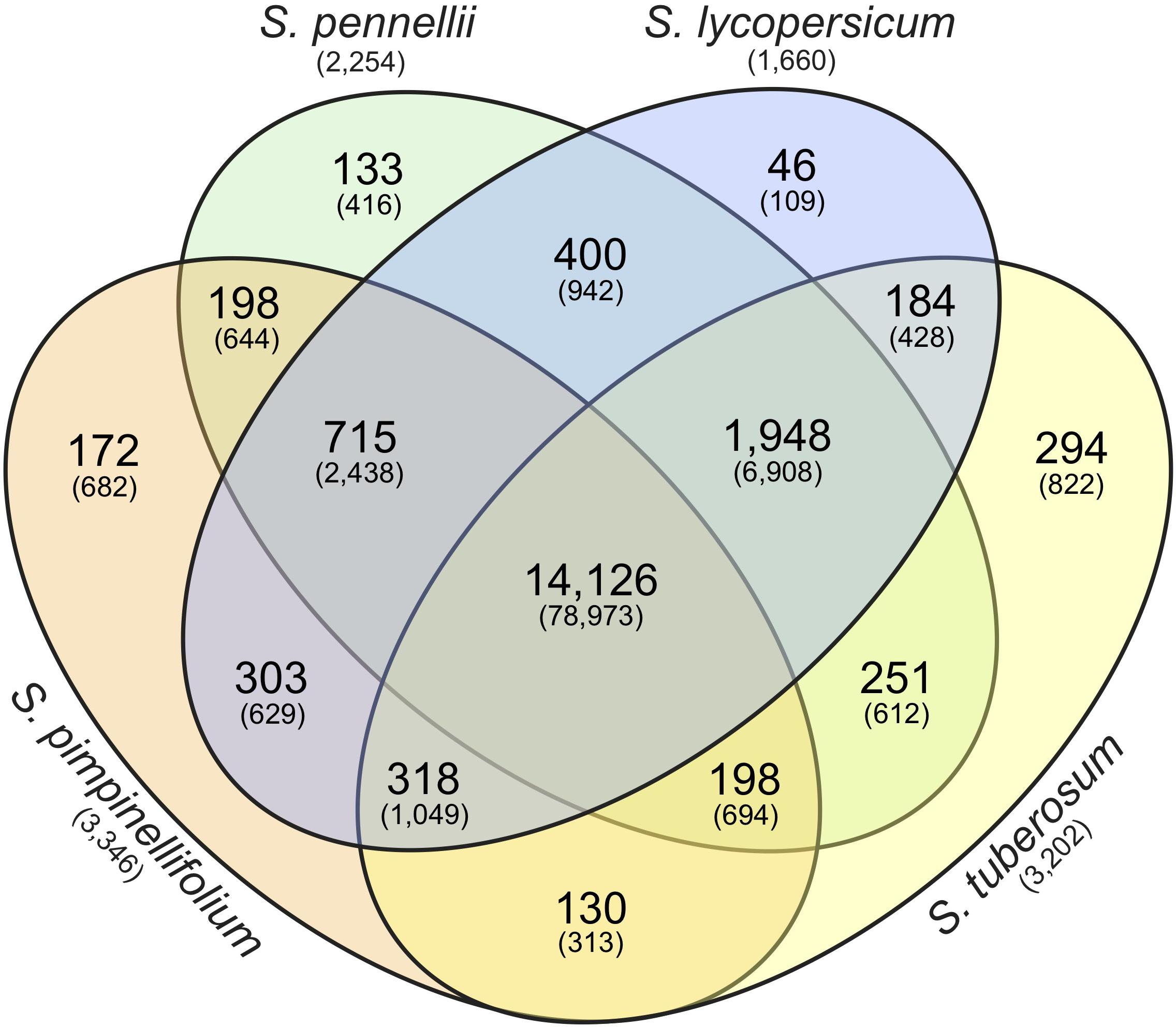
FIGURE 2. Identification of orthologous gene clusters in S. pimpinellifolium, S. pennellii, S. lycopersicum, and S. tuberosum. The Venn diagram represents the number of protein-coding genes and gene clusters shared between, or distinct to, the indicated species. The number in each sector of the diagram indicates the number of homologous clusters and the numbers in parentheses indicate the total number of genes contained within the associated clusters. The numbers in parentheses below the species names indicate the number of species-specific singletons (genes with no homologs).
Of particular interest is the identity of genes encoding the 644 proteins identified as being specific to the two wild tomato species, which are both described as being more tolerant to abiotic stresses than cultivated tomato (e.g., Bolger A. et al., 2014; Rao et al., 2015). This increased tolerance may be due to retention of ancestral Lycopersicon genes that were lost during domestication of cultivated tomato. Within this set of wild tomato-specific genes, we identified 34 S. pimpinellifolium genes with high confidence functional annotations. Specifically, we identified genes with high homology to oxidoreductases [FQR1-like NAD(P)H dehydrogenase (SPi16852.1) and tropinone reductase I (SPi19065.1)], calcium sensors (calmodulin-like protein 3 (SPi15382.1) and WRKY transcription factors (SPi13765.1 and SPi20050.1) that may be involved in abiotic stress tolerance in ‘LA0480’. FQR1-like NAD(P)H dehydrogenases have been linked to ST (Laskowski et al., 2002; Song et al., 2016), while tropinone reductase I has been suggested to play roles in salt stress and drought tolerance (Taji et al., 2004; Shaar-Moshe et al., 2015). The roles of WRKY transcription factors and calmodulins (reviewed by Chen et al., 2012) in abiotic stress tolerance are not well defined, but numerous studies have suggested roles for these proteins in salt, drought, heat and cold tolerance (Reddy et al., 2011; Chen et al., 2012; Niu et al., 2012; Virdi et al., 2015).
Structural Genomic Variation Between S. pimpinellifolium ‘LA0480’ and S. lycopersicum ‘Heinz 1706’
We investigated structural variation between the S. pimpinellifolium and S. lycopersicum genomes by identifying copy number variations (CNVs) due to duplication or deletion of genomic regions in either genome. We mapped S. pimpinellifolium and S. lycopersicum (SRA accession: SRR404081) short reads to the S. lycopersicum reference genome and identified regions of the S. lycopersicum genome with significantly increased coverage of either S. pimpinellifolium or S. lycopersicum reads after normalizing for differences in sequencing depth (Figure 3). CNV windows were identified as 276 bp sections of the S. lycopersicum genome where there was one log2-fold difference between the number of S. pimpinellifolium and S. lycopersicum mapped reads. CNV regions were called where there was at least 1,000 bp of contiguous CNV window coverage. We identified a total of 79,585 CNV regions, with 17,271 and 62,314 regions with higher and lower CNV, respectively, in S. pimpinellifolium (Supplementary Table S15). The average length of these CNV regions is 3,024 bp, covering a total of 241 Mb (29.5%) of the S. lycopersicum genome. In S. pimpinellifolium, we observed substantially more low than high CNV regions, presumably because of the decreased mapping rate of the S. pimpinellifolium reads onto the S. lycopersicum reference genome as a result of sequence divergence between S. pimpinellifolium and S. lycopersicum. Thus, only regions with high CNVs in S. pimpinellifolium were analyzed further.
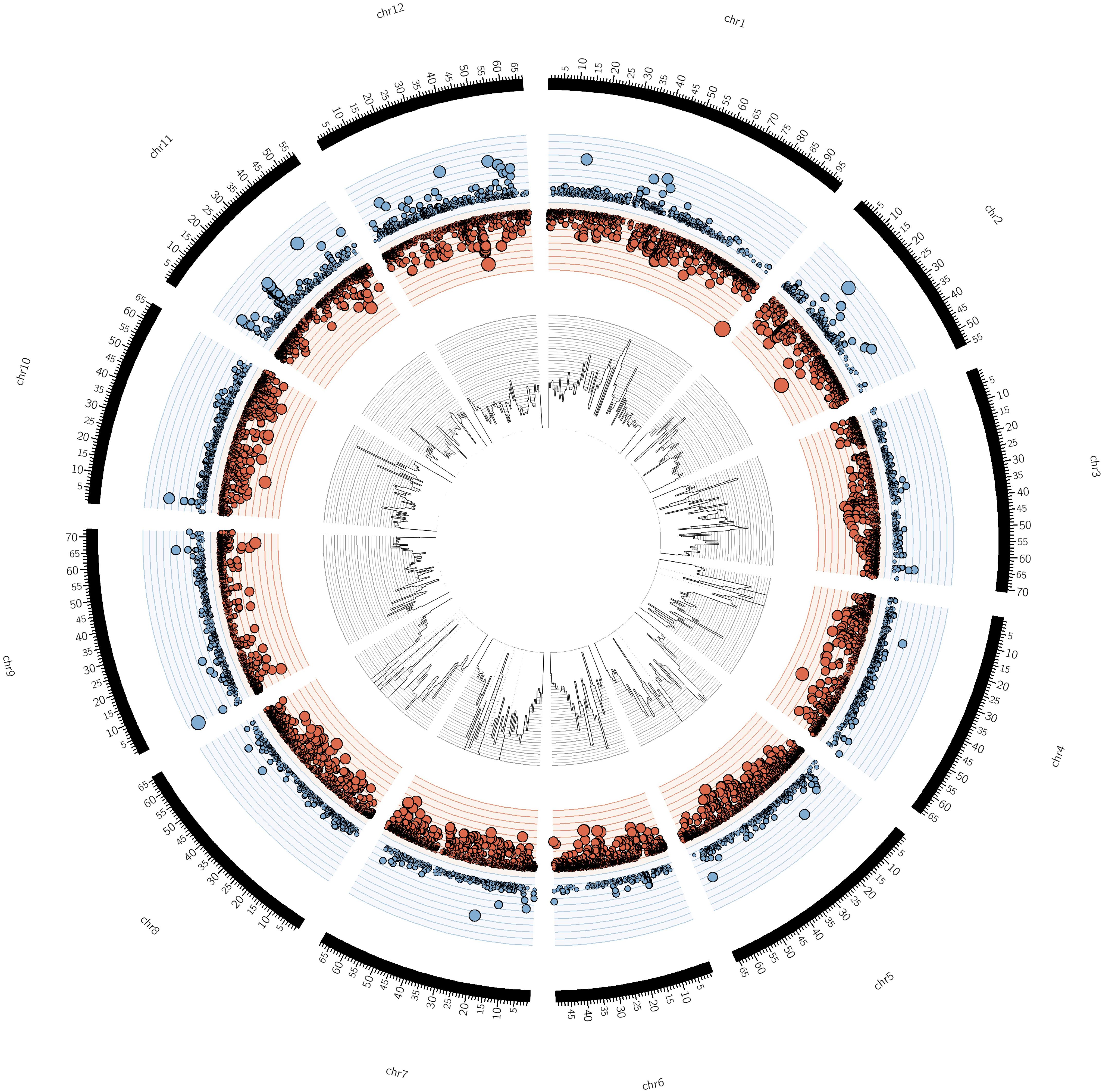
FIGURE 3. Circular representation of S. pimpinellifolium genome structure in comparison with S. lycopersicum. From the outside to the inside: The outer layer represents the 12 chromosomes of S. lycopersicum, with the axis scale in Mb. The second layer (blue/red) represents the scatter plot of copy number variant (CNVs) regions with blue and red circles denoting high and low copy variants, respectively in S. pimpinellifolium relative to S. lycopersicum. The size of the circles is proportional to the absolute value of the log2 CNV. The y-axis scale on the second layer corresponds to the log2 CNV ranging from –10 to 10. The innermost layer represents the histogram of SNPs in 1 Mb bins. The y-axis scale on the innermost layer represents the SNP distribution between the two species, which ranges from 0 to 19,095 SNPs.
We identified 1,809 genes within these S. lycopersicum CNV regions, which is a comparable result to previous studies (Swanson-Wagner et al., 2010; Cao et al., 2011; Zheng et al., 2011). Our analysis also indicated that 29.5% of the S. lycopersicum genome corresponds to CNV regions in S. pimpinellifolium. The proportion of the genome covered with CNV regions in this inter-species comparison is higher than what has been reported in intra-species comparisons (e.g., Belo et al., 2010; Cao et al., 2011; Yu et al., 2011), where values are typically less than 5%. This difference is presumably due to the increased sequence divergence between the two tomato species investigated here.
As the identified CNVs represent regions of the genome that are substantially different between the two species, we investigated the S. lycopersicum genes that are within these CNV regions. We identified a total of 264 S. lycopersicum genes that may have a duplication of the corresponding regions in S. pimpinellifolium (Supplementary Table S16), including genes that may play roles in abiotic or biotic stress tolerance. In particular, we identified one gene related to abiotic stresses tolerance such as drought and salt (LOC543714) (Islam and Wang, 2009); three genes related to leaf rust resistance (LOC101267807, LOC101268104 and LOC101254899) (Qin et al., 2012); and two genes related to late blight resistance (LOC101264157 and LOC101258147) (Nowicki et al., 2012). Additionally, we identified a number of S. lycopersicum transcription factors that may be duplicated in S. pimpinellifolium (e.g., LOC101259210, LOC101259230, LOC101262802, and LOC104649092). The identification of S. lycopersicum genes with roles in abiotic and biotic stress tolerance, that correspond to duplicated S. pimpinellifolium genes, provides candidates for further investigation as these genes that may underlie the increased stress tolerance in S. pimpinellifolium.
