 Raúl Ortells-Fabra1†
Raúl Ortells-Fabra1† Carolina Gallego-Giraldo1†
Carolina Gallego-Giraldo1† Maria Angeles Forner-Giner2†
Maria Angeles Forner-Giner2† Alberto Urbaneja1†
Alberto Urbaneja1† Meritxell Pérez-Hedo3*†
Meritxell Pérez-Hedo3*†- 1Instituto Valenciano de Investigaciones Agrarias (IVIA), Centro de Protección Vegetal y Biotecnología, Unidad de Entomología, Moncada, Spain
- 2Instituto Valenciano de Investigaciones Agrarias (IVIA), Centro de Citricultura y Producción Vegetal, Moncada, Spain
- 3Instituto de Biología Molecular y Celular de Plantas (IBMCP), Consejo Superior de Investigaciones Científicas, Universitat Politècnica de València, Valencia, Spain
Herbivore-induced plant volatiles (HIPVs) are known to activate immune signaling in plants; however, their effectiveness can vary depending on the genotype and the signaling pathway involved. In this study, we evaluated the transcriptional response of four citrus rootstocks (Carrizo citrange, Forner-Alcaide 5 (FA5), Forner-Alcaide 74 (FA74), and Microcitrus australasica) to six synthetic HIPVs [(Z)-3-hexen-1-ol, (Z)-3-hexenyl acetate, (Z)-3-hexenyl butyrate, (Z)-3-hexenyl propanoate, methyl jasmonate, and methyl salicylate]. We focused on genes associated with the salicylic acid (SA) and jasmonic acid (JA) pathways, as well as the susceptibility gene CsPUB21. Overall, the SA pathway was more consistently activated than the JA pathway, with upstream and intermediate genes induced across most genotypes and treatments. In contrast, downstream markers showed more variable expression, suggesting that synthetic HIPVs may induce a primed rather than fully activated defense state. Among the volatiles tested, (Z)-3-hexenyl propanoate and (Z)-3-hexen-1-ol were the most effective, activating genes in both pathways. Importantly, these two compounds also consistently repressed CsPUB21 expression, a gene recently associated with huanglongbing (HLB) susceptibility, through coordinated transcriptional and post-translational regulation. Carrizo citrange showed the strongest transcriptional response, while FA74 exhibited more moderate activation, emphasizing the influence of genetic background on HIPV perception and signaling. These findings highlight the potential of selected synthetic HIPVs as sustainable defense priming agents capable of enhancing citrus immunity by simultaneously activating immune pathways and repressing susceptibility genes such as CsPUB21. This dual mode of action offers promising tools for the integrated management of HLB and other citrus diseases.
1 Introduction
Plants and herbivorous insects are engaged in a continuous evolutionary battle in which plants have evolved sophisticated defense strategies to mitigate herbivory (Kessler and Baldwin, 2002; Howe and Jander, 2008; War et al., 2012). Among these defenses, the production of herbivore-induced plant volatiles (HIPVs) and oviposition-induce plant volatiles (OIPVs) which represent reliable cues associated with presence of host eggs laid on plant tissues, a subset of volatile organic compounds (VOCs), plays a pivotal role in plant immunity (Dicke, 2009; Dicke and Baldwin, 2010). These airborne chemical signals serve dual functions in plant defense: they attract natural enemies of the herbivores (indirect defense) and act as inter-plant alarm signals. Upon detecting HIPVs, neighboring plants activate immune responses, particularly through the jasmonic acid (JA) and salicylic acid (SA) signaling pathways, which orchestrate the expression of defense genes (Turlings and Erb, 2018). This ability of plants to “eavesdrop” on the distress signals of their neighbors underscores the ecological significance of HIPVs in plant-insect interactions and provides a basis for innovative pest management strategies (Frost et al., 2008; Turlings and Erb, 2018).
Recent studies have demonstrated that the exogenous application of specific HIPVs can prime or induce defense responses in various plant species, thereby enhancing their resistance to pests and pathogens in an environmentally friendly manner. For instance, priming seeds with the indole volatile (emitted by maize during herbivory) boosted the resistance of Arabidopsis thaliana and Medicago truncatula plants against the beet armyworm Spodoptera exigua (Fab.) (Lepidoptera: Noctuidae) and the pea aphid Acyrthosiphon pisum (Harris) (Hemiptera: Aphididae), without compromising plant growth (Maurya et al., 2022). In maize, exposure to green leaf volatiles such as (Z)-3-hexenyl acetate is known to prime stronger anti-herbivore defenses upon subsequent attack (Engelberth et al., 2004). In tomato, foliar application of (Z)-3-hexen-1-ol was shown to induce both JA- and SA-mediated defenses directly, improving the plant’s resistance to the tobacco whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) while also increasing the emission of volatiles that attract whitefly parasitoids (Yang et al., 2020). Similarly, treating tomato plants with (Z)-3-hexenyl butyrate triggers early defense signaling events, including Ca2+ influx, activation of mitogen-activated protein kinases, and a rapid burst of reactive oxygen species, ultimately leading to defense responses such as stomatal closure (López-Gresa et al., 2018). Notably, the induction of these defenses by (Z)-3-hexenyl butyrate has been validated, resulting in enhanced resistance of crops to infections by the potato late blight disease, Phytophthora infestans, in potato and the rod-shaped, gram-negative bacterium Pseudomonas syringae in tomato (Payá et al., 2024).
Building on this concept, herbivore-induced volatiles are being explored as practical tools in agriculture. The use of slow-release dispensers emitting synthetic HIPVs has yielded promising results in pest control. For example, the green leaf volatile (Z)-3-hexenyl propanoate released in greenhouses significantly reduced infestations of key pests, such as the South American pinworm Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) in tomato (Pérez-Hedo et al., 2021a) and the foxglove aphid Aulacorthum solani (Kaltenbach) (Hemiptera: Aphididae) in sweet pepper (Depalo et al., 2022). (Z)-3-hexenyl propanoate-exposed tomato crops induced overexpression of anti-herbivore defense genes, indicating activation of the jasmonate pathway and accumulation of defense metabolites in the plant (Pérez-Hedo et al., 2021a). Beyond annual crops, volatile defense elicitors have also been tested in perennial systems; for instance, exogenous application of methyl jasmonate (a naturally occurring VOC) in grapevine triggered the production of pathogenesis-related proteins and phytoalexins, resulting in increased resistance against powdery mildew in the vineyard (Belhadj et al., 2006). Taken together, these examples demonstrate the potential of synthetic HIPVs to enhance plant immunity and contribute to sustainable, integrated pest management, thereby reducing reliance on synthetic pesticides and their associated risks.
Citrus crops, which are of enormous global economic importance, are under intense pressure from a variety of pests and diseases that can severely impact yield and fruit quality (Urbaneja et al., 2020). Traditionally, citrus pest management has relied on chemical insecticides, a practice that provides short-term control but entails environmental and human health risks (Tudi et al., 2021). This has created an urgent need for sustainable alternatives. Inducing citrus defenses via HIPVs represents a novel approach to this challenge. To date, however, citrus plants have been largely overlooked in studies of VOC-induced resistance, possibly due to a historical focus on model plants (e.g., Arabidopsis and tomato) and the complexity of citrus biology, including its perennial growth and diverse genetics in rootstocks. Only recently has evidence emerged that citrus could respond to airborne defense cues. Recently, Pérez-Hedo et al. (2024a) demonstrated that exposing the rootstock Carrizo citrange [a hybrid of Citrus sinensis (L.) Osb. × Poncirus trifoliata (L.) Raf.] to (Z)-3-hexenyl propanoate activates their immune machinery. Exposed citrus plants showed strong upregulation of defense-related genes associated with both the SA and JA pathways, and this molecular response was manifested in reduced performance of the South African mealybug Delottococcus aberiae (De Lotto) (Hemiptera: Pseudococcidae) and the two-spotted mite Tetranychus urticae Koch (Acari: Tetranychidae) and increased attraction of those pests’ natural enemies (Pérez-Hedo et al., 2024a). This finding highlights the feasibility of enhancing citrus resistance through external volatile cues, opening the door to more ecologically based pest management strategies in orchards.
Despite this advance, it remains unknown how different synthetic HIPV compounds compare in their ability to induce defense responses in citrus, or whether such induction is consistent across different citrus genotypes. In the present study, we addressed this gap by investigating the effects of six HIPVs [(Z)-3-hexen-1-ol, (Z)-3-hexenyl acetate, (Z)-3-hexenyl butyrate, (Z)-3-hexenyl propanoate, methyl jasmonate, and methyl salicylate] on the expression of eight defense-related genes in four citrus rootstock species (Forner-Alcaide 5 (C. reshni Hort. Ex Tan. x P. trifoliata), Forner-Alcaide 74 (C. reshni x P. trifoliata), Carrizo citrange, and Microcitrus australasica). To capture the hierarchical structure of the defense signaling, we selected genes representing different functional levels within the SA and JA signaling pathways: two genes acting upstream in biosynthesis, one intermediate regulatory component, and one downstream marker of pathway activation. This approach enables us to evaluate whether synthetic HIPVs impact distinct stages of signaling and defense activation across different genotypes.
In addition, we incorporated the susceptibility gene CsPUB21 into our analysis. This gene encodes a U-box E3 ubiquitin ligase involved in the degradation of the transcription factor MYC2, a central regulator of JA-mediated defense responses. Recent studies have shown that CsPUB21 expression correlates positively with susceptibility to huanglongbing (HLB) (Zhao et al., 2025), the most devastating disease in citrus (Pérez-Hedo et al., 2025); and that its downregulation enhances MYC2 stability and resistance to infection. Given the known induction of MYC2 by synthetic HIPVs (Pérez-Hedo et al., 2024a), we explored whether volatile exposure could also affect CsPUB21 expression. This could reveal whether HIPV-mediated priming activates defense signaling and also attenuates the expression of negative regulators, with potential implications for improving citrus resilience to HLB.
