 Milena Carvalho
Milena Carvalho Jonathan A. Haws1,2
Jonathan A. Haws1,2- 1ICArEHB, Faculdade de Ciências Humanas e Sociais, Universidade do Algarve, Campus de Gambelas, Faro, Portugal
- 2Department of Anthropology, University of Louisville, Louisville, KY, United States
- 3Department of Anthropology, University of New Mexico, Albuquerque, NM, United States
Recent revisions of Neanderthal behavioral models call into question the notion of inflexible subsistence strategies. Here, we present new zooarchaeological and taphonomic data from Level JJ at Lapa do Picareiro (Portuguese Estremadura), dating to ~51.5–42.5 ka cal BP, to evaluate Neanderthal prey selection, mobility, and adaptive responses to climatic fluctuations during MIS 3. Our zooarchaeological and taphonomic analysis of macro-mammalian faunal assemblages—divided into three stratigraphic subunits—reveals a consistent emphasis on red deer exploitation, supplemented by occasional hunting of ibex, chamois, aurochs, and horse. Despite paleoenvironmental disturbances associated with Heinrich Stadial (HS) 5, the stable abundance and processing of red deer indicate that these key resources remained reliable, supporting a localized foraging territory estimated at 225–400 km2. In contrast, the lithic record shows a shift in raw material procurement before and after the climatic event, suggesting adaptive technological responses. Overall, our findings imply that Neanderthals at Picareiro employed brief, intermittent site occupations while maintaining resilient, regionally adapted subsistence and mobility strategies in the face of environmental variability.
1 Introduction
Neanderthal behavioral models, particularly in the realm of subsistence, have undergone considerable revision in the last few decades (e.g., Blasco et al., 2022). Earlier views that Neanderthal subsistence was inflexible and narrow in diet breadth (e.g., Straus, 1992; Richards et al., 2000; Grayson and Delpech, 2002; Patou-Mathis, 2000) and that this may have contributed to their demise (e.g., Fa et al., 2013) have been challenged by emerging zooarchaeological evidence at numerous sites tri-peninsular southern Europe (e.g., Blasco and Peris, 2012; Blasco et al., 2016a; Carvalho et al., 2018; Blasco et al., 2022; Finlayson et al., 2012; Peresani et al., 2011; Nabais and Zilhão, 2019; Boneta Jiménez et al., 2025; Cortés-Sánchez et al., 2011; Villa et al., 2020; Nabais et al., 2023; Livraghi et al., 2021; Rendu et al., 2023; Terlato et al., 2019), France (e.g., Hardy and Moncel, 2011; Cochard et al., 2012; Morin and Laroulandie, 2012; Hardy et al., 2013; Pelletier et al., 2019; Daujeard et al., 2017; Marin et al., 2020), the Levant (e.g., Stiner, 1994; Blasco et al., 2016b, 2019; Orbach and Yeshurun, 2021; Yaroshevich et al., 2023;) and elsewhere in Eurasia (e.g., Evins, 1982; Gaudzinski-Windheuser and Niven, 2009; Van Neer and Wouters, 2009; Darlas and Psathi, 2016; Stepanchuk et al., 2017; Goffette et al., 2024) of a wide array of foraging behaviors and adaptive strategies, from exploiting small game, and marine resources to targeting one or two larger ungulates. This dietary diversity suggests that Neanderthals displayed flexibility in foraging tactics and expertly exploited their surrounding environments, contradicting the idea that limited prey selection had a role in their demise.
In Iberia, where the timing of the last Neanderthal occupations is contentious (e.g., Wood et al., 2013; Higham et al., 2014), prey selection behaviors among Middle and Upper Paleolithic human populations were similar (Altuna, 1992; Straus, 2013; Yravedra, 2013; Marín-Arroyo and Sanz-Royo, 2021) and may have been driven by bioclimatic region (Jones and Carvalho, 2023) and biomass availability (Vidal-Cordasco et al., 2022). Differences in land use and mobility, rather than prey selection, may have been a key behavioral separation between Middle and Upper Paleolithic humans. Perhaps unsurprisingly, scales of analysis influence the patterns observed. Based on cluster and nonmetric multidimensional scaling analyses of anthropogenic faunal assemblages dated 60–30 kya from all over Iberia, Neanderthals and modern human prey selection was similar and influenced by the availability of mammalian taxa in different bioclimatic regions on a peninsular scale (Jones and Carvalho, 2023). Differences in prey selection between the two human populations were hypothesized by the authors to be driven by mobility patterns, wherein Neanderthals had high foraging mobility and shorter site occupation while Upper Paleolithic peoples exploited more localized hunting patches and occupied sites more intensely, for longer durations (Jones and Carvalho, 2023). At the regional scale, however, evidence varies. A converse pattern has been observed in northern Spain (Marín-Arroyo and Sanz-Royo, 2021), with early modern humans highly mobile on the landscape with short occupations at sites while Neanderthals remained at sites for longer durations. In the Iberian Mediterranean basin, however, evidence points to highly mobile Neanderthal populations who left behind accumulations of materials at sites indicative of short duration occupations (Martínez-Moreno et al., 2004; Real et al., 2018; Rosell et al., 2017; Vallverdú et al., 2005; Zilhão and Villaverde, 2008). As in Iberia, variation in Neanderthal foraging mobility throughout Eurasia seems to be shaped by regional environmental and potentially cultural factors (see Marin et al., 2020 and references therein) and fits into two categories proposed by Stiner (2013): high mobility and short-term site occupations or lower mobility and more intensive site occupations. The potential for local adaptations to climate and resource fluctuations suggest detailed, multi-layered site-based analyses could reveal nuances in Neanderthal subsistence, mobility and landscape use not visible at broader geographic scales.
Prey choice offers a valuable lens for reconstructing Neanderthal mobility and land use. The species composition of a faunal assemblage can reflect the habitats Neanderthals exploited for food procurement and has been widely used to infer land use and mobility strategies (e.g., Villaverde et al., 1996; Daujeard et al., 2017; Marín et al., 2019). Investigating the ecological requirements of the taxa selected by Neanderthals allows for the identification of potential habitats in which Neanderthals could find specific prey items. Calculating the distance between the site and the nearest habitat in which a taxon can be encountered results in a minimum subsistence mobility radius that Neanderthals frequented. For example, the presence of ibex (Capra pyrenaica) and chamois (Rupicapra rupicapra) in an assemblage suggests hunting in alpine or mountainous regions (Acevedo and Cassinello, 2009; Nesti et al., 2010). Red deer (Cervus elaphus), roe deer (Capreolus capreolus), and wild boar (Sus scrofa), particularly when found together, point to movements through forested landscapes. While red deer are ecologically flexible (Straus, 1981; Sommer et al., 2008), their coexistence with roe deer and wild boar in an assemblage strengthens this interpretation (Mauget, 1981; Danilkin, 1995). Aurochs (Bos primigenius) are thought to have inhabited semi-open riverine areas (Hall, 2008) and the presence of horses (Equus sp.) may indicate exploitation of open, colder habitats (Berger, 1986).
Analyzing the ecological requirements of other hunted taxa in a similar way can then build a wider picture of minimum potential movements made on the landscape by Neanderthals. It is important to consider, however, that Portuguese Estremadura during the Pleistocene did not have the same ecological conditions as today's, however, some habitat types such as altitude, river valleys, are consistent between the Pleistocene and modern times.
For Portuguese Estremadura, where complex landscapes provide access to several ecotonal zones, investigations into subsistence, foraging mobility and landscape use are particularly intriguing. The region is home to numerous well-known Middle Paleolithic sites that have provided information on Neanderthal subsistence, mobility and land use (e.g., Zilhão et al., 2010; Nabais and Zilhão, 2019; Carvalho et al., 2018; Fernández-Laso et al., 2015; Zilhão et al., 2020; Nabais et al., 2023; Haws et al., 2020a). Understanding the prey selection behaviors of Neanderthals provides a multidimensional perspective on Neanderthal lifeways helping to better understand their ecological niche, social organization and survival strategies in dynamic and often challenging environments. For example, evidence of selective hunting or opportunistic foraging can provide clues about their flexibility in survival strategies across different landscapes. It can also provide information past ecological conditions and whether Neanderthals practiced sustainable hunting or over-exploited specific resources, which may have influenced local fauna and ecosystems.
In Portuguese Estremadura, where current understanding of Neanderthal subsistence is both chronologically fragmented and based on only three sites, Neanderthal diets focused mostly red deer and ibex, with inputs from a wide -array of other ungulates and small prey (e.g., Fernández-Laso et al., 2015; Nabais, 2018; Carvalho et al., 2024; Nabais and Zilhão, 2019; Boneta Jiménez et al., 2025; Cardoso and Cascalheira, 2024). The consistent abundance of red deer in these non-contemporaneous anthropic assemblages could reflect stability in the red deer populations. Such stability may relate to the refugial nature of southern Europe and Iberia in particular (e.g., Figueiral and Terral, 2002; Gomez and Lunt, 2007); conversely, it could relate to the ecological flexibility of red deer (e.g., Straus, 1981). Regardless, it suggests Neanderthal hunting was not sufficiently intensive to negatively impact red deer populations. During MIS 5, however, intensive exploitation of tortoises by Neanderthals evidenced at Gruta da Oliveira may have led to a localized extinction of this food resource at the beginning of MIS 4. This finding supports the notion of large enough Neanderthal population for overexploitation to occur (Nabais and Zilhão, 2019).
Lapa do Picareiro (Figure 1), a Paleolithic cave site situated in mountainous terrain of Estremadura, offers a unique opportunity to investigate these facets of behavior among the last Neanderthals in the region. The site contains a chronologically constrained multi-layer sequence of late Middle Paleolithic occupations with rich faunal assemblages, making it ideal for examining how Neanderthals navigated local landscapes and resources over time. With archaeological evidence spanning multiple occupation phases and accompanied by paleoenvironmental records, Lapa do Picareiro builds upon previous investigations into the relationship between subsistence choices and foraging mobility in Portuguese Estremadura.
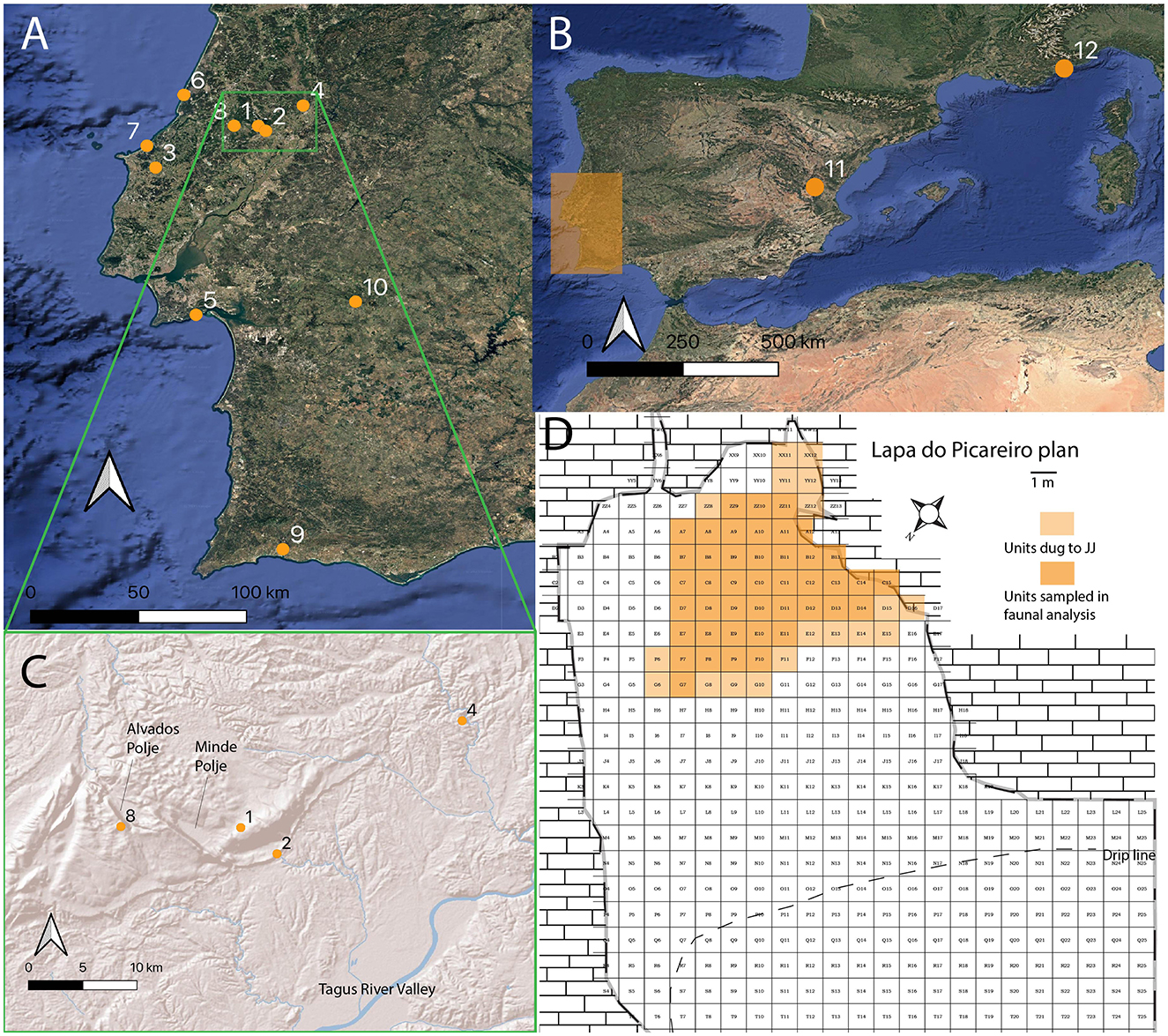
Figure 1. (A, B) Map showing the location of sites mentioned in the text: (1) Lapa do Picareiro located on the Serra d'Aire mountain, (2) Gruta da Oliveira, (3) Gruta Nova da Columbeira, (4) Gruta do Caldeirão, (5) Gruta da Figueira Brava, (6) Mira Nascente, (7) Praia Rei Cortiço, (8) Buraca Gloriosa, (9) Gruta da Companheira, (10) Gruta do Escoural, (11) Abrigo de la Quebrada, (12) Grotte du Lazaret. (C) Terrain inset map showing local geographic features near Picareiro mentioned in the text. (D) Site plan of Picareiro showing were Level JJ has been excavated and what units were sampled for this study.
This paper contributes to the growing regional record of Neanderthal subsistence and mobility strategies in Portuguese Estremadura by presenting new zooarchaeological and taphonomic data from multiple Middle Paleolithic occupations at Lapa do Picareiro. Using macro-mammalian faunal assemblages from three subunits of Level JJ, the latest Middle Paleolithic deposits at the site, we investigate subsistence and foraging mobility through time. These data are complemented by Picareiro's paleoenvironmental records, which allow for an evaluation of Neanderthal responses to climate change during MIS 3 (Benedetti et al., 2019; Carvalho et al., 2022).
We address the following research questions:
1. How did Neanderthals structure their subsistence at Picareiro, particularly in terms of prey selection?
2. What does the reconstructed Neanderthal subsistence, particularly their contribution to the faunal assemblages and prey choices, reveal about their movements across the landscape?
3. Do we see continuity or change and Neanderthal subsistence and foraging mobility in level JJ at Picareiro if change is observed, can this adaptation be linked to specific climate episodes?
2 Materials and methods
2.1 Study area and site
Portuguese Estremadura, a predominantly karstic region in central Portugal, is a key area for Middle Paleolithic research (Cardoso and Cascalheira, 2024), encompassing sites spanning MIS 5–3, such as Lapa do Picareiro (e.g., Bicho et al., 2003, 2009; Benedetti et al., 2019; Haws, 2006; Haws et al., 2020a, 2021), Gruta do Caldeirão (e.g., Zilhão et al., 2021a), Gruta da Figueira Brava (e.g., Antunes and Cunha, 1992; Raposo and Cardoso, 2000), Gruta da Oliveira (e.g., Zilhão et al., 2010, 2021b), Mira Nascente (e.g., Haws et al., 2020b), and Praia Rei Cortiço (e.g., Haws et al., 2010, 2020b). This region serves as a large-scale ecotone, where diverse marine ecosystems and terrestrial Mediterranean bioclimatic zones overlap (Haws et al., 2010). Its mosaic of montane islands, plains, and valleys supports Thermo- and Meso-Mediterranean vegetation and fauna, fed by the Tagus River Basin, associated drainages, and natural springs (Quézel, 1985).
