 Vanesa Eleonora Tossi1,2†Jose Javier Regalado1,2†Jesica Iannicelli3,4Leandro Ezequiel Laino1
Vanesa Eleonora Tossi1,2†Jose Javier Regalado1,2†Jesica Iannicelli3,4Leandro Ezequiel Laino1 Hernan Pablo Burrieza5,6
Hernan Pablo Burrieza5,6 Alejandro Salvio Escandón3
Alejandro Salvio Escandón3 Sandra Irene Pitta-Álvarez1,2*
Sandra Irene Pitta-Álvarez1,2*- 1Laboratorio de Cultivo Experimental de Plantas y Microalgas, Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
- 2Instituto de Micología y Botánica, CONICET-Universidad de Buenos Aires, Buenos Aires, Argentina
- 3Instituto de Genética “Ewald A. Favret,” Instituto Nacional de Tecnología Agropecuaria, Buenos Aires, Argentina
- 4CONICET-Consejo Nacional de Investigaciones Científicas y Tecnológicas, Buenos Aires, Argentina
- 5Laboratorio de biología del desarrollo de las plantas, Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
- 6Instituto de Biodiversidad y Biología Experimental, CONICET-Universidad de Buenos Aires, Buenos Aires, Argentina
Ultraviolet-B radiation (UV-B, 280–315 nm) is an important environmental signal that regulates growth and development in plants. Two dose-dependent UV-B response pathways were described in plants: a specific one, mediated by UVR8 (the specific UV-B receptor) and an unspecific one, activated by the oxidative damage produced by radiation. The constitutively expressed receptor appears inactive as a dimer, with the two monomers dissociating upon UV-B irradiation. The monomer then interacts with COP1, an ubiquitin ligase, hindering its ability to poly-ubiquitinate transcriptional factor HY5, thus averting its degradation and activating the photomorphogenic response. HY5 induces the synthesis of proteins RUP1 and RUP2, which interact with UVR8, releasing COP1, and inducing the re-dimerization of UVR8. This mechanism has been thoroughly characterized in Arabidopsis, where studies have demonstrated that the UVR8 receptor is key in UV-B response. Although Arabidopsis importance as a model plant many mechanisms described in this specie differ in other plants. In this paper, we review the latest information regarding UV-B response mediated by UVR8 in different species, focusing on the differences reported compared to Arabidopsis. For instance, UVR8 is not only induced by UV-B but also by other agents that are expressed differentially in diverse tissues. Also, in some of the species analyzed, proteins with low homology to RUP1 and RUP2 were detected. We also discuss how UVR8 is involved in other developmental and stress processes unrelated to UV-B. We conclude that the receptor is highly versatile, showing differences among species.
Introduction
Approximately 7% of all solar radiation that reaches the Earth's surface is ultraviolet light (UV) (Frohnmeyer and Staiger, 2003). UV radiation is divided into three wavelength ranges: UV-A (315–400 nm), UV-B (280–315 nm) and UV-C (100–280 nm) (Björn, 2015). UV-C, the most energetic of the three, is completely absorbed by the ozone (O3) layer. UV-A is not attenuated by ozone, but it is the least damaging and acts as a photomorphogenic signal (Björn, 1996). Approximately 95% of UV-B radiation is absorbed by the ozone layer and reaches the Earth's surface with an average intensity of 1 W.m−2 (Cejka et al., 2011). Depending on the dosage, UV-B radiation can either act as a signal or cause damage. Due to its high energy, UV-B can break chemical bonds and produce highly reactive molecules.
The natural dynamic balance of ozone levels was abruptly interrupted by anthropogenic emission of ozone-depleting substances (ODS) (from the 1960's to the end of the 1990's), with a decrease in ozone levels and an increase in UV-B radiation reaching the Earth's surface (Steinbrecht et al., 2018). Thirty years after the Montreal Treaty, which banned ODS emission worldwide, the ozone layer shows signs of recovery (Ball et al., 2018; Steinbrecht et al., 2018). Nonetheless, this recovery has not been reported in the areas where practically all inhabitants and cultivars are located Chipperfield et al. (2017), Weber et al. (2018). Some papers have even reported a decrease in stratospheric ozone in these latitudes since 1998 (Ball et al., 2018).
The most common effects induced by UV-B radiation in plants are biomass decrease (Tevini and Teramura, 1989; Vandenbussche et al., 2018), alterations in cuticle and epidermis (Tevini and Steinmüller, 1987), abnormal growth (Teramura and Sullivan, 1994; Searles et al., 2001), and damages to photosystems I and II (Pang and Hays, 1991; Liu et al., 2013). These effects can affect both cultivar quality and yield (Bais et al., 2018). It has been estimated that cultivar growth decreases approximately 1% for each 3% increase in UV-B radiation (Ballaré et al., 2011), but this could be more dramatic in UV-B sensitive cultivars (Hakala et al., 2002; Gao et al., 2003; Hidema and Kumagai, 2006; Zhua and Yang, 2015). UV-B radiation could also have effects on food quality, by affecting levels and composition of vitamins, fatty acids, polyphenols, and flavonoids or anthocyanins, thus modifying nutritional and organoleptic characteristics (Choudhary and Agrawal, 2016; Reddy et al., 2016; Tripathi and Agrawal, 2016; Nguyen et al., 2017; Wang et al., 2017; Wu et al., 2017).
Knowing how plants react to UV-B radiation is therefore crucial to establish damage reducing strategies for cultivars. In the past 20 years, there have been great advances in this area, but most of the work has been done using Arabidopsis thaliana as a model plant. One of the outstanding advances has been the characterization of the first specific UV-B photoreceptor: UV RESISTANCE LOCUS 8 (UVR8) (Rizzini et al., 2011; Christie et al., 2012; Wu et al., 2012).
The aim of this article is to review the current documented knowledge concerning plants' responses to UV-B radiation, venturing beyond the information available for Arabidopsis thaliana and focusing on other species. To achieve this, we spotlighted the responses mediated by the UVR8 receptor and the UV-B response repressors, RUP proteins (REPRESSOR OF UV-B PHOTOMORPHOGENESIS). Furthermore, we analyzed the participation of UVR8 in diverse stresses, fruit development, and UV-B-independent responses.
UVR8-mediated UV-B Signaling in Arabidopsis
Protein UV RESPONSE LOCUS 8 (UVR8) has been characterized as the UV-B radiation receptor (Kliebenstein et al., 2002; Brown et al., 2005; Rizzini et al., 2011; Christie et al., 2012; Jenkins, 2014). As shown in Figure 1, in presence of UV-B, UVR8 changes its quaternary structure from homodimer to active monomer, translocating from the cytoplasm to the nucleus, where it is functional (Kaiserli and Jenkins, 2007; Rizzini et al., 2011; Christie et al., 2012; Qian et al., 2016; Yin et al., 2016). Constitutively photomorphogenic 1 (COP1) is part of the E3 ubiquitin ligase complex, where it interacts with SUPPRESSOR OF PHYA (SPA1), poly-ubiquitinating the transcriptional factors ELONGATED HYPOCOTYL 5 (HY5) and HY5 HOMOLOG (HYH), which are subsequently degraded via proteasome (Lau and Deng, 2012; Huang et al., 2014). In presence of UV-B radiation, the UVR8 monomer interacts with COP1 and disengages COP1-SPA from the E3 ubiquitin ligase complex, avoiding ubiquitination and subsequent degradation of HY5 and HYH (Favory et al., 2009; Cloix et al., 2012). HY5 levels increase, inducing its own expression and regulating the expression of key genes in UV-B response (Brown et al., 2005; Binkert et al., 2014).
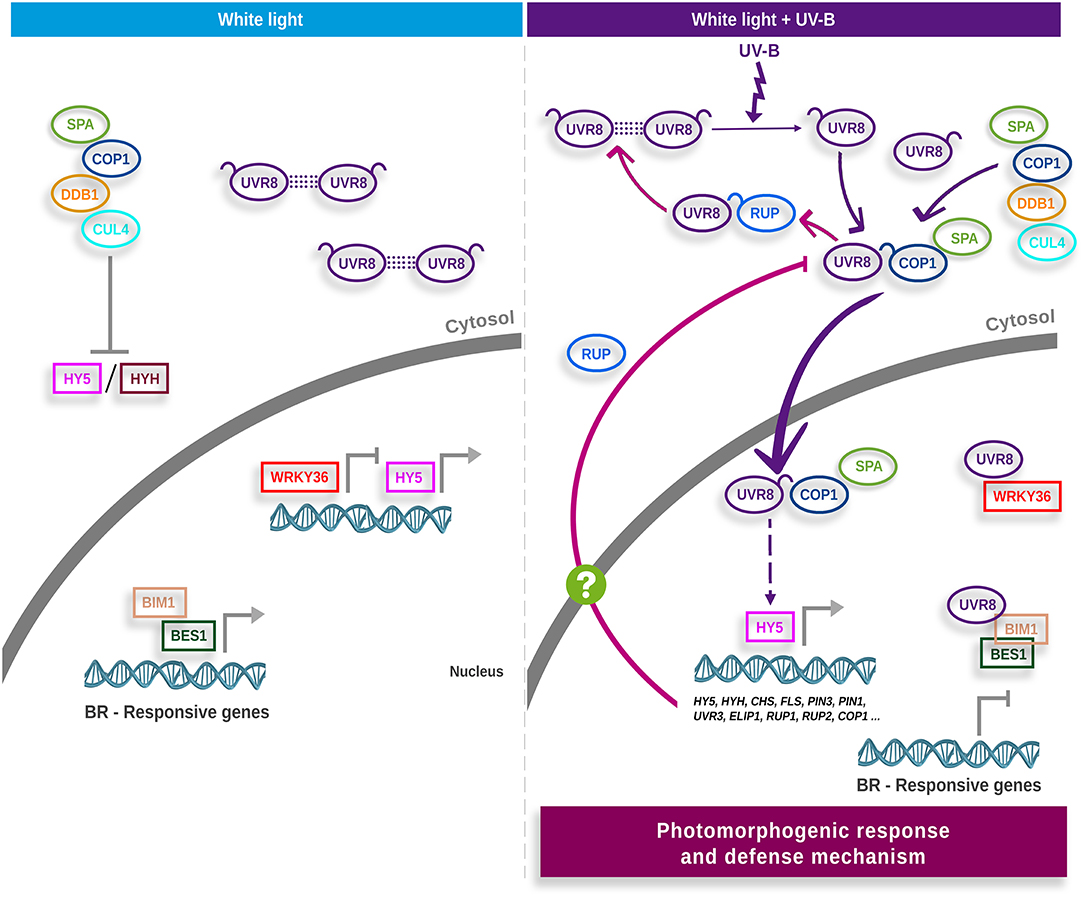
Figure 1. UVR8-mediated signal transduction model in Arabidopsis. In white light (left panel), the UV-B photoreceptor UVR8 homodimer and E3 ubiquitin ligase complex are located in the cytosol. The E3 ubiquitin ligase promotes degradation of the HY5 and HYH transcription factors. HY5, acting redundantly with HYH, mediates transcriptional responses. Transcription factors HY5, WRKY36, BIM1 and the functional BES1 are localized in the nucleus. HY5 binds to its own promoter to activate HY5 transcription, and WRKY36 also binds to the HY5 promoter to inhibit its transcription. BIM1 and BES1 act together to induce the expression of brassinosteroid (BR)-responsive genes. When plants are exposed to UV-B (right panel), the UVR8 homodimer is dissociated into active monomers. Monomeric UVR8 binds to COP1–SPA and elicits the COP1–SPA dissociation from the CUL4–DDB1 E3 ubiquitin ligase complex precluding HY5/HYH degradation. UVR8–COP1–SPA travels to the nucleus and facilitates HY5 protein stabilization and enhances the binding of HY5 to its own promoter. UVR8 monomer interacts with WRKY36 to inhibit WRKY36 binding to the HY5 promoter hence removing the inhibition of HY5 expression. In addition, the UVR8 monomer interacts with BIM1 and the functional dephosphorylated BES1 to inhibit their binding to the BR-induced genes involved in cell elongation, thus repressing the expression of BR-induced elongation genes and further repressing the BR-promoted plant growth. HY5 induces the transcription of key genes in the photomorphogenic response and defense mechanism. RUP1 y RUP2 are two of the genes induced by HY5. RUP proteins (RUP1 and RUP2) negatively regulate UVR8 by binding to the C27 region, displacing COP1, and promoting re-dimerization of the photoreceptor (pink arrows). RUP could act both in the nucleus and in the cytosol. Currently, the mechanism that transports RUP between the cytosol and the nucleus is unknown. The figure is based on the models proposed by Jenkins (2017) and Liang et al. (2019). UV-B, ultraviolet-B radiation; UVR8, UV RESISTANCE LOCUS 8; COP1, CONSTITUTIVELY PHOTOMORPHOGENIC 1; HY5, ELONGATED HYPOCOTYL 5; HYH, HY5 HOMOLOG; SPA1, SUPPRESSOR OF PHYA; DDB1, DNA DAMAGE-BINDING PROTEIN 1; CUL4, CULLIN 4; WRKY36, WRKY DNA-BINDING PROTEIN 36; BES1, BRI1-EMS-SUPPRESSOR1; BIM1, BES1-INTERACTING MYC-LIKE 1; BRs, Brassinosteroids; RUP1 and RUP2, REPRESSOR OF UV-B PHOTOMORPHOGENESIS 1 and 2; FLS, FLAVONOL SYNTHASE; UVR3, UV REPAIR DEFECTIVE 3; ELIP1, EARLY LIGHT-INDUCIBLE PROTEIN 1; CHS, CHALCONE SYNTHASE.
