 Isabel Vicente Muñoz
Isabel Vicente Muñoz Sabrina Sarrocco
Sabrina Sarrocco Luca Malfatti
Luca Malfatti Riccardo Baroncelli
Riccardo Baroncelli Giovanni Vannacci
Giovanni Vannacci- 1Department of Agriculture, Food and Environment, University of Pisa, Pisa, Italy
- 2Spanish-Portuguese Center for Agricultural Research (CIALE), Department of Microbiology and Genetics, University of Salamanca, Villamayor, Spain
Background
Fungal pathogens are the main factors responsible for the most severe diseases affecting plants, leading to significant reduction in yield and crop quality and causing enormous economic losses worldwide. It is estimated that around 30% of the emerging diseases are caused by fungi (Giraud et al., 2010) thus requiring new strategies to improve their management. Biological control approach, frequently referred to the use of non-pathogenic microbial antagonists or products derived from their metabolism, represents a valid and promising alternative under a more ecological perspective to reduce the activities and to control populations of target pathogens (Singh, 2016). However, although the use of antagonists belonging to species different from that of the pathogen has been successfully reported, the use of competitors belonging to the same species of the pathogen is not widespread. A biocontrol strategy based on competition for space and nutrients and/or the induction of plant defenses against virulent pathogens performed by attenuated or avirulent pathogens (Ghorbanpour et al., 2018) could, therefore, be considered a valid alternative.
The Results So Far
Veloso et al. (2015) reported the use of an avirulent isolate of Fusarium oxysporum to reduce Verticillium wilt severity in pepper, through competition and induction of the plant defense responses. A similar approach was described by Salazar et al. (2012) for the management of anthracnose in strawberries. The avirulent isolate F7 of Colletotrichum fragariae conferred full protection from the infection caused by C. acutatum and also enhanced plant resistance against Botrytis cinerea through the induction of plant defense responses. Similarly, the use of an attenuated Verticillium nigrescens isolate reduced cotton wilt caused by a virulent isolate of V. dahliae (Vagelas and Leontopoulos, 2015).
(Aimé et al., 2013) used an avirulent isolate of F. oxysporum to combat F. oxysporum f. sp. lycopersici to reduce Fusarium wilt by priming a Salicylic-dependant signaling defense on tomato plants. The use of an avirulent strain of Valsa mali var. mali reduced the infection rate of apple tree canker caused by the virulent strain LXS080601 from 97 to 41% (Zhang et al., 2014) on apple callus. In 1993 as regards the mycotoxigenic fungi, Cotty and Bayman (1993) suggested the use of a non-aflatoxigenic isolate of Aspergillus flavus to control the development of aflatoxigenic strains in maize kernels by competitive exclusion and this strategy today is commercially applied in several countries (Ojiambo et al., 2018).
However, the selection of suitable isolates to be used as potential antagonists from the local fungal community often takes (long) time for identification and screening. Selection within a great number of isolates based on morphological, physiological and genetic features is usually required, followed by an in vivo screening against the pathogen on a real disease scenario. An interesting alternative to easily and quickly obtain new genotypes able to act as biocontrol agents, could be the induction of genetic mutations in the virulent genotypes, providing new avirulent strains that can compete directly with the virulent ones or induce plant defense responses (Ghorbanpour et al., 2018). The application of genetic transformation techniques to silencing genes putatively involved in pathogenicity has been widely used to uncoil the role of these genes in the establishment and development of the infection processes (Johnson et al., 2018). However, the disruption of a gene function usually involves the integration in the genome of foreign DNA sequences used as reporter genes in order to select transformants, leading to the generation of antibiotic-resistant or fluorescent strains. These genetic modifications represent a major constraint for their use in field.
