 Rodrigo Cortés-Antiquera
Rodrigo Cortés-Antiquera Marisol Pizarro
Marisol Pizarro Rodrigo A. Contreras
Rodrigo A. Contreras Hans Köhler
Hans Köhler Gustavo E. Zúñiga
Gustavo E. Zúñiga- 1Departamento de Biologia, Facultad de Química y Biología, Universidad de Santago de Chile, Santiago, Chile
- 2Centro para el Desarrollo de la Nanociencia y la Nanotecnología (CEDENNA), Santiago, Chile
Deschampsia antarctica Desv, is the most successful colonizing species of a cold continent. In recent years due to climate change, the frequency of heat waves has increased in Antarctica, registering anomalous high temperatures during the summer of 2020. However, the populations of D. antarctica are responding positively to these events, increasing in number and size throughout the Antarctic Peninsula. In this work, the physiological and biochemical responses of D. antarctica plants grown in vitro (15 ± 1°C) and plants subjected to two heat shock treatments (23 and 35°C) were evaluated. The results obtained show that D. antarctica grown in vitro is capable of tolerating heat shock treatments; without showing visible damage to its morphology, or changes in its oxidative state and photosynthetic performance. These tolerance responses are primarily mediated by the efficient role of enzymatic and non-enzymatic antioxidant systems that maintain redox balance at higher temperatures. It is postulated that these mechanisms also operate in plants under natural conditions when exposed to environmental stresses.
Introduction
Antarctica is considered the most severe ecosystem in the world (Robinson et al., 2003). The temperature experiences daily fluctuations of −10°C to +15°C during summer (Convey, 2013). These extreme climatic conditions explain the scarce plant biodiversity that exists in Antarctica (Convey, 2006), it has allowed that only two vascular plants have naturally colonized the Maritime Antarctica, Colobanthus quitensis (Kunth) Bartl caryophyllaceae and Deschampsia antarctica Desv poaceae.
Both Antarctic plants differ in their reproductive capacities (Gielwanoska and Szczuka, 2005), being D. antarctica the most successful colonizer of the Antarctic continent, compared to C. quitensis (Cavieres et al., 2016). D. antarctica is the most abundant and widely distributed throughout maritime Antarctica (Smith, 2003; Sáez et al., 2018). The plasticity of D. antarctica is a key to response to fluctuating environmental conditions (Gielwanoska et al., 2005). Anatomical modifications of the leaf anatomy present similitudes with Kranz anatomy associated with carbon metabolism, C4, furthermore presenting characteristics [R] more xeromorphic and ultrastructure differences in chloroplasts and mitochondria (Romero et al., 1999; Mantovani and Vieira, 2000; Gielwanoska and Szczuka, 2005), that improves the water-use efficiency (Montiel et al., 1999) and photosynthetic efficiency (Xiong et al., 1999; Sáez et al., 2017, 2018).
The effects of climate change are evident throughout the Antarctic Peninsula, recording the fastest atmospheric warming in the last 50 years (Turner and Overland, 2009). In this period, the average temperature in Antarctica increased by 3.7°C, a value much higher than the rest of the world (Turner et al., 2013). During the summer of 2020 in Antarctica, heatwaves were recorded at the Marambio base on Seymour Island, on the east side of the Antarctic peninsula, exceeding 20°C (Robinson et al., 2020). These warmer temperatures in Antarctica have produced dramatic changes on the continent: longer growing seasons with high temperatures, the retreat of glaciers, an increment of ice-free areas, and increased frequency of rainfall (Cavieres et al., 2016; Sáez et al., 2018). Warmer temperatures do not affect the landscapes, but could also affect the biota of the continent, changing the distribution and abundance of populations. In nature, any abrupt and short temperature increase of 5°C or more above the optimal growth temperature can be considered as a heat shock (Bita and Gerats, 2013). If the time of exposure to thermal shock is prolonged, it can irreversibly damage the stability of proteins, membranes, cytoskeletal structures, and RNA, affecting the growth and development of plants (Driendoks et al., 2015).
Although these negative effects of global warming on plants, populations of D. antarctica are increasing in number and size along the Antarctic Peninsula: King George Island (Torres-Mellado et al., 2011), Byers Peninsula, Livingston Island (Vera et al., 2013), Isla Robert (Torres-Mellado et al., 2011) and even in Argentine islands (Fowbert and Smith, 1994). The distribution of D. antarctica is highly influenced by climatic conditions (Parnikoza et al., 2011), studies carried out in ground conditions show that at warmer temperatures (13°C and 19°C), D. antarctica obtains the maximum CO2 assimilation rate, improving its photosynthetic performance and growth (Edwards and Smith, 1988; Sáez et al., 2017, 2018). This may explain the reproductive success of D. antarctica even in warmer conditions. It has been suggested that in the future, high temperatures could induce morphological and anatomical changes in Antarctic plants to maximize light capture and growth (Cavieres et al., 2016). Due to their successful adaptation to climate change and its rapid spread, D. antarctica has been suggested as an ecological marker for global warming and a valuable resource for understanding stress tolerance (Lee et al., 2012).
Plants have developed different mechanisms to tolerate heat shock: maintaining the stability of cell membranes, ROS uptake, antioxidant biosynthesis, accumulation of compatible osmolytes, induction of intracellular signaling cascades and the action of molecular chaperones (Wahid et al., 2007; Bita and Gerats, 2013). Several studies on D. antarctica have shown the effective role of antioxidants to respond to various types of abiotic stress such as osmotic stress (Zamora et al., 2010) and UV-B radiation stress (Sequeida et al., 2012; Köhler et al., 2017). While in heat shock events, the accumulation of heat shock protein 70 (Hsp70) in leaves has been described (Reyes et al., 2003) and the crucial high affinity of Rubisco for CO2 to optimize carbon assimilation (Sáez et al., 2018). Studies of in situ heating have provided important information on the response to thermal shock in D. antarctica. Working with plants grown in vitro allows to control environmental variables and expose D. antarctica to unusual conditions of heat shock, which contribute to clearly analyze the responses generated. We hypothesize that D. antarctica has efficient antioxidant protection mechanisms in response to heat shock treatment, being able to maintain normal physiology, photosynthetic performance, and redox state balance.
The objective of this work was to evaluate the capacity of D. antarctica cultivated in vitro to tolerate heat shock treatment, analyzing the effect of temperature on the redox balance, photosynthetic performance, and various enzymatic like SOD, POD, CAT, APX, and non-enzymatic antioxidant activities.
Materials and Methods
Plant Material
Plants of D. antarctica plants were grown in vitro at the Plant Physiology and Biotechnology Laboratory (Universidad Santiago de Chile). The plant material was micropropagated using Murashige and Skoog basal medium, supplemented with 20 g/L of sucrose, 5 mM myo-inositol, 2 mg/L naphthaleneacetic acid (NAA), 0.8 mg/L benzyl amino purine (BAP) and 4 g/L of activated carbon. The pH of the medium was adjusted to 5.6 and 6 g/L of agar-agar was added as a gelling agent. The culture medium was sterilized in an autoclave at 121°C and a pressure of 15 pounds. The in vitro plants were kept under controlled conditions at 15 ± 1°C and 16:8 photoperiods (16 h of light and 8 h of night) in a culture chamber at 70% of relative humidity. All in vitro plants used in the experiments were two-months old.
Lethal Temperature 50
The lethal temperature (LT50) is defined as the temperature necessary to kill 50% of the leaf tissue and was determined in D. antarctica grown in vitro. 100 mg of plant tissue were resuspended in 5 mL of deionized water in a Falcon tube and kept in a block heater (Thermo Scientific, Spain) for 3 h in a temperature range of 15–100°C. After This period, the plants were kept at room temperature for 10 min, and the maximum photosynthetic efficiency of photosystem II (PSII) (Fv/Fm) of the plant was determined as a survival parameter. The degree of damage caused by heat treatments was calculated considering 100% damage at a temperature of 100°C. The data obtained in the treatments were used in the determination of the LT50 in D. antarctica plants grown in vitro.
Heat Shock Treatment
Two heat shock treatments were used at 23 and 35°C, (8 and 20°C above the optimum growth temperature under laboratory conditions, respectively, for D. antarctica plants grown in vitro. This selection temperature was based on field temperature data recorded by our group in the South Shetlands Islands over the past 5 years (data not yet published). The plants were removed from the culture chamber at 15°C and exposed to the thermal shock treatments for 2 h (8–10 am) using a heating bath (Memmert, Germany) for a period of 7 days. The relative humidity inside the culture vessel was over 90%. The analyzes for both treated and control plants were performed on days 1, 2, 3, and 7. The aerial growth of the seedlings was evaluated immediately after the respective samplings, using ImageJ software, the samples were frozen in liquid nitrogen and stored at −80°C for later analysis.
Oxidative Stress Parameters
ROS’s total content was measured by fluorometric quantification of dichlorodihydrofluorescein-diacetate (DCDHF-DA) oxidized by the presence of ROS. 50 mg of fresh plant tissue were incubated in 1 mL of 10 mM DCDHF-DA in Tris–HCl buffer (50 mM, pH 8.0) for 1 h at 37°C in dark conditions. Then, the tissue was washed twice with a 50 mM EDTA solution to remove the remains of the incubation solution, and the tissue was ground under liquid nitrogen to a powder and extracted into 1 ml of Tris–HCl buffer (50 mM, pH 8.0) and centrifuged at 10,000 rpm for 10 min. The obtained supernatant was used to determine the fluorescence, using an excitation wavelength of 488 nm and an emission wavelength of 535 nm in a microplate reader spectrophotometer (TECAN, Infinite2000pro, Austria). The ROS content was determined using an oxidized DCF-DA calibration curve (Ross et al., 2008).
