 Mayra J. Esparza-Araiza1
Mayra J. Esparza-Araiza1 Bernardo Bañuelos-Hernández1
Bernardo Bañuelos-Hernández1 Gerardo R. Argüello-Astorga1
Gerardo R. Argüello-Astorga1 José P. Lara-Ávila2Paul H. Goodwin3
José P. Lara-Ávila2Paul H. Goodwin3 María I. Isordia-Jasso1Rosalba Castillo-Collazo1
María I. Isordia-Jasso1Rosalba Castillo-Collazo1 Alejandra Rougon-Cardoso4
Alejandra Rougon-Cardoso4 Ángel G. Alpuche-Solís1*
Ángel G. Alpuche-Solís1*- 1División Biología Molecular, Instituto Potosino de Investigación Científica y Tecnológica A.C., San Luis Potosí, México
- 2Facultad de Agronomía y Veterinaria, Universidad Autónoma de San Luis, San Luis Potosí, México
- 3School of Environmental Sciences, University of Guelph, Guelph, ON, Canada
- 4Laboratory of Agrogenomic Sciences, Universidad Nacional Autónoma de México, ENES-León, León, México
Clavibacter michiganensis subsp. michiganensis (Cmm) causes bacterial wilt and canker of tomato. Currently, no Solanum lycopersicum resistant varieties are commercially available, but some degree of Cmm resistance has been identified in Solanum peruvianum. Previous research showed up-regulation of a SUMO E2 conjugating enzyme (SCEI) transcript in S. peruvianum compared to S. lycopersicum following infection with Cmm. In order to test the role of SCEI in resistance to Cmm, a fragment of SCEI from S. peruvianum was cloned into a novel virus-induced gene-silencing (VIGS) vector based on the geminivirus, Tomato Mottle Virus (ToMoV). Using biolistic inoculation, the ToMoV-based VIGS vector was shown to be effective in S. peruvianum by silencing the magnesium chelatase gene, resulting in leaf bleaching. VIGS with the ToMoV_SCEI construct resulted in ~61% silencing of SCEI in leaves of S. peruvianum as determined by quantitative RT-PCR. The SCEI-silenced plants showed unilateral wilting (15 dpi) and subsequent death (20 dpi) of the entire plant after Cmm inoculation, whereas the empty vector-treated plants only showed wilting in the Cmm-inoculated leaf. The SCEI-silenced plants showed higher Cmm colonization and an average of 4.5 times more damaged tissue compared to the empty vector control plants. SCEI appears to play an important role in the innate immunity of S. peruvianum against Cmm, perhaps through the regulation of transcription factors, leading to expression of proteins involved in salicylic acid-dependent defense responses.
Introduction
Clavibacter michiganensis subsp. michiganensis (Cmm) is a Gram-positive plant bacterial pathogen belonging to the order Actinomycetales in family Microbacteraceae (Gartemann et al., 2008). It is the causal agent of bacterial wilt and canker of tomato (Solanum lycopersicum), which occurs worldwide (Eichenlaub and Gartemann, 2011). Bacterial wilt and canker has been reported to cause losses of tomato production as high as 84% in the U.S.A. and Canada (Strider, 1969; Gleason et al., 1993). Cmm can infect through wounds entering the xylem vessels producing enzymes, such as, endocellulases, polygalacturonases, pectin methylesterases, xylanases, serine proteases, and endo-1,4-glycosidases, that render the xylem non-functional (Carlton et al., 1998; Jahr et al., 1999, 2000). The symptoms begin as a unilateral wilting of leaves, followed by a generalized wilting of all the leaves and the cankers development on the stem. Cankers on young plants are particularly damaging as they can result in plant death. Cmm also infects fruit, which results in necrotic spots called bird's eyes (Gartemann et al., 2003).
Control of bacterial canker is difficult. Antibiotics are effective but can lead to selection of resistant bacterial populations (Strider, 1969; Gartemann et al., 2003). Cultural control can be achieved using certified disease-free seeds and strict hygienic measures, such as the removal and destruction of infected plants and compost biofumigation with compost (Gartemann et al., 2003). However, resistance is a desirable trait. Although there are no Cmm-resistant tomato cultivars commercially available, resistance has been identified in several wild tomato species, such as S. pimpinelifolium, S. peruvianum, and S. habrochaites (van Heusden et al., 1999; Kabelka et al., 2002; Coaker and Francis, 2004).
Using cDNA-AFLP analysis, a number of genes were found that were up-regulated in Cmm resistant S. peruvianum in comparison to Cmm susceptible S. lycopersicum plants following inoculation with Cmm (Lara-Ávila et al., 2012). One of those genes was the SUMO E2 conjugating enzyme SCE1 (SCEI), which encodes an enzyme involved in protein modification through sumoylation, which is a post-translational modification that covalently conjugates the small ubiquitin-like modifier (SUMO) protein to lysines on target proteins. Proteins labeled with SUMO are then modified by the addition of small chemical groups, such as sugars and lipids or by the covalent attachment of other proteins. Sumoylation is a multistep process mediated by E1 (SUMO activating enzyme), then E2 SCEI (SUMO conjugating enzyme) and finally E3 (SUMO ligase) (Berndsen and Wolberger, 2014).
SCEI has been shown to increase during plant-pathogen interactions. Expression of SCEI increased during Pseudomonas syringae pv. tomato (Pst) infection of tomato plants, along with several hormones, such as salicylic acid (SA) and jasmonic acid (JA), which are key signaling molecules in innate immunity (Miura and Hasegawa, 2010; van den Burg et al., 2010; Park et al., 2011). Overexpression of SCEI in Arabidopsis did not have obvious effects on plant development but increased expression of abscisic acid (ABA)-responsive genes following ABA treatment. ABA is also linked to innate immunity through its positive effects on callose deposition (Lois et al., 2003). In addition, SCEI promotes SUMO conjugation, which affects innate immunity due to its involvement in SA-dependent resistance to bacterial pathogens (van den Burg et al., 2010; Park et al., 2011). Also some pathogen effectors, which suppress innate immunity, such as AvrBst of Xanthomonas campestris pv. vesicatoria, act as desumoylation enzymes, indicating a link between sumoylation and innate immunity (Xia, 2004).
One approach to evaluate the role of SCEI and sumoylation in plant disease resistance is to down-regulate its expression. Virus-induced gene silencing (VIGS) involves the production of dsRNA that directs DICER complexes for degradation of desired sequences resulting in effective plant gene silencing (Liu et al., 2002; Robertson, 2004; Galun, 2005; Cai et al., 2007). In this study, a novel VIGS vector was developed for wild tomato species by modifying a Tomato Mottle Virus (ToMoV), which is a Begomovirus that infects many wild tomato species, such as S. peruvianum (Polston et al., 1993) and S. habrochaites, and does not cause a drastic phenotypic effect on either of those species (Esparza-Araiza et al., unpublished). ToMoV contains two single-stranded circular DNAs (DNA A and B). DNA A has the genes Rep for virus replication, C4 for infectivity and suppression of posttranscriptional gene silencing, Trap for transactivation of BC1 and BV1, Ren for increased multiplication efficiency and CP for viral capsid. DNA B has the genes BC1 and BV1 for viral movement (Jeske, 2009) (Figure 1).
