 Kristina Jaškūnė1*
Kristina Jaškūnė1* Andrius Aleliūnas1
Andrius Aleliūnas1 Gražina Statkevičiūtė1
Gražina Statkevičiūtė1 Vilma Kemešytė2
Vilma Kemešytė2 Bruno Studer3
Bruno Studer3 Steven Yates3*
Steven Yates3*- 1Laboratory of Genetics and Physiology, Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, Akademija, Lithuania
- 2Department of Grass Breeding, Institute of Agriculture, Lithuanian Research Centre for Agriculture and Forestry, Akademija, Lithuania
- 3Department of Environmental Systems Science, Molecular Plant Breeding, Institute of Agricultural Sciences, ETH Zurich, Zurich, Switzerland
Global warming is predicted to impact many agricultural areas, which will suffer from reduced water availability. Due to precipitation changes, mild summer droughts are expected to become more frequent, even in temperate regions. For perennial ryegrass (Lolium perenne L.), an important forage grass of the Poaceae family, leaf growth is a crucial factor determining biomass accumulation and hence forage yield. Although leaf elongation has been shown to be temperature-dependent under normal conditions, the genetic regulation of leaf growth under water deficit in perennial ryegrass is poorly understood. Herein, we evaluated the response to water deprivation in a diverse panel of perennial ryegrass genotypes, employing a high-precision phenotyping platform. The study revealed phenotypic variation for growth-related traits and significant (P < 0.05) differences in leaf growth under normal conditions within the subgroups of turf and forage type cultivars. The phenotypic data was combined with genotypic variants identified using genotyping-by-sequencing to conduct a genome-wide association study (GWAS). Using GWAS, we identified DNA polymorphisms significantly associated with leaf growth reduction under water deprivation. These polymorphisms were adjacent to genes predicted to encode for phytochrome B and a MYB41 transcription factor. The result obtained in the present study will increase our understanding on the complex molecular mechanisms involved in plant growth under water deficit. Moreover, the single nucleotide polymorphism (SNP) markers identified will serve as a valuable resource in future breeding programs to select for enhanced biomass formation under mild summer drought conditions.
Introduction
Abiotic stress has detrimental effects on agriculture worldwide, threatening food security as huge yield losses are already being reported (Daryanto et al., 2016; Fahad et al., 2017). Moreover, concerns are rising about the effect climate shifts may impose on grassland productivity, and consequently feed production (Ergon et al., 2018). Climate models predict an increase in dry spells in the future, the key components being water deficit caused by the changes in precipitation patterns and temperature increase (Boyer, 1982; Lobell and Field, 2007; Salekdeh et al., 2009; Moore and Lobell, 2015).
Drought is a major limitation to crop productivity, crop irrigation consumes large amounts of water resources (Condon et al., 2004). Plant breeding for drought tolerant cultivars is considered as an important measures to mitigate the effects of drought on food and feed production, however, direct selection for yield under water stress resulted in limited genetic gain for drought tolerance (Hu and Xiong, 2014; Kumar et al., 2014; Mwadzingeni et al., 2016; Sandhu and Kumar, 2017). Plant phenotypes are inherently complex, reflecting the interactions of the genotype with environmental factors (Forsman, 2014). These interactions influence plant growth and development, which affect biomass accumulation and hence yield formation. A very important aspect to consider when breeding for drought tolerant crops is the severity and duration of the stress, as increased survival under severe drought does not mean improved performance under mild drought (Skirycz et al., 2011). When faced with water limitation during summer droughts in temperate environments, plants might employ alternative strategies, like drought escape, and drought avoidance (Kooyers, 2015). In either case, the outcome is reduced vegetative growth and biomass accumulation (Farooq et al., 2012). Plants can adapt to water stress in different ways, they might choose between survival, meaning reduced water use and growth halt, or continued slow growth (Claeys and Inzé, 2013). The latter option offers a competitive advantage in biomass accumulation, assuming the stress is temporary but can threaten survival if the stress persists. From a practical point of view, aiming to obtain stable yield under mild drought, a forage grass cultivar continuing slower growth after sensing the water deprivation is preferable to the one that opts for the survival strategy and halts its growth early.
Throughout the decades of perennial ryegrass (Lolium perenne L.) breeding, progress has been made in many quality-related traits (Humphreys et al., 2010; Sampoux et al., 2011, 2013). Drought adaptation of cultivars and natural ecotypes, reflected both in aboveground biomass production under field conditions and controlled environments, has been the focus of many studies (Korte and Chu, 1983; Kemesyte et al., 2017; Bothe et al., 2018; Lee et al., 2019). However, most of the earlier studies used destructive phenotyping methods or were limited to measuring the traits at start and end of the stress. This might lead to missing important quantitative trait loci (QTL) in association studies (Bac-Molenaar et al., 2016) as well as limited understanding of the overall complexity of temporal plant response to the environment. Recently, Yates et al. (2019) described a dynamic phenotyping method as a non-destructive and precise diagnostic tool to profile leaf growth under water deficit stress in situ. This method allows modeling leaf growth and pinpointing when a plant reduces and arrests leaf growth due to water deficit and thus has the potential to unravel the genetic mechanisms underlying these responses by QTL analysis. In this study, we aimed at: (i) determining the genotypic response of leaf growth to temperature and soil water availability, (ii) identifying genome-wide DNA sequence polymorphisms using genotyping-by-sequencing (GBS) for population description, and (iii) g genetic loci associated with the response to water deficit stress by performing genome-wide association study (GWAS).
Materials and Methods
Plant Material and Growth Conditions
A perennial ryegrass association panel consisting of 197 genotypes was used throughout the water deficit experiments. The panel was chosen on the basis of earlier experiments for drought (Jonavičienė et al., 2014) and freezing tolerance (Aleliūnas et al., 2015), where results revealed a high genotypic diversity among the panel individuals as well as a high phenotypic variability for drought-related traits. Most of the cultivars were of European origin, except one genotype coming from Japan, one genotype from New Zealand and six genotypes from the United States. The majority of ecotypes were of Lithuanian and Ukrainian origin, 35 and 55, respectively, but a few of the ecotypes were from Latvia (two), Poland (two), Slovakia (two), Denmark (one), and the Kaliningrad Region of Russian Federation (ten). A detailed description of the association mapping panel is presented in Supplementary Tables 1, 2.
For the leaf growth under water deficit experiment, plants were vegetatively propagated into four clonal replicates, each consisting of 20 tillers, and grown in plastic pots (ø15 cm and 12 cm height) filled with 450 g of commercial potting mix substrate (“Spezialmischung 209,” RICOTER Erdaufbereitung AG, Aarberg, Switzerland) under regular irrigation and fertilization in a greenhouse. 4 to 6 weeks after clonal propagation, plants were transferred into a climate chamber (Conviron, 86 Winnipeg, Canada) under controlled conditions with a light/dark photoperiod of 16/8 h and a light intensity of 275 μmol photosynthetically active radiation (PAR) m–2 s–1. The climate chamber was equipped with a 2:1 mixture of fluorescent lamps of two types (T5 FQ 54W/840 HO, Osram GmbH, Munich, Germany and T5 FH054W/GRO G5 F 54W, Havells Sylvania Europe Ltd, London, United Kingdom). The day/night temperature was 25/15°C, and relative air humidity was set to 50%.
Water Deficit Treatment and Leaf Growth Measurements
To measure the phenotypic response to water deficit, leaf growth was assessed non-destructively using a largely automated phenotyping platform as described by Yates et al. (2019). Briefly, the tip of a young growing leaf was attached with a hair pin to a string and kept taut using weights of 20 g. White plastic beads (ø20 mm, 7 g) were threaded onto the strings and placed on the growth array to provide artificial landmarks for image-based marker tracking. Images of the growth array were taken every 2 min with a LupusNET HD camera (LUPUS-Electronics® Gmbh, Landau, Germany) and analyzed with the LLT software (Nagelmüller et al., 2016).
To induce a water deficit stress, perennial ryegrass plants were deprived of water for 130 h. After the stress treatment, plants were re-watered and grown for additional 35 h to confirm that water limitation was the cause of leaf growth arrest, but not leaf age. Soil moisture deficit was measured using an integrated wireless microclimate sensing system (WiFi Plant sensor, Koubachi, Switzerland). Each sensor was calibrated individually and soil moisture data was measured at a depth of 7 cm every 4 h. The meristem temperatures of six plants per experiment were measured with a K type thermocouple (GTF 300, Greisinger, Germany; ø 0.2 mm) inserted into the tiller at meristem height.