S. pimpinellifolium Shows an Enrichment in Classes of Genes Related to Stress Responses
To identify classes of genes that are overrepresented in S. pimpinellifolium, with respect to S. lycopersicum, we annotated both genomes with KEGG Ontology (KO) terms using DEAP and performed an enrichment analysis (Figure 4). We discuss only those KO terms that are enriched in S. pimpinellifolium because the S. lycopersicum and S. pimpinellifolium genome assemblies have different levels of fragmentation, which could lead to the under-representation of some KO terms in the S. pimpinellifolium assembly. While correction for multiple testing using the Benjamini–Hochberg method detected only one significant enrichment, the KO analysis still highlights KO terms that are likely to be biologically relevant (on the basis of high fold change or absolute difference), if not statistically significant. Therefore, we discuss the top-ranking terms, but non-significant KO terms should be considered with caution.
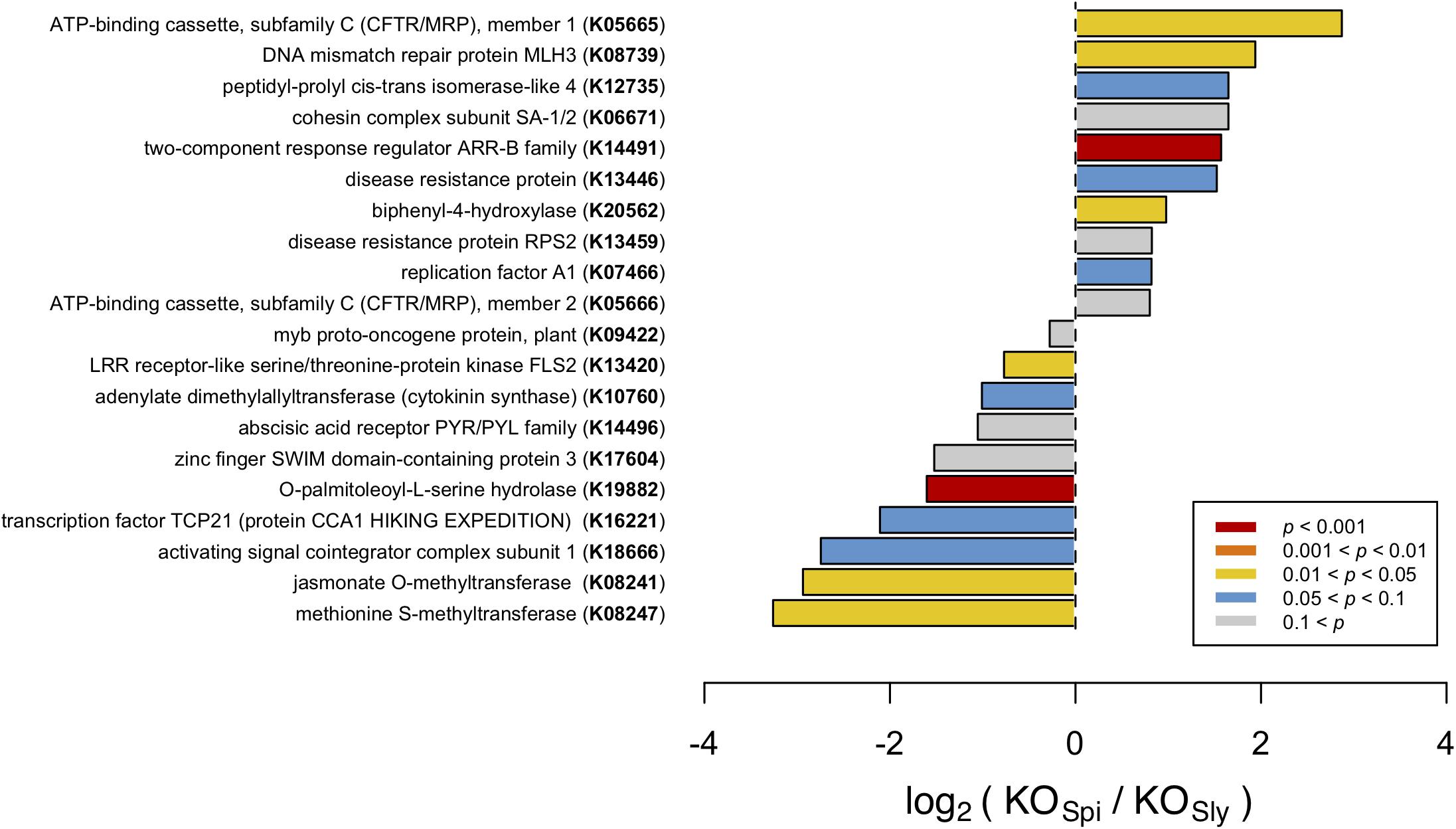
FIGURE 4. Comparison of KO term frequency in S. pimpinellifolium (KOSpi) and S. lycopersicum (KOSly) genomes, presented as the ratio on a log2 scale. Bars are color-coded based on the P-values from a Fisher’s exact test-based enrichment analysis (corrected for multiplicity using the Bonferroni method); the top 20 entries with the highest P values are presented. Entries are ordered based on log2 values.
Our analysis detected multiple KO terms that are enriched in S. pimpinellifolium with respect to S. lycopersicum, several of which, according to KEGG classification, pertain to biological processes associated with biotic and abiotic stress tolerance, such as ‘two-component response regulator ARR-B family’ (K14491; P-value < 3E-05), ‘biphenyl-4-hydroxylase’ (K20562; P-value < 0.025), ‘DNA mismatch repair protein MLH3’ (K08739; P-value < 0.035) and ‘ATP-binding cassette, subfamily C (CFTR/MRP), member 1’ (K05665, P-value < 0.04). To elucidate the downstream biological relevance of such enrichments, we further investigated the functions of genes annotated with the corresponding KOs.
The KO term ‘two-component response regulator ARR-B family’ (K14491) denotes members of the Type-B response regulators (RR-B), a class of transcription factors that are the essential and final effectors in cytokinin (CK) signal transduction (Mason et al., 2005). We observed 51 and 18 occurrences of this KO term in S. pimpinellifolium and S. lycopersicum, respectively. This was the only enriched KO term with a p-value that passed the Bonferroni threshold. RR-Bs have been implicated in pathogen defense, acting as a bridge between cytokinin signaling and salicylic acid and jasmonic acid immune response pathways (Choi et al., 2010; Argueso et al., 2012). Moreover, cytokinins are involved in salinity responses (Tran et al., 2007; Ghanem et al., 2008; Mason et al., 2010), with overexpression of cytokinin biosynthesis genes in S. lycopersicum shown to increase ST (Ghanem et al., 2011; Žižková et al., 2015). These results suggest that the apparent expansion of RR-Bs in S. pimpinellifolium could contribute toward the increased pathogen resistance and stress tolerance of the species.
The ‘Biphenyl-4-hydroxylase’ (K20562) KO term was detected 32 and 17 times in the S. pimpinellifolium and S. lycopersicum genomes, respectively. Biphenyl-4-hydroxylases (B4H) have only recently been identified and cloned in rowan (Sorbus aucuparia) and apple (Malus spp.) and were characterized as cytochrome P450 736A proteins that catalyze the 4-hydroxylation of a biphenyl scaffold toward the biosynthesis of biphenyl phytoalexins such as aucuparin in response to pathogen attack (Khalil et al., 2013; Sircar et al., 2015). Research into biphenyl phytoalexins is somewhat scarce, possibly due to the absence of B4H in the model organism Arabidopsis, with most studies restricted to the Malinae subtribe of the subfamily Amygdaloideae (e.g., apple and pear) (Kokubun et al., 1995; Hüttner et al., 2010; Chizzali and Beerhues, 2012; Chizzali et al., 2012, 2016; Sircar et al., 2015). As such, the observed presence and, indeed, expansion of B4H-related genes in S. pimpinellifolium, could be related to the increased pathogen resistance of S. pimpinellifolium and represents an interesting target for further studies.
In Arabidopsis, AtMLH3 (MutL protein homolog 3) regulates the rate of chromosome crossover during meiosis in reproductive tissues (Franklin et al., 2006; Jackson et al., 2006). We identified 11 and 3 occurrences of the corresponding KO term, ‘DNA mismatch repair protein MLH3’ (K08739), in the S. pimpinellifolium and S. lycopersicum, genomes respectively. A recent study on Crucihimalaya himalaica, an Arabidopsis relative that grows in the extreme environment of the Qinghai-Tibet Plateau, showed that the C. himalaica MLH3 homologue was under strong positive selection and may play a role in the repair of DNA damage caused by high UV radiation (Qiao et al., 2016). This could point toward a role for MLH3-like genes repair of DNA damage (e.g., ROS-induced) caused by abiotic stress in S. pimpinellifolium.
‘ATP-binding cassette, subfamily C (CFTR/MRP), member 1’ (K05665) is enriched in S. pimpinellifolium, which has seven occurrences of this KO term against one in S. lycopersicum, suggesting an expansion of the ATP-binding cassette subfamily C (ABCC) protein superfamily in the wild tomato. ABC proteins encode transmembrane transporters and soluble proteins with crucial functions, and are ubiquitous across all kingdoms of life having a particularly high presence in plants (Andolfo et al., 2015). ABCCs have been implicated in various transport processes in plants, such as vacuolar compartmentalization of glutathione conjugates, glucuronides and anthocyanins, as well as ATP-gated chloride transport, and the regulation of ion channels in guard cells (Martinoia et al., 2002; Goodman et al., 2004; Klein et al., 2006; Suh et al., 2007; Verrier et al., 2008). Although the function of ABC proteins is difficult to determine from sequence similarity alone, we noted that the sole protein annotated with this KO in S. lycopersicum, namely XP_004248540, bears greatest sequence identity to Arabidopsis MRP9 (or ABCC9) and human SUR2 (sulfonylurea receptor 2), which are regulators of potassium channel activity (Rea, 2007). Because this class of genes has established roles in membrane transport, particularly of chloride and potassium, we hypothesize that the high number of ABCC annotated proteins in S. pimpinellifolium might contribute to its higher ST. However, further analyses are required to determine the precise functions of these proteins and the extent of their involvement in such processes.
To complement the results of our comparative genomics analyses, we also undertook a literature-guided approach whereby genes with established roles in ST were examined.
Analysis of Candidate Genes That May Confer Salt Tolerance
Given the higher ST of S. pimpinellifolium and the broad and substantial knowledge of genes that contribute to ST in tomato and other related species, we undertook a CG approach to identify potentially important genes in S. pimpinellifolium. Based on the literature search summarized by Roy et al. (2014), we selected 15 CGs of primary interest (Table 3 and Supplementary Table S17) that have been overexpressed in at least one plant species and were quantifiably shown to increase phenotypic performance under salt-stress conditions. Based on these CGs, we identified the corresponding S. pimpinellifolium orthologs based on published functional reports [e.g., SlNNX1 (Gálvez et al., 2012)], OrthoMCL grouping (Figure 2), and reciprocal BLAST-P (Supplementary Figure S9). Only CGs with orthologs that met these stringent criteria were considered for further analysis.
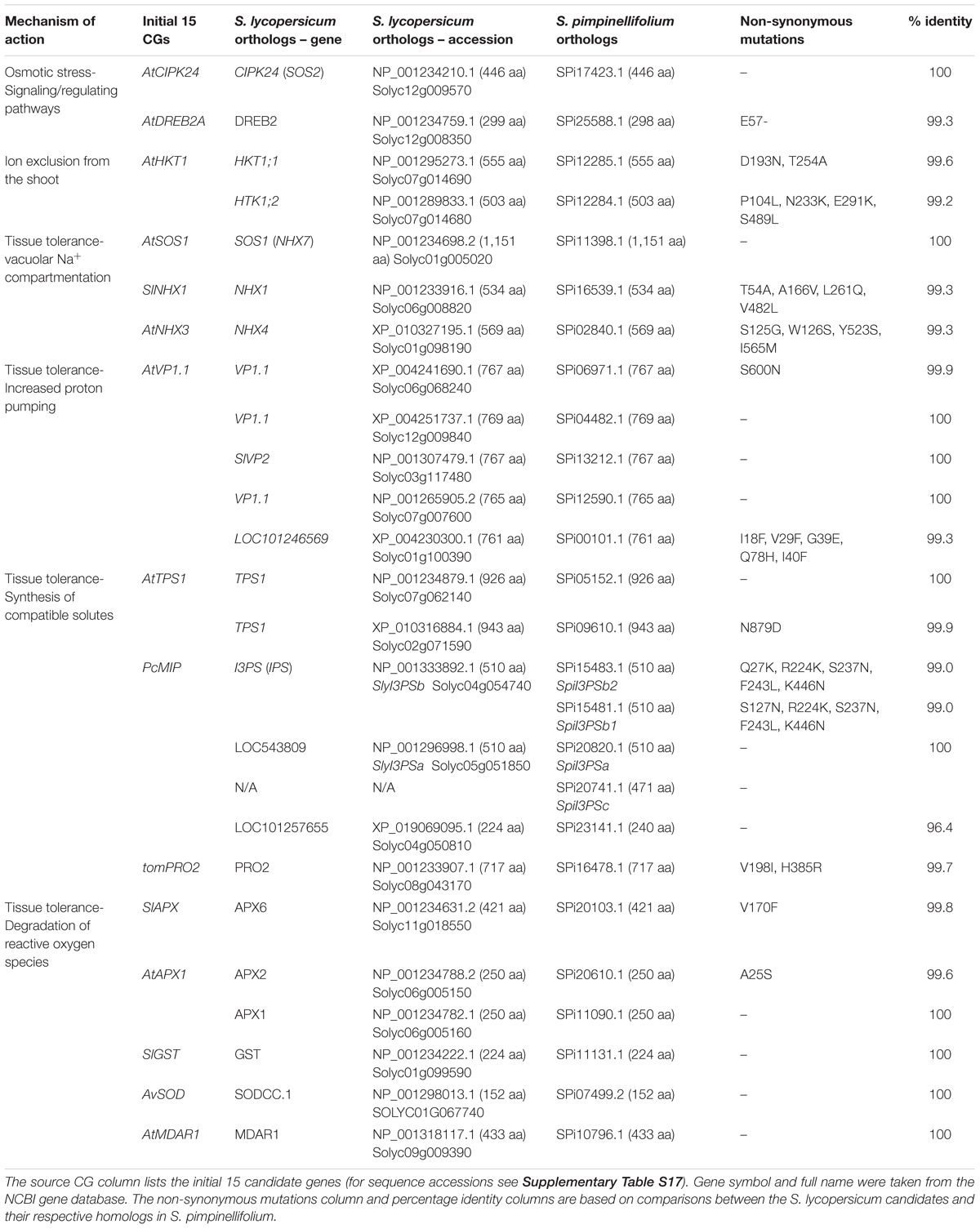
TABLE 3. List of candidate genes for salinity tolerance in S. pimpinellifolium and S. lycopersicum.