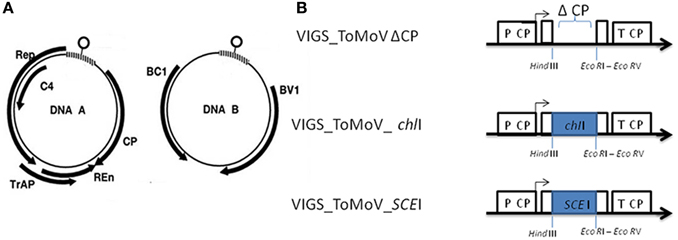
Figure 1. ToMoV silencing vector. (A) Diagram of the five genes of ToMoV component A: replication-associated protein (Rep), transcription activator protein (Trap), replication enhancer (Ren), coat protein (CP), and the two genes of ToMoV component B: movement proteins (BC1 and BV1) (Figure 1A modified from Gutierrez, 2002). (B) Organization of the modified ToMoV vector used in VIGS of ChII and SCEI genes. The capsid's promoter is shown as P CP. The capsid protein gene (CP) was deleted by reverse PCR to generate Δ CP, and portions of ChII and SCEI were cloned into this region with EcoRI and HindIII.
The goals of this study were to determine the effectiveness of a modified ToMoV vector for VIGS in S. peruvianum and to use the modified ToMoV as a vector for VIGS of SCEI. The effectiveness of a modified ToMoV VIGS vector was demonstrated by silencing a magnesium chelatase gene (ChlI) since its silencing causes leaf bleaching due to the suppression of the magnesium chelatase complex which is required for chlorophyll production (Cai et al., 2007). A portion of the SCEI gene from S. peruvianum was cloned into the modified ToMoV vector, introduced into S. peruvianum by particle bombardment, and then the SCEI-silenced plants were evaluated for their degree of resistance to Cmm.
Materials and Methods
Virus, Bacterial Strain, Plant Material, Inoculum, and Challenge Procedure
The sequence of ToMoV was obtained from a strain isolated from tomato leaves in Florida, USA in 2003 (GenBank accession nos. NC_00193 and NC_001939). Strain 1387 of Cmm was isolated from a commercial tomato field near Queretaro, Mexico in 2012 by J. P. Lara-Avila, Universidad Autonoma de San Luis Potosi, MX. Cmm was grown in 802 medium broth (polypeptone 1 g L−1, yeast extract 2 g L−1, MgSO4·7H2O 0.92 g L−1) at 28°C for 48 h shaking at 190 rpm. The bacterial suspension was diluted to 5 × 107 CFU/mL−1 (A620 = 0.2), and 0.5 mL was injected with an insulin syringe into the lower side of the first true leaves of 2 month-old of S. peruvianum. Seeds of S. peruvianum accession LA2172 were obtained from Tomato Genetics Resources Center, Davis, California, and seeds of S. lycopersicum cv. Ailsa Craig were obtained from the University of Nottingham, UK. Plants were grown in a commercial soil mixture substrate (Sunshine Mix #6, Sun Grow Horticulture, Vancouver, BC, CA), in individual pots in growth chambers at 25°C with a 16 h/8 h light/dark regime and then transferred to a greenhouse.
Silencing Vector Design and Cloning of ChII and ScEI Genes
DNA of ToMoV A and B components were extracted from tomato leaves by R. F. Rivera-Bustamante, CINVESTAV, IPN, Mexico. The complete DNA of the ToMoV A and B components were cloned into pBluescript [pBS II SK (+∕−)] (Stratagene, La Jolla, CA, USA) using ApaI digestion. These constructs were named pBS_ToMoV A and pBSToMoV B. The pBS_ToMoV A construct was modified by removing a 51-bp fragment between a XhoI (668 bp position) and BamHI sites (719 bp position), as it would have interfered with the use of the multiple cloning site of pBS II SK (+∕−). The cloned ToMoV A capsid gene was then modified by removing a 657 bp region using PCR with the forward primer 5′-CTGAATTCAAGCTTTGCACT CATGCGTCTAACCCTG-3′ and reverse primer 5′-TCGAATTCGATATCCC ATGGCAAATCACGCTTAGGC-3′ that flanked the capsid gene. The forward primer was designed with EcoRI and HindIII restriction sites and the reverse primer was designed with EcoRI and EcoRV restriction sites. The PCR conditions were 7 min at 95°C, followed by 35 cycles of 1 min at 94°C, 1 min at 55°C, 5.5 min at 72°C, and 10 min at 72°C. The PCR product was purified, digested with EcoRI enzyme and ligated subsequently to pBS II SK(+/–) (Stratagene, La Jolla, CA, USA). The ligation mix was used to transform E. coli Top 10 strain (Invitrogen, Carlsbad, CA, USA). This construct was named ToMoVΔCP.
For VIGS of ChII, a 249-bp portion of the chelatase gene was amplified using genomic DNA of S. peruvianum LA2172 with the forward primer 5′-CTGCAGGAATTCCTC CAGAGCCAAATCACCTC-3′ and reverse primer 5′-AAGCTTAG ATTCCAACGGATCCTTCC- 3′. The forward and reverse primers were designed with EcoRI and HindIII restriction sites, respectively. These primers were designed based on the S. lycopersicum's chelatase sequence (XM_004248092.1). The PCR conditions were 5 min at 94°C, 50 s at 94°C, 40 s at 60°C, 50 s at 72°C for 35 cycles, and then 5 min at 72°C. The PCR product was purified and ligated to ToMoVΔCP-VIGS following digestion with EcoRI and HindIII. This plasmid was named ToMoV_ChII.
For VIGS of SCEI, genomic DNA of S. peruvianum LA2172 was used as a template in PCR with forward primer 5′-CTCGAATTCTCCTCAATGAAG ACAGTGGTTGG-3′and reverse primer 5′-ATAAAGCTTCA CCCTCTTTCGGTACTCCA-3′, containing EcoRI and HindIII restriction sites, respectively. Theses primers were designed based on the sequence (emb|CAE45567.1) Lara-Ávila et al. (2012). The 169-bp PCR product and the ToMoVΔCP-VIGS vector were digested with EcoRI and HindIII and then ligated. This plasmid was named ToMoV_SCEI. The identity of all constructs was confirmed by restriction analysis with EcoRI and HindIII and sequencing.
Plant Inoculation with the ToMoV VIGS Vector
ToMoV virus A and B components were mixed in a 1:1 ratio (1 μg of component A: ToMoVΔCP, ToMoV_ChII or ToMoV_SCEI and 1 μg of component B: ToMoV B) after their digestion from the pBS II SK vector with ApaI. Twenty-two day-old plants were treated with the virus mixture using the Biolistic® PDS-1000He gun (Bio Rad, Hercules, CA, USA) at low pressure (Carrillo-Tripp et al., 2006). The plants were then maintained for 60 days post-treatment (dpt) in a greenhouse at 25–30°C before inoculation with Cmm. This time period was chosen because ToMoV_ChII inoculated plants showed bleaching of all leaves by 60 days, which indicated that silencing had occurred. By 15 days post inoculation (dpi) with Cmm, symptoms of leaf wilting and necrosis were observed and recorded by scanning excised damaged leaves on a flat-bed scanner. A tif file was created and the number of pixels of damaged tissue was quantified by Scion Image (Scion Corporation, Frederick, MD, USA) (Wijekoon et al., 2008). Statistical analysis was based on T-test with unpaired data with Graph Pad Prism® V.5 (GraphPad, San Diego, CA, USA), and a statistically significant result was considered to be P < 0.01.