We hypothesized that exposure to these volatiles would differentially activate the citrus immune signaling network, specifically, the JA and/or SA defense pathways, as evidenced by enhanced transcription of genes across different functional levels. Understanding these dynamics may guide the identification of the most effective synthetic HIPVs and responsive citrus genotypes for use in stress management and resistance-oriented breeding strategies.
2 Materials and methods
2.1 Plant material
Four citrus rootstocks were selected for this study: Forner-Alcaide 5 (FA-5) and Forner-Alcaide 74 (FA-74), both hybrids of Cleopatra mandarin (Citrus reshni) and Poncirus trifoliata, developed at the Valencian Institute of Agricultural Research (IVIA); Carrizo citrange (CC), a hybrid of Poncirus trifoliata and Citrus sinensis, and Microcitrus australasica (F.Muell.) Swingle (Microcitrus) a hybrid of Citrus reticulata Blanco × Microcitrus australis. The rootstocks selected for this study were chosen based on their agronomic importance and relevance in citrus breeding, as well as their contrasting defence profiles. Carrizo Citrange is the most widely used rootstock in Spanish citrus production. FA5 has gained considerable popularity in recent years and is currently the most planted, while FA74 is expected to enter the market soon. Finally, Microcitrus australasica is increasingly being included in breeding programs due to its natural resistance to key diseases, including huanglongbing (HLB).
Seedlings were grown in 8 × 8 × 8 cm plastic pots filled with a mixture of 70% black peat and 30% perlite. All citrus plants were pesticide-free and watered twice a week. One of the waterings was supplemented with 2% citrus-specific fertilizer composed of NH4H2PO4 at 0.115 g/L, KNO3 at 0.065 g/L, Ca(NO3)2 at 1.25 g/L, and synthetic chelating at 0.018 g/L [Sequestrene® (Syngenta NK 138Fe, Basel, Switzerland)] (Dahmane et al., 2022). Plants were maintained under controlled environmental conditions (25 ± 1°°C, 60% relative humidity, and a 14:10 h light:dark photoperiod). After approximately three months of growth, when they had developed 8 to 9 fully expanded leaves and reached a height of around 30 cm, the plants were considered physiologically suitable and selected for use in the bioassays.
2.2 Citrus plant’s exposure to synthetic GLVs
Six synthetic standards of volatile compounds were selected for this study based on their known roles in plant defence signaling: (Z)-3-hexen-1-ol (Z3C6OH), (Z)-3-hexenyl acetate (Z3-HA), (Z)-3-hexenyl butyrate (Z3-HB), (Z)-3-hexenyl propanoate (Z3-HP), methyl jasmonate (MeJA), and methyl salicylate (MeSA). All compounds were purchased from Sigma-Aldrich (St. Louis, MO, United States) with a reported purity of ≥98%. The (Z)-3-hexenyl derivatives were confirmed to contain >95% of the (Z)-isomer. Methyl jasmonate was used as a commercial mix of cis- and trans-isomers, and methyl salicylate was used in its standard, non-chiral form. All compounds were applied in pure (neat) form using low-density polyethylene (LDPE) diffusers as described below (Pérez-Hedo et al., 2021b; Riahi et al., 2022).
For each rootstock, an independent experiment was conducted under identical environmental conditions to evaluate the effect of six synthetic HIPVs and one mock control. Each treatment was assigned to a separate climate-controlled incubator (Sanyo MLR-350H, Sanyo, Japan), containing seven plants (biological replicates) of a single genotype. This design prevented cross-exposure between treatments and avoided inter-plant signaling effects. All incubators were run in parallel and maintained under the same conditions: 25 ± 1°C, 60% relative humidity, and a 14:10 h light:dark photoperiod. Each incubator was equipped with fluorescent lighting (15-watt white fluorescent tubes, FL15W, Sanyo Electric Co., Ltd.) positioned on the chamber ceiling. The light intensity at shelf level was approximately 160 µmol m²s¹ (photosynthetically active radiation, PAR), measured using a quantum light sensor. This setup ensured homogeneous light distribution across all trays and chambers. Each incubator was assigned to one of the seven treatments (six synthetic HIPVs and one mock), and seven individual plants of the corresponding rootstock were placed inside, serving as the biological replicates. Volatile application was achieved using low-density polyethylene (LDPE) polymer diffusers (Kartell, Fisher Scientific SL, Madrid, Spain) (Pérez-Hedo et al., 2021a), each containing 2 ml of the designated volatile compound in pure (neat) form and suspended within the incubator chamber. Plants were exposed continuously to the volatiles for 48 hours before sample collection.
2.3 RNA extraction and gene expression analysis
Apical tissue samples were collected from each plant 48 hours after exposure to synthetic HIPV. The samples were immediately flash-frozen in liquid nitrogen and ground to a fine powder for RNA extraction using NZYol reagent (NZYTech, Lisbon, Portugal). One microgram (µg) of total RNA from each sample was treated with the TURBO DNA-free™ Kit (Ambion®, Life Technologies, CA, USA) to eliminate any contaminating genomic DNA. Complementary DNA (cDNA) was synthesized by reverse transcription using the PrimeScript™ RT Reagent Kit (TAKARA Bio, CA, USA), following the manufacturer’s instructions.
Quantitative real-time PCR (qPCR) was performed using the LightCycler® 480 System (Roche Molecular Systems, Inc., Switzerland) with NZYSpeedy qPCR Green Master Mix (2×) (NZYTech, Lisbon, Portugal), as described by Bouagga et al. (2018). Relative gene expression was calculated using the comparative Ct method (ΔΔCt) after validating primer efficiencies through the Relative Standard Curve Method (Applied Biosystems). The reference gene CsGAPC1 (Glyceraldehyde-3-phosphate dehydrogenase, formerly GAPDH) was used for normalization. Primer sequences are listed in Table 1.
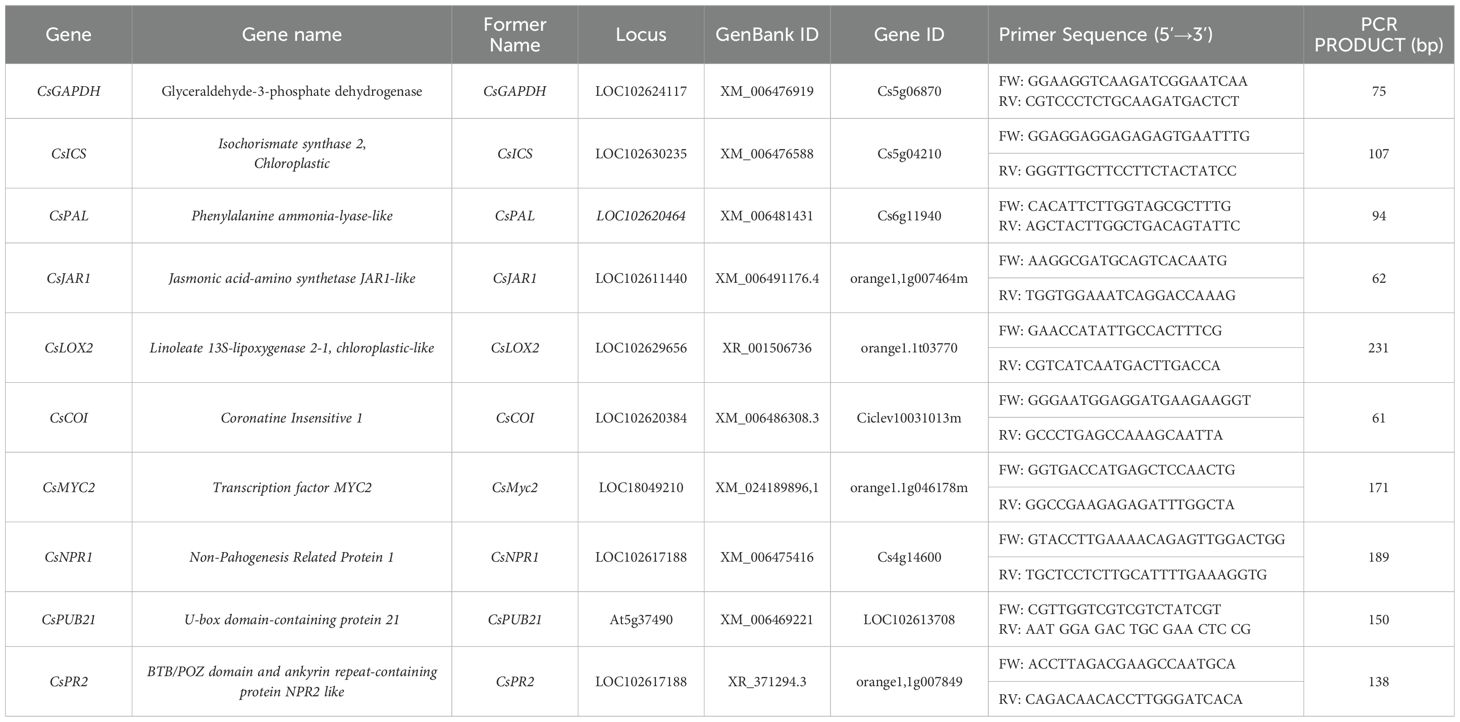
Table 1. List of genes and primers used in qPCR assays.