We identified 24 putative orthologs in the S. lycopersicum genome that matched the selected 15 CGs. The AtAVP1.1 gene has five orthologs in S. lycopersicum, PcMIP has three, while AtHTK1, AtTPS1, and AtAPX1 have two orthologs each. The remaining ten CGs have one-to-one orthologous relationships. After establishing these S. lycopersicum orthologs, we investigated potential orthologs in S. pimpinellifolium (Table 3) and S. pennellii (Supplementary Table S17 and Supplementary Figures S10–S24). All of the S. lycopersicum genes have an identical number of orthologs in S. pimpinellifolium and S. pennellii except for the inositol-3-phosphate synthase (I3PS) gene. In terms of percentage identity, we observed a high similarity between the S. lycopersicum candidates and the corresponding orthologs in S. pimpinellifolium (>99%, with 11 out of 24 reaching 100% similarity).
In the S. lycopersicum genome, we identified two copies of I3PS (SlyI3PSa and SlyI3PSb) as well as a truncated pseudogene (LOC101257655), while in the S. pimpinellifolium genome we identified four copies of I3PS as well as a truncated pseudogene. S. pimpinellifolium harbors one copy of SpiI3PSa, with 100% identity to SlyI3PSa, two copies of SpiI3PSb (SpiI3PSb1 and SpiI3PSb2), with more than 99% identity to SlyI3PSb, as well as SpiI3PSc (Table 3). At the DNA level, this fourth copy, SpiI3PSc, which is supported by RNA-seq evidence, is highly similar to the SpiI3PSb genes but contains two short frameshifts (not shown). As such, SpiI3PSc putatively encodes a protein product that is shorter than the other SpiI3PS genes due to the deletion of 36 amino acid residues.
To further investigate the relationships between the tomato I3PS genes, we built a DNA-based phylogenetic tree of the I3PS gene family using seven species of the Solanaceae, with Arabidopsis MIPS genes utilized as outgroups (Figure 5 and Supplementary Figure S25). Our results show a clear separation of the gene family into three distinct clades, namely the Arabidopsis, the “A” type and the “B” type clades, with the separation between the two gene types in Solanaceae being supported by high bootstrap values. In the “A-clade,” we observed a single-copy I3PSa gene for all Solanaceae species except for tobacco, which has two IP3Sa genes. The “B-clade” includes a single-copy of the I3PSb gene for all the Solanaceae species except for tobacco and S. pimpinellifolium, which both harbor two copies. This clade also contains SpiI3PSc (SPi20741), which appears to be a divergent I3PSb gene grouping with S. pennellii. However, the placement of these two sequences is unclear, as indicated by the low bootstrap value of 0.56. While the exact placement of SpiI3PSc and SpeI3PSb within the “B-clade” is unclear, alignment of the protein sequences (Supplementary Figure S26) suggests that SpiI3PSc is an atypical form of the I3PS protein.
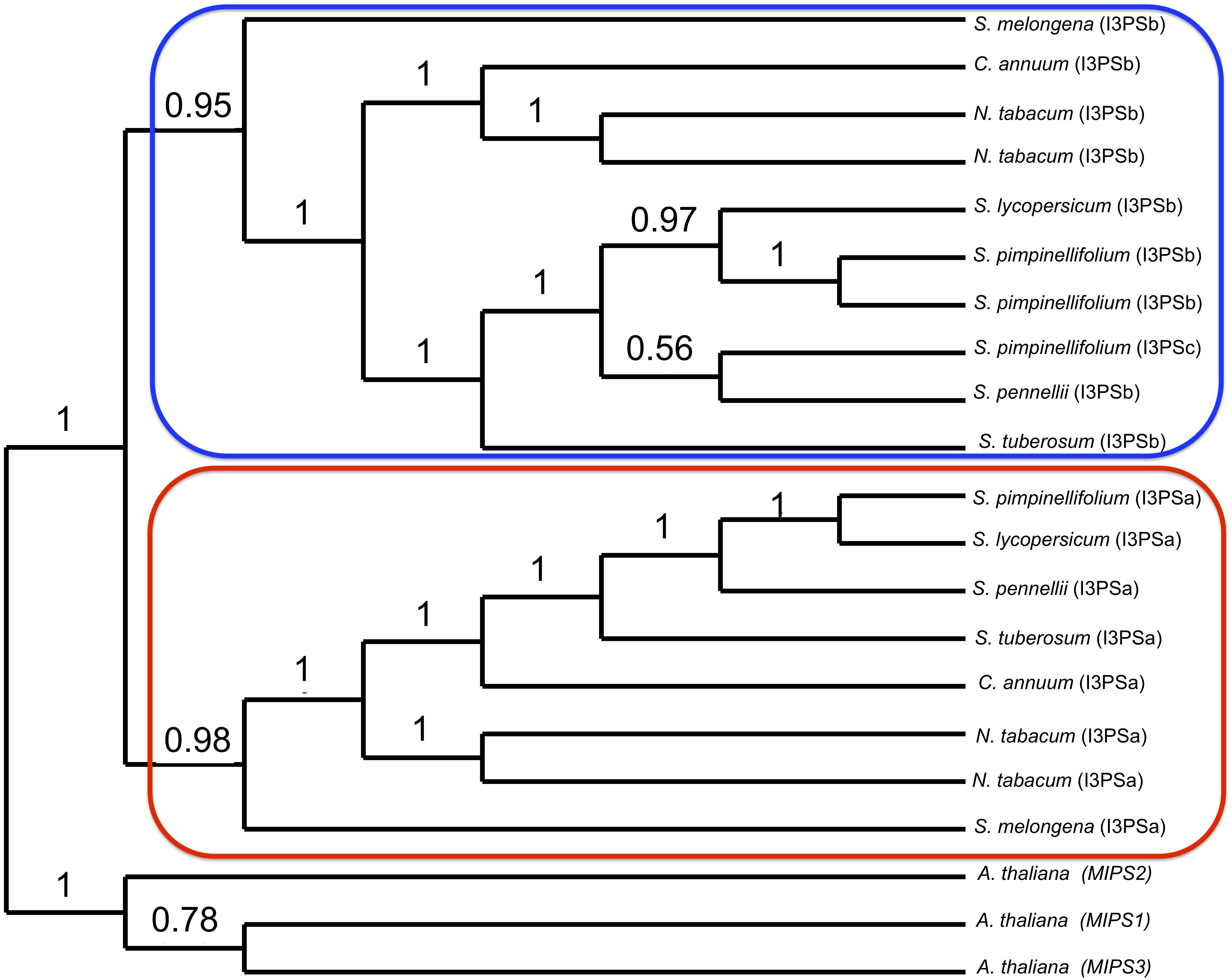
FIGURE 5. Phylogenetic analysis of the inositol-3-phosphate synthase (I3PS) gene family in the Solanaceae family. Node values represent the percentage of 100 bootstrap replicates that support the topology. The I3PSa and I3PSb genes are encircled in red and blue, respectively. A. thaliana MIPS genes were used as outgroups.
I3PS (EC:5.5.1.4) is a key enzyme in the inositol phosphate metabolism, which contributes to cell wall and membrane biogenesis, generates second messengers and signaling molecules, and provides compounds involved in abiotic stress response, phosphate storage in seeds, etc. (Bohnert et al., 1995). I3PS is a NAD+-dependent enzyme that catalyzes the first step in the production of all inositol-containing compounds by converting glucose-6-phosphate (Glc6P) to D-myo-inositol-3-phosphate (Ins3P) (Majumder et al., 2003; Stieglitz et al., 2005), which is subsequently dephosphorylated by the inositol monophosphatase (EC:3.1.3.25) enzyme to myo-inositol (Stieglitz et al., 2007). Myo-inositol is the substrate of the phosphatidylinositol synthase (PIS) (EC:2.7.8.11, CDP-diacylglycerol-inositol-3-phosphatidyltransferase) that forms the phospholipid phosphatidylinositol (PtIns), an abundant phospholipid in non-photosynthetic membranes (Harwood, 1980; Boss and Im, 2012). The inositol moiety of PtIns can be targeted at the 3, 4, or 5 positions by specific kinases, leading to a variety of polyphosphoinositides, such as PtdIns3P, PtdIns4P, PtdIns5P, PtdIns(4, 5)P2, PtdIns(3, 5)P2 and PtdIns(3, 4)P2 (Boss and Im, 2012; Krishnamoorthy et al., 2014). Phosphoinositides are involved in different cellular and developmental processes and contribute responses to various stresses. For instance, it was shown that the overexpression of phosphatidylinositol synthase in maize leads to increased drought tolerance by triggering ABA biosynthesis and modulation of the lipid composition of membranes (Liu et al., 2013). Myo-inositol is also a precursor of soluble signaling molecules, such as InsP6 (myo-Inositol hexakisphosphate also known as phytate) that acts as a second messenger triggering the release of Ca2+ from intracellular stores in guard cells (Lemtiri-Chlieh et al., 2003), as well as ascorbic acid, a powerful reducing agent that is involved in scavenging reactive oxygen species under stress (reviewed by Akram et al., 2017). Moreover, this compound plays a pivotal role in ST by promoting the accumulation of its derivatives, such as D-pinitol and D-ononitol, as compatible solutes and thus protecting cells from osmotic imbalance (e.g., Nelson et al., 1998, 1999). The accumulation of compatible solutes in the cell cytosol is critical for tissue tolerance, a key mechanism that involves the sequestration of Na + ions in the vacuole (Tester and Davenport, 2003; Munns and Tester, 2008).
Transgenic rice, tobacco and Indian mustard plants expressing the I3PS gene (PcINO1) from Porteresia coarctata, a halophytic wild rice, under the CaMV 35S promoter were reported to have enhanced ST due to a substantial increase in inositol levels (Majee et al., 2004; Das-Chatterjee et al., 2006). Likewise, we suggest that the additional SpiI3PS gene copies identified in the wild tomato, S. pimpinellifolium, may contribute to its higher ST when compared to cultivated tomato. However, further studies are necessary to validate the relative importance of each copy of SpiI3PS in S. pimpinellifolium.
Assessing the Role of I3PS in Salinity Tolerance of S. pimpinellifolium
To investigate if the four copies of I3PS in S. pimpinellifolium are functional, we first aligned the sequences of the eight I3PS proteins identified in the Lycopersicon species, namely two copies from S. lycopersicum (SlyI3PSa and SlyI3PSb), two copies from S. pennellii (SpeI3PSa and SpeI3PSb) and four copies from S. pimpinellifolium (SpiI3PSa, SpiI3PSb1, SpiI3PSb2, and SpiI3PSc) (Supplementary Figure S26). The Lycopersicon I3PSs protein sequences align well, except for SpiI3PSc, which showed a deletion of 36 amino acid residues (Figures 6A,B). To evaluate if the four S. pimpinellifolium proteins are catalytically functional, we used computational 3D molecular structure modeling.
