 Sílvia Castro1*†
Sílvia Castro1*† Mariana Castro1†
Mariana Castro1† Victoria Ferrero2,3
Victoria Ferrero2,3 Joana Costa1
Joana Costa1 Daniela Tavares1
Daniela Tavares1 Luis Navarro2
Luis Navarro2 João Loureiro1
João Loureiro1- 1Centre for Functional Ecology and Department of Life Sciences, University of Coimbra, Coimbra, Portugal
- 2Department of Plant Biology, Faculty of Science, University of Vigo, Vigo, Spain
- 3Department of Ecology and Evolutionary Biology, University of Toronto, Toronto, ON, Canada
Biological invasions offer optimal scenarios to study evolutionary changes under contemporary timescales. After long-distance dispersal, exotic species have to cope with strong mate limitation, and shifts toward uniparental reproduction have been hypothesized to be selectively advantageous. Oxalis pes-caprae is a clonal tristylous species native to South Africa, and invasive in Mediterranean regions worldwide. It reproduces sexually and asexually but the importance of each strategy differs between ranges. Native populations reproduce mostly sexually while in invasive ones asexual reproduction is the prevailing strategy due to the dominance of pentaploid monomorphic populations. Nevertheless, two contrasting scenarios have been observed after introduction: transition toward clonality, and re-acquisition of sexuality fueled by multiple introductions of compatible mates. Here, we aimed to assess evolutionary changes of reproductive traits in O. pes-caprae invasive populations and evaluate whether these traits could be related with invasion success and prevalence of certain forms in the western Mediterranean basin. Sexual and asexual reproduction traits were quantified under optimal conditions in a common garden experiment including native and invasive sexual, predominately asexual, and obligated asexual individuals. Different reproductive, ecological, and genetic constraints created by long-distance dispersal seem to have generated different selective pressures in sexual and asexual traits, with our results supporting evolutionary changes in invasive populations of O. pes-caprae. Native plants had higher sexual fitness, while a transition toward clonality was clear for invasive forms, supporting clonal reproduction as a major trait driving invasion. Differences were also observed among invasive plants, with sexual forms having increased dispersal potential; thus, they are expected to be in advantage in comparison with predominantly asexual and obligated asexual plants, and may become widespread in the future. Historical processes, like the initial introduction of predominantly asexual forms followed by sexual forms more recently, could be in the origin of current distribution patterns of O. pes-caprae in the western Mediterranean. This study shows that invasion processes are very dynamic and that ecological and genetic constraints determined by the invasion process may originate different reproductive strategies that are likely to determine invasion success.
Introduction
Biological invasions are a serious threat to biodiversity and have long been recognized to comprise significant ecological and evolutionary consequences, not only for the communities being invaded, but also for the invasive species themselves (Brown and Eckert, 2005; Barrett et al., 2008; Pyšek et al., 2012; Oduor, 2013). For these reasons, since the seminal works of Elton (1958) and Baker and Stebbins (1965), biological invasions have attracted much attention of researchers in an attempt to identify traits that might confer an advantage during colonization of new habitats (Pyšek and Richardson, 2007; Hayes and Barry, 2008; van Kleunen et al., 2010) and to understand the biotic and abiotic factors that determine invasion success (e.g., Souza et al., 2011; Wisz et al., 2013). It also became clear that the introduction of sub-populations in new ecological scenarios generates valuable oportunities to study evolutionary transitions over contemporary time scales (e.g., Sakai et al., 2001; Brown and Eckert, 2005; Barrett et al., 2008; Prentis et al., 2008). These studies contribute to a better understanding of the factors triggering a successful invasion, and provide new insights on the evolutionary history of specific traits, such as those related to the reproductive system (Barrett et al., 2008; Barrett, 2011).
Reproduction is one of the key factors involved in the successful establishment and spread of a given organism after long-distance dispersal (e.g., Sakai et al., 2001; Barrett et al., 2008; Hayes and Barry, 2008; Castro-Díez et al., 2014; Moravcová et al., 2015). Reproductive modes determine the production, dispersal and genetic composition of propagules, thus influencing the genetic and demographic structure of populations, as well as the dispersal ability and evolutionary potential of introduced individuals or sub-populations, that, by its turn, will also determine the reproductive strategy (Sakai et al., 2001; Novak and Mack, 2005; Barrett et al., 2008; Ness et al., 2010). Flowering plants exhibit an outstanding diversity of reproductive strategies, from sexual to asexual modes and from self-compatible to obligated outcrossers (reviewed in Barrett, 2002), that frequently occur in combination and reveal liability under certain ecological and genetic stressful conditions (Dorken and Eckert, 2001; Eckert, 2002; Goodwillie et al., 2005; Herben et al., 2015). Thus, the relative contribution of each strategy to the fitness of a population/individual is expected to vary under the novel conditions and will play a major role in the establishment and spread of the introduced individual(s) (e.g., Brown and Eckert, 2005; Lui et al., 2005; Barrett et al., 2008; Silvertown, 2008). Sexual reproduction provides the possibility for increasing genetic diversity through recombination, thus contributing not only to ameliorate loss of genetic diversity due to founder events, but also to fuel the opportunities for local adaptation and the ability of colonizers to respond to unpredictable environmental fluctuations in the new range(s) (Eckert, 2002; Novak and Mack, 2005; reviewed in Barrett, 2011). Despite the clear advantages of sexuality, asexual reproduction might be favored under unreliable circumstances, as it provides reproductive assurance and enables the persistence of individuals in unfavorable habitats for sexual reproduction or avoids the costs associated with sexual reproduction, allowing small populations and adaptive genotypes to rapidly establish and spread (Eckert, 2002; Barrett, 2015; reviewed in Vallejo-Marín et al., 2010).