The Genome-Editing Era: State of the Art and Perspectives for the Management of Plant Diseases
The arrival of the CRISPR-Cas9 (Clustered Regularly Interspaced Short Palindromic Repeats – CRISPR Associated protein 9) genome-editing technique enabled researchers to modify genomic sequences in a more precise way (Knott and Doudna, 2018). CRISPR-Cas9 Type II system uses two principal components for gene targeting and cleavage: the RNA guide (sgRNA) and the Cas9 endonuclease. The sgRNA consists of a simple chimeric strand of RNA, which leads Cas9 up to the localization in the genome of the target gene, whose expression has to be blocked. Cas9 is able to bind the DNA and to produce a double strand break (DSB) in the target gene. The DSB then induces the activation of one of the DNA cellular reparation systems, the Non-Homologous End-Joining (NHEJ) system, that can re-join the ends of the DSB without introducing errors or, unlikely, giving rise to insertions or deletions of nucleotides during the repair. These InDels led to changes in the gene reading frame producing non-sense sequences or causing the appearance of premature stop codons, thus blocking the transcription of the target gene (Bono et al., 2015). In most cases the application of the technique in filamentous fungi consisted of a proof of concept of its feasibility (see Table 1). The system has the advantage that once implemented in the organism, it is possible to change the target gene by changing the sgRNA spacer sequence, as well as silencing several genes simultaneously by transforming the cell with different sgRNAs along with Cas9 (Hsu et al., 2015). Nevertheless, the most interesting advantage is that it allows to perform marker-free deletions by using transient expression plasmids that can self-replicate only under antibiotic pressure (Katayama et al., 2015, Nødvig et al., 2015; Schuster et al., 2016; Zhang et al., 2016; Liu et al., 2017; Wenderoth et al., 2017; Weyda et al., 2017; Wang et al., 2018). The use of CRISPR-Cas not only provides a time-saving path to perform genomic functional analyses, but also could provide new fungal genotypes, that can be used as potential competitors of plant pathogens and/or in the priming of plant defense responses.
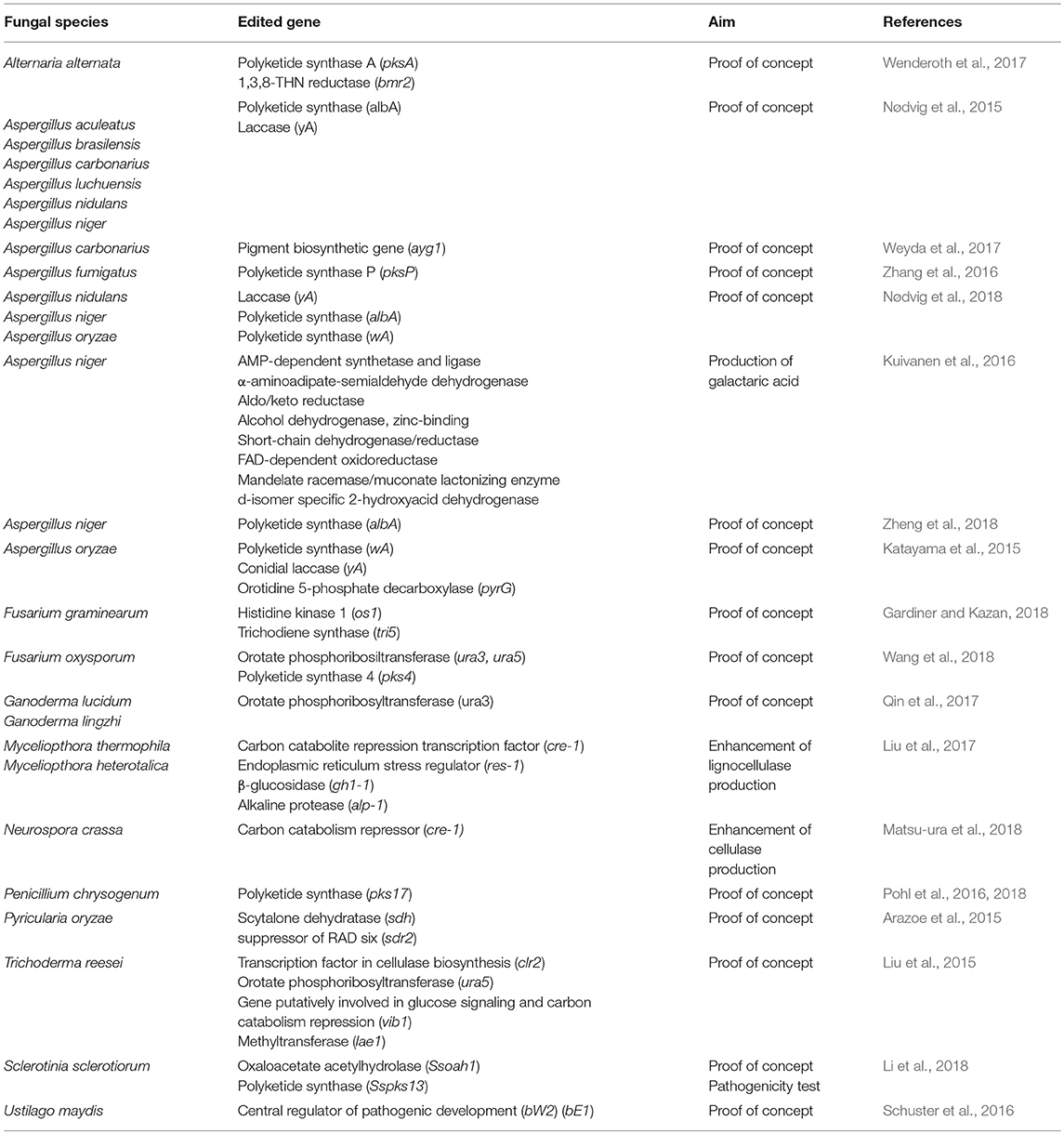
Table 1. Application of CRISPR-Cas9 for gene-silencing in filamentous fungi.