RNA Isolation and Quantitative RT-PCR Analysis
In order to quantify SCEI mRNA from silenced and empty vector control plants, total RNA was isolated using Trizol (Invitrogen, Carlsbad, CA, USA) from 40 dpt S. peruvianum (62 days old plants) inoculated either with ToMoVΔCP or ToMoV_SCEI. After treatment with DNAse I (Invitrogen, Carlsbad, CA, USA), RNA was quantified by a NanoDrop ND-1000 UV-Vis spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA) according to the manufacturer's instructions. cDNA synthesis and quantitative real time PCR analysis were performed using the iScript™ One-Step RT-PCR kit with SYBR® Green (Bio-Rad Laboratories, Hercules, CA, USA). The 20 μL reactions contained 100 ng of total RNA, 12.5 μL of 2x SYBR Green RT-PCR reaction mix, 200 nM of each primer listed below and 1 μL of iScript MMlV reverse transcriptase. Quantification was based on a cycle threshold value with expression level of SCE1 gene normalized to actin gene (Accession no. FJ532351). The SCEI forward primer 5′-TTGCTAAGCCGGA GACACTT-3′ and reverse primer 5′-ACACTTTGGC GGTTTACTCG-3′ were designed outside the targeted region for silencing. For actin, the forward primer was 5′-CCTCACCGAGAGAGGTTACA TGT-3′ and reverse primer was 5′-CATGTCGCGGACAATTTCC3′. The RT-PCR conditions were 10 min at 50°C (cDNA synthesis), 5 min at 95°C (iScript MMLV reverse transcriptase inactivation), followed by 40 PCR cycles of 10 s at 95°C and 30 s at 60°C. Melting curves were performed by 80 cycles of 1 min at 95°C, 1 min at 55°C, and 10 s at 55°C increasing the temperature by 0.5°C per cycle of 10 s each. Absence of contaminating genomic DNA was confirmed by PCR of RNA samples without cDNA synthesis. PCRs were performed on an Applied Biosystems 7500 Fast Real-Time Real-Time-PCR system version 2.0, and the data was analyzed with the Applied Biosystems 7500 software V.2.0. Three biological replicates were analyzed with three technical replicates per biological replicate. Statistical analysis was based on T-tests with unpaired data with Graph Pad Prism® V.5 (GraphPad, San Diego, CA, USA), and a statistically significant result was considered to be P < 0.01.
Cmm DNA Detection in Inoculated Plant Tissue by PCR
Total DNA was isolated from S. peruvianum plants at 10 and 20 dpi with ToMoVΔCP or ToMoV_SCEI based on a modified protocol of Dellaporta et al. (1983), and then quantified with a NanoDrop ND-1000 UV-Vis Spectrophotometer (NanoDrop Technologies Inc., Wilmington, DE, USA). A 233-bp of the endo-1,4-beta-glucosidase gene, Cel-A (HQ636581; Lara-Ávila et al., unpublished) was amplified using 100 ng of the DNA as template and forward primer 5′-ATCAAGCAGATGG GGTTCAC-3′ and reverse primer 5′-TCCGGATACTGCGA TGTGTA-3′. The PCR conditions were 5 min at 94°C, and then 50 s at 94°C, 40 s at 60°C, 50 s at 72°C for 35 cycles followed by 5 min at 72°C.
Cmm DNA Quantification in Inoculated Plant Tissue by Quantitative PCR
Total DNA was isolated and quantified as above from S. peruvianum plants inoculated either with ToMoVΔCP or ToMoV_SCEI. For a 20 μL reaction, 100 ng of the DNA was added as template to 10 μL SYBR Green RT-PCR master mix (Applied Biosystems, Carlsbad, CA, USA) and 200 nM each of forward primer 5′-GAGCCAAGCCAC TGATCTTC-3′ and reverse primer 5′-CGTTCT CGTAGAGGCGGTAG-3′ to generate a 219 bp portion of the tomatinase, endo-1,4-beta-glycosidase constitutive gene, TomA, of Cmm (AF393183.1; Flügel et al., 2012), RT-PCR, melting curve, quantification and data analysis were performed as per SCEI described previously. A standard curve was created based on the concentrations of a cloned version of TomA in the pGEM-T Easy vector (Promega, Madison, WI, USA) using 2.86 × 102 to 2.86 × 108 copies/ng. The correlation coefficient between the cycle threshold value and the concentration of the cloned TomA was of 0.999. Statistical analysis was based on a correlation test with unpaired data with Graph Pad Prism® V.5 (GraphPad, San Diego, CA, USA), and a statistically significant result was considered to be P < 0.01.
Scanning Electron Microscopy
One-half centimeter stem samples were excised and fixed with glutaraldehyde 3% in Sörensen buffer (100 mM sodium phosphate pH 7.4). After washing three times in buffer, the samples were immersed in 1% osmium tetraoxide (OsO4) in Sörensen buffer for 2 h, washed three times with Sörensen buffer, and then dehydrated with ethanol 30, 50, 70, 90, 95% and absolute ethanol for 15 min each, and then incubated two additional times in absolute ethanol for 15 min. Critical point dried were done in a Tousimis Samdri-PVT-3D (Tousimis Research, Rockville, MA), mounted and gold coated sputter in Cressington model 108auto (Cressington Scientific Instruments, Watford, UK) and examined in a FEI model Quanta 200 SEM (FEI, Brno, Czech Republic).
Results
Silencing ChII Using the ToMoV VIGS Vector in S. peruvianum
In order to develop a VIGS vector based on ToMoV, the coat protein contained in component A of ToMoV was removed, and a cloning site was added at the same location (Figure 1). A portion of the magnesium chelatase gene from S. peruvianum was cloned into the ToMoV VIGS vector (ToMoV_ChII) and introduced by bombardment into 22 day old S. peruvianum. Typical leaf bleaching indicating silencing of ChlI first appeared at 10 dpi and spread from the bombarded leaf until the whole plant was showing patchy bleaching at 40 dpi (Figure 2). Control plants inoculated with an empty ToMoV vector (ToMoVΔCP) did not show any bleaching symptoms. Similar results were obtained when ChlI was silenced in S. habrochaites, S. lycopersicum cv. Micro Tom, S. lycopersicum cv. Ailsa Craig, and Nicotiana benthamiana (data not shown).

Figure 2. Phenotype of silencing of ChII gene in S. peruvianum. (A) Phenotype of leaf bleaching at 10 dpi. (B) Close up of a bleached leaf at 10 dpi. (C) Close up of a bleached leaf 40 dpi. (D) Bleaching phenotype at 40 dpi.
Silencing of SCEI Using the ToMoV VIGS Vector in S. peruvianum
To silence SCEI, a 269 bp portion of the gene from S. peruvianum was amplified and cloned into the ToMoV VIGS vector (Figure 1B). Expression of SCEI at 40 dpi for S. peruvianum inoculated with ToMoV_SCEI was 0.61 of the value of plants inoculated with the empty vector (Figure 3) (t-test, p < 0.001) indicating silencing. The plant morphology and flowers of S. peruvianum inoculated with ToMoV_SCEI was identical to that of empty vector control plants (Figures 4A,B). Thus, silencing of SCEI did not have any apparent effect on the healthy plant phenotype under the conditions used in these experiments.
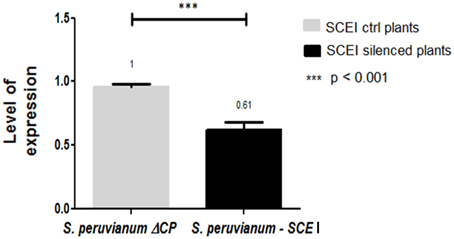
Figure 3. Expression level of the SCEI gene in S. peruvianum plants after silencing with ToMoV – VIGS vector. Real Time one step RT-PCR amplifications were performed using 100 ng of total RNA with the iScript™ One-Step RT-PCR kit using SYBER® Green. Quantification was based on the cycle threshold value with the expression level of SCEI normalized to that of S. peruvianum actin.