Long-distance dispersal is frequently associated with strong founder effects and loss of genetic diversity, thus exposing founder individual(s) to strong mate limitation both at the establishment of the first viable population(s) and during range expansion (Baker, 1955, 1965; Stebbins, 1957). This is particularly relevant in obligated outcrossing species, such as self-incompatible or heterostylous plants, in which compatible mates might be lost during long-distance dispersal (e.g., Ornduff, 1987; Hollingsworth and Bailey, 2000; Barrett et al., 2008; Zhang et al., 2010). Under this scenario, a switch to uniparental reproduction, either through self-fertilization or increased asexual reproduction, might be selectively advantageous and foster invasion (reviewed in Pannell et al., 2015). Transitions to asexual reproduction or selfing have been documented for several introduced species, such as the clonals Fallopia japonica in the UK (Hollingsworth and Bailey, 2000), Eichhornia crassipes in China (Zhang et al., 2010), Oxalis pes-caprae in Mediterranean regions (Baker, 1965; Ornduff, 1987), and Arundo donax in Australia (Haddadchi et al., 2013), and the self-compatibles Echinochloa microstachya in Australia (Barrett and Husband, 1990), Echium plantagineum in Australia and Canary Islands (Petanidou et al., 2012), and Gomphocarpus physocarpus in Australia (Ward et al., 2012). Additionally, higher rates of uniparental reproduction in introduced and in invasive species compared with natives or with species that failed to establish, have been reported by several studies (Mulligan and Findlay, 1970; Rambuda and Johnson, 2004; Silvertown, 2008; van Kleunen et al., 2008; Marco et al., 2010). Nevertheless, comparative studies of plant reproductive strategies in native and invaded ranges are scarce (but see Brown and Eckert, 2005; Lavergne and Molofsky, 2007; Petanidou et al., 2012).
Sexual and asexual reproductive strategies frequently co-occur in flowering plants and, although this dual strategy was proven to be advantageous (Silander, 1985; Bengtsson and Ceplitis, 2000; Van Drunen et al., 2015), it can also lead to allocation trade-offs and antagonist interactions between reproductive modes, such as the interference generated by clonal growth in the opportunities for mating (Handel, 1985; Vallejo-Marín et al., 2010; Barrett, 2015; Van Drunen et al., 2015). By reducing the number of mating partners and by increasing the opportunities for geitonogamous pollen dispersal, clonal growth interferes with sexual reproduction in reducing not only the offspring sired, but also its quality and fitness (e.g., Handel, 1985; Charpentier, 2002; Somme et al., 2014; but see Van Drunen et al., 2015). Allocation trade-offs occur when the production of sexual and asexual structures compete for the resources available from the total resource pool (van Kleunen et al., 2002; Thompson and Eckert, 2004; Liu et al., 2009), or through the replacement of sexual structures by asexual ones or vice-versa (e.g., production of inflorescences instead of vegetative shoots, Geber et al., 1992; production of bulbils in the inflorescences instead of flowers, Ronsheim and Bever, 2000; or production of flowers from meristems that in previous years resulted in vegetative tissue, Savinykh, 2003). Therefore, it is expected that differential fitness of the two strategies will affect the balance between sexual and asexual reproduction in the population over time (Silvertown, 2008; Vallejo-Marín et al., 2010; Van Drunen et al., 2015). If strong trade-offs between investment in sexual and asexual reproduction occur, rapid clonal expansion may limit allocation to flowering and seed production (Vallejo-Marín et al., 2010). However, evidence for fitness trade-offs between sexual and asexual reproduction is ambiguous (Van Drunen et al., 2015). Although several studies support a trade-off between the two strategies (e.g., van Kleunen et al., 2002; Thompson and Eckert, 2004; Liu et al., 2009; Van Drunen and Dorken, 2012), studies at the genet level are scarce and many of them failed to detect such trade-offs between reproductive strategies (Vallejo-Marín et al., 2010; Van Drunen and Dorken, 2012).
Oxalis pes-caprae is a clonal tristylous species native to South Africa and invasive throughout all Mediterranean regions of the world. This species reproduces by two contrasting strategies: asexually through the profuse production of bulbs (Pütz, 1994; Vilà et al., 2006) and sexually trough a highly specialized mechanism, tristyly and heteromorphic self-incompatibility system (Ornduff, 1987) that promote cross-fertilization and increased genetic diversity (Barrett, 2002). Sexual and asexual reproduction occurs in both native and invaded areas, but the contribution of each reproductive mode differs between ranges (Castro et al., 2007, 2013; Ferrero et al., 2015). In the native range, isoplethic populations occur (Ornduff, 1987; Turketti, 2010; Ferrero et al., 2015), i.e., populations with similar proportions of the three reciprocal style morphs (long-, mid-, and short-styled morphs, hereafter L-, M-, and S-morph, respectively), indicating that populations are in equilibrium and that sexual reproduction is expected to be the main reproductive mode. In the invaded range, two different scenarios appear to be occurring. Until very recently, the main scenario was a transition toward clonality in which the pentaploid (5x) S-morph was the dominant form, and thus asexual reproduction through bulbs has been pointed as the prevailing mechanism of reproduction and spread (Baker, 1965; Ornduff, 1987; Castro et al., 2007). Additionally, a complete sterile double-flowered form was also reported to be successfully spreading in south western Iberian Peninsula (Castro et al., 2007). However, we have recently detected the re-acquisition of sexual reproduction likely fueled by multiple introductions of compatible mating partners [tetraploid (4x) L-, M-, and S-morph individuals; Castro et al., 2013; Ferrero et al., 2015]. This is the first study exploring the role of reproductive traits in the invasion success of O. pes-caprae.
The objective of this study was to quantify changes in reproductive traits in invasive populations of O. pes-caprae and evaluate whether these differences could explain the prevalence of some floral forms in the invaded range of the western Mediterranean basin and be involved in the invasion success of this species. We compared the investment in sexual and asexual reproduction between native and invasive individuals, and among sexual (4x L-, M-, and S-morph), predominately asexual (5x S-morph) and obligated asexual individuals (4x sterile double-flowered form) found in the invaded range. Based on the invasion history of O. pes-caprae and on a trade-off hypothesis between allocation to sexual and asexual reproduction, we expected that, in the invaded range, selection has promoted individuals with an increased capacity for investment in asexual reproduction in detriment of sexual reproduction, especially among the mostly clonal forms; in sexual forms the trade-off between the two strategies might be more dependent of the environmental context. Still, the low sexual success of sexual forms in the invaded area (due to low mate availability and/or genetic depauperated populations; Castro et al., 2013; Ferrero et al., 2015) may generate a context promoting asexuality in comparison with sexual forms from native populations. Thus, we hypothesized that asexual forms would have significantly higher asexual potential than sexual forms in order to become dominant in the invaded range, and that both would have significantly higher asexual potential than sexual forms from the native area where sexual reproduction prevails. Our findings are discussed in the light of biological invasions and of the role of reproductive traits in successful invasion.