One possible scenario for the application of CRISPR-Cas9 silenced mutants could be Fusarium Head Blight (FHB), one of the most destructive diseases of grain cereal crops worldwide caused by different Fusarium spp., with F. graminearum and F. culmorum as the most common and aggressive agents. In FHB, while yield loss derives from sterility of infected florets, grain quality reduction is mainly due to the accumulation of trichothecenes—coded by the fungal tri genes cluster—highly toxic for humans and animals. Previous studies reported that iRNA (interference RNA) Δtri6 mutants of F. culmorum showed reduced disease indices ranging from 40 to 80% on durum wheat (Scherm et al., 2011). In addition, classic knocked-out Δtri5 and Δtri6 mutants of F. graminearum were unable to spread the disease to the adjacent spikelets and grains on wheat and corn, respectively, and also induced plant defense responses (Ravensdale et al., 2014). Likewise, Δmap1 mutants of F. graminearum showed two-fold reduction of mycotoxin production and were unable to produce perithecia as well as to penetrate in wheat tissues, while the ability to colonize the straw was not affected (Urban et al., 2003). Considering that competition for space and nutrients between virulent and non-virulent strains could reduce the disease, the field release of non-virulent CRISPR-mutant strains of F. graminearum and F. culmorum might help to control the incidence of FHB
Another contribution of CRISPR-Cas9 is the production of well-known antagonists with enhanced biocontrol aptitudes achieved through genome-editing (Vicente Muñoz et al., 2017). For example, species belonging to genus Trichoderma have been considered outstanding biocontrol agents able to reduce the disease severity (Sarrocco et al., 2013), not only by constraining the growth of the phytopathogens (Sarrocco et al., 2009), even killing them, but also by eliciting the plant defense responses (Fiorini et al., 2016; Sarrocco et al., 2017). One of the mechanisms through which these fungi can antagonize phytopathogenic fungi is the release of a wide arsenal of cell-wall degrading enzymes and secondary metabolites such as antibiotics, among others (Khalid, 2017).
Genetic engineering of the metabolic pathways that trigger the biosynthesis of secreted proteins and secondary compounds could provide new fungal strains with enhanced biocontrol activity. Previous studies reported that it is possible to achieve the same effect through the silencing of negative regulatory elements, signal-transduction components or genes belonging to contiguous metabolic networks, thus, redirecting metabolite flow and biosynthesis or supressing the feedback inhibition by which its production could be regulated (Bailey, 1991). For example, Δtvk1 mutants of T. virens displayed enhanced biocontrol activity against R. solani, in addition to an increased expression of mycoparasitism-related genes and overproduction of lytic enzymes (Mendoza-Mendoza et al., 2003). Likewise, four knockout mutants in SSCPs (small secreted cysteine rich proteins)-encoding genes of T. virens showed greater ability to induce ISR (Induced Systemic Resistance) on corn against Cochliobolus heterostrophus than the wild type (Lamdan et al., 2015). Another example of biocontrol enhanced ability was described by Reithner et al. (2005) in T. atroviride, in which the Δtga1 mutants exhibited an overproduction of antifungal secondary metabolites. Similarly, the Δtmk1 mutants of T. atroviride showed overproduction of 6-pentyl-pyrone and peptaibols, resulting in an enhanced antifungal activity and increased protection of bean plants against Rhizoctonia solani (Reithner et al., 2007). On the other hand, biosynthesis of secondary metabolites is often carried out by clustered genes whose expression could be induced by environmental conditions. However, in many cases these clusters are silent and their activation cannot be achieved (Osbourn, 2010). Bok et al. (2009) demonstrated that the silencing of a transcription factor involved in the methylation of lysine 4 of the histone H3 in Aspergillus nidulans activated the expression of cryptic clusters and yielded novel secondary metabolites. The silencing of ace1 gene induces the up-regulation of four polyketide biosynthetic gene clusters in T. atroviride, leading to an increase in the production of antibiotics and other secondary metabolites that clearly enhanced its potential as biocontrol agent against F. oxysporum and R. solani (Fang and Chen, 2018). Following this approach, it is possible to induce the activation of unknown clusters in beneficial fungi by using CRISPR-Cas9, allowing the discovery of new secondary metabolites that could interact with plants or phytopathogens. This could result in new interesting biocontrol strains to be released in field avoiding the introduction of transgenes in the environment.