To obtain a comprehensive overview of defense signaling, we selected eight genes that encompass different functional components of the SA and JA pathways. For the SA pathway, we evaluated CsICS (Isochorismate synthase) and CsPAL (Phenylalanine ammonia-lyase) as two upstream biosynthetic enzymes, CsNPR1 (Non-pathogenesis-related protein 1) as an intermediate regulatory gene, and CsPR2 (Pathogenesis-related protein 2) as a downstream marker of SA pathway activation. For the JA pathway, we included two upstream genes, CsLOX2 (Lipoxygenase 2) and CsJAR1 (Jasmonate resistant 1), CsCOI (Coronatine insensitive 1) as an intermediate regulatory component, and CsMYC2 (Transcription factor MYC2) as a downstream effector. In addition, we included CsPUB21, a gene encoding a U-box E3 ubiquitin ligase recently identified as a negative regulator of JA signaling through MYC2 degradation (Table 1).
2.4 Statistical analysis
Results are presented as mean ± standard error (SE). Gene expression data were analyzed using one-way ANOVA, followed by Tukey’s post hoc test for mean separation at a significance level of P < 0.05. Statistical analyses were performed using GraphPad Prism version 10.4 (GraphPad Software, Boston, MA, USA). To generate heatmaps, data analysis was performed using R version 4.4.3. Gene expression data from pairwise combinations were transformed into a matrix format using the dcast function to calculate the mean expression values for each combination. Heatmaps were generated using the ComplexHeatmap package, employing a white-to-dark color gradient defined by the colorRamp2 function from the circlize package. Hierarchical clustering was performed on both rows and columns using Pearson correlation distance (1 - r) and complete linkage. The final visualization displayed annotated expression values within each cell.
3 Results
3.1 Expression of defense-related genes involved in the SA and JA pathways
3.1.1 Gene expression in Carrizo citrange
Within the SA pathway, the upstream gene CsICS (Figure 1A) showed higher expression levels in all volatile treatments, except for MeSA (F6,39 = 5.077; P < 0.009). For the other upstream gene, CsPAL (Figure 1B), all treatments resulted in significantly higher expression (F6,41 = 25.92; P < 0.001), with Z3C6OH inducing the highest expression level. For the intermediate gene CsNPR1 (Figure 1C), only MeSA, MeJA, and (Z)-3-HP induced significantly higher expression levels (F6,41 = 2.985; P = 0.018). Expression of the downstream marker CsPR2 (Figure 1D) did not show significant differences in response to any of the volatile treatments (F6,41 = 0.583; P = 0.090).
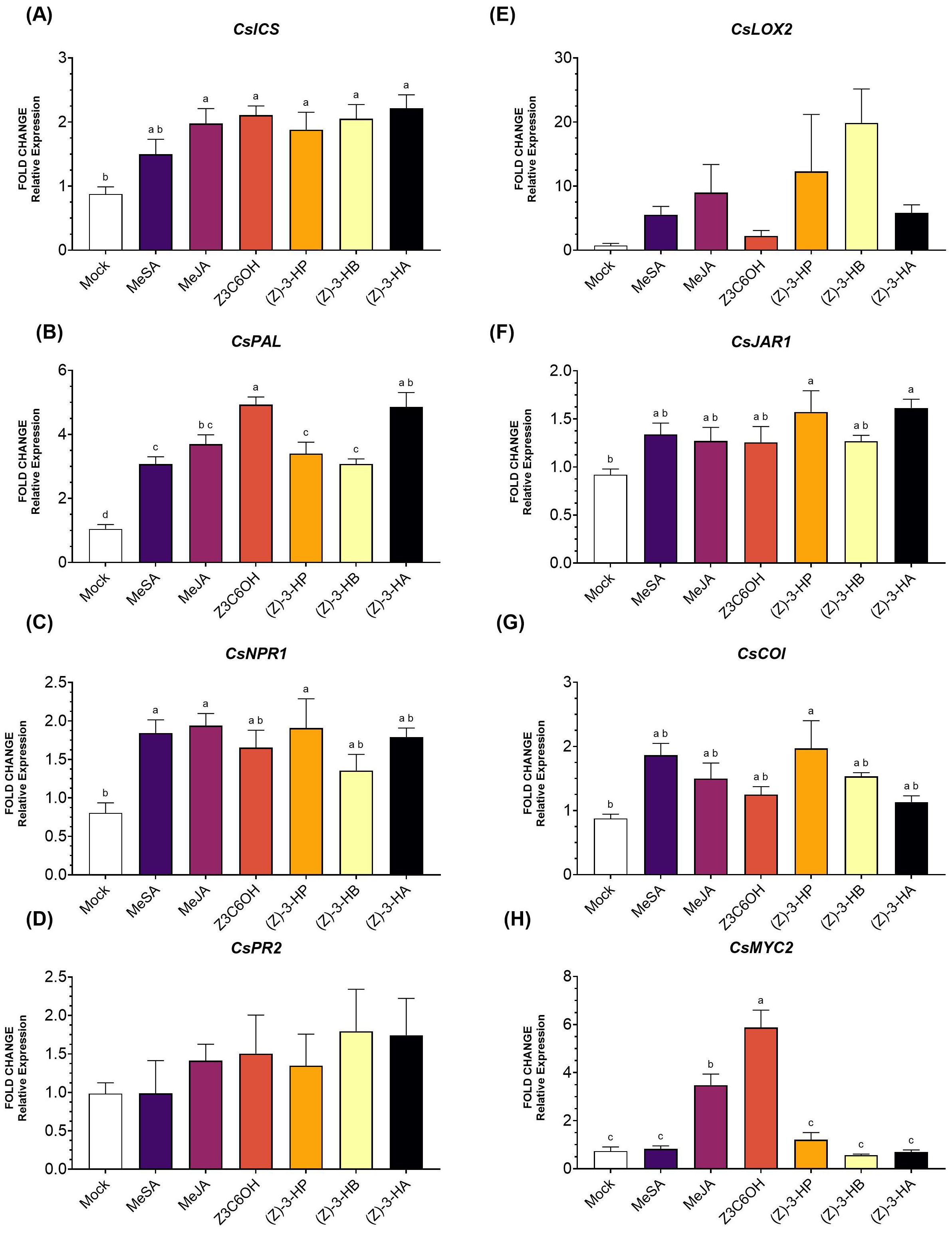
Figure 1. Expression of defense-related genes involved in the salicylic acid (SA) and jasmonic acid (JA) signaling pathways in Carrizo citrange rootstock exposed six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (A) CsICS, (B) CsPAL, (C) CsNPR1, (D) CsPR2, (E) CsLOX2, (F) CsJAR1, (G) CsCOI, (H) CsMYC2. Letters indicate significant differences between treatments based on Tukey’s test (P < 0.05).
In the JA pathway, CsLOX2 expression (Figure 1E) was higher in all HIPV treatments; however, the high variability of the data resulted in no significant differences (F6,28 = 2.402; P = 0.061). Expression of CsJAR1 (Figure 1F) was significantly increased only by (Z)-3-HP and (Z)-3-HA (F6,40 = 2.725; P = 0.028); other volatile caused moderate but non-significant increases. The expression of the intermediate marker gene CsCOI (Figure 1G) was only significantly induced by (Z)-3-HP (F6,46 = 2.973; P = 0.017). The transcription factor MYC2 (Figure 1H) was strongly upregulated by Z3C6OH and MeJA, with Z3C6OH inducing the highest expression level and showing a significant difference from all other treatments (F6,36 = 30.44; P < 0.001). Expression levels for the remaining HIPVs were not significantly different from the mock.
3.1.2 Gene expression in Forner Alcaide 5.
Within the SA pathway, the gene CsICS (Figure 2A) was significantly upregulated by MeSA, MeJA, Z3C6OH, (Z)-3-HP, and (Z)-3-HA (F6,42 = 16.54; P < 0.001). The strongest induction was observed for (Z)-3-HP, followed by (Z)-3-HA. The volatile (Z)-3-HB did not differ significantly from the mock. For the gene CsPAL (Figure 2B), expression was significantly higher in all treatments (F6,38 = 44.17; P < 0.001), with Z3C6OH showing the highest induction, followed by (Z)-3-HA. For the gene CsNPR1 (Figure 2C), expression was significantly higher only in the MeSA, MeJA, and (Z)-3-HP treatments (F6,47 = 10.08; P < 0.001). Although some variation was observed, the expression of the CsPR2 did not show significant differences in response to volatile treatments (Figure 2D) (F6,44 = 1.289; P = 0.285).