The statistical analysis was implemented with the open source R statistical environment (version 3.1.0; R Development Core Team, 2005). Leaf growth under water limitation was estimated using the TriPhase function described by Yates et al. (2019). The function dissects complex growth processes into qualitative traits, described as leaf thermal growth (a), leaf growth reduction point (Σ), and the leaf growth arrest point (σ). The average phenotype for each trait per genotype was determined using least-square means implemented in “lsmeans” package for R (Lenth, 2016). Student’s t-test was used to compare the means of different growth traits and the Pearson’s correlation coefficients between different phenotypes were estimated. Broad sense heritability of the traits was determined using the “heritability” package of R (Kruijer et al., 2014).
Genotyping-by-Sequencing and SNP Discovery
High-molecular-weight DNA was extracted from fresh leaf material using the method described by Mayjonade et al. (2016). The DNA was quality checked on a 1% agarose gel and quantified with a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, United States). The association panel was divided into three groups with equal number of individuals in each to produce GBS libraries. One hundred ng of DNA of each individual was digested by PstI (Thermo Fisher Scientific, Waltham, MA, United States) and then unique barcodes were ligated using T4 DNA ligase (Thermo Fisher Scientific, Waltham, MA, United States). The libraries were then pooled into a single tube and PCR-amplified using a common primer with a HiSeq adapter. After quality checking and cleaning using Qiagen spin columns (QIAGEN Sciences Inc, United States), the samples were sequenced by 126 bp single-end reads on a Illumina HiSeq 2500 (Illumina, Inc, San Diego, United States) sequencing system at the Functional Genomics Center Zurich (Zurich, Switzerland).
The quality of raw sequence data was checked using FastQC (v0.11, DePristo et al., 2011). After the initial quality check, reads of 164 genotypes were demultiplexed into separate files using saber1 with no mismatches allowed. Illumina adapter sequences were removed using Cutadapt (v1.18; Martin, 2011) and reads shorter than 40 bp were discarded. The reads were mapped to the perennial ryegrass draft genome assembly (Byrne et al., 2015) using BWA-MEM (Li, 2013). Bi-allelic variants were called using Genome Analysis Toolkits (GATK, v.3.8-1-0, McKenna et al., 2010) and then hard-filtered using vcftools (v.0.1.14, Auton et al., 2011) with minimum depth of 5 and minimum quality (GQ) score of 40. After removing single nucleotide polymorphisms (SNPs) with low-quality scores, variant sites were additionally filtered on allele frequencies by excluding variants with a minor allele frequency (MAF) below 0.05. Afterward, variant sites with more than 50% of missing data in the population were excluded.
Linkage Disequilibrium, Population Structure and GWAS
Linkage disequilibrium (LD) in the association panel was expressed as squared Pearson’s correlation coefficient (r2) on all the marker-tagged scaffolds larger than 20 kbp. For LD analysis, SNPs with an estimated MAF lower than 0.05 were removed. Squared correlation coefficient between the variant sites was evaluated using vcftools (v0.1.16), while the subsequent analysis was performed using R.
Population structure of the perennial ryegrass association panel was investigated using principal component analysis (PCA). PCA was conducted using “prcomp” function implemented in stats package for R. For PCA, genotypic data matrix was used with missing data replaced using mean imputation.
Marker-trait associations were tested using the mean phenotypic values. A GWAS was performed for each trait separately using a single marker regression (SMR, Lenth, 2016) and three complementing methods: (1) FarmCPU (Fixed and random model Circulating Probability Unification, Liu et al., 2016) implemented in GAPIT software package2, (2) multi-locus mixed model (MLMM, Segura et al., 2012), and (3) Bayesian-information and Linkage-disequilibrium Iteratively Nested Keyway (BLINK, Zhang et al., 2018) for R. The first three principal components (PC) were provided as covariate variables. For the SMR analysis, each SNP marker was tested with the phenotype as the response variable and the genotype scores as predictor variable. SNPs were retained when more than 100 data points for both the phenotype and the genotype were available. Alleles were removed if their MAF was less than 0.05 If a SNP met these criteria and was still polymorphic, a regression model using the “lm” function in R was fitted and the significance (P-value) and the variance explained (R2) was extracted.
To correct for multiple testing, the P-values were adjusted using both FDR (Benjamini and Hochberg, 1995) and Bonferroni corrections. An annotation file from the Perennial Ryegrass Genome Sequencing Project (available online at: http://185.45.23.197:5080/ryegrassdata/GENE_ANNOTATIONS/Lp_Annotation_V1.1.mp.gff3) was used to retrieve predicted gene positions and sequence information.
To identify the putative location of markers on chromosome level, scaffolds harboring significant marker-trait associations were anchored to the pseudomolecules of Hordeum vulgare genome (Mascher et al., 2017) using LAST3.
Results
Phenotyping of Leaf Growth Under Water Deficit Conditions
Substantial variation for leaf growth under water deficit was detected within the perennial ryegrass panel (Figure 1). In the whole association panel, the values for leaf thermal growth rate (a) ranged from 0.001 to 0.147 mm.h–1.°C–1 with a mean a value of 0.06 mm.h–1.°C–1. Growth reduction point (Σ) varied between 1.368–4.432 log10 (hPa) with a mean Σ value of 2.38 log10 (hPa). The growth arrest point (σ) was lower and varied between 1.87–5.94 log10 (hPa) with a mean σ value of 3.89 log10 (hPa). The comparison between ecotype and cultivar groups in the association panel revealed no significant differences (P > 0.05) for all assessed traits, but significant differences were found when comparing the subgroups. Under optimal conditions, the averaged thermal growth rate of forage type cultivars was a = 0.07 mm.h–1.°C–1, while the turf type cultivars grow significantly (P < 0.01) slower with a = 0.05 mm.h–1.°C–1. A significant difference was also observed between the turf subgroup and both ecotype groups originating from maritime and continental climate zones (P < 0.05). Overall, the heritability values for the measured traits were high: leaf thermal growth rate (a) h2 = 0.67, growth reduction point (Σ) h2 = 0.72, and growth arrest point (σ) h2 = 0.71.