Conclusions
The availability of novel or the improvement of known techniques that are safer for people and the environment is of outmost importance to guarantee food safety and security especially in those countries where famine is still an important issue (Vurro et al., 2010). A novel technique that allows the production of precise knock-out mutants without the insertion of foreign DNA in a saprotrophic/pathogenic fungus opens new possibilities of controlling plant pathogens. The use of such edited fungal strains needs a correct strategy to minimize possible risks. The major risk related to the release of a mutant strain is the rise, in the field, of novel combinations of pathogenesis/fitness related genes following the sexual or parasexual cycle. The genetic background of an edited isolate and its wild type is exactly the same, except for the edited gene, thus novel combinations of genes are not conceivable. Anyway, to further reduce such possibility, we can imagine a strategy of deployment that includes 1) the gene edit of the most prevalent genotype of the pathogen/saprotroph in the release area; 2) the editing of more than one gene in the same metabolic pathway and 3) the editing also of the idiomorphs and /or the HET genes, to make sexual or parasexual recombination (including the re-gain of virulence) even less likely.
Anyway, the application of novel techniques and the release of new products need, as usual, to be evaluated for their safety and to be accepted by populations. A recent sentence of the Court of Justice of the EU stated that edited organisms, even if they do not contain alien DNA, have to be subjected to the rules set up for Genetically Modified Organisms. This is not the place to discuss this issue, but it is high time, in EU at least, to reconsider the whole GMO legislation.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Aimé, S., Alabouvette, C., Steinberg, C., and Olivain, C. (2013). The endophytic strain Fusarium oxysporum Fo47: a good candidate for priming the defense responses in tomato roots. Mol. Plant Microbe Interact. 26, 918–926. doi: 10.1094/MPMI-12-12-0290-R
Arazoe, T., Miyoshi, K., Yamato, T., Ogawa, T., Ohsato, S., Arie, T., et al. (2015). Tailor-made CRISPR/Cas system for highly efficient targeted gene replacement in the rice blast fungus. Biotech. Bioeng. 112, 2543–2549. doi: 10.1002/bit.25662
Bailey, J. (1991). Toward a science of metabolic engineering. Science 252, 1668–1675. doi: 10.1126/science.2047876
Bok, J. W., Chiang, Y. M., Szewczyk, E., Reyes-Dominguez, Y., Davidson, A. D., Sanchez, J. F., et al. (2009). Chromatin-level regulation of biosynthetic gene clusters. Nat. Chem. Biol. 5, 462–464. doi: 10.1038/nchembio.177
Bono, J. M., Olesniky, E. C., and Matzkin, L. M. (2015). Connecting genotypes, phenotypes and fitness: harnessing the power of CRISPR/Cas9 genome editing. Mol. Ecol. 24, 3810–3822. doi: 10.1111/mec.13252
Cotty, P. J., and Bayman, P. (1993). Competitive exclusion of a toxigenic strain of Aspergillus flavus by an atoxigenic strain. Phytopathology 83, 1283–1287. doi: 10.1094/Phyto-83-1283
Fang, C., and Chen, X. (2018). Potential biocontrol efficacy of Trichoderma atroviride with cellulase expression regulator ace1 gene knock-out. Biotechnology 8:302. doi: 10.1007/s13205-018-1314-z