During the Late Pleistocene, Estremadura was characterized by steppe and open mountain-type habitats, dominated by pine woodlands (González-Sampériz et al., 2010). These environments fluctuated between open and closed forested conditions in response to climatic variability (Carvalho et al., 2022). The region's proximity to the Atlantic Ocean, where ice-rafted debris floated during Heinrich events (Heinrich, 1988), suggests that cold and arid conditions during such events and other stadials could have been extreme, though direct evidence for their severity remains limited.
Neanderthal subsistence in Portuguese Estremadura is poorly understood due to the limited number of archaeofaunal assemblages directly linked to human activity (Nabais, 2018). However, evidence points to a broad diet dominated by cervids, supplemented by ibex and other ungulates such as horse and aurochs. Additionally, Neanderthals exploited small prey like rabbits (Mein and Antunes, 2000; Carvalho et al., 2018), tortoises (Nabais and Zilhão, 2019; Boneta Jiménez et al., 2025), crustaceans (Nabais et al., 2023), mollusks, and other marine resources (Zilhão et al., 2020). It has been suggested that this dietary breadth reflects a decreased need for high mobility strategies (Nabais, 2018).
Prey selection and raw material procurement strategies have led to the hypothesis that Neanderthals in this region practiced residential mobility within relatively restricted territories (Raposo, 1995; Zilhão, 2001). However, the scale of these territories remains poorly defined. A 87Sr isotope study from Gruta da Oliveira and Gruta do Caldeirão suggests Neanderthal territories may have covered ~600 km2, while later Upper Paleolithic humans operated within smaller territories of around 300 km2 (Linscott et al., 2023). These estimates are based on limited data, and it is unlikely that territory sizes or mobility patterns remained static over tens of thousands of years, particularly given the cycles of climatic change during this period. To better understand the subsistence and foraging mobility of Estremaduran Neanderthals, we need multi-occupation, site-based subsistence records with good chronological control, and stratigraphic integrity to assess the behavioral and temporal variability of Neanderthals.
Lapa do Picareiro (Figure 1) is one such site, located on the west-facing slope of the Serra d'Aire about 100 km northeast of Lisbon. It is the highest-altitude Paleolithic cave site in Portugal (570 m a.s.l.) and lies ~40 km from the modern coastline. Since 1994, excavations have revealed large faunal assemblages, lithic artifacts, and charcoal spanning more than 40 stratigraphic units, covering occupations from the Middle Paleolithic to the Bronze Age (Benedetti et al., 2019; Bicho et al., 2000, 2003, 2006, 2009; Bicho, 2004; Haws, 2006; Haws et al., 2019, 2020a, 2021). Substantial radiocarbon dating with no significant inversions provide robust chronological control for these sequences. The Middle Paleolithic deposits are well-stratified, minimally disturbed, and contain evidence of multiple Neanderthal occupations associated with small lithic assemblages, primarily characterized by discoidal technology (Haws et al., 2020a).
The lithic evidence suggests that most Paleolithic occupations at Picareiro were short-term, potentially due to the site's challenging accessibility, though more intensive occupations occurred during the Magdalenian (Bicho et al., 2003, 2006, 2009; Bicho and Haws, 2012). However, it remains unclear whether the faunal record supports similarly short-term Neanderthal occupations. With one of the largest Paleolithic faunal assemblages in Portugal, Picareiro has yielded tens of thousands of macro-, meso- and micro-mammal remains from Middle Paleolithic deposits alone. Additionally, abundant remains of birds, amphibians, and micromammals push the total number of skeletal remains even higher. Preliminary taphonomic studies indicate these assemblages were deposited by humans, carnivores, and raptorial birds (Haws et al., 2020a; Carvalho et al., 2023, 2024).
The stratigraphic sequence at Picareiro provides a strong paleoclimate signal. Variations in magnetic susceptibility and clast sizes align closely with the NGRIP curve (50–45 ka) and correspond to deep-sea pollen and sea surface temperature records (Benedetti et al., 2019). Isotopic zooarchaeology has identified Greenland Stadial and Interstadial signals in the sequence (Carvalho et al., 2022), corroborated by speleothem data from Buraca Gloriosa, a cave located <10 km away (Figure 1) (Denniston et al., 2018; Thatcher et al., 2020). Most archaeological horizons at Picareiro can be linked to specific climate events within MIS 3 (Benedetti et al., 2019), making the site ideal for examining how subsistence, foraging mobility, and landscape use responded to climatic downturns.
2.2 Level JJ
The assemblages analyzed in this study come from Level JJ, which represents the last Middle Paleolithic occupations at the site. This deposit, over 1 meter thick, dates to ~51.5–42.5 ka cal BP based on radiocarbon assays (Haws et al., 2020a). The lithic assemblage of Level JJ is comprised of primarily flakes and shatter, with few cores and one retouched piece (Haws et al., 2020a). Level JJ contains evidence of multiple Neanderthal occupation phases, with distinct lower and upper lithic horizons separated by a 25 cm-thick layer known as the JJ Dark Layer. This layer, composed of dark reddish-brown, organic-rich sediment, shows a notable decrease in lithic artifacts and faunal remains compared to the deposits above and below.
Picareiro's level JJ offers the opportunity to examine temporal and behavioral variability, as multiple phases of occupation have been observed. Level JJ and its associated faunal assemblages were divided into three subunits: Lower JJ (LJJ), JJ Dark Layer (JJD), and Upper JJ (UJJ), based on concentrations of archaeological materials (see Figure 2). These divisions, supported by stratigraphic integrity and age control, allow for assessments of subsistence and foraging mobility over time.
1. Lower JJ (LJJ): The base of Level JJ, ~62 cm thick, and can be characterized by medium clasts in a matrix of very muddy reddish brown fine sediment (Benedetti et al., 2019). Radiocarbon dates range from 51.5 to 44.1 ka cal BP. This subunit contains a lithic artifact concentration dominated by quartzite, quartz, and chert. Magnetic susceptibility data identifies a signal corresponding to Greenland Interstadial 13 near the top of LJJ. Stable isotope zooarchaeology suggests a temperate forest environment with stable vegetation, transitioning to slightly more forested conditions during this period (Carvalho et al., 2022).
2. JJ Dark Layer (JJD): Situated directly above LJJ, this 25 cm-thick subunit contains medium to large clasts in an organic- and clay-rich matrix that is dark reddish brown. It has been radiocarbon dated to 45.4–44.2 ka cal BP. It contains few lithic artifacts (n = 4) and limited charcoal remains but thousands of skeletal remains of mammals (ranging from micro- to large-sized), amphibians, and birds. Magnetic susceptibility and isotopic data indicate that JJD formed during Heinrich Event 5, a period of environmental disturbance (Benedetti et al., 2019; Carvalho et al., 2022).
3. Upper JJ (UJJ): The top 45 cm of Level JJ contains medium to large clasts in a reddish brown clay-rich matrix. It represents the latest Middle Paleolithic occupation at Picareiro, dating to 44.9–42 ka cal BP. The lithic assemblage is characterized by quartz and quartzite, reflecting a shift in raw material procurement strategies that may indicate adaptation to the climatic instability of Heinrich Event 5 (Carvalho et al., 2022). Magnetic susceptibility data suggests signals for Greenland Stadial/Interstadial phases 12, 11, and 10, while isotopic data indicates fluctuating vegetation regimes, transitioning between open and closed temperate forests (Benedetti et al., 2019; Carvalho et al., 2022).

Figure 2. Composite image of a stratigraphic profile showing the dark layer in Level JJ.
2.3 Methodology
2.3.1 Excavation methodology
The cave was mapped using a 1 × 1 m grid system. Each unit was excavated according to natural stratigraphy and 5 cm artificial levels within the geologic strata. All artifacts, bones (larger than 2 cm), and features were mapped in three dimensions using a total station. All sediment was sieved through nested 2 and 4 mm mesh screens to separate the finer sediment from the larger fraction which is mostly limestone éboulis. The larger fraction was sorted in the field. The remaining sediment in the 2 mm screen was water-sieved in the laboratory in order to recover small bones, lithics, and macrobotanical remains. This allowed for the recovery of small rabbit and micromammal elements, fish bones, shell fragments, stone chippage, personal ornaments and charcoal.
2.3.2 Zooarchaeological and taphonomic analysis
To reconstruct late Neanderthal subsistence and foraging mobility, we conducted zooarchaeological and taphonomic analyses on a sample of macro-mammalian remains from the three subunits of Level JJ. The skeletal remains analyzed included both in situ piece-plotted specimens and those recovered through sieving, with different taphonomic data recorded for each recovery method.
To investigate Neanderthal subsistence at Picareiro, we generated a comprehensive zooarchaeological and taphonomic dataset for the faunal assemblages of each subunit. Prey selection is assessed through species composition of each subunit. Identification of faunal remains was performed at the Interdisciplinary Center for Archaeology and Evolution of Human Behaviour (ICArEHB) at the Universidade do Algarve. Taxonomic and anatomical identifications were made using skeletal reference collections at the ICArEHB OsteoArchaeology Lab, supplemented by digital and printed atlases (e.g., Alimen and Lavocat, 1966; Hillson, 1996; Pales and Lambert, 1971; Schmid, 1972). Data entry was facilitated using the E5 configurable data entry program for archaeology (McPherron, 2022).
For each specimen, we recorded taxonomy, skeletal anatomy, portion, landmarks, and side, identifying remains to the lowest taxonomic level possible. Indeterminate fragments were categorized by morphology and size into large (300+ kg), medium (60–300 kg), or small (30–60 kg) ungulate size groups, based on cortical bone thickness and specimen dimensions.
We used the Number of Identified Specimens (NISP) as the primary analytical unit, following Grayson (1984) and Lyman (2008). Because the Picareiro dataset combines faunal remains from both piece-plotted and sieved material, which were excavated in arbitrary spits, and excavations are ongoing (i.e., the analysis presented here is based on a sample), derived measures like Minimum Number of Individuals or Minimum Number of Elements likely to be misleading. While NISP can also be misleading (in particular, issues of differential fragmentation and diagenesis can artificially inflate its numbers), in datasets with limitations such as the Picareiro dataset has, it is the preferred unit so long as these potential impacts are taken into account in drawing any conclusions (see Discussion in Lyman, 2008).
To distinguish between the contributions of different taphonomic agents, we recorded bone surface modifications and overall bone condition to distinguish what remains were deposited by humans vs. non humans. Observations included both anthropogenic modifications (e.g., cut marks, percussion marks, breakage patterns, fragmentation, and burning) (Blumenschine, 1988; Pérez Ripoll, 1992; Fisher, 1995) and non-human modifications (e.g., digestive traces from both carnivores and raptorial birds, gnaw marks, and punctures) based on guidelines from Andrews (1990), Cruz-Uribe (1991), Fisher (1995), and Hockett (1996).
Human-related bone surface modifications provide insight into the prey selection behaviors of Neanderthals as well as the carcass processing strategies taking place at the site. Impact and percussion marks were recorded following the guidelines outlined by Vettese et al. (2020), with totals summarized for each subunit. All potential cut marks and burn marks were examined under a Dino-Lite Edge portable microscope (model: MSAM7915MZT) and Nikon microscope (model:SMZ800N). Burn color was recorded only for piece-plotted remains according to Cáceres (2002). Breakage patterns were classified as green, dry, modern and indeterminate fracture types for the most dominant fracture edge type for each specimen but were only recorded for piece plotted remains. Green fractures are indicative of fresh bone breakage during human or carnivore processing while dry fractures signal post-depositional breakage, a common observation in natural accumulations (Villa and Mahieu, 1991).
To assess the degree of fragmentation, which can indicate intensive human manipulation of bones (e.g., Binford, 1978), we measured and categorized piece-plotted remains into five size classes: 0–2.5, 2.5–5, 5–10, 10–20, and >20 cm. This analysis provides insight into the intensity of Neanderthal carcass processing and subsistence behaviors.
Species richness (the number of different taxa in an assemblage) provides insights into diet breadth, while species evenness (the relative abundance of each taxon) can reveal patterns of environmental use and hunting frequency (Jones, 2004). For example, a low evenness value could indicate reliance on a single or dominant prey species, suggesting systematic targeted hunting in specific environments. Conversely, higher evenness might suggest more generalized foraging or seasonal variation in prey choice. Variations in species richness and evenness can also reflect differences in environmental conditions, or broader hunting strategies (Jones, 2004; Grayson and Delpech, 2002; Lyman, 2008; Starkovich, 2017; Stiner, 2001). Assemblages with high evenness might indicate consistence use of diverse habitats, while changes in evenness could suggest shifts in Neanderthal mobility, resource availability or climatic influences. By integrating these measures, we aim to better understand the relationship between Neanderthal subsistence and their movements on the landscape. We assess species evenness through the reciprocal of Simpson's Dominance Index (Simpson, 1949; Faith and Du, 2018).
Do we see continuity or change and Neanderthal subsistence and foraging mobility in level JJ at Picareiro if change is observed, can this adaptation be linked to specific climate episodes?
Besides prey selection, we also assess the continuity (or lack thereof) of carcass processing strategies observed among the subunits. Smaller fragments can be an indicator of greater processing effort or post-depositional fragmentation. We thus assess fragmentation size using a Mann-Whitney U Test for meaningful comparisons between two subunits (VanPool and Leonard, 2011). This test evaluates whether the distributions of fragment sizes differ significantly between groups based on ranks, which is particularly relevant for assessing relative fragmentation intensities. In this analysis we use Mann-Whitney to identify whether more intensive bone processing occurred in specific subunits. We set alpha at 0.05 and used the PAST4 software (Hammer et al., 2001) for all statistical tests.
3 Results
Of the 5,371 specimens analyzed in this study (Table 1), 1,159 came from LJJ, 813 in JJD, and 3,399 in UJJ. The highly fragmented assemblages made identifiability low, with the most abundant taxonomic categories being the M-sized ungulate: 42.5% of the NISP in LJJ, 80.9% in JJD and 55.8% in UJJ fall within this category. Of the remains identified to species or genus level, red deer was the most dominant in all three subunits, representing 9.1% of the LJJ assemblage, 2.7% in JJD and 5.8% in UJJ (Figure 3). Ibex was the second most dominant taxon identified in LJJ (1.4%) and UJJ (5.8%). Lower representations of other ungulates like aurochs, chamois, roe deer, wild boar, and horse were observed in all assemblages. Similar values of the Simpson's reciprocal index in LJJ (1/D = 1.80), JJD (1/D = 1.74) and UJJ (1/D = 1.67) indicate consistent interassemblage evenness.