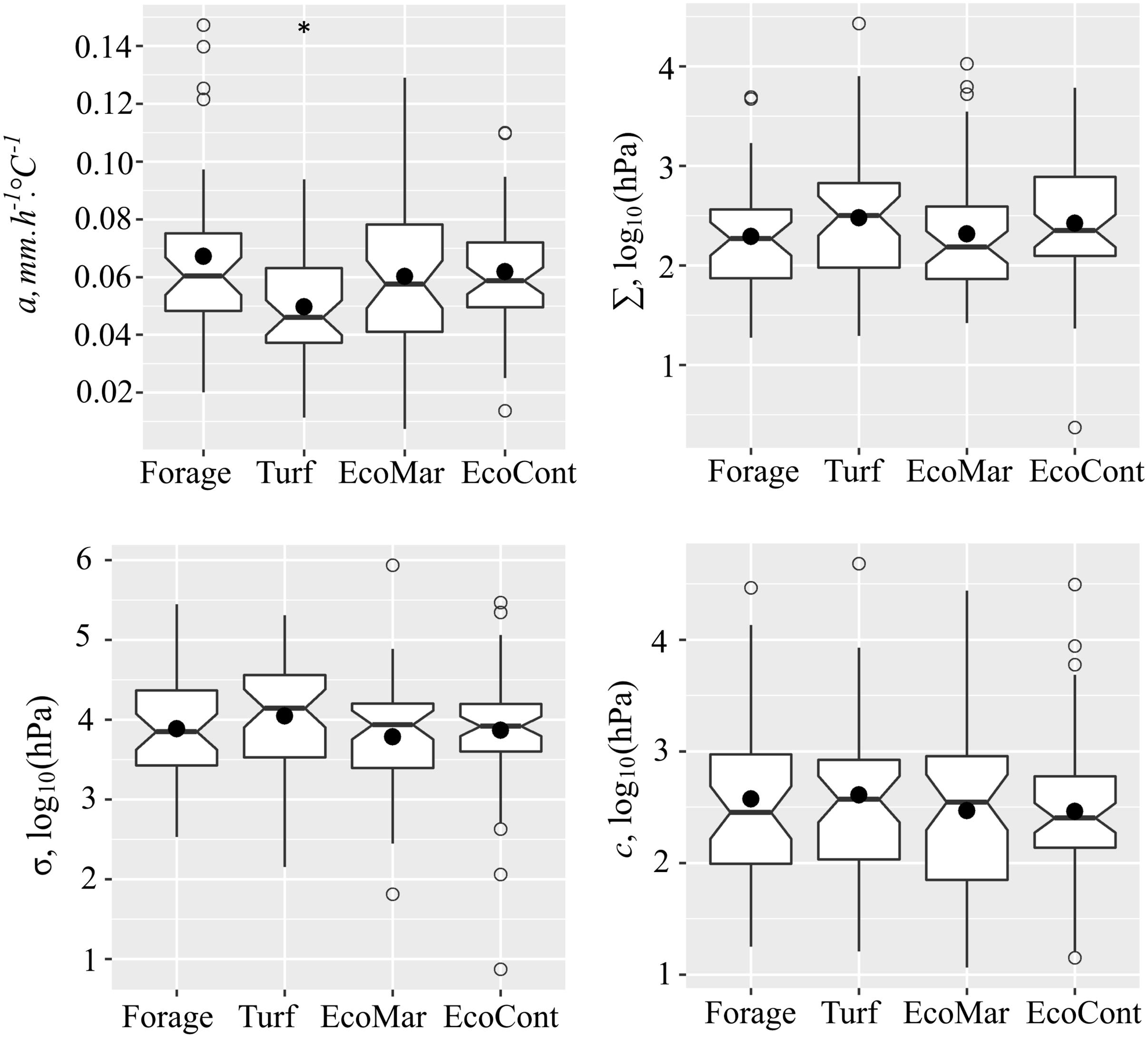
Figure 1. Variation of thermal leaf growth (a), leaf growth reduction point (Σ), leaf growth arrest point (σ), and water deprivation tolerance (c) between the forage cultivar (n = 43), turf cultivar (n = 46), maritime origin ecotype (EcoMar, n = 50) and continental origin ecotype (EcoCont, n = 58) groups in the perennial ryegrass panel. *indicates significant difference at P < 0.05 (Student’s t-test).
Variable correlations between the thermal growth, growth reduction point (Σ), and the growth arrest point (σ) were obtained in the subgroups (Figure 2). Strong to moderate positive and significant correlation was observed between growth reduction and growth arrest points in both ecotype groups and turf type cultivar group, ranging from r = 0.62 to r = 0.46, P < 0.05. Leaf growth arrest point strongly correlated with tolerance of genotypes in all groups (r ranging from 0.50 to 0.61, P < 0.05).
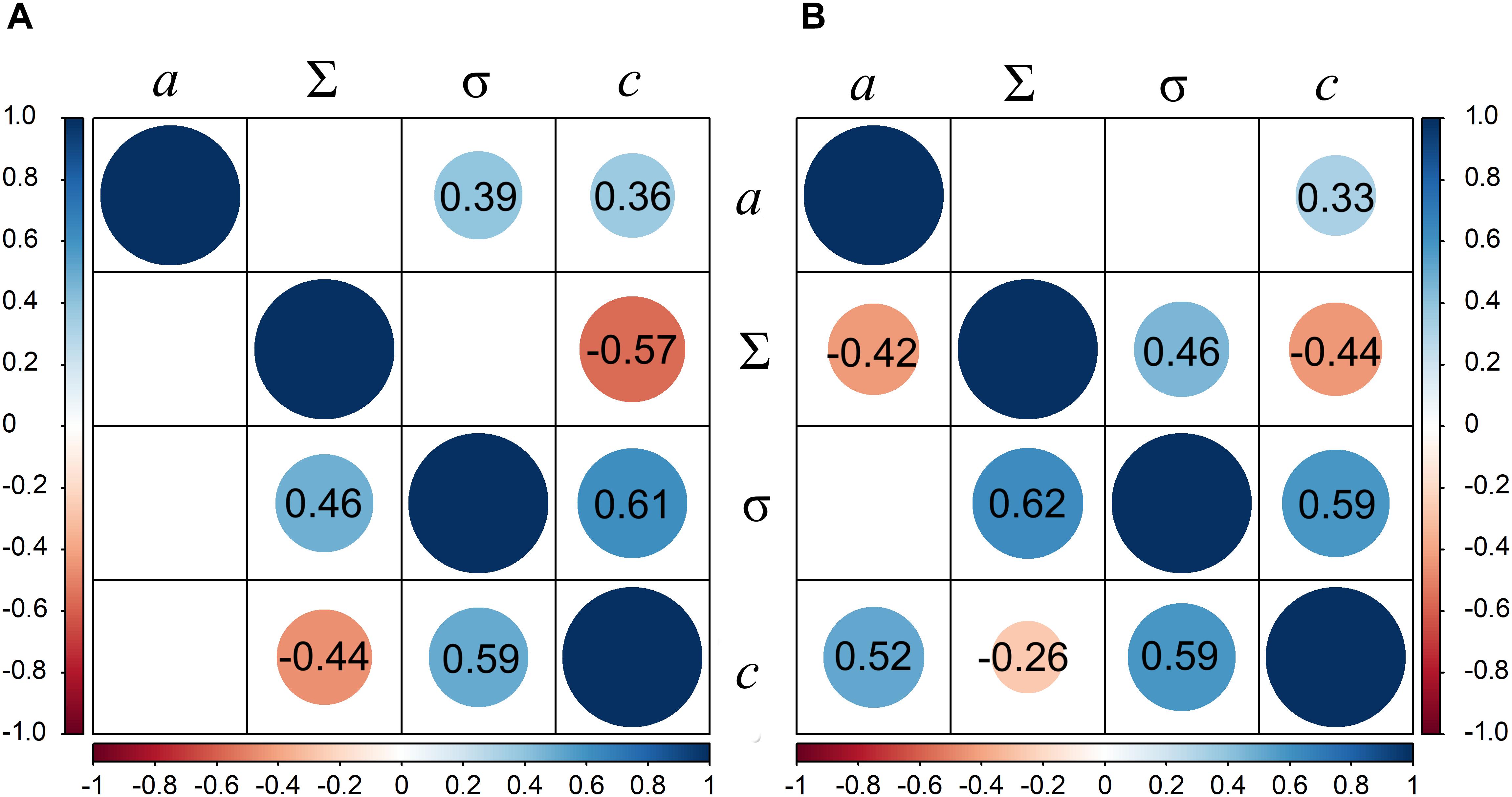
Figure 2. Correlation between thermal leaf growth (a), the leaf growth reduction point (Σ), the leaf growth arrest point (σ), and water deprivation tolerance (c) traits in perennial ryegrass panel; In (A) upper triangle represents forage cultivar group and lower triangle represents turf. (B) Upper triangle represents group of ecotypes of maritime origin and lower triangle represents group of ecotypes of continental origin.
GBS, Population Structure and LD
Sequencing of the GBS libraries yielded over 573 million reads. The number of reads per sample ranged from 257,518 to 3,667,767 reads with an average reads number of 1,528,212 ± 604,999. After removing variant sites with missing rates ≥0.5 and minor allele frequency <0.05, 21,648 markers were identified and used for further analyses.
A rapid LD decay was observed in the perennial ryegrass association panel using GBS-PstI markers. LD was measured by the r2 and the average distance between the SNPs with LD ≥ 0.5 was determined to be 712 bp. The distance for SNPs with LD ≥ 0.25 was on average 1,378 bp.
Genetic structure in perennial ryegrass association panel was visualized using PCA (Figure 3). The first and second PC accounted from 3.0 to 4.0% and from 1.9 to 2.4% of the observed genetic variance, respectively. The first PC (Figure 3A) did not discriminate between ecotypes and cultivars, but the second PC shown some separation. The subgroups of forage and turf type cultivars were not clearly separated (Figure 3B), while for the ecotype group, a clear separation of the genotypes was observed regarding the country of origin (Figure 3C). Moreover, a high correlation was detected between PC1 and geographical latitude (r = 0.87, P < 0.01). Genotypes for the neighboring countries of Lithuania, Latvia and Kaliningrad region of Russia formed a distinct cluster, while Ukrainian, Polish and Slovakian ecotypes formed a separate cluster. Interestingly, two Ukrainian ecotypes were found to be distantly related to other genotypes with an obvious separation on PC2 (Figure 3C). PC1, in Figure 3C, clearly separated ecotypes originating from the Baltic Sea region from the ecotype group of Ukrainian and Slovakian which represented continental climate zone.