Fiorini, L., Guglielminetti, L., Mariotti, L., Curadi, M., Picciarelli, P., Scartazza, A., et al. (2016). Trichoderma harzianum T6776 modulates a complex metabolic network to stimulate tomato cv Micro-Tom growth. Plant Soil 400, 351–366. doi: 10.1007/s11104-015-2736-6
Gardiner, D. M., and Kazan, K. (2018). Selection is required for efficient Cas9-mediated genome editing in Fusarium graminearum. Fungal Biol. 122, 131–137. doi: 10.1016/j.funbio.2017.11.006
Ghorbanpour, M., Omidvari, M., Abbaszadeh-Dahaji, P., Omidvar, R., and Kariman, K. (2018). Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biol. Cont. 117, 147–157. doi: 10.1016/j.biocontrol.2017.11.006
Giraud, T., Gladieux, P., and Gavrilets, S. (2010). Linking the emergence of fungal plant diseases with ecological speciation. Trends Ecol. Evol. 25, 387–395. doi: 10.1016/j.tree.2010.03.006
Hsu, P. D., Lander, E. S., and Zhang, F. (2015). Development and applications of CRISPR-Cas9 for genome engineering. Cell 157, 1262–1278. doi: 10.1016/j.cell.2014.05.010
Johnson, E. T., Proctor, R. H., Dunlap, C. A., and Busman, M. (2018). Reducing production of fumonisin mycotoxins in Fusarium verticillioides by RNA interference. Myc. Res. 34, 29–37. doi: 10.1007/s12550-017-0296-8
Katayama, T., Tanaka, Y., Okabe, T., Nakamura, H., Fuji, W., Kitamoto, K., et al. (2015). Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotech. Lett. 38, 637–642. doi: 10.1007/s10529-015-2015-x
Khalid, S. A. (2017). Trichoderma as biological control weapon against soil borne plant pathogens. Afr. J. Biotech. 16, 2299–2306. doi: 10.5897/AJB2017.16270
Knott, G. J., and Doudna, J. A. (2018). CRISPR-Cas guides the future of genetic engineering. Science. 361, 866–869. doi: 10.1126/science.aat5011
Kuivanen, J., Wang, Y. M. J., and Richard, P. (2016). Engineering Aspergillus niger for galactaric acid production: elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Micr. Cell Fact. 15:210. doi: 10.1186/s12934-016-0613-5
Lamdan, N. L., Shalaby, S., Ziv, T., Kenerley, C. M., and Horwitz, B. A. (2015). Secretome of Trichoderma interacting with maize roots: role in induced systemic resistance. Mol. Cell. Prot. 14, 1054–1063. doi: 10.1074/mcp.M114.046607
Li, J., Zhang, Y., Zhang, Y., Yu, P. L., Pan, H., and Rollins, J. A. (2018). Introduction of large sequence inserts by CRISPR-Cas9 to create pathogenicity mutants in the multinucleate filamentous pathogen Sclerotinia sclerotiorum. mBio 9:e00567–18. doi: 10.1128/mBio.00567-18
Liu, Q., Gao, R., Li, J., Lin, L., Zhao, J., Sun, W., et al. (2017). Development of a genome-editing CRISPR/Cas9 system in thermophilic fungal Myceliophthora species and its application to hyper-cellulase production strain engineering. Biotech. Biofuels 10:1. doi: 10.1186/s13068-016-0693-9
Liu, R., Chen, L., Jiang, Y., Zhou, Z., and Zou, G. (2015). Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Disc. 1:15007. doi: 10.1038/celldisc.2015.7
Matsu-ura, T., Dovzhenok, A. A., Coradetti, S. T., Subramanian, K. R., Meyer, D. R., Kwon, J. J., et al. (2018). Synthetic gene network with positive feedback loop amplifies cellulase gene expression in Neurospora crassa. ACS Synth. Biol. 7, 1395–1405. doi: 10.1021/acssynbio.8b00011
Mendoza-Mendoza, A., Pozo, M. J., Grzegorski, D., Martinez, P., Garcia, J. M., Olmedo-Monfil, V., et al. (2003). Enhanced biocontrol activity of Trichoderma through inactivation of a mitogen-activated protein kinase. Proc Natl Acad Sci U.S.A. 100, 15965–15970. doi: 10.1073/pnas.2136716100