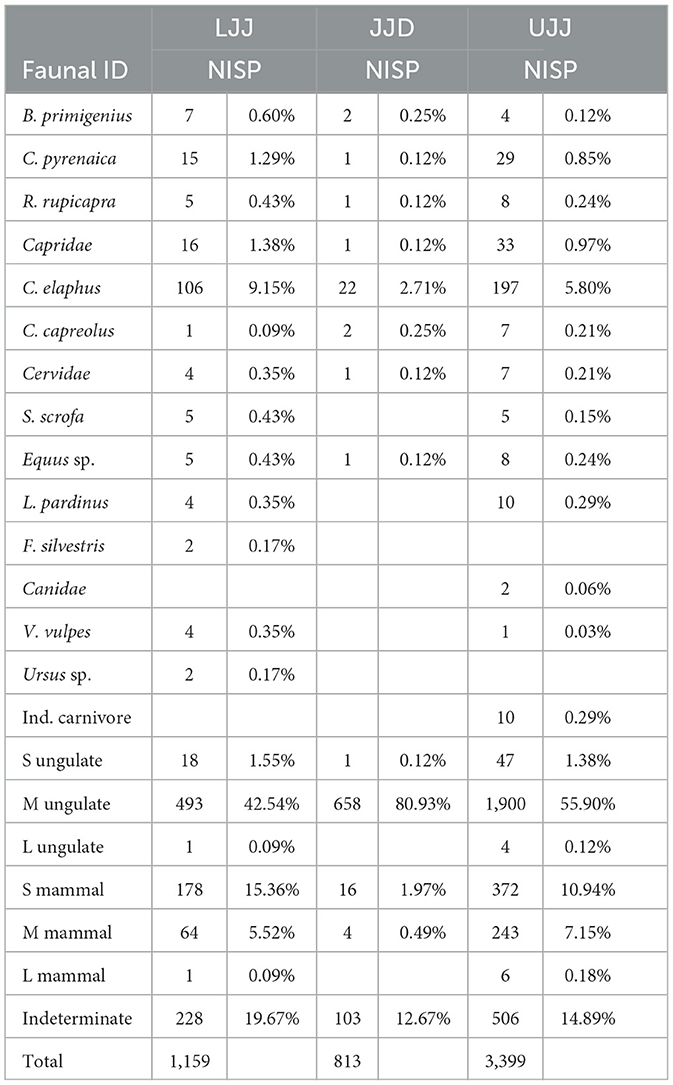
Table 1. Numbers of identified specimens (NISP) and percentages of taxa identified in LJJ, JJD and UJJ units.
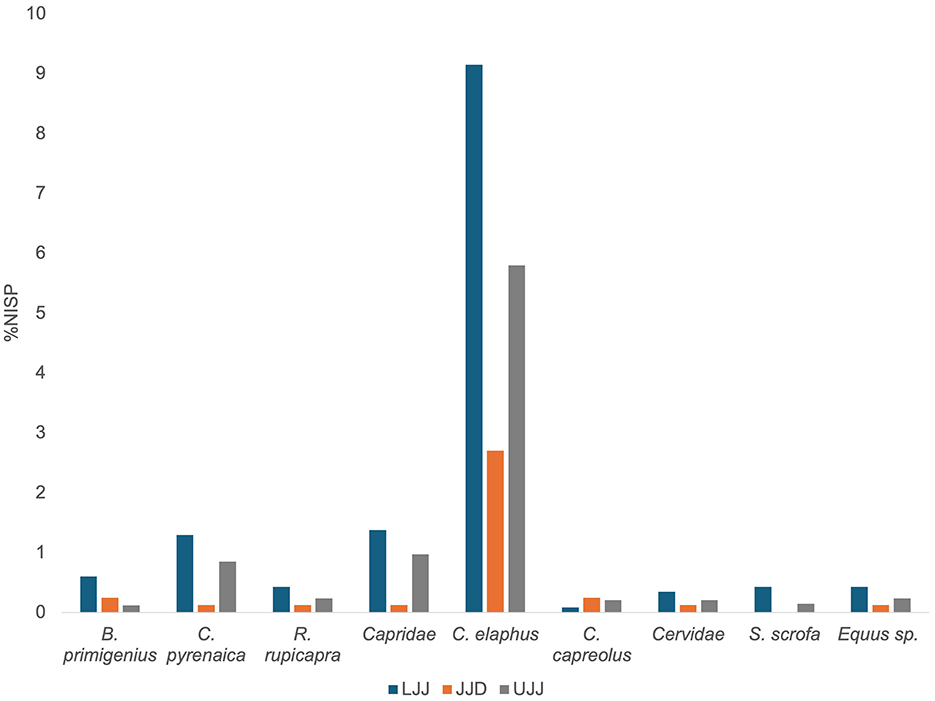
Figure 3. %NISP of ungulate taxa identified in the LJJ, JJD, and UJJ assemblages. Ungulate taxa identified were B. primigenius (aurochs), C. pyrenaica (Pyrenean ibex), R. rupicapra (chamois), C. elaphus (red deer), C. capreolus (roe deer), S. scrofa (wild boar), and Equus sp. (horse sp.).
Carnivore remains are rare. Low incidences of lynx (L. pardinus), wild cat (F. silvestris), canids, including red fox (V. vulpes) and bear (Ursus sp.) were identified in the LJJ and UJJ assemblages. No carnivores were identified in the JJD assemblage.
The representation of skeletal elements of ungulate taxa is presented in Table 2 (see Supplementary Table 1 for carnivore skeletal element representation). In the case of ungulates, elements of the appendicular skeleton are the most abundant (Table 2). For the most dominant taxon, red deer, metapodials and podials are the most abundant skeletal elements in all three assemblages (Figure 4), though cranial, axial and appendicular elements are all represented. In addition to the metapodials and podials, in all assemblages, the mandible, humerus, tibia and vertebrae were common in all three assemblages. Red deer scapulae were only encountered in the UJJ assemblage.
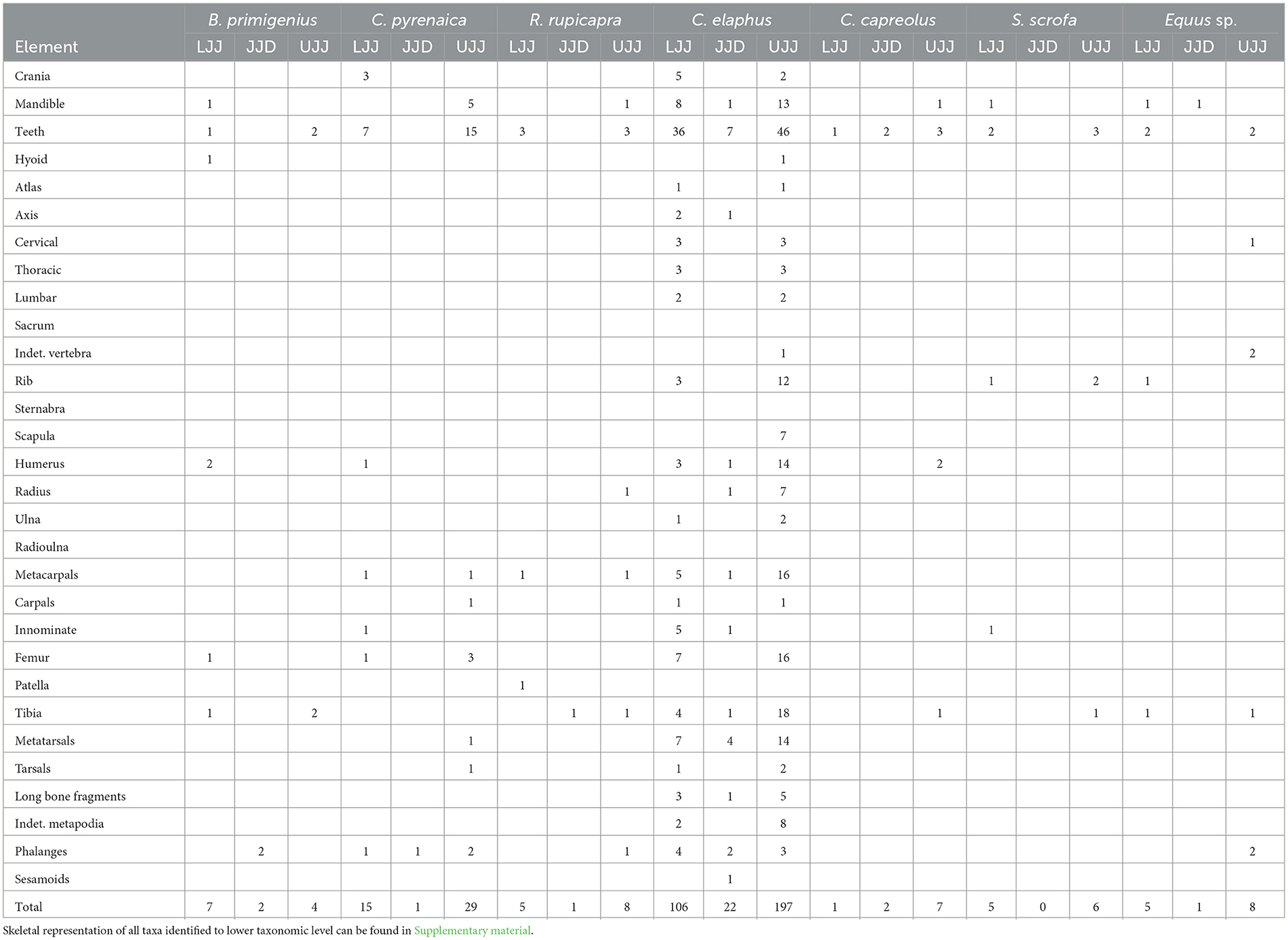
Table 2. Number of identified specimens of skeletal element representation of the ungulate taxa identified in the LJJ, JJD, and UJJ assemblages.
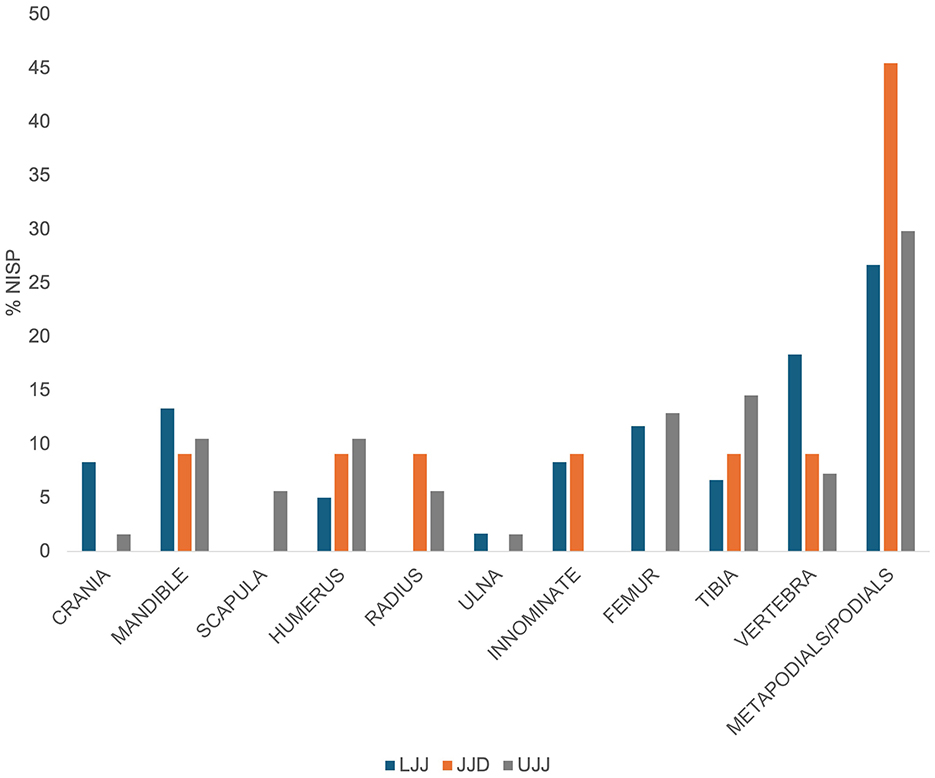
Figure 4. %NISP of aggregated skeletal element representation of Cervus elaphus.
The bone surface modifications and breakage patterns typically associated with human (i.e., cutmarks, percussion marks, and burn marks) are present in all three assemblages to varying degrees (Table 3; Figure 5). The highest incidence of cutmarked specimens was observed in LJJ (2.4% of NISP), followed by UJJ (1.7%) then JJD (0.5%). In the JJD assemblage, cutmarks are only observed on red deer and in the medium-sized mammal and ungulate categories. In addition to red deer, cutmarks were encountered on aurochs and horse specimens in the LJJ and UJJ assemblages and ibex and chamois in UJJ (Figure 6). In the assemblages, cutmarks are most frequent on the appendicular and cranial portions of the skeleton, with cutmarks observed on a few ribs and thoracic vertebrae in the C. elaphus and medium taxon size category (Supplementary Table 2). Supplementary data on the frequency of cutmarks by skeletal element is available in Supplementary Table 2.
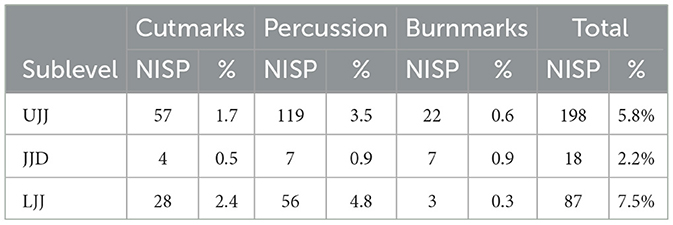
Table 3. Number of identified specimens displaying presence of cutmarks, percussion marks, or burn marks in the LJJ, JJD, and UJJ assemblages.

Figure 5. Types of bone surface modifications typically encountered in the Level JJ assemblages. (A, B) Burned bones (different stages of burning); (C) Percussion mark and green spiral fracture; (D) Cutmarks on rib fragment; (E) Punctured and digested phalanx; (F) Crush mark resulting in a puncture.
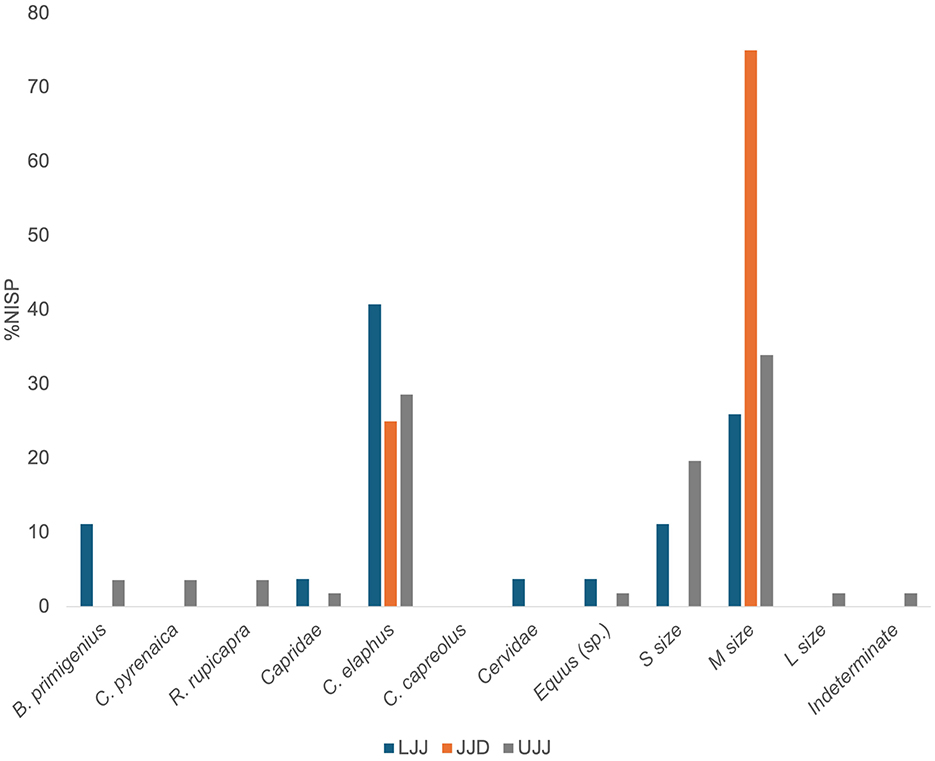
Figure 6. %NISP carrying cutmarks by taxon. Cutmarks were encountered on B. primigenius (aurochs), C. pyrenaica (Pyrenean ibex), R. rupicapra (chamois), C. elaphus (red deer), C. capreolus (roe deer), S. scrofa (wild boar), Equus sp. (horse sp.), cervids, caprids, small, medium and large ungulates, and indeterminate fragments.
Percussion marks (notches, conchoidal fractures, etc.) were the most common anthropic marker encountered in the assemblages, with the highest incidences occurring in LJJ (4.8%), followed by UJJ (3.5%) and <1% of the JJD assemblage showing percussion marks. In all three assemblages red deer, small and medium sized mammal specimens showed the most evidence of percussion (Figure 7). Burning was the least common anthropic bone surface modification, with <1% of total NISP of each assemblage showing any evidence of burning (Figure 8). Of the plotted specimens with recorded burn color and stage, two exhibited stage 2 (brown), five showed stage 3 (black, charred), and one displayed stage 4 (gray). Anthropic bone surface modifications on red deer are most observed on the appendicular elements and mandibles, with occasional anthropic marks observed on the axial portion of the skeleton (Figure 9).