Figure 4. Phenotypes of S. peruvianum and S. lycopersicum plants inoculated with ToMoV VIGS vector prior and 20 days post-infection with Cmm. (A) Phenotype of SCEI-silenced S. peruvianum plant without Cmm inoculation. (B) non-silenced S. peruvianum plant without Cmm inoculation (negative control). (C) non-silenced S. lycopersicum plant inoculated with Cmm. (D) empty vector-treated S. peruvianum plants inoculated with Cmm. (E) SCEI-silenced S. peruvianum plant inoculated with Cmm. (F) Close up of the Cmm infection site on the S. peruvianum leaf for an empty vector-treated plant. (G) Close up of the S. peruvianum leaf located above the Cmm infection site for the same empty vector-treated plant, (H) Close up of the Cmm infection site on the S. peruvianum leaf for a SCEI-silenced plant. (I) Close up of the S. peruvianum leaf located above the Cmm infection site for the same SCEI-silenced plant showing unilateral wilting.
SCEI Silencing is Associated with Disease Susceptibility and Increased Cmm Growth in S. peruvianum Plants
Cmm-inoculated S. peruvianum silenced for SCEI first showed unilateral wilting of leaves at 15 dpi (Figures 4H,I), and the wilting spread to all leaves at 20 dpi resulting in plant death (Figure 4E). Plants inoculated with the empty vector showed necrosis only in the leaf that was inoculated with Cmm (Figure 4F), and the rest of the plant appeared healthy without any symptoms typical of Cmm infection for up to 20 dpi (Figures 4D,G). In contrast, the susceptible cultivar, S. lycopersicum cv. Ailsa Craig, showed unilateral wilting of leaves at 15 dpi, and the wilting spread to all leaves at 20 dpi (Figure 4C). The amount of damaged tissue in S. peruvianum plants due to Cmm infection varied between 12.57% to almost 25% in plants inoculated with ToMoVΔCP, whereas it was between 70.53 and 100% in plants inoculated with ToMoV_SCEI (t-test, p < 0.01) (Figure 5). Thus, the percentage of necrosis was approximately five times lower in the empty vector control than in the SCEI-silenced plants.
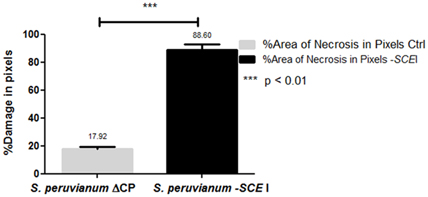
Figure 5. Damaged leaf tissues in empty vector-treated or SCEI-silenced S. peruvianum plants at 20 dpi with Cmm.
The presence of Cmm in inoculated plants was confirmed by PCR using specific primers for Cmm Cel-A using DNA obtained from ~3 cm above the inoculation site (Figure 6). SEM of non-infected S. peruvianum and S. lycopersicum plants showed that the parenchymal tissues appeared intact, healthy and without bacterial cells (Figures 7A,B). However, in S. peruvianum inoculated with ToMoVΔCP and challenged with Cmm, some bacteria were observed in the parenchymal tissue with little to no parenchymal tissue damage (Figures 7C,E). In contrast, S. peruvianum inoculated with ToMoV_SCEI and challenged with Cmm had more bacteria in the parenchymal tissue and more parenchymal tissue damage when compared with empty vector-inoculated control plants (Figure 7F). However, the number of bacteria and the level of parenchymal tissue damage were less than in the susceptible S. lycopersicum cv. Ailsa Craig, where the highest number of Cmm and the most parenchymal tissue damage was visible (Figure 7D). The quantity of bacteria observed by SEM correlated with the visible symptoms observed in empty vector inoculated S. peruvianum, SCEI silenced S. peruvianum, and susceptible S. lycopersicum, cv. Alisa Craig.
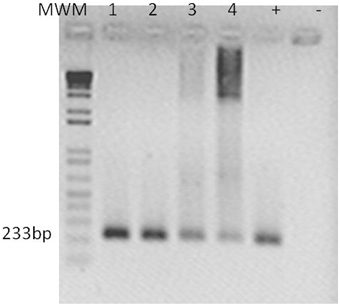
Figure 6. Cmm detection in S. peruvianum tissues by PCR. An amplicon of 233 bp was obtained using Cel-A primers por Cmm detection at 10 and 20 dpi. Lanes show PCR products shown with template of: (1) leaf tissue of SCEI-silenced S. peruvianum at 10 dpi, (2) leaf tissue of SCEI-silenced S. peruvianum at 20 dpi, (3) leaf tissue of empty vector treated S. peruvianum at 10 dpi, (4) leaf tissue of empty vector treated S. peruvianum at 20 dpi. Plasmid of pGEM T-Easy with Cel-A fragment. (-) Negative control. 100 ng of DNA were taken for each reaction.

Figure 7. Scanning electron microscopy of longitudinal cut of a stem of non-treated, empty vector and SCEI-silenced Solanum plants with and without Cmm challenge. (A) S. peruvianum without inoculation of Cmm showing no bacteria or tissue damage (B) S. lycopersicum without inoculation of Cmm showing no bacteria or tissue damage (C) non-VIGS treated S. peruvianum at 10 dpi with Cmm showing some bacterial but limited parenchymal tissue damage. (D) S. lycopersicum at 10 dpi with Cmm revealing more bacterial structures with greater parenchymal tissue damage. (E) empty-vector treated S. peruvianum at 10 dpi with Cmm with some bacterial structures and limited parenchymal tissue damage. (F) SCEI-silenced S. peruvianum at 10 dpi with Cmm revealing considerable bacterial structures and parenchymal tissue damage. Magnification 6000x, scale bar ~10 μm.
Real time PCR of TomA of Cmm was performed to quantify the Cmm population. At 10 dpi, the Cmm population reached 3.77 × 108 CFU per g of stem in SCEI-silenced S. peruvianum, whereas the population only reached 6.80 × 107 CFU per g of stem in the empty vector control S. peruvianum (t-test, p < 0.05) (Table 1). At 20 dpi, the population in the S. peruvianum SCEI-silenced plants increased to 1.23 × 109 CFU per g of stem, which was significantly higher than the 1.11 × 108 CFU per g of stem in the empty vector silenced plants (t-test, p < 0.05). Thus, Cmm populations were about 5–10 times higher due to SCEI silencing in S. peruvianum. At 10 dpi, the susceptible S. lycopersicum cv. Ailsa Craig had a significantly higher population of 4.63 × 108 CFU per g of stem, and at 20 dpi, the Cmm population reached 3.00 × 1010 CFU per g of stem, which as significantly higher than in S. peruvianum for both the SCEI-silence and empty vector control. These population differences correlated well with the amount of necrosis observed in the different plants.
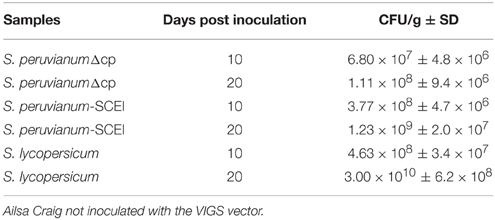
Table 1. Cmm populations estimated by qRT-PCR of the constitutive Cmm TomA gene in inoculated stems of S. peruvianum inoculated with ToMoVΔCP or ToMoV_SCEI or S. lycopersicum cv.