Lipid peroxidation was estimated by measuring the concentration of malondialdehyde (MDA) by the thiobarbituric acid reactive substances (TBARS) assay. 50 mg of fresh plant tissue was ground in liquid nitrogen to a powder and suspended in 1 ml of 1% trichloroacetic acid (TCA). Then the homogenate was centrifuged at 13,000 rpm for 5 min. 250 μL of the supernatant was mixed with 1 mL of 0.5% thiobarbituric acid (TBA) in 20% TCA. The mixture was boiled for 30 min and cooled to room temperature. The MDA formed was quantified at 532 nm and 600 nm in a microplate reader spectrophotometer (TECAN, Infinite2000pro, Austria), using a molar extinction coefficient of 155 mM-1 cm-1 (Ederli et al., 2004).
Photosynthetic Efficiency
Chlorophyll fluorescence was analyzed using the portable fluorometer PEA (Hansatech, England). The leaves were held in commercial leaf clips (Hansatech, England) that allow keeping the samples in the dark before the measurement (30 min) (Stirbet and Govindjee, 2011).
From the fluorescence induction signal from 10 μs to 3 s the instrument determines initial (Fo) and maximum (Fm) fluorescence and the variable fluorescence (Fv) at specified time intervals.
Furthermore, it calculates specific parameters such as the maximum efficiency of PSII (Fv/Fm), maximum quantum yield PSII (Φ PSII), and electron transport rate (ETR) (Stirbet and Govindjee, 2011).
Photosynthetic Pigments Content
The total content of chlorophyll and carotenoids was determined by the method of Wellburn and Lichtenthaler (1984). 100 mg of fresh plant tissue was used and macerated with liquid nitrogen in 1 mL of cold 80% acetone. The mixture was centrifuged at 13,000 rpm for 10 min and the supernatant was recovered. Each sample was diluted 1:10 and absorbance was measured at 470 nm, 649 nm, and 665 nm in a microplate reader spectrophotometer (TECAN, Infinite2000pro, Austria). Chlorophyll concentration was determined using Eqs. (1) and (2), and the ratio between chlorophyll was determined using Eq. (3). Carotenoids concentration was determined using Eq. (4)
Enzyme Extraction and Quantification
The protein extraction was conducted using 100 mg of fresh plant tissue and was ground to a powder and suspended in 1 mL of sodium phosphate buffer (50 mM, pH 7.5), and then the mixture was centrifuged at 10,000 rpm for 10 min at 4°C. The supernatant was recovered, and the total protein content was determined according to Bradford method (Bradford, 1976). The absorbance was measured at 595 nm on a UV-Vis spectrophotometer (Agilent 8453, Santa Clara, CA, United States). The total protein content was calculated using Bovine Serum Albumin (BSA) as a calibration standard (Bradford, 1976).
Antioxidant Enzymatic Activity
Superoxide dismutase activity (SOD, EC 1.15.1) was determined by using the photoinhibition of nitro-blue tetrazolium assay (NBT) (Beauchamp and Fridovich, 1971). A reaction mixture was prepared using 600 μL of sodium phosphate buffer (50 mM, pH 7.5), 10 μL of 10 mM EDTA, 100 μL of 130 mM methionine, 10 μL of 2 mM riboflavin, 200 μL of 3 mM of nitroblue tetrazolium (NBT) in 70% dimethylformamide (DMF) and 100 μL of protein extract. The mixture was incubated under white light for 15 min at room temperature (a blank kept in the dark). Absorbance at 560 nm was determined where one enzymatic unit (EU) was considered as the capacity to inhibit 50% of photochemical reduction of NBT (Köhler et al., 2017).
Total peroxidase class III activity (POD, EC 1.11.1.7) was determined by tetrahydro-guaiacol (THG) formation using peroxide and guaiacol as substrates (Pinhero et al., 1997). A reaction mixture that contained 1 mL of sodium phosphate buffer (50 mM, pH 7.5), 10 μL of protein extract, 5 μL of 100 vol. hydrogen peroxide and 5 μL of guaiacol was prepared. Absorbance at 470 nm was recorded after a reaction time of 1 min indicating POD activity in terms of tetrahydroguaiacol (THG) formation. POD activity was calculated using molar extinction of THG, ε = 26.6 mM-1 cm-1 (Köhler et al., 2017).
Ascorbate peroxidase (APX, EC 1.11.1.11) was determined by ascorbate consumption for 60 s (Lima et al., 2002). A reaction mixture that contained 1 mL of sodium phosphate buffer (50 mM, pH 7.5), 20 μL of protein extract, 5 μL of 100 vol. hydrogen peroxide and 40 μL of 10 mM sodium ascorbate was prepared. Absorbance at 290 nm was recorded after the reaction had proceeded for 1 min indicating APX activity in terms of ascorbate consumption. APX activity was calculated using molar extinction of ascorbate, ε = 2.8 mM-1 cm-1 (Köhler et al., 2017).
Catalase activity (CAT, EC 1.11.1.6) was determined by H2O2 consumption for 60 s (Lima et al., 2002). A reaction mixture that contained 900 μL of sodium phosphate buffer (50 mM, pH 7.5), 10 μL of protein extract and 3 μL of 100 vol. hydrogen peroxide was prepared. Absorbance at 240 nm was recorded after a reaction time of 1 min indicating CAT activity in terms of hydrogen peroxide consumption. CAT activity was calculated using molar extinction of hydrogen peroxide, ε = 39.4 mM-1 cm-1 (Köhler et al., 2017).
Hydroalcoholic Extracts Preparation
Hydroalcoholic extracts were prepared using 100 mg fresh plant tissue and were ground to a powder and suspended in 1 mL ethanol (85% v/v) and sonicated at 50-60 Hz for 3 h at room temperature. Extracts were filtered in a 0.45 um pore filter (Millipore, Billerica, MA, United States) and used for non-enzymatic antioxidant assays (Contreras et al., 2015).
Total Phenolic Content
The total phenolics content was determined by the modified Folin-Ciocalteu colorimetric method (Asami et al., 2003). A reaction mixture was prepared with 115 μL of deionized water, 20 μL of Folin-Ciocalteu reagent diluted (1:10), and 5 μL of hydroalcoholic extract. The mixture was incubated in the dark for 15 min at room temperature, then 60 μL of sodium carbonate (7% w / v) was added and incubated for 1 h at 35°C. The absorbance was measured at 740 nm in a microplate reader spectrophotometer (TECAN, Infinite2000pro, Austria). The total phenol content was expressed in gallic acid equivalents (GA) per gram of fresh weight (FW).
Non-enzymatic Antioxidant Activity
Antioxidant activity of hydroalcoholic extracts was measured by the bleaching of 1,1-Difenil-2-Picril-Hidrazil (DPPH) cation radical (Brand-Williams et al., 1995). A reaction mixture with 100 μL DPPH 100 mM (A517 = 0,7 − 0,8) and 10 μL protein extract was prepared. Absorbance at 515 nm was measured for 10 min at 37°C (TECAN, Infinite2000pro, Austria). The antioxidant activity were expressed as DPPH consumption percentage (%), where DPPH 100 mM was used as a control reference (Naik et al., 2005).
Statistical Analysis
All experiments were performed in triplicate, for statistically significant differences we used two-way ANOVA with multiple comparisons. Post statistical analysis were performed using Tukey’s post-test (P < 0.05).
Results
LT50 Determination
Two-month-old plants were used in the determination of LT50 (Figure 1). To determine heat tolerance, we calculated LT50 values based on the measurement of maximum PSII efficiency (Fv/Fm) in D. antarctica cultured in vitro. From this analysis, it was determined that the LT50 in D. antarctica shoots, grown under laboratory conditions at 15°C, was 52°C (Figure 2).

Figure 1. In vitro grown D. antarctica plants used in the experiments. Each culture vessel is considered a biological sample.