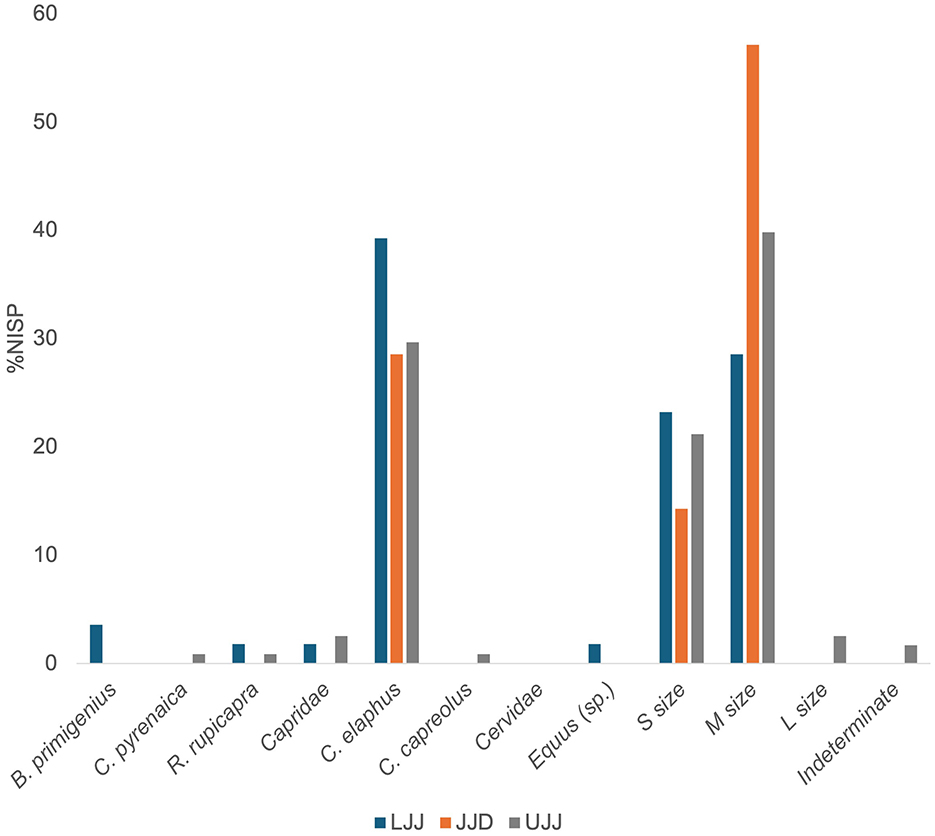
Figure 7. %NISP carrying percussion mark by taxon. Percussion marks were encountered on were encountered on B. primigenius (aurochs), C. pyrenaica (Pyrenean ibex), R. rupicapra (chamois), C. elaphus (red deer), C. capreolus (roe deer), S. scrofa (wild boar), Equus sp. (horse sp.), cervids, caprids, small, medium and large ungulates, and indeterminate fragments.
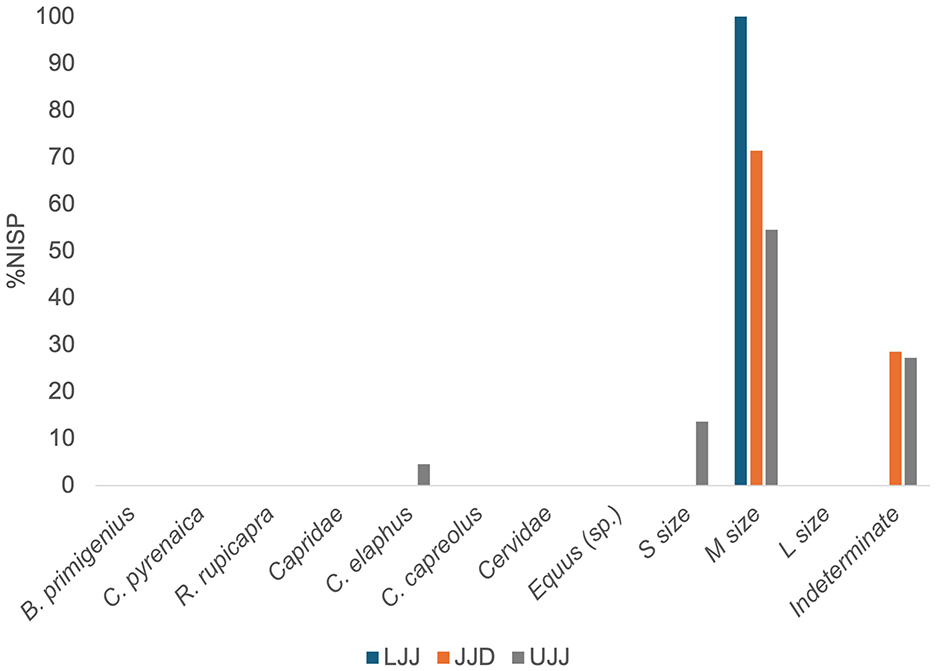
Figure 8. %NISP carrying burn marks by taxon. Burn marks were only encountered on C. elaphus (red deer), small medium and large ungulate categories, and indeterminate fragments.
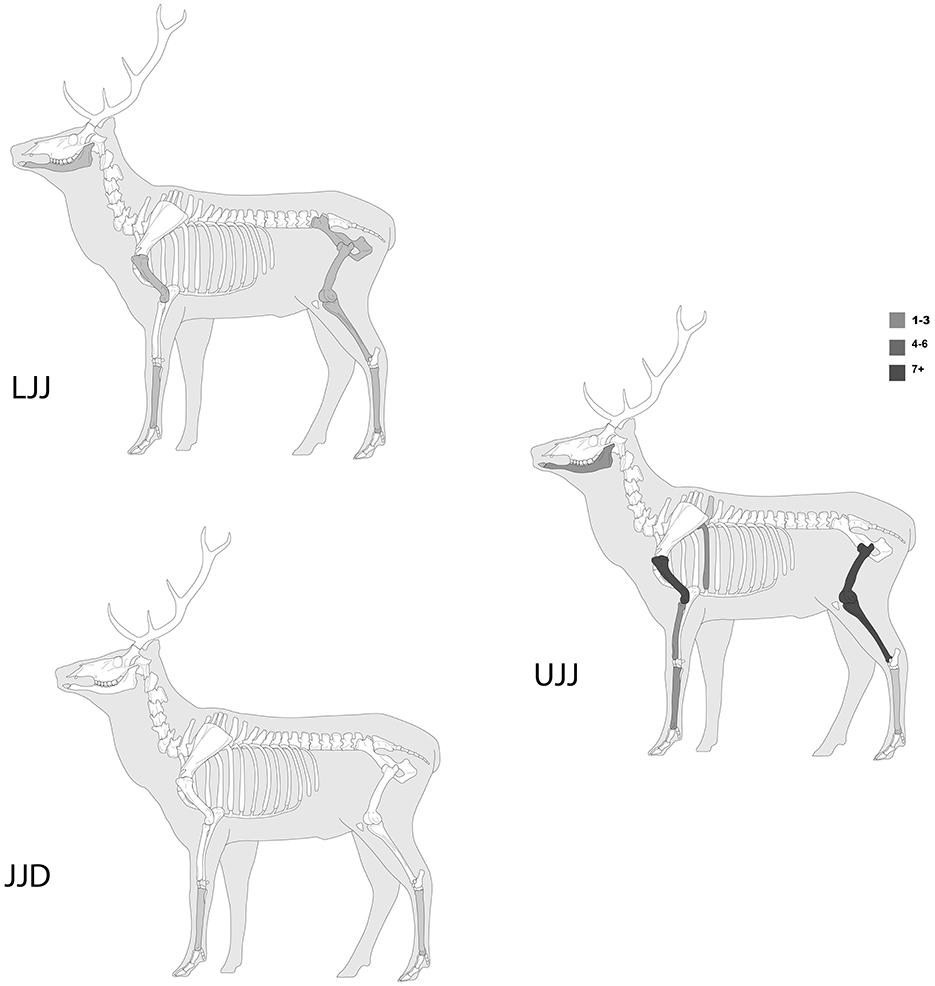
Figure 9. Schematic showing the frequency of red deer elements with anthropic modifications (cut marks, burning, or percussion marks) in LJJ, JJD, and UJJ. Only specimens with anthropic modifications are represented.
The assemblages are all highly fragmented (Figure 10). The Mann-Whitney U test showed no significant results (LLJ to JJD: U = 2.5, p = 0.1316, r = 0.17; JJD to UJJ: U = 2, p = 0.1034, r = 0.08; LJJ to UJJ: U = 6.5, p = 0.2492, r = 0.045; a Bonferroni correction was applied for all tests). Breakage patterns are dominated by green fractures in all assemblages, with more than 60% of the piece plotted remains showing green fractures and roughly 20% showing dry fractures (Figure 11) in all subunits.
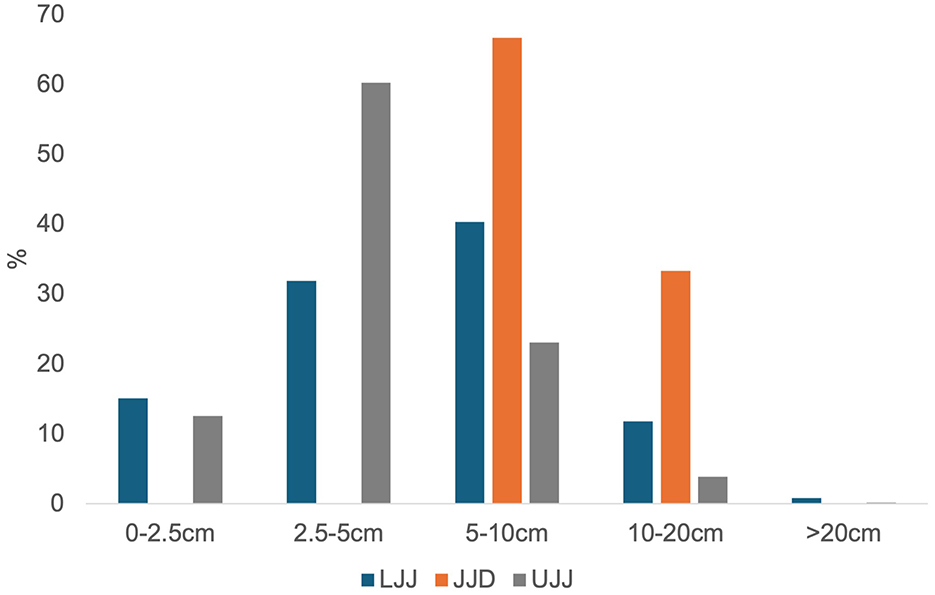
Figure 10. Percentage of piece plotted remains identified per specimen size category of LJJ, JJD, and UJJ assemblages.
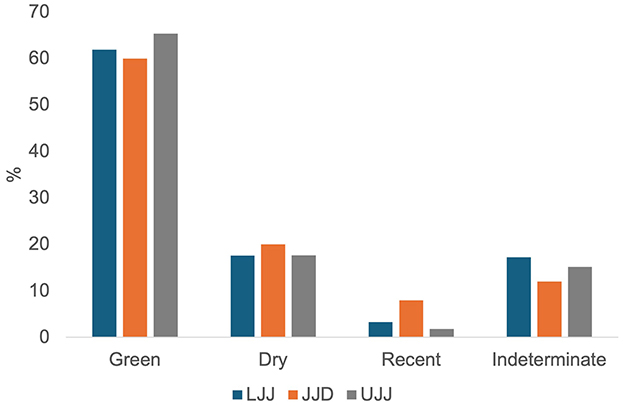
Figure 11. Percentage of piece plotted remains in breakage pattern profiles categories in LJJ, JJD, and UJJ assemblages.
Non-human bone surface modifications were encountered in small quantities, with similar proportions in the JJD and UJJ assemblages (Table 4; Figure 5), with the highest frequencies of non-human marks in LJJ (2.5% punctures, 1.2% gnawing, 3.6% digested).
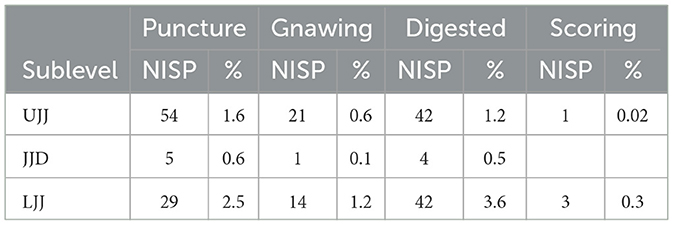
Table 4. Number of identified specimens displaying the presence of punctures, gnawing, digestion and carnivore scoring.
4 Discussion
4.1 How did Neanderthals structure their subsistence at Picareiro, particularly in terms of prey selection?
The results presented here suggest that Neanderthals contributed significantly to the formation of the faunal assemblages at Picareiro, although non-human processes also played a lesser role in JJD and UJJ. Non-human modifications, such as punctures and digested remains, were most prevalent in the LJJ assemblage, though in small quantities. Generally, the higher frequency of anthropic marks compared to non-human indicators in the assemblages suggests that humans were the primary agents in shaping their accumulation.
The level JJ assemblages at Picareiro also displayed similar species richness and evenness, suggesting similar taxonomic composition. Prey selection during the late Middle Paleolithic at Picareiro is preferential toward red deer across all three occupation phases. Red deer remains showed consistent evidence of anthropic modification, reflecting a potential preference for the prey item typically observed among Neanderthals at Iberian sites (e.g., Straus, 2013; Marín et al., 2017; Marín-Arroyo and Sanz-Royo, 2021; see also references in Carvalho et al., 2022). Though a slight preference for red deer by Neanderthals seems evident at Picareiro, given that the small- and medium-sized categories also displayed high incidence of anthropic marks in all three subunits and the high degree of fragmentation observed in the assemblages, it is possible future proteomic and zooarchaeological analyses will increase the proportions of non-red deer ungulates into their diet. It is also possible future proteomic and zooarchaeological analysis of the heavily fragmented portion of the assemblage reflects a higher incidence of red deer, demonstrating a hunting strategy that is heavily focused on one prey item like that identified at sites in more northern latitudes where reindeer dominate assemblages (80–90% of NISP) (e.g., Costamagno et al., 2006; Discamps and Faivre, 2017; Rendu et al., 2023). Though rare, other ungulates also displayed anthropic bone surface modifications in all subunits, suggesting supplementation of a red deer-focused diet by aurochs, ibex, chamois, and horse. Currently, small and medium-sized categories do not provide much insight into prey selection.
At Picareiro, if the preference for red deer as a dietary resource is confirmed with future data, this would be indicative of a hunting behavior characterized by selection and not opportunism. The concept of selection refers to a human population's ability to recognize its needs and devise targeted strategies to meet them effectively (Rendu, 2022). This process involves anticipating future requirements, planning activities and utilizing appropriate technological tools to achieve these objectives (Rendu, 2022). Thus, at Picareiro, it seems that late Neanderthals occupying Picareiro had knowledge of the presence of red deer populations in this area, and perhaps visited the site in part to exploit this resource. The riverine valleys to the south of Picareiro, intermontane valleys, temperate forested environments (Carvalho et al., 2022), and polje that may have been a paleolake during the Late Pleistocene (Rodrigues, 1991) may have supported relatively stable red deer populations that Neanderthals could exploit within a 20 km radius of the site. On the other side of the coin, if future data suggest no preference for red deer but an exploitation of various ungulates in equal or similar proportions, an opportunistic hunting strategy would be assumed. Given Picareiro's location in an ecotone with proximity to various habitats, it could be that Neanderthals visited the site as a camp to exploit diverse hunting patches.