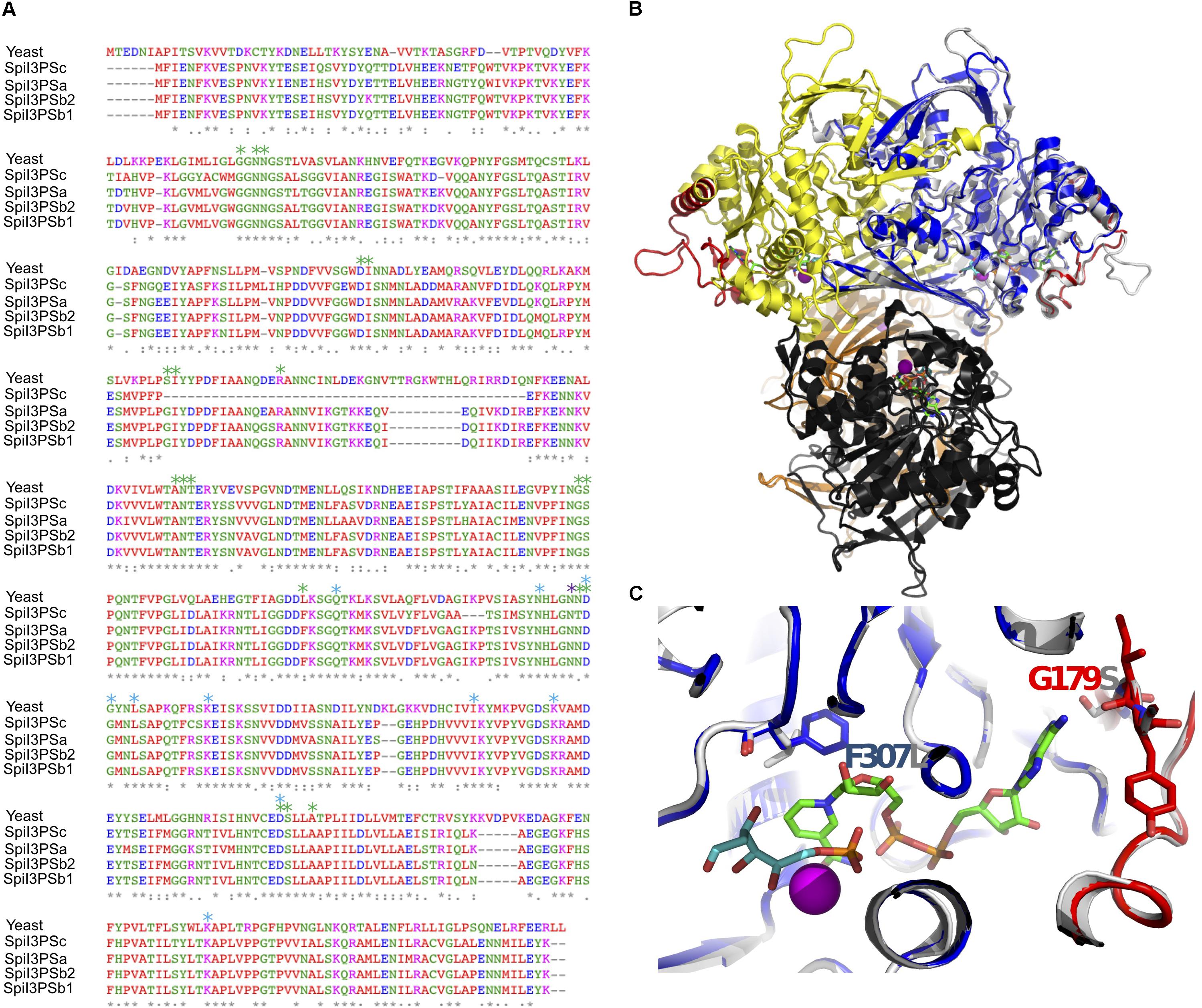
FIGURE 6. Structural evaluation of the catalytic activity of S. pimpinellifolium I3PS proteins. (A) Multiple sequence alignment. Yeast MIP protein. Asterisks label residues involved in binding to NAD (green), DG6 (cyan) and NH4 (magenta). (B) Overall view of the MIP tetramer (PDB: 1jki); individual monomers are shown in gray and yellow (dimer A) and orange and black (dimer B). Red: regions deleted in SpiI3PSc. Blue: homology model of SpiI3PSa superposed. NAD is shown as stick model with green carbons, and DG6 as stick model with cyan carbons, and NH4 as magenta sphere; (C) Detail of the binding site. Colors as in (B). Side chains discussed in the text are shown.
The 3D structures of the four S. pimpinellifolium I3PS proteins were inferred with high confidence by homology modeling based on the ∼55% identical yeast MIP 1-L-myo-inositol-1-phosphate synthase (Stein and Geiger, 2002). When the S. pimpinellifolium I3PS sequences were superimposed onto this tetrameric and catalytically competent model structure (1jki) in complex with nicotinamide adenine dinucleotide (NAD), ammonium (NH4+) and the inhibitor 2-deoxy-glucitol-6-phosphate (DG6), we observed that the short deletions/insertions of one to three residues in SpiI3PSa and SpiI3PSb are distant from the active site (Figure 6A), and thus unlikely to affect the catalytic function. All MIP residues that form the ligand and cofactor binding sites are strictly conserved, except for F307 and G179 (numbering based on SpiI3PSa), which replace, respectively, a leucine and a serine in MIP (Figures 6A,C). Analysis of the homology models strongly suggested that these two substitutions can be accommodated by the 3D environment and do not affect binding and turnover of NAD (Figure 6C). Conversely, SpiI3PSc showed a deletion of 36 residues (red regions in Figure 6B) that could potentially affect the structure of the “lid” that covers the site that binds NAD. While this deletion might not completely abolish catalytic activity, it may result in a lower affinity for NAD and/or a more rapid (but possibly less efficient) substrate turnover. The dimerization and tetramerization interface of yeast MIP was intact and preserved in Spil3PSc, suggesting that all of the I3PS enzymes in S. pimpinellifolium form stable and functional tetramers. We therefore conclude that the I3PS enzymes in S. pimpinellifolium are functional; thus, supporting the critical relevance of I3PS catalytic function.
Given the apparent increase of I3PS gene copy number in S. pimpinellifolium, we examined the broader inositol-related pathway in S. pimpinellifolium using DEAP. We observed that the S. pimpinellifolium genome contains all the genes necessary for 1-phosphatidyl-1D-myo-inositol and myo-inositol cycling according to the inositol phosphate metabolism reference pathway in KEGG5 (verified on 20th of July, 2017). The two key enzymes involved in these cycling processes are Inositol 3-kinase (EC:2.7.1.64) and CDP-diacylglycerol-inositol 3-phosphatidyltransferase (EC:2.7.8.11) (Figure 7), with both enzymes regulating the speed at which the central compound, myo-Inositol, and its derivatives are produced. We also observed that the two entry points into the inositol pathway are present in S. pimpinellifolium inositol pathway, namely: (1) I3PS (EC:5.5.1.4), which catalyzes the conversion of Glc6P to D-myo-inositol-3-phosphate, and (2) phosphatidylinositol-3-phosphatase enzyme (EC:3.1.3.64), which catalyzes the conversion D-myo-inositol 1,3-bisphosphate to myo-inositol 1-phosphate. Thus, the inositol phosphate metabolism pathway in S. pimpinellifolium appears to be complete in terms of entry points and main compounds required for the synthesis of myo-inositol. The gene copy-number between species is the same for the majority of the enzymes present in the inositol pathway, with the notable exceptions being I3PS (EC:5.5.1.4), inositol-phosphate phosphatase (EC: 3.1.3.25) and phosphatidylinositol 4-kinase (EC: 2.7.1.67), which have higher gene copy-number in S. pimpinellifolium relative to S. lycopersicum and S. pennellii (Figure 7, Supplementary Table S19, and Supplementary Figure S27). These changes may not only lead to an increased myo-inositol content but also to modulation of the pattern and concentration of phosphatidylinositols and soluble polyphosphoinositols. Moreover, changes in inositol metabolism will likely impact the concentration of other metabolites (Liu et al., 2013; Kusuda et al., 2015).
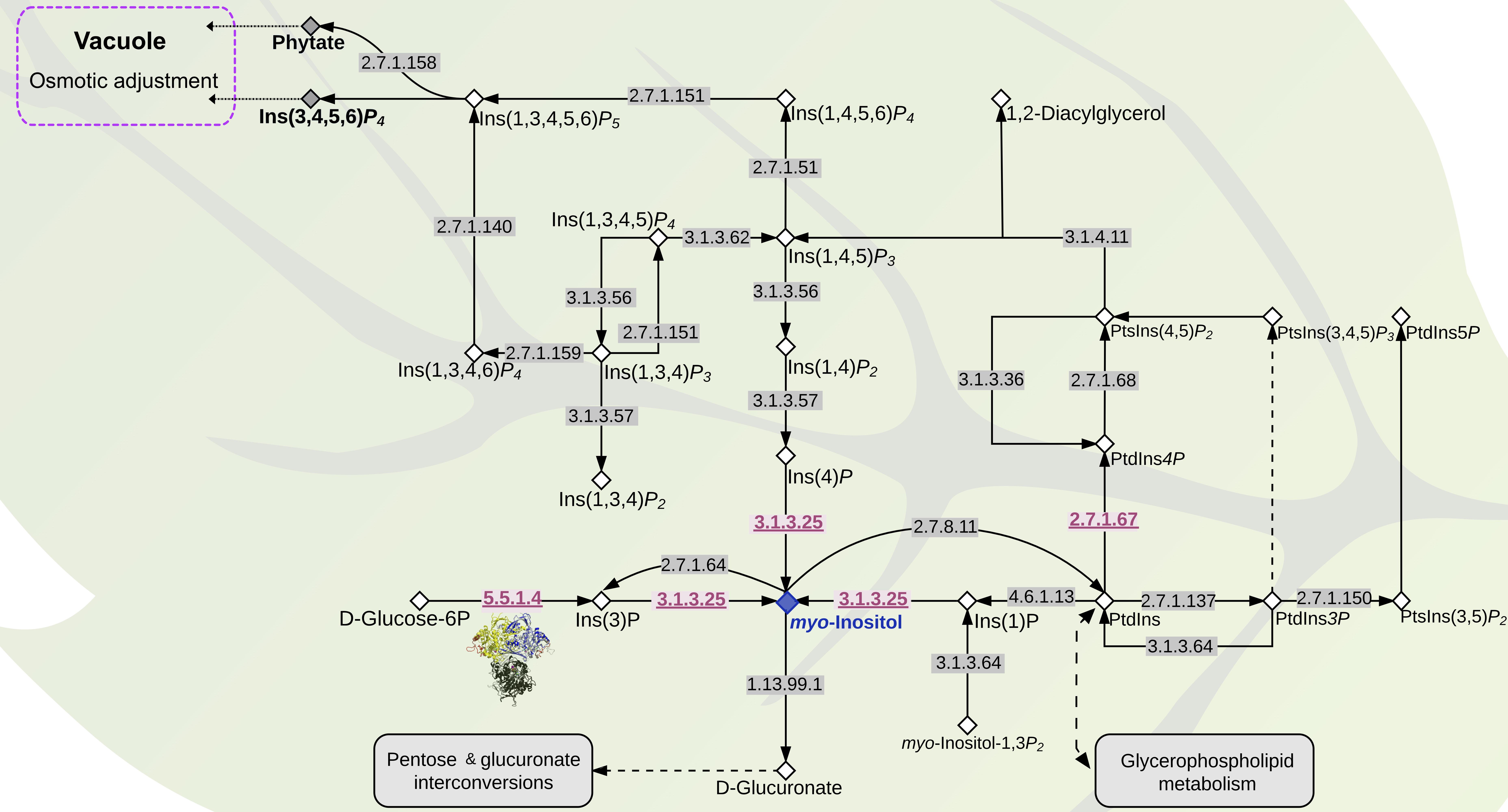
FIGURE 7. The inositol metabolism pathway in S. pimpinellifolium and S. lycopersicum. The pathway was adapted from the KEGG inositol metabolism pathway (map00562- http://www.genome.jp/kegg/pathway/map/map00562.html). Compounds are represented with diamonds, myo-inositol is shown in a blue diamond whereas phytate and Ins(1,3,4,5)P4 are represented by a gray diamond. Enzymes are represented with their EC numbers placed directly on arrows. Enzymes with gene copy numbers higher in S. pimpinellifolium than in S. lycopersicum are underlined and colored in red (Supplementary Table S18). Compound abbreviations were taken from the ChEBI database (Hastings et al., 2013): Ins(1)P: Inositol 1-phosphate; PtdIns: Phosphatidyl-1D-myo-inositol; PtdIns3P: 1-Phosphatidyl-1D-myo-Inositol-3P; PtsIns(3,5)P2: 1-phosphatidyl-1D-myo-inositol 3,5-bisphosphate; PtdIns5P: 1-phosphatidyl-1D-myo-inositol 5-phosphate; PtsIns(3,4,5)P3: 1-phosphatidyl-1D-myo-inositol 3,4,5-trisphosphate; PtsIns(4,5)P2: 1-phosphatidyl-1D-myo-inositol 4,5-bisphosphate; Ins(1,4,5)P3: 1D-myo-inositol 1,4,5-trisphosphate; Ins(1,3,4,5)P4: 1D-myo-inositol 1,3,4,5-P4; Ins(1,4,5,6)P4: 1D-myo-inositol 1,3,4,5-P4; Ins(1,3,4,5,6)P5: 1D-myo-inositol 1,3,4,5,6-P5.