Discussion
VIGS is a tool that has been used successfully for the analysis of gene function without the time-consuming need to generate mutants or transgenic plants (Liu et al., 2002; Burch-Smith et al., 2004). For S. lycopersicum, there have been several viruses used for VIGS, such as Potato Virus X (Giliberto et al., 2005) and DNA1 component of tobacco curly shoot virus (TbCSV) (Huang et al., 2009), but the most commonly used VIGS vector has been based on Tobacco Rattle Virus (TRV) (Liu et al., 2002). However, there is one report of VIGS with the wild tomato, S. peruvianum, using TRV (Senthil-Kumar et al., 2007). In this work, a new VIGS vector based in ToMoV was developed, which showed silencing of genes in S. peruvianum and other Solanum species. The vector produced no or barely any viral symptoms in those Solanum species, and was an effective VIGS vector in S. peruvianum based on silencing a chelatase gene that blocks chlorophyll production resulted in large areas of yellowed and bleached tissues, similar to the phenotype reported in other plant species with chelatase silencing (Ekengren et al., 2003; Burch-Smith et al., 2004; Cai et al., 2007). An advantage of the ToMoV VIGS vector over TRV vectors is that silencing of genes with ToMoV was effective in S. peruvianum and other Solanum species at temperatures ranging from 22 to 30°C (data not shown). However, for TRV, slightly higher or lower temperatures from 21°C can greatly affect VIGS in tomato (Cai et al., 2007). Based on these results, the ToMoV VIGS vector was used to silence our gene of interest in S. peruvianum to determine if it may be involved in plant defenses against Cmm.
SCEI was selected for VIGS because Lara-Ávila et al. (2012) demonstrated that expression of this gene was highly up-regulated in the early stage of Cmm infection in S. peruvianum, which was followed by a decline to basal levels later in the infection, suggesting a role in the early defense response. In contrast, S. lycopersicum showed no significant change in SCEI expression levels following Cmm infection. All S. lycopersicum genotypes are relatively susceptible to Cmm, but S. peruvianum has a high level of resistance to this bacterial pathogen (van Ooijen et al., 1994). Inoculation of S. peruvianum, either on its own or with the empty ToMoV vector, only produced a rapid localized cell death at the inoculation site, whereas susceptible S. lycopersium cv. Ailsa Craig and SCEI-silenced S. peruvianum showed a necrotic phenotype of the leaves ranging from 70 to 100%. In addition, the population size of Cmm in S. peruvianum without treatment or treated with the empty ToMoV vector was much lower than in the SCEI-silenced S. peruvianum or S. lycopersicum cv. Ailsa Craig, whose populations reached levels similar to those of Cmm reported on susceptible S. lycopersicum cv. Moneymaker and S. lycopersicum cv. Jet Star by Sen et al. (2013) and Carlton et al. (1998), respectively.
Plant innate immunity is a multi-step process beginning with pathogen recognition. One type of innate immunity is based on recognition of Pathogen-Associated Molecular Patterns (PAMPs), which are invariant epitopes within molecules that are fundamental to the pathogens fitness, widely distributed among different microorganisms (Schwessinger and Zipfel, 2008). Another type of innate immunity is based on recognition of effectors, which generally are secreted by pathogens to manipulate or reprogram host defenses (Zipfel, 2008). The two types of recognition result in PAMP triggered immunity (PTI) and effector triggered immunity (ETI), which is usually stronger and longer than PTI and is often associated with the hypersensitive response (HR) at the infection site (Tsuda and Katagiri, 2010; Meng and Zhang, 2013). Both ETI and PTI initiate common signaling pathways differing in length and amplitude, such as an oxidative burst, activation of transcription factors, and MAP kinases and the production of plant hormones (Chisholm et al., 2006). Eventually, the plant responds through the expression of defense genes resulting in the production of various PR proteins and antimicrobial compounds and structures that limit pathogen spread and reproduction (Kaup et al., 2005; Saracco et al., 2007; Miura and Hasegawa, 2010; van den Burg et al., 2010; Balaji et al., 2011). The observation of rapid localized necrosis at the Cmm inoculation site in S. peruvianum indicates that the HR and ETI may be involved in Cmm resistance. In this study, SCEI-silenced plants did not have this localized cell death and clearly failed to restrict pathogen growth, similar to the susceptible S. lycopersicum, indicating that ETI could be compromised with diminished SCEI.
SUMOylation is a key process in plants as it provides post-translational modification of proteins involved in nuclear-cytosolic transport, transcriptional regulation, apoptosis, protein stability, response to stress and progression through the cell cycle and is controlled by SUMO pathway through regulation of transcription (Yang and Sharrocks, 2006; Enserink, 2015). The importance of SUMOylation in ETI can be inferred from studies of certain pathogen effectors. Hotson et al. (2003) found that the XopD effector of Xanthomonas campestris pathovar vesicatoria (Xcv) functions as cysteine protease with plant-specific SUMO substrate specificity. Roden et al. (2004) showed that the AvrXv4 effector of Xcv possesses SUMO isopeptidase activity, suggesting that SUMO conjugation system may be a key target for plant pathogen effectors. Therefore, some pathogen effectors act to hydrolyse SUMO-conjugated proteins to alter host cell signaling events, presumably for the pathogen's benefit (Hotson et al., 2003).
Several studies have suggested that SUMO plays an important role in pathogen defense responses (Lois et al., 2003; Saracco et al., 2007; van den Burg et al., 2010). One of the early defense genes with increased expression is SCEI (Pitzschke et al., 2009; van den Burg and Takken, 2010). Increased SCEI expression may be required in innate immunity because MAP kinase signaling and SUMOylation appear to converge to regulate the same targets that participate in signaling that controls defense gene expression (Yang and Sharrocks, 2006; Miller et al., 2010). Defense signaling involves a number of WRKY transcription factors, many of which have been identified as SUMOylation targets after phosphorylation by MAPKs. For example, WRKY3, WRKY4, WRKY6, WRKY33, WRKY72, and other WRKY transcription factors in Arabidopsis act as activators and/or repressor of defense gene expression and are also SUMO targets (Bethke et al., 2009; Popescu et al., 2009; Bhattarai et al., 2010; Miller et al., 2010; van den Burg and Takken, 2010). Therefore, silencing of the S. peruvianum SCEI gene by VIGS in this study may have made the SUMOylation mechanism on WRKYs or other MAP kinase targets non-functional, allowing for increased multiplication and development of disease symptoms by Cmm.
Increased levels of SCEI in plants, also occurs following abiotic stresses, such as salinity, drought, and cold. This was observed following salinity and drought stress in Spartina alterniflora (Karan and Subudhi, 2012). Lois et al. (2003) and Kurepa et al. (2003) also reported that high expression of SCE correlated with ABA mediated stress responses in different tissues of Arabidopsis, suggesting that sumoylation by SUMO1/2 played an early role in the plant stress response. ABA is well known for mediating plant stress responses to salinity, drought and cold (Karan and Subudhi, 2012). Thus, SCEI could also be involved in early stress responses following Cmm infection. One of the first responses to bacterial infections is a burst of ROS, and sumoylation is regulated by ROS (Zipfel and Robatzek, 2010; Park et al., 2011). On other hand, the overexpression of a SUMO gene in Arabidopsis resulted in activation of SA signaling following infections with Pseudomonas syringae pv tomato DC3000 and enhanced resistance to PstDC3000 (Panstruga et al., 2009; van den Burg et al., 2010; Xiong and Wang, 2013). While SA and ROS are both signals in plant defense response, there is an antagonistic interaction between ROS and SA signaling (Xu and Brosché, 2014). Therefore, silencing SCEI in S. peruvianum could have made sumoylation non-functional, which could be affecting SA levels in response to ROS and thus affecting defense responses to Cmm infection. Baysal et al. (2003) and Balaji et al. (2008) found that SA is induced by acidbenzolar-S methyl (ASM) increased resistance in S. lycopersicum seeds. The best protection against Cmm was obtained when the ASM had been applied 3 days before the Cmm infection. If the SA response was not sufficiently activated, then Cmm may have been able to avoid SA-dependent defenses allowing it to reproduce to higher levels, spreading in the plant and eventually killing it.