Nødvig, C. S., Hoof, J. B., Kogle, M. E., Jarczynska, Z. D., Lehmbeck, J., Klitgaard, D. K., et al. (2018). Efficient oligo nucleotide mediated CRISPR-Cas9 gene editing in Aspergilli. Fung. Gen. Biol. 115, 78–89. doi: 10.1016/j.fgb.2018.01.004
Nødvig, C. S., Nielsen, J. B., Kogle, M. E., and Mortensen, U. H. (2015). A CRISPR-Cas9 system for genetic engineering of filamentous fungi. PLoS ONE 10:e0133085. doi: 10.1371/journal.pone.0133085
Ojiambo, P. S., Battilani, P., Cary, J. W., Blum, B. H., and Carbone, I. (2018). Cultural and genetic approaches to manage aflatoxin contamination: recent insights provide opportunities for improved control. Phytopath 108, 1024–1037. doi: 10.1094/PHYTO-04-18-0134-RVW
Osbourn, A. (2010). Secondary metabolic gene clusters: evolutionary toolkits for chemical innovation. Trends Gen. 26, 449–457. doi: 10.1016/j.tig.2010.07.001
Pohl, C., Kiel, J. A. K. W., Driessen, A. J. M., Bovenberg, R. A. L., and Nygård, Y. (2016). CRISPR/Cas9 based genome editing of Penicillium chrysogenum. Synth. Biol. 5, 754–764. doi: 10.1021/acssynbio.6b00082
Pohl, C., Mózsik, L., Driessen, A. J. M., Bovenberg, R. A. L., and Nygård, Y. I. (2018). Genome editing in Penicillium chrysogenum using Cas9 ribonucleoprotein particles. Synth. Biol. 1772, 213–232. doi: 10.1007/978-1-4939-7795-6_12
Qin, H., Xiao, H., Zou, G., Zhou, Z., and Zhong, J. J. (2017). CRISPR-Cas9 assisted gene disruption in the higher fungus Ganoderma species. Proc. Biochem. 56, 57–61. doi: 10.1016/j.procbio.2017.02.012
Ravensdale, M., Rocheleau, H., Wang, L., Nasmith, C., Ouellet, T., and Subramaniam, R. (2014). Components of priming-induced resistance to fusarium head blight in wheat revealed by two distinct mutants of Fusarium graminearum. Mol. Plant Path. 5, 948–956. doi: 10.1111/mpp.12145
Reithner, B., Brunner, K., Schuhmacher, R., Peissl, I., Seidl, V., Krska, R., et al. (2005). The G protein α subunit Tga1 of Trichoderma atroviride is involved in chitinase formation and differential production of antifungal metabolites. Fung. Gen. Biol. 42, 749–760. doi: 10.1016/j.fgb.2005.04.009
Reithner, B., Schuhmacher, R., Stoppacher, N., Pucher, M., Brunner, K., and Zeilinger, S. (2007). Signaling via the Trichoderma atroviride mitogen-activated protein kinase Tmk1 differentially affects mycoparasitism and plant protection. Fung. Gen. Biol. 44, 1123–1133. doi: 10.1016/j.fgb.2007.04.001
Salazar, S. M., Grellet, C. F., Chalfoun, N. R., Castagnaro, A. P., and Díaz Ricci, J. C. (2012). Avirulent strain of Colletotrichum induces a systemic resistance in strawberry. Eur. J. Plant Path. 135, 877–888. doi: 10.1007/s10658-012-0134-y
Sarrocco, S., Guidi, L., Fambrini, S., Degl'Innocenti, E., and Vannacci, G. (2009). Competition for cellulose exploitation between Rhizoctonia solani and two Trichoderma isolates in the decomposition of wheat straw. J. Plant Path. 91, 331–338. doi: 10.4454/jpp.v91i2.962
Sarrocco, S., Matarese, F., Baroncelli, R., Seidl-Seiboth, V., Kubicek, C. P., Vannacci, G., et al. (2017). The constitutive endopolygalacturonase TvPG2 regulates the induction of plant systemic resistance by Trichoderma virens. Phytopath ology 107, 537–544. doi: 10.1094/PHYTO-03-16-0139-R
Sarrocco, S., Matarese, F., Moncini, L., Pachetti, G., Ritieni, A., Moretti, et al. (2013). Biocontrol of fusarium head blight by spike application of Trichoderma gamsii. J. Plant Pathol. S1, 19–27.