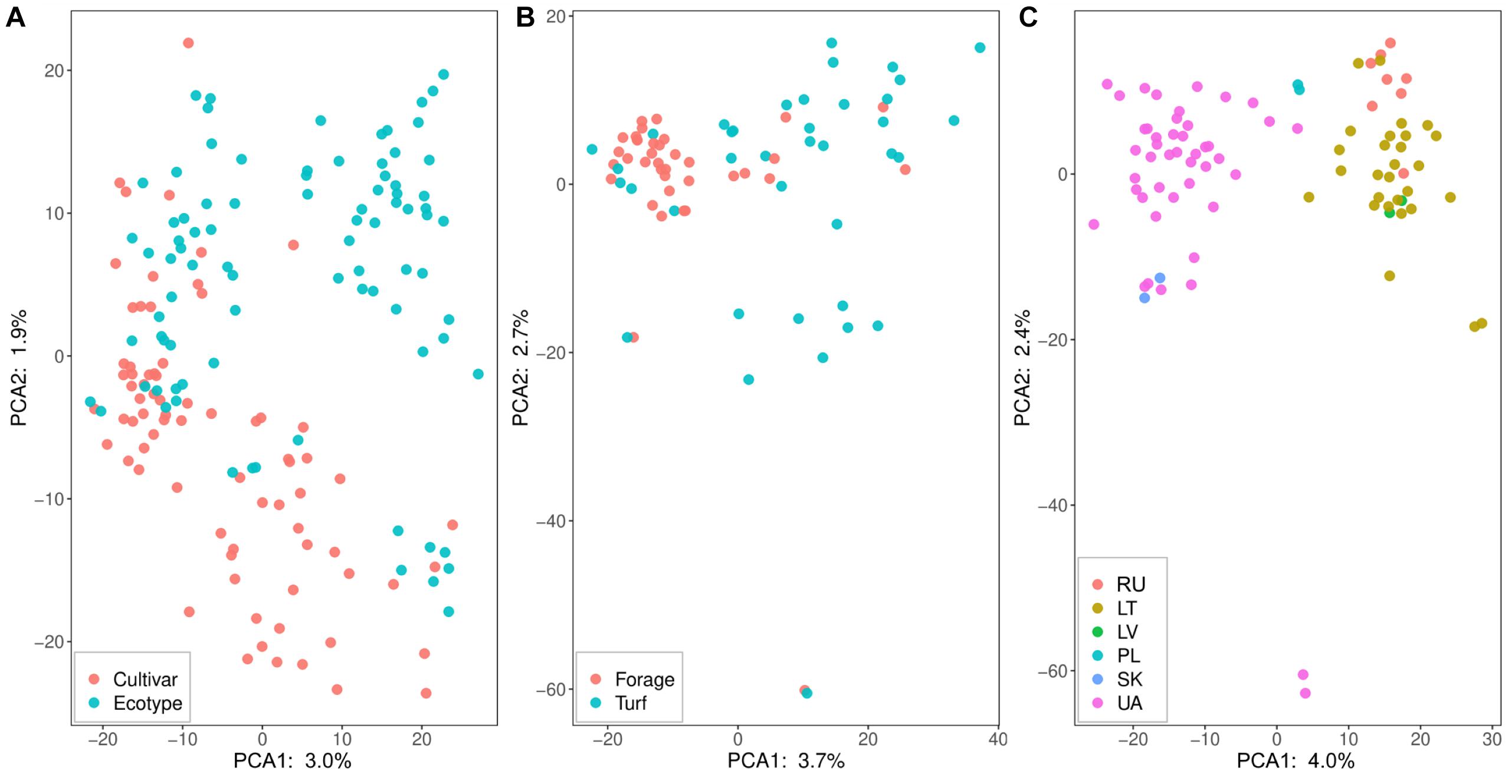
Figure 3. Principal component analysis (PCA) of 197 perennial ryegrass cultivars and ecotypes based on genotyping-by-sequencing data. (A) shows the dispersion of cultivars and ecotypes where cultivars are indicated in red and ecotypes in blue dots. The dispersion of cultivars based on the type is shown in (B), where forage type cultivars are represented in red dots and blue dots indicate turf type cultivars. (C) demonstrates the distribution of the ecotypes according to their country of origin. Countries are presented in two-letter codes: RU, Kaliningrad region, Russian Federation; LT, Lithuania; LV, Latvia; PL, Poland; SK, Slovakia; and UA, Ukraine.
GWAS for Biomass Formation
Marker trait associations were tested individually using SMR and three multi-QTL approaches: FarmCPU, BLINK, and MLMM. No significant marker trait associations were found after correcting for multiple testing, using SMR. This is attributable to the incomplete genotypic data from GBS and rare alleles coupled with strict multiple testing. Although three SNPs were significantly associated with the growth reduction point Σ, after correction for multiple testing, when using FarmCPU, BLINK, and MLMM. These markers are in close proximity or within predicted genes (Table 1 and Figure 4). However, no SNPs were significantly associated with other traits. The significant MYB41 loci found by FarmCPU had the lowest P-value (P = 6.35E–06) when tested by SMR model. Given that both Bayesian and frequentist procedures, using FarmCPU and SMR statistical approaches, respectively, identified the same loci suggests they are important.
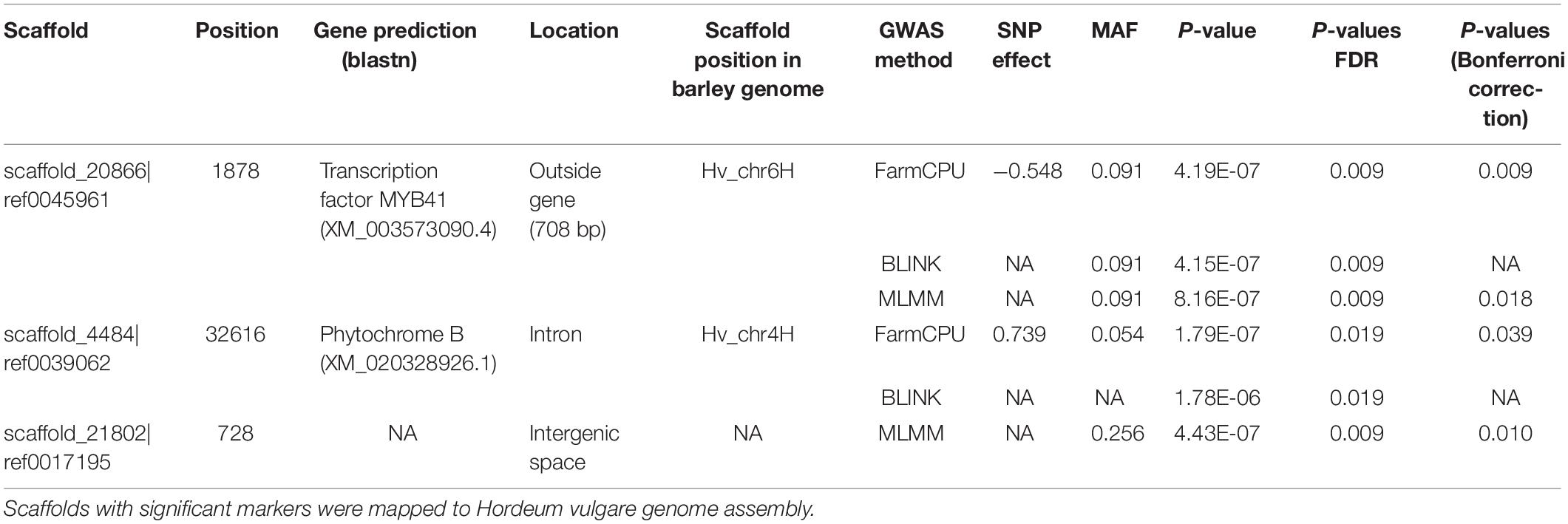
Table 1. Description of the most significant marker trait associations for the growth decrease trait on genomic scaffolds.