Silencing SCEI could also have affected the defense response in S. peruvianum to Cmm by altering other factors in the plant, such as plant morphology. Null mutations of SCEI in Arabidopsis resulted in embryo lethality (Park et al., 2011), but in Arabidopsis, mutants with moderately reduced SCEI levels showed a normal phenotype suggesting that partially reduced levels of SCEI can be tolerated under non-stressed conditions (Saracco et al., 2007). Using VIGS to silence SCEI in S. peruvianum did not cause visible aberrant developmental effects, and thus this explanation appears unlikely for the loss of Cmm resistance in S. peruvianum following SCEI silencing.
While S. lycopersicum is susceptible to Cmm, this is not due to the lack of SCEI genes. The partial sequence of SCEI in S. peruvianum had 100% nucleotide identity with a S. lycopersicum sequence (Solyc02g093110) and lesser nucleotide identity with other SCEI genes from S. lycopersicum (Figure S1). Since sumoylation is involved in many processes other than pathogen resistance, S. lycopersicum must have a number of SCEI genes in order to survive. Although the coding region of the SCEI in S. peruvianum in this study and Solyc02g093110 may be identical, the results of Lara-Ávila et al. (2012) show that they are regulated very differently following Cmm inoculation. It is unknown at which stage that innate immunity to Cmm differs between S. peruvianum and S. lycopersicum, but SCEI regulation is a possibility. If so, then transgenic S. lycopersicum with Solyc02g093110 regulated by the promoter region of the SCEI gene from S. peruvianum could result in greater induction following infection leading to greater resistance.
In this work, a novel VIGS-vector with ToMoV was developed, which did not produce severe viral symptoms and was able to silence genes in S. peruvianum. With this vector, it was possible to determine that SCEI is important in the defense of S. peruvianum against Cmm, possibly because SCEI impacted ETI through the effects of sumoylation on transcription factors, like WRKYs, and/or the production of the defense hormones, like SA. This study only examined silencing of SCEI in S. peruvianum in its relationship to innate immunity to Cmm, but SCEI may also play roles in resistance to other diseases or pests. For example, S. peruvianum has resistance against root-knot nematode (Yaghoobi et al., 2005) and tomato leaf curl virus (Anbinder et al., 2009). Therefore, altering expression of SCEI may be a strategy to increase resistance not only against Cmm but also against several other diseases. Based on our findings, one biotechnological approach to improve the Cmm resistance on commercial tomato cultivars is the overexpression of SCEI gene either using constitutive promoters or the promoter region of the SCEI gene from S. peruvianum which may activate the SA signaling pathway following infection of Cmm, resulting in an enhanced resistance to this bacterial disease.
Author Contributions
ME, BB, GA, JL, and AA design the study. ME, BB, and MI collected the data. ME, BB, GA, AA, and BB analyzed the data. ME, BB, GA, AA, PG, and AR interpreted the data. ME wrote the first draft and all co-authors contributed substantially to revisions.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge to Fordecyt-CONACYT 2012-02-193512 and Fundación Produce San Luis Potosí for funding. We are very grateful to Dr. Olga Araceli Patrón Soberano (Div. Biologia Molecular IPICYT, San Luis Potosí, MX) for her technical support with the scanning electron microscopy, PG and his laboratory members (University of Guelph, Ontario, CA) for their technical support, and also for CONACYT fellowship 161730 granted to ME for her PhD studies.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2015.01019
References
Anbinder, I., Reuveni, M., Azari, R., Paran, I., Nahon, S., Shlomo, H., et al. (2009). Molecular dissection of Tomato leaf curl virus resistance in tomato line TY172 derived from Solanum peruvianum. Theor. Appl. Genet. 119, 519–530. doi: 10.1007/s00122-009-1060-z
Balaji, V., Mayrose, M., Sherf, O., Jacob-Hirsch, J., Eichenlaub, R., Iraki, N., et al. (2008). Tomato transcriptional changes in response to Clavibacter michiganensis subsp. michiganensis reveal a role for ethylene in disease development. Plant Physiol. 146, 1797–1809. doi: 10.1104/pp.107.115188
Balaji, V., Sessa, G., and Smart, C. D. (2011). Silencing of host basal defense response-related gene expression increases susceptibility of Nicotiana benthamiana to Clavibacter michiganensis subsp. michiganensis. Phytopathology 101, 349–357. doi: 10.1094/PHYTO-05-10-0132
Baysal, O., Soylu, E., and Soylu, S. (2003). Induction of defense-related enzymes and resistance by the plant activator acibenzolar-S-methyl in tomato seedlings against bacterial canker caused by Clavibacter michiganensis ssp. michiganensis. Plant Physiol. 52, 747–753. doi: 10.1111/j.1365-3059.2003.00936.x
Berndsen, C. E., and Wolberger, C. (2014). New insights into ubiquitin E3 ligase mechanism. Nat. Struct. Mol. Biol. 21, 301–307. doi: 10.1038/nsmb.2780
Bethke, G., Unthan, T., Uhrig, J. F., Pöschl, Y., Gust, A. A., Scheel, D., et al. (2009). FLg22 regulates the release of ethylene response factor substrate from MAP kinase 6 in Arabidopsis thaliana via ethylene signaling. Proc. Natl. Acad. Sci. U.S.A. 16, 8067–8072. doi: 10.1073/pnas.0810206106
Bhattarai, K. K., Atamian, H. S., Kaloshian, I., and Eulgem, T. (2010). Worky72-type transcription factors contribute to basal immunity in tomato and Arabidopsis as well as gene-for-gene resistance mediated by the tomato R gene Mi-1. Plant J. 63, 229–240. doi: 10.1111/j.1365-313X.2010.04232.x
Burch-Smith, T. M., Anderson, J. C., Martin, G. B., and Dinesh-Kumar, S. P. (2004). Applications and advantages of virus-induced gene silencing for gene function studies in plants. Plant J. 39, 743–746. doi: 10.1111/j.1365-313X.2004.02158.x
Cai, X., Wang, C., Xu, Y., Xu, Q., Zheng, Z., and Zhou, X. (2007). Efficient gene silencing induction in tomato by a viral satellite DNA vector. Virus Res. 125, 169–175. doi: 10.1016/j.virusres.2006.12.016
Carlton, W. M., Braun, E. J., and Gleason, M. L. (1998). Ingress of Clavibacter michiganenesis subsp. michiganensis into tomato leaves through hydathodes. Phytopathology 88, 525–529. doi: 10.1094/PHYTO.1998.88.6.525
Carrillo-Tripp, J., Shimada-Beltrán, H., and Rivera-Bustamante, R. (2006). Use of geminiviral vectors for functional genomics. Curr. Opin. Plant Biol. 9, 209–215. doi: 10.1016/j.pbi.2006.01.012