Materials and Methods
Plant Species
Oxalis pes-caprae L. (Oxalidaceae), Bermuda buttercup, is a geophyte with a deeply buried annual bulb that produces subterranean stems bearing a rosette of leaves and several inflorescences of yellow flowers arranged in umbellate cymes (Vilà et al., 2006; Sánchez-Pedraja, 2015). It is a tristylous species with a heteromorphic self-incompatibility system (Ornduff, 1987). Thus, the production of viable offspring only occurs after legitimate pollination between individuals with reciprocal style morphs. Double-flowered sterile individuals have also been frequently observed in the western Mediterranean basin (Castro et al., 2007) and sporadically in South Africa (Salter, 1944; Suda and Oberlander, personal communication). The Bermuda buttercup has a high capacity for asexual reproduction through a profuse production of bulbs. The main bulb produces a fasciculate root with contractile properties that grows deeper in the soil some centimeters each year (Pütz, 1994), and later in the season or under stressful conditions (e.g., soil perturbation), the subterranean stems produce a high number of small bulbs (Young, 1968; Verdaguer et al., 2010; authors personal observations). Furthermore, O. pes-caprae is a polyploid species, with diploid (2n = 2x = 14 chromosomes), tetraploid (2n = 4x = 28 chromosomes), and pentaploid (2n = 5x = 35 chromosomes) individuals. In South Africa, all cytotypes have been reported, although the 5x cytotype appears to be extremely rare (Ornduff, 1987; te Beest et al., 2012; Ferrero et al., 2015). Contrarily, the 5x is the dominant cytotype in the invaded ranges worldwide, although in Australia and recently in the western Mediterranean region, the 4x has also been reported (Symon, 1961; Michael, 1964; Castro et al., 2007, 2013).
The Bermuda buttercup was introduced into the Mediterranean basin in the end of the 18th century, most probably multiple times (Vignoli, 1937; Galil, 1968; Signorini et al., 2011), and spread widely afterward. The species was soon recognized as a weed in several Mediterranean areas (e.g., Sicily, Hildebrand, 1887; Canary Islands, Morris, 1895; Algeria, Ducellier, 1914; Balearic Islands, Knoche, 1922; Malta and neighbor islands, Borg, 1927; Tunisia, Chabrolin, 1934), including Portugal where it was described as abundant in orchards (Henriques, 1920; Vasconcelos and Moreira, 1976). Early introductions of the plant occurred due to its ornamental value, and later through soil movement in agriculture, horticulture, and gardening (Michael, 1964; Signorini et al., 2011). Still, the routes of (repeated) introduction to the Mediterranean basin and other invaded regions are not clear.
Field Sampling
Extensive field sampling for bulb harvesting was conducted during February and March 2010 in the invaded range of the western Mediterranean basin (MB), and during August 2011 in the native area, South Africa (SA). All the necessary permits for plant collection were obtained. In the invaded range, an additional effort was made to sample throughout the regions where trimorphic populations and the sterile double-flowered form are more common (Castro et al., 2007, 2013). All floral forms and cytotypes found in this invaded range were included in our study, i.e., the 4x L-morph, 4x M-morph, 4x S-morph, 5x S-morph, and the 4x sterile double-flowered individuals (Supplementary Table 1). In the native range, the field sampling was conducted across most of the latitudinal and longitudinal distribution of the species (Salter, 1944; Supplementary Table 1). Our extensive sampling in the native range confirmed previous results showing that the 5x S-morph and the double-flowered individuals are extremely rare in South Africa (Ferrero et al., 2015). For this reason, only 4x L-morph, 4x M-morph, and 4x S-morph native plants were included in this comparative study. In each population, we sampled bulbs from 10 individuals per floral form, separated at least 5-m apart to avoid re-sampling clones of the same individual. Sampled populations were characterized for style morph frequency and cytotype composition as described in Castro et al. (2013).
Common Garden Experiment
To investigate if there were differences in sexual and asexual reproductive traits between native and invasive plants, we conducted a common garden experiment at the Botanical Garden of the University of Coimbra, where individual plants from both areas were grown outdoors under similar optimal conditions. To remove potential maternal effects, bulbs from SA and the MB were grown for one and two generations, respectively, before sexual and asexual investment traits were measured. In June 2012, all bulbs were harvested and stored in paper bags. During September 2012, they were weighed and weight values were recorded as initial bulb weight. The analysis of the dispersion of the initial bulb weight allowed us to select one bulb per individual matching similar overall mean weights (mean ± SD, 0.463 ± 0.086 g). In total, 338 bulbs were selected, representing 29 populations and 137 individuals from the native area, and 13 populations and 201 individuals from the invaded range (Supplementary Table 1). This selection reflected the different reproductive strategies found in SA and MB: sexual (4x L-, M-, and S-morphs), predominately asexual (5x S-morph), and obligated asexual (4x sterile double-flowered form).