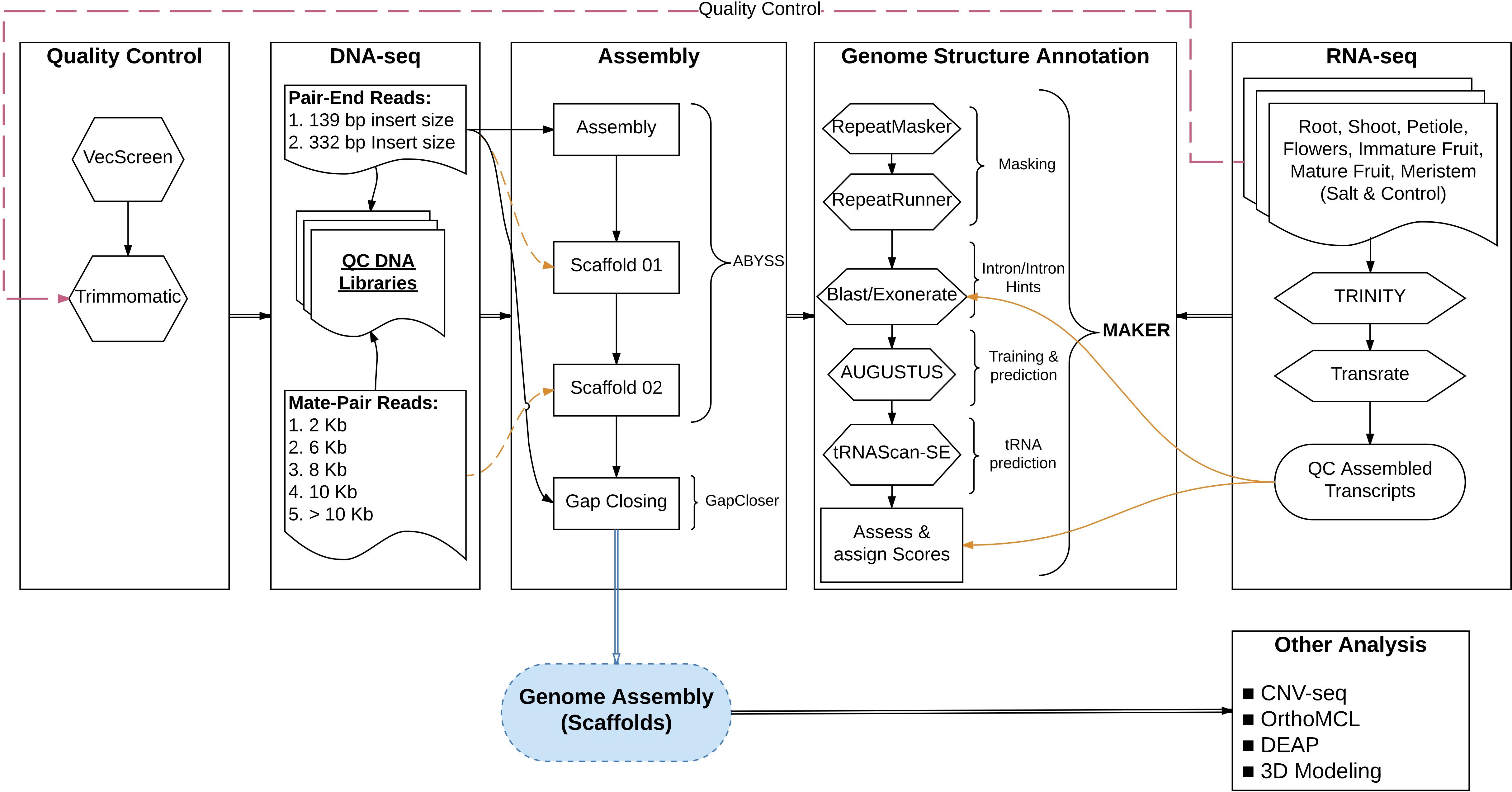
FIGURE 8. Schematic overview of the main tools used for the genome sequence assembly and annotation of S. pimpinellifolium ‘LA0480’. The diagram outlines the workflow and the main tools that were used in the different stages of assembly, gene model annotation and functional annotation.
To assess the expression level of the inositol metabolism pathway genes we explored RNA-seq on leaf samples from S. pimpinellifolium plants grown under control or saline conditions (Supplementary Table S20, and the complete expression dataset for Inositol phosphate metabolism pathway can be found in Data Sheet 3). To normalize the expression levels of the target genes, we examined several tomato reference genes (Supplementary Table S21), and selected tubulin-beta as an adequate reference gene on the basis of its stability between treatments (Supplementary Figure S28). Our analysis suggested that I3PS (EC:5.5.1.4) and inositol-1,4,5-trisphosphate 5-phosphatase gene (EC:3.1.3.56) are up-regulated under salt stress in S. pimpinellifolium (Supplementary Tables S22, S23 and Supplementary Figure S29). In cultivated tomato, a previous study showed that myo-inositol production increases under salt stress (Sacher and Staples, 1985); however, to our knowledge, there is no available expression data of the key genes involved in the inositol pathway under salt stress in this species. The up-regulation of I3PS under salinity has been observed in previous studies using Lotus japonicus (Sanchez et al., 2008a), Mesembryanthemum crystallinum (Nelson et al., 1998) and Populus euphratica (Brosché et al., 2005). The increased accumulation of inositol under salinity stress has been observed in salt-tolerant species such as Eutrema salsugineum (formerly known as Thellungiella halophila), relative to the closely related Arabidopsis thaliana (Gong et al., 2005). This metabolic response resulted from increased expression levels of genes involved in the inositol pathway (Gong et al., 2005). The presence of higher levels of inositol in salt-tolerant species has been suggested as an adaptive response of salt-tolerant species by a metabolic anticipation of stress (Sanchez et al., 2008b).
Next, we analyzed the myo-inositol content in the leaf tissues of ‘LA0480’ and ‘Heinz 1706’ from hydroponically grown plants, under control and saline conditions. Our results showed a significant increase in the amount of myo-inositol produced under saline conditions in both species, but no significant difference in this response was observed between S. pimpinellifolium and S. lycopersicum (Supplementary Figure S30 – bottom panels). The quantification of myo-inositol in both species was unable to shed light on the importance of the extra copy-numbers of I3PS in S. pimpinellifolium. Thus, we hypothesize that the higher ST of S. pimpinellifolium may be related to differences in expression or function of downstream compounds in the inositol pathway, such as different polyphosphoinositides that are involved in signaling pathways, or differences in D-glucuronate that leads to sugar interconversions and/or ascorbic acid levels. For example, in Arabidopsis, the overexpression of the purple acid phosphatase gene (AtPAP15), a phytase that hydrolyzes phytate to myo-inositol and free phosphate, led to the accumulation of ascorbic acid in the shoot and an increase in ST (Zhang, 2008). Additionally, downstream inositol derivatives such as Ins(1,4,5)P3, PtsIns(4,5)P2, and PtdIns4P (Figure 7) have been shown to play a role in abiotic stress signaling (reviewed by Munnik and Nielsen, 2011). For instance, Ins(1,4,5)P3, has been suggested to contribute to drought tolerance in tomato (Khodakovskaya et al., 2010), and could also play a role in ST. Furthermore, phosphoinositide phospholipase C (EC:3.1.4.11) expression has been shown to increase in response to salinity stress in both rice and Arabidopsis and is required for stress-induced Ca2+ signaling and for controlling Na+ accumulation in leaves (Munnik and Nielsen, 2011; Li et al., 2017). Similarly, in S. pimpinellifolium, the higher copy number of phosphatidylinositol 4-kinase (EC:2.7.1.67) (Figure 7 and Supplementary Table S19) and the increased expression of 1-phosphatidylinositol-4-phosphate 5-kinase (EC:2.7.1.68), phosphatidylinositol phospholipase C (EC:3.1.4.11) as well as phosphatidylinositol 4-kinase (EC:2.7.1.67) (Supplementary Table S22) may be involved in the increased ST of S. pimpinellifolium.
Because the accumulation of myo-inositol in the cytoplasm of cells under stress is thought to be related to the tissue tolerance mechanism (Nelson et al., 1999; Munns, 2005; Roy et al., 2014), we investigated the Na and K concentration in the same tissues (Supplementary Figure S30, top and middle panels). We observed that Na accumulates to higher levels in S. pimpinellifolium compared to S. lycopersicum, yet S. pimpinellifolium is more salt-tolerant; thus, reinforcing the idea that tissue tolerance is the main mechanism of ST in this species. Interestingly, other tomato wild relatives besides S. pimpinellifolium, namely S. pennellii, S. peruvianum and S. galapagense also accumulate higher concentrations of Na while being more salt-tolerant than cultivated tomato (Tal, 1971; Santa-Cruz et al., 1999; Almeida et al., 2014), which may also suggest that tissue tolerance could be the main mechanism of ST in these wild species.
Although much research has been conducted into the biochemistry of inositol-related pathways, we are still far from fully understanding their underlying complexity. Specifically, to our knowledge, the link between these pathway derivatives and stress-response mechanisms have not been fully elucidated. As such, further studies on the role of these derivatives in processes such as Ca2+ signaling, osmoprotection and maintenance of membrane integrity are expected to reveal the basis of the higher ST of S. pimpinellifolium compared with S. lycopersicum.
Conclusion
Solanum pimpinellifolium has the potential to increase the genetic diversity of cultivated tomato. Despite the availability of a draft genome sequence of S. pimpinellifolium, limited progress has been made toward unlocking the genetic potential of this species. Our work provides the basis to accelerate the improvement of cultivated tomato by presenting the genome sequence and annotation of the salt-tolerant S. pimpinellifolium accession ‘LA0480’. Our genome analysis shows that S. pimpinellifolium is enriched in genes involved in biotic and abiotic stress responses in comparison to cultivated tomato. Moreover, we demonstrate the increased ST of ‘LA0480,’ and suggest that it could be related to the inositol-related pathways. The expansion of inositol-3-phosphate synthase gene copies in S. pimpinellifolium, which encodes a key enzyme in the inositol pathway, may contribute to its higher ST when compared to S. lycopersicum. Future studies are necessary to validate the role of I3PS in ST in tomato, for instance by using genetic tools (e.g., gene knockout and overexpression) and metabolic profiling by quantifying inositol derivatives. Altogether, our work will enable geneticists and breeders to further explore genes that underlie agronomic traits as well as stress-tolerance mechanisms in S. pimpinellifolium, and to use this knowledge to improve cultivated tomato.
Accession Numbers
Raw data for the DNA-seq, RNA-seq as well as the assembled genome are available under the NCBI BioProject accession PRJNA390234. The assembled genome sequence and annotations are also available through the KAUST library repository at DOI: https://doi.org/10.25781/KAUST-4KWTX. The raw data, assemblies and annotation are available on the SolGenomics website.
Author Contributions
RR, SB, MM, and DL conceived and designed the analyses and managed particular components of the project. RR and SB performed the bioinformatics analyses, which included the compilation of genome scaffolds, and annotation and genomic analyses. MM produced and analyzed the field data, performed the KO enrichment analysis, and oversaw its biological context for data interpretation. RR performed the phylogenetic analyses. DL produced the OrthoMCL results. SB and DL performed the CG analyses. IA and AK developed the computational tool Dragon Eukaryotic Analyses Platform (DEAP). ME and SA-B analyzed the inositol pathway and provided its biological context. SA performed the computational structure–function analysis of the I3PS protein. YP performed the myo-inositol quantification and analyzed the Na and K concentration, and SA-B provided its metabolic context. MS performed the field trial supervision and phenotypic data collection. MM, CM, and SS prepared the materials and undertook sequencing activities. DL and YH contributed to the bioinformatics and genomic analyses. RR, SB, MM, DL, and SN organized the manuscript, analyzed the data, and wrote the article. SN, MT, and VB designed the research, supervised the project, and reviewed the article. All the authors contributed to the writing of the paper.
Funding
This publication is based upon work supported by the King Abdullah University of Science and Technology (KAUST) Office of Sponsored Research (OSR) under Award No. 2302-01-01 and KAUST Base Research Funds to VB Grant No. BAS/1/1606-01-01 and to MT Grant No. BAS/1/1038-01-01.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank John Hanks and Craig Kapfer for the great assistance with the computational resources and the installation of the many bioinformatics tools. Genome sequencing was performed at the biological core laboratories of KAUST. All the computational analyses were performed on Dragon and Snapdragon computer clusters of the Computational Bioscience Research Center (CBRC) at King Abdullah University of Science and Technology (KAUST). We thank Gabriele Fiene (KAUST) for her assistance with the field trial and phenotypic data collection. We also thank Hajime Ohyanagi for his comments on the phylogenetic analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2018.01402/full#supplementary-material
Footnotes
- ^https://solgenomics.net/
- ^https://phytozome.jgi.doe.gov
- ^pymol.org
- ^http://www.cbrc.kaust.edu.sa/dmap
- ^http://www.genome.jp/kegg/pathway/map/map00562.html
References
Akram, N. A., Shafiq, F., and Ashraf, M. (2017). Ascorbic acid - a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Front. Plant Sci. 8:613. doi: 10.3389/fpls.2017.00613
Almeida, P., Feron, R., De Boer, G. J., and De Boer, A. H. (2014). Role of Na +, K +, Cl-, proline and sucrose concentrations in determining salinity tolerance and their correlation with the expression of multiple genes in tomato. AoB Plants 6:plu039. doi: 10.1093/aobpla/plu039
Andolfo, G., Ruocco, M., Di Donato, A., Frusciante, L., Lorito, M., Scala, F., et al. (2015). Genetic variability and evolutionary diversification of membrane ABC transporters in plants. BMC Plant Biol. 15:51. doi: 10.1186/s12870-014-0323-2
Argueso, C. T., Ferreira, F. J., Epple, P., To, J. P., Hutchison, C. E., Schaller, G. E., et al. (2012). Two-component elements mediate interactions between cytokinin and salicylic acid in plant immunity. PLoS Genet. 8:e1002448. doi: 10.1371/journal.pgen.1002448
Arnold, K., Bordoli, L., Kopp, J., and Schwede, T. (2006). The swiss-model workspace: a web-based environment for protein structure homology modelling. Bioinformatics 22, 195–201. doi: 10.1093/bioinformatics/bti770
Azzi, L., Deluche, C., Gévaudant, F., Frangne, N., Delmas, F., Hernould, M., et al. (2015). Fruit growth-related genes in tomato. J. Exp. Bot. 66, 1075–1086. doi: 10.1093/jxb/eru527
Bai, Y. L., and Lindhout, P. (2007). Domestication and breeding of tomatoes: what have we gained and what can we gain in the future? Ann. Bot. 100, 1085–1094. doi: 10.1093/aob/mcm150
Bairoch, A., and Apweiler, R. (2000). The swiss-prot protein sequence database and its supplement TrEMBL in 2000. Nucleic Acids Res. 28, 45–48. doi: 10.1093/nar/28.1.45
Belo, A., Beatty, M. K., Hondred, D., Fengler, K. A., Li, B., and Rafalski, A. (2010). Allelic genome structural variations in maize detected by array comparative genome hybridization. Theor. Appl. Genet. 120, 355–367. doi: 10.1007/s00122-009-1128-9
Blanca, J., Cañizares, J., Cordero, L., Pascual, L., Diez, M. J., and Nuez, F. (2012). Variation revealed by SNP genotyping and morphology provides insight into the origin of the tomato. PLoS One 7:e48198. doi: 10.1371/journal.pone.0048198
Bohnert, H. J., Nelson, D. E., and Jensen, R. G. (1995). Adaptations to environmental stresses. Plant Cell 7, 1099–1111. doi: 10.1105/tpc.7.7.1099
Bolarin, M. C., Estan, M. T., Caro, M., Romero-Aranda, R., and Cuartero, J. (2001). Relationship between tomato fruit growth and fruit osmotic potential under salinity. Plant Sci. 160, 1153–1159. doi: 10.1016/S0168-9452(01)00360-0
Bolarin, M. C., Fernandez, F. G., Cruz, V., and Cuartero, J. (1991). Salinity tolerance in 4 Wild tomato species using vegetative yield salinity response curves. J. Am. Soc. Hortic. Sci. 116, 286–290.