Chisholm, S. T., Coaker, G., Day, B., and Staskawicz, B. J. (2006). Host-Microbe Interactions: shaping the evolution of the plant immune response. Cell 124, 803–814. doi: 10.1016/j.cell.2006.02.008
Coaker, G. L., and Francis, D. M. (2004). Mapping, genetic effects, and epistatic interaction of two bacterial canker resistance QTLs from Lycopsersicon hirsutum. Theor. Appl. Genet. 108, 1047–1055. doi: 10.1007/s00122-003-1531-6
Dellaporta, S. L., Wood, J., and Hicks, J. B. (1983). A plant DNA minipreparation: version, II. Plant. Mol. Biol. Rep. 1, 19–21. doi: 10.1007/BF02712670
Eichenlaub, R., and Gartemann, K. H. (2011). The Clavibacter michiganensis subspecies: molecular investigation of gram-positive bacterial plant pathogens. Annu. Rev. Phytopathol. 49, 445–464. doi: 10.1146/annurev-phyto-072910-095258
Ekengren, S. K., Liu, Y., Schiff, M., Dinesh-Kumar, S. P., and Martin, G. B. (2003). Two MAPK cascades, NPR1, and TGA transcription factors play a role in Pto-mediated disease resistance in tomato. Plant J. 36, 905–917. doi: 10.1046/j.1365-313X.2003.01944.x
Enserink, J. M. (2015). SUMO and the cellular stress response. Cell Div. 10, 4. doi: 10.1186/s13008-015-0010-1
Flügel, M., Becker, A., Gartemann, K. H., and Eichenlaub, R. (2012). Analysis of the interaction of Clavibacter michiganensis subsp. michiganensis with its host plant tomato by genome-wide expression profiling. J. Biotechnol. 160, 42–45. doi: 10.1016/j.jbiotec.2012.01.023
Galun, E. (2005). RNA silencing in plants. In Vitro Cell. Dev. Biol. 41, 113–123. doi: 10.1079/IVP2004619
Gartemann, K. H., Abt, B., Bekel, T., Burger, A., Engemann, J., Flügel, M., et al. (2008). The genome sequence of the tomato-pathogenic actinomycete Clavibacter michiganensis subsp. michiganensis NCPPB382 reveals a large island involved in pathogenicity. J. Bacteriol. 190, 2138–2149. doi: 10.1128/JB.01595-07
Gartemann, K. H., Kirchner, O., Engemann, J., Gräfen, I., Eichenlaub, R., and Burger, A. (2003). Clavibacter michiganensis subsp. michiganensis: first steps in the understanding of virulence of a gram-positive phytopathogenic bacterium. J. Biotechnol. 106, 179–191. doi: 10.1016/j.jbiotec.2003.07.011
Giliberto, L., Perrotta, G., Pallara, P., Weller, J. L., and Fraser, P. D., Bramley, P. M., et al. (2005). Manipulation of the blue light photoreceptor cryptochrome 2 in tomato affects vegetative development, flowering time, and fruit antioxidant content. Plant Physiol. 137, 199–208. doi: 10.1104/pp.104.051987
Gleason, M. L., Gitaitis, R. D., and Ricker, M. D. (1993). Recent progress in understanding and controlling bacterial canker of tomato in Eastern North America. Plant Dis. 77, 1069–1076. doi: 10.1094/PD-77-1069
Gutierrez, C. (2002). Strategies for geminivirus DNA replication and cell cycle interference. Physiol. Mol. Plant Pathol. 20, 219–230. doi: 10.1006/pmpp.2002.0401
Hotson, A., Chosed, R., Shu, H., Orth, K., and Mudgett, M. B. (2003). Xanthomonas type III effector XopD targets SUMO-conjugated proteins in planta. Mol. Microbiol. 50, 377–389. doi: 10.1046/j.1365-2958.2003.03730.x
Huang, C., Xie, Y., and Zhou, X. (2009). Efficient virus-induced gene silencing in plants using a modified geminivirus DNA1 component. Plant Biotechnol. J. 7, 254–265. doi: 10.1111/j.1467-7652.2008.00395.x
Jahr, H., Bahro, R., Burger, A., Ahlemeyer, J., and Eichenlaub, R. (1999). Interactions between Clavibacter michiganensis and its host plants. Environ. Microbiol. 1, 113–118. doi: 10.1046/j.1462-2920.1999.00011.x
Jahr, J., Dreier, J., Meletzuz, D., Bahro, R., and Eichenlaub, R. (2000). The endo β-1,4-glucanase Cel-A of Clavibacter michiganensis subsp. michiganensis is a pathogenicity determinant required for induction of bacterial wilt of tomato. Mol. Plant Microbe Interact. 13, 703–714. doi: 10.1094/MPMI.2000.13.7.703
Kabelka, E., Franchino, B., and Francis, D. M. (2002). Two loci from Lycopersicon hirsutum LA407 confer resistance to strains of Clavibacter michiganensis subsp. michiganensis. Phytopathology 92, 504–510. doi: 10.1094/PHYTO.2002.92.5.504
Karan, R., and Subudhi, P. J. (2012). A stress inducible SUMO conjugating enzyme gene (SaSce9) from a grass halophyte Spartina alterniflora enhances salinity and drought stress tolerance in Arabidopsis. BMC Plant Biol. 12:187. doi: 10.1186/1471-2229-12-187
Kaup, O., Gräfen, I., Zellermann, E. M., Eichenlaub, R., and Gartemann, K. H. (2005). Identification of tomatinase in the tomato-pathogenic actinomycete Clavibacter michiganensis subsp. michiganensis NCPPB382. Mol. Plant Microbe Interact. 18, 1090–1098. doi: 10.1094/MPMI-18-1090
Kurepa, J., Walker, J. M., Smalle, J., Gosink, M. M., Davis, S. J., Durham, T. J., et al. (2003). The small Ubiquity-like modifier (SUMO) protein modification system in Arabidopsis. J. Biol. Chem. 278, 6862–6872. doi: 10.1074/jbc.M209694200
Lara-Ávila, P., Isordia-Jasso, I., Castillo-Collazo, R., Simpson, J., and Alpuche-Solís, A. (2012). Gene expression analysis during interaction of tomato and related wild species with Clavibacter michiganensis subsp. michiganensis. Plant Mol. Biol. Rep. 30, 498–511. doi: 10.1007/s11105-011-0348-8
Liu, Y., Schiff, M., and Dinesh-Kumar, P. S. (2002). Virus-induced gene silencing in tomato. Plant J. 31, 777–786. doi: 10.1046/j.1365-313X.2002.01394.x
Lois, M. L., Lima, C. D., and Chua, N. H. (2003). Small ubiquitin-like modifier modulates abscisic signaling in Arabidopsis. Plant Cell 15, 1347–1359. doi: 10.1105/tpc.009902
Meng, X., and Zhang, S. (2013). MAPK cascades in plant disease resistance signaling. Annu. Rev. Phytopathol. 51, 245–266. doi: 10.1146/annurev-phyto-082712-102314
Miller, M. J., Barrett-Wilt, G. A., Hua, Z., and Vierstra, R. D. (2010). Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 107, 16512–16517. doi: 10.1073/pnas.1004181107
Miura, K., and Hasegawa, P. M. (2010). Sumoylation and other ubiquitin-like post-translational modifications in plants. Cell 20, 223–232. doi: 10.1016/j.tcb.2010.01.007
Panstruga, R., Parker, J. E., and Schulze-Lefert, P. (2009). SnapShot: plant immune response pathways. Cell 136, 978. doi: 10.1016/j.cell.2009.02.020
Park, H. J., Kim, W. Y., Park, H. C., Lee, S. Y., Bohnert, H. J., and Yun, D. J. (2011). SUMO and SUMOylation in Plants. Mol. Cell. 32, 305–316. doi: 10.1007/s10059-011-0122-7
Pitzschke, A., Schikora, A., and Hirt, H. (2009). MAPK cascade signaling networks in plant defense. Curr. Opin. Plant Biol. 12, 421–426. doi: 10.1016/j.pbi.2009.06.008
Polston, J. E., Hiebert, E., McGovern, R. J., Stansly, P. A., and Schuster, D. J. (1993). Host range of tomato mottle virus, a new geminivirus infecting tomato in Florida. Plant Dis. 77, 1181–1184.