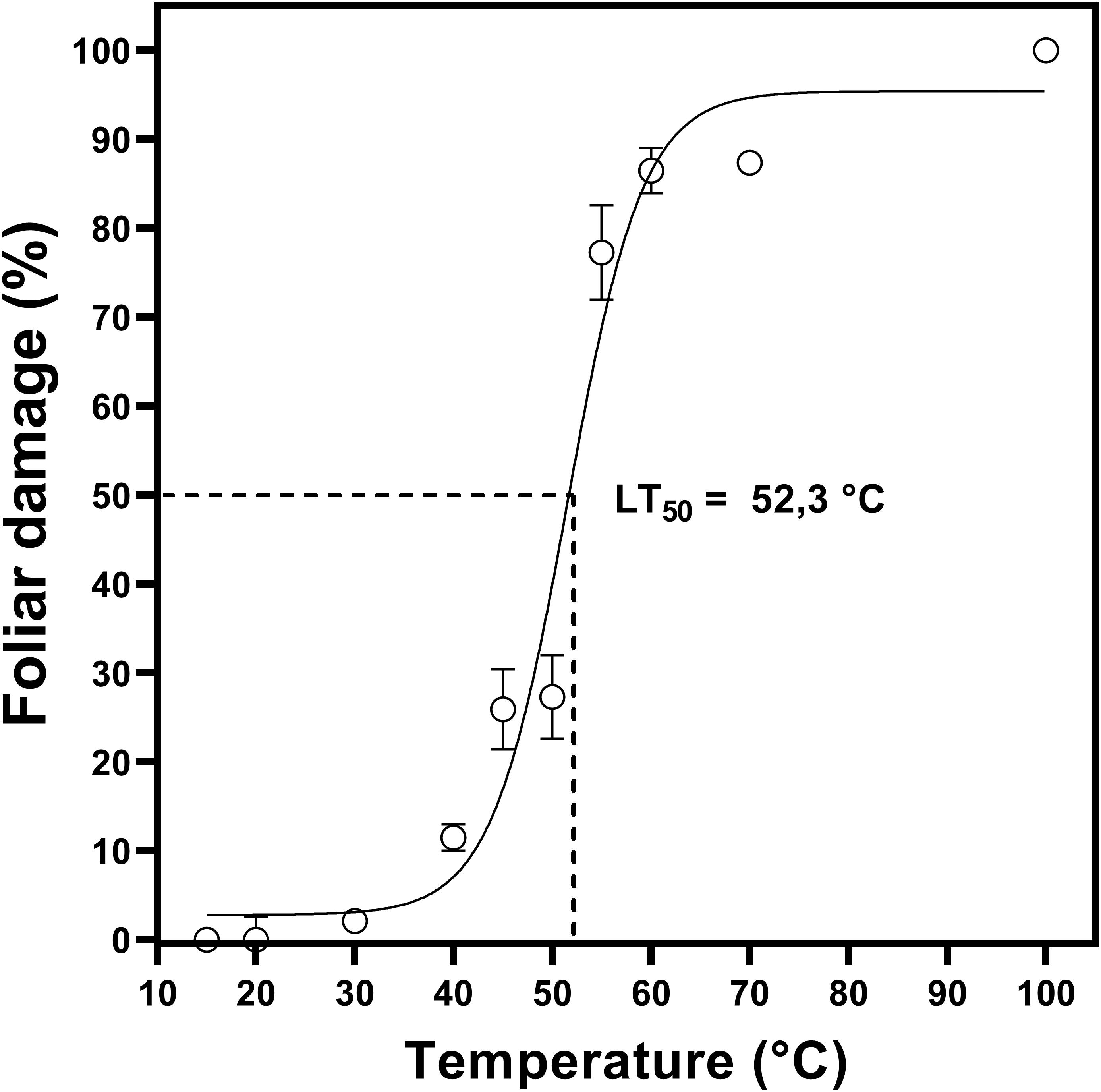
Figure 2. Sublethal temperature 50 (LT50) in D. antarctica cultivated in vitro. The photosynthetic parameter Fv/Fm was expressed like foliar damage (%). Each point represent means of 3 biological replicates (N = 3; ± standard error of the mean).
Heat Shock Effect on Physiology and Photosynthetic Efficiency
From the normal growth temperature, the determined LT50 value, and the temperature data recorded in Antarctica, two thermal shock treatments were applied (23 and 35°C). The plants treated at both temperatures did not present morphological differences concerning the control (Figure 3A), maintaining their coloration and leaf shape. When evaluating the effect of the treatment on plant growth, no significant differences were observed in the plants’ aerial length (P < 0.05) (Figure 3B).
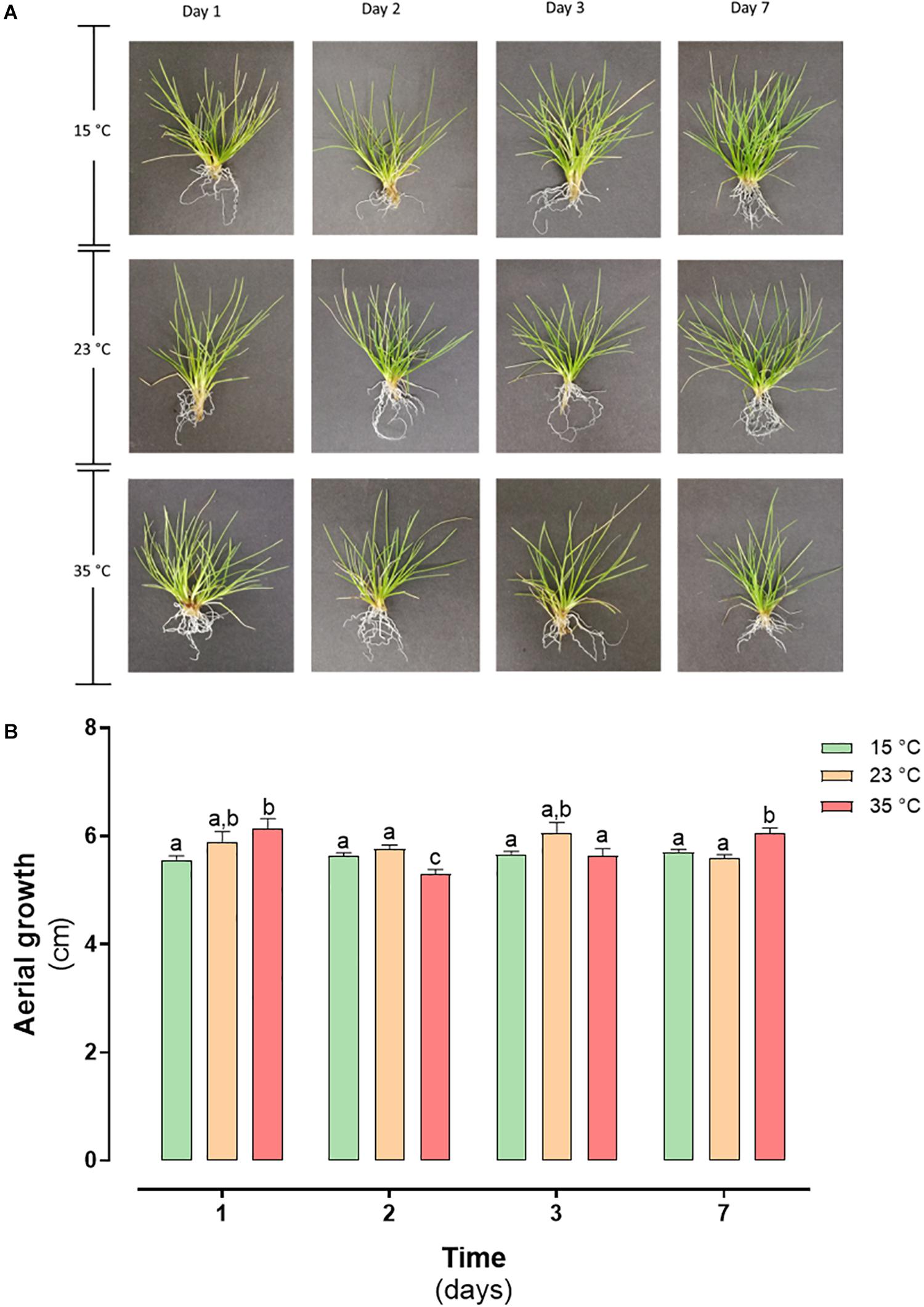
Figure 3. Heat shock effect in morphology and growth in D. antarctica control (15°C) and heat shock treatments (23°C and 35°C) cultivated in vitro. (A) Photographs of D. antarctica expose to heat shock treatments from day 1 at day 7 and (B) aerial growth. Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
As shown in Figure 4, we examined three photosynthetic parameters: chlorophyll fluorescence (Fv/Fm), quantum yield of photosystem II (ΦPSII) and electron transport rate (ETR). The maximal photochemical efficiency (Fv/Fm) was in the range 0.70–0.80 at both temperatures of heat shock (Figure 4A). The effective photochemical efficiency (ΦPSII) and electron transport rate (ETR) did not show any significant changes in D. antarctica exposed to heat shock at 23 and 35°C compared with the control (Figures 4B,C, respectively). These results on D. antarctica are comparable with previous results reported in nonstressed plants.