Bolger, A., Scossa, F., Bolger, M. E., Lanz, C., Maumus, F., Tohge, T., et al. (2014). The genome of the stress-tolerant wild tomato species Solanum pennellii. Nat. Genet. 46, 1034–1038. doi: 10.1038/ng.3046
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Boss, W. F., and Im, Y. J. (2012). Phosphoinositide signaling. Annu. Rev. Plant Biol. 63, 409–429. doi: 10.1146/annurev-arplant-042110-103840
Bougouffa, S., Radovanovic, A., Essack, M., and Bajic, V. B. (2014). DEOP: a database on osmoprotectants and associated pathways. Database 2014:bau100. doi: 10.1093/database/bau100
Brooks, C., Nekrasov, V., Lippman, Z. B., and Van Eck, J. (2014). Efficient gene editing in tomato in the first generation using the clustered regularly interspaced short palindromic repeats/CRISPR-associated9 system. Plant Physiol. 166, 1292–1297. doi: 10.1104/pp.114.247577
Brosché, M., Vinocur, B., Alatalo, E. R., Lamminmäki, A., Teichmann, T., Ottow, E. A., et al. (2005). Gene expression and metabolite profiling of Populus euphratica growing in the Negev desert. Genome Biol. 6:R101. doi: 10.1186/gb-2005-6-12-r101
Cagas, C. C., Lee, O. N., Nemoto, K., and Sugiyama, N. (2008). Quantitative trait loci controlling flowering time and related traits in a Solanum lycopersicum × S. pimpinellifolium cross. Sci. Hortic. 116, 144–151. doi: 10.1016/j.scienta.2007.12.003
Cantarel, B. L., Korf, I., Robb, S. M., Parra, G., Ross, E., Moore, B., et al. (2008). MAKER: an easy-to-use annotation pipeline designed for emerging model organism genomes. Genome Res. 18, 188–196. doi: 10.1101/gr.6743907
Cao, J., Schneeberger, K., Ossowski, S., Gunther, T., Bender, S., Fitz, J., et al. (2011). Whole-genome sequencing of multiple Arabidopsis thaliana populations. Nat. Genet. 43, 956–963. doi: 10.1038/ng.911
Capel, C., Yuste-Lisbona, F. J., Lopez-Casado, G., Angosto, T., Cuartero, J., Lozano, R., et al. (2016). Multi-environment QTL mapping reveals genetic architecture of fruit cracking in a tomato RIL Solanum lycopersicum x S. pimpinellifolium population. Theor. Appl. Genet. 130, 213–222. doi: 10.1007/s00122-016-2809-9
Chen, A.-L., Liu, C.-Y., Chen, C.-H., Wang, J.-F., Liao, Y.-C., Chang, C.-H., et al. (2014). Reassessment of QTLs for late blight resistance in the tomato accession L3708 using a restriction site associated DNA (RAD) linkage map and highly aggressive isolates of Phytophthora infestans. PLoS One 9:e96417. doi: 10.1371/journal.pone.0096417
Chen, F., Foolad, M., Hyman, J., Clair, D. S., and Beelaman, R. (1999). Mapping of QTLs for lycopene and other fruit traits in a Lycopersicon esculentum × L. pimpinellifolium cross and comparison of QTLs across tomato species. Mol. Breed. 5, 283–299. doi: 10.1023/A:1009656910457
Chen, L., Song, Y., Li, S., Zhang, L., Zou, C., and Yu, D. (2012). The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta 1819, 120–128. doi: 10.1016/j.bbagrm.2011.09.002
Chevenet, F., Brun, C., Banuls, A. L., Jacq, B., and Christen, R. (2006). Treedyn: towards dynamic graphics and annotations for analyses of trees. BMC Bioinformatics 7:439. doi: 10.1186/1471-2105-7-439
Chizzali, C., and Beerhues, L. (2012). Phytoalexins of the pyrinae: biphenyls and dibenzofurans. Beilstein J. Org. Chem. 8, 613–620. doi: 10.3762/bjoc.8.68
Chizzali, C., Khalil, M. N., Beuerle, T., Schuehly, W., Richter, K., Flachowsky, H., et al. (2012). Formation of biphenyl and dibenzofuran phytoalexins in the transition zones of fire blight-infected stems of Malus domestica cv.‘Holsteiner Cox’and Pyrus communis cv.‘Conference’. Phytochemistry 77, 179–185. doi: 10.1016/j.phytochem.2012.01.023
Chizzali, C., Swiddan, A. K., Abdelaziz, S., Gaid, M., Richter, K., Fischer, T. C., et al. (2016). Expression of biphenyl synthase genes and formation of phytoalexin compounds in three fire blight-infected Pyrus communis cultivars. PLoS One 11:e0158713. doi: 10.1371/journal.pone.0158713
Choi, J., Huh, S. U., Kojima, M., Sakakibara, H., Paek, K.-H., and Hwang, I. (2010). The cytokinin-activated transcription factor ARR2 promotes plant immunity via TGA3/NPR1-dependent salicylic acid signaling in Arabidopsis. Dev. Cell 19, 284–295. doi: 10.1016/j.devcel.2010.07.011
Cuartero, J., and Fernandez-Munoz, R. (1999). Tomato and salinity. Sci. Hortic. 78, 83–125. doi: 10.1016/S0304-4238(98)00191-5
Cuartero, J., Yeo, A. R., and Flowers, T. J. (1992). Selection of donors for salt-tolerance in tomato using physiological traits. New Phytol. 121, 63–69. doi: 10.1111/j.1469-8137.1992.tb01093.x
Das-Chatterjee, A., Goswami, L., Maitra, S., Dastidar, K. G., Ray, S., and Majumder, A. L. (2006). Introgression of a novel salt-tolerant L-myo-inositol 1-phosphate synthase from Porteresia coarctata (Roxb.) Tateoka (PcINO1) confers salt tolerance to evolutionary diverse organisms. FEBS Lett. 580, 3980–3988. doi: 10.1016/j.febslet.2006.06.033
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36, W465–W469. doi: 10.1093/nar/gkn180
Doganlar, S., Frary, A., Ku, H.-M., and Tanksley, S. D. (2002). Mapping quantitative trait loci in inbred backcross lines of Lycopersicon pimpinellifolium (LA1589). Genome 45, 1189–1202. doi: 10.1139/g02-091
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Estan, M. T., Villalta, I., Bolarin, M. C., Carbonell, E. A., and Asins, M. J. (2009). Identification of fruit yield loci controlling the salt tolerance conferred by Solanum rootstocks. Theor. Appl. Genet. 118, 305–312. doi: 10.1007/s00122-008-0900-6
Fernandez-Pozo, N., Menda, N., Edwards, J. D., Saha, S., Tecle, I. Y., Strickler, S. R., et al. (2015). The sol genomics network (SGN)-from genotype to phenotype to breeding. Nucleic Acids Res. 43, D1036–D1041. doi: 10.1093/nar/gku1195
Foolad, M. (1999). Comparison of salt tolerance during seed germination and vegetative growth in tomato by QTL mapping. Genome 42, 727–734. doi: 10.1139/g98-163
Foolad, M., and Chen, F. (1999). RFLP mapping of QTLs conferring salt tolerance during the vegetative stage in tomato. Theor. Appl. Genet. 99, 235–243. doi: 10.1007/s001220051229
Foolad, M., Chen, F., and Lin, G. (1998). RFLP mapping of QTLs conferring salt tolerance during germination in an interspecific cross of tomato. Theor. Appl. Genet. 97, 1133–1144. doi: 10.1007/s001220051002
Foolad, M., Zhang, L., and Lin, G. (2001). Identification and validation of QTLs for salt tolerance during vegetative growth in tomato by selective genotyping. Genome 44, 444–454. doi: 10.1139/g01-030
Foolad, M. R., and Lin, G. Y. (1997). Genetic potential for salt tolerance during germination in Lycopersicon species. HortScience 32, 296–300.
Franklin, F., Higgins, J., Sanchez-Moran, E., Armstrong, S., Osman, K., Jackson, N., et al. (2006). Control of meiotic recombination in Arabidopsis: role of the MutL and MutS homologues. Biochem. Soc. Trans. 34, 542–544. doi: 10.1042/BST0340542
Gálvez, F. J., Baghour, M., Hao, G., Cagnac, O., Rodríguez-Rosales, M. P., and Venema, K. (2012). Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol. Biochem. 51, 109–115. doi: 10.1016/j.plaphy.2011.10.012
Ghanem, M. E., Albacete, A., Martinez-Andujar, C., Acosta, M., Romero-Aranda, R., Dodd, I. C., et al. (2008). Hormonal changes during salinity-induced leaf senescence in tomato (Solanum lycopersicum L.). J. Exp. Bot. 59, 3039–3050. doi: 10.1093/jxb/ern153
Ghanem, M. E., Albacete, A., Smigocki, A. C., Frébort, I., Pospíšilová, H., Martínez-Andújar, C., et al. (2011). Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 62, 125–140. doi: 10.1093/jxb/erq266
Gianfagna, T., Logendra, L., Durner, E., and Janes, H. (1997). Improving tomato harvest index by controlling crop height and side shoot production. Life Support Biosph. Sci. 5, 255–261.
Gilmour, A. R., Gogel, B., Cullis, B., Thompson, R., and Butler, D. (2009). ASReml User Guide Release 3.0. Hemel Hempstead: VSN International Ltd.
Gong, Q., Li, P., Ma, S., Indu Rupassara, S., and Bohnert, H. J. (2005). Salinity stress adaptation competence in the extremophile Thellungiella halophila in comparison with its relative Arabidopsis thaliana. Plant J. 44, 826–839. doi: 10.1111/j.1365-313X.2005.02587.x
Goodman, C. D., Casati, P., and Walbot, V. (2004). A multidrug resistance–associated protein involved in anthocyanin transport in Zea mays. Plant Cell 16, 1812–1826. doi: 10.1105/tpc.022574
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Harwood, J. L. (1980). “Plant acyl lipids: structure, distribution and analysis,” in The Biochemistry of Plants, ed. P. K. Stumpf (New York, NY: Academic Press), 1–55.