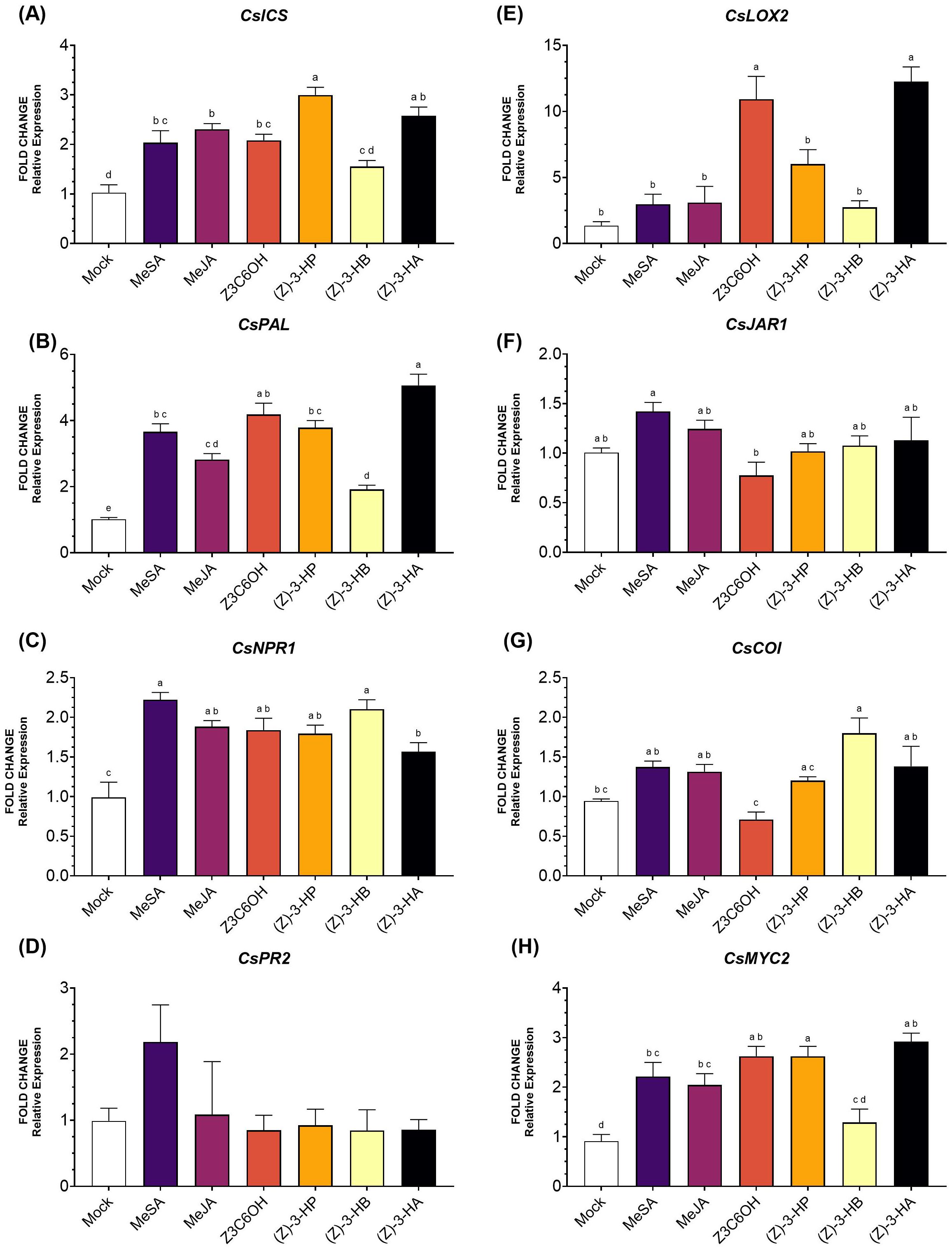
Figure 2. Expression of defense-related genes involved in the salicylic acid (SA) and jasmonic acid (JA) signaling pathways in Forner-Alcaide 5 (FA5) rootstock exposed six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (A) CsICS, (B) CsPAL, (C) CsNPR1, (D) CsPR2, (E) CsLOX2, (F) CsJAR1, (G) CsCOI, (H) CsMYC2. Letters indicate significant differences between treatments based on Tukey’s test (P < 0.05).
In the JA pathway, CsLOX2 expression (Figure 2E) was induced by all HIPV treatments, but only Z3C6OH and (Z)-3-HA induced a statistically significant increase (F6,35 = 16.79; P < 0.001). The expression of CsJAR1 (Figure 2F) was not significantly upregulated by any volatile treatment (F6,44 = 3.523; P = 0.006). Interestingly, CsJAR1 expression was reduced after Z3C6OH exposure, although this difference was not statistically significant. However, it was significantly lower than the expression observed following MeSA exposure. The expression of CsCOI (Figure 2G) was only significantly induced by (Z)-3-HB (F6,46 = 6.164; P < 0.001). Interestingly, the expression level under Z3C6OH treatment, which was not significantly different from the mock treatment, was also lower than that observed for other volatiles such as MeSA, MeJA, (Z)-3-HB, and (Z)-3-HA. Finally, CsMYC2 (Figure 2H) was significantly upregulated by all synthetic GLVs except (Z)-3-HB, with the strongest induction observed in response to (Z)-3-HP (F6,41 = 16.16; P < 0.001).
3.1.3 Gene expression in Forner Alcaide 74.
Within the SA pathway, CsICS (Figure 3A) was significantly upregulated by Z3C6OH, (Z)-3-HP, (Z)-3-HB, and (Z)-3-HA (F6,44 = 7.604; P < 0.001). Among these, (Z)-3-HP induced the highest expression levels. MeSA and MeJA also increased CsICS expression, although the differences were not statistically significant. For the gene CsPAL (Figure 3B), all synthetic GLVs treatments significantly increased expression, with MeSA, Z3C6OH, and (Z)-3-HA showing the strongest inductions (F6,40 = 22.06; P < 0.001). CsNPR1 expression (Figure 3C) increased significantly after exposure to MeJA, Z3C6OH, and (Z)-3-HP (F6,40 = 4.734; P = 0.001). Despite some changes in expression levels of CsPR2 (Figure 3D), the high variability observed between replicates resulted in no statistically significant differences across HIPV treatments (F6,40 = 1.600; P = 0.177).
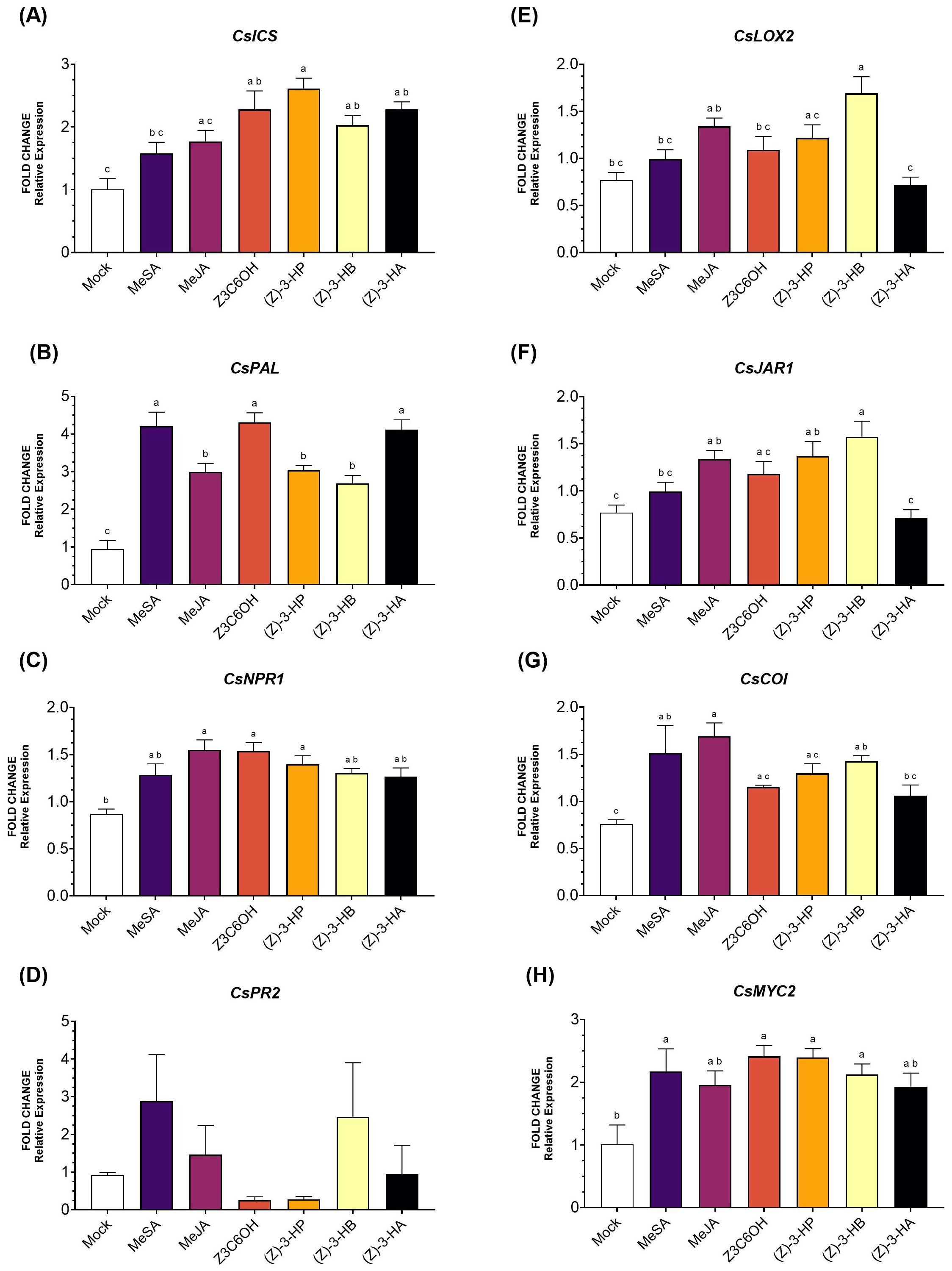
Figure 3. Expression of defense-related genes involved in the salicylic acid (SA) and jasmonic acid (JA) signaling pathways in Forner-Alcaide 74 (FA74) rootstock exposed six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (A) CsICS, (B) CsPAL, (C) CsNPR1, (D) CsPR2, (E) CsLOX2, (F) CsJAR1, (G) CsCOI, (H) CsMYC2. Letters indicate significant differences between treatments based on Tukey’s test (P < 0.05).
In the JA pathway, CsLOX2 expression (Figure 3E) was significantly upregulated only by (Z)-3-HB (F6,41 = 7.110; P < 0.001). Although other volatiles- except for (Z)-3-HA- also triggered higher expression than the control, the differences were not statistically significant. Similarly, CsJAR1 expression (Figure 3F) was significantly increased in response to MeJA, (Z)-3-HP, and (Z)-3-HB treatments (F6,37 = 7.488; P < 0.001). In the case of CsCOI (Figure 3G), exposure to MeSA, MeJA, and (Z)-3-HB led to its overexpression (F6,37 = 4.661; P = 0.002). Finally, all volatile upregulated CsMYC2 (Figure 3H), but only MeSA, Z3C6OH, (Z)-3-HP, and (Z)-3-HB showed significantly higher expression (F6,39 = 4.145; P = 0.003).