Bulbs were individually planted ∼2.0 cm below the soil surface in 2 L plastic pots (9.6 cm × 9.6 cm × 21.5 cm) filled with commercial substrate, and pots were randomized at the beginning of the experiment. Before flowering, plants were covered with a mosquito net to avoid undesirable pollination. To characterize sexual and asexual reproduction performance, we measured the following traits: (a) bulb viability, (b) occurrence of flowering; (c) floral display; (d) biomass invested in sexual and asexual structures; and (e) production of diaspores through sexual and asexual means (fruit, seed, and bulb production). During the flowering peak, we classified each individual as either vegetative or reproductive, and we collected one flower per inflorescence, when produced, into individual paper bags for later estimation of: (a) mean flower weight and (b) total weight investment in flowers per plant. Inflorescences were periodically monitored, and were collected when senescent, allowing us to simultaneously assess: (a) the total number of flowers produced per plant, i.e., floral display, and (b) the total investment in inflorescences measured as dry weight. Total and average investments in the production of sexual structures were estimated for each plant. Fruit and seed production were obtained by cross pollinating three flowers per plant using reciprocal style morphs of the same area of origin. Fruit set was calculated as the proportion of flowers that developed into fruits and seed production as the mean number of seeds per fruit. We calculated a measure of sexual potential for each plant by multiplying the total number of flowers produced by the mean fruit and seed production. Bulbs were harvested by the end of the season. The investment in asexual structures was quantified by assessing the number of bulbs produced per plant, and total and mean bulb weight per plant.
Statistical Analysis
Data was grouped according to the following criteria: (a) area of origin (South Africa native range, SA, and invaded range of the Mediterranean Basin, MB) and (b) reproductive strategy. We defined reproductive strategy as: sexual, including 4x plants with morphologically functional flowers (Sex); predominantly asexual, 5x S-morph plants that reproduce mostly asexually despite some sporadic ability to produce viable offspring (Asex; Castro et al., 2013; Costa et al., 2014, in press); and, obligated asexual, 4x double-flowered sterile mutants (St). Accordingly, all individuals were assigned to one of the following groups: South African 4x sexual forms (SA4xSex), Mediterranean basin 4x sexual forms (MB4xSex), Mediterranean basin 5x predominantly asexual form (MB5xAsex) and Mediterranean basin 4x obligated asexual form (MB4xSt, double-flowered sterile mutant).
The above groups were defined as fixed factor in generalized linear mixed models (GLMM) to assess differences in sexual and asexual traits. GLMMs enabled us to model variables that did not completely fulfill the assumptions of a standard linear model and had the advantage to allow the incorporation of random factors in the models (Bolker et al., 2009). Although the initial bulb weight was fairly homogenous, this variable was included as covariate to account for possible differences caused by bulb weight. Population and individual were defined as random factors and these were removed from the models whenever their variance was lower than the variance of the residuals (Bolker et al., 2009). When both random factors were removed, a generalized linear model (GLM) was used instead (Supplementary Table 2). A binomial distribution with a logit link function was used to model bulb viability and probability of flowering; a Poisson distribution with a log link function was used to model the number of flowers, inflorescences and bulbs per plant; and a Gaussian distribution with an identity link function was used to model the mean number of flowers per inflorescence, mean flower and inflorescence weight, total flower and inflorescence weight per plant, total weight of sexual structures per plant, fruit set (arcsine transformed), mean seed production, sexual potential, mean and total bulb weight. In all cases, differences between least-square means were tested pairwise through multiple comparisons. To evaluate the existence of trade-offs between sexual and asexual investments, correlations between the amount of biomass invested in sexual and asexual structures were calculated for the entire dataset and for each group separately. All statistical analyses were performed in R version 3.1.1 (R Core Team, 2014) using the packages “car” for GLMs and Type-III analysis of variance (Fox and Weisberg, 2015), “nlme” for linear and non-linear mixed models (Pinheiro et al., 2015), and “multcomp” for multiple comparisons after Type-III analysis of variance (Hothorn et al., 2008), and “stats” for GLMs (R Core Team, 2014).
Results
Bulb Viability and Probability of Flowering
Results from all statistical analyses are summarized in Supplementary Table 2. Bulb viability was high, varying between 88% in invasive sexual individuals (MB4xSex) and 94% in native sexual individuals (SA4xSex), with no statistically significant differences being observed among groups (χ23,338 = 3.49, P = 0.322; Supplementary Figure 1A).
The probability of producing floral structures differed among groups (χ23,305 = 11.23, P = 0.011), with the obligated asexual individuals (MB4xSt) having a significantly higher probability to remain vegetative than the other invasive groups (i.e., MB4xSex and MB5xAsex; P < 0.05), while native sexual plants had intermediate values between the two extremes (Supplementary Figure 1B).
Sexual Traits: Floral Display
Floral display differed among groups in terms of the number of flowers per inflorescence (χ23,276 = 19.07, P < 0.001), number of inflorescences per plant (χ23,276 = 47.89, P < 0.001) and total number of flowers per plant (χ23,276 = 31.68, P < 0.001; Figures 1A–C). The native sexual plants and the invasive predominantly asexual plants (MB5xAsex) produced inflorescences with more flowers than the invasive sexual individuals (MB4xSex; P < 0.05), while the obligated asexual form (MB4xSt) had a lower and highly variable mean value not differing from the other three groups (Figure 1A). The groups with ability for sexual reproduction (SA4xSex and MB4xSex), even if only sporadically (MB5xAsex), produced significantly more inflorescences than the obligated asexual individuals, resulting in larger total floral display per plant (P < 0.05; Figures 1B,C).