One of the genes induced by HY5 encodes for RUP (REPRESSOR OF UV-B PHOTOMORPHOGENESIS) proteins, which participate in the negative feedback regulation by binding to UVR8 and promoting its re-dimerization (Gruber et al., 2010; Heijde and Ulm, 2013). When plants are grown in diurnal periods, UVR8 reaches a dimer/monomer equilibrium in which RUP proteins are crucial for correct dimerization (Findlay and Jenkins, 2016). Other proteins also participate in the negative feedback regulation of UV-B: SALT TOLERANCE/BBX24 (STO/BBX24), RADICAL-INDUCED CELL DEATH1 (RCD1) and DWD HYPERSENSITIVE TO UV-B 1 (DHU1) (Jiang et al., 2009, 2012a,b; Kim et al., 2017). BBX24 binds to RCD1 and interacts with HY5 and COP1, stabilizing COP1, reducing HY5 accumulation and inhibiting its activity as a transcriptional factor (Jiang et al., 2012a). DHU1 sequesters COP1, found in the complex UVR8-COP1, diminishing the UV-B response (Kim et al., 2017).
On the other hand, it has been demonstrated that UVR8 monomers can interact with the transcriptional factors WRKY DNA-BINDING PROTEIN 36 (WRKY36), BRI1-EMS-SUPPRESSOR1 (BES1) and BES1-INTERACTINGMYC-LIKE 1 (BIM1) in the nucleus, as part of a signaling cascade in response to UV-B (Favory et al., 2009; Liang et al., 2018; Yang et al., 2018a). Free WRKY36 binds to the HY5 promoter and blocks its transcription. When UVR8 interacts with WRKY36, this factor is removed and HY5 can induce its expression (Yang et al., 2018a). BES1 and BIM1 are part of the Brassinosteroids' (BRs) signaling cascade (Yin et al., 2005). BES 1 interacts with BIM1 and binds to DNA, inducing BRs response genes (Yin et al., 2005; Belkhadir and Jaillais, 2015; Liang et al., 2018). The monomer UVR8 binds preferentially to dephosphorylated BES1, which is the active form, and by precluding its interaction with DNA, promotes one of the classic responses to UV-B: the inhibition of hypocotyl growth (Vert and Chory, 2006; Jiang et al., 2015; Liang et al., 2018).
UVR8: from Green Algae to Higher Plants
UVR8 is functional as a receptor because of amino acid sequences that are key for UV-B perception, homodimerization, and COP1 interaction. UV-B perception is achieved through tryptophan residues (O'Hara and Jenkins, 2012; Ulm and Jenkins, 2015). UVR8 monomers have three Gly-Trp-Arg-His-Thr motifs (GWRHT) that form a tryptophan triade (W233, W285, and W337), where W285 is the main UV-B sensor. Once the tryptophans absorb UV-B radiation, the dimer dissociates into its monomers (Christie et al., 2012; Wu et al., 2012). The interaction of UVR8 with COP1 may occur either in a UV-B dependent fashion, through a β-propeller domain, or constitutively, via core VP (Val-Pro) in the UVR8C27 domain (Cloix et al., 2012; Yin et al., 2015). By analyzing the amino acid sequences and focusing on the key motifs for UVR8 function, UVR8 sequences have been identified in green algae, bryophytes, lycophytes, and angiosperms (Rizzini et al., 2011; Wu et al., 2012; Fernández et al., 2016). These motifs are highly conserved from green algae to higher plants. At the moment, no UVR8 homologs in gymnosperms have been found or characterized, making it an issue worth exploring. Characterization studies for the UVR8 receptor have been performed in Arabidopsis (Rizzini et al., 2011; Cloix et al., 2012; Wu et al., 2012), and, in recent years, studies of UVR8 in other species have been undertaken. In green algae, such as Chlamydomonas reinhardtii (Cr), CrUVR8 dimers dissociate upon UV-B irradiation and re-form when transferred to white light. Interaction between CrUVR8 and CrCOP1 in Chlamydomonas has been confirmed in yeast two-hybrid assays. In addition, CrUVR8 complements the Arabidopsis uvr8 mutant, indicating that the molecular mechanism of action characterized in Arabidopsis is well-conserved (Tilbrook et al., 2016).
Soriano et al. (2018) studied the structure and function of UVR8 in two bryophytes, the moss Physcomitrella patens and the liverwort Marchantia polymorpha. Unlike Arabidopsis, which only has one UVR8 gene, P. patens expresses two UVR8 genes that encode functional proteins, whereas the single M. polymorpha UVR8 gene expresses two transcripts by alternative splicing, and the resulting proteins are functional. In Physcomitrella, the UVR8 dimer also dissociates in the presence of UV-B (as in Arabidopsis), but in its absence, both dimers and monomers were detected. However, in M. polymorpha, the UVR8 monomer is present independently of UV-B treatment and the dimer UVR8 is barely detected.
UVR8 is translocated from the cytoplasm into the nucleus, where it accumulates rapidly when Arabidopsis plants are exposed to UV-B (Kaiserli and Jenkins, 2007; Yin et al., 2016). Contrary to Arabidopsis, both M. polymorpha GFP-UVR8 fusion proteins encoded in uvr8 are found in the nucleus, independently of UV-B treatment. Nevertheless, both Marchantia and Physcomitrella UVR8 complement Arabidopsis uvr8 mutants, thus reinstating HY5 transcript levels, the accumulation of the chalcone-synthase enzyme (CHS), and the suppression of hypocotyl growth in response to UV-B.
In Angiosperms such as Chrysanthemum morifolium (Cm), Populus euphratica (Pe), and Malus domestica (Md), heterologous expression of CmUVR8, PeUVR8, and MdUVR8 rescued the deficient phenotype of uvr8 mutants in Arabidopsis in response to UV-B. In addition, the CmUVR8-CmCOP1, and MdUVR8-MdCOP1 interactions were confirmed using a yeast two-hybrid assay. PeUVR8-AtCOP1 interaction was verified using bimolecular fluorescence complementation (BiFC) assay (Mao et al., 2015; Zhao et al., 2016; Yang et al., 2018b).
In agreement with previous studies in Arabidopsis, overexpression of UVR8 in Solanum lycopersicum (Sl) increased tolerance to UV-B, and gene silencing in tomato lines (SlUVR8Ri) promoted UV-B hypersensibility (Li H. et al., 2018). UV-B response gene induction, such as HY5 and CHS, as well as anthocyanin accumulation were repressed in SlUVR8Ri, indicating that SlUVR8 plays an essential role in UV-B response. GFP-SlUVR8 proteins expressed in Nicotiana showed cytoplasm to nucleus translocation as reported for Arabidopsis (Li H. et al., 2018).
In all the species mentioned, the UVR8 receptor was functional and behaved similarly to the receptor described for Arabidopsis. This confirms the predicted presence of UVR8 in green algae, bryophytes, and angiosperms by bioinformatics analyses. The existence of UVR8 in ancestral organisms, such as green algae, could be related to the elevated UV-B dose in the primitive atmosphere and the ensuing self-protection required by the first photosynthetic plants (Jenkins, 2017).
UVR8 Expression in Different Plant Species
The expression of the UVR8 gene in Arabidopsis (AtUVR8) is ubiquitous and constitutive in plant tissues, and this includes roots, which do not receive direct light (Rizzini et al., 2011; Jenkins, 2017). Arabidopsis has only one copy of the UVR8 gene, while several species contain at least two UVR8 genes (Brown et al., 2005; Fernández et al., 2016). Although, at the moment, there is no evidence of functional differences between UVR8 proteins within analyzed species, the possession of multiple genes provides the potential for differential expression regulation.
As can be observed in Table 1, in species such as P. euphratica, S. lycopersicum, Betula platyphylla, C. morifolium, or Malus domestica, the UVR8 gene is expressed in all the tissues, as in Arabidopsis, but the transcript levels differ significantly between tissues (Mao et al., 2015; Li H. et al., 2018; Yang et al., 2018b). In Populus, expression of PeUVR8 is higher in shoots and leaves, declines in stems, and is weak in roots (Mao et al., 2015). In Solanum SlUVR8 expression levels are higher in flowers than in the other tissues analyzed (Li X. et al., 2018). The expression of BpUVR8 in Betula is highest in leaves and significantly reduced in xylem, phloem, and inflorescences (Li H. et al., 2018). In Chrysanthemum, expression is higher in leaves, followed by flowers and weakest in roots. MdUVR8 gene expression in apple trees is higher in stems, lower in fruits and roots, and very low in leaves and flowers (Zhao et al., 2016). Besides changes in expression levels among apple tissues, the abundance of the transcript MdUVR8 also varies throughout the development of the fruit (Henry-Kirk et al., 2018). In Vitis vinifera, expression of VvUVR8 in the grape berry varies, and it is four to five times higher in pre-verasion than in post-verasion (Liu et al., 2015).
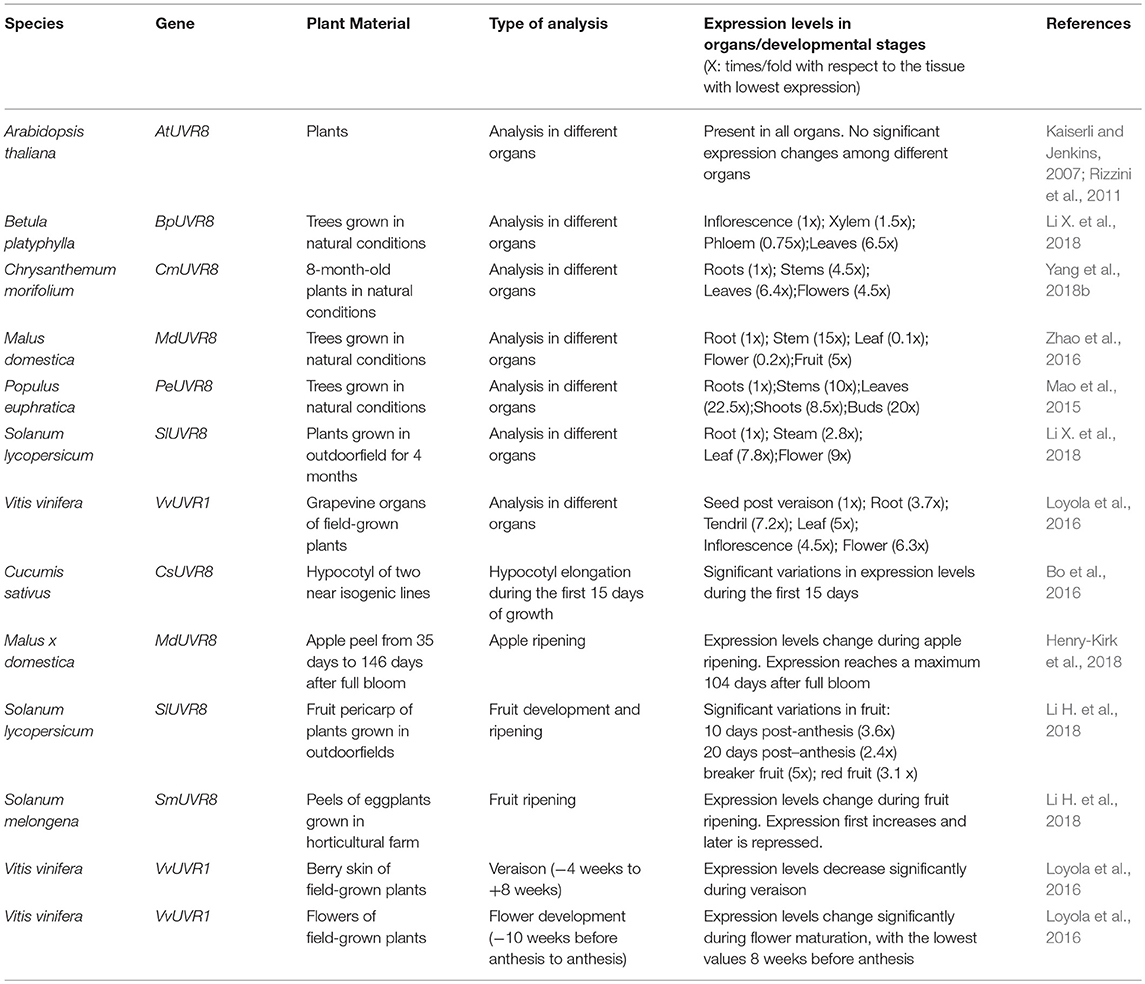
Table 1. UVR8 gene expression in different organs and developmental stages in diverse species.