Hastings, J., De Matos, P., Dekker, A., Ennis, M., Harsha, B., Kale, N., et al. (2013). The ChEBI reference database and ontology for biologically relevant chemistry: enhancements for 2013. Nucleic Acids Res. 41, D456–D463. doi: 10.1093/nar/gks1146
Heberle, H., Meirelles, G. V., Da Silva, F. R., Telles, G. P., and Minghim, R. (2015). InteractiVenn: a web-based tool for the analysis of sets through venn diagrams. BMC Bioinformatics 16:169. doi: 10.1186/s12859-015-0611-3
Hüttner, C., Beuerle, T., Scharnhop, H., Ernst, L., and Beerhues, L. (2010). Differential effect of elicitors on biphenyl and dibenzofuran formation in Sorbus aucuparia cell cultures. J. Agric. Food Chem. 58, 11977–11984. doi: 10.1021/jf1026857
Islam, M. S., and Wang, M. H. (2009). Expression of dehydration responsive element-binding protein-3 (DREB3) under different abiotic stresses in tomato. BMB Rep. 42, 611–616. doi: 10.5483/BMBRep.2009.42.9.611
Jackson, N., Sanchez-Moran, E., Buckling, E., Armstrong, S. J., Jones, G. H., and Franklin, F. C. H. (2006). Reduced meiotic crossovers and delayed prophase I progression in AtMLH3-deficient Arabidopsis. EMBO J. 25, 1315–1323. doi: 10.1038/sj.emboj.7600992
Jones, P., Binns, D., Chang, H. Y., Fraser, M., Li, W., Mcanulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Kevei, Z., King, R. C., Mohareb, F., Sergeant, M. J., Awan, S. Z., and Thompson, A. J. (2015). Resequencing at ≥ 40-fold depth of the parental genomes of a Solanum lycopersicum × S. pimpinellifolium recombinant inbred line population and characterization of frame-shift InDels that are highly likely to perturb protein function. G3 5, 971–981. doi: 10.1534/g3.114.016121
Khalil, M. N., Beuerle, T., Müller, A., Ernst, L., Bhavanam, V. B., Liu, B., et al. (2013). Biosynthesis of the biphenyl phytoalexin aucuparin in Sorbus aucuparia cell cultures treated with Venturia inaequalis. Phytochemistry 96, 101–109. doi: 10.1016/j.phytochem.2013.09.003
Khodakovskaya, M., Sword, C., Wu, Q., Perera, I. Y., Boss, W. F., Brown, C. S., et al. (2010). Increasing inositol (1,4,5)-trisphosphate metabolism affects drought tolerance, carbohydrate metabolism and phosphate-sensitive biomass increases in tomato. Plant Biotechnol. J. 8, 170–183. doi: 10.1111/j.1467-7652.2009.00472.x
Klein, M., Burla, B., and Martinoia, E. (2006). The multidrug resistance-associated protein (MRP/ABCC) subfamily of ATP-binding cassette transporters in plants. FEBS Lett. 580, 1112–1122. doi: 10.1016/j.febslet.2005.11.056
Kokubun, T., Harborne, J. B., Eagles, J., and Waterman, P. G. (1995). Dibenzofuran phytoalexins from the sapwood tissue of Photinia, Pyracantha and Crataegus species. Phytochemistry 39, 1033–1037. doi: 10.1016/0031-9422(95)00128-T
Krishnamoorthy, P., Sanchez-Rodriguez, C., Heilmann, I., and Persson, S. (2014). Regulatory roles of phosphoinositides in membrane trafficking and their potential impact on cell-wall synthesis and re-modelling. Ann. Bot. 114, 1049–1057. doi: 10.1093/aob/mcu055
Krzywinski, M., Schein, J., Birol, I., Connors, J., Gascoyne, R., Horsman, D., et al. (2009). Circos: an information aesthetic for comparative genomics. Genome Res. 19, 1639–1645. doi: 10.1101/gr.092759.109
Kusuda, H., Koga, W., Kusano, M., Oikawa, A., Saito, K., Hirai, M. Y., et al. (2015). Ectopic expression of myo-inositol 3-phosphate synthase induces a wide range of metabolic changes and confers salt tolerance in rice. Plant Sci. 232, 49–56. doi: 10.1016/j.plantsci.2014.12.009
Laskowski, M. J., Dreher, K. A., Gehring, M. A., Abel, S., Gensler, A. L., and Sussex, I. M. (2002). FQR1, a novel primary auxin-response gene, encodes a flavin mononucleotide-binding quinone reductase. Plant Physiol. 128, 578–590. doi: 10.1104/pp.010581
Lemtiri-Chlieh, F., Macrobbie, E. A., Webb, A. A., Manison, N. F., Brownlee, C., Skepper, J. N., et al. (2003). Inositol hexakisphosphate mobilizes an endomembrane store of calcium in guard cells. Proc. Natl. Acad. Sci. U.S.A. 100, 10091–10095. doi: 10.1073/pnas.1133289100
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows–Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Li, L., Stoeckert, C. J. Jr., and Roos, D. S. (2003). OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res. 13, 2178–2189. doi: 10.1101/gr.1224503
Li, L., Wang, F., Yan, P., Jing, W., Zhang, C., Kudla, J., et al. (2017). A phosphoinositide-specific phospholipase C pathway elicits stress-induced Ca2 + signals and confers salt tolerance to rice. New Phytol. 214, 1172–1187. doi: 10.1111/nph.14426
Lin, K.-H., Yeh, W.-L., Chen, H.-M., and Lo, H.-F. (2010). Quantitative trait loci influencing fruit-related characteristics of tomato grown in high-temperature conditions. Euphytica 174, 119–135. doi: 10.1007/s10681-010-0147-6
Liu, M., Gomes, B. L., Mila, I., Purgatto, E., Peres, L. E. P., Frasse, P., et al. (2016). Comprehensive profiling of ethylene response factor expression identifies ripening-associated ERF genes and their link to key regulators of fruit ripening in tomato. Plant Physiol. 170, 1732–1744. doi: 10.1104/pp.15.01859
Liu, X., Zhai, S., Zhao, Y., Sun, B., Liu, C., Yang, A., et al. (2013). Overexpression of the phosphatidylinositol synthase gene (ZmPIS) conferring drought stress tolerance by altering membrane lipid composition and increasing ABA synthesis in maize. Plant Cell Environ. 36, 1037–1055. doi: 10.1111/pce.12040
Lowe, T. M., and Eddy, S. R. (1997). tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 25, 955–964. doi: 10.1093/nar/25.5.0955
Luckwill, L. (1943). The Genus Lycopersicon, An Historical, Biological and Taxonomic Survey of the Wild and Cultivated Tomatoes. Aberdeen: Aberdeen Univ.Press.
Luo, R., Liu, B., Xie, Y., Li, Z., Huang, W., Yuan, J., et al. (2012). SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. Gigascience 1:18. doi: 10.1186/2047-217X-1-18
Majee, M., Maitra, S., Dastidar, K. G., Pattnaik, S., Chatterjee, A., Hait, N. C., et al. (2004). A novel salt-tolerant L-myo-inositol-1-phosphate synthase from Porteresia coarctata (Roxb.) Tateoka, a halophytic wild rice: molecular cloning, bacterial overexpression, characterization, and functional introgression into tobacco-conferring salt tolerance phenotype. J. Biol. Chem. 279, 28539–28552. doi: 10.1074/jbc.M310138200
Majumder, A. L., Chatterjee, A., Ghosh Dastidar, K., and Majee, M. (2003). Diversification and evolution of L-myo-inositol 1-phosphate synthase. FEBS Lett. 553, 3–10. doi: 10.1016/S0014-5793(03)00974-8
Martinoia, E., Klein, M., Geisler, M., Bovet, L., Forestier, C., Kolukisaoglu, Ü, et al. (2002). Multifunctionality of plant ABC transporters–more than just detoxifiers. Planta 214, 345–355. doi: 10.1007/s004250100661
Mason, M. G., Jha, D., Salt, D. E., Tester, M., Hill, K., Kieber, J. J., et al. (2010). Type-B response regulators ARR1 and ARR12 regulate expression of AtHKT1;1 and accumulation of sodium in Arabidopsis shoots. Plant J. 64, 753–763. doi: 10.1111/j.1365-313X.2010.04366.x
Mason, M. G., Mathews, D. E., Argyros, D. A., Maxwell, B. B., Kieber, J. J., Alonso, J. M., et al. (2005). Multiple type-B response regulators mediate cytokinin signal transduction in Arabidopsis. Plant Cell 17, 3007–3018. doi: 10.1105/tpc.105.035451
Miller, J. C., and Tanksley, S. D. (1990). RFLP analysis of phylogenetic relationships and genetic variation in the genus Lycopersicon. Theor. Appl. Genet. 80, 437–448. doi: 10.1007/BF00226743
Munnik, T., and Nielsen, E. (2011). Green light for polyphosphoinositide signals in plants. Curr. Opin. Plant Biol. 14, 489–497. doi: 10.1016/j.pbi.2011.06.007
Munns, R. (2005). Genes and salt tolerance: bringing them together. New Phytol. 167, 645–663. doi: 10.1111/j.1469-8137.2005.01487.x
Munns, R., and Tester, M. (2008). Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59, 651–681. doi: 10.1146/annurev.arplant.59.032607.092911
Nakano, H., Sasaki, K., Mine, Y., Takahata, K., Lee, O., and Sugiyama, N. (2016). Quantitative trait loci (QTL) controlling plant architecture traits in a Solanum lycopersicum × S. pimpinellifolium cross. Euphytica 211, 353–367. doi: 10.1007/s10681-016-1744-9
Nelson, D. E., Koukoumanos, M., and Bohnert, H. J. (1999). Myo-inositol-dependent sodium uptake in ice plant. Plant Physiol. 119, 165–172. doi: 10.1104/pp.119.1.165
Nelson, D. E., Rammesmayer, G., and Bohnert, H. J. (1998). Regulation of cell-specific inositol metabolism and transport in plant salinity tolerance. Plant Cell 10, 753–764. doi: 10.2307/3870662
Ni, J., Bai, S., Gao, L., Qian, M., Zhong, L., and Teng, Y. (2017). Identification, classification, and transcription profiles of the B-type response regulator family in pear. PLoS One 12:e0171523. doi: 10.1371/journal.pone.0171523
Niu, C. F., Wei, W., Zhou, Q. Y., Tian, A. G., Hao, Y. J., Zhang, W. K., et al. (2012). Wheat WRKY genes TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Environ. 35, 1156–1170. doi: 10.1111/j.1365-3040.2012.02480.x
Nowicki, M., Foolad, M. R., Nowakowska, M., and Kozik, E. U. (2012). Potato and tomato late blight caused by Phytophthora infestans: an overview of pathology and resistance breeding. Plant Dis. 96, 4–17. doi: 10.1094/PDIS-05-11-0458
Pedley, K. F., and Martin, G. B. (2003). Molecular basis of Pto-mediated resistance to bacterial speck disease in tomato. Annu. Rev. Phytopathol. 41, 215–243. doi: 10.1146/annurev.phyto.41.121602.143032
Peralta, I., and Spooner, D. (2000). Classification of wild tomatoes: a review. Kurtziana 28, 45–54.
Peralta, I. E., Knapp, S., and Spooner, D. M. (2005). New species of wild tomatoes (Solanum section Lycopersicon: Solanaceae) from Northern Peru. Syst. Bot. 30, 424–434. doi: 10.1600/0363644054223657
Peralta, I. E., Spooner, D. M., and Knapp, S. (2008). Taxonomy of tomatoes: a revision of wild tomatoes (Solanum section Lycopersicon) and their outgroup relatives in sections Juglandifolia and Lycopersicoides. Syst. Bot. Monogr. 84, 1–186.
Pitblado, R., and Kerr, E. (1979). Resistance to bacterial speck (Pseudomonas tomato) in tomato. Acta Hortic. 100, 379–382. doi: 10.17660/ActaHortic.1980.100.40
Qiao, Q., Wang, Q., Han, X., Guan, Y., Sun, H., Zhong, Y., et al. (2016). Transcriptome sequencing of Crucihimalaya himalaica (Brassicaceae) reveals how Arabidopsis close relative adapt to the qinghai-tibet plateau. Sci. Rep. 6:21729. doi: 10.1038/srep21729
Qin, B., Chen, T., Cao, A., Wang, H., Xing, L., Ling, H., et al. (2012). Cloning of a conserved receptor-like protein kinase gene and its use as a functional marker for homoeologous group-2 chromosomes of the triticeae species. PLoS One 7:e49718. doi: 10.1371/journal.pone.0049718
Rao, E. S., Kadirvel, P., Symonds, R. C., and Ebert, A. W. (2013). Relationship between survival and yield related traits in Solanum pimpinellifolium under salt stress. Euphytica 190, 215–228. doi: 10.1007/s10681-012-0801-2
Rao, E. S., Kadirvel, P., Symonds, R. C., Geethanjali, S., Thontadarya, R. N., and Ebert, A. W. (2015). Variations in DREB1A and VP1.1 genes show association with salt tolerance traits in wild tomato (Solanum pimpinellifolium). PLoS One 10:e0132535. doi: 10.1371/journal.pone.0132535
Rea, P. A. (2007). Plant ATP-binding cassette transporters. Annu. Rev. Plant Biol. 58, 347–375. doi: 10.1146/annurev.arplant.57.032905.105406
Reddy, A. S., Ali, G. S., Celesnik, H., and Day, I. S. (2011). Coping with stresses: roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell 23, 2010–2032. doi: 10.1105/tpc.111.084988
Rick, C. M., Fobes, J. F., and Holle, M. (1977). Genetic variation in Lycopersicon pimpinellifolium: evidence of evolutionary change in mating systems. Plant Syst. Evol. 127, 139–170. doi: 10.1007/BF00984147
Rick, C. M., Holle, M., and Thorp, R. W. (1978). Rates of cross-pollination in Lycopersicon pimpinellifolium: impact of genetic variation in floral characters. Plant Syst. Evol. 129, 31–44. doi: 10.1007/BF00988982
Roy, S. J., Negrão, S., and Tester, M. (2014). Salt resistant crop plants. Curr. Opin. Biotechnol. 26, 115–124. doi: 10.1016/j.copbio.2013.12.004
Sacher, R. F., and Staples, R. C. (1985). Inositol and sugars in adaptation of tomato to salt. Plant Physiol. 77, 206–210. doi: 10.1104/pp.77.1.206
Salinas, M., Capel, C., Alba, J. M., Mora, B., Cuartero, J., Fernandez-Munoz, R., et al. (2013). Genetic mapping of two QTL from the wild tomato Solanum pimpinellifolium L. controlling resistance against two-spotted spider mite (Tetranychus urticae Koch). Theor. Appl. Genet. 126, 83–92. doi: 10.1007/s00122-012-1961-0
Sanchez, D. H., Lippold, F., Redestig, H., Hannah, M. A., Erban, A., Kramer, U., et al. (2008a). Integrative functional genomics of salt acclimatization in the model legume Lotus japonicus. Plant J. 53, 973–987.