The skeletal representation across all taxonomic categories is dominated by appendicular elements, with cranial fragments being less frequent and axial elements rarely present. For red deer, the most abundant taxon and the one showing the highest incidences of anthropic bone surface modifications, this pattern holds true (Figure 9). This suggests that the appendicular skeleton, which includes meat-rich and marrow-bearing parts, was frequently transported to the cave for processing and consumption. Cranial fragments, which are nutrient and calorie rich, were also brought to the cave.
Axial elements are nearly absent across all ungulate taxa, with only one or two specimens identified for horse and wild boar, and slightly more for red deer (Table 1). This pattern may reflect selective transport of high-utility body parts to the cave, as carrying whole carcasses up the steep terrain would have been challenging. However, zooarchaeological studies emphasize that skeletal element abundances must be evaluated in light of density-mediated attrition, as post-depositional processes can heavily bias skeletal representation. Low-survival elements such as vertebrae and ribs are highly susceptible to weathering, microbial decay, and mechanical destruction from trampling, significantly reducing their likelihood of preservation (Lyman, 1994, 2021; Stiner, 2002; Faith and Thompson, 2018). This attritional overprint can obscure original transport decisions, meaning that the skeletal pattern observed at Picareiro may not solely reflect human selection. While carnivore modifications are not frequent in the assemblage, gnawing and digestion traces suggest that occasional scavenging may have further contributed to the depletion of fragile axial elements. Despite this, the presence of elements representing the entire red deer skeleton at Picareiro indicates that whole carcasses may have occasionally been brought to the cave. Even so, given the potential effects of post-depositional attrition, we focus our behavioral interpretations primarily on high-survival skeletal elements to minimize taphonomic bias. The palimpsestic nature of the site may also contribute to the observed pattern, as repeated occupations could have resulted in differential preservation and secondary displacement of skeletal elements.
Analysis of anthropic bone surface modifications at Lapa do Picareiro reveals various carcass processing activities. Cutmarks are most frequently observed on meat-rich limb elements (Supplementary Table 2), suggesting meat fileting and stripping (e.g., Binford, 1978, 1981; Bunn et al., 1986; Dominguez-Rodrigo, 1997). Cranial fragments with cutmarks indicate attempts to access nutrient- and fat-rich resources, such as the brain (e.g., Lyman, 2005). Mandibular cutmarks on red deer, aurochs, and ibex suggest butchering focused on tongue and cheek meat extraction (e.g., Dominguez-Rodrigo, 1997) or as part of the marrow extraction process (Blumenschine and Marean, 1993). Less commonly, cutmarks on an ibex phalanx and a red deer incisor root suggest specialized butchery practices. Cutmarks on phalanges, which are low-yield elements with little muscle tissue, may indicate hide removal (Binford, 1981) or intensive carcass processing (Lyman, 2005). Similarly, cutmarks on tooth roots are rare but may reflect removal of mandibular or maxillary flesh, particularly tongue and cheek meat, leaving traces on the mandible, maxilla, and occasionally the tooth roots (e.g., Dominguez-Rodrigo, 1997). Cutmarks on red deer and medium-sized taxa ribs suggest activities related to fileting and meat removal (e.g., Binford, 1981), trimming fat or sinew (Lupo and O'Connell, 2002), or accessing internal organs.
At Picareiro, the presence of cutmarks on meat- and nutrient-rich portions of the skeleton suggests that Neanderthals had primary access to carcasses, rather than scavenging remains left by other predators. However, both ethnographic and archaeological evidence indicate that butchery strategies varied significantly, depending on environmental conditions, dietary needs, and cultural preferences (e.g., Dominguez-Rodrigo, 1997; Lyman, 2005; Lupo and O'Connell, 2002). On-going analyses on cutmark location and morphology will help unravel carcass exploitation and transport strategies.
Archaeological and ethnographic evidence points to humans as primary depositional agents in assemblages with processed long bones and skulls to extract marrow and contents of the brain cavity (e.g. Blumenschine, 1995; Blumenschine and Marean, 1993; Dominguez-Rodrigo, 1997). Percussion marks, typically associated with marrow extraction activities (see Discussion in Vettese et al., 2020) were identified in all three subunits indicating that Neanderthals at Picareiro maximized their calorie intake from these carcasses.
Evidence of burning which can be an indicator roasting or using bone fuel (e.g., Binford, 1978; Costamagno et al., 2006; David, 1990) is severely limited. Of the piece plotted remains where burning color or stage are recorded, two brown specimens (Stage 2) indicated exposure to low-temperature fires (200–300°C) which may be indicative of low-intensity or indirect heat exposure, five charred black (stage 3) specimens suggesting exposure to fires with moderate heat (300–500°C), and one gray calcined (stage 4) bone that was burned at higher temperatures (~600°C and above) (Cáceres, 2002). With such scarce data, little can be said about whether burn marks observed in the Picareiro assemblage relate to intentional cooking activities or simply their proximity to fires. Spatial analysis and categorization of the burned specimens encountered during sieving will be critical for resolving this issue, but at this time, we cannot confidently interpret these remains as being a result of anthropogenic burning.
The slightly higher frequencies of red deer exploitation at Picareiro mirrors other Neanderthal-affiliated zooarchaeological assemblages in Portuguese Estremadura where red deer is the taxon that is most exploited (e.g., Cardoso et al., 2002; Zilhão et al., 2010; Brugal and Raposo, 1999). However, given the very limited and temporally fragmented subsistence record in the region, whether this is a regional pattern needs further investigation through the analysis of Neanderthal-affiliated zooarchaeological assemblages from other sites. Picareiro is unique, however, in that its faunal assemblages relating to the Middle Paleolithic studied thus far show no evidence of extinct Pleistocene fauna which are present at Gruta da Oliveira, a site located on the other side of the Serra d'Aire, during MIS5 (Zilhão et al., 2010). In addition, preliminary results of an ongoing analysis of meso-fauna from Level JJ at Picareiro (Carvalho et al., 2023, 2024) indicate that with the exception of rabbits, exploitation of other small prey items such as tortoises or aquatic taxa by Neanderthals have not been identified thus far. This is in stark contrast to sites like Figueira Brava and Gruta da Oliveira, where regular exploitation of tortoises and marine prey by Neanderthals has been identified (e.g., Nabais et al., 2023; Nabais and Zilhão, 2019; Zilhão et al., 2020). Future analyses at Picareiro and recently excavated sites like Gruta da Companheira (e.g., Barbieri et al., 2023) and Gruta do Escoural will provide a better picture on Neanderthal subsistence behaviors in westernmost Iberia.
Neanderthals at Picareiro appear to have targeted prey similar to that found at other Iberian sites, primarily focusing on equids, cervids, and caprids. This pattern seems largely driven by the availability of these animals in the surrounding landscape (e.g., Eixea et al., 2020; Marín-Arroyo and Sanz-Royo, 2021; Yravedra et al., 2016; Nabais, 2018; Sánchez-Hernández et al., 2020a,b). For example, red deer are often the most common taxon in zooarchaeological assemblages across many Iberian regions (e.g., Blasco, 2008; Blasco and Peris, 2012; Salazar-García et al., 2013; Marín-Arroyo and Sanz-Royo, 2021; Ruiz et al., 2021; Yravedra et al., 2016). In contrast, sites in high-altitude alpine settings frequently exhibit a dominance—or even a specialization (exceeding 70% of the assemblage)—of either ibex or chamois (e.g., Yravedra et al., 2014; Real et al., 2019; Yravedra and Cobo-Sánchez, 2015; Yravedra, 2013). Additionally, in more open Pleistocene environments, such as at Abrigo de la Quebrada in Valencia, there is a clear preference for equid remains (Real et al., 2019). These observations support the hypothesis that, like other Neanderthal groups across Iberia, those at Picareiro exploited locally available fauna, with their prey selection primarily influenced by the immediate environmental context (see Discussion in Jones and Carvalho, 2023).
4.2 What does the reconstructed Neanderthal subsistence reveal about their movements across the landscape?
Assessing the duration of site occupations often involves analyzing site formation processes and the density of archaeological materials, the presence of living floors or structures, and the types of modifications observed on those materials (e.g., Stiner, 2013; Marín et al., 2019). However, most archaeological sites are palimpsests—composites of materials accumulated through repeated occupations rather than discrete single events (Bailey, 2008; Bailey and Galanidou, 2009), meaning that distinguishing between short- and long-term occupations isn't straightforward, and requires multiple lines of evidence (Moncel and Rivals, 2011). Lithic indices proposed for assessing site occupation durations based on tool the presence of retouched tools (Riel-Salvatore and Barton, 2004) have shown contradicting results (Bicho and Cascalheira, 2020), demonstrating the need for multiple lines of evidence.
At Lapa do Picareiro, the evidence for occupation appears somewhat contradictory. The small lithic assemblage from Level JJ—dominated by flakes, some shatter, and very few formal or retouched tools (Haws et al., 2020a)—suggests a low-density human presence. Moreover, the predominance of flakes over other tool types may indicate a segmented chaîne opératoire, a pattern often associated with short-term occupations (Naudinot and Marchand, 2020), where a more segmented sequence typically reflects briefer site use. However, further detailed lithic analyses are needed at Picareiro to confirm this interpretation.
In addition, the limited input from both humans and non-human agents (such as carnivores and predatory birds), along with the presence of carnivore taxa within the assemblages, supports the idea of intermittent human activity interspersed with animal use of the cave. This, combined with the general absence of features like stacked hearths, structures, or living floors—elements that would signal more intensive, long-term occupation—reinforces the hypothesis that the cave was used for relatively short durations.
It is important to note, however, that the site's low sedimentation rate (0.1–0.3 mm a−1; Benedetti et al., 2019) indicates a palimpsest deposit. Thus, additional spatial, lithic, and geoarchaeological data are required to conclusively determine the duration of occupations at Picareiro. Based on the current evidence, we hypothesize that the occupations were brief rather than intensive.
Neanderthal prey selection at Picareiro reflects a primarily local mobility strategy center on exploiting the site's location in an ecotonal environment. Red deer, the dominant prey species across all subunits, though sometimes used as indicator of forested environments, have been shown to display dietary and ecological flexibility, foraging in riverine valleys, forests and alpine environments during the winter (Schmid, 1972; Hofmann, 1989; Fonseca, 1998; Azorit et al., 2012; Berlioz et al., 2017; Gebert and Verheyden-Tixier, 2001; Sánchez-Hernández et al., 2020a,b; Straus, 1981). The location of Picareiro (Figure 1) fits this description as it is characterized by a large-scale ecotone with mountainous, valleys and plains. Several water sources, the Poljes of Mira-Minde (>5 km away) and Alvados (20 km away), and the Tagus river valley and its tributaries could have supported red deer and other ungulate populations.
Red deer populations are generally sedentary, with some modern populations migrating seasonally to access food, reduce competition, or avoid snow in higher latitudes (e.g., Mysterud et al., 2011). However, archaeological evidence suggests that such movements were relatively limited in the past (Steele, 2002). 87Sr analysis of red deer from MIS 6 at Lazaret Cave (France) indicates that while some individuals moved seasonally, their travel distances did not exceed 20 km (Barakat et al., 2023). In Portuguese Estremadura, similar 87Sr studies further support the presence of red deer year-round during MIS 5, with no evidence of long-distance migration (Linscott et al., 2023). Given the c. 680 m elevation of the Serra d'Aire, any localized movements by red deer in this region were likely minor and driven by food availability rather than adverse climatic conditions (Linscott et al., 2023). If red deer populations were present year-round during both MIS 5 and MIS 3, this continuity suggests a longstanding healthy demography among Pleistocene red deer populations in the region, with localized groups persisting in the region across climatic fluctuations. These findings indicate that Neanderthals at Picareiro had a stable and predictable red deer resource throughout the year, reinforcing the interpretation that this species played an important role in their subsistence strategy.
The presence of ibex and occasional chamois in the assemblages suggests that Neanderthals may have frequented mountainous environments to hunt these species (Acevedo and Cassinello, 2009; Nesti et al., 2010). While ibex has been found at Paleolithic sites in Iberia across a range of settings, from montane to coastal environments, chamois are typically associated with steeper, high-altitude habitats (Yravedra and Cobo-Sánchez, 2015). Although Neanderthals specialized in hunting alpine taxa in classic rocky, high-altitude settings, sites where ibex and chamois appear in smaller frequencies do not necessarily need to be mountainous (Yravedra and Cobo-Sánchez, 2015), as seen at Picareiro. Nevertheless, the inclusion of these taxa in the assemblage raises the possibility of movements to local mountainous areas such as the Serra d'Aire, where Picareiro is located, or to other nearby ranges like Montejunto or Serra da Estrela, situated 30–40 km southwest and northeast of the site, respectively.
The presence of aurochs and horse among the anthropic remains suggests that Neanderthals ventured into more open environments to hunt these species. Aurochs, which typically forage in low-lying, semi-open riverine areas (Hall, 2008), may have drawn Neanderthals to the riverine plains of the Tagus, located approximately 10–15 km southeast of the site. Horses, on the other hand, could have been encountered in nearby patches of open landscapes, as Portuguese Estremadura during the Late Pleistocene was characterized by steppe-like vegetation and open mountain habitats dominated by pine woodlands (González-Sampériz et al., 2010). Isotopic zooarchaeology indicates that this region fluctuated between mosaics of semi-closed forests, open vegetation, and much drier steppe environments over time (Carvalho et al., 2022).
All of the prey items that Neanderthals procured at Picareiro were presumably available within a 15–20 km radius from the site, with patches of high-altitude environments from which ibex and chamois could be procured 30–40 km away. If Neanderthals did not travel to Montejunto or Serra da Estrela to procure prey given the existence of habitats suitable for the ungulates consumed by Neanderthals no more than 15–20 km away, it is possible that Neanderthals had a minimum subsistence territory of 225–400 km2. However, if longer distance trips were required for encountering ungulate populations, subsistence territories could be as large as 1,600 km2, assuming Neanderthals occasionally traveled to Montejunto or Serra da Estrela, to hunt ibex and chamois. However, considering ibex's ecological versatility, along with local mobility signal inferred by the presence of red deer and presence of aurochs and horse that may have been available within a 15 km radius, it is likely that Neanderthals primarily exploited a much more localized environment. The smaller estimated territory of 225–400 km2 is smaller than the 600 km2 territory proposed for Neanderthals at Gruta da Oliveira, based on 87Sr studies of human and ungulate remains (Linscott et al., 2023). Because 87Sr values provide direct evidence of the territory of an animal during one point of its life, it is not unreasonable to assume that our estimation, based on the proximity of patches in which targeted prey were encountered, is an underestimation.
4.3 Do we see continuity or change and Neanderthal subsistence and foraging mobility in Level JJ at Picareiro if change is observed, can this adaptation be linked to specific climate episodes?
Currently available paleoenvironmental indicators from Picareiro point to some sort of environmental disturbance linked with Heinrich Event 5 during the deposition of the JJD subunit (Benedetti et al., 2019; Carvalho et al., 2022). Though based on limited data, paleoenvironmental reconstructions based on isotope zooarchaeology point to rapidly oscillating environments from more open to more closed temperate forests in JJD. Thus, if we observe changes in Neanderthal subsistence before, during and after the deposition of JJD, it could be that ecological instability in Heinrich Event 5 forced Neanderthals to change their subsistence to adapt. However, if continuity is observed, this would indicate resilience in Neanderthal subsistence behaviors and/or population stability among targeted taxa. In other words, though regional environmental conditions may have oscillated, continuity in Neanderthal subsistence would suggest this part of Portuguese Estremadura consistently supported animal populations.
The absence of statistically significant differences in prey selection across the three JJ assemblages indicates that Neanderthal subsistence and mobility strategies remained largely consistent during the late Middle Paleolithic at Lapa do Picareiro. Across all subunits, the persistent exploitation of red deer underscores the resilience of these strategies—red deer served as a dependable resource despite climatic instability, likely due to the species' inherent ecological stability. This continuity is particularly striking given the isotopic evidence of environmental disturbance in red deer remains from JJD (Carvalho et al., 2022).