The receptor's constitutive presence allows a rapid response to mitigate the damage caused by UV-B (Rizzini et al., 2011). Arabidopsis UVR8 over-expressing mutants are more tolerant and adapt better to UV-B (Favory et al., 2009; Morales et al., 2013). If we assume that higher UVR8 expression correlates with an increase in protein levels, differential expression in the diverse tissues could be associated with different degrees of UV-B tolerance throughout the plant.
AtUVR8 expression levels do not vary significantly in response to UV-B (Kliebenstein et al., 2002; Tohge et al., 2011; Fasano et al., 2014). Similarly, VvUVR8 expression does not change with UV-B treatment in berry skin (Carbonell-Bejerano et al., 2014). Whereas in M. polymorpha and P. patens, UVR8 expression remains constant in presence of UV-B (Soriano et al., 2018), there are some species where UVR8 expression varies significantly in response to UV-B (see Table 2). In birch tree, after 6 h exposure to UV-B, BpUVR8 expression increases 3.5 times with respect to the control, with a maximum peak at 9 h (4.65 times more than in the control) (Li J. et al., 2018). In Antarctic moss Pohlia mutans, expression levels of genes involved in the UV-B signaling pathway, PnUVR8, PnHY5 and PnCOP1, increase significantly after 3 h treatments with UV-B (Li et al., 2019). In another Antarctic species, such as Colobanthus quitensis, the increase in the transcription of UVR8 and COP1 is dose-dependent, reaching its peak 6 h after UV-B exposure (Contreras et al., 2019).
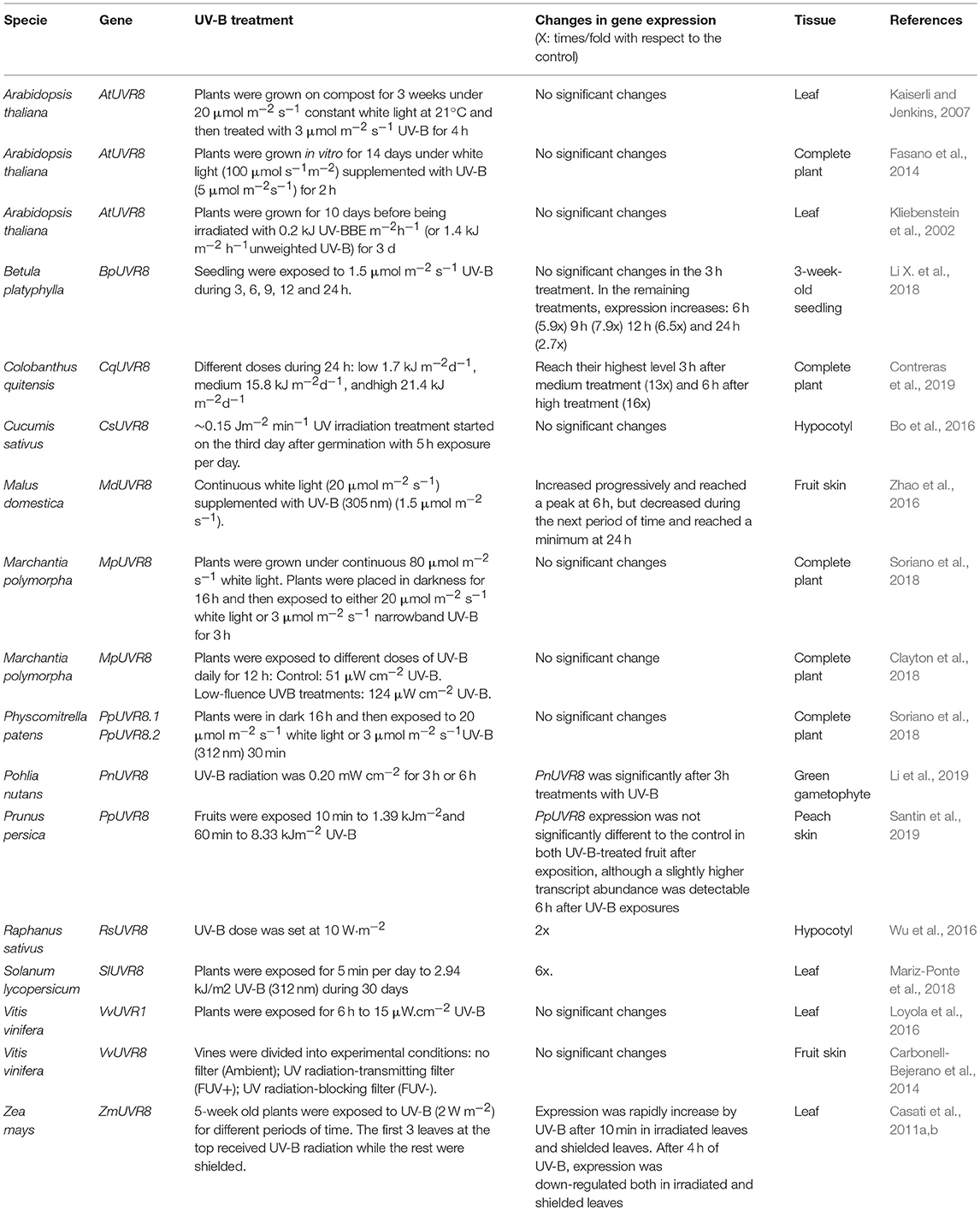
Table 2. UVR8 gene expression in different plants treated with UV-B.
UVR8 expression is repressed in maize leaves irradiated with UV-B for 1 h (Casati et al., 2011a). However, in the leaves shielded from UV-B, UVR8 expression is induced as part of a systemic response. The induction observed in the shielded leaves is overturned when the plants are exposed to UV-B more than 4 h (Casati et al., 2011a). In eggplant, as in maize, expression of the UVR8 transcript is induced with 0.5 h UV-B exposure and repressed after 4 and 5 h (Li J. et al., 2018).
In radish sprout hypocotyls, the UVR8 expression is induced significantly by UV-B (2 times with respect to the control) (Wu et al., 2016). Expression levels of PpUVR8 did not vary substantially in peach skin during treatment with UV-B, but increased slightly after exposure, reaching the highest peak at 6 h (Santin et al., 2019). In conclusion, there are some species in which UVR8 expression in response to UV-B is constant, as in Arabidopsis, but in most cases the expression levels vary.
Presence of RUP 1 and RUP2 in Different Species
RUP1, RUP2, and COP1 are members of the WD40-repeat protein family. RUP1 and RUP2 contain seven WD40-repeats with apparently no additional domains (Van Nocker and Ludwig, 2003; Li et al., 2013). In the presence of UV-B, levels of RUP proteins increase and their relative abundance (compared to COP1) interrupts the interaction UVR8-COP1, contributing to the reversal of UVR8 from monomer to homodimer. As a result, RUP proteins switch off the signaling cascade, thus averting problems in plant growth and development (Gruber et al., 2010; Heijde et al., 2013). In plants growing in diurnal photoperiods under natural light, a UVR8 dimer/monomer photo-equilibrium is established. In that state, RUP1 and RUP2 play a crucial role in the reversion of monomer to dimer, allowing for the plant's optimum growth and development in that specific environment (Findlay and Jenkins, 2016).
In addition to RUP proteins, STO/BBX24, RCD1, and DHU1 have also been characterized as negative regulators in the Arabidopsis UV-B signaling cascade (Jiang et al., 2009, 2012a,b; Kim et al., 2017). Most studies concerning the characterization of RUP proteins in response to UV-B have been performed in Arabidopsis (Gruber et al., 2010; Heijde et al., 2013; Vanhaelewyn et al., 2016). AtRUP1 (385aa) and AtRUP2 (368aa) proteins have a 64% identity (Favory et al., 2009; Supplementary Table 1) and both fulfill the same functions in the UV-B response (Vanhaelewyn et al., 2016). In order to search for RUP proteins in terrestrial plants, a blastp (blast protein, ncbi) was carried out using AtRUP1 (OAO92149.1) and AtRUP2 (OAO92900.1) as query sequences. As can be observed in Figure 2 and Supplementary Table 1, there are few RUP1 sequences with a high percent identity (91% to 64%). The sequences identified as RUP1 derive from species of the Brassicaceae family, as Arabidopsis. The one exception is a Tarenaya hassleriana sequence, which belongs to a Brassicaceae sister family. The high percent identity values are to be expected since the sequences derive from the same family.
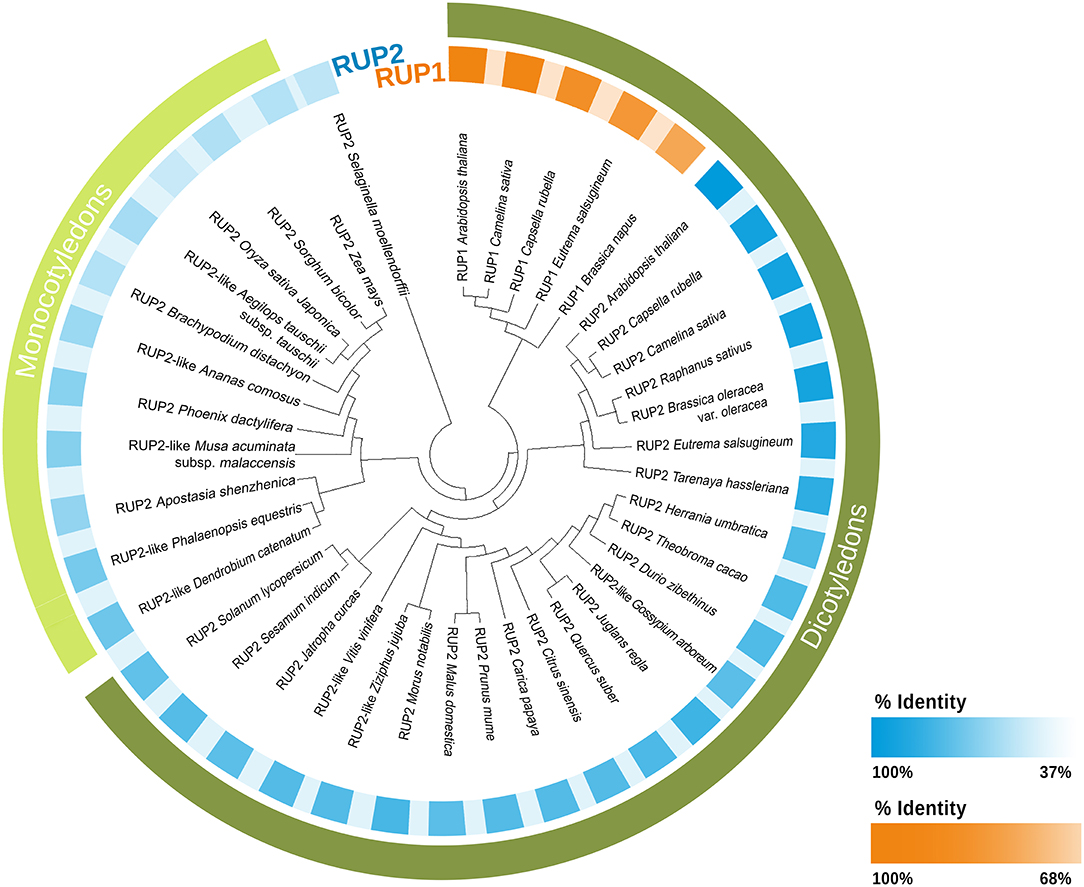
Figure 2. Phylogram of RUP1 and RUP2 proteins. RUPs proteins sequences of plant species and the percentage identity were obtained from the NCBI database (http://www.ncbi.nlm.nih.gov/) using the BLASTP method. The query sequences used were AtRUP1(OAO92149.1) and AtRUP2 (OAO92900.1). The tree was built using Maximum Likelihood method and JTT matrix-based model (Jones et al., 1992). Initial tree obtained by Neighbor-Join and BioNJ algorithms. Distances estimated using JTT. Evolutionary analyses were conducted in MEGA X (Kumar et al., 2018). The highest log likelihood: −15691.88. To make this tree we used some representative sequence of each species. For more information about sequences see Supplementary Table 1.
Recently, in experiments with mutants obtained through CRISPR/Cas9, it has been demonstrated that UV-B response in M. polymorpha includes many common components with Arabidopsis, among them RUP1 (Mapoly0094-s0072) (Clayton et al., 2018). In contrast to the RUP1 sequences detected, there is a higher number of RUP2 sequences detected (Figure 2 and Supplementary Table 1). The phylogram shows that RUP1 and RUP2 from monocotyledons and dicotyledons belong to different clusters (Figure 2).
The percent identity in the sequences identified as RUP2 in dicotyledons (93%-53%) is higher than in monocotyledons (lower than 55%). Due to the low percent identity of RUP2 proteins in monocotyledons and the fact that RUP1 sequences have not been detected in this group, it could be hypothesized that RUP proteins would not be part of the switch-off mechanism in monocotyledons.