Sanchez, D. H., Siahpoosh, M. R., Roessner, U., Udvardi, M., and Kopka, J. (2008b). Plant metabolomics reveals conserved and divergent metabolic responses to salinity. Physiol. Plant. 132, 209–219. doi: 10.1111/j.1399-3054.2007.00993.x
Santa-Cruz, A., Acosta, M., Rus, A., and Bolarin, M. C. (1999). Short-term salt tolerance mechanisms in differentially salt tolerant tomato species. Plant Physiol. Biochem. 37, 65–71. doi: 10.1016/S0981-9428(99)80068-0
Shaar-Moshe, L., Hubner, S., and Peleg, Z. (2015). Identification of conserved drought-adaptive genes using a cross-species meta-analysis approach. BMC Plant Biol. 15:111. doi: 10.1186/s12870-015-0493-6
Sievers, F., Wilm, A., Dineen, D., Gibson, T. J., Karplus, K., Li, W., et al. (2011). Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 7:539. doi: 10.1038/msb.2011.75
Sim, S.-C., Durstewitz, G., Plieske, J., Wieseke, R., Ganal, M. W., Van Deynze, A., et al. (2012a). Development of a large SNP genotyping array and generation of high-density genetic maps in tomato. PLoS One 7:e40563. doi: 10.1371/journal.pone.0040563
Sim, S.-C., Van Deynze, A., Stoffel, K., Douches, D. S., Zarka, D., Ganal, M. W., et al. (2012b). High-density SNP genotyping of tomato (Solanum lycopersicum L.) reveals patterns of genetic variation due to breeding. PLoS One 7:e45520. doi: 10.1371/journal.pone.0045520
Simão, F. A., Waterhouse, R. M., Ioannidis, P., Kriventseva, E. V., and Zdobnov, E. M. (2015). BUSCO: assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 31, 3210–3212. doi: 10.1093/bioinformatics/btv351
Simpson, J. T., Wong, K., Jackman, S. D., Schein, J. E., Jones, S. J., and Birol, I. (2009). ABySS: a parallel assembler for short read sequence data. Genome Res. 19, 1117–1123. doi: 10.1101/gr.089532.108
Sircar, D., Gaid, M., Chizzali, C., Reckwell, D., Kaufholdt, D., Beuerle, T., et al. (2015). Biphenyl 4-hydroxylases involved in aucuparin biosynthesis in rowan and apple are CYP736A proteins. Plant Physiol. 168, 428–442. doi: 10.1104/pp.15.00074
Smith-Unna, R., Boursnell, C., Patro, R., Hibberd, J. M., and Kelly, S. (2016). TransRate: reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 26, 1134–1144. doi: 10.1101/gr.196469.115
Song, X., Fang, J., Han, X., He, X., Liu, M., Hu, J., et al. (2016). Overexpression of Quinone reductase from Salix matsudana Koidz enhances salt tolerance in transgenic Arabidopsis thaliana. Gene 576, 520–527. doi: 10.1016/j.gene.2015.10.069
Spooner, D. M., Peralta, I. E., and Knapp, S. (2005). Comparison of AFLPs with other markers for phylogenetic inference in wild tomatoes [Solanum L. section Lycopersicon (Mill.) Wettst.]. Taxon 54, 43–61. doi: 10.2307/25065301
Stanke, M., Steinkamp, R., Waack, S., and Morgenstern, B. (2004). AUGUSTUS: a web server for gene finding in eukaryotes. Nucleic Acids Res. 32, W309–W312. doi: 10.1093/nar/gkh379
Stein, A. J., and Geiger, J. H. (2002). The crystal structure and mechanism of 1-L-myo-inositol- 1-phosphate synthase. J. Biol. Chem. 277, 9484–9491. doi: 10.1074/jbc.M109371200
Stieglitz, K. A., Roberts, M. F., Li, W., and Stec, B. (2007). Crystal structure of the tetrameric inositol 1-phosphate phosphatase (TM1415) from the hyperthermophile, Thermotoga maritima. FEBS J. 274, 2461–2469. doi: 10.1111/j.0014-2956.2007.05779.x
Stieglitz, K. A., Yang, H., Roberts, M. F., and Stec, B. (2005). Reaching for mechanistic consensus across life kingdoms: structure and insights into catalysis of the myo-inositol-1-phosphate synthase (mIPS) from Archaeoglobus fulgidus. Biochemistry 44, 213–224. doi: 10.1021/bi048267o
Suh, S. J., Wang, Y.-F., Frelet, A., Leonhardt, N., Klein, M., Forestier, C., et al. (2007). The ATP binding cassette transporter AtMRP5 modulates anion and calcium channel activities in Arabidopsis guard cells. J. Biol. Chem. 282, 1916–1924. doi: 10.1074/jbc.M607926200
Swanson-Wagner, R. A., Eichten, S. R., Kumari, S., Tiffin, P., Stein, J. C., Ware, D., et al. (2010). Pervasive gene content variation and copy number variation in maize and its undomesticated progenitor. Genome Res. 20, 1689–1699. doi: 10.1101/gr.109165.110
Taji, T., Seki, M., Satou, M., Sakurai, T., Kobayashi, M., Ishiyama, K., et al. (2004). Comparative genomics in salt tolerance between Arabidopsis and Arabidopsis-related halophyte salt cress using Arabidopsis microarray. Plant Physiol. 135, 1697–1709. doi: 10.1104/pp.104.039909
Tal, M. (1971). Salt tolerance in the wild relatives of the cultivated tomato: responses of Lycopersicon esculentum, L. peruvianum, and L. esculentum minor to sodium chloride solution. Aust. J. Agric. Res. 22, 631–638. doi: 10.1071/AR9710631
Tanksley, S. D. (2004). The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. Plant Cell 16, S181–S189. doi: 10.1105/tpc.018119
Tanksley, S. D., Grandillo, S., Fulton, T. M., Zamir, D., Eshed, Y., Petiard, V., et al. (1996). Advanced backcross QTL analysis in a cross between an elite processing line of tomato and its wild relative L. pimpinellifolium. Theor. Appl. Genet. 92, 213–224. doi: 10.1007/BF00223378
Tanksley, S. D., and McCouch, S. R. (1997). Seed banks and molecular maps: unlocking genetic potential from the wild. Science 277, 1063–1066. doi: 10.1126/science.277.5329.1063
Tarailo-Graovac, M., and Chen, N. (2009). Using repeatmasker to identify repetitive elements in genomic sequences. Curr. Protoc. Bioinformatics 4, 4.10.1–4.10.14. doi: 10.1002/0471250953.bi0410s25
Tester, M., and Davenport, R. (2003). Na + tolerance and Na + transport in higher plants. Ann. Bot. 91, 503–527. doi: 10.1093/aob/mcg058
Thapa, S. P., Miyao, E. M., Davis, R. M., and Coaker, G. (2015). Identification of QTLs controlling resistance to Pseudomonas syringae pv. tomato race 1 strains from the wild tomato, Solanum habrochaites LA1777. Theor. Appl. Genet. 128, 681–692. doi: 10.1007/s00122-015-2463-7
The Tomato Genome Consortium (2012). The tomato genome sequence provides insights into fleshy fruit evolution. Nature 485, 635–641. doi: 10.1038/nature11119
The 100 Tomato Genome Sequencing Consortium, Aflitos, S., Schijlen, E., De Jong, H., De Ridder, D., Smit, S., et al. (2014). Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 80, 136–148. doi: 10.1111/tpj.12616
Tieman, D., Zhu, G., Resende, M. F., Lin, T., Nguyen, C., Bies, D., et al. (2017). A chemical genetic roadmap to improved tomato flavor. Science 355, 391–394. doi: 10.1126/science.aal1556
Tran, L.-S. P., Urao, T., Qin, F., Maruyama, K., Kakimoto, T., Shinozaki, K., et al. (2007). Functional analysis of AHK1/ATHK1 and cytokinin receptor histidine kinases in response to abscisic acid, drought, and salt stress in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 104, 20623–20628. doi: 10.1073/pnas.0706547105
Verrier, P. J., Bird, D., Burla, B., Dassa, E., Forestier, C., Geisler, M., et al. (2008). Plant ABC proteins – a unified nomenclature and updated inventory. Trends Plant Sci. 13, 151–159. doi: 10.1016/j.tplants.2008.02.001
Villalta, I., Reina-Sanchez, A., Bolarin, M. C., Cuartero, J., Belver, A., Venema, K., et al. (2008). Genetic analysis of Na + and K + concentrations in leaf and stem as physiological components of salt tolerance in tomato. Theor. Appl. Genet. 116, 869–880. doi: 10.1007/s00122-008-0720-8
Víquez-Zamora, M., Caro, M., Finkers, R., Tikunov, Y., Bovy, A., Visser, R. G., et al. (2014). Mapping in the era of sequencing: high density genotyping and its application for mapping TYLCV resistance in Solanum pimpinellifolium. BMC Genomics 15:e1152. doi: 10.1186/1471-2164-15-1152
Virdi, A. S., Singh, S., and Singh, P. (2015). Abiotic stress responses in plants: roles of calmodulin-regulated proteins. Front. Plant Sci. 6:809. doi: 10.3389/fpls.2015.00809
Xiao, H., Jiang, N., Schaffner, E., Stockinger, E. J., and Van Der Knaap, E. (2008). A retrotransposon-mediated gene duplication underlies morphological variation of tomato fruit. Science 319, 1527–1530. doi: 10.1126/science.1153040
Xie, C., and Tammi, M. T. (2009). CNV-seq, a new method to detect copy number variation using high-throughput sequencing. BMC Bioinformatics 10:80. doi: 10.1186/1471-2105-10-80
Yu, P., Wang, C., Xu, Q., Feng, Y., Yuan, X., Yu, H., et al. (2011). Detection of copy number variations in rice using array-based comparative genomic hybridization. BMC Genomics 12:372. doi: 10.1186/1471-2164-12-372
Zhang, L. P., Lin, G. Y., and Foolad, M. R., (2003). “QTL comparison of salt tolerance during seed germination and vegetative stage in a Lycopersicum esculentum X L. pimpinellifolium RIL population,” in Proceedings of the XXVI International Horticultural Congress: Environmental Stress and Horticulture Crops (Toronto: ISHS Acta Horticulturae), 59–67. doi: 10.17660/ActaHortic.2003.618.5
Zhang, Y. (2008). I-TASSER server for protein 3D structure prediction. BMC Bioinformatics 9:40. doi: 10.1186/1471-2105-9-40
Zheng, L. Y., Guo, X. S., He, B., Sun, L. J., Peng, Y., Dong, S. S., et al. (2011). Genome-wide patterns of genetic variation in sweet and grain sorghum (Sorghum bicolor). Genome Biol. 12:R114. doi: 10.1186/gb-2011-12-11-r114
Zhong, S., Fei, Z., Chen, Y.-R., Zheng, Y., Huang, M., Vrebalov, J., et al. (2013). Single-base resolution methylomes of tomato fruit development reveal epigenome modifications associated with ripening. Nat. Biotechnol. 31, 154–159. doi: 10.1038/nbt.2462
Žižková, E., Dobrev, P. I., Muhovski, Y., Hošek, P., Hoyerová, K., Haisel, D., et al. (2015). Tomato (Solanum lycopersicum L.) SlIPT3 and SlIPT4 isopentenyltransferases mediate salt stress response in tomato. BMC Plant Biol. 15:85. doi: 10.1186/s12870-015-0415-7
Keywords: wild tomato, Solanum pimpinellifolium, genome analysis, salinity tolerance, inositol 3-phosphate synthase
Citation: Razali R, Bougouffa S, Morton MJL, Lightfoot DJ, Alam I, Essack M, Arold ST, Kamau AA, Schmöckel SM, Pailles Y, Shahid M, Michell CT, Al-Babili S, Ho YS, Tester M, Bajic VB and Negrão S (2018) The Genome Sequence of the Wild Tomato Solanum pimpinellifolium Provides Insights Into Salinity Tolerance. Front. Plant Sci. 9:1402. doi: 10.3389/fpls.2018.01402
Received: 29 April 2018; Accepted: 04 September 2018;
Published: 04 October 2018.
Edited by:
Henry T. Nguyen, University of Missouri, United StatesReviewed by:
Marina Tucci, Consiglio Nazionale delle Ricerche (CNR), ItalyAureliano Bombarely, Virginia Tech, United States
Copyright © 2018 Razali, Bougouffa, Morton, Lightfoot, Alam, Essack, Arold, Kamau, Schmöckel, Pailles, Shahid, Michell, Al-Babili, Ho, Tester, Bajic and Negrão. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vladimir B. Bajic, dmxhZGltaXIuYmFqaWNAa2F1c3QuZWR1LnNh Sónia Negrão, c29uaWEubmVncmFvQHVjZC5pZQ==
†Shared first authorship
‡Present address: Salim Bougouffa, King Abdullah University of Science and Technology (KAUST), Core Labs, Thuwal, Saudi Arabia; Damien J. Lightfoot, KAUST Environmental Epigenetic Program, Division of Biological and Environmental Science and Engineering, King Abdullah University of Science and Technology, Thuwal, Saudi Arabia; Craig T. Michell, Department of Environmental and Biological Sciences, University of Eastern Finland, Joensuu, Finland; Sónia Negrão, UCD School of Biology and Environmental Science, Science Centre West, University College Dublin (UCD), Dublin, Ireland