3.1.4 Gene expression in microcitrus
Within the SA pathway, the gene CsICS (Figure 4A) was significantly upregulated by all HIPV treatments, except for MeSA (F6,36 = 19.18; P < 0.001). Z3C6OH triggered the highest expression levels. For the gene CsPAL (Figure 4B), expression was significantly increased by MeSA, Z3C6OH, and (Z)-3-HP, with Z3C6OH showing the strongest response (F6,40 = 17.64; P < 0.001). The remaining volatiles, MeJA and (Z)-3-HB, also resulted in higher expression levels than the mock, but the differences were not statistically significant. CsNPR1 expression increased after exposure to all volatiles except (Z)-3-HA (F6,43 = 23.37; P < 0.001). As observed for the two upstream genes, Z3C6OH induced the highest activation, followed by (Z)-3-HP (Figure 4C). No statistically significant differences were observed in CsPR2 expression between synthetic GLVs treatments and the mock control (F6,46 = 1.777; P = 0.128) (Figure 4D).
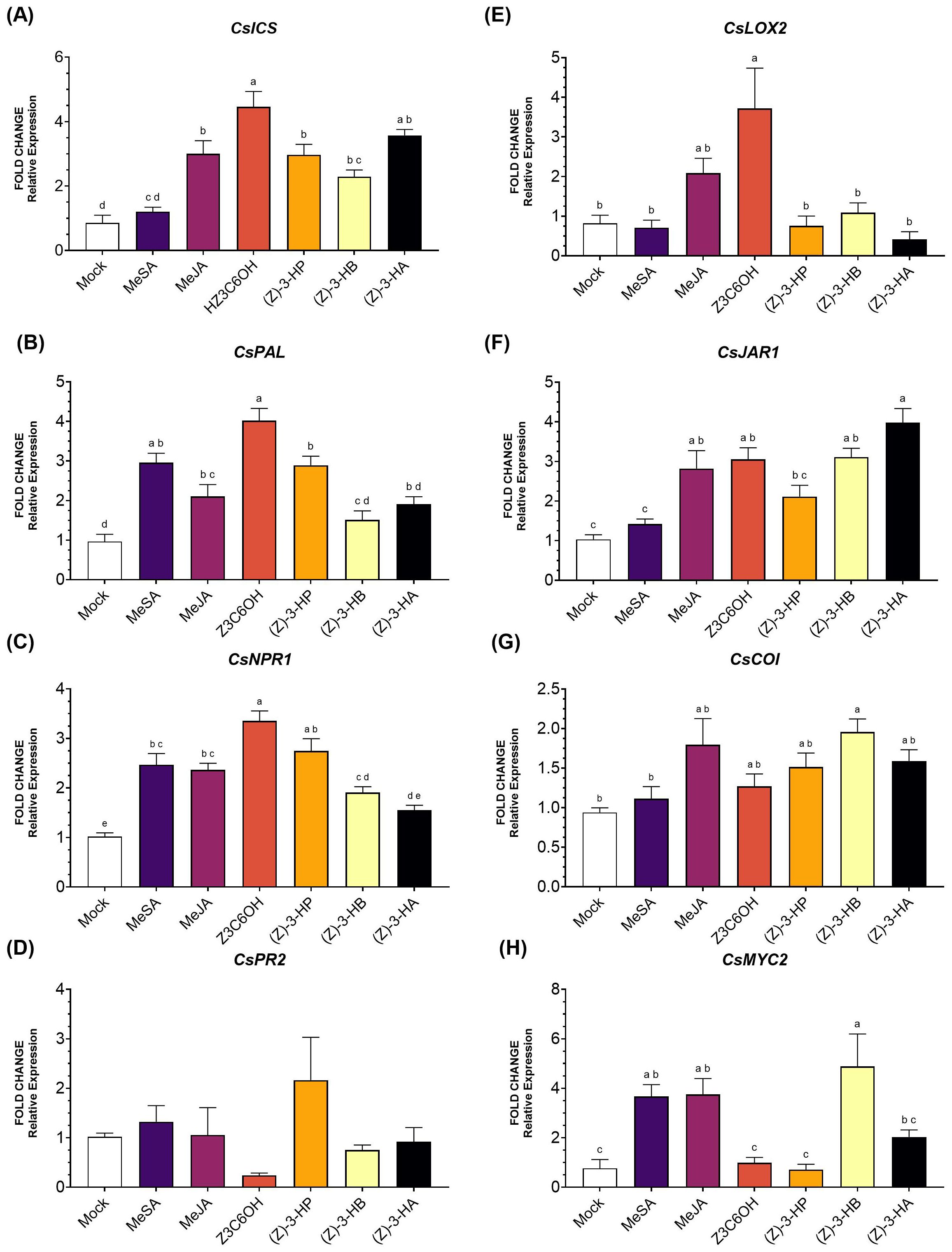
Figure 4. Expression of defense-related genes involved in the salicylic acid (SA) and jasmonic acid (JA) signaling pathways in Microcitrus rootstock exposed to six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (A) CsICS, (B) CsPAL, (C) CsNPR1, (D) CsPR2, (E) CsLOX2, (F) CsJAR1, (G) CsCOI, (H) CsMYC2. Letters indicate significant differences between treatments based on Tukey’s test (P < 0.05).
In the JA pathway, CsLOX2 expression (Figure 4E) was significantly upregulated only by Z3C6OH (F6,34 = 7.699; P < 0.001). The expression of CsJAR1 (Figure 4F) was significantly increased by MeJA, MeSA, (Z)-3-HB, and (Z)-3-HA (F6,40 = 15.90; P < 0.001). CsCOI (Figure 4G) was significantly upregulated only in response to (Z)-3-HB exposure (F6,44 = 3.511; P = 0.007). Finally, CsMYC2 (Figure 4H) showed significantly higher expression only in response to MeSA, MeJA, and (Z)-3-HB. At the same time, the other volatiles did not differ significantly from the mock treatment (F6,33 = 9.146; P < 0.001).
3.2 Expression of the susceptibility-related gene CsPUB21
Expression patterns of CsPUB21 varied across citrus genotypes in response to HIPV treatments (Figure 5). In Carrizo citrange (Figure 5A), transcript levels were significantly reduced by MeJA, Z3C6OH, and (Z)-3-HP, which showed the lowest expression values among treatments (F6,28 = 6.695; P = 0.004). (Z)-3-HA maintained high expression levels comparable to the control, while MeSA and (Z)-3-HB resulted in intermediate values. In FA5 (Figure 5B), all treatments except MeSA and (Z)-3-HA significantly reduced CsPUB21 expression compared to the mock (F6,28 = 8.213; P < 0.001), with the lowest levels observed for MeJA, Z3C6OH, (Z)-3-HP, and (Z)-3-HB. In FA74 (Figure 5C), all volatiles tested resulted in significant downregulation of CsPUB21 compared to the control (F6,28 = 9.884; P < 0.001), with Z3C6OH exhibiting the strongest repression. In contrast, in Microcitrus (Figure 5D), no significant differences were observed between treatments (F6,28 = 2.016; P = 0.097).
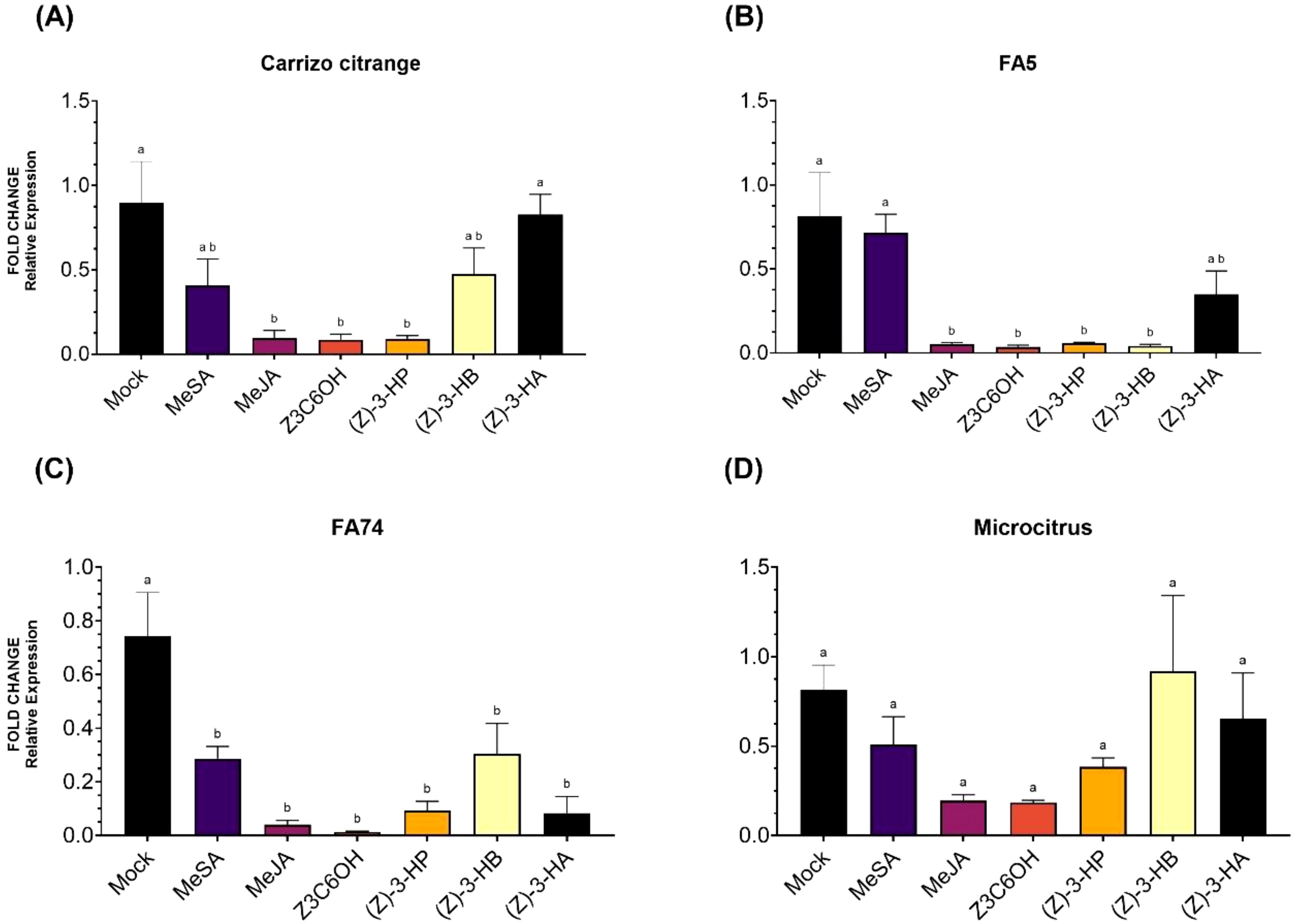
Figure 5. Relative expression of the susceptibility gene CsPUB21 in citrus rootstocks (Carrizo citrange, FA5, FA74, and Microcitrus) exposed to six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (A) Carrizo citrange, (B) Forner-Alcaide 5 (FA5), (C) Forner-Alcaide 74 (FA74), (D) Microcitrus. Letters indicate significant differences between treatments based on Tukey’s test (P < 0.05).