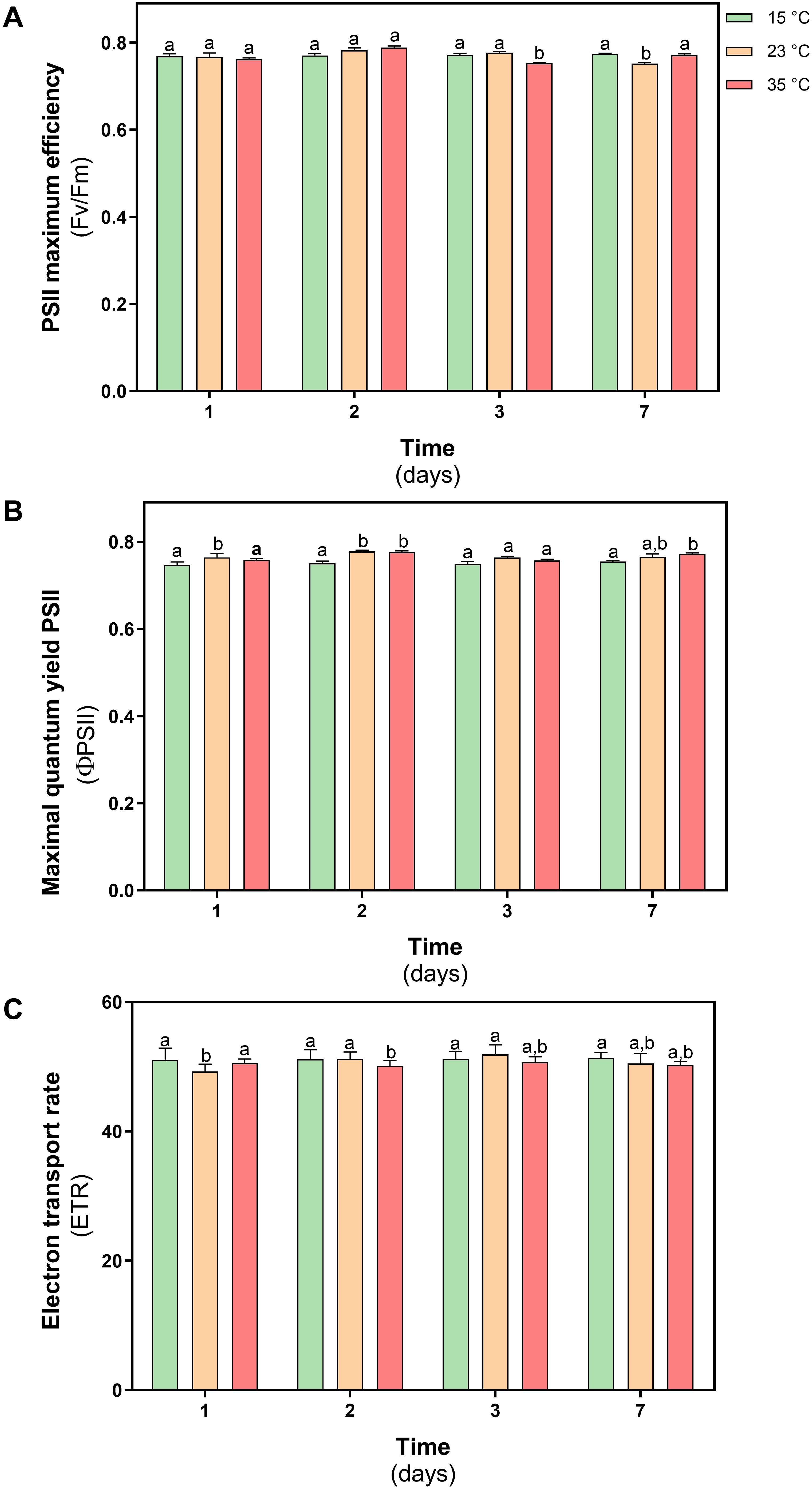
Figure 4. Photosynthetic efficiency parameters in D. antarctica control (15°C) and heat shock treatments (23°C and 35°C) cultivated in vitro. (A) PSII maximun efficiency (Fv/Fm), (B) maximal quantum yield PSII (ΦPSII), and (C) electron transport rate (ETR). Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
With the purpose of complementing the previous results, the levels of photosynthetic pigments in D. antarctica were measured at both treatments. The results obtained show that the Chl-a / Chl-b ratio in plants subjected to thermal treatments remained within the range (1.8–2.5) described for nonstressed plants (Figure 5A). Additionally, the content of carotenoid pigments in D. antarctica showed a significant increase on day 2 of treatment (47% at 23°C and 21% at 35°C) but decreased at the end of the experiment reaching basal values (P < 0.05) (Figure 5B).
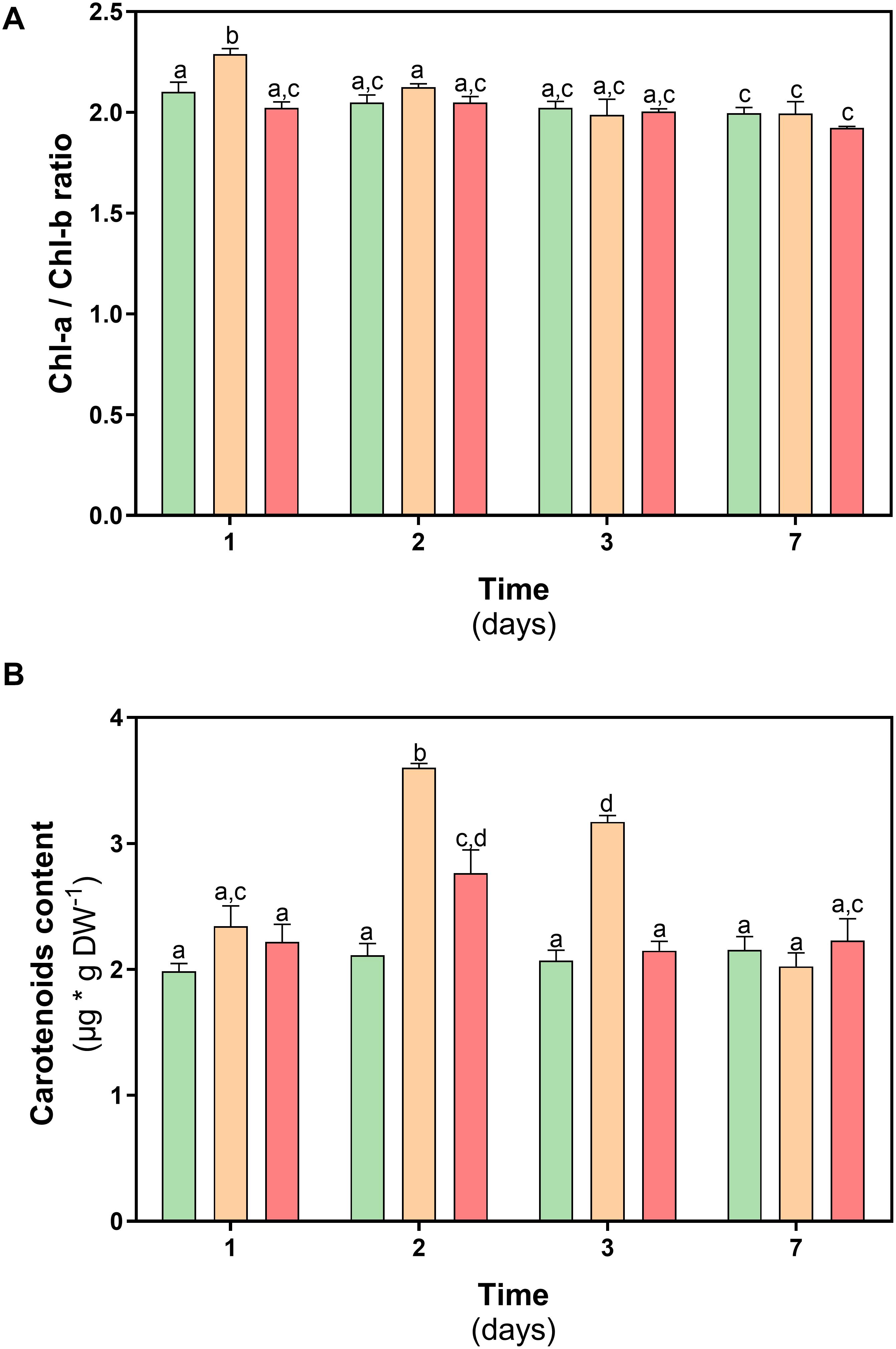
Figure 5. Photosynthetic pigments content in D. antarctica control (15°C) and heat shock treatments (23 and 35°C) cultivated in vitro. (A) chl-a/chl-b ratio and (B) carotenoids content. Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
Homeostasis Redox
One of the effects caused by the increase in temperature in plants is to alter redox homeostasis. For this reason, the levels of reactive oxygen species (ROS) and the levels of membrane lipoperoxidation (MDA) was evaluated in D. antarctica plants subjected to both thermal shock treatments. In plants treated at 23°C, an increase in ROS levels was observed on day 1 of treatment (25%). However, on day 7 of treatment, no significant differences were observed with respect to the control plants. In the plants treated at 35°C, a progressive decrease in ROS levels was observed during the treatment, reaching a 50% decrease on day 7 of treatment (P < 0.05) (Figure 6A). The MDA content in both treatments showed an increase on day 1, reaching values of 19 and 47% at 23 and 35°C, respectively. Then the MDA levels decreased over time in both treatments, reaching the levels of the control plants (Figure 6B).
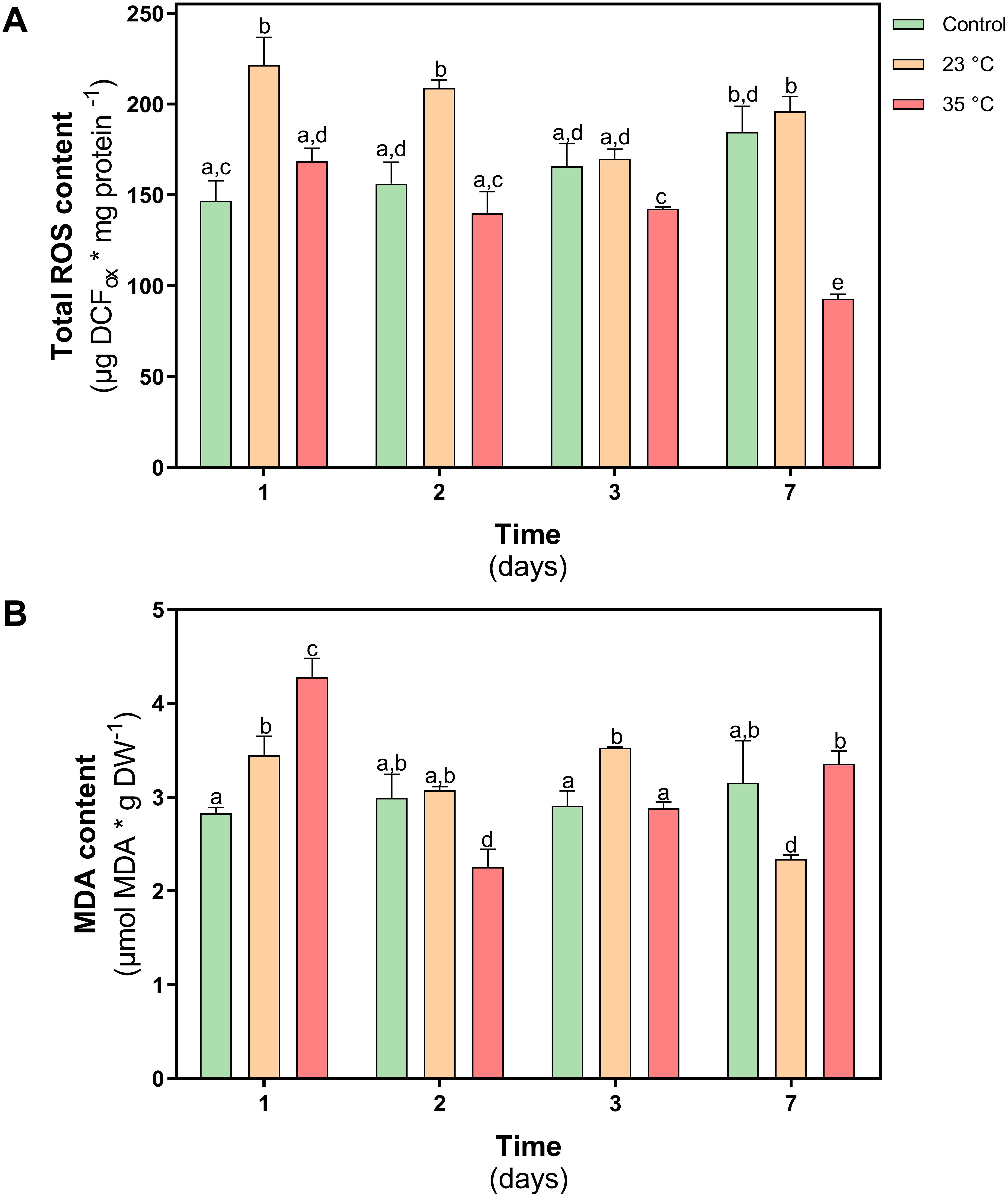
Figure 6. Oxidative stress parameters in D. antarctica control (15°C) and heat shock treatments (23°C and 35°C) cultivated in vitro. (A) Total ROS content and (B) membrane peroxidation (malondialdehyde content). Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
Enzymatic Antioxidant System
The activity of superoxide dismutase (SOD) did not show differences in their activity at day 1, while through time only at 35°C showed a decrease of 13% at day 7 respect to control and under 23°C treatment SOD had similar levels of activity (Figure 7A).