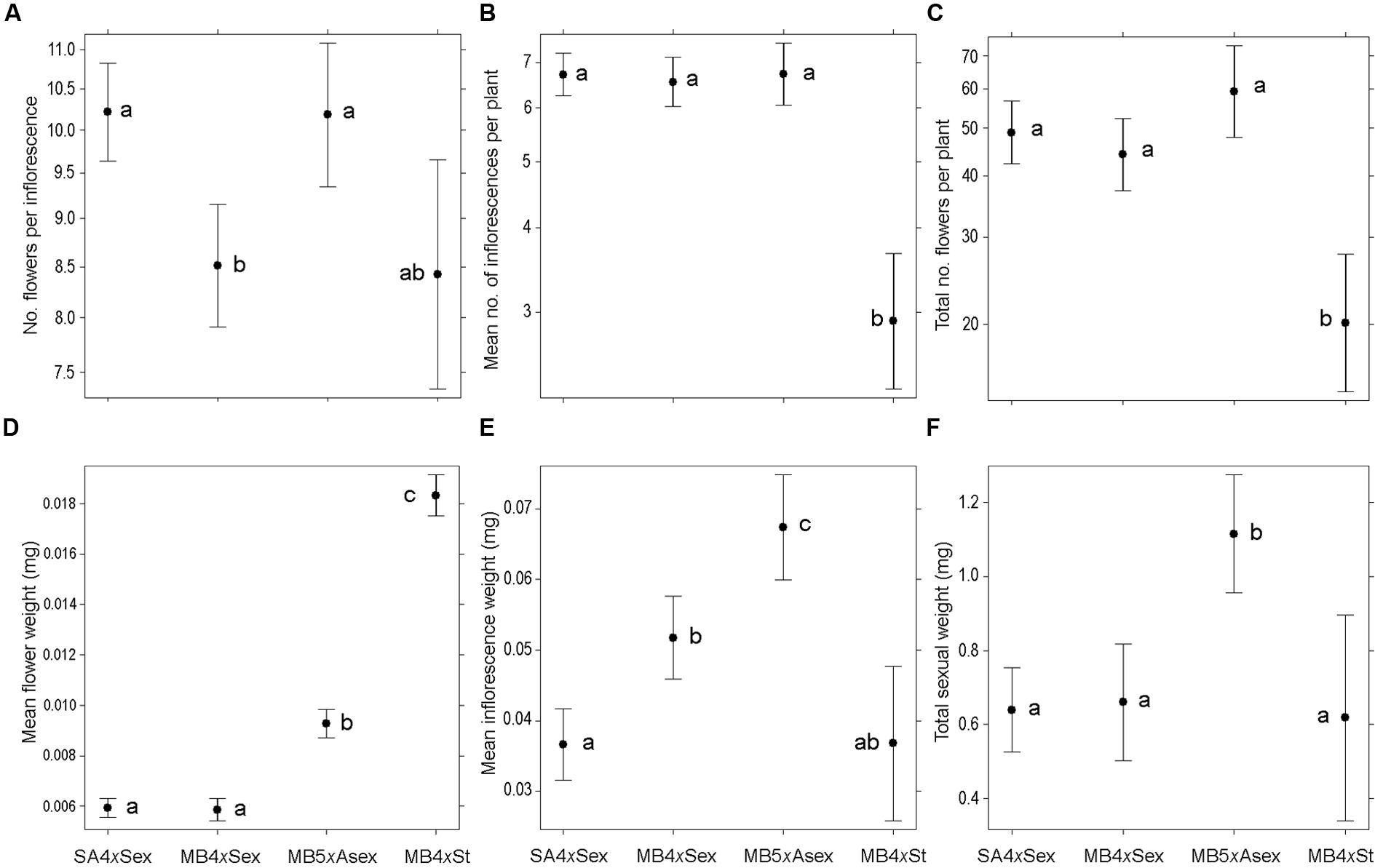
FIGURE 1. Floral display and biomass invested in sexual reproductive structures among different forms of Oxalis pes-caprae from its native and invaded areas and with distinct reproductive strategies: South African 4x sexual forms (SA4xSex), Mediterranean basin 4x sexual forms (MB4xSex), Mediterranean basin 5x predominantly asexual form (MB5xAsex), and Mediterranean basin 4x obligated asexual form (MB4xSt, sterile double-flowered form). (A) Mean number of flowers per inflorescence; (B) Mean number of inflorescences per plant; (C) Total number of flowers per plant; (D) Mean flower weight (mg); (E) Mean inflorescence weight (mg); (F) Total weight of sexual structures per plant (mg). Values are given as model-adjusted back-transformed least-square means and 95% confident intervals. Significant differences among factors are indicated with different letters (P < 0.05).
Significant differences among groups were also observed in the biomass invested for the production of sexual structures (Figures 1D–F, Supplementary Figure 1), namely in the mean flower and inflorescence weight (χ23,276 = 820.80, P < 0.001 and χ23,276 = 51.78, P < 0.001, respectively; Figures 1D–E), total flower and inflorescence weight per plant (χ23,276 = 28.08, P < 0.001 and χ23,276 = 44.39, P < 0.001, respectively; Supplementary Figures 1C,D), and total weight of sexual structures per plant (χ23,276 = 32.30, P < 0.001; Figure 1F). Obligated asexual individuals (MB4xSt) had significantly heavier flowers, followed by the MB5xAsex, and the native and invasive sexual individuals had lower flower weights (P < 0.05; Figure 1D). The same trend was observed for total flower weight per plant, except for the obligated asexual individuals (MB4xSt) which produced less inflorescences (Figure 1B) and consequently less flowers (Figure 1C), lower total flower weight (Figure 1F) and reduced investment in total flower biomass (Supplementary Figure 1C). A different scenario was found for mean inflorescence weight, which was significantly lower for the obligated asexual individuals and sexual native plants. Sexual invasive individuals presented intermediate inflorescence weight, and invasive predominantly asexual plants had significantly heavier inflorescences (P < 0.05; Figure 1E; a similar pattern is observed for the total inflorescence weight per plant; Supplementary Figure 1D). Despite the differences in the number and biomass of reproductive structures among all groups, the total investments in the production of sexual structures per plant did not differ among groups except for the MB5xAsex, which presented significantly higher weights (P < 0.05; Figure 1F).
Production of Dispersal Units: Sexual and Asexual Strategies
Sexual fitness differed significantly among groups (fruit set: χ22,251 = 47.38, P < 0.001; seed production: χ22,246 = 89.44, P < 0.001; and sexual potential: χ22,251 = 15.77, P < 0.001). Fruit and seed production were significantly lower in invasive plants than in native ones and, among invasive groups, it was significantly lower in the predominantly asexual individuals (P < 0.05; Figures 2A,B). The calculation of a measure of sexual potential revealed that native sexual individuals had a significantly higher success than invasive plants (P < 0.05); however, no significant differences were detected in sexual potential between sexual invasive and predominantly asexual invasive individuals (Figure 2C).