3.3 Gene expression heatmaps
Figure 6 illustrates the transcriptional responses of defense-related genes under different volatile treatments and across citrus genotypes. The clustering analysis in Figure 6A reveals that the Z3C6OH and MeJA groups cluster together, as do the (Z)-3-HP and (Z)-3-HA groups, indicating that these volatiles elicit similar gene expression patterns. In contrast, MeSA and (Z)-3-HB form independent branches, suggesting distinct transcriptional profiles compared to the other HIPVs.
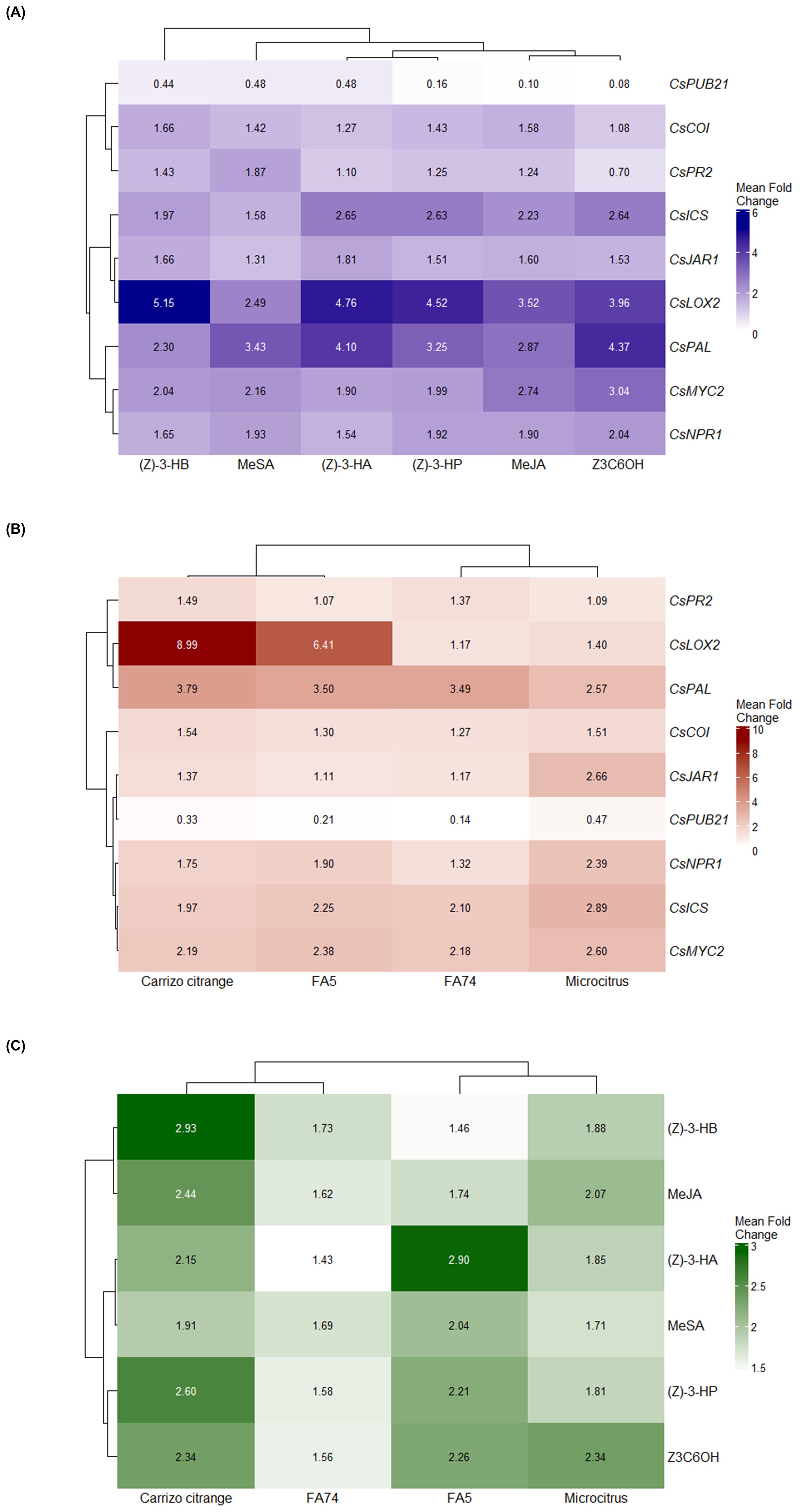
Figure 6. Heatmaps showing hierarchical clustering of fold-change expression levels of defense-related genes in citrus rootstocks exposed to synthetic HIPVs. Log2 fold-change (FC) values were calculated from qPCR data for eight defense-related genes (CsICS, CsPAL, CsNPR1, CsPR2, CsLOX2, CsJAR1, CsCOI, CsMYC2, and CsPUB21) following exposure to synthetic HIPVs. (A) Gene expression profiles under six synthetic HIPV treatments: methyl salicylate (MeSA), methyl jasmonate (MeJA), (Z)-3-hexenyl hexanoate (Z3C6OH), (Z)-3-hexenyl propanoate [(Z)-3-HP], (Z)-3-hexenyl butanoate [(Z)-3-HB], and (Z)-3-hexenyl acetate [(Z)-3-HA]. (B) Expression patterns across four citrus genotypes: Carrizo citrange, FA5, FA74, and Microcitrus. (C) Combined heatmap showing average gene expression responses across both synthetic HIPV treatments and genotypes. Hierarchical clustering was performed using Euclidean distance and complete linkage. Color scales represent log2 FC values, with darker shades indicating stronger induction levels.
In Figure 6B, the hierarchical clustering of genotypes reveals that Carrizo and FA5 cluster together, indicating comparable and elevated transcriptional responses to volatile exposure. In contrast, FA74 and Microcitrus form a separate cluster, with Microcitrus exhibiting higher overall induction levels than FA74.
Figure 6C further displays the clustering of both genotypes and volatiles based on average gene expression levels. Among the volatiles, (Z)-3-HP and Z3C6OH form one cluster, while MeSA and (Z)-3-HA form another, indicating shared activation profiles within each pair. MeJA and (Z)-3-HB appear as separate branches, suggesting unique patterns of gene induction. These results demonstrate both compound- and genotype-dependent variation in transcriptional responses to synthetic HIPVs.
4 Discussion
Our results demonstrate that synthetic GLVs can activate immune signaling pathways in citrus rootstocks, with responses depending on both the hormone pathway and the genotype. Notably, we observed the downregulation of CsPUB21, a key susceptibility gene linked to MYC2 regulation and HLB sensitivity.
Exposure to synthetic HIPVs generally triggered the activation of upstream and intermediate genes in the SA pathway, including CsICS, CsPAL, and CsNPR1, as indicated by their fold-change values in Figure 6A. In contrast, CsPR2 displayed much lower and more variable expression, suggesting that volatile exposure may selectively enhance early steps of the signaling cascade without fully activating downstream defense effectors. This pattern supports the hypothesis that these volatiles induce a primed state, in which plants are metabolically prepared to respond more rapidly and effectively to subsequent stress (Engelberth et al., 2004; Ton et al., 2006; Frost et al., 2008; Erb et al., 2015). However, this interpretation remains hypothetical and would need to be validated through challenge experiments involving actual biotic stress to confirm that the plants are indeed primed for enhanced defence.
In addition to priming, other regulatory mechanisms may explain the limited activation of downstream defense genes such as CsPR2. Antagonistic hormonal crosstalk, particularly from JA or ABA, could interfere with SA signaling or limit the activity of key regulators such as NPR1. Notably, NPR1 requires interaction with TGA transcription factors to activate SA-responsive genes, and their availability or activity may be constrained under certain hormonal conditions (Zhang et al., 1999). These multilayered controls, including feedback loops and energy-saving strategies, help fine-tune immune responses, allowing the plant to balance growth and defense depending on environmental cues (Huot et al., 2014; Zhou et al., 2015).