One hypothesis is that mechanisms such as the ones mediated by STO/BBX24 and DHU1 suffice to switch off the UV-B response. Another possibility is the existence of proteins fulfilling the same functions as RUPs, but with a low percent identity. Yet another proposition is that the negative regulation mechanism could be based on direct repression of UVR8 expression or UVR8 monomer degradation, whenever necessary. Research has demonstrated that, in several species, UVR8 gene expression is regulated in the presence of UV-B (Table 2). In maize, for example, Casati et al. (2011a) reported that, after extended exposure to UV-B, UVR8 expression was repressed both in exposed and shielded leaves, possibly as part of negative feedback.
There are proteins that have a low percent identity but still exhibit conserved domains that allow them to fulfill the same functions. RUP proteins have seven WD40-repeats with apparently no additional domains (Gruber et al., 2010). It was observed that the C27 region of UVR8 interacts with the WD40 region of COP1 and RUP1/2 (Gruber et al., 2010; Cloix et al., 2012). It would be interesting to examine the degree of conservation of the WD40 domain among RUP2 proteins in monocotyledons and dicotyledons, and if it can interact with UVR8 as it does in Arabidopsis.
UVR8 in Fruit Developmental Stages
UV-B is an environmental signal that is perceived by plants, modulating growth, development, and metabolism in different organs (Tilbrook et al., 2013). By studying Arabidopsis mutants, the participation of UVR8 in many of the responses triggered by UV-B in leaf (Wargent et al., 2009), hypocotyl (Liang et al., 2019), root (Wan et al., 2018), and flower (Dotto et al., 2018) has been determined, but further research in fruit is required. UV-B promotes flavonoids and anthocyanins accumulation in Arabidopsis and other species (Lois, 1994; Ubi et al., 2006). In Arabidopsis leaves, synthesis of flavonoids and anthocyanins in response to solar UV-B radiation is regulated by the UVR8 receptor (Morales et al., 2013). The production of other pigments, such as carotenoids, is also induced by UV-B (Becatti et al., 2009). Furthermore, UV-B radiation increases fruit color in several fruit trees like grapes, apples, peaches, and blueberries, among others (Zhao et al., 2016; Escobar et al., 2017; Henry-Kirk et al., 2018). Fruit nutritional quality frequently depends on the content of those pigments, and their composition varies throughout the fruit's development and ripening (Wang et al., 2008).
In sunlight conditions, anthocyanins accumulate in the pericarp of Litchi chinensis. In Litchi, eight putative UVR8 encoding genes were identified, and their expression increases differentially when the fruit is exposed to UV radiation. The incremental expression of UVR8 is accompanied by the up-regulation of key genes in the anthocyanin biosynthesis pathway. In the skin of apples and eggplants, UVR8 expression increases in response to UV-B, together with a rise in flavonoid and anthocyanin levels. During fruit ripening, UVR8 expression changes (increments and decrements) were reported, suggesting physiological changes do not play a minor role in its regulation (Henry-Kirk et al., 2018; Santin et al., 2019). From this information we can infer that, in the presence of solar UV-B, UVR8 is involved in the skin coloring of litchi, apple and eggplant throughout fruit development.
Li H. et al. (2018) demonstrated that, in response to UV-B, UVR8 in tomato (SlUVR8) induces the expression of genes and the accumulation of light-absorbing compounds in leaves. Tomato plants containing silenced UVR8 (SIUVR8Ri) presented pale-green fruits when grown under natural sunlight. In contrast, in UVR8 over-expressing plants (SlUVR8OE), the fruits exhibited a darker green color compared to the wild-type (WT). Furthermore, the accumulation of starch, carotenoids, and lycopenes was higher in SlUVR8OE and lower in SIUVR8Ri fruits. The number of chloroplasts per cell was the same in WT and transgenic plants, but plastid size and thylakoid granules increased. The transcriptional factor GOLDEN2-LIKE (GLK2) determines chlorophyll accumulation and chloroplast development in fruits, inducing expression of genes associated to photosynthesis (Waters et al., 2008). When treated with UV-B radiation, SlUVR8 increased both GLK2 accumulation and that of its target genes (Li H. et al., 2018).
Pre and postharvest treatments with UV-B radiation have been employed to induce fruit ripening and pigment accumulation (Castagna et al., 2013). In grape berries, UV-B modifies the quantity and quality of flavonoids, improving their organoleptic properties (Martínez-Lüscherab et al., 2014). UV-B induction of secondary metabolites is not only beneficial to plants but also to human health (Schreiner et al., 2012).
In fruits exposed to UV-B, chloroplasts are larger and starch accumulation is higher, hence influencing the nutritional quality of the fruit in a UVR8 dependent pathway. In addition, it has been observed that, in fruit, UVR8 could induce the synthesis of pigments that absorb UV-B. Since UV-B induces pigment increase and metabolite accumulation, and this impacts directly on production and fruit quality, it would be interesting to examine if these processes are mediated by UVR8.
Different Inducers of UVR8 Expression
In Arabidopsis, UVR8 expression increases slightly in response to various abiotic stresses, except for the important increase when osmotic and salt stress are involved (Fasano et al., 2014; Table 3). Expression of the UVR8 transcript and protein levels increase in Arabidopsis plants grown in MS medium with NaCl and polyethylene glycol (PEG) for 24 h. In B. platyphylla, treatments with ABA and NaCl induced UVR8's promotor activity, resulting in an increase in BpUVR8 transcript levels (Li X. et al., 2018). Expression of UVR8 gene is induced in foxtail millet cultivar (cv. Prasad) 24 h after salt stress, while in cv. Lepakshi it decreases continuously (Puranik et al., 2011). In leaves of two drought-tolerant barley genotypes (Martin and Hordeum spontaneum 41-1 cultivars), UVR8 transcript levels are higher than in a drought-sensitive genotype (Moroc9-75), but the levels do not change when the three genotypes are exposed to drought stress (Guo et al., 2009). Interestingly, UVR8 expression in barley and millet differ among cultivars of the same species (Guo et al., 2009; Puranik et al., 2011). Under waterlogging conditions, ethylene promotes the formation of lysigenous aerenchyma in the root cortex of Zea mays. ZmUVR8 transcription is induced by waterlogging, mainly in the cortical cells (Rajhi et al., 2011). In radish sprouts, UV-B, cadmium, chilling, and salinity induced the expression of UVR8 (Wu et al., 2016). Additionally, exogenous addition of H2O2 and sodium nitroprusside (SNP, an NO donor) also induced UVR8 expression. This response is inhibited by dimethylthiourea (a H2O2 scavenger) and cPTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3 oxide, a NO scavenger) (Wu et al., 2016).
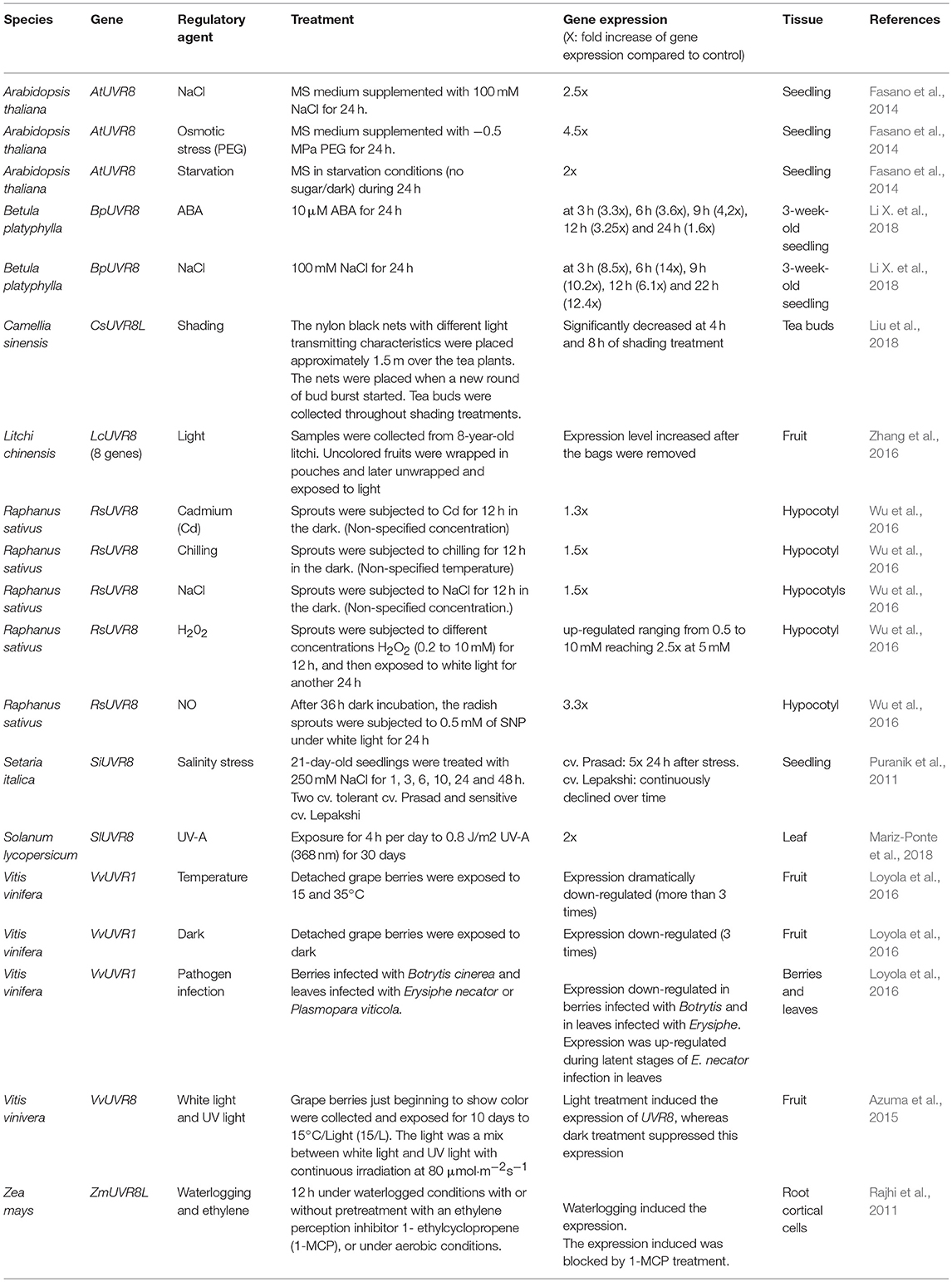
Table 3. Regulatory agents of UVR8 expression in different plants.
In Litchi fruits, differential expression of multiple UVR8 genes has been reported. Although the expression of all these genes is induced by light, maximum transcript levels are achieved at different times (Zhang et al., 2016). White light induces the expression of the UVR8 gene in radish and grape berries, while darkness suppresses it only in grape berries (Azuma et al., 2015; Wu et al., 2016). Loyola et al. (2016), who refer to VvUVR8 as UVR1, showed that UVR8 expression is subject to shading and elevated temperature in Vitis vinifera (grape) berries.
As in Arabidopsis, under shading treatment, the expression of UVR8 does not change significantly in Camellia sinensis tea buds. Interestingly, a unigene annotated as UVR8 LIKE decreases its expression with shading treatment (Liu et al., 2018).
As previously mentioned, UVR8 gene expression changes in response to UV-B radiation and other agents that provoke several stresses (see Tables 2, 3). Based in this information, we suggest that UVR8 acts as the common factor in the cross-talk among multiple stresses.
UVR8-independent UV-B Responses
UVR8 is the only UV-B receptor characterized so far (Rizzini et al., 2011), and it has been shown that it mediates a large number of UV-B specific responses (Wargent et al., 2009; Tossi et al., 2014; Li H. et al., 2018). In Arabidopsis leaves exposed to low levels of UV-B, UVR8 regulates the expression of a wide number of key genes in the photomorphogenic and acclimation response. Conversely, under high UV-B doses, the regulation of WRKY, FAD oxide reductase, and UDPgtfp expression is UVR8-independent (Brown and Jenkins, 2008). The expression of these genes varies in response to several abiotic stresses and is induced by H2O2 (Inzé et al., 2011; Chen et al., 2012). ARIADNE 12 (ARI12) belongs to a family of proteins with E3 Ubiquitin ligase activity induced by UV-B (Eisenhaber et al., 2007). In low UV-B fluence rate conditions, ARI12 transcription is induced only in WT and not in the uvr8 mutants, indicating that gene induction is mediated by the UVR8 receptor. On the contrary, with high UV-B fluence rates, ARI12 transcript levels increased in both genotypes. Thus, expression of ARI12 is induced through a UVR8-dependent pathway in low UV-B fluence rate conditions and by an independent one in high fluence rates (Lang-Mladek et al., 2011). Mitogen-activated protein kinase (MAPK) networks are activated by diverse stresses (Holley et al., 2003). Gonzalez Besteiro et al. (2011) demonstrated that UV-B stress activates the signaling cascade mediated by MAPK1 independently from UVR8. When exposed to UV-B doses that cause acute UV-B stress, the MKP1 mutant (mkp1) is hypersensitive to radiation, while the uvr8 mutant is not. On the contrary, at low UV-B doses, the uvr8 mutant is impaired, but the mkp1 mutant is not. This indicates that, in response to damaging UV-B doses, MKP1 has a main role, while UVR8 is more important at low levels of UV-B light. Thus, the mkp1 and uvr8 mutants allow the genetic dissection of two UV-B response pathways that coordinately determine plant UV-B tolerance.