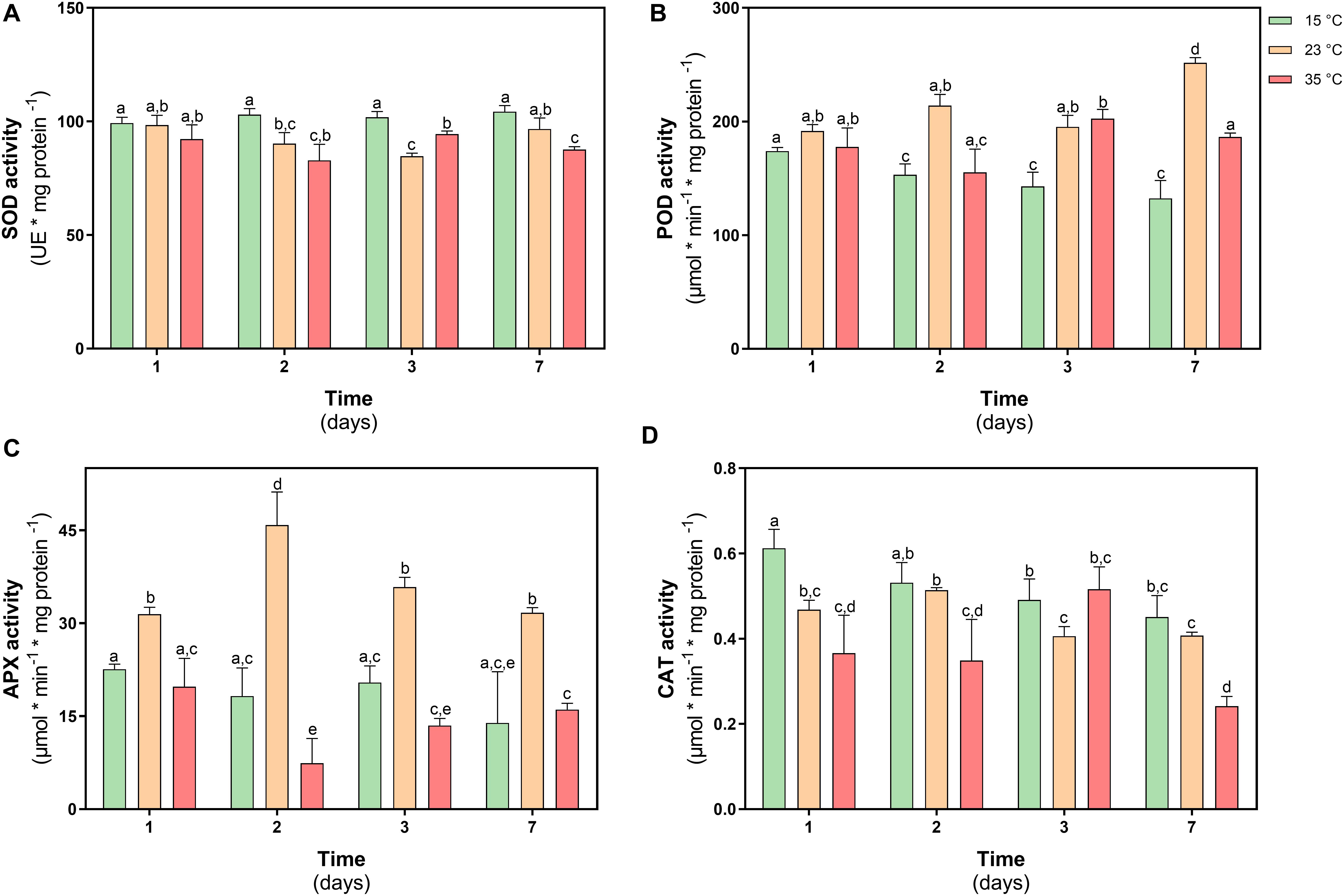
Figure 7. Antioxidant enzyme activity in D. antarctica control (15°C) and heat shock treatments (23°C and 35°C) cultivated in vitro. (A) superoxide dismutase (SOD) activity, (B) total peroxidases type III (POD) activity, (C) ascorbate peroxidase (APX) activity, and (D) catalase (CAT) activity. Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
The activity of type III peroxidases in D. antarctica (POD), showed an increase in their activity of 38% on day 7 in plants treated at 23°C, while in plants treated at 35°C an increase was only observed on day 3 treatment equivalent to 8% (Figure 7B). The activity of the APX enzyme only showed changes in the treatment at 23°C, increasing 33 and 60% on days 1 and 2 of treatment, respectively, with respect to the control plants (Figure 7C). The activity of the CAT enzyme showed a more erratic behavior during the treatment. This activity showed a decrease on days 1 and 2 with respect to the control plants (18 and 35%, respectively) (Figure 7D).
Non-enzymatic Antioxidant System
The content of total phenolic compounds was determined in both treatments. In D. antarctica plants subjected to 35°C, an increase of 10% was observed on day 1 with respect to the control, however, in plants treated at 23°C, a 45% decrease was observed on day 1. At day 7 of treatment, the values of the content of total phenolic compounds did not show differences with the levels of control plants (Figure 8A).
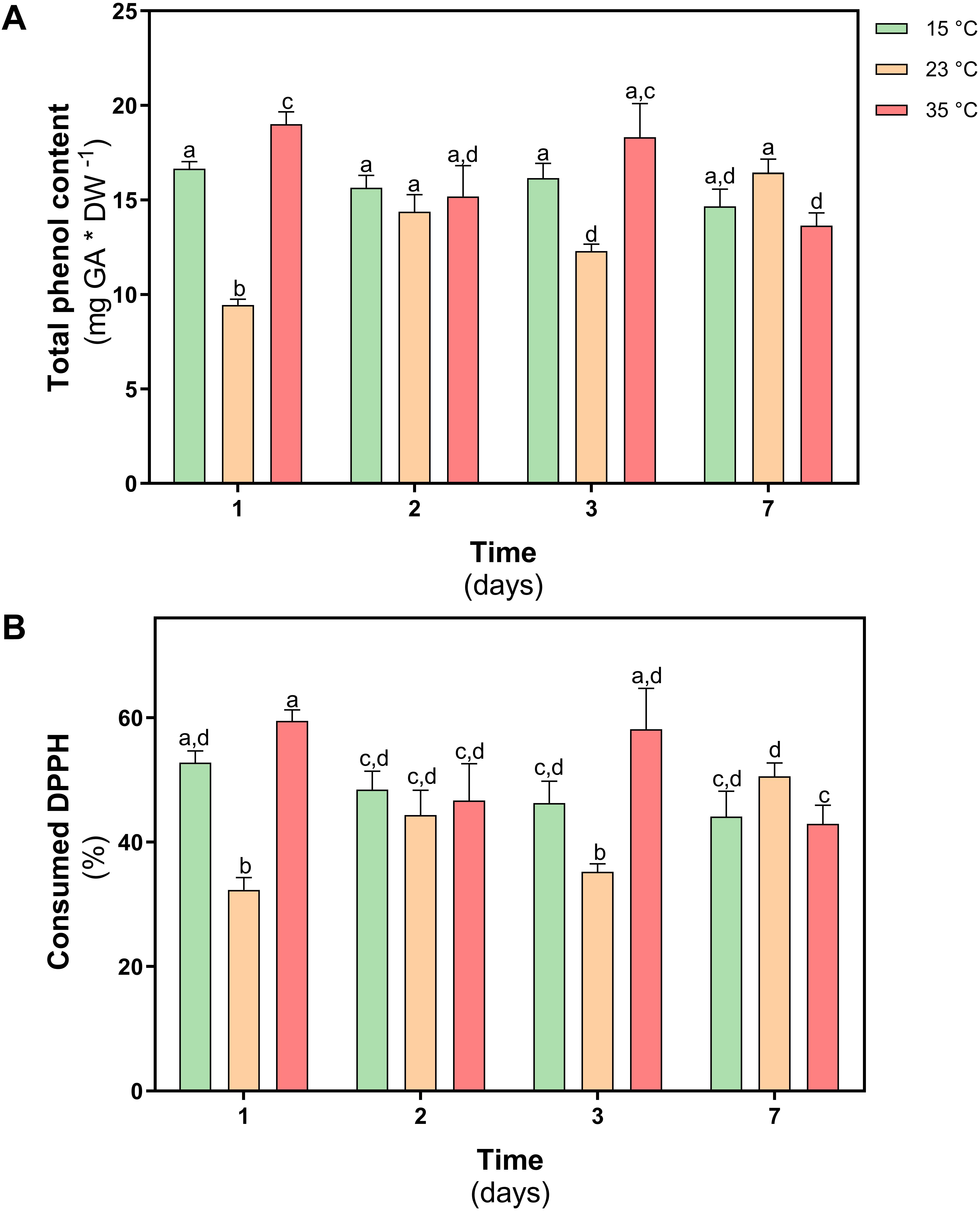
Figure 8. Non-enzymatic antioxidant activity in D. antarctica control (15°C) and heat shock treatments (23 and 35°C) cultivated in vitro. (A) total phenol content and (B) the consumption DPPH radical. Each bar represent means of 3 biological replicates (N = 3; ± standard error of the mean). Significant differences between treatments are indicated by letters (P < 0.05).