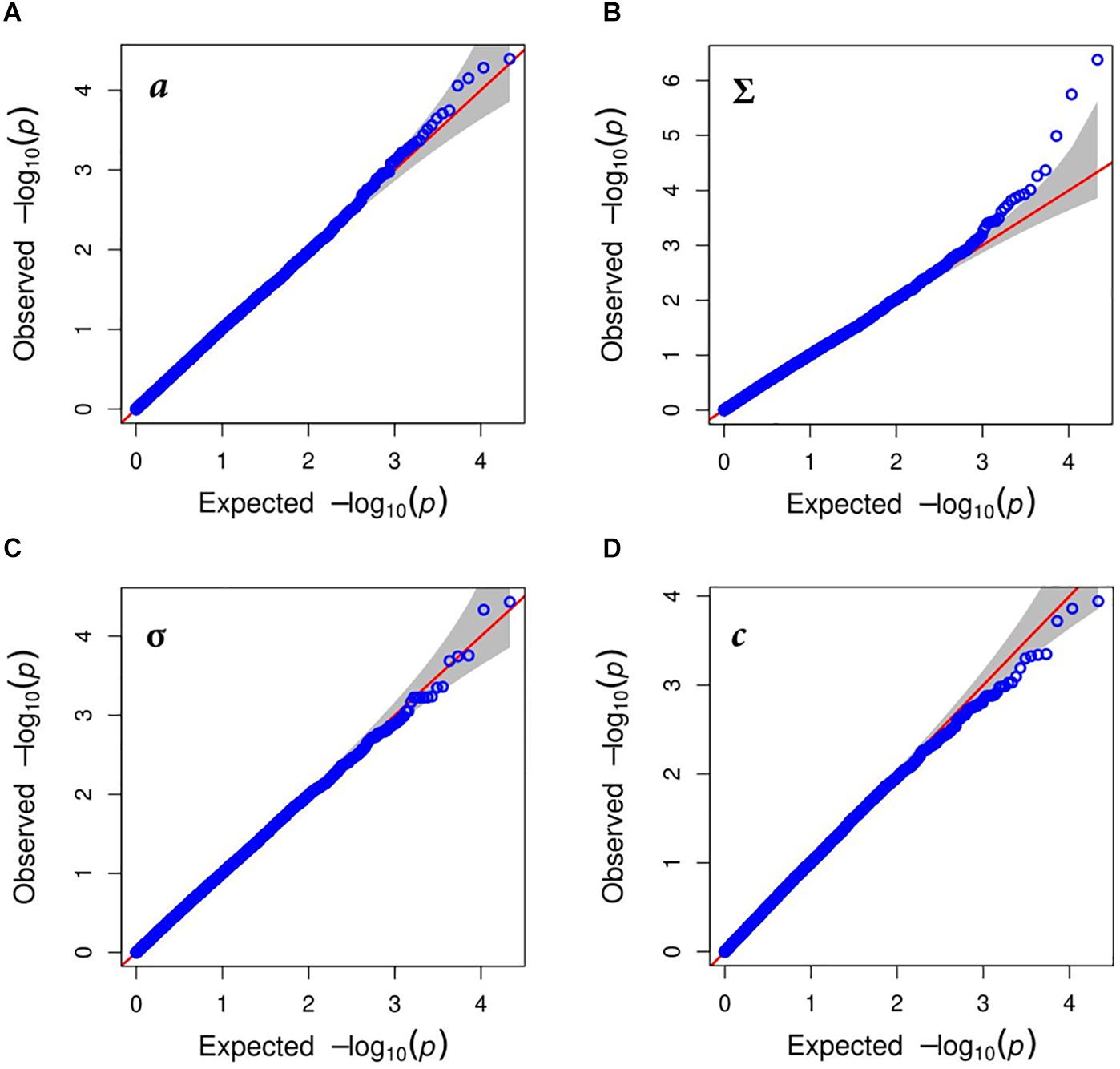
Figure 4. Quantile-quantile (QQ)-plots for biomass formation under water deficit condition traits after correction for population structure: (A) shows thermal leaf growth (a) of perennial ryegrass under optimal growth conditions; leaf growth reduction point (Σ) presented in (B); in (C) the leaf growth arrest point (σ) shown and tolerance to water deprivation (c) traits is given in (D).
The location of the marker-tagged scaffolds at the chromosome level was revealed by BLAST alignment to the barley H. vulgare genome (Mascher et al., 2017). Two of the scaffolds which harbored the most significant markers were anchored to chromosomes 4 and 6 in barley. The allele substitution effects for the significant markers associated with growth reduction ranged from −0.548 to 0.739 log10 (hPa). The most significant SNP (P = 4.19E–07) with an estimated effect size of −0.548 log10 (hPa) was located on a small genomic scaffold 20866| ref0045961 at position 1,878 bp. This scaffold harbored a single gene prediction for MYB41 transcription factor (TF), while the marker was of 708 bp away from the gene. The second most significant SNP (P = 1.79E–07), estimated effect of 0.739 log10 (hPa) and was located at 32,616 bp of scaffold 4484| ref0039062 within the intron of predicted gene with a high sequence homology to Phytochrome B. The third most significant marker (P = 4.436E–07) was on scaffold 21802| ref0017195 at position 728 bp. However, there were no gene predictions in close proximity to the marker.
Discussion
This study revealed the wide range of growth response to water deficit in a diverse panel of perennial ryegrass in a changing environment. The most sensitive genotypes limited the growth already under moist conditions [Σ < 1.50 log10 (hPa)], while some of the genotypes maintained leaf growth under lethal conditions [Σ > 4.00 log10 (hPa)]. The slower growth of genotypes representing turf cultivars obtained in the study reflects the decades of selective breeding aiming in opposite directions for forage and amenity grasses. The results show that turf types have shorter leaves because they grow slower. Though this may also be attributable to reduced growth duration, which was not evaluated in this study. The shorter, narrower leaves and lower plant height of turf genotypes was already confirmed in previous studies on plant architecture by Statkevičiūtė et al. (2015, 2018). The forage varieties in contrast have been selected for high biomass, for which leaf growth is an important factor (Wilkins, 1991; Barre et al., 2006).
Natural ecotypes are considered a source of stress resistance. Furthermore, they can possess high yielding potential, comparable to that of registered cultivars (Bachmann-Pfabe et al., 2018). The ecotypes in our study demonstrated similar growth patterns to the forage cultivar group. The significant correlation between the growth reduction (Σ) and the growth arrest (σ) points was estimated in all genotype groups except the forage cultivars, but it is worth noting that this correlation was strongest in the continental climate ecotype group. Extensive studies of natural perennial ryegrass diversity in Europe demonstrated that modern cultivars are mostly related to north-western Europe ecotypes (Blanco-Pastor et al., 2019), leaving the vast majority of the natural genetic variation unexploited. This was confirmed in our study as most ecotypes, especially the ones originating from Ukraine, genetically separated from the cultivars with very little overlap. Furthermore, the latitudinal position correlated highly with the first principle component of genetic structure suggesting latitude was a prominent force shaping the diversity of wild-growing perennial ryegrass populations. From a breeder’s perspective, combining an ecotype that responds late to water stress with elite germplasm that has high thermal growth rate would result in a high yielding drought tolerant cultivars.
Three significant marker trait associations were found with growth reduction under water deficit, with all of them passing Bonferroni correction. For two of these markers are in close proximity to predicted genes with sequence homology to PhytochromeB (PhyB) and MYB41. Both genes have well established functions within plant kingdom.
The phytochrome family is a part of the plant circadian clock, they initiate and mediate a complex photomorphogenesis signal cascade when activated by the red light, with the PhyB being the main photoreceptor (Nagy and Schäfer, 2002; Srivastava et al., 2019). The role of PhyB in plant growth and development has been known for decades when abnormal elongation of various tissues, from stems to root hair, has been demonstrated in Arabidopsis thaliana phyB mutant (Reed et al., 1993). Transgenic plants overexpressing PhyB had increased transpiration rate caused by higher stomata density and stomatal index, leading to enhanced photosynthesis but reduced water-use efficiency (Boccalandro et al., 2009). The development of rice phyB mutants did not differ from WT plants under well-watered conditions; but exhibited higher tolerance when drought stress was applied. This was mainly due to reduced transpiration and stomatal density compared to WT plants (Liu et al., 2012). PhyB was also shown to enhance ABA sensitivity under water shortage in A. thaliana (González et al., 2012). Moreover, the role of PhyB was shown in other abiotic stresses, like heat and cold (He Y. et al., 2016; Song et al., 2017). Plant hormone biosynthesis and signaling pathway genes, as well as abiotic stress-related genes were found to be regulated by PhyB in wheat (Pearce et al., 2016).