Popescu, S. C., Popescu, G. V., Bachan, S., Zhang, Z., Gerstein, M., Snyder, M., et al. (2009). MAPK target networks in Arabidopsis thaliana revealed using functional protein microarrays. Genes Dev. 23, 80–92. doi: 10.1101/gad.1740009
Robertson, D. (2004). VIGS Vector for gene silencing: many targets, many tools. Annu. Rev. Plant Biol. 55, 495–519. doi: 10.1146/annurev.arplant.55.031903.141803
Roden, J., Eardley, L., Htson, A., Cao, Y., and Mudgett, M. B. (2004). Characterization of the Xanthomonas AvrXv4 effector, a SUMO protease translocated into plant cells. Mol. Plant Microbe Interact. 7, 633–643. doi: 10.1094/MPMI.2004.17.6.633
Saracco, S. A., Miller, M. J., Kurepa, J., and Vierstra, R. D. (2007). Genetic analysis of SUMOylation in Arabidopsis: conjugation of SUMO1 and SUMO2 to nuclear proteins is essential. Plant Physiol. 145, 119–134. doi: 10.1104/pp.107.102285
Schwessinger, B., and Zipfel, C. (2008). News from the frontline: recent insights into PAMP-triggered immunity in plants. Curr. Opin. Plant Biol. 11, 389–395. doi: 10.1016/j.pbi.2008.06.001
Sen, Y., Feng, Z., Vandenbroucke, H., van der Wolf, J., Viser, R. G. F., and van Heusden, A. W. (2013). Screening for new sources of resistance to Clavibacter michiganensis subsp. michiganensis (Cmm)in tomato. Euphytica 190, 309–317. doi: 10.1007/s10681-012-0802-1
Senthil-Kumar, M., Hema, R., Anand, A., Kang, L., Udayakumar, M., and Mysore, K. S. (2007). A systematic study to determine the extent of gene silencing in Nicotiana benthamina and other Solanaceae species when heterologous gene sequences are used for virus-induced gene silencing. New Phytol. 176, 782–791. doi: 10.1111/j.1469-8137.2007.02225.x
Strider, D. L. (1969). Bacterial Canker of Tomato Caused by Corynebacterium michiganense, A Literature Review and Bibliography. Raleigh, NC: North Carolina Agricultural Experiment Station. No. 193.
Tsuda, K., and Katagiri, F. (2010). Comparing signaling mechanisms engaged in pattern-triggered and effector-triggered immunity. Curr. Opin. Plant Biol. 13, 459–465. doi: 10.1016/j.pbi.2010.04.006
van den Burg, H. A., Kini, R. K., Schuurink, R. C., and Takken, F. L. (2010). Arabidopsis Small Ubiquitin-like modifer paralogs have distinct functions in development and defense. Plant Cell 22, 1998–2016. doi: 10.1105/tpc.109.070961
van den Burg, H. A., and Takken, F. L. W. (2010). SUMO-MAPK- and resistance protein-signaling converge at transcription complex that regulate plant innate immunity. Plant Signal. Behav. 5, 1597–1601. doi: 10.4161/psb.5.12.13913
van Heusden, A. W., Koornneef, M., Voorrips, R. E., Brüggemann, W., Pet, G., Vrielink-van Ginke, R., et al. (1999). Three QTLs from Lycopersicon peruvianum confer a high level of resistance to Clavibacter michiganensis subsp. michiganensis. Theor. Appl. Genet. 99, 1068–1074. doi: 10.1007/s001220051416
van Ooijen, J. W., Sandbrink, J. M., Vrielink, M., Verkerk, R., Zabel, P., and Lindhout, P. (1994). An RFLP linkage map of Lycopersicon peruvianum. Theor. Appl. Genet. 89, 1007–1013. doi: 10.1007/BF00224531
Wijekoon, C. P., Goodwin, P. H., and Hsiang, T. (2008). Quantifying fungal infection of plant leaves by digital image analysis using Scion Image software. J. Microbiol. Methods 74, 94–101. doi: 10.1016/j.mimet.2008.03.008
Xia, Y. (2004). Proteases in pathogenesis and plant defence. Cell. Microbiol. 6, 905–913. doi: 10.1111/j.1462-5822.2004.00438.x
Xiong, R., and Wang, A. (2013). SCE1, the SUMO-Conjugating enzyme in plants that interacts with Nlb, the RNA-dependent RNA polymerase of turnip mosaic virus, is required for viral infection. J. Virol. 87, 4704–4715. doi: 10.1128/JVI.02828-12
Xu, E., and Brosché, M. (2014). Salicylic acid signaling inhibits apoplastic reactive oxygen species signaling. BMC Plant. Biol. 14:155. doi: 10.1186/1471-2229-14-155
Yaghoobi, J., Yates, J. L., and Williamson, V. M. (2005). Fine mapping of the nematode resistance gene Mi-3 in Solanum peruvianum and construction of a S. lycopersicum DNA contig spanning the locus. Mol. Genet. Genomics 274, 60–69. doi: 10.1007/s00438-005-1149-2
Yang, S.-H., and Sharrocks, A. D. (2006). “Interplay of the SUMO and MAPK kinase pathways,” in The Histone Code and Beyond, eds S. L. Berger, O. Nakanishi, and B. Haendler (Grenoble: ESRF Press), 193–209.
Zipfel, C. (2008). Pattern recognition receptors in plant innate immunity. Curr. Opin. Immunol. 20, 10–16. doi: 10.1016/j.coi.2007.11.003
Keywords: SUMO-conjugating enzyme, virus-induced gene-silencing, Tomato Mottle Virus, bacterial canker, Clavibacter michiganensis subsp. michiganensis
Citation: Esparza-Araiza MJ, Bañuelos-Hernández B, Argüello-Astorga GR, Lara-Ávila JP, Goodwin PH, Isordia-Jasso MI, Castillo-Collazo R, Rougon-Cardoso A and Alpuche-Solís ÁG. (2015) Evaluation of a SUMO E2 Conjugating Enzyme Involved in Resistance to Clavibacter michiganensis Subsp. michiganensis in Solanum peruvianum, Through a Tomato Mottle Virus VIGS Assay. Front. Plant Sci. 6:1019. doi: 10.3389/fpls.2015.01019
Received: 09 August 2015; Accepted: 04 November 2015;
Published: 17 December 2015.
Edited by:
Libo Shan, Texas A&M University, USACopyright © 2015 Esparza-Araiza, Bañuelos-Hernández, Argüello-Astorga, Lara-Ávila, Goodwin, Isordia-Jasso, Castillo-Collazo, Rougon-Cardoso and Alpuche-Solís. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ángel G. Alpuche-Solís, YWxwdWNoZUBpcGljeXQuZWR1Lm14