All this information demonstrates that the participation of UVR8 in the UV-B response is UV-B dose-dependent. UVR8 mediates several responses to low doses of UV-B, while high UV-B doses trigger other adaptive mechanisms. However, other findings suggest there are specific responses to low doses of UV-B that are independent of UVR8. Exposure to UV-B inhibits leaf growth in various plant species (Liu et al., 1995; Searles et al., 2001) due to a reduction in the number of epidermal cells. In both uvr8-2 and WT plants, the number of epidermal cells per leaf is reduced by UV-B, which suggests that the control of epidermal cell division is independent of UVR8 (Wargent et al., 2009). Moreover, by exposing uvr8 mutants to outdoor conditions, it was observed that UV-B radiation affected morphology by a UVR8-independent mechanism (Coffey et al., 2017). At the DNA level, UV-B radiation induces cyclobutane pyrimidine dimer (CPD) formation, which inhibits transcription and replication, and induces mutations. The activity of CPD photolyases is thus essential for protecting genome integrity from UV-B radiation. Expression of the CPD photolyase (PHR) gene is mediated by both UVR8-dependent and UVR8-independent pathways. In uvr8-6 mutants, the expression of AtPHR is induced by low doses of UV-B, which generate CPDS and not ROS (Li et al., 2015).
These responses, which appear at UV-B doses that do not produce oxidative stress and are independent of UVR8, would suggest the existence of new and/or different UV-B receptor/s.
Discussion
UVR8 in Green Algae, Bryophytes and Angiosperms
The UV-B light receptor has been thoroughly characterized in Arabidopsis since Kliebenstein first identified it in 2002 (Kliebenstein et al., 2002). Over the last 4 years, research on UVR8 has been conducted for other plant species such as apple, tomato, grape, etc. (Loyola et al., 2016; Zhao et al., 2016; Li H. et al., 2018). Employing bioinformatic analyses, the protein's functional motifs have shown to be widely conserved in green algae, bryophytes, lycophytes and angiosperms (Fernández et al., 2016), but it has not been found in gymnosperms yet. Fernández et al. (2016) suggest that this might be due to the absence of whole genome sequences for gymnosperm species. Gymnosperms are more tolerant than Angiosperms to UV-B injury because their epidermal cells and leaf anatomy are more effective at attenuating UV-B radiation (Bassman et al., 2001). In some outdoor-grown gymnosperms, such as Scots pine and Norway spruce, UV-B radiation has no significant effects on growth or secondary compounds (Turtola et al., 2006). Based on this information, we propose that UVR8 sequences in gymnosperm have not been detected because either the receptor is unnecessary for this group or there is another receptor.
The UVR8 gene copy number changes between plant species. Chlorophytes contain a single copy of UVR8, same as in Arabidopsis, but bryophytes contain two copies. As in bryophytes, 40% of monocot species contain two UVR8 copies and 32% of dicots contain more than one UVR8 copies (Brown et al., 2005; Fernández et al., 2016). The difference in the UVR8 gene copy number among plant species is an interesting subject of debate. There are several articles that show that an increase in gene copy number and polyploidy contribute in the adaptation to stress (te Beest et al., 2011; Soltis et al., 2015; Panchy et al., 2016).
Chlorophytes mark the first appearance of the UVR8 gene in the Viridiplantae kingdom (Fernández et al., 2016). Chlorophytes live in water bodies where UV-B is 100% filtered within the first meter of depth (Zellmer, 1995). This group is able to float, thus allowing green algae to regulate the UV-B doses to which they are exposed. The bryophytes were the first terrestrial plants; they were exposed to high UV-B doses and could not avoid the radiation. In this condition, the presence of two UVR8 gene copies could give them an additional UV-B stress tolerance. When higher plants colonized the earth, some of them grew in places characterized by high UV-B doses and others by low UV-B doses. Under this heterogeneous scenario, the loss or gain of a gene copy could be an adaptive advantage. Interestingly, a study of the genome of the marine Angiosperm Zostera marina claims that UVR8 is not present (Olsen et al., 2016). This plant is found in mostly shallow coastal soft bottom environments with reasonably high water clarity to allow growth at a depth enough to filter almost all UV-B light (Short and Coles, 2001).
It will be interesting to evaluate a potential correlation between the number of UVR8 gene copies and the tolerance to UV-B stress or even the diversity of different eco-geographical niches a species can colonize. Elucidating if higher copy numbers of UVR8 are linked to an increase in the protein levels and if the multiple copies have redundant roles are also important challenges.
At the protein level, UVR8 has been characterized in Arabidopsis and in others plants species with similar results. However, contrary to Arabidopsis, in M. polymorpha, the UVR8 receptor is mainly present as a monomer in absence of UV-B, probably due to the poor structural stability of the dimer (Soriano et al., 2018). Furthermore, the subcellular localization differs from Arabidopsis, with MpUVR8 found constitutively in the nucleus (Soriano et al., 2018). Despite these differences, both proteins are functional.
Functional motifs that regulate protein stability and localization are conserved between species (Soriano et al., 2018). The observed differences could be attributed to small variations in amino acid sequences localized outside these motifs. This information suggests that amino acid sequences important to UVR8 behavior could remain unidentified.
UVR8 Gene Is Differentially Expressed in Plant Organs and Throughout Fruit Development
UVR8 expression is ubiquitous in every species analyzed so far (Rizzini et al., 2011; Jenkins, 2017; Table 1). Although in some species UVR8 expression is constitutive, in other species the expression varies in a tissue- and development-dependent fashion (Table 1). In fruits, for example, UVR8 expression is induced during development (Table 1). Some plant species have several UVR8 genes, e.g., litchi fruits (Fernández et al., 2016; Zhang et al., 2016). The UVR8 gene copy number could account for the differential expression observed in this plant. It will be thus interesting to analyze differences in the relative levels of either UVR8 transcripts during development.
UVR8 is expressed and UVR8 protein is detected in all plant organs (Rizzini et al., 2011). Transcript levels are higher in leaf, flower and fruit than in root (Table 1). This difference between plant organs could be associated with the necessity of the plant to protect itself from UV-B. On the other hand, the detection of UVR8 transcript in root, an organ that does not receive UV-B light, suggests other functions for UVR8 beyond light perception. Recently, using a Nicotiana silenced roots (irNaUVR8) approach, it has been demonstrated that UVR8 mediates colonization of Deinococcus (Santhanam et al., 2017).
Throughout fruit developmental stages, UVR8 transcript levels change (Table 1), indicating the possible participation of UVR8 in this process. Li H. et al. (2018) have determined that UVR8 improves chloroplast development in the fruit through the regulation of SlGLK2 in tomato grown outdoors. The study of UVR8 in different species has allowed the identification of new UVR8 functions demonstrating that UVR8 not only participates in the perception of UV-B radiation.
UV-B Regulates UVR8 Transcript Expression in Different Plant Species
UVR8 expression is constitutive in Arabidopsis plants exposed to UV-B (Kaiserli and Jenkins, 2007; Fasano et al., 2014). However, changes in the levels of the UVR8 transcript were detected in several plants treated with UV-B (Table 2). The levels of UVR8 transcripts induced by UV-B increase in the first hours of treatment, but later decrease (Table 2). In Zea mays, UVR8 expression was transient and also systemic in shielded leaves of UV-B exposed plants (Casati et al., 2011a). This information suggests the existence of a mechanism regulating UVR8 gene expression by UV-B.
UVR8 transcript level induction is an energetically efficient mechanism in most plants, since the receptor is synthesized only when required. This, however, implies a slower adaptation to UV-B acute irradiation. In Arabidopsis, in turn, constitutive expression and dynamic regulation, which depends on the UVR8 dimer-monomer equilibrium, allows for an immediate –although more energetically expensive- response (Rizzini et al., 2011). Selection of one strategy in lieu of the other could be due to a difference in tolerance to UV-B and, in some instances, both modes of UV-B acclimation could be active together, depending on the levels of perceived radiation.
RUP1 and RUP2 are negative feedback regulators of the UV-B signaling cascade. Both proteins interact with UVR8-COP1, facilitating UVR8 redimerization (Gruber et al., 2010; Heijde et al., 2013). In monocotyledons, we found RUP2 sequences with percentages of similarity lower than 50% compared to Arabidopsis, but no sequences similar to RUP1 (Figure 2). In this group, UVR8 activity regulation could be a balance between UVR8 synthesis and degradation and re-dimerization mediated by RUPs proteins would not be necessary. Nonetheless, the coexistence of both mechanisms of UVR8 activity regulation in some plants cannot be ruled out.
UVR8 Participates in Response to Multiple Stresses
Several reports have demonstrated that UVR8 participates in diverse stresses, often in combination with UV-B (Demkura and Ballaré, 2012; Fasano et al., 2014; Santhanam et al., 2017). The metabolites produced in UV-B response mediated by UVR8 are beneficial when the plant faces other stresses (Demkura and Ballaré, 2012). As shown in Tables 2, 3, UVR8 expression is induced by UV-B, salinity, starvation, pathogen infection, waterlogging, UV-A, chilling, etc. The regulation of UVR8 transcript levels in response to different stresses suggests a possible participation of the receptor. The UVR8 transcript expression in leaves of two barley genotypes tolerant to drought are higher in cultivars Martin and Hordeum spontaneum 41-1 compared to a genotype sensitive to drought (Moroc9-75) (Guo et al., 2009). The correlation between the UVR8 expression levels—stress tolerance, and the evaluation of the role of UVR8 in those stresses have not yet been determined.
ABA plays an important role both in plant development and in response to abiotic stresses (Tuteja, 2007; Trivedi et al., 2016). In plants exposed to high doses of UV-B, ABA production is induced, activating NADPH oxidase and generating H2O2, and also increasing NO production, which abates the damage caused by UV-B (Tossi et al., 2009, 2012). In radish hypocotyls, UV-B, H2O2 and nitric oxide (NO) induce UVR8 expression, whereas a chemical trap for H2O2 represses it (Wu et al., 2016). UVR8 transcript expression is induced by salinity, chilling and osmotic stress (Table 3). Interestingly, these stresses are regulated by ABA (Tuteja, 2007). Treatments with ABA, salt stress, high temperatures and UV-B increase the activity of the BpUVR8 promoter in transgenic tobacco leaves (Li X. et al., 2018). ABA could play a key role in the induction of UVR8 expression in abiotic stresses and development. The interaction between ABA and UVR8 should be thoroughly examined. The identification of specific motives in UVR8 promoters of different species could be an approach to identify other regulatory agents of UVR8 transcript expression.
Conclusion
The advantage of working with Arabidopsis is evident and, in this particular case, it has facilitated the characterization of UVR8. However, results gathered from different plant species suggest that the results obtained with Arabidopsis cannot be extended wholly to all plants due to the extant diversity. The study of UVR8 physiology in several species beyond Arabidopsis has enabled to establish that:
- UVR8 is highly conserved and is functional in green algae, bryophytes and angiosperms.
- There are species that present more than one copy of UVR8 gene, and in some species the gene can undergo alternative splicing, generating two UVR8 functional proteins.
- The UVR8 gene is expressed differentially in different organs and throughout fruit development.
- UVR8 is involved in several pathways other than UV-B perception. In tomato fruit, for instance, UVR8 regulates the development of the chloroplast and the synthesis of chlorophyll, carotenoids, starch and lycopenes, thus influencing its nutritional quality. In Nicotiana roots, it mediates colonization by Deinococcus.
- UVR8 transcript expression is regulated by UV-B, H2O2, ABA and other stresses.
The information obtained using different plant species show that UVR8 is a versatile molecule involved in perception of UV-B, as well as in developmental and stress processes. However, there is a lack of knowledge regarding UVR8 activity and its importance in these responses. Some important questions that arise are, among others, the following:
• How is UVR8 regulated during these processes?
• Is there a correlation between the UVR8 transcript levels and stress tolerance?
• How does negative feedback work in plant species where RUP 1 has not been detected and RUP2 has a low similarity percentage?
• How does UVR8 participate in developmental processes under natural growth environments?
Only a comprehensive consideration of UVR8 physiology in different species will allow us to bring light into the mechanisms and importance of this key modulator of several aspects involving plant evolution, development and survival. In addition the information derived from the analysis of the conservation and/or the variability of the responses in different plants might be harnessed to improve cultivar management by means of biotechnological approaches.
Author Contributions
VT and JR conceived the work and wrote the manuscript, except sections UVR8: from green algae to higher plants and UVR8 in fruit developmental stages. JI wrote section UVR8 in fruit developmental stages UVR8-independent UV-B responses. LL contributed in writing the Discussion and reviewing critically the manuscript. HB wrote section UVR8: from green algae to higher plants UVR8: from green algae to higher plants. AE supervised the writing of the manuscript. SP-Á coordinated the work and carried out the native English edition of the manuscript.