The MYB proteins comprise one of the largest TF transcription factor families in plants and are known to be involved in multiple functions, including plant development, growth, metabolism, cell fate, and abiotic stress response (Dubos et al., 2010; Baldoni et al., 2015; Li et al., 2015). The R2R3-MYB family, which is the biggest MYB transcription factor family in plants, has been studied in various plant species, especially Arabidopsis (Dubos et al., 2010; Du et al., 2012a, b; Katiyar et al., 2012; He et al., 2019). However, not much is known about the MYB TFs in ryegrasses. A number of MYB TFs were demonstrated to coordinate drought and salt stress responses in cotton, sheepgrass and rice with various effects; some were down-regulated during stress, some up-regulated, or differentially expressed in different plant organs (Chen et al., 2015; Zhu et al., 2015; He Q. et al., 2016; Zhao et al., 2019). In Arabidopsis, the expression of MYB41 was only detectable under drought, salt and ABA treatment. Transgenic plants over-expressing this gene were more sensitive to desiccation and had smaller cells among other phenotypic effects. Furthermore, the expression of genes regulating lipid metabolism, cell wall modifications and cell expansion was altered (Cominelli et al., 2008). The increased expression of MYB41 in osmotic stress response was confirmed in a study by Lippold et al. (2009), although the effect of MYB41 over-expression on plant growth were less drastic. Involvement of MYB41 in cell-wall associated lipid metabolism as well as activation of MYB41 promoter in the endodermis under abiotic stress was observed in both Arabidopsis and Nicotiana benthamiana (Kosma et al., 2014). These results suggest that MYB41 might confer osmotic stress resistance by regulating cell wall and leaf cuticle deposition, yet the exact mode of action of this particular TF is not defined due to the complexity of the gene regulation network it is involved in. A number of other MYB TFs modulate plant stress resistance in a similar manner to PhyB by regulating size, density, and opening rates of stomata (Baldoni et al., 2015; Chen et al., 2015; Butt et al., 2017).
Taken together, this study combines precision phenotyping of leaf growth in response to water stress, with genome wide association. The PhyB and MYB41 genes, identified in this study, are known to modulate abiotic stress response in many plant species. Given the sub-gene LD level in this population, suggests they are the causal genes underlying the detected QTL. To our knowledge this is the first time these genes have been associated with perennial ryegrass leaf growth under drought stress. The study also has practical applications for plant breeding; depicting how different, natural, alleles might be used to improve drought tolerance and what effect they have. In addition, the genes found highlight the potential of the phenotyping method. Namely, that using leaf growth as a proxy for stress response is a tool that can be used to dissect a complex trait into fundamental components. Thus, improving our understanding of plant behavior under water stress and subsequently, yielding potential in the regions experiencing frequent albeit mild summer droughts. Cultivars, purposely selected for a high tolerance, in terms of growth reduction and arrest by our phenotyping model, might provide a reliable feed source in the changing environment of the future world.
Data Availability Statement
The data is shared on https://doi.org/10.6084/m9.figshare.12433799.v3 and https://doi.org/10.6084/m9.figshare.12433715.v3.
Author Contributions
KJ, SY, and BS conceived the study. SY and KJ conducted the experiments. KJ, AA, and SY performed data analysis. VK and GS assisted in data analysis and interpretation. KJ, GS, and AA drafted the manuscript, which was improved by SY, VK, and BS. All authors read and approved the final manuscript.
Funding
This work was funded by the Research Council of Lithuania Grant No. S-MIP-17-24 (GrowGene) and Sciex-NMSch, a Scientific Exchange Program between Switzerland and the New Member States of the European Union (Project Code 13.032).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Prof. Achim Walter from ETH Zurich for providing access to infrastructure of the Crop Science group. Also, we want to thank Dr. Norbert Kirchgessner and Dr. Frank Liebisch for assistance in the setup of the phenotyping experiment and data analysis.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.570204/full#supplementary-material
Footnotes
References
Aleliūnas, A., Jonavičienė, K., Statkevičiūtė, G., Vaitiekūnaitė, D., Kemešytė, V., Lübberstedt, T., et al. (2015). Association of single nucleotide polymorphisms in LpIRI1 gene with freezing tolerance traits in perennial ryegrass. Euphytica 204, 523–534. doi: 10.1007/s10681-014-1330-y
Auton, A., Albers, C. A., Banks, E., Marth, G. T., Lunter, G., McVean, G., et al. (2011). The variant call format and VCF tools. Bioinformatics 27, 2156–2158. doi: 10.1093/bioinformatics/btr330
Bachmann-Pfabe, S., Willner, E., and Dehmer, K. J. (2018). Ex-situ evaluations of Lolium perenne L. ecotypes collected in Bulgaria, Croatia, Spain and Ireland reveal valuable breeding material. Genet. Resour. Crop Evol. 65, 1423–1439. doi: 10.1007/s10722-018-0623-8
Bac-Molenaar, J. A., Granier, C., Keurentjes, J. J. B., and Vreugdenhil, D. (2016). Genome-wide association mapping of time-dependent growth responses to moderate drought stress in Arabidopsis. Plant Cell Environ. 39, 88–102. doi: 10.1111/pce.12595
Baldoni, E., Genga, A., and Cominelli, E. (2015). Plant MYB transcription factors: their role in drought response mechanisms. Int. J. Mol. Sci. 16, 15811–15851. doi: 10.3390/ijms160715811
Barre, P., Emile, J.-C., Betin, M., Surault, F., Ghesquière, M., and Hazard, L. (2006). Morphological characteristics of perennial ryegrass leaves that influence short-term intake in dairy cows. Agron. J. 98, 978–985. doi: 10.2134/agronj2005.0213
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 57, 289–300. Available online at: http://www.jstor.org/stable/2346101
Blanco-Pastor, J. L., Manel, S., Barre, P., Roschanski, A. M., Willner, E., Dehmer, K. J., et al. (2019). Pleistocene climate changes, and not agricultural spread, accounts for range expansion and admixture in the dominant grassland species Lolium perenne L. J. Biogeogr. 46, 1451–1465. doi: 10.1111/jbi.13587
Boccalandro, H. E., Rugnone, M. L., Moreno, J. E., Ploschuk, E. L., Serna, L., Yanovsky, M. J., et al. (2009). Phytochrome B enhances photosynthesis at the expense of water-use efficiency in arabidopsis. Plant Physiol. 150, 1083–1092. doi: 10.1104/pp.109.135509
Bothe, A., Westermeier, P., Wosnitza, A., Willner, E., Schum, A., Dehmer, K. J., et al. (2018). Drought tolerance in perennial ryegrass (Lolium perenne L.) as assessed by two contrasting phenotyping systems. J. Agron. Crop Sci. 204, 375–389. doi: 10.1111/jac.12269
Boyer, J. S. (1982). Plant productivity and environment. Science 218, 443–448. doi: 10.1126/science.218.4571.443