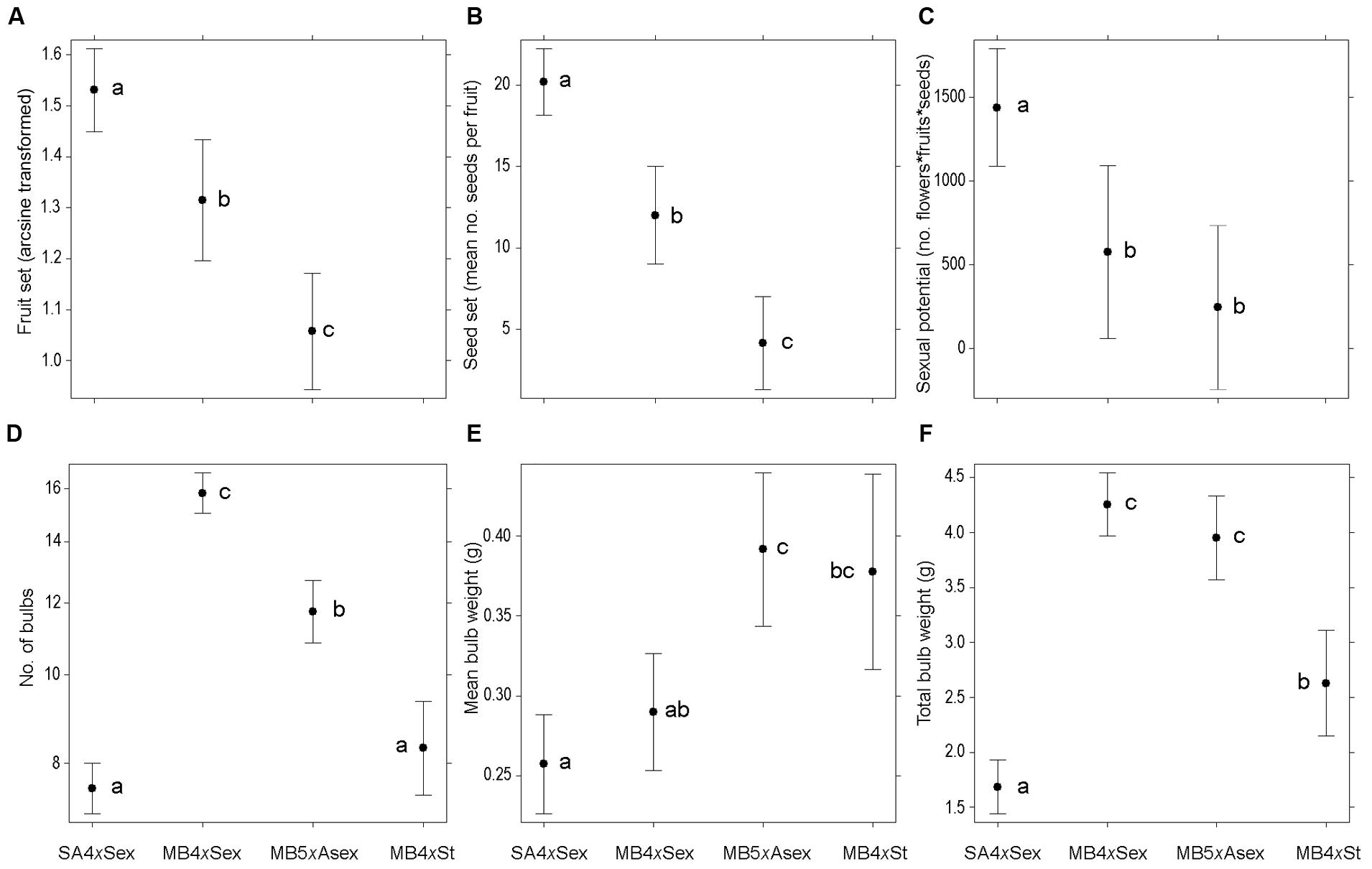
FIGURE 2. Production of sexual and asexual diaspores among different forms of Oxalis pes-caprae from its native and invaded areas and with distinct reproductive strategies: South African 4x sexual forms (SA4xSex), Mediterranean basin 4x sexual forms (MB4xSex), Mediterranean basin 5x predominantly asexual form (MB5xAsex) and Mediterranean basin 4x obligated asexual form (MB4xSt, sterile double-flowered form). (A) Fruit set, given as arcsine of the proportion of flowers developing into fruit; (B) Seed production, given as the mean number of seeds per fruit; (C) Sexual potential, given as a measure of sexual ability of the plant obtained multiplying the number of flower produced in a plant by the fruit and seed production; (D) Mean number of bulbs per plant; (E) Mean bulb weight (mg); (F) Total weight of the bulbs produced per plant (mg). Values are given as model-adjusted back-transformed least-square means and 95% confident intervals. Significant differences among factors are indicated with different letters (P < 0.05).
Asexual traits also differed significantly among groups, either measured as number of bulbs (χ23,305 = 350.13, P < 0.001), mean bulb weight (χ23,305 = 27.36, P < 0.001) or total bulb weight per plant (χ23,305 = 211.13, P < 0.001). Invasive sexual plants produced more bulbs per plant than predominantly asexual individuals, which also produced more bulbs than native sexual and invasive obligated asexual individuals (P < 0.05; Figure 2D). However, predominantly asexual individuals had significantly heavier bulbs than native and invasive sexual plants (P < 0.05), while obligated asexuals had fairly heavy, but highly heterogeneous bulbs that did not differ significantly from the other invasive groups (Figure 2E). There was a clear and significantly higher investment in total bulb weight by the invasive sexual and predominantly asexual plants than the other groups, as well as in the obligated asexual individuals in comparison with native sexual plants (P < 0.05; Figure 2F).
Trade-off between Sexual and Asexual Investment
No trade-off was observed in the biomass invested in sexual and asexual structures. On the contrary, the production of sexual structures was positively correlated with the biomass invested in the production of bulbs, except for invasive sexuals and obligated asexuals (total: r = 0.214, P < 0.001; analyses by group: SA4xSex: r = 0.286, P = 0.0182; MB4xSex: r = -0.0223, P = 0.800; MB5xAsex: r = 0.449, P < 0.001; MB4xSt: r = 0.183, P = 0.381; Figure 3).