Although Heinrich events may have fragmented Neanderthal territories in other parts of southern Europe during periods of climatic downturn (Melchionna et al., 2018), the subsistence behavior at Picareiro appears unaffected. In contrast, the lithic record from the site reveals a shift in raw material procurement strategies: before Heinrich Event 5, there was a broader use of quartz, quartzite, and chert, whereas after the event, the assemblage relies primarily on quartz and quartzite, with chert largely omitted (Carvalho et al., 2022). Despite this change in lithic practices, the faunal evidence confirms a stable pattern of red deer exploitation—a trend that is consistent with other Iberian sites where red deer remain the preferred prey even amid cycles of environmental change (e.g., Jones et al., 2019; Marín-Arroyo and Sanz-Royo, 2021).
5 Conclusion
Our results suggest Neanderthals were the primary agents shaping the Level JJ faunal assemblages at Picareiro, with a consistent preference on red deer, supplemented occasionally by other ungulates such as aurochs, ibex, chamois, and horse. Skeletal remains were primarily appendicular and cranial elements, reflecting the transport of meat-rich and marrow-bearing parts to the cave for processing, were largely absent because they either were not transported to the site, or suffered attrition due to site formation processes. Cutmarks and percussion marks reveal butchering practices such as defleshing, fileting and marrow extraction, suggesting efficient resource use. The stable ecological conditions of surrounding riverine valleys and temperate forests likely supported red deer populations, enabling Neanderthals to exploit this key resource through variable hunting strategies while adapting their behaviors to the challenges of the landscape and environment.
The evidence from Lapa do Picareiro suggests that Neanderthal occupations were likely brief and characterized by intermittent use rather than long-term, intensive habitation. Although the small lithic assemblage and the absence of clear structural or living floor features point to low-density, short-term site use, the palimpsest nature of the deposits and the need for multiple lines of evidence caution against definitive conclusions. At the same time, the subsistence data—marked by the consistent exploitation of red deer alongside occasional hunting of ibex, chamois, aurochs, and horse—indicates that Neanderthals were effectively exploiting a diverse, ecotonal environment within a relatively localized territory, potentially ranging from 225 to 400 km2.
Despite environmental disturbances during Heinrich Event 5, the evidence from Picareiro demonstrates that Neanderthal subsistence strategies remained remarkably resilient. While shifts in lithic raw material procurement signal adaptive responses to changing conditions, the consistent exploitation of red deer across assemblages underscores a stable, reliable resource base. This continuity suggests that, even amid oscillating environments, the landscape of Portuguese Estremadura consistently supported key prey populations, enabling Neanderthals to maintain effective subsistence and mobility strategies throughout the late Middle Paleolithic. These findings highlight the dynamic interplay between ecological stability, environmental pressures, and Neanderthal subsistence and mobility strategies in the late Middle Paleolithic.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because this study analyzes archaeological animal remains that are tens of thousands of years old. Since no research is conducted on modern or live specimens, no ethical approval is required for research.
Author contributions
MC: Conceptualization, Data curation, Formal analysis, Investigation, Writing – original draft, Writing – review & editing, Funding acquisition, Methodology, Project administration, Visualization. JH: Conceptualization, Funding acquisition, Investigation, Methodology, Supervision, Writing – review & editing. EJ: Conceptualization, Investigation, Methodology, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work at Lapa do Picareiro was funded by U.S. National Science Foundation (NSF) awards to JH (BCS-1420299, BCS-1724997, BCS-2150625) and Michael Benedetti (BCS-1420453, BCS-1725015, BCS-2150662). The analysis was funded by a Latin American and Iberian Institute PhD Fellowship (University of New Mexico) to MC. MC is currently a beneficiary of a CEEC – 5th Edition research contract promoted by the Portuguese FCT (reference: 2022. 06405.CEECIND). Open-access publication is funded by the European Union (ERC-CoG-2021, FINISTERRA, 101045506). Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency. Neither the European Union nor the granting authority can be held responsible for them.
Acknowledgments
We wanted to thank ICArEHB (Universidade do Algarve) for hosting M.C. and the University of New Mexico's Latin American and Iberian Institute for financial support to M.C. during the zooarchaeological analysis in this study. In addition, we would like to thank Lawrence G. Straus for his mentorship during the development and execution of this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fearc.2025.1545249/full#supplementary-material
References
Acevedo, P., and Cassinello, J. (2009). Biology, ecology and status of Iberian ibex Capra pyrenaica: a critical review and research prospectus. Mamm. Rev. 39, 17–32. doi: 10.1111/j.1365-2907.2008.00138.x
Alimen, M.-H., and Lavocat, R. (1966). Atlas de préhistoire. v.3. Faunes et flores préhistoriques de l'Europe occidentale. Paris: N. Boubée.
Altuna, J. (1992). Asociaciones de macromamíferos del Pleistoceno superior en el Pirineo occidental y el Cantábrico. The Late Quaternary of the Pyrenean Region. Bilbao: Universidad del Pais Vasco.
Antunes, M. T., and Cunha, A. S. (1992). Neanderthalian remains fromFigueira Brava cave, Portugal. Geobios 25, 681–692. doi: 10.1016/0016-6995(92)80108-P
Azorit, C., Tellado, S., Oya, A., and Moro, J. (2012). Seasonal and specific diet variations in sympatric red and fallow deer of southern Spain: a preliminary approach to feeding behaviour. Anim. Prod. Sci. 52:720. doi: 10.1071/AN12016
Bailey, G. (2008). “Time perspectivism: origins and consequences,” in Time in Archaeology: Time Perspectivism Revisited, eds. S. Holdaway and L. Wandsnider (Salt Lake City, UT: University of Utah Press), 13–30.
Bailey, G., and Galanidou, N. (2009). Caves, palimpsests and dwelling spaces: examples from the Upper Palaeolithic of south-east Europe. World Archaeol. 41:215e241. doi: 10.1080/00438240902843733
Barakat, S., Le Corre, M., Willmes, M., Cohen, J., Vuillien, M., Desclaux, E., et al. (2023). Laser ablation strontium isotopes and spatial assignment show seasonal mobility in red deer (Cervus elaphus) at Lazaret Cave, France (MIS 6). Front. Ecol. Evol. 11:988837. doi: 10.3389/fevo.2023.988837
Barbieri, A., Regala, F. T., Cascalheira, J., and Bicho, N. (2023). The sediment at the end of the tunnel: geophysical research to locate the Pleistocene entrance of Gruta da Companheira (Algarve, Southern Portugal). Archaeol. Prospect. 30, 117–134. doi: 10.1002/arp.1881
Benedetti, M. M., Haws, J. A., Bicho, N. F., Friedl, L., and Ellwood, B. B. (2019). Late Pleistocene site formation and paleoclimate at Lapa do Picareiro, Portugal. Geoarchaeology 34, 698–726. doi: 10.1002/gea.21735
Berger, J. (1986). Wild Horses of the Great Basin: Social Competition and Population Size. Chicago, IL: University of Chicago Press.
Berlioz, E., Azorit, C., Blondel, C., Ruiz, M. S. T., and Merceron, G. (2017). Deer in an arid habitat: dental microwear textures track feeding adaptability. Hystrix 28, 222–230. doi: 10.4404/HYSTRIX-28.2-12048
Bicho, N. (2004). “The middle paleolithic occupation of Southern Portugal,” in Settlement Dynamics of the Middle Paleolithic and Middle Stone Age, vol. II, ed N. Conard (T?bingen: Kerns Verlag), 513–531.
Bicho, N., and Cascalheira, J. (2020). “Use of lithic assemblages for the definition of short-term occupations in hunter-gatherer prehistory,” in Short-Term Occupations in Paleolithic Archaeology: Definition and Interpretation, eds. J. Cascalheira, and A. Picin (Cham: Springer International Publishing), 19–38.
Bicho, N., and Haws, J. (2012). The Magdalenian in central and southern Portugal: human ecology at the end of the Pleistocene. Quat. Int. 272–273, 6–16. doi: 10.1016/j.quaint.2012.02.055
Bicho, N., Haws, J., and Hockett, B. (2006). Two sides of the same coin—rocks, bones and site function of Picareiro Cave, central Portugal. J. Anthropol. Archaeol. 25, 485–499. doi: 10.1016/j.jaa.2006.03.009
Bicho, N. F., Haws, J., Gibaja, J. F., and Hockett, B. (2009). Lapa do Picareiro, un asentamiento de caza magdaleniense en la Estremadura portuguesa. Complutum 20, 71–82.
Bicho, N. F., Haws, J., Hockett, B., and Markova, A. K. (2003). Paleoecologia e ocupa..o humana da Lapa do Picareiro: resultados preliminares. Revista portuguesa de Arqueologia 6, 49–81.
Bicho, N. F., Hockett, B., Haws, J., et al. (2000). Hunter–gatherer subsistence at the end of the Pleistocene: preliminary results from Picareiro Cave, Central Portugal. Antiquity 74, 500–506. doi: 10.1017/S0003598X00059834
Binford, L. R. (1981). Behavioral archaeology and the “Pompeii premise”. J. Anthropol. Res. 37, 195–208. doi: 10.1086/jar.37.3.3629723
Blasco, R. (2008). Human consumption of tortoises at level IV of Bolomor Cave (Valencia, Spain). J. Archaeol. Sci. 35, 2839–2848. doi: 10.1016/j.jas.2008.05.013
Blasco, R., Cochard, D., Colonese, A. C., Laroulandie, V., Meier, J., Morin, E., et al. (2022). “Small animal use by Neanderthals,” in Updating Neanderthals: Understanding Behavioural Complexity in the Late Middle Palaeolithic, eds. F. Romagnoli, F. Rivals, and S. Benazzi (London: Academic Press), 123–143.
Blasco, R., and Peris, J. F. (2012). A uniquely broad spectrum diet during the Middle Pleistocene at Bolomor Cave (Valencia, Spain). Quat. Int. 252, 16–31 doi: 10.1016/j.quaint.2011.03.019
Blasco, R., Rosell, J., Rufà, A., Marco, A. S., and Finlayson, C. (2016a). Pigeons and choughs, a usual resource for the Neanderthals in Gibraltar. Quat. Int. 421, 62–77. doi: 10.1016/j.quaint.2015.10.040
Blasco, R., Rosell, J., Sánchez-Marco, A., Gopher, A., and Barkai, R. (2019). Feathers and food: human-bird interactions at Middle Pleistocene Qesem Cave, Israel. J. Hum. Evol. 136:102653. doi: 10.1016/j.jhevol.2019.102653
Blasco, R., Rosell, J., Smith, K. T., Maul, L. C., Sañudo, P., Barkai, R., et al. (2016b). Tortoises as a dietary supplement: a view from the Middle Pleistocene site of Qesem Cave, Israel. Quat. Sci. Rev. 133, 165–182. doi: 10.1016/j.quascirev.2015.12.006
Blumenschine, R. J. (1988). An experimental model of the timing of hominid and carnivore influence on archaeological bone assemblages. J. Archaeol. Sci. 15, 483–502. doi: 10.1016/0305-4403(88)90078-7
Blumenschine, R. J. (1995). Percussion marks, tooth marks and experimental determinations of the timing of hominid and carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge, Tanzania. J. Hum. Evol. 29, 21–52.
Blumenschine, R. J., and Marean, C. W. (1993). “A carnivore's view of archaeological bone assemblages,” in From Bones to Behavior: Ethnoarchaeological and Experimental Contributions to the Interpretation of Faunal Remains. Occasional Paper 21, ed. J. Hudson (Carbondale: Center for Archaeological Investigations, Southern Illinois University), 273–300.
Boneta Jiménez, I., Cardoso, J. L., and Pérez-García, A. (2025). The turtles from the middle Paleolithic site of Gruta Nova da Columbeira (Bombarral, Portugal): update through an archaeozoological perspective. Anat. Rec. 308, 1677–1691. doi: 10.1002/ar.25229
Brugal, J. P., and Raposo, L. (1999). Foz do Enxarrique (Ródão, Portugal): preliminary results of the analysis of a bone assemblage from a Middle Palaeolithic open site. Monogr. Römisch Germanischen Zentralmuseums 42, 367–379.
Bunn, H. T., Kroll, E. M., Ambrose, S. H., Behrensmeyer, A. K., Binford, L. R., Blumenschine, R. J., et al. (1986). Systematic butchery by Plio/Pleistocene hominids at Olduvai Gorge, Tanzania [and comments and reply]. Curr. Anthropol. 27, 431–452. doi: 10.1086/203467
Cáceres, I. (2002). Tafonomía de yacimientos antrópicos en karst. Complejo Galería (Sierra de Atapuerca, Burgos), Vanguard Cave (Gibraltar) y Abric Romaní (Capellades, Barcelona) (Ph.D. thesis). Dpt. Història i Geografia. Rovira i Virgili, Tarragona, 1–304.
Cardoso, J. L., and Cascalheira, J. (2024). 40,000 years later: what we know about the presence of Neanderthals in Portuguese territory and their extinction. Estudos Arqueológicos de Oeiras 34, 9–52. doi: 10.5281/zenodo.12731989
Cardoso, J. L., Raposo, L., and Ferreira, V. (2002). A Gruta Nova da Columbeira Bombarral. Bombarral: Câmara Municipal do Bombarral.
Carvalho, M., Benedetti, M., Cascalheira, J., Friedl, L., and Haws, J. (2023). “Neanderthals and early modern humans in western Iberia: Taphonomy and diet at Lapa do Picareiro (central Portugal),” in Poster Presented at the European Society for the Study of Human Evolution Annual Meeting, Aarhus, Denmark, 21–22 September (Aarhus).
Carvalho, M., Ellis, G., Benedetti, M., Friedl, L., Cascalheira, J., Bicho, N., et al. (2024). “Human diet and ecology during the Middle to Upper Paleolithic Transition in Portuguese Estremadura: The case from Lapa do Picareiro,” in Paper presented at the European Association of Archaeologists Annual Meeting, Rome, Italy, 28–31 August (Rome).
Carvalho, M., Jones, E. L., Ellis, M. G., Cascalheira, J., Bicho, N., Meiggs, D., et al. (2022). Neanderthal palaeoecology in the late Middle Palaeolithic of western Iberia: a stable isotope analysis of ungulate teeth from Lapa do Picareiro (Portugal). J. Quat. Sci. 37, 300–319. doi: 10.1002/jqs.3363
Carvalho, M., Pereira, T., and Manso, C. (2018). Rabbit exploitation in the Middle Paleolithic at Gruta Nova da Columbeira, Portugal. J. Archaeol. Sci. 21, 821–832. doi: 10.1016/j.jasrep.2018.09.003
Cochard, D., Brugal, J. P., Morin, E., and Meignen, L. (2012). Evidence of small fast game exploitation in the Middle Paleolithic of Les Canalettes Aveyron, France. Quat. Int. 264, 32–51. doi: 10.1016/j.quaint.2012.02.014
Cortés-Sánchez, M., Morales-Muñiz, A., Simón-Vallejo, M. D., Lozano-Francisco, M. C., Vera-Peláez, J. L., Finlayson, C., et al. (2011). Earliest known use of marine resources by Neanderthals. PLoS ONE 6:e24026. doi: 10.1371/journal.pone.0024026
Costamagno, S., Liliane, M., Cédric, B., Bernard, V., and Bruno, M. (2006). Les Pradelles (Marillac-le-Franc, France): a mousterian reindeer hunting camp? J. Anthropol. Archaeol. 25, 466–484. doi: 10.1016/j.jaa.2006.03.008
Cruz-Uribe, K. (1991). Distinguishing hyena from hominid bone accumulations. J. Field Archaeo. 18, 467–486. doi: 10.2307/530408
Danilkin, A. (1995). Behavioural Ecology of Siberian and European Roe Deer. Netherlands: Springer. Available online at: https://books.google.it/books?id=ugPa5Siepr4C
Darlas, A., and Psathi, E. (2016). “The middle and upper Paleolithic on the western coast of the Mani peninsula (southern Greece),” in Paleoanthropology of the Balkans and Anatolia (Dordrecht: Springer), 95–117.