Funding
This work was supported by grants from the Consejo Nacional de Investigaciones Científicas y Técnicas, the Agencia Nacional de Promoción Científica y Tecnológica and Universidad de Buenos Aires.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Mariana Valente and Lic. Axel Rizzo for helping us with the edition of the Figures. We also thank Dr. Juan Pablo Cordoba for helping with the manuscript revision and for his helpful comments.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2019.00780/full#supplementary-material
References
Azuma, A., Fujii, H., Shimada, T., and Yakushiji, H. (2015). Microarray analysis for the screening of genes inducible by light or low temperature in post-veraison grape berries. Hort. J. 84, 214–226. doi: 10.2503/hortj.MI-041
Bais, A. F., Lucas, R. M., Bornman, J. F., Williamson, C. E., Sulzberger, B., Austin, A. T., et al. (2018). Environmental effects of ozone depletion, UV radiation and interactions with climate change: UNEP environmental effects assessment panel, update 2017. Photochem. Photobiol. Sci. 17, 127–179. doi: 10.1039/c7pp90043k
Ball, W. T., Alsing, J., Mortlock, D. J., Staehelin, J., Haigh, J. D., Peter, T., et al. (2018). Evidence for a continuous decline in lower stratospheric ozone offsetting ozone layer recovery. Atmos. Chem. Phys. 18, 1379–1394. doi: 10.5194/acp-18-1379-2018
Ballaré, C. L., Caldwell, M. M., Flint, S. D., Robinson, S. A., and Bornman, J. F. (2011). Effects of solar ultraviolet radiation on terrestrial ecosystems. Patterns, mechanisms, and interactions with climate change. Photochem. Photobiol. Sci. 10, 226–241. doi: 10.1039/c0pp90035d
Bassman, J., Robberecht, R., and Edwards, G. (2001). Effects of enhanced UV-B radiation on growth and gas exchange in Populus deltoides Bartr. ex Marsh. Int. J. Plant Sci. 162, 103–110. doi: 10.1086/317901
Becatti, E., Petroni, K., Giuntini, D., Castagna, A., Calvenzani, V., Serra, G., et al. (2009). Solar UV-B radiation influences carotenoid accumulation of tomato fruit through both ethylene-dependent and -independent mechanisms. J. Agric. Food Chem. 57, 10979–10989. doi: 10.1021/jf902555x.
Belkhadir, Y., and Jaillais, Y. (2015). The molecular circuitry of brassinosteroid signaling. N Phytol. 206, 522–540. doi: 10.1111/nph.13269
Binkert, M., Kozma-Bognár, L., Terecskei, K., De Veylder, L., Nagy, F., and Ulm, R. (2014). UV-B-responsive association of the Arabidopsis bZIP transcription factor ELONGATED HYPOCOTYL5 with target genes, including its own promoter. Plant Cell 26, 4200–4213. doi: 10.1105/tpc.114.130716
Björn, L. O. (1996). Effects of ozone depletion and increased UV-B on terrestrial ecosystems. Int. J. Environ. Stud. 51, 217–243. doi: 10.1080/00207239608711082
Björn, L. O. (2015). Ultraviolet-A, B and C. UV4Plants Bulletin 1, 17–18. doi: 10.19232/uv4pb.2015.1.12
Bo, K., Wang, H., Pan, Y., Behera, T. K., Pandey, S., Wen, C., et al. (2016). SHORT HYPOCOTYL1 encodes a SMARCA3-Like Chromatin Remodeling Factor Regulating Elongation. Plant Physiol. 172, 1273–1292. doi: 10.1104/pp.16.00501
Brown, B., and Jenkins, G. (2008). UV-B signaling pathways with different fluence-rate response profiles are distinguished in mature Arabidopsis leaf tissue by requirement for UVR8, HY5, and HYH. Plant Physiol. 146, 576–588. doi: 10.1104/pp.107.108456
Brown, B. A., Cloix, C., Jiang, G. H., Kaiserli, E., Herzyk, P., Kliebenstein, D. J., et al. (2005). A UV-B-specific signaling component orchestrates plant UV protection. Proc. Natl Acad. Sci. U.S.A. 102, 18225–18230. doi: 10.1073/pnas.0507187102
Carbonell-Bejerano, P., Diago, M. P., Martínez-Abaigar, J., Martínez-Zapater, J. M., Tardáguila, J., and Núñez-Olivera, E. (2014). Solar ultraviolet radiation is necessary to enhance grapevine fruit ripening transcriptional and phenolic responses. BMC Plant Biol. 14:183. doi: 10.1186/1471-2229-14-183
Casati, P., Campi, M., Morrow, D. J., Fernandes, J. F., and Walbot, V. (2011a). Transcriptomic, proteomic and metabolomic analysis of UV-B signaling in maize. BMC Genom 12:321. doi: 10.1186/1471-2164-12-321
Casati, P., Morrow, D. J., Fernandes, J., and Walbot, V. (2011b). Rapid maize leaf and immature ear responses to UV-B radiation. Front. Plant Sci. 2:33. doi: 10.3389/fpls.2011.00033
Castagna, A., Chiavaro, E., Dall'Asta, C., Rinaldi, M., Galaverna, G., and Ranieria, A. (2013). Effect of postharvest UV-B irradiation on nutraceutical quality and physical properties of tomato fruits. Food Chem. 137, 151–158. doi: 10.1016/j.foodchem.2012.09.095
Cejka, C., Ardan, T., Širc, J., Michálek, J., Beneš, J., Brunová, B., et al. (2011). Hydration and transparency of the rabbit cornea irradiated with UVB-doses of 0.25 J/cm2 and 0.5 J/cm2 compared with equivalent UVB radiation exposure reaching the human cornea from sunlight. Curr. Eye Res. 36, 607–613. doi: 10.3109/02713683.2011.574332
Chen, L., Song, Y.u., Li, S., Zhang, L., Zou, C., and Yu, D. (2012). The role of WRKY transcription factors in plant abiotic stresses. Biochim. Biophys. Acta. 1819, 120–128. doi: 10.1016/j.bbagrm.2011.09.002
Chipperfield, M. P., Bekki, S., Dhomse, S., Harris, N. R. P., Hassler, B., Hossaini, R., et al. (2017). Detecting recovery of the stratospheric ozone layer. Nature 549, 211–218. doi: 10.1038/nature23681
Choudhary, K. K., and Agrawal, S. B. (2016). Assessment of fatty acid profile and seed mineral nutrients of two soybean (Glycine max L.) cultivars under elevated ultraviolet-B: Role of ROS, pigments and antioxidants. Photochem. Photobiol. 92, 134–143. doi: 10.1111/php.12544
Christie, J. M., Arvai, A. S., Baxter, K. J., Heilmann, M., Pratt, A. J., O'Hara, A., et al. (2012). Plant UVR8 photoreceptor senses UV-B by tryptophan mediated disruption of cross-dimer salt bridges. Science 335, 1492–1496. doi: 10.1126/science.1218091
Clayton, W. A., Albert, N. W., Thrimawithana, A. H., McGhie, T. K., Deroles, S. C., Schwinn, K. E., et al. (2018). UVR8-mediated induction of flavonoid biosynthesis for UVB tolerance is conserved between the liverwort Marchantia polymorpha and flowering plants. Plant J. 96, 503–517. doi: 10.1111/tpj.14044
Cloix, C., Kaiserli, E., Heilmann, M., Baxter, K., Brown, B., O'Hara, A., et al. (2012). C-terminal region of the UV-B photoreceptor UVR8 initiates signaling through interaction with the COP1 protein. Proc. Natl. Acad. Sci. U.S.A. 109, 16366–16370. doi: 10.1073/pnas.1210898109
Coffey, A., Prinsen, E., Jansen, M., and Conway, J. (2017). The UVB photoreceptor UVR8 mediates accumulation of UV absorbing pigments, but not changes in plant morphology, under outdoor conditions. Plant Cell Environ. 40, 2250–2260. doi: 10.1111/pce.13025
Contreras, R. A., Pizarro, M., Köhler, H., Zamora, P., and Zúñiga, G. E. (2019). UV-B shock induces photoprotective flavonoids but not antioxidant activity in Antarctic Colobanthus quitensis (Kunth) Bartl. Environ. Exper. Bot. 159, 179–190. doi: 10.1016/j.envexpbot.2018.12.022
Demkura, P., and Ballaré, C. (2012). UVR8 mediates UV-B-induced Arabidopsis defense responses against Botrytis cinerea by controlling sinapate accumulation. Mol. Plant 5, 642–652. doi: 10.1093/mp/sss025
Dotto, M., Gómez, M. S., Soto, M. S., and Casati, P. (2018). UV-B radiation delays flowering time through changes in the PRC2 complex activity and miR156 levels in Arabidopsis thaliana. Plant Cell Environ. 41, 1394–1406. doi: 10.1111/pce.13166
Eisenhaber, B., Chumak, N., Eisenhaber, F., and Hauser, M. T. (2007). The ring between ring fingers (RBR) protein family. Genome Biol. 8:209. doi: 10.1186/gb-2007-8-3-209
Escobar, A., de Oliveira Silva, F., Acevedo, P., Nunes-Nesi, A., Alberdi, M., and Reyes-Díaz, M. (2017). Different levels of UV-B resistance in Vaccinium corymbosum cultivars reveal distinct backgrounds of phenylpropanoid metabolites. Plant Physiol. Biochem. 118, 541–550. doi: 10.1016/j.plaphy.2017.07.021
Fasano, R., Gonzalez, N., Tosco, A., Dal Piaz, F., Docimo, T., Serrano, R., et al. (2014). Role of Arabidopsis UV RESISTANCE LOCUS 8 in plant growth reduction under osmotic stress and low levels of UV-B. Mol. Plant 7, 773–791. doi: 10.1093/mp/ssu002
Favory, J. J., Stec, A., Gruber, H., Rizzini, L., Oravecz, A., Funk, M., et al. (2009). Interaction of COP1 and UVR8 regulates UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. EMBO J. 28, 591–601. doi: 10.1038/emboj.2009.4
Fernández, M. B., Tossi, V., Lamattina, L., and Cassia, R. (2016). A comprehensive phylogeny reveals functional conservation of the UV-B photoreceptor UVR8 from green algae to higher plants. Front. Plant Sci. 7:1698. doi: 10.3389/fpls.2016.01698
Findlay, K., and Jenkins, G. (2016). Regulation of UVR8 photoreceptor dimer/monomer photo-equilibrium in Arabidopsis plants grown under photoperiodic conditions. Plant Cell. Environ. 39, 1706–1714. doi: 10.1111/pce.12724
Frohnmeyer, H., and Staiger, D. (2003). Ultraviolet-B radiation-mediated responses in plants. Balancing damage and protection. Plant physiol. 133, 1420–1428. doi: 10.1104/pp.103.030049
Gao, W., Zheng, Y. F., Slusser, J. R., and Heisler, G. M. (2003). Impact of enhanced ultraviolet-B irradiance on cotton growth, development, yield, and qualities under field conditions. Agric. Forest Meteorol. 120, 241–248. doi: 10.1016/j.agrformet.2003.08.019
Gonzalez Besteiro, M., Bartels, S., Albert, A., and Ulm, R. (2011). Arabidopsis MAP kinase phosphatase 1 and its target MAP kinases 3 and 6 antagonistically determine UV-B stress tolerance, independent of the UVR8 photoreceptor pathway. Plant J. 68, 727–737. doi: 10.1111/j.1365-313X.2011.04725.x
Gruber, H., Heijde, M., Heller, W., Albert, A., Seidlitz, H. K., and Ulm, R. (2010). Negative feedback regulation of UV-B-induced photomorphogenesis and stress acclimation in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 107, 20132–20137. doi: 10.1073/pnas.0914532107
Guo, P., Baum, M., Grando, S., Ceccarelli, S., Bai, G., Li, R., et al. (2009). Differentially expressed genes between drought-tolerant and drought-sensitive barley genotypes in response to drought stress during the reproductive stage. J. Exp. Bot. 60, 3531–3544. doi: 10.1093/jxb/erp194
Hakala, K., Jauhiainen, L., Koskela, T., Kayhko, P., and Vorne, V. (2002). Sensitivity of crops to increased ultraviolet radiation in northern growing conditions. J. Agron. Crop Sci. 188, 8–18. doi: 10.1046/j.1439-037x.2002.00536.x
Heijde, M., Binkert, M., Ruohe, Y., Ares-Orpel, F., Rizzini, L., Van De Slijke, E., et al. (2013). Constitutively active UVR8 photoreceptor variant in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 110, 20326–20331. doi: 10.1073/pnas.1314336110
Heijde, M., and Ulm, R. (2013). Reversion of the Arabidopsis UV-B photoreceptor UVR8 to the homodimeric ground state. Proc. Natl. Acad. Sci. U.S.A. 110, 1113–1118. doi: 10.1073/pnas.1214237110