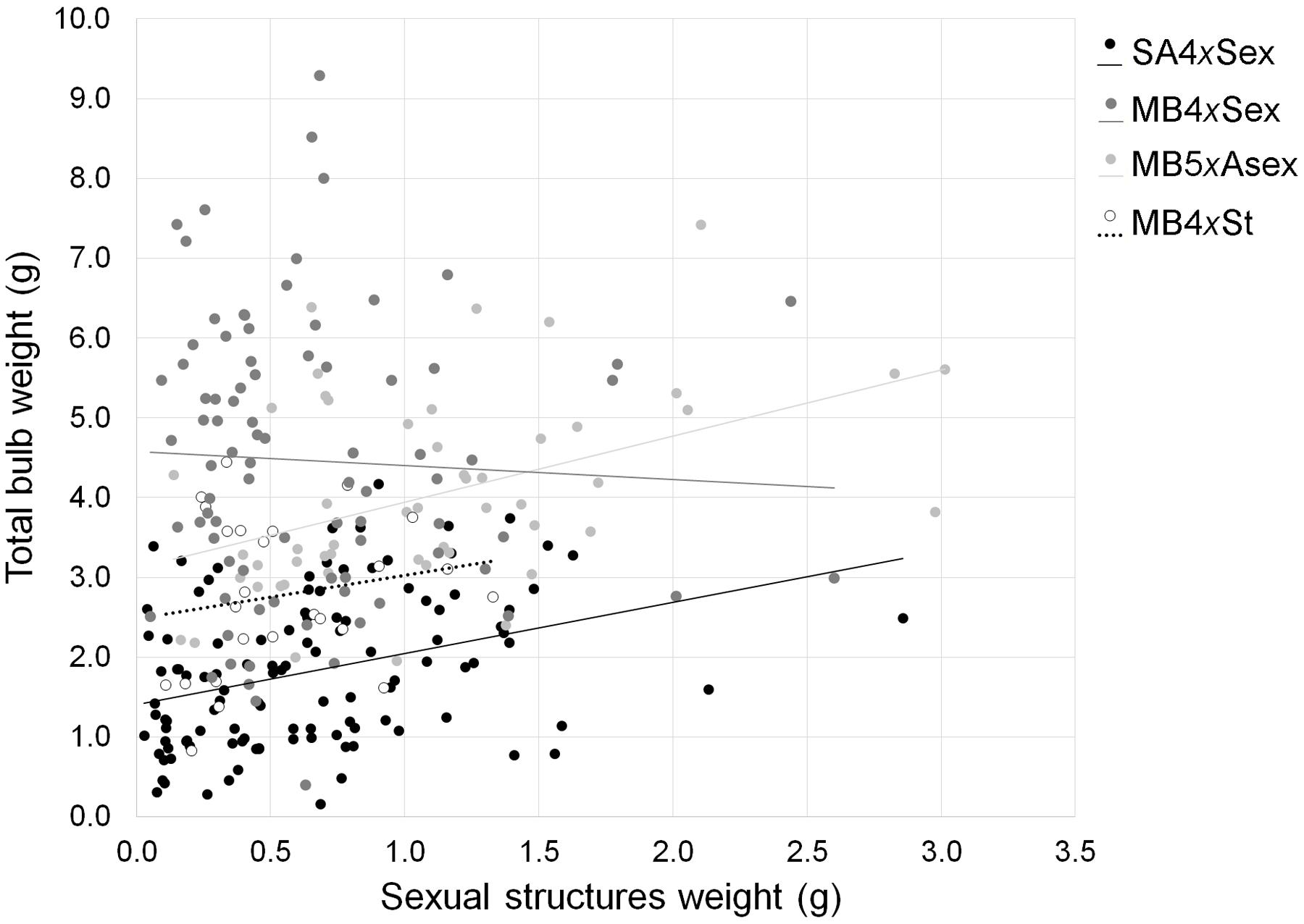
FIGURE 3. Correlation between biomass invested in the production of sexual and asexual structures among different forms of Oxalis pes-caprae from its native and invaded areas and with distinct reproductive strategies: South African 4x sexual forms (SA4xSex), Mediterranean basin 4x sexual forms (MB4xSex), Mediterranean basin 5x predominantly asexual form (MB5xAsex) and Mediterranean basin 4x obligated asexual form (MB4xSt, sterile double-flowered form).
Discussion
Our results indicate the occurrence of evolutionary changes in the reproductive traits of invasive populations of O. pes-caprae. Indeed, most of the traits evaluated, differed between native and invasive populations and among individuals with different reproductive strategies when grown in the same environment. In particular, we found that: (1) overall, plants with sexual ability (including the 5x S-morph) had higher probability of flowering and larger floral displays than the sterile forms; (2) the total investment in the production of floral structures was significantly higher in the 5x form than in the remaining forms; this suggests an effect of the ploidy level in the overall size of the structures and, in the case of the sterile double-flowered form, a trade-off between the number of flowering structures and the resources needed to produce them (i.e., heavier sterile flowers resulting in lower number of inflorescences); (3) differences in the production of bulbs and seeds revealed that native plants had higher sexual fitness, while a transition toward clonality was clear for the invasive forms; (4) differences were also observed among invasive individuals, with the sexual forms producing more dispersal units (seeds and small bulbs), the predominantly asexual form producing an inter-medium number of large bulbs, and the sterile form being apparently less aggressive and producing less, yet large, bulbs; (5) finally, no trade-off between sexual and asexual investments was observed. Below, we discuss our results in light of the complex invasion history of O. pes-caprae and draw hypotheses on how reproductive traits could have been involved in the invasion success and in the prevalence of some forms in the invaded range of the western Mediterranean region.
Reproduction: Traits and Strategies
Reproduction determines the number and genetic composition of dispersal units, being vital for the establishment and spread of plant populations after long-distance dispersal (e.g., Sakai et al., 2001; Ness et al., 2010). The relative contribution of different reproductive modes varies depending on the ecological and genetic factors under which colonizers are subjected (e.g., Dorken and Eckert, 2001; Eckert, 2002; Herben et al., 2015). Our results showed remarkable differences in several reproductive traits between ranges and among forms with different reproductive strategies. Overall, a transition to clonality was observed among invasive plants: native individuals had a higher sexual fitness than invasive ones, which in contrast had higher asexual fitness than natives. These observations matched our expectations mainly by two reasons, described below.