To determine the antioxidant role of phenolic compounds in D. antarctica, the scavenger capacity of the free radical DPPH was evaluated in ethanolic extracts obtained from treated and control plants. The scavenger capacity of the extract obtained in plants treated at 23°C, decreased by 35% on day 1 of treatment, but on day 7 it did not show differences with the control group (Figure 8B). Under the 35°C treatment, a slight increase of the antioxidant activity was observed on day 1, but no differences were observed at the end of the treatment on day 7.
Discussion
High temperature is an important abiotic stress factor for plants that is not restricted to tropical areas and desert belts but also may play an important role in colder regions like the Antarctica (Convey, 2006). The Antarctic Peninsula has experienced rapid warming in recent decades. The warming may in turn cause critical overheating as the heat trapping nature can get fatal in situations with high solar irradiation input, combined to restricted transpiration and calm winds causing situations where the thermal high temperature thresholds can get exceeded.
High temperature induces metabolic imbalances that can cause an oxidative stress in plant cells. This effect results in the generation and accumulation of ROS, promoting oxidation of cellular components, hindering metabolic activities and affecting organelle integrity (Suzuki et al., 2012).
In the present study, was assessed with the hypothesis that D. antarctica has efficient antioxidant protection mechanisms in response to heat shock treatment, being able to maintain normal Physiology, photosynthetic performance, and redox state balance.
The results have shown that D. antarctica has a high maximum heat tolerance of 52°C (LT50). Traditionally, the method used to determine the TL50 is based on the measurement of electron leakage (EL). The method used in this work is based on the measurement of the maximum photosynthetic efficiency (Fv/Fm). Determining chlorophyll fluorescence is a good alternative for EL measurement because of their noninvasive and rapid performance, as well as their potential for estimating LT50 Fv/Fm. This method is sensible, non-invasive, early, fast and easy to use (Ehlert and Hincha, 2008). Previously, Reyes et al. (2003), reported that D. antarctica plants, grown in growth chambers at 4 and 13°C, presented a LT50 of 48.3°C when analyzing the membrane damage. This value is similar to the one determined in this work, although the methods used are totally different.
The value of LT50 determined is surprisingly high for a species that has lived in a rather cold environment. Due to global warming that affects the Antarctic region and the increasing heat waves it can be suggested that this species has the potential to tolerate warm temperatures.
Global warming has led to an increase in soil temperature, major abiotic stress that poses a difficult threat to plants. Photosynthesis is among the plant cell functions that are very sensitive to stress at high temperatures and often interfere before other cell functions deteriorate. The major targets for high-temperature stress are Photosystem II (PSII), and oxygen evolution complex (OEC), and ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco) (Mathur et al., 2014). Previous studies have shown that the optimal photosynthetic temperature in D. antarctica is in a range between 13 and 19°C (Edwards and Smith, 1988; Xiong et al., 1999). Additionally, the carboxylase activity of Rubisco in D. antarctica has not shown differences at 5, 15, and 25°C (Sáez et al., 2017).
Related to the hypothesis of this work, the results show D. antarctica is able to tolerate a heat shock treatment of 23°C and 35°C. Elevated temperatures could also determine a loss in the content of photosynthetic pigments. It has been reported that due to the effect of heat, chlorophyllases and peroxidase are induced in plants (Rossi et al., 2017). However, in D. antarctica, the values of the chlorophyll a/b ratio remained in the range 1.8–2.5, which is characteristic of nonstressed plants (Yu et al., 2015). The null effect observed in the photosynthetic performance which means that the thylakoid membranes have maintained their functionality, without showing damage by lipid oxidation (lipoperoxidation), due to the accumulation of ROS. Photosynthetic activity at 35°C has not been previously reported in D. antarctica and shows its plasticity under unfavorable conditions such as those that are affecting the Antarctic region.
A key factor in tolerance to thermal stress is the ability of plants to rapidly control oxidative damage (Tognetti et al., 2012). The PSII complex is one of the most thermolabile components, mainly the D1 protein due to ROS attack (Wang et al., 2018). The results of this work show D. antarctica maintained a normal functioning of their photosynthetic machinery to both shock treatments. This was due at least in part to the efficient enzymatic antioxidant activity, which allowed it to control H2O2 levels, described as a potent inhibitor of photosynthesis in plants due de oxidation of the core components of PSII, like protein D1, and for the oxidation of other biomolecules like lipids, proteins among others (Foyer and Shigeoka, 2011). APX is one of the key enzymes to protect photosynthetic activity because it is located inside the chloroplasts that detoxify the H2O2 produced by SOD activity using ascorbate as a substrate (Roach and Krieger-Liszkay, 2014). Our results show an induction of APX activity at 23°C but not at 35°C. This is important, since it shows that this enzyme is activated only at certain stress thresholds.
Since there was no damage to D. antarctica in both treatments, it can be suggested that other mechanisms operate at 35°C instead of APX activity, which would act at 23°C. At 23°C during the first days of treatment, an increase in carotenoids levels is observed, a situation that does not occur at 35°C. Since the carotenoid level showed an increase in the first days of treatment at 23°C, its role may be related to the photoprotective properties. Its antioxidant activity (Nizar et al., 2015), alpha, beta carotenoids, and xanthophylls are agents stabilizers of the thylakoid membrane and reaction centers of the photosystems; these molecules are essential to prevent the leakage of ions from the tissues and act as antioxidants when they are free in the cell, protecting the membranes of the thylakoids from oxidation by the ROS attack (Latowski et al., 2011).
Another enzyme that showed significant activity at 23°C is POD. This enzyme has previously been reported to play an important role in the tolerance of D. antarctica to osmotic stress (Zamora et al., 2010) and UV-B radiation (Köhler et al., 2017). In addition to their antioxidant function, some apoplastic POD isoforms are related to the thickening of the cell wall and the polymerization of lignin (De Gara, 2004). Therefore, POD activity also contributes to maintaining low ROS levels, membrane integrity, and metabolic functionality under heat shock conditions. Gielwanoska et al., 2005, described that in D. antarctica, adverse environmental conditions stimulate growth, altering the lignification of the cell wall. In a field study subjecting D. antarctica plants to a passive heating treatment, it was shown that after the treatment the plants showed a higher fiber content, mainly cellulose, and higher sclerophylly index (Sáez et al., 2018). The catalase activity did not show significant changes in the heat shock treatment. This could be explained by its low affinity to H2O2 compared to peroxidases, which are present in most organelles (Zamora et al., 2010), and increases in CAT activity are also related to the photorespiration process in plants (Foyer et al., 2009).
As previously mentioned, the activity of the APX and POD enzymes was sufficient to maintain the oxidative state in D. antarctica in the treatment at 23°C. On the other hand, at 35°C these enzymes were not induced, and no damage was observed, that implies another control mechanism must be operating against ROS accumulation. In this sense, an increase in the content of phenolic compounds and an increase of the free radical trapping activity were shown. A severe heat shock of 35°C activates signaling cascades based on lipid and Ca+2 inputs that regulate gene expression (Saidi et al., 2011). Among these genes, those involved with certain secondary metabolites related to the acquisition of thermotolerance in plants will be activated (Bita and Gerats, 2013). The chemical structure of phenolic compounds allows them to act as free-radical species trappers through two mechanisms, by electron transfer (SET, Single Electron Transfer) or hydrogen transfer (HAT, Hydrogen Atom Transfer) (Ahmad et al., 2010). For this reason, the accumulation of phenolic compounds within chloroplasts contributes to the detoxification of ROS and free radicals, maintaining the functionality of the chloroplast (Zhao and Zou, 2002), which would explain the low APX activity in D. antarctica at 35°C. compared to treatment at 23°C.
The efficient control of ROS in plants subjected to heat shock resulted in the control of MDA levels and in the maintenance of photosynthesis under conditions of thermal stress (Chen et al., 2010). Chlorophyll stability has been proposed to be a key characteristic in maize varieties classified as thermotolerant (Shirdelmoghanloo et al., 2016). The definition of homochlorophilic plants has been proposed to those species that have the capacity to maintain the integrity of photosystems and the integrity of chlorophyll under conditions of oxidative imbalance, induced by heat shock (Tuba and Lichtenthaler, 2011). In this work, we demonstrated that D. antarctica achieved maintaining all essential physiological parameters despite heat shock treatment with similar features like thermotolerant species (Vierling, 1991; Nagarajan et al., 2010; Scafaro et al., 2010;.). Photosynthetic activity at 35°C has not been previously reported in D. antarctica and shows its plasticity under unfavorable conditions such as those that are affecting the Antarctic region.