When examining the JA-related genes in the heatmap (Figure 6), CsLOX2 and CsMYC2 stand out as the most strongly induced genes within the JA signaling group. CsLOX2, which participates in the initial steps of JA biosynthesis, showed consistent upregulation across treatments, particularly under (Z)-3-HA and (Z)-3-HP. However, this induction does not necessarily imply full activation of the jasmonic acid pathway, as reflected by the low expression levels of intermediate genes such as CsJAR1 and CsCOI. It is possible that CsLOX2 is acting independently, or that its activation does not culminate in the production of bioactive JA derivatives, since LOX genes can participate in various oxylipin-related processes. CsMYC2 also exhibited strong expression, especially in response to Z3C6OH and MeJA, suggesting that HIPVs may preferentially enhance late-stage components of the JA cascade. These findings support the hypothesis of transcriptional reprogramming centered on MYC2, independently of the full activation of the upstream JA biosynthetic machinery. As proposed by Song et al. (2022), MYC2 functions as a central integrator of stress signals and can activate defense genes in response to specific stimuli, including HIPVs, even in the absence of complete JA biosynthesis. Such activation may represent a priming-like state, allowing the plant to prepare for future attack with minimal metabolic cost. Supporting this hypothesis, we found that the expression of CsPUB21, an E3 ubiquitin ligase responsible for MYC2 degradation (Zhao et al., 2025), was consistently repressed across all volatile treatments and genotypes. Fold-change values remained below 0.5 in every case. This suggests that HIPVs may activate MYC2 at the transcriptional level and reduce its proteolytic turnover, contributing to its stabilization. This dual regulatory effect, involving the activation of MYC2 and the suppression of its negative regulator, adds a new layer of control to volatile-mediated defense priming in citrus.
Among the four genotypes evaluated, Carrizo and FA5 exhibited the most robust transcriptional responses to HIPV exposure, as observed in their clustering and expression levels in Figure 6B, with strong induction of genes in both the SA and JA pathways. This suggests a higher sensitivity to volatile cues and more efficient signal transduction in these rootstocks. In contrast, FA74 showed the weakest activation overall, particularly within the JA pathway, while Microcitrus displayed a broad but moderate activation pattern. These genotypic differences may have practical implications for selecting rootstocks that enhance pest and disease resistance under integrated management strategies.
The observed variability in gene activation among citrus rootstocks likely reflects their distinct genetic backgrounds, which influence volatile perception, basal defense status, and signaling capacity. The stronger response of Carrizo citrange and FA5 may be attributed to their P. trifoliata lineage, known for its role in conferring resistance to both biotic and abiotic stressors (Forner-Giner et al., 2020; Peng et al., 2020; Wang et al., 2024). Recent studies have shown that P. trifoliata contributes to enhanced expression of defense-related genes in response to synthetic HIPVs (Pérez-Hedo et al., 2024a). In parallel, species of the Microcitrus genus, such as Microcitrus australasica, are naturally resistant to a broad range of citrus pathogens, including CLas, the causal agent of HLB (Huang et al., 2021). This resistance has positioned Microcitrus as a valuable genetic resource for both conventional breeding and biotechnological approaches (Alquézar et al., 2021).
Interestingly, despite its strong resistance phenotype, Microcitrus australasica exhibited the highest CsPUB21 expression among the four genotypes and showed no significant repression of this gene following volatile treatment. This suggests a fundamentally different regulatory context for CsPUB21 in this species. According to Zhao et al. (2025), Microcitrus lacks a helitron insertion in the CsPUB21 promoter that is present in susceptible species such as C. sinensis, which reduces its responsiveness to JA-mediated induction via MYC2 binding sites. Additionally, although Microcitrus expresses low levels of the dominant-negative paralog PUB21DN, this mechanism may still partially mitigate PUB21 activity. Together, these findings suggest that in Microcitrus, resistance may not depend on the transcriptional repression of PUB21, but instead on structural and functional divergence that diminishes its immunosuppressive role.
SA plays a central role in defense against biotrophic pathogens, and its importance in limiting CLas colonization and symptom progression is increasingly supported by experimental evidence (Liu et al., 2023; Pérez-Hedo et al., 2024b; Yang et al., 2024). Based on this, priming SA-related defenses prior to pathogen exposure emerges as a promising strategy to improve citrus tolerance to HLB. Our findings support this approach, showing that selected HIPVs can pre-activate components of both SA and JA pathways while simultaneously repressing CsPUB21, a gene associated with increased HLB susceptibility. This dual action may reinforce plant immunity and promote a sustained defense-ready state. Applied in nurseries or during early orchard establishment, synthetic HIPV treatments could serve as a proactive tool to strengthen the plant’s immune system before infection occurs, contributing to more effective and sustainable HLB management strategies. This is a critical question for future research, especially in the context of field applications where transient exposure or pulsed treatments may be more practical. Encouragingly, studies in other crops using a single, transient biotic stimulus, such as the brief exposure to herbivory by the predatory mirid Nesidiocoris tenuis Reuter (Hemiptera: Miridae), have shown that defence responses can persist for up to 14 days (Bouagga et al., 2018). Whether a similar persistence can be achieved with synthetic volatiles in citrus is currently unknown and will be addressed in future work aimed at characterizing the temporal window of the primed state.
Among the tested volatiles, (Z)-3-HP and Z3C6OH consistently stood out for their robust transcriptional impact and for promoting CsPUB21 repression, a pattern confirmed by their grouping and high average expression values in Figure 6C. These dual effects, which enhance immune signaling and reduce the expression of a key susceptibility gene, position them as promising agents for defense priming. This aligns with our previous transcriptomic study with (Z)-3-HP, which showed broad transcriptional activation of genes related to plant immunity, stress tolerance, and redox balance in citrus (Pérez-Hedo et al., 2024a), reinforcing the potential of this compound for integrated pest and disease management.
Overall, our findings underscore the potential of (Z)-3-HP and Z3C6OH as robust priming agents to enhance citrus defense responses. Their consistent activation of SA and JA signaling, combined with the suppression of the susceptibility gene CsPUB21, suggests a coordinated mechanism that strengthens immunity at multiple regulatory levels. Given their broad-spectrum effects across genotypes, these synthetic HIPVs represent promising candidates for sustainable citrus protection (Farag et al., 2005; Yang et al., 2020; Pérez-Hedo et al., 2021b, 2024a). However, their practical implementation requires further validation under field conditions and in grafted plants, considering that rootstock-scion interactions may influence responsiveness.
Future work will focus on consolidating and expanding the validation of synthetic HIPVs under field conditions. We have been conducting field experiments for the past three years across three commercial citrus orchards in eastern Spain. These trials are evaluating the long-term effects of selected volatile compounds on disease resistance (including HLB), pest population dynamics, and agronomic performance. Treated trees are being compared with untreated controls to assess differences in disease incidence, physiological stress, and fruit yield. Building on these results, upcoming studies will explore the effectiveness of pulsed and transient exposure strategies, as well as rootstock–scion interactions that may influence responsiveness to synthetic HIPVs. This ongoing work aims to establish practical, low-input protocols for integrating HIPV-based priming into sustainable citrus production systems.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
RO: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. CG: Data curation, Formal analysis, Investigation, Methodology, Software, Writing – review & editing. MF: Funding acquisition, Investigation, Resources, Writing – review & editing. AU: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing. MP: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was partially supported by project no. PID2020-113234RR-I00 from the Spanish Ministry of Science and Innovation through the State Research Agency (co-financed by the European Regional Development Fund, ERDF) and by the IVIA-52202F project from the Valencian Institute of Agricultural Research (IVIA) of the Valencian Government (GVA) (this project is eligible for co-financing by the European Union through the ERDF Operational Program). RO-F was supported by a Ph.D. contract from the Spanish Ministry of Science and Innovation (PRE2021-100879).
Conflict of interest
MP and AU are inventors of the Spanish Patent No. P202030330 entitled “Use of (Z)-3-hexenyl esters and method for protecting plants against pests".