Butt, H. I., Yang, Z., Gong, Q., Chen, E., Wang, X., Zhao, G., et al. (2017). GaMYB85, an R2R3 MYB gene, in transgenic Arabidopsis plays an important role in drought tolerance. BMC Plant Biol. 17:142. doi: 10.1186/s12870-017-1078
Byrne, S. L., Nagy, I., Pfeifer, M., Armstead, I., Swain, S., Studer, B., et al. (2015). A synteny-based draft genome sequence of the forage grass Lolium perenne. Plant J. 84, 816–826. doi: 10.1111/tpj.13037
Chen, T., Li, W., Hu, X., Guo, J., Liu, A., and Zhang, B. (2015). A cotton MYB transcription factor, GbMYB5, is positively involved in plant adaptive response to drought stress. Plant Cell Physiol. 56, 917–929. doi: 10.1093/pcp/pcv019
Claeys, H., and Inzé, D. (2013). The agony of choice: how plants balance growth and survival under water-limiting conditions. Plant Physiol. 162, 1768–1779. doi: 10.1104/pp.113.220921
Cominelli, E., Sala, T., Calvi, D., Gusmaroli, G., and Tonelli, C. (2008). Over-expression of the Arabidopsis AtMYB41 gene alters cell expansion and leaf surface permeability. Plant J. 53, 53–64. doi: 10.1111/j.1365-313X.2007.03310.x
Condon, A. G., Richards, R. A., Rebetzke, G. J., and Farquhar, G. D. (2004). Breeding for high water-use efficiency. J. Exp. Bot. 55, 2447–2460. doi: 10.1093/jxb/erh277
Daryanto, S., Wang, L., and Jacinthe, P.-A. (2016). Global synthesis of drought effects on maize and wheat production. PLoS One 11:e0156362. doi: 10.1371/journal.pone.0156362
DePristo, M. A., Banks, E., Poplin, R., Garimella, K. V., Maguire, J. R., Hartl, C., et al. (2011). A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43:491. doi: 10.1038/ng.806
Du, H., Feng, B. R., Yang, S. S., Huang, Y. B., and Tang, Y. X. (2012a). The R2R3-MYB transcription factor gene family in maize. PLoS One 7:e37463. doi: 10.1371/journal.pone.0037463
Du, H., Yang, S. S., Liang, Z., Feng, B. R., Liu, L., Huang, Y. B., et al. (2012b). Genome-wide analysis of the MYB transcription factor superfamily in soybean. BMC Plant Biol. 12:106. doi: 10.1186/1471-2229-12
Dubos, C., Stracke, R., Grotewold, E., Weisshaar, B., Martin, C., and Lepiniec, L. (2010). MYB transcription factors in Arabidopsis. Trends Plant Sci. 15, 573–581. doi: 10.1016/j.tplants.2010.06.005
Ergon, Å, Seddaiu, G., Korhonen, P., Virkajärvi, P., Bellocchi, G., Østrem, L., et al. (2018). How can forage production in Nordic and Mediterranean Europe adapt to the challenges and opportunities arising from climate change? To cite this version: HAL Id: hal-01635214 the challenges and opportunities arising from climate change? Eur. J. Agron. 92, 97–106. doi: 10.1016/j.eja.2017.09.016
Fahad, S., Bajwa, A. A., Nazir, U., Anjum, S. A., Farooq, A., Zohaib, A., et al. (2017). Crop production under drought and heat stress: plant responses and management options. Front. Plant Sci. 8:1147. doi: 10.3389/fpls.2017.01147
Farooq, M., Hussain, M., Wahid, A., and Siddique, K. H. M. (2012). “Drought stress in plants: an overview,” in Plant Responses to Drought Stress: from Morphological to Molecular Features, ed. R. Aroca (Berlin: Springer Berlin Heidelberg), 1–33. doi: 10.1007/978-3-642-32653-0_1
Forsman, A. (2014). Rethinking phenotypic plasticity and its consequences for individuals, populations and species. Heredity (Edinb). 115, 276–284. doi: 10.1038/hdy.2014.92
González, C. V., Ibarra, S. E., Piccoli, P. N., Botto, J. F., and Boccalandro, H. E. (2012). Phytochrome B increases drought tolerance by enhancing ABA sensitivity in Arabidopsis thaliana. Plant Cell Environ. 35, 1958–1968. doi: 10.1111/j.1365-3040.2012.02529.x
He, C., Teixeira da Silva, J. A., Wang, H., Si, C., Zhang, M., Zhang, X., et al. (2019). Mining MYB transcription factors from the genomes of orchids (Phalaenopsis and Dendrobium) and characterization of an orchid R2R3-MYB gene involved in water-soluble polysaccharide biosynthesis. Sci. Rep. 9, 1–19. doi: 10.1038/s41598-019-49812
He, Q., Jones, D. C., Li, W., Xie, F., Ma, J., Sun, R., et al. (2016). Genome-wide identification of R2R3-MYB genes and expression analyses during abiotic stress in Gossypium raimondii. Sci. Rep. 6:22980. doi: 10.1038/srep22980
He, Y., Li, Y., Cui, L., Xie, L., Zheng, C., Zhou, G., et al. (2016). Phytochrome B negatively affects cold tolerance by regulating OsDREB1 gene expression through phytochrome interacting factor-like protein OsPIL16 in rice. Front. Plant Sci. 7:1963. doi: 10.3389/fpls.2016.01963
Hu, H., and Xiong, L. (2014). Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 65, 715–741. doi: 10.1146/annurev-arplant-050213
Humphreys, M., Feuerstein, U., Vandewalle, M., and Baert, J. (2010). “Ryegrasses,” in Fodder Crops and Amenity Grasses, eds B. Boller, U. K. Posselt, and F. Veronesi (New York, NY: Springer New York), 211–260. doi: 10.1007/978-1-4419-0760-8_10
Jonavičienė, K., Statkevičiūtė, G., Kemešytė, V., and Brazauskas, G. (2014). Genetic and phenotypic diversity for drought tolerance in perennial ryegrass (Lolium perenne L.). Zemdirbyste Agricult. 101, 411–418. doi: 10.13080/z-a.2014.101.052
Katiyar, A., Smita, S., Lenka, S. K., Rajwanshi, R., Chinnusamy, V., and Bansal, K. C. (2012). Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis. BMC Genomics 13:544. doi: 10.1186/1471-2164-13
Kemesyte, V., Statkeviciute, G., and Brazauskas, G. (2017). Perennial ryegrass yield performance under abiotic stress. Crop Sci. 57, 1935–1940. doi: 10.2135/cropsci2016.10.0864
Kooyers, N. J. (2015). The evolution of drought escape and avoidance in natural herbaceous populations. Plant Sci. 234, 155–162. doi: 10.1016/j.plantsci.2015.02.012
Korte, C. J., and Chu, A. C. P. (1983). “Some effects of drought on perennial ryegrass swards,” in Proceedings of the New Zealand Grassland Association, Blenheim, 211–216. doi: 10.33584/jnzg.1983.44.1625
Kosma, D. K., Murmu, J., Razeq, F. M., Santos, P., Bourgault, R., Molina, I., et al. (2014). AtMYB41 activates ectopic suberin synthesis and assembly in multiple plant species and cell types. Plant J. 80, 216–229. doi: 10.1111/tpj.12624
Kruijer, W., Boer, M. P., Malosetti, M., Flood, P. J., Engel, B., Kooke, R., et al. (2014). Marker-based estimation of heritability in immortal populations. Genetics 199, 379–398. doi: 10.1534/genetics.114.167916
Kumar, A., Dixit, S., Ram, T., Yadaw, R. B., Mishra, K. K., and Mandal, N. P. (2014). Breeding high-yielding drought-tolerant rice: genetic variations and conventional and molecular approaches. J. Exp. Bot. 65, 6265–6278. doi: 10.1093/jxb/eru363
Lee, M., Howard-Andrews, V., and Chester, M. (2019). Resistance of multiple diploid and tetraploid perennial ryegrass (Lolium perenne L.) Varieties to Three Projected Drought Scenarios for the UK in 2080. Agronomy 9:159. doi: 10.3390/agronomy9030159
Lenth, R. V. (2016). Least-squares means: The R package lsmeans. J. Stat. Softw. 69. doi: 10.18637/jss.v069.i01
Li, C., Ng, C. K. Y., and Fan, L. M. (2015). MYB transcription factors; active players in abiotic stress signaling. Environ. Exp. Bot. 114, 80–91. doi: 10.1016/j.envexpbot.2014.06.014
Li, H. (2013). Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv [Preprint]. 1303.3997
Lippold, F., Sanchez, D. H., Musialak, M., Schlereth, A., Scheible, W. R., Hincha, D. K., et al. (2009). AtMyb41 regulates transcriptional and metabolic responses to osmotic stress in Arabidopsis. Plant Physiol. 149, 1761–1762. doi: 10.1104/pp.108.134874