The results allow us to suggest that tolerance to a shock of 35°C is strongly influenced by the accumulation of phenolic compounds. The ability of the plant to accumulate phenolic compounds is considered an evolutionary advantage, which allows it to tolerate adverse environmental conditions (Edreva et al., 2008). It has been reported that under abiotic stress conditions, an activation of the transcription of key enzymes of the phenylpropanoid pathway such as phenylalanine ammonia lyase (PAL), chalcone synthase (CHS), chalcone isomerase (CHI) and flavonol synthase (FLS) are induced (Sharma et al., 2019).
Conclusion
The results shown in this work allow us to indicate that D. antarctica plants grown in vitro are capable of tolerating thermal shock conditions of 23 and 35°C. These temperatures are 8 and 20°C higher than the temperature at which the plants are grown in the growth chambers. The tolerance shown is related to efficient enzymatic and non-enzymatic antioxidant systems, which facilitate proper cell function by controlling ROS levels.
This response of D. antarctica to heat shock could be key to its success against the warming that affects the Antarctic continent.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
RC-A designed and conducted all experiments in the laboratory at University of Santiago and wrote the first draft of the manuscript. MP, RC, and HK collaborated in some analysis. MP, RC, and GZ collected samples in Antarctica. All authors contributed equally to the discussion and revision of the final version of the manuscript.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding
This work has been supported by fellowship ANID-PFCHA/Doctoral scholarship/2019 N°21191483 (RC-A), Chilean Antarctic Institute (INACh) RT_14-17 project (GZ), and Proyecto Basal CEDENNA AFB180001 (GZ). The financial support of DICYT, University of Santiago de Chile is also appreciated.
Acknowledgments
We appreciate the logistical support provided by the staff from the Professor Julio Escudero Station (INACH), King George Island. We also wish to give special thanks to Elizabeth Barria and Cristina Vargas for all their support in the in vitro culture of D. antarctica.
References
Ahmad, P., Jaleel, C. A., Salem, M. A., Nabi, G., and Sharma, S. (2010). Roles of enzymatic and noenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 30, 161–175. doi: 10.3109/07388550903524243
Asami, D., Hong, Y., Barrett, D., and Mitchell, A. (2003). Comparison of the total phenolic and ascorbic acid content of freeze-dried and air-dried conventional, organic, and sustainable agricultural practices. J. Agricult. Food Chem. 51, 1237–1241.
Beauchamp, C., and Fridovich, I. (1971). Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 44, 276–287.
Bita, C. E., and Gerats, T. (2013). Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 4:273. doi: 10.3389/fpls.2013.00273
Bradford, M. M. (1976). A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 72, 248–254. doi: 10.1006/abio.1976.9999
Brand-Williams, W., Cuvelier, M. E., and Berset, C. (1995). Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 28, 25–30.
Cavieres, L. A., Sáez, P., Sanhueza, C., Sierra-Almeida, A., Rabert, C., Corcuera, L. J., et al. (2016). Ecophysiological traits of Antarctic vascular plants: the importance in the responses to climate change. Plant Ecol. 217, 343–358. doi: 10.1007/s11258-016-0585-x
Chen, J., Wang, P., Mi, H. L., Chen, G. Y., and Xu, D. Q. (2010). Reversible association of ribulose-1,5-biphosphate carboxylase/oxygenase activase with thylakoid membrane depends upon the ATP level and pH in rice without heat stress. J. Exp. Bot. 61, 2939–2950. doi: 10.1093/jxb/erq122
Contreras, R. A., Köhler, H., Pizarro, M., and Zúñiga, G. E. (2015). In vitro cultivars of Vaccinium corymbosum L. (Ericaceae) are a source of antioxidant phenolics. Antioxidants 4, 281–292.
Convey, P. (2006). Antarctic terrestrial ecosystems: responses to environmental changes. Polarforsch 75, 101–111.
Convey, P. (2013). “Antarctic ecosystems,” in Encyclopedia of Biodiversity, 2nd Edn, eds S. A. Levin and S. A. Levin (San Diego, CA: Elsevier), 179–188.
De Gara, L. (2004). Class III peroxidases and ascorbate metabolism in plants. Phytochem. Rev. 3, 195–205. doi: 10.1023/B:PHYT.0000047795.82713.99
Driendoks, N., Xu, J., Peters, J. L., Park, S., and Rieu, I. (2015). Multi-level interactions between heat shock factors, heat shock proteins, and the redox system regulate acclimation to heat. Front. Plant Sci. 6:999. doi: 10.3389/fpls.2015.00999
Ederli, L., Reale, L., Ferranti, F., and Pasqualini, S. (2004). Responses induced by high concentration of cadmium in Phragmites australis roots. Physiol. Plant 121, 66–74. doi: 10.1111/j.0031-9317.2004.00295.x
Edreva, A., Velikova, V., Tsonev, T., Dagnon, S., Gürel, A., Aktas, L., et al. (2008). Stress-protective role of secondary metabolites: diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 34, 67–78.
Edwards, J. A., and Smith, R. I. L. (1988). Photosynthesis and respiration of Colobanthus quitensis and Deschampsia antarctica from the Maritime Antarctic. Br. Antarct. Surv. B 81, 43–63.
Ehlert, B., and Hincha, D. (2008). Chlorophyll fluorescence imaging accurately quantifies freezing damage and cold acclimation responses in Arabidopsis leaves. Plant Methods 4:12. doi: 10.1186/1746-4811-4-12
Fowbert, J. A., and Smith, R. I. L. (1994). Rapid population increases in native vascular plants in the Argentine Island, Antarctic Peninsula. Arct. Antarct. Alp. Res. 26, 290–296. doi: 10.2307/1551941
Foyer, C. H., Bloom, A. J., Queval, G., and Noctor, G. (2009). Photorespiratory metabolism: genes, mutants, energetics, and redox signaling. Annu. Rev. Plant Biol. 60, 455–484. doi: 10.1146/annurev.arplant.043008.091948
Foyer, C. H., and Shigeoka, S. (2011). Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 155, 93–100. doi: 10.1104/pp.110.166181
Gielwanoska, I., and Szczuka, E. (2005). New ultrastructural features of organelles in leaf cells of Deschampsia antarctica. Desv. Polar Biol. 28, 951–955. doi: 10.1007/s00300-005-0024-2
Gielwanoska, I., Szczuka, E., Bednara, J., and Górecki, R. (2005). Anatomical features and ultrastructure of Deschampsia antarctica (Poaceae) leaves from different growing habitats. Ann. Bot. 96, 1109–1119. doi: 10.1093/aob/mci262
Köhler, H., Contreras, R. A., Pizarro, M., Cortés-Antiquera, R., and Zúñiga, G. E. (2017). Antioxidant responses induced by UVB radiation in Deschampsia antarctica Desv. Front. Plant Sci. 8:921. doi: 10.3389/fpls.2017.00921
Latowski, D., Kuczynska, P., and Strzalka, K. (2011). Xantohpyll cycle a mechanism protecting plants against oxidative stress. Redox Rep. 16, 78–90. doi: 10.1179/174329211X13020951739938
Lee, J., Noh, E. K., Choi, H. S., Shin, S. C., Park, H., and Lee, H. (2012). Transcriptome sequencing of the Antarctic vascular plant Deschampsia antarctica Desv. under abiotic stress. Planta 237, 823–836. doi: 10.1007/s00425-012-1797-5
Lima, A. L. S., DaMatta, F. M., Pinheiro, H. A., Totola, M. R., and Loureiro, M. E. (2002). Photochemical responses and oxidative stress in twi clones of Coffea canephora under water deficit conditions. Environ. Exp. Bot. 47, 239–247. doi: 10.1016/S0098-8472(01)00130-7
Mantovani, A., and Vieira, R. C. (2000). Leaf micromorphology of Antarctic pearwort Colobanthus quitensis (Kuntz) Bartl. Polar Biol. 28, 531–538. doi: 10.1007/s003000000117
Mathur, S., Agrawal, D., and Jajoo, A. (2014). Photosynthesis: response to high temperature stress. J. Photochem. Photobiol. B 137, 116–126. doi: 10.1016/j.jphotobiol.2014.01.010
Montiel, P., Smith, A., and Keller, D. (1999). Photosynthetic responses of selected Antarctic plants to solar radiation in the southern maritime Antartic. Polar Res. 18, 229–235. doi: 10.1111/j.1751-8369.1999.tb00298
Nagarajan, S., Jagadish, S. V. K., Prasad, A. S. H., Thomar, A. K., Anand, A., Pal, M., et al. (2010). Local climate affects growth, yield and grain quality of aromatic and non-aromatic rice in northwestern India. Agric. Ecosyst. Environ. 138, 274–281. doi: 10.1016/j.agee.2010.05.012
Naik, G. H., Priyandarsini, K. I., and Mohan, H. (2005). Evaluating the antioxidant activity of different plant extracts and herbal formulations. Res. Chem. Intermediat. 31, 145–151. doi: 10.1163/1568567053146823
Nizar, N., Li, L., Lu, S., Khin, N. C., and Pogson, B. J. (2015). Carotenoid metabolism in plants. Mol. Plant 8, 68–82. doi: 10.1016/j.molp.2014.12.007
Parnikoza, I., Kozerestska, I., and Kunakh, V. (2011). Vascular plants of the maritime Antarctic: origin and adaptation. Am. J. Plant Sci. 2, 381–395. doi: 10.4236/ajps.2011.23044
Pinhero, R., Rao, M., Paliyath, G., Murr, D., and Fletcher, R. (1997). Changes in activities of antioxidant enzymes and their relationship to genetic and paclobutrazol-induced chilling tolerance of maize seedling. Plant Physiol. 114, 695–704.