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Some paragraphs were checked at the grammatical level in English.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alquézar, B., Carmona, L., Bennici, S., and Peña, L. (2021). Engineering of citrus to obtain huanglongbing resistance. Curr. Opin. Biotechnol. 70, 196–203. doi: 10.1016/j.copbio.2021.06.003
Belhadj, A., Saigne, C., Telef, N., Cluzet, S., Bouscaut, J., Corio-Costet, M.-F., et al. (2006). Methyl Jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator. J. Agric. Food Chem. 54, 9119–9125. doi: 10.1021/jf0618022
Bouagga, S., Urbaneja, A., Rambla, J. L., Flors, V., Granell, A., Jaques, J. A., et al. (2018). Zoophytophagous mirids provide pest control by inducing direct defences, antixenosis and attraction to parasitoids in sweet pepper plants. Pest Manag. Sci. 74, 1286–1296. doi: 10.1002/ps.4838
Dahmane, M., Urbaneja, A., Ruíz-Rivero, O., Alonso-Valiente, M., and Pérez-Hedo, M. (2022). The zoophytophagous predator Pilophorus clavatus (Hemiptera: Miridae) induces plant defences in citrus. J. Pest Sci. 95, 1519–1530. doi: 10.1007/s10340-022-01558-5
Depalo, L., Urbaneja, A., Gallego, C., Fournarakos, A., Alonso, M., and Pérez-Hedo, M. (2022). Eliciting sweet pepper plant resistance to Aulacorthum solani and attractiveness on Aphelinus abdominalis by exposure to (Z)-3-hexenyl propanoate. Entomol. Gen. 42, 743–749. doi: 10.1127/entomologia/2022/1595
Dicke, M. (2009). Behavioural and community ecology of plants that cry for help. Plant Cell Environ. 32, 654–65. doi: 10.1111/j.1365-3040.2008.01913.x
Dicke, M and Baldwin, I. T. (2010). The evolutionary context for herbivore-induced plant volatiles: beyond the 'cry for help'. Trends Plant Sci. 15, 167–75. doi: 10.1016/j.tplants.2009.12.002
Engelberth, J., Alborn, H. T., Schmelz, E. A., and Tumlinson, J. H. (2004). Airborne signals prime plants against insect herbivore attack. Proc. Natl. Acad. Sci. 101, 1781–1785. doi: 10.1073/pnas.0308037100
Erb, M., Veyrat, N., Robert, C. A. M., Xu, H., Frey, M., Ton, J., et al. (2015). Indole is an essential herbivore-induced volatile signal that primes maize. Nat. Commun. 6, 6273. doi: 10.1038/ncomms7273
Farag, M. A., Fokar, M., Abd, H., Zhang, H., Allen, R. D., and Paré, P. W. (2005). (Z)-3-Hexenol induces defense genes and downstream metabolites in maize. Planta 220, 900–909. doi: 10.1007/s00425-004-1404-5
Forner-Giner, M. A., Continella, A., and Grosser, J. W. (2020). Citrus rootstock breeding and selection. Eds. Gentile, A., La Malfa, S., and Deng, Z. (Cham, Switzerland: Springer), 49–74. doi: 10.1007/978-3-030-15308-3_5
Frost, C. J., Mescher, M. C., Carlson, J. E., and De Moraes, C. M. (2008). Plant defense priming against herbivores: Getting ready for a different battle. Plant Physiol. 146, 818–824. doi: 10.1104/pp.107.113027
Howe, G. A. and Jander, G. (2008). Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 59, 41–66. doi: 10.1146/annurev.arplant.59.032607.092825
Huang, C.-Y., Araujo, K., Sánchez, J. N., Kund, G., Trumble, J., Roper, C., et al. (2021). A stable antimicrobial peptide with dual functions of treating and preventing citrus huanglongbing. Proc. Natl. Acad. Sci. 118, e2019628118. doi: 10.1073/pnas.2019628118
Huot, B., Yao, J., Montgomery, B. L., and He, S. Y. (2014). Growth–defense tradeoffs in plants: a balancing act to optimize fitness. Mol. Plant 7, 1267–1287. doi: 10.1093/mp/ssu049
Kessler, A. and Baldwin, I. T. (2002). Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 53, 299–328. doi: 10.1146/annurev.arplant.53.100301.135207
Liu, Y., Dong, L., Ran, D., Wang, S., Qu, R., Zheng, L., et al. (2023). A comparative analysis of three Rutaceae species reveals the multilayered mechanisms of citrus in response to huanglongbing disease. J. Plant Growth Regul. 42, 7564–7579. doi: 10.1007/s00344-023-11032-w
López-Gresa, M. P., Payá, C., Ozáez, M., Rodrigo, I., Conejero, V., Klee, H., et al. (2018). A new role for green leaf volatile esters in tomato stomatal defense against Pseudomonas syringe pv. tomato. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01855
Maurya, A. K., Pazouki, L., and Frost, C. J. (2022). Priming seeds with indole and (Z)-3-hexenyl acetate enhances resistance against herbivores and stimulates srowth. J. Chem. Ecol. 48, 441–454. doi: 10.1007/s10886-022-01359-1
Payá, C., Belda-Palazón, B., Vera-Sirera, F., Pérez-Pérez, J., Jordá, L., Rodrigo, I., et al. (2024). Signalling mechanisms and agricultural applications of (Z)-3-hexenyl butyrate-mediated stomatal closure. Hortic. Res. 11, uhad248. doi: 10.1093/hr/uhad248
Peng, Z., Bredeson, J. V., Wu, G. A., Shu, S., Rawat, N., Du, D., et al. (2020). A chromosome-scale reference genome of trifoliate orange (Poncirus trifoliata) provides insights into disease resistance, cold tolerance and genome evolution in Citrus. Plant J. 104, 1215–1232. doi: 10.1111/tpj.14993
Pérez-Hedo, M., Alonso-Valiente, M., Vacas, S., Gallego, C., Pons, C., Arbona, V., et al. (2021a). Plant exposure to herbivore-induced plant volatiles: a sustainable approach through eliciting plant defenses. J. Pest Sci. 94, 1221–1235. doi: 10.1007/s10340-021-01334-x
Pérez-Hedo, M., Alonso-Valiente, M., Vacas, S., Gallego, C., Rambla, J. L., Navarro-Llopis, V., et al. (2021b). Eliciting tomato plant defenses by exposure to herbivore induced plant volatiles. Entomol. Gen. 41, 209–218. doi: 10.1127/entomologia/2021/1196
Pérez-Hedo, M., Gallego-Giraldo, C., Forner-Giner, M. A., Ortells-Fabra, R., and Urbaneja, A. (2024a). Plant volatile-triggered defense in citrus against biotic stressors. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1425364
Pérez-Hedo, M., Hoddle, M. S., Alferez, F., Tena, A., Wade, T., Chakravarty, S., et al. (2025). Huanglongbing (HLB) and its vectors: recent research advances and future challenges. Entomol. Gen. 45, 17–35. doi: 10.1127/entomologia/2024/3081
Pérez-Hedo, M., Urbaneja, A., and Alférez, F. (2024b). Homobrassinolide delays huanglongbing progression in newly planted citrus (Citrus sinensis) trees. Plants 13, 1229. doi: 10.3390/plants13091229
Riahi, C., González-Rodríguez, J., Alonso-Valiente, M., Urbaneja, A., and Pérez-Hedo, M. (2022). Eliciting plant defenses through herbivore-induced plant volatiles’ exposure in sweet peppers. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.776827
Song, S., Cao, Y., Dai, J., Li, G., Manzoor, M.A., Chen, C., and Deng, H. (2022). The multifaceted roles of MYC2 in plants: toward transcriptional reprogramming and stress tolerance by jasmonate signaling. Front. Plant Sci. 13:868874. doi: 10.3389/fpls.2022.868874
Ton, J., D’Alessandro, M., Jourdie, V., Jakab, G., Karlen, D., Held, M., et al. (2006). Priming by airborne signals boosts direct and indirect resistance in maize. Plant J. 49, 16–26. doi: 10.1111/j.1365-313X.2006.02935.x
Tudi, M., Daniel Ruan, H., Wang, L., Lyu, J., Sadler, R., Connell, D., et al. (2021). Agriculture development, pesticide application and its impact on the environment. Int. J. Environ. Res. Public Health 18, 1112. doi: 10.3390/ijerph18031112
Turlings, T. C. J. and Erb, M. (2018). Tritrophic interactions mediated by herbivore-induced plant volatiles: mechanisms, ecological relevance, and application potential. Annu. Rev. Entomol. 63, 433–452. doi: 10.1146/annurev-ento-020117-043507
Urbaneja, A., Grout, T. G., Gravena, S., Wu, F., Cen, Y., and Stansly, P. A. (2020). “Citrus pests in a global world,” in The Genus Citrus (Cambridge, MA, US: Elsevier), 333–348. doi: 10.1016/B978-0-12-812163-4.00016-4
Wang, X., Ji, H., Zhong, L., Zeng, W., Ouyang, Z., and Li, R. (2024). A transcriptome analysis of Poncirus trifoliata, an Aurantioideae species tolerant to Asian citrus psyllid, has identified potential genes and events associated with psyllid resistance. Insects 15, 589. doi: 10.3390/insects15080589
War, A. R., Paulraj, M. G., Ahmad, T., Ahad, A., Hussain, B., Ignacimuthu, S., et al. (2012). Mechanisms of plant defense against insect herbivores. Plant Signal Behav. 7, 1306–1320. doi: 10.4161/psb.21663
Yang, K., Hu, B., Zhang, W., Yuan, T., and Xu, Y. (2024). Recent progress in the understanding of Citrus huanglongbing: from the perspective of pathogen and citrus host. Mol. Breed. 44, 77. doi: 10.1007/s11032-024-01517-1
Yang, F., Zhang, Q., Yao, Q., Chen, G., Tong, H., Zhang, J., et al. (2020). Direct and indirect plant defenses induced by (Z)-3-hexenol in tomato against whitefly attack. J. Pest Sci. 93, 1243–1254. doi: 10.1007/s10340-020-01234-6
Zhang, Y., Fan, W., Kinkema, M., Li, X., and Dong, X. (1999). Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Acad. Sci. 96, 6523–6528. doi: 10.1073/pnas.96.11.6523
Zhao, P., Yang, H., Sun, Y., Zhang, J., Gao, K., Wu, J., et al. (2025). Targeted MYC2 stabilization confers citrus Huanglongbing resistance. Science 388, 191–198. doi: 10.1126/science.adq7203
Keywords: jasmonic acid signaling, salicylic acid pathway, CsPUB21, volatile organic compounds, defense gene expression, biotic stress, abiotic stress, citrus rootstocks
Citation: Ortells-Fabra R, Gallego-Giraldo C, Forner-Giner MA, Urbaneja A and Pérez-Hedo M (2025) Genotype-dependent responses to HIPV exposure in citrus: repression of CsPUB21 and activation of SA/JA signaling. Front. Plant Sci. 16:1605151. doi: 10.3389/fpls.2025.1605151
Received: 02 April 2025; Accepted: 21 May 2025;
Published: 25 June 2025.
Edited by:
Islam S. Sobhy, Cardiff University, United KingdomReviewed by:
Maria Carolina Blassioli Moraes, Brazilian Agricultural Research Corporation (EMBRAPA), BrazilJozsef Vuts, Rothamsted Research, United Kingdom
Copyright © 2025 Ortells-Fabra, Gallego-Giraldo, Forner-Giner, Urbaneja and Pérez-Hedo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Meritxell Pérez-Hedo, bWVwZWhlQGlibWNwLnVwdi5lcw==
†ORCID: Raúl Ortells-Fabra, orcid.org/0000-0001-7752-7749
Carolina Gallego-Giraldo, orcid.org/0000-0002-3385-3072
Maria Angeles Forner-Giner, orcid.org/0000-0003-3237-0893
Alberto Urbaneja, orcid.org/0000-0001-5986-3685
Meritxell Pérez-Hedo, orcid.org/0000-0003-3411-0761