Henry-Kirk, R., Plunkett, B., Hall, M., McGhie, T., Allan, A., Wargent, J., et al. (2018). Solar UV light regulates flavonoid metabolism in apple (Malus x domestica). Plant Cell Environ. 41, 675–688. doi: 10.1111/pce.13125
Hidema, J., and Kumagai, T. (2006). Sensitivity of rice to ultraviolet-B radiation. Ann. Bot. 97, 933–942. doi: 10.1093/aob/mcl044
Holley, S. R., Yalamanchili, R. D., Moura, D. S., Ryan, C. A., and Stratmann, J. W. (2003). Covergence of signaling pathways induced by systemin, oligosaccharide elicitors, and ultraviolet-B radiation at the level of mitogen-activated protein kinases in Lycopersicon peruvianum suspension-cultured cells. Plant Physiol. 132, 1728–1738. doi: 10.1104/pp.103.024414
Huang, X., Yang, P., Ouyang, X., Chen, L., and Deng, X. (2014). Photoactivated UVR8-COP1 module determines photomorphogenic UV-B signaling output in Arabidopsis. PLoS Genet. 10:e1004218. doi: 10.1371/journal.pgen.1004218
Inzé, A., Vanderauwera, S., Hoeberichts, F. A., Vandorpe, M., Van Gaever, T., and Van Breugsegem, F. (2011). A subcellular localization compendium of hydrogen peroxide-induced proteins. Plant Cell Environ. 35, 308–320. doi: 10.1111/j.1365-3040.2011.02323.x
Jenkins, G. I. (2014). The UV-B photoreceptor UVR8, from structure to physiology. Plant Cell 26, 21–37. doi: 10.1105/tpc.113.119446
Jenkins, G. I. (2017). Photomorphogenic responses to ultraviolet-B light. Plant Sell Environ. 40, 2544–2557. doi: 10.1111/pce.12934
Jiang, J., Zhang, C., and Wang, X. (2015). A recently evolved isoform of the transcription factor BES1 promotes brassinosteroid signaling and development in Arabidopsis thaliana. Plant Cell 27, 361–374. doi: 10.1105/tpc.114.133678
Jiang, L., Wang, Y., Björn, L. O., and Li, S. (2009). Arabidopsis radical induced cell death1 is involved in UV-B signaling. Photochem. Photobiol. Sci. 8, 838–846. doi: 10.1039/b901187k
Jiang, L., Wang, Y., Li, Q., Bjorn, L., He, J., and Li, S. (2012a). Arabidopsis STO/BBX24 negatively regulates UV-B signaling by interacting with COP1 and repressing HY5 transcriptional activity. Cell Res. 22, 1046–1057. doi: 10.1038/cr.2012.34
Jiang, L., Wang, Y., Olof Björn, L., He, J. X., and Li, S. S. (2012b). Sensing of UV-B radiation by plants. Plant Signal. Behav. 7, 999–1003. doi: 10.4161/psb.20815
Jones, D. T., Taylor, W. R., and Thornton, J. M. (1992). The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 8, 275–282. doi: 10.1093/bioinformatics/8.3.275
Kaiserli, E., and Jenkins, G. I. (2007). UV-B promotes rapid nuclear translocation of the Arabidopsis UV-B specific signaling component UVR8 and activates its function in the nucleus. Plant Cell 19, 2662–2673. doi: 10.1105/tpc.107.053330
Kim, S. H., Kim, H., Chung, S., and Lee, J. H. (2017). DHU1 negatively regulates UV-B signaling via its direct interaction with COP1 and RUP1. Biochem. Biophys. Res. Commun. 491, 285–290. doi: 10.1016/j.bbrc.2017.07.110
Kliebenstein, D. J., Lim, J. E., Landry, L. G., and Last, R. L. (2002). Arabidopsis UVR8 regulates ultraviolet-B signal transduction and tolerance and contains sequence similarity to human Regulator of Chromatin Condensation 1. Plant Physiol. 130, 234–243. doi: 10.1104/pp.005041
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 35, 1547–9. doi: 10.1093/molbev/msy096
Lang-Mladek, C., Xie, L., Nigam, N., Chumak, N., Binkert, M., Neubert, S., et al. (2011). UV-B signaling pathways and fluence rate dependent transcriptional regulation of ARIADNE12. Physiol. Plantarum 145, 527–539. doi: 10.1111/j.1399-3054.2011.01561.x
Lau, O., and Deng, X. (2012). The photomorphogenic repressors COP1 and DET1, 20 years later. TIPS 17, 584–593. doi: 10.1016/j.tplants.2012.05.004
Li, C., Liu, S., Zhang, W., Chen, K., and Zhang, P. (2019). Transcriptional profiling and physiological analysis reveal the critical roles of ROS-scavenging system in the Antarctic moss Pohlia nutans under Ultraviolet-B radiation. Plant Physiol. Biochem. 134, 113–122. doi: 10.1016/j.plaphy.2018.10.034
Li, H., Li, Y., Deng, H., Sun, X., Wang, A., Tang, X., et al. (2018). Tomato UV-B receptor SlUVR8 mediates plant acclimation to UV-B radiation and enhances fruit chloroplast development via regulating SlGLK2. Sci. Rep. 8:6097. doi: 10.1016/s41598-018-24309-y
Li, J., He, Y. J., Zhou, L., Liu, Y., Jiang, M., Ren, L., et al. (2018). Transcriptome profiling of genes related to light-induced anthocyanin biosynthesis in eggplant (Solanum melongena L.) before purple color becomes evident. BMC Genomics 19:201. doi: 10.1186/s12864-018-4587-z
Li, J., Yang, L., Jin, D., Nezames, C. D., Terzaghi, W., and Deng, X. W. (2013). UV-B-induced photomorphogenesis in Arabidopsis. Protein Cell. 4, 485–492. doi: 10.1007/s13238-013-3036-7
Li, N., Teranishi, M., Yamaguchi, H., Matsushita, T., Watahiki, M., Tsuge, T., et al. (2015). UV-B-Induced CPD photolyase gene expression is regulated by UVR8-dependent and -independent pathways in arabidopsis. Plant Cell Physiol. 56, 2014–2023. doi: 10.1093/pcp/pcv121
Li, X., Ma, M., Shao, W., Wang, H., Fan, R., Chen, X., et al. (2018). Molecular cloning and functional analysis of a UV-B photoreceptor gene, BpUVR8 (UV resistance locus 8), from birch and its role in ABA response. Plant Sci. 274, 294-308. doi: 10.1016/j.plantsci.2018.06.006
Liang, T., Mei, S., Shi, C., Yang, Y., Peng, Y., Ma, L., et al. (2018). UVR8 interacts with BES1 and BIM1 to regulate transcription and photomorphogenesis in Arabidopsis. Dev. cell 44, 512–523. doi: 10.1016/j.devcel.2017.12.028
Liang, T., Yang, Y.u., and Liu, H. (2019). Signal transduction mediated by the plant UV-B photoreceptor UVR8. N. Phytol. 221, 1247–1252. doi: 10.1111/nph.15469
Liu, L., Gitz, D., and Mcclure, J. (1995). Effects of UV-B on flavonoids, ferulic acid, growth and photosynthesis in barley primary leaves. Physiol. Plantarum 93, 725–733. doi: 10.1111/j.1399-3054.1995.tb05123.x
Liu, L., Li, Y., She, G., Zhang, X., Jordan, B., Chen, Q., et al. (2018). Metabolite profiling and transcriptomic analyses reveal an essential role of UVR8-mediated signal transduction pathway in regulating flavonoid biosynthesis in tea plants (Camellia sinensis) in response to shading. BMC Plant Biol. 18:233. doi: 10.1186/s12870-018-1440-0
Liu, L., Scott, G., Winefield, C., and Jordan, B. (2015). From UVR8 to flavonol synthase: UV-B-induced gene expression in Sauvignon blanc grape berry. Plant Cell Environ. 38, 905–919. doi: 10.1111/pce.12349
Liu, X., Yue, M., Ji, Q., and He, J. (2013). Effects of ultraviolet-B radiation on primary photophysical process in photosystem II: a fluorescence spectrum analysis,” in Proceedings of the 15th International Conference on Photosynthesis. Photosynthesis Research for Food, Fuel and the Future (Berlin: Springer, 642–649.
Lois, R. (1994). Accumulation of UV-absorbing flavonoids induced by UV-B radiation in Arabidopsis thaliana L. Planta 194, 498–503. doi: 10.1007/BF00714462
Loyola, R., Herrera, D., Mas, A., Wong, D. C., Höll, J., Cavallini, E., et al. (2016). The photomorphogenic factors UV-B RECEPTOR 1, ELONGATED HYPOCOTYL 5, and HY5 HOMOLOGUE are part of the UV-B signalling pathway in grapevine and mediate flavonol accumulation in response to the environment. J. Exp. Bot. 67, 5429–5445. doi: 10.1093/jxb/erw307
Mao, K., Wang, L., Li, Y. Y., and Wu, R. L. (2015). Molecular cloning and functional analysis of UV RESISTANCE LOCUS 8 (PeUVR8) from Populus euphratica. PLoS ONE 10:e0132390. doi: 10.1371/journal.pone.0132390
Mariz-Ponte, N., Mendes, R. J., Sario, S., Ferreira de Oliveira, J. M. P., Melo, P., and Santos, C. (2018). Tomato plants use non-enzymatic antioxidant pathways to cope with moderate UV-A/B irradiation: a contribution to the use of UV-A/B in horticulture. J. Plant Physiol. 221, 32–42 doi: 10.1016/j.jplph.2017.11.013
Martínez-Lüscherab, J., Torres, N., Hilberta, G., Richard, T., Sánchez-Díaz, M., Delrot, S., et al. (2014). Ultraviolet-B radiation modifies the quantitative and qualitative profile of flavonoids and amino acids in grape berries. Phytochem. 102, 106–114. doi: 10.1016/j.phytochem.2014.03.014
Morales, L., Brosché, M., Vainonen, J., Jenkins, G., Wargent, J., Sipari, N., et al. (2013). Multiple roles for UV RESISTANCE LOCUS8 in regulating gene expression and metabolite accumulation in Arabidopsis under solar ultraviolet radiation. Plant Physiol. 161, 744–759. doi: 10.1104/pp.112.211375
Nguyen, C. T. T., Lim, S., Lee, J. G., and Lee, E. J. (2017). VcBBX, VcMYB21, and VcR2R3MYB transcription factors are involved in UV-B-induced anthocyanin biosynthesis in the peel of harvested blueberry fruit. J. Agric. Food Chem. 65, 2066–2073. doi: 10.1021/acs.jafc.6b05253
O'Hara, A., and Jenkins, G. I. (2012). In vivo function of tryptophans in the Arabidopsis UV-B photoreceptor UVR8. Plant Cell 24, 3755–3766. doi: 10.1105/tpc.112.101451
Olsen, J. L., Rouze, R., Verhelst, B., Lin, Y. C., Bayer, T., Collen, J., et al. (2016). The genome of the seagrass Zostera marina reveals angiosperm adaptation to the sea. Nature 530, 331–335. doi: 10.1038/nature16548
Panchy, N., Lehti-Shiu, M., and Shiu, S. H. (2016). Evolution of gene duplication in plants. Plant Physiol. 171, 2294–2316. doi: 10.1104/pp.16.00523
Pang, Q., and Hays, J. B. (1991). UVB-Inducible and temperature-sensitive photoreactivation of cyclobutane pyrimidine dimers in Arabidopsis thaliana. Plant Physiol. 95, 536–543. doi: 10.1104/pp.95.2.536
Puranik, S., Jha, S., Srivastava, P., Sreenivasulu, N., and Prasad, M. (2011). Comparative transcriptome analysis of contrasting foxtail millet cultivars in response to short-term salinity stress. J. Plant Physiol. 168, 280–287. doi: 10.1016/j.jplph.2010.07.005
Qian, C., Mao, W., Liu, Y., Ren, H., Lau, O. S., Ouyang, X., et al. (2016). Dual-source nuclear monomers of UV-B light receptor direct photomorphogenesis in Arabidopsis. Mol. plant 9, 1671–1674. doi: 10.1016/j.molp.2016.10.005
Rajhi, I., Yamauchi, T., Takahashi, H., Nishiuchi, S., Shiono, K., Watanabe, R., et al. (2011). Identification of genes expressed in maize root cortical cells during lysigenous aerenchyma formation using laser microdissection and microarray analyses. N. Phytol. 190, 351–368. doi: 10.1111/j.1469-8137.2010.03535.x
Reddy, K. R., Patro, H., Lokhande, S., Bellaloui, N., and Gao, W. (2016). Ultraviolet-B radiation alters soybean growth and seed quality. Food Nutr. Sci. 7, 55–66. doi: 10.4236/fns.2016.71007
Rizzini, L., Favory, J. J., Cloix, C., Faggionato, D., O'Hara, A., Kaiserli, E., et al. (2011). Perception of UV-B by the Arabidopsis UVR8 protein. Science 332, 103–106. doi: 10.1126/science.1200660