Liu, J., Zhang, F., Zhou, J., Chen, F., Wang, B., and Xie, X. (2012). Phytochrome B control of total leaf area and stomatal density affects drought tolerance in rice. Plant Mol. Biol. 78, 289–300. doi: 10.1007/s11103-011-9860
Liu, X., Huang, M., Fan, B., Buckler, E. S., and Zhang, Z. (2016). Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 12:e1005767. doi: 10.1371/journal.pgen.1005767
Lobell, D. B., and Field, C. B. (2007). Global scale climate–crop yield relationships and the impacts of recent warming. Environ. Res. Lett. 2:014002. doi: 10.1088/1748-9326/2/1/014002
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 17:10. doi: 10.14806/ej.17.1.200
Mascher, M., Gundlach, H., Himmelbach, A., Beier, S., Twardziok, S. O., Wicker, T., et al. (2017). A chromosome conformation capture ordered sequence of the barley genome. Nature 544:427. doi: 10.1038/nature22043
Mayjonade, B., Gouzy, J., Donnadieu, C., Pouilly, N., Marande, W., Callot, C., et al. (2016). Extraction of high-molecular-weight genomic DNA for long-read sequencing of single molecules. Biotechniques 61, 203–205. doi: 10.2144/000114460
McKenna, A., Hanna, M., Banks, E., Sivachenko, A., Cibulskis, K., Kernytsky, A., et al. (2010). The genome analysis toolkit: a mapreduce framework for analyzing next-generation DNA sequencing data. Genome Res. 20, 1297–1303. doi: 10.1101/gr.107524.110.20
Moore, F. C., and Lobell, D. B. (2015). The fingerprint of climate trends on European crop yields. Proc. Natl. Acad. Sci. U.S.A. 112, 2670–2675. doi: 10.1073/pnas.1409606112
Mwadzingeni, L., Shimelis, H., Dube, E., Laing, M. D., and Tsilo, T. J. (2016). Breeding wheat for drought tolerance: progress and technologies. J. Integr. Agric. 15, 935–943. doi: 10.1016/S2095-3119(15)61102
Nagelmüller, S., Kirchgessner, N., Yates, S., Hiltpold, M., and Walter, A. (2016). Leaf Length Tracker: a novel approach to analyse leaf elongation close to the thermal limit of growth in the field. J. Exp. Bot. 67, 1897–1906. doi: 10.1093/jxb/erw003
Nagy, F., and Schäfer, E. (2002). Phytochromes control photomorphogenesis by differentially regulated, interacting signaling pathways in higher plants. Annu. Rev. Plant Biol. 53, 329–355. doi: 10.1146/annurev.arplant.53.100301.135302
Pearce, S., Kippes, N., Chen, A., Debernardi, J. M., and Dubcovsky, J. (2016). RNA-seq studies using wheat PHYTOCHROME B and PHYTOCHROME C mutants reveal shared and specific functions in the regulation of flowering and shade-avoidance pathways. BMC Plant Biol. 16:141. doi: 10.1186/s12870-016-0831
R Development Core Team (2005). R: A Language and Environment for Statistical Computing. Available online at: http://www.r-project.org
Reed, J. W., Nagpal, P., Poole, D. S., Furuya, M., and Chory, J. (1993). Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell 5, 147–157. doi: 10.2307/3869581
Salekdeh, G. H., Reynolds, M., Bennett, J., and Boyer, J. (2009). Conceptual framework for drought phenotyping during molecular breeding. Trends Plant Sci. 14, 488–496. doi: 10.1016/j.tplants.2009.07.007
Sampoux, J.-P., Baudouin, P., Bayle, B., Béguier, V., Bourdon, P., Chosson, J.-F., et al. (2011). Breeding perennial grasses for forage usage: an experimental assessment of trait changes in diploid perennial ryegrass (Lolium perenne L.) cultivars released in the last four decades. F. Crop. Res. 123, 117–129. doi: 10.1016/j.fcr.2011.05.007
Sampoux, J. P., Baudouin, P., Bayle, B., Béguier, V., Bourdon, P., Chosson, J. F., et al. (2013). Breeding perennial ryegrass (Lolium perenne L.) for turf usage: an assessment of genetic improvements in cultivars released in Europe, 1974–2004. Grass Forage Sci. 68, 33–48. doi: 10.1111/j.1365-2494.2012.00896.x
Sandhu, N., and Kumar, A. (2017). Bridging the rice yield gaps under drought: QTLs, genes, and their use in breeding programs. Agronomy 7, 1–27. doi: 10.3390/agronomy7020027
Segura, V., Vilhjálmsson, B. J., Platt, A., Korte, A., Seren, Ü., Long, Q., et al. (2012). An efficient multi-locus mixed-model approach for genome-wide association studies in structured populations. Nat. Genet. 44, 825–830. doi: 10.1038/ng.2314
Skirycz, A., Vandenbroucke, K., Clauw, P., Maleux, K., De Meyer, B., Dhondt, S., et al. (2011). Survival and growth of Arabidopsis plants given limited water are not equal. Nat. Biotechnol. 29, 212–214. doi: 10.1038/nbt.1800
Song, J., Liu, Q., Hu, B., and Wu, W. (2017). Photoreceptor phyB involved in Arabidopsis temperature perception and heat-tolerance formation. Int. J. Mol. Sci. 18:1194. doi: 10.3390/ijms18061194
Srivastava, D., Shamim, M., Kumar, M., Mishra, A., Maurya, R., Sharma, D., et al. (2019). Role of circadian rhythm in plant system: an update from development to stress response. Environ. Exp. Bot. 162, 256–271. doi: 10.1016/j.envexpbot.2019.02.025
Statkevičiūtė, G., Aleliūnas, A., Kemešytė, V., Pašakinskienė, I., Lübberstedt, T., and Brazauskas, G. (2015). Association analysis of five candidate genes with plant height and dry matter yield in perennial ryegrass. Plant Breed. 134, 454–460. doi: 10.1111/pbr.12280
Statkevičiūtė, G., Kemešytė, V., Aleliūnas, A., Jonavičienė, K., and Brazauskas, G. (2018). LpBRI1 polymorphism association with flag leaf architecture in perennial ryegrass. Zemdirbyste 105, 33–38. doi: 10.13080/z-a.2018.105.005
Wilkins, P. W. (1991). Breeding perennial ryegrass for agriculture. Euphytica 52, 201–214. doi: 10.1007/BF00029397
Yates, S., Jaškūnė, K., Liebisch, F., Nagelmüller, S., Kirchgessner, N., Kölliker, R., et al. (2019). Phenotyping a dynamic trait: leaf growth of perennial ryegrass under water limiting conditions. Front. Plant Sci. 10:344. doi: 10.3389/fpls.2019.00344
Zhang, Z., Liu, X., Zhou, Y., and Summers, R. M. (2018). BLINK: a package for the next level of genome-wide association studies with both individuals and markers Meng Huang. Gigascience 8, 1–12. doi: 10.1093/gigascience/giy154
Zhao, P., Hou, S., Guo, X., Jia, J., Yang, W., Liu, Z., et al. (2019). A MYB-related transcription factor from sheepgrass, LcMYB2, promotes seed germination and root growth under drought stress. BMC Plant Biol. 19:564. doi: 10.1186/s12870-019-2159
Keywords: leaf growth, drought tolerance, genome-wide association study, dynamic phenotyping, Lolium perenne L., Phytochrome B, MYB41
Citation: Jaškūnė K, Aleliūnas A, Statkevičiūtė G, Kemešytė V, Studer B and Yates S (2020) Genome-Wide Association Study to Identify Candidate Loci for Biomass Formation Under Water Deficit in Perennial Ryegrass. Front. Plant Sci. 11:570204. doi: 10.3389/fpls.2020.570204
Received: 06 June 2020; Accepted: 12 November 2020;
Published: 08 December 2020.
Edited by:
Soren K. Rasmussen, University of Copenhagen, DenmarkReviewed by:
Leif Skot, Aberystwyth University, United KingdomNa Luo, South China Agricultural University, China
Copyright © 2020 Jaškūnė, Aleliūnas, Statkevičiūtė, Kemešytė, Studer and Yates. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kristina Jaškūnė, a3Jpc3RpbmEuamFza3VuZUBsYW1tYy5sdA==; a3Jpc3RpbmEuam9uYXZpY2llbmVAbGFtbWMubHQ=; Steven Yates, c3RldmVuLnlhdGVzQHVzeXMuZXRoei5jaA==