Daujeard, C., Vettese, D., Britton, K., Béarez, P., Boulbes, N., Crégut-Bonnoure, E., et al. (2017). Neanderthal selective hunting of reindeer? The case study of Abri du Maras (south-eastern France). Archaeol. Anthropol. Sci. 11, 985–1011. doi: 10.1007/s12520-017-0580-8
David, B. (1990). How Was This Bone Burnt? in Problem Solving in Taphonomy: Archaeological and Palaeontological Studies From Europe, Africa and Oceania, Vol. 2, eds. S. Solomon, I. Davidson and D. Watson (Brisbane, QLD: Tempus), 65–79.
Denniston, R. F., Houts, A. N., Asmerom, Y., Wanamaker, A. D Jr., Haws, J. A., Polyak, V. J., et al. (2018). A stalagmite test of North Atlantic SST and Iberian hydroclimate linkages over the last two glacial cycles. Clim. Past 14, 1893–1913. doi: 10.5194/cp-14-1893-2018
Discamps, E., and Faivre, J. P. (2017). Substantial biases affecting Combe-Grenal faunal record cast doubts on previous models of Neanderthal subsistence and environmental context. J. Archaeol. Sci. 81, 128–132. doi: 10.1016/j.jas.2017.03.009
Dominguez-Rodrigo, M. (1997). Meat-eating by early hominids at the FLK 22Zinjanthropussite, Olduvai Gorge (Tanzania): an experimental approach using cut-mark data. J. Hum. Evol. 33, 669–690. doi: 10.1006/jhev.1997.0161
Eixea, A., Chacón, M. G., Bargalló, A., Sanchis, A., Romagnoli, F., Vaquero, M., et al. (2020). Neanderthal spatial patterns and occupation dynamics: a focus on the central region in Mediterranean Iberia. J. World Prehist. 33, 267–324. doi: 10.1007/s10963-020-09143-3
Evins, M. A. (1982). The fauna from Shanidar Cave: Mousterian wild goat exploitation in northeastern Iraq. Paléorient 8, 37–58. doi: 10.3406/paleo.1982.4308
Fa, J. E., Stewart, J. R., Lloveras, L., and Vargas, J. M. (2013). Rabbits and hominin survival in Iberia. J. Hum. Evol. 64, 233–241. doi: 10.1016/j.jhevol.2013.01.002
Faith, J. T., and Du, A. (2018). The measurement of taxonomic evenness in zooarchaeology. Archaeol. Anthropol. Sci. 10, 1419–1428. doi: 10.1007/s12520-017-0467-8
Faith, J. T., and Thompson, J. C. (2018). “Low-survival skeletal elements track attrition, not carcass transport behavior in Quaternary large mammal assemblages,” in Zooarchaeology in Practice: Case Studies in Methodology and Interpretation in Archaeofaunal Analysis, eds. C. M. Giovas, and M. J. LeFebvre (Cham: Springer), 109–126.
Fernández-Laso, M. C., Brugal, J. P., and Raposo, L. (2015). Gruta Nova da Columbeira (Bombarral, Portugal): un modelo de ocupación en cueva durante el Paleolítico Medio. Resultados del estudio del registro de macromamíferos. Trabajos de Prehistoria 72, 304–326. doi: 10.3989/tp.2015.12156
Figueiral, I., and Terral, J. F. (2002). Late quaternary refugia of Mediterranean taxa in the Portuguese Estremadura: charcoal based palaeovegetation and climatic reconstruction. Quat. Sci. Rev. 21, 549–558. doi: 10.1016/S0277-3791(01)00022-1
Finlayson, C., Brown, K., Blasco, R., Rosell, J., Negro, J. J., et al. (2012). Birds of a feather: neanderthal exploitation of raptors and corvids. PLoSONE 7:e45927. doi: 10.1371/journal.pone.0045927
Fisher, J. W. (1995). Bone surface modifications in zooarchaeology. J. Archaeol. Method Theory 2, 7–68. doi: 10.1007/BF02228434
Fonseca, M. M. (1998). Plasticity of mating behaviour in red deer (Cervus elaphus) in a Mediterranean environment (Doctoral dissertation). University College London, London, United Kingdom.
Gaudzinski-Windheuser, S., and Niven, L. (2009). “Hominin subsistence patterns during the Middle and Late Paleolithic in Northwestern Europe,” in The Evolution of Hominin Diets: Integrating Approaches to the Study of Palaeolithic Subsistence, eds. J.-J. Hublin, and M. P. Richards (Dordrecht: Springer), 99–111.
Gebert, C., and Verheyden-Tixier, H. (2001). Variations of diet composition of red deer (Cervus elaphus L.) in Europe. Mamm. Rev. 31, 189–201. doi: 10.1111/j.1365-2907.2001.00090.x
Goffette, Q., Rots, V., Abrams, G., Pirson, S., Di Modica, K., Bray, F., et al. (2024). Neanderthal exploitation of birds in north-western Europe: Avian remains from Scladina Cave (Belgium). Front. Environ. Archaeol. 3:1441926. doi: 10.3389/fearc.2024.1441926
Gomez, A., and Lunt, D. H. (2007). “Refugia within refugia: patterns of phylogeographic concordance in the Iberian Peninsula,” in Phylogeography of Southern European Refugia, eds. S. Weiss, and N. Ferrand (Dordrecht: Springer), 155–188.
González-Sampériz, P., Leroy, S. A., Carrión, J. S., Fernández, S., García-Antón, M., Gil-García, M. J., et al. (2010). Steppes, savannahs, forests and phytodiversity reservoirs during the Pleistocene in the Iberian Peninsula. Rev. Palaeobot. Palynol. 162, 427–457. doi: 10.1016/j.revpalbo.2010.03.009
Grayson, D. K., and Delpech, F. (2002). Specialized early Upper Palaeolithic hunters in southwestern France? J. Archaeol. Sci. 29, 1439–1449. doi: 10.1006/jasc.2002.0806
Hall, S. J. (2008). A comparative analysis of the habitat of the extinct aurochs and other prehistoric mammals in Britain. Ecography 31, 187–190. doi: 10.1111/j.0906-7590.2008.5193.x
Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontol. Electro. 4:9.
Hardy, B. L., and Moncel, M. H. (2011). Neanderthal use of fish, mammals, birds, starchy plants and wood 125-250,000 years ago. PLoS ONE 6:e23768. doi: 10.1371/journal.pone.0023768
Hardy, B. L., Moncel, M. H., Daujeard, C., Fernandes, P., Béarez, P., Desclaux, E., et al. (2013). Impossible Neanderthals? Making string, throwing projectiles and catching small game during Marine Isotope Stage 4 (Abri du Maras, France). Quat. Sci. Rev. 82, 23–40 doi: 10.1016/j.quascirev.2013.09.028
Haws, J., Benedetti, M., Carvalho, M., and Ellis, G. (2021). Human adaptive responses to climate andenvironmental change during the Gravettian of Lapa do Picareiro (Portugal). Quat. Int. 587, 4–18. doi: 10.1016/j.quaint.2020.08.009
Haws, J. A. (2006). “Late Upper Paleolithic large mammal exploitation at Lapa do Picareiro,” in Animais na Pré-história e Arqueologia da Península Ibérica. Actas IV Congreso de Arqueología Peninsula Vol. 3, ed. N. F. Bicho (Faro: Universidade do Algarve), 179–195.
Haws, J. A., Benedetti, M. M., Cascalheira, J. M., Bicho, N. F., et al. (2019). “Human occupation during the Late Pleniglacial at Lapa do Picareiro (Portugal),” in Human Adaptations to the Last Glacial Maximun: The Solutrean and Its Neighbors, eds I. Schmidt, J. Cascalheira, N. Bicho, and G. C., Weniger (Cambridge: Cambridge Scholar Publishing), 188.
Haws, J. A., Benedetti, M. M., Funk, C. L., Bicho, N. F., Daniels, J. M., Hesp, P. A., et al. (2010). Coastal wetlands and the Neanderthal settlement of Portuguese Estremadura. Geoarchaeology 25, 709–744. doi: 10.1002/gea.20330
Haws, J. A., Benedetti, M. M., Funk, C. L., Bicho, N. F., Pereira, T., Marreiros, J., et al. (2020b). Late Pleistocene landscape and settlement dynamics of Portuguese Estremadura. J. Field Archaeol. 45, 222–248. doi: 10.1080/00934690.2020.1733780
Haws, J. A., Benedetti, M. M., Talamo, S., Bicho, N., Cascalheira, J., Ellis, M. G., et al. (2020a). The early Aurignacian dispersal of modern humans into westernmost Eurasia. Proc. Natl. Acad. Sci. U. S. A. 117, 25414–25422. doi: 10.1073/pnas.2016062117
Heinrich, H. (1988). Origin and consequences of cyclic ice rafting in the northeast Atlantic Ocean during the past 130,000 years. Quat. Res. 29, 142–152. doi: 10.1016/0033-5894(88)90057-9
Higham, T., Douka, K., Wood, R., et al. (2014). The timing and spatiotemporal patterning of Neanderthal disappearance. Nature 512, 306–309. doi: 10.1038/nature13621
Hillson, S. (1996). Mammal Bones and Teeth: An Introductory Guide to Methods of Identification. London: Institute of Archaeology, University College London.
Hockett, B. S. (1996). Corroded, thinned and polished bones created by golden eagles (Aquila chrysaetos): taphonomic implications for archaeological interpretations. J. Archaeol. Sci. 23, 587–591. doi: 10.1006/jasc.1996.0055
Hofmann, R. R. (1989). Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78, 443–457. doi: 10.1007/BF00378733
Jones, E., and Carvalho, M. (2023). Ecospaces of the middle to upper paleolithic transition: the archaeofaunal record of the iberian Peninsula. J. Hum. Evol. 177:103331. doi: 10.1016/j.jhevol.2023.103331
Jones, E. L. (2004). Dietary evenness, prey choice, and human–environment interactions. J. Archaeol. Sci. 31, 307–317. doi: 10.1016/j.jas.2003.08.011
Jones, J. R., Richards, M. P., Reade, H., Bernaldo de Quirós, F., and Marín-Arroyo, A. B. (2019). Multi-Isotope investigations of ungulate bones and teeth from El Castillo and Covalejos caves (Cantabria, Spain): implications for paleoenvironment reconstructions across the Middle-Upper Palaeolithic transition. J. Archaeol. Sci. Rep. 23, 1029–1042. doi: 10.1016/j.jasrep.2018.04.014
Linscott, B., Pike, A. W., Angelucci, D. E., Cooper, M. J., Milton, J. S., Matias, H., et al. (2023). Reconstructing Middle and Upper Paleolithic human mobility in Portuguese Estremadura through laser ablation strontium isotope analysis. Proc. Nat. Acad. Sci. U. S. A. 120:e2204501120. doi: 10.1073/pnas.2204501120
Livraghi, A., Fanfarillo, G., Dal Colle, M., Romandini, M., and Peresani, M. (2021). Neanderthal ecology and the exploitation of cervids and bovids at the onset of MIS4: a study on De Nadale cave, Italy. Quat. Int. 586:24–41. doi: 10.1016/j.quaint.2019.11.024
Lupo, K. D., and O'Connell, J. F. (2002). Cut and tooth mark distributions on large animal bones: ethnoarchaeological data from the Hadza and their implications for current ideas about early human carnivory. J. Archaeol. Sci. 29, 85–109. doi: 10.1006/jasc.2001.0690
Lyman, R. L. (2005). Analyzing cut marks: lessons from artiodactyl remains in the northwestern United States. J. Archaeol. Sci. 32, 1722–1732. doi: 10.1016/j.jas.2005.06.003
Lyman, R. L. (2021). “Bone density and bone attrition,” in Manual of Forensic Taphonomy, eds. J. T. Pokines and S. A. Symes (Boca Raton, FL: CRC Press), 79–102.
Marin, J., Daujeard, C., Saladi,é, P., Rodríguez-Hidalgo, A., Vettese, D., Rivals, F., et al. (2020). Neanderthal faunal exploitation and settlement dynamics at the Abri du Maras, level 5 (south-eastern France). Quat. Sci. Rev. 243:106472. doi: 10.1016/j.quascirev.2020.106472
Marín, J., Rodríguez-Hidalgo, A., Vallverdú, J., de Soler, B. G., Rivals, F., Rabuñal, J. R., et al. (2019). Neanderthal logistic mobility during MIS3: zooarchaeological perspective of abric romaní level P (Spain). Quat. Sci. Rev. 225:106033. doi: 10.1016/j.quascirev.2019.106033
Marín, J., Saladié, P., Rodríguez-Hidalgo, A., and Carbonell, E. (2017). Neanderthal hunting strategies inferred from mortality profiles within the Abric Romaní sequence. PLoS ONE 12:e0186970. doi: 10.1371/journal.pone.0186970
Marín-Arroyo, A. B., and Sanz-Royo, A. (2021). What Neanderthals and AMH ate: reassessment of the subsistence across the Middle–Upper Palaeolithic transition in the Vasco-Cantabrian region of SW Europe. J. Quat. Sci. 37, 320–334. doi: 10.1002/jqs.3291
Martínez-Moreno, J., Mora, R., and De la Torre, I. (2004). “Methodological approach for understanding Middle Palaeolithic settlement dynamics at La Roca dels Bous (Noguera, Catalunya, Northeast Spain),” in Settlement Dynamics of the Middle Paleolithic and Middle Stone Age, Vol. 2, ed. N. J. Conard (Tübingen: Kerns Verlag), 393–413.
Mauget, R. (1981). “Behavioural and reproductive strategies in wild forms of Sus scrofa (European wild boar and feral pigs),” in The Welfare of Pigs, ed. W. Sybesma (The Hague: Martinus Nijhoff Publishers), 3–13.
McPherron, S. P. (2022). E5 [Computer software]. Available online at: https://github.com/surf3s/E5
Mein, P., and Antunes, M. T. (2000). “Gruta da Figueira Brava: petits mammifères – Insectívora, Chiroptera, Lagomorpha,” in Últimos Neandertais em Portugal – evidência, odontológica e outra, ed M. T. Antunes (Lisboa: Academia das Ciências de Lisboa), 163–177.
Melchionna, M., Di Febbraro, M., Carotenuto, F., Rook, L., Mondanaro, A., Castiglione, S., et al. (2018). Fragmentation of Neanderthals' pre-extinction distribution by climate change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 496, 146–154. doi: 10.1016/j.palaeo.2018.01.031
Moncel, M.-H., and Rivals, F. (2011). On the question of short-term neanderthal site occupations: payre, France (MIS 8-7), and Taubach/Weimar, Germany (MIS 5). J. Anthropol. Res. 67, 47–75. doi: 10.3998/jar.0521004.0067.104
Morin, E., and Laroulandie, V. (2012). Presumed symbolic use of diurnal raptors by Neanderthals. PLoS ONE 7:e32856. doi: 10.1371/journal.pone.0032856
Mysterud, A., Loe, L. E., Zimmermann, B., Bischof, R., Veiberg, V., and Meisingset, E. (2011). Partial migration in expanding red deer populations at northern latitudes–a role for density dependence? Oikos 120, 1817–1825. doi: 10.1111/j.1600-0706.2011.19439.x
Nabais, M. (2018). Neanderthal subsistence in Portugal: what evidence?. Archaeol. Int. 21, 95–100. doi: 10.5334/ai-376
Nabais, M., Dupont, C., and Zilhão, J. (2023). The exploitation of crabs by last interglacial Iberian Neanderthals: The evidence from Gruta da Figueira Brava (Portugal). Front. Environ. Archaeol. 2:1097815. doi: 10.3389/fearc.2023.1097815
Nabais, M., and Zilhão, J. (2019). The consumption of tortoise among last interglacial Iberian Neanderthals. Quat. Sci. Rev. 217, 225–246. doi: 10.1016/j.quascirev.2019.03.024
Naudinot, N., and Marchand, G. (2020). Take Shelter! Short-Term Occupations of the Late Paleolithic and the Mesolithic in the French Far West. Short-Term Occupations in Paleolithic Archaeology: Definition and Interpretation. Springer Nature, 121–146.