Santhanam, R., Oh, Y., Kumar, R., Weinhold, A., Luu, V. T., Groten, K., et al. (2017). Specificity of root microbiomes in native-grown Nicotiana attenuata and plant responses to UVB increase Deinococcus colonization. Mol. Ecol. 26, 2543–2562. doi: 10.1111/mec.14049
Santin, M., Lucini, L., Castagna, A., Rocchetti, G., Hauser, M. T., and Ranieri, A. (2019). Comparative “phenol-omics” and gene expression analyses in peach (Prunus persica) skin in response to different postharvest UV-B treatments. Plant Physiol. Biochem. 135, 511–519. doi: 10.1016/j.plaphy.2018.11.009
Schreiner, M., Mewis, I., Huyskens-Keil, S., Jansen, M., Zrenner, R., Winkler, J., et al. (2012). UV-b-induced secondary plant metabolites - potential benefits for plant and human health. Crit. Rev. Plant Sci. 31, 229–240. doi: 10.1080/07352689.2012.664979
Searles, P., Flint, S., and Caldwell, M. (2001). A meta analysis of plant field studies simulating stratospheric ozone depletion. Oecologia 127, 1–10. doi: 10.1007/s004420000592
Soltis, P., Marchant, D., Van de Peer, Y., and Soltis, D. (2015). Polyploidy and genome evolution in plants. Curr Opin Genet Dev. 35, 119–125. doi: 10.1016/j.gde.2015.11.003
Soriano, G., Cloix, C., Heilmann, M., Núñez-Olivera, E., Martínez-Abaigar, J., and Jenkins, G. I. (2018). Evolutionary conservation of structure and function of the UVR8 photoreceptor from the liverwort Marchantia polymorpha and the moss Physcomitrella patens. N. Phytol. 161, 151–162. doi: 10.1111/nph.14767
Steinbrecht, W., Hegglin, M. I., Harris, N., and Weber, M. (2018). Is global ozone recovering? C. R. Geoscience 350, 368–375. doi: 10.1016/j.crte.2018.07.012
te Beest, M., Le Roux, J. J., Richardson, D. M., Brysting, A. K., Suda, J., Kubešová, M., et al. (2011). The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot. 109, 19–45. doi: 10.1093/aob/mcr277
Teramura, A. H., and Sullivan, J. H. (1994). Effects of UVB radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 39, 463–473. doi: 10.1007/BF00014599
Tevini, M., and Steinmüller, D. (1987). Influence of light, UVB radiation, and herbicides on wax biosynthesis of cucumber seedling. J. Plant Physiol. 131, 111–121. doi: 10.1016/S0176-1617(87)80272-9
Tevini, M., and Teramura, A. H. (1989). UVB effects on terrestrial plants. Photochem. Photobiol. 50, 479–487. doi: 10.1111/j.1751-1097
Tilbrook, K., Arongaus, A., Binkert, M., Heijde, M., Yin, R., and Ulm, R. (2013). The UVR8 UV-B photoreceptor: perception, signaling and response. Arabidopsis Book 11:e0164. doi: 10.1199/tab.0164
Tilbrook, K., Dubois, M., Crocco, C. D., Yin, R., Chappuis, R., Allorent, G., et al. (2016). UV-B perception and acclimation in Chlamydomonas reinhardtii. Plant Cell 28, 966–983. doi: 10.1105/tpc.15.00287
Tohge, T., Kusano, M., Fukushima, A., Saito, K., and Fernie, A. R. (2011). Transcriptional and metabolic programs following exposure of plants to UV-B irradiation. Plant Signal. Behav. 6, 1987–1992. doi: 10.4161/psb.6.12.18240
Tossi, V., Cassia, R., Bruzzone, S., Zocchi, E., and Lamattina, L. (2012). ABA says NO to UV-B: a universal response? TIPS 17, 510–517. doi: 10.1016/j.tplants.2012.05.007
Tossi, V., Lamattina, L., and Cassia, R. (2009). An increase in the concentration of abscisic acid is critical for nitric oxide-mediated plant adaptive responses to UV-B irradiation. N. Phytol. 181, 871–879. doi: 10.1111/j.1469-8137.2008.02722.x
Tossi, V., Lamattina, L., Jenkins, G. I., and Cassia, R. O. (2014). Ultraviolet-B-induced stomatal closure in Arabidopsis is regulated by the UV RESISTANCE LOCUS8 photoreceptor in a nitric oxide-dependent mechanism. Plant Physiol. 164, 2220–2230. doi: 10.1104/pp.113.231753
Tripathi, R., and Agrawal, S. B. (2016). Effect of supplemental UV-B on yield, seed quality, oil content and fatty acid composition of Brassica campestris L. under natural field conditions. Qual. Assur. Saf. Crops Foods 8, 11–20. doi: 10.3920/QAS2013.0290
Trivedi, D. K., Gill, S. S., and Tuteja, N. (2016). “Abscisic acid (ABA): biosynthesis, regulation, and role in abiotic stress tolerance”, in Abiotic Stress Response in Plants, eds N. Tuteja and S. S. Gill (Weinheim: Wiley-VCH Verlag GmbH & Co.), 311–322. doi: 10.1002/9783527694570.ch15
Turtola, S., Sallas, L., Holopainen, J. K., Julkunen-Tiitto, R., and Kainulainen, P. (2006). Long-term exposure to enhanced UV-B radiation has no significant effects on growth or secondary compounds of outdoor-grown Scots pine and Norway spruce seedlings. Environ. pollut. 144, 166–171. doi: 10.1016/j.envpol.2005.12.025
Tuteja, N. (2007). Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2, 135–138. doi: 10.4161/psb.2.3.4156
Ubi, B. E., Honda, C., Bessho, H., Kondo, S., Wada, M., Kobayashi, S., et al. (2006). Expression analysis of anthocyanin biosynthetic genes in apple skin: effect of UV-B and temperature. Plant Sci. 170, 571–578. doi: 10.1016/j.plantsci.2005.10.009
Ulm, R., and Jenkins, G. I. (2015). Q&A: how do plants sense and respond to UV-B radiation? BMC Biol. 13:45. doi: 10.1186/s12915-015-0156-y
Van Nocker, S., and Ludwig, P. (2003). The WD-repeat protein superfamily in Arabidopsis: conservation and divergence in structure and function. BMC Genomics 4:50. doi: 10.1186/1471-2164-4-50
Vandenbussche, F., Yu, N., Li, W., Vanhaelewyn, L., Hamshou, M., Van Der Straeten, D., et al. (2018). An ultraviolet B condition that affects growth and defense in Arabidopsis. Plant Sci. 268, 54–63. doi: 10.1016/j.plantsci.2017.12.005
Vanhaelewyn, L., Schumacher, P., Poelman, D., Fankhauser, C., Van Der Straeten, D., and Vandenbussche, F. (2016). REPRESSOR OF ULTRAVIOLET-B PHOTOMORPHOGENESIS functionallows efficient phototropin mediated ultraviolet-B phototropism in etiolated seedlings. Plant Sci. 252, 215–221. doi: 10.1016/j.plantsci.2016.07.008
Vert, G., and Chory, J. (2006). Downstream nuclear events in brassinosteroid signalling. Nature 441, 96–100. doi: 10.1038/nature04681
Wan, J., Zhang, P., Wang, R., Sun, L., Wang, W., Zhou, H., et al. (2018). UV-B radiation induces root bending through the flavonoid-mediated auxin pathway in Arabidopsis. Front. Plant Sci. 9:618. doi: 10.3389/fpls.2018.00618
Wang, H., Gui, M., Tian, X., Xin, X., Wang, T., and Li, J. (2017). Effects of UV-B on vitamin C, phenolics, flavonoids and their related enzyme activities in mung bean sprouts (Vigna radiata). Int. J. Food Sci. Technol. 52, 827–833. doi: 10.1111/ijfs.13345
Wang, S., Chen, C., Sciarappa, W., Wang, C., and Camp, M. (2008). Fruit quality, antioxidant capacity, and flavonoid content of organically and conventionally grown blueberries. J. Agric. Food Chem. 56, 5788–5794. doi: 10.1021/jf703775r
Wargent, J., Gegas, V., Jenkins, G., Doonan, J., and Paul, N. (2009). UVR8 in Arabidopsis thaliana regulates multiple aspects of cellular differentiation during leaf development in response to ultraviolet B radiation. N. Phytol. 183, 315–326. doi: 10.1111/j.1469-8137.2009.02855.x
Waters, M., Moylan, E., and Langdale, J. (2008). GLK transcription factors regulate chloroplast development in a cell autonomous manner. Plant Cell 56, 432–444. doi: 10.1111/j.1365-313X.2008.03616.x
Weber, M., Coldewey-Egbers, M., Fioletov, V. E., Frith, S. M., Wild, J. D., Burrows, J. P., et al. (2018). Total ozone trends from 1979 to 2016 derived from five merged observational datasets – the emergence into ozone recovery. Atmos. Chem. Phys. 18, 2097–2117. doi: 10.5194/acp-18-2097-2018
Wu, D., Hu, Q., Yan, Z., Chen, W., Yan, C. Y., Huang, X., et al. (2012). Structural basis of ultraviolet-B perception by UVR8. Nature 484, 214–219. doi: 10.1038/nature10931
Wu, G., Bornman, J. F., Bennett, S. J., Clarke, M. W., Fang, Z., and Johnson, S. K. (2017). Individual polyphenolic profiles and antioxidant activity in sorghum grains are influenced by very low and high solar UV radiation and genotype. J. Cereal Sci. 77, 17–23. doi: 10.1016/j.jcs.2017.07.014
Wu, Q., Su, N., Zhang, X., Liu, Y., Cui, J., and Liang, Y. (2016). Hydrogen peroxide, nitric oxide and UV RESISTANCE LOCUS8 interact to mediate UV-B-induced anthocyanin biosynthesis in radish sprouts. Sci. Rep. 6:29164. doi: 10.1038/srep29164
Yang, Y., Liang, T., Zhang, L., Shao, K., Gu, X., Shang, R., et al. (2018a). UVR8 interacts with WRKY36 to regulate HY5 transcription and hypocotyl elongation in Arabidopsis. Nat. Plants 4, 98–107. doi: 10.1038/s41477-017-0099-0
Yang, Y., Yang, X., Jang, Z., Chen, Z., Ruo, X., Jin, W., et al. (2018b). UV RESISTANCE LOCUS 8 From Chrysanthemum morifolium Ramat (CmUVR8) plays important roles in UV-B signal transduction and UV-B-induced accumulation of flavonoids. Front. Plant Sci. 9:955. doi: 10.3389/fpls.2018.00955
Yin, R., Arongaus, A. B., Binkert, M., and Ulm, R. (2015). Two distinct domains of the UVR8 photoreceptor interact with COP1 to initiate UV-B signaling in Arabidopsis. Plant Cell 27, 202–213. doi: 10.1105/tpc.114.133868
Yin, R., Skvortsova, M. Y., Loubéry, S., and Ulm, R. (2016). COP1 is required for UV-B–induced nuclear accumulation of the UVR8 photoreceptor. Proc. Natl. Acad. Sci. U.S.A. 113, E4415–E4422. doi: 10.1073/pnas.1607074113
Yin, Y., Vafeados, D., Tao, Y., Yoshida, S., Asami, T., and Chory, J. (2005). A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell 120, 249–259. doi: 10.1016/j.cell.2004.11.044
Zellmer, I. (1995). “UV-B-tolerance of alpine and arctic Daphnia,” in Cladocera as Model Organisms in Biology, eds P. Larsson and L. J. Weider (Dordrecht: Springer, 153–159.
Zhang, H. N., Li, W. C., Wang, H. C., Shi, S. Y., Shu, B., Liu, L. Q., et al. (2016). Transcriptome profiling of light-regulated anthocyanin biosynthesis in the pericarp of litchi. Front. Plant Sci. 7:963. doi: 10.3389/fpls.2016.00963
Zhao, C., Mao, K., You, C. X., Zhao, X. Y., Wang, S. H., Li, Y. Y., et al. (2016). Molecular cloning and functional analysis of a UV-B photoreceptor gene, MdUVR8 (UV Resistance Locus 8), from apple. Plant Sci. 247, 115–126. doi: 10.1016/j.plantsci.2016.03.006
Keywords: UV-B, UVR8, RUP1, RUP2, Arabidopsis, terrestrial plants, photomorphogenic response, gene expression
Citation: Tossi VE, Regalado JJ, Iannicelli J, Laino LE, Burrieza HP, Escandón AS and Pitta-Álvarez SI (2019) Beyond Arabidopsis: Differential UV-B Response Mediated by UVR8 in Diverse Species. Front. Plant Sci. 10:780. doi: 10.3389/fpls.2019.00780
Received: 09 March 2019; Accepted: 28 May 2019;
Published: 18 June 2019.
Edited by:
Antonella Castagna, University of Pisa, ItalyCopyright © 2019 Tossi, Regalado, Iannicelli, Laino, Burrieza, Escandón and Pitta-Álvarez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandra Irene Pitta-Álvarez, c2FuZHJhcGl0dGEtYWx2YXJlekBjb25pY2V0Lmdvdi5hcg==; c3BpdHRhMTk1OUBnbWFpbC5jb20=
†These authors have contributed equally to this work