Reyes, M. A., Corcuera, L. J., and Cardemil, L. (2003). Accumulation of Hsp70 in Deschampsia antartica Desv leaves under thermal stress. Antarct. Sci. 15, 342–352. doi: 10.1017/S0954102003001366
Roach, T., and Krieger-Liszkay, A. (2014). Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 15, 351–362. doi: 10.2174/1389203715666140327105143
Robinson, S., Klekociuk, A. R., King, D. H., Pizarro, M., Zúñiga, G. E., and Bergstrom, D. M. (2020). The 2019/2020 summer of Antarctic heatwaves. Glob. Change Biol. 26, 3178–3180. doi: 10.1111/gcb.15083
Robinson, S. A., Wasley, J., and Tobin, A. K. (2003). Living on the edge – plants and global change in continental and maritime Antarctica. Glob. Change Biol. 9, 1681–1717. doi: 10.1046/j.1365-2486.2003.00693.x
Romero, M., Casanova, A., Iturra, G., Reyes, A., Montenegro, G., and Alberdi, M. (1999). Lead anatomy of Deschampsia antarctica (Poaceae) from the Maritime Antarctic and its plastic response to changes in growth conditions. Rev. Chil. Hist. Nat. 72, 411–425.
Ross, C., Puglisi, M. P., and Paul, V. J. (2008). Antifungal defenses of seagrasses from the Indian River Lagoon. Florida. Aquat. Bot. 88, 134–141. doi: 10.1016/j.aquabot.2007.09.003
Rossi, S., Burgess, P., Jerpersen, D., and Huang, B. (2017). Heat-induced senescence associated with chlorophyll metabolism in Bentgrass lines differing in heat tolerance. Crop. Sci. 57, S169–S178. doi: 10.2135/cropsci2016.06.0542
Sáez, P., Bravo, L., Cavieres, L., Vallejos, V., Sanhueza, C., Font-Carrascosa, M., et al. (2017). Photosynthetic limitations in two Antarctic vascular plants: importance of leaf anatomical traits and Rubisco kinetic parameters. J. Exp. Bot. 68, 2871–2883. doi: 10.1093/jxb/erx148
Sáez, P., Cavieres, L. A., Galmés, J., Gil-Pelegrin, E., Peguero-Pina, J. J., Sancho-Knapik, D., et al. (2018). In situ warming in the Antarctic: effects on growth and photosynthesis in Antarctic vascular plants. New Phytol. 218, 1406–1418. doi: 10.1111/nph.15124
Saidi, Y., Finka, A., and Goloubinoff, P. (2011). Heat perception and signalling in plants: a tortuous path to thermotolerance. New Phytol. 190, 556–565. doi: 10.1111/j.1469-8137.2010.03571.x
Scafaro, A. P., Haynes, P. A., and Atwell, B. J. (2010). Physiological and molecular changes in Oryza meridionalis Ng., a heat-tolerant species of wild rice. J. Exp. Bot. 61, 191–202. doi: 10.1093/jxb/erp294
Sequeida, A., Tapia, E., Ortega, M., Zamora, P., Castro, A., Montes, C., et al. (2012). Production of phenolic metabolites by Deschampsia antarctica shoots using UV-B treatments during cultivation in a photobiorector. Electron. J. Biotech 15:8. doi: 10.2225/vol15-issue4-fulltext-7
Sharma, A., Shahzad, B., Rehman, A., Bhardwaj, R., Landi, M., and Zheng, B. (2019). Response of phenylpropanoid pathway and the role of polyphenols in plant under abiotic stress. Molecules 24:2452. doi: 10.3390/molecules24132452
Shirdelmoghanloo, H., Cozzolino, D., Lohrased, I., and Collins, N. C. (2016). Truncation of grain filling in wheat (Triticum aestivum) triggered by brief heat stress during early grain filling: association with senescence responses and reductions in steam reserves. Funct. Plant Biol. 43, 919–930. doi: 10.1071/FP15384
Smith, R. I. L. (2003). “The enigma of Colobanthus quitensis and Deschampsia antarctica in Antarctica,” in Antarctic Biology in a Global Context, eds A. H. L. Huiskes, W. W. C. Gieskes, J. Rozema, R. M. L. Schorno, S. M. van der Vies, and W. J. Wolff (Leiden: Backhuys Publishers), 234–239.
Stirbet, A., and Govindjee. (2011). On the relation between the Kautsky effect (chlorophyll a fluorescence induction) and Photosystem II: basics and applications of the OJIP fluorescence transient. J. Photochem. Photobiol. B 104, 236–257.
Suzuki, N., Koussevitzky, S., Mittler, R., and Miller, G. (2012). ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 35, 259–270. doi: 10.1111/j.1365-3040.2011.02336.x
Tognetti, V. B., Mühlenbock, P. E. R., and Van Breusegem, F. (2012). Stress homeostasis – the redox and auxin perspective. Plant Cell Environ. 35, 321–333. doi: 10.1111/j.1365-3040.2011.02324.x
Torres-Mellado, G. A., Jaña, R., and Casanova-Katny, M. A. (2011). Antarctic hairgrass expansion in the South Shetland archipelago and Antarctic Peninsula revisited. Polar. Biol. 34, 1679–1688. doi: 10.1007/s00300-011-1099-6
Tuba, Z., and Lichtenthaler, H. K. (2011). “Ecophysiology of homoiochlorophyllous and poikilochlorophyllous desiccation-tolerant plants and vegetations,” in Plant Desiccation Tolerance, Ecological Studies, Vol. 215, eds U. Luttge, E. Beck, and D. Bartels (Heidelberg: Springer), 157–183. doi: 10.1007/978-3-642-19106-0_9
Turner, J., Hosking, J., Scott, P., and Marshall, G. J. (2013). Temporal and spatial evolution of the Antarctic sea ice prior to the September 2012 record maximum extent. Geophys. Res. Lett. 40, 5894–5898. doi: 10.1002/2013GL058371
Turner, J., and Overland, J. (2009). Contrasting climate change in the two polar regions. Polar Res. 28, 146–164. doi: 10.1111/j.1751-8369.2009.00128.x
Vera, M. L., Fernández-Teruel, T., and Quesada, A. (2013). Distribution and reproductive capacity of Deschampsia antarctica and Colobanthus quitensis on Byers Peninsula, Livingston Island, South Shetland Islands, Antarctica. Antarct. Sci. 25, 292–302. doi: 10.1017/S0954102012000995
Vierling, E. (1991). The roles of heat shock proteins in plants. Annu. Rev. Plant Biol. 42, 579–620. doi: 10.1146/annurev.pp.42.060191.003051
Wahid, A., Gelani, S., Asharf, M., and Foolad, M. (2007). Heat tolerance in plants: an overview. Environ. Exp. Bot. 61, 199–223. doi: 10.1016/j.envexpbot.2007.05.011
Wang, Q. L., Chen, J. Q., He, N. Y., and Guo, F. Q. (2018). Metabolic reprogramming in chloroplasts under heat stress in plants. Int. J. Mol. Sci. 19:849. doi: 10.3390/ijms19030849
Wellburn, A. R., and Lichtenthaler, H. (1984). “Formulae and program to determine total carotenoids and chlorophylls A and B of leaf extracts in different solvents,” in Advances in Photosynthesis Research. Advances in Agricultural Biotechnology, Vol. 2, ed. C. Sybesma (Dordrecht: Springer).
Xiong, F. S., Ruhland, C. T., and Day, T. A. (1999). Photosynthetic temperature response of the Antarctic vascular plants Colobanthus quitensis and Deschampsia antarctica. Physiol. Plant. 106, 276–286. doi: 10.1034/j.1399-3054.1999.106304.x
Yu, K., Gnyp, M. L., Gao, L., and Miao, Y. (2015). Estimate leaf chlorophyll of rice using reflectance indices and partial least squares. Photogr. Fernerk, Geoinform. 10, 45–54. doi: 10.1127/pfg/2015/0253
Zamora, P., Rasmussen, S., Pardo, A., Prieto, H., and Zúñiga, G. E. (2010). Antioxidant responses of in vitro shoots of Deschampsia antarctica to polyethylene glycol treatment. Antartc. Sci. 22, 163–169. doi: 10.1017/S0954102009990733
Keywords: Antarctica, climate change, oxidative stress–related enzymes, peroxidases, photosynthesis
Citation: Cortés-Antiquera R, Pizarro M, Contreras RA, Köhler H and Zúñiga GE (2021) Heat Shock Tolerance in Deschampsia antarctica Desv. Cultivated in vitro Is Mediated by Enzymatic and Non-enzymatic Antioxidants. Front. Plant Sci. 12:635491. doi: 10.3389/fpls.2021.635491
Received: 30 November 2020; Accepted: 01 February 2021;
Published: 23 February 2021.
Edited by:
Heng Zhang, Shanghai Center for Plant Stress Biology, Shanghai Institute for Biological Sciences, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Hyoungseok Lee, Korea Polar Research Institute, South KoreaAntonio Batista Pereira, Universidade Federal do Pampa - UNIPAMPA, Brazil
Copyright © 2021 Cortés-Antiquera, Pizarro, Contreras, Köhler and Zúñiga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gustavo E. Zúñiga, Z3VzdGF2by56dW5pZ2FAdXNhY2guY2w=