Nesti, I., Posillico, M., and Lovari, S. (2010). Ranging behaviour and habitat selection of Alpine chamois. Ethol. Ecol. Evol. 22, 215–231. doi: 10.1080/03949370.2010.502316
Orbach, M., and Yeshurun, R. (2021). The hunters or the hunters: human and hyena prey choice divergence in the Late Pleistocene Levant. J. Hum. Evol. 160:102572. doi: 10.1016/j.jhevol.2019.01.005
Pales, L., and Lambert, C. (1971). Atlas ostéologique pour servir à l'identification des mammifères du quaternaire. Paris: Éditions du Centre national de la recherche scientifique.
Patou-Mathis, M. (2000). Neanderthal subsistence behaviours in Europe. Int. J. Osteoarchaeol. 10, 379–395. doi: 10.1002/1099-1212(200009/10)10:5<379::AID-OA558>3.0.CO;2-4
Pelletier, M., Desclaux, E., Brugal, J. P., and Texier, P. J. (2019). The exploitation of rabbits for food and pelts by last interglacial Neandertals. Quat. Sci. Rev. 224:105972. doi: 10.1016/j.quascirev.2019.105972
Peresani, M., Fiore, I., Gala, M., Romandini, M., and Tagliacozzo, A. (2011). Late Neandertals and the intentional removal of feathers as evidenced from bird bone taphonomy at Fumane Cave 44 ky BP, Italy. Proc. Nat. Acad. Sci. U. S. A. 108, 3888–3893. doi: 10.1073/pnas.1016212108
Pérez Ripoll, M. (1992). Marcas de carniceria, fracturas intenciona-das y mordeduras de carnivoros en huesos prehistoricos del mediterráneo español. Alicante: Instituto de Cultura Juan Gil Albert (Coleccion Patrimonio 15).
Quézel, P. (1985). “Definition of the Mediterranean region and the origin of its flora,” in Plant Conservation in the Mediterranean Area, ed. C. Gómez-Campo (Dordrecht: Dr. W. Junk Publishers), 9–24.
Raposo, L. (1995). Ambientes, territorios y subsistencia en el Paleolítico Medio de Portugal. Complutum, 6, 57–78.
Raposo, L., and Cardoso, J. L. (2000). “Mousterian industries of the Gruta da Figueira Brava,” in Memórias da Academia das Ciências de Lisboa (Lisbon: Academia das Ciências de Lisboa), 319–337.
Real, C., Eixea, A., Sanchis, A., Morales, J. V., Klasen, N., Zilhão, J., et al. (2018). Abrigo de la Quebrada level IV (Valencia, Spain): interpreting a middle palaeolithic palimpsest from a zooarchaeological and lithic perspective. J. Paleolit. Archaeol. 3, 187–224. doi: 10.1007/s41982-018-0012-z
Real, C., Sanchis, A., Eixea, A., and Villaverde, V. (2019). Neanderthal subsistence and short-term human occupation patterns during MIS 5. New data from Abrigo de la Quebrada (Valencia, Spain). J. Archaeol. Sci. 28:102056. doi: 10.1016/j.jasrep.2019.102056
Rendu, W. (2022). “Selection versus opportunism: a view from Neanderthal subsistence strategies,” in Updating Neanderthals: Understanding Behavioural Complexity in the Late Middle Palaeolithic, eds. F. Romagnoli, F. Rivals, and S. Benazzi (Amsterdam: Academic Press; Elsevier), 109–122.
Rendu, W., Renou, S., Koliasnikova, A., Baumann, M., Plisson, H., Discamps, E., et al. (2023). Neanderthal subsistence at Chez-Pinaud Jonzac (Charente-Maritime, France): a kill site dominated by reindeer remains, but with a horse-laden diet? Front. Ecol. Evol. 10:1085699h. doi: 10.3389/fevo.2022.1085699
Richards, M. P., Pettitt, P. B., Trinkaus, E., Smith, F. H., Paunović, M., and Karavanić, I. (2000). Neanderthal diet at Vindija and Neanderthal predation: the evidence from stable isotopes. Proc. Nat. Acad. Sci. U. S. A. 97, 7663–7666. doi: 10.1073/pnas.120178997
Riel-Salvatore, J., and Barton, C. M. (2004). Late Pleistocene technology, economic behavior, and land-use dynamics in southern Italy. Am. Antiq. 69, 257–274. doi: 10.2307/4128419
Rodrigues, M. L. (1991). Depósitos e evolução quaternária das vertentes nas depressões de Minde e de Alvados (Maciço Calcário Estremenho, Portugal). Finisterra XXVI, 5–26. doi: 10.18055/Finis1907
Rosell, J., Blasco, R., Rivals, F., Chacon, M. G., Arilla, M., Camaros, E., et al. (2017). A resilient landscape at teixoneres cave (MIS 3; Moi a, Barcelona, Spain): the Neanderthals as disrupting agent. Quat. Int. 435:195e210. doi: 10.1016/j.quaint.2015.11.077
Ruiz, M. N., Benito-Calvo, A., Alonso-Alcalde, R., Alonso, P., de la Fuente, H., Santamaría, M., et al. (2021). Late Neanderthal subsistence strategies and cultural traditions in the northern Iberia Peninsula: Insights from Prado Vargas, Burgos, Spain. Quat. Sci. Rev. 254:106795. doi: 10.1016/j.quascirev.2021.106795
Salazar-García, D. C., Power, R. C., Serra, A. S., Villaverde, V., Walker, M. J., and Henry, A. G. (2013). Neanderthal diets in central and southeastern Mediterranean Iberia. Quat. Int. 318, 3–18. doi: 10.1016/j.quaint.2013.06.007
Sánchez-Hernández, C., Gourichon, L., Blasco, R., Carbonell, E., Chacón, G., Galván, B., et al. (2020b). High-resolution Neanderthal settlements in mediterranean Iberian Peninsula: A matter of altitude? Quat. Sci. Rev. 247:106523. doi: 10.1016/j.quascirev.2020.106523
Sánchez-Hernández, C., Gourichon, L., Soler, J., Soler, N., Blasco, R., Rosell, J., et al. (2020a). Dietary traits of ungulates in northeastern Iberian Peninsula: did these Neanderthal preys show adaptive behaviour to local habitats during the Middle Palaeolithic? Quat. Int. 557, 47–62. doi: 10.1016/j.quaint.2020.01.008
Schmid, E. (1972). Atlas of animal bones. For prehistorians, archaeologists and Quaternary geologists. Knochenatlas. Für Prähistoriker, Archäologen und Quartärgeologen. Amsterdam: Elsevier.
Sommer, R. S., Zachos, F. E., Street, M., Jöris, O., Skog, A., and Benecke, N. (2008). Late quaternary distribution dynamics and phylogeography of the red deer (Cervus elaphus) in Europe. Quat. Sci. Rev. 27, 714–733. doi: 10.1016/j.quascirev.2007.11.016
Starkovich, B. M. (2017). Paleolithic subsistence strategies and changes in site use at Klissoura Cave 1 (Peloponnese, Greece). J. Hum. Evol. 111, 63–84. doi: 10.1016/j.jhevol.2017.04.005
Steele, T. E. (2002). Red deer: Their ecology and how they were hunted by Late Pleistocene hominids in Western Europe (Doctoral dissertation). Stanford University, Stanford, CA, United States.
Stepanchuk, V. N., Vasilyev, S. V., Khaldeeva, N. I., Kharlamova, N. V., and Borutskaya, S. B. (2017). The last Neanderthals of Eastern Europe: Micoquian layers IIIa and III of the site of Zaskalnaya VI (Kolosovskaya), anthropological records and context. Quat. Int. 428, 132–150. doi: 10.1016/j.quaint.2015.11.042
Stiner, M. C. (1994). Honor Among Thieves: A Zooarchaeological Study of Neandertal Ecology. Princeton, NJ: Princeton Univ. Press.
Stiner, M. C. (2001). Thirty years on the “Broad Spectrum Revolution” and Paleolithic demography. Proc. Natl. Acad. Sci. U. S. A. 98, 6993–6996. doi: 10.1073/pnas.121176198
Stiner, M. C. (2002). On in situ attrition and vertebrate body part profiles. J. Archaeol. Sci. 29, 979–991. doi: 10.1006/jasc.2001.0798
Stiner, M. C. (2013). An unshakable middle paleolithic? Trends versus conservatism in the predatory niche and their social ramifications. Curr. Anthropol. 54, S288–S304. doi: 10.1086/673285
Straus, L. G. (1992). Iberia Before the Iberians: The Stone Age Prehistory of Cantabrian Spain. Albuquerque: University of New Mexico Press.
Straus, L. G. (2013). “Iberian archaeofaunas and hominin subsistence during marine isotope stages 4 and 3,” in Zooarchaeology and Modern Human Origins, eds. J. L. Clark, J. D. Speth (Dordrecht: Springer), 97–128.
Terlato, G., Livraghi, A., Romandini, M., and Peresani, M. (2019). Large bovids on the Neanderthal menu: exploitation of Bison priscus and Bos primigenius in northeastern Italy. J Archaeol. Sci. Rep. 25, 129–143. doi: 10.1016/j.jasrep.2019.04.006
Thatcher, D. L., Wanamaker, A. D., Denniston, R. F., Ummenhofer, C. C., Regala, F. T., Jorge, N., et al. (2020). Linking the karst record to atmospheric, precipitation, and vegetation dynamics in Portugal. Chem. Geol. 558:119949. doi: 10.1016/j.chemgeo.2020.119949
Vallverdú, J., Allue, E., Bischoff, J. L., C aceres, I., Carbonell, E., Cebri a, A., et al. (2005). Short human occupations in the middle palaeolithic level iof the abric Romaní rock-shelter (Capellades, Barcelona, Spain). J. Hum. Evol. 48:157e174. doi: 10.1016/j.jhevol.2004.10.004
Van Neer, W., and Wouters, W. (2009). “Les restes de Poissons des niveaux du Paléolithique moyen et supérieur de la grotte Walou,” in La grotte Walou à Trooz (Belgique), eds. S. Pirson, C. Draily, and M. Toussaint (Namur: Etudes et Documents Archéologie 21; Département du Patrimoine & Institut du Patrimoine Wallon), 68–81.
VanPool, T. L., and Leonard, R. D. (2011). Quantitative Analysis in Archaeology. Hoboken, NJ: John Wiley & Sons.
Vettese, D., Blasco, R., Cáceres, I., Gaudzinski-Windheuser, S., Moncel, M.-H., Hohenstein, U. T., et al. (2020). Towards an understanding of hominin marrow extraction strategies: a proposal for a percussion mark terminology. Archaeol. Anthropol. Sci. 12:48. doi: 10.1007/s12520-019-00972-8
Vidal-Cordasco, M., Ocio, D., Hickler, T., and Marín-Arroyo, A. B. (2022). Ecosystem productivity affected the spatiotemporal disappearance of Neanderthals in Iberia. Nat. Ecol. Evol. 6, 1644–1657. doi: 10.1038/s41559-022-01861-5
Villa, P., and Mahieu, E. (1991). Breakage patterns of human long bones. J. Hum. Evol. 21, 27–48. doi: 10.1016/0047-2484(91)90034-S
Villa, P., Soriano, S., Pollarolo, L., Smriglio, C., Gaeta, M., D'orazio, M., et al. (2020). Neandertals on the beach: Use of marine resources at Grotta dei Moscerini (Latium, Italy). PLoS ONE 15:e0226690. doi: 10.1371/journal.pone.0226690
Villaverde, V., Martínez, R., Guillem, P. M., and Fumanal, M. P. (1996). “Mobility and the role of small game in the paleolithic of the central region of the Spanish Mediterranean: a comparison of Cova Negra with other paleolithic deposits,” in The Last Neanderthals, the First Anatomically Modern Humans, eds. E. Carbonell and M. Vaquero (Tarragona: Universitat Rovira i Virgili), 267–288.
Wood, R. E., Barroso-Ruíz, C., Caparrós, M., Jordá Pardo, J. F., Galván Santos, B., and Higham, T. F. (2013). Radiocarbon dating casts doubt on the late chronology of the Middle to Upper Palaeolithic transition in southern Iberia. Proc. Nat. Acad. Sci. U. S. A. 110, 2781–2786. doi: 10.1073/pnas.1207656110
Yaroshevich, A., Oron, M., and Sharon, G. (2023). Big-game hunting during the late Middle Paleolithic in the Levant: insights into technology and behavior from Nahal Mahanayeem Outlet, Upper Jordan River, Israel. J. Archaeol. Sci. 47:103777. doi: 10.1016/j.jasrep.2022.103777
Yravedra, J. (2013). “New contributions in the subsistence of Middle-Upper Palaeolithic in northern Spain,” in Zooarchaeology and Modern Human Origins: Human Hunting Behavior during the Later Pleistocene, eds. J. Clark and J. D. Speth (New York, NY: Springer), 77–95.
Yravedra, J., and Cobo-Sánchez, L. (2015). Neanderthal exploitation of ibex and chamois in southwestern Europe. J. Hum. Evol. 78, 12–32. doi: 10.1016/j.jhevol.2014.10.002
Yravedra, J., Gómez-Castanedo, A., Aramendi-Picado, J., Montes-Barquín, R., and Sanguino-González, J. (2016). Neanderthal and Homo sapiens subsistence strategies in the Cantabrian region of northern Spain. Archaeol. Anthropol. Sci. 8:779–803. doi: 10.1007/s12520-015-0253-4
Yravedra, J., Gómez-Castanedo, A., Picado, J. A., and Preysler, J. B. (2014). Specialised hunting of Iberian ibex during Neanderthal occupation at El Esquilleu Cave, northern Spain. Antiquity 88, 1035–1049. doi: 10.1017/S0003598X00115303
Zilhão, J. (2001). “Middle Paleolithic settlement patterns in Portugal,' in Settlement Dynamics of the Middle Paleolithic and Middle Stone Age, ed N. Conard (T?bingen: Kerns Verlag), 597–608.
Zilhão, J., Angelucci, D., Argant, J., Brugal, J. P., Carrión, J. S., Carvalho, R., et al. (2010). “Humans and hyenas in the middle Paleolithic of Gruta da Oliveira (Almonda karstic system, Torres Novas, Portugal),” in Actas de la 1ª Reunión de científicos sobre cubiles de hiena (y otros grandes carnívoros) en los yacimientos arqueológicos de la Península Ibérica (Murcia: Museo Arqueológico de Murcia), 298–308.
Zilhão, J., Angelucci, D. E., Arnold, L. J., Demuro, M., Hoffmann, D. L., and Pike, A. W. (2021b). A Revised, Last Interglacial Chronology for the Middle Palaeolithic Sequence of Gruta da Oliveira (Almonda karst system). Lisbon: UNIARQ - Centro de Arqueologia da Universidade de Lisboa.
Zilhão, J., Angelucci, D. E., Arnold, L. J., d'Errico, F., Dayet, L., Demuro, M., et al. (2021a). Revisiting the Middle and Upper Palaeolithic archaeology of Gruta do Caldeirão (Tomar, Portugal). PLoS ONE 16:e0259089. doi: 10.1371/journal.pone.0259089
Zilhão, J., Angelucci, D. E., Igreja, M. A., Arnold, L. J., Badal, E., Callapez, P., et al. (2020). Last Interglacial Iberian Neandertals as fisher-hunter-gatherers. Science 367:eaaz7943. doi: 10.1126/science.aaz7943
Keywords: Neanderthals, taphonomy, zooarchaeology, Iberia, Paleolithic
Citation: Carvalho M, Haws JA and Jones EL (2025) Late Neanderthal subsistence and foraging mobility at Lapa do Picareiro: a zooarchaeological and taphonomic analysis of Level JJ. Front. Environ. Archaeol. 4:1545249. doi: 10.3389/fearc.2025.1545249
Received: 14 December 2024; Accepted: 27 May 2025;
Published: 01 July 2025.
Edited by:
Mariana Nabais, Institut Català de Paleoecologia Humana i Evolució Social (IPHES), SpainReviewed by:
Alessandra Livraghi, UMR5608 Travaux de Recherches Archéologiques sur les Cultures, les Espaces et les Sociétés (TRACES), FranceKeiko Kitagawa, University of Tübingen, Germany
Copyright © 2025 Carvalho, Haws and Jones. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Milena Carvalho, bW1jYXJ2YWxob0B1YWxnLnB0