Scherm, B., Orr,ú, M., Balmas, V., Spanu, F., Azara, E., Delogu, G., et al. (2011). Altered trichothecene biosynthesis in TRI6-silenced transformants of Fusarium culmorum influences the severity of crown and foot rot on durum wheat seedlings. Mol. Plant Pathol. 12, 759–771. doi: 10.1111/j.1364-3703.2011.00709.x
Schuster, M., Schweizer, G., Reissmann, S., and Kahmann, R. (2016). Genome editing in Ustilago maydis using the CRISPR–Cas system. Fung. Gen. Biol. 89, 3–9. doi: 10.1016/j.fgb.2015.09.001
Singh, S. (2016). Role of nonpathogenic fungi in inducing systemic resistance in crop plants against phytopathogens. Micr. Inoc. Sust. Agric. Prod. 69–83. doi: 10.1007/978-81-322-2644-4_5
Urban, M., Mott, E., Farley, T., and Hammond-Kosack, K. (2003). The Fusarium graminearum MAP1 gene is essential for pathogenicity and development of perithecia. Mol. Plant Pathol. 4, 347–359. doi: 10.1046/j.1364-3703.2003.00183.x
Vagelas, J., and Leontopoulos, S. (2015). Cross-protection of cotton against Verticillium wilt by Verticillium nigrescens. Emir. J. Food Agric. 27, 687–691. doi: 10.9755/ejfa.2015-04-047
Veloso, J., Alabouvette, C., Olivain, C., Flors, V., Pastor, V., García, et al. (2015). Modes of action of the protective strain Fo47 in controlling Verticillium wilt of pepper. Plant Pathol. 65, 997–1007. doi: 10.1111/ppa.12477
Vicente Muñoz, I., Sarrocco, S., and Vannacci, G. (2017). CRISPR-CAS for the genome editing of two Trichoderma spp. beneficial isolates. J. Plant Pathol. 99:S63. doi: 10.1038/srep45763
Vurro, M., Bonciani, B., and Vannacci, G. (2010). Emerging infectious diseases of crop plants in developing countries: impact on agriculture and socio-economic consequences. Food Sec. 2, 113–132. doi: 10.1007/s12571-010-0062-7
Wang, Q., Cobine, P. A., and Coleman, J. J. (2018). Efficient genome editing in Fusarium oxysporum based on CRISPR/Cas9 ribonucleoprotein complexes. Fung. Gen. Biol. 117, 21–29. doi: 10.1016/j.fgb.2018.05.003
Wenderoth, M., Pinecker, C., Vo,ß, B., and Fischer, R. (2017). Establishment of CRISPR/Cas9 in Alternaria alternata. Fung. Gen. Biol. 101, 55–60. doi: 10.1016/j.fgb.2017.03.001
Weyda, I., Yang, L., Vang, J., Ahring, B. K., Lübeck, M., and Lübeck, P. S. (2017). A comparison of Agrobacterium-mediated transformation and protoplast-mediated transformation with CRISPR-Cas9 and bipartite gene targeting substrates, as effective gene targeting tools for Aspergillus carbonarius. J. Micrbiol. Methods 135, 26–34. doi: 10.1016/j.mimet.2017.01.015
Zhang, C., Meng, X., Wei, X., and Lu, L. (2016). Highly efficient CRISPR mutagenesis by microhomology-mediated end joining in Aspergillus fumigatus. Fungal Gen. Biol. 86, 47–57. doi: 10.1016/j.fgb.2015.12.007
Zhang, Q., Wang, C., Yong, D., Li, G., Dong, X., and Li, B. (2014). Induction of resistance mediated by an attenuated strain of Valsa mali var.mali using pathogen-apple callus interaction system. Sci. World J. 2014:201382. doi: 10.1155/2014/201382
Keywords: genome editing, CRISPR-Cas9, filamentous fungi, biocontrol, plant diseases management, fungal pathogens, beneficial fungi
Citation: Muñoz IV, Sarrocco S, Malfatti L, Baroncelli R and Vannacci G (2019) CRISPR-Cas for Fungal Genome Editing: A New Tool for the Management of Plant Diseases. Front. Plant Sci. 10:135. doi: 10.3389/fpls.2019.00135
Received: 29 November 2018; Accepted: 28 January 2019;
Published: 15 February 2019.
Edited by:
Ivan Baccelli, Istituto per la Protezione sostenibile delle Piante, ItalyReviewed by:
Kemal Kazan, Commonwealth Scientific and Industrial Research Organization (CSIRO), AustraliaCopyright © 2019 Muñoz, Sarrocco, Malfatti, Baroncelli and Vannacci. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sabrina Sarrocco, c2FicmluYS5zYXJyb2Njb0B1bmlwaS5pdA==