Firstly, genetic diversity of native populations is higher than that of invasive ones (Ferrero et al., 2015), and this is expected to affect the sexual fitness. Sexual reproduction is the main mechanism of reproduction in the native range, where floral polymorphism promotes outcrossing and thus genetic diversity and frequency dependent selection governs isoplethic populations (Ornduff, 1987; Turketti, 2010; Ferrero et al., 2015). Contrarily, invasive populations are highly constrained by the scarcity of compatible mates (Castro et al., 2007, 2013), being dominated by 5x S-morphs with residual sexual reproduction (Costa et al., 2014, in press), and thus with low recombination probabilities. Additionally, strong founder effects led to a decrease in genetic diversity of invasive populations (Ferrero et al., 2015). Altogether, these factors significantly impacted genetic composition of invasive populations and, consequently, plant fitness under outcross pollinations. Genetic depauperation after long distance dispersal has been described in several other invasive species (e.g., Dlugosch and Parker, 2008; Zhang et al., 2010), and although multiple introductions can ameliorate their consequences (Novak and Mack, 2005; Dlugosch and Parker, 2008; Simberloff, 2009), negative impacts of low genetic diversity for plant reproduction have been shown (Barrett, 2002; Crawford and Whitney, 2010). However, genetic bottlenecks do not necessarily hinder the adaptive potential of invasive species (Barrett et al., 2008; Dlugosch and Parker, 2008; Rollins et al., 2013).
Secondly, under unfavorable conditions for sexual reproduction in invasive populations, i.e., strong mate limitation (either due to monomorphic populations or due to the predominance of asexual 5x individuals; Baker, 1965; Ornduff, 1987; Castro et al., 2007, 2013; Ferrero et al., 2015), we expected that selection would benefit individuals with an increased capacity for investment in asexual reproduction. Indeed, uniparental reproduction has been proposed to be selectively advantageous under scenarios of strong mate limitation, such as invasions, rapid range expansion, island colonization and meta-population dynamics (Baker’s Law; Baker, 1965; Pannell et al., 2015). Our results corroborate this prediction: regardless of the reproductive strategy, invasive O. pes-caprae forms invested significantly more in asexual reproduction than natives. An enhancement in clonal reproduction in invasive populations in comparison with natives has also been observed in several other species. For example, invasive Butomus umbellatus individuals were more likely to produce bulbils than native individuals (Brown and Eckert, 2005); rapid selection of genotypes with stronger vegetative growth was observed in Phalaris arundinacea (Lavergne and Molofsky, 2007), and greater vegetative reproduction in the invasive ranges of Achillea millefolium and Hypericum perforatum than on their native ranges (Beckmann et al., 2009). Interestingly, differences in bulb production have also been observed among invasive populations of O. pes-caprae in the Mediterranean basin, with insular populations having higher dispersal potential than continental ones (Vilà and Gimeno, 2006). Clonal reproduction was one of the traits identified by Baker (1965) for the “ideal weed” and there are several studies addressing invasive species traits that support this reproductive strategy as one of the features involved with successful invasions (e.g., Pyšek and Richardson, 2007; Silvertown, 2008; Marco et al., 2010).
As expected, individuals with the ability to reproduce sexually, including the predominantly asexual plants, invested more in the production of floral structures than the sterile form. This is in accordance with a strategy to promote sexual reproduction either by investing in attractive floral display (i.e., larger inflorescences, inflorescences with larger flower displays) or by producing more sexual potential units (Barrett, 2002). A trade-off between the number of inflorescences produced and the considerable amount of energy necessary to produce double-flowers might explain the reduced floral display of the sterile double-flowered form (see below). Interestingly, the 5x form produced larger flowers and larger inflorescences that resulted in a higher biomass investment without a detrimental impact in the floral display. The production of larger reproductive structures is likely related with the ploidy level, since polyploidy is hypothesized to drive significant changes in cell size and, consequently, in overall organ size (Levin, 2002). Apparently, this higher investment did not lead to allocation trade-offs since the 5x cytotype produced similar floral display to other sexual forms.
Besides the differences in the production of sexual and asexual diaspores detected between ranges, different strategies were also observed among invasive forms. As expected, sexual fitness was higher in the 4x sexual forms than in the 5x form, and null in the sterile double-flowered form where the sexual organs were replaced by petals due to a mutation in the genes responsible for the floral development (Weigel and Meyerowitz, 1994). The lower sexual fitness of the 5x individuals after outcrossing is mainly due to its odd ploidy level; although these 5x individuals are able to produce some viable gametes, they also produce unviable gametes with variable ploidies (Vignoli, 1937; Signorini et al., 2013; Costa et al., 2014), diminishing significantly the production of offspring through seed. However, and as described above, the 5x individuals produced slightly larger floral displays and bigger floral structures; these features increase the number of gametes and the attractiveness of the plants for pollinators, which might contribute to ameliorate the low sexual potential of the 5x individuals. Additionally, differences in the asexual traits were also detected among invasive forms. Under optimal resource conditions, the obligated asexuals invested significantly less in bulb production than the other invasive forms, producing larger bulbs but in smaller amounts (like native plants) than the other invasive forms. Interestingly, the sexual and predominately asexual forms allocated a similar amount of energy to the production of bulbs; however, while the former invested resources in producing many small bulbs, the later invested in less but larger bulbs. These patterns agree with trade-off models for propagule number and size which predict that in optimal environments it is preferable to maximize offspring quantity, whereas in stressful conditions (such as the limitation of sexual partners) it is preferable to invest in offspring quality (Smith and Fretwell, 1974; Sadras, 2007). Based on this model, we could hypothesize that selective pressures during the invasion of the predominantly asexual form might have benefited larger bulbs, while selective pressures over asexual propagule production are not expected to be so strong for the sexual forms that have an additional reproductive mode (Costa et al., unpublished results). Oxalis pes-caprae bulb weight has been pointed as an important feature of the invasion process, especially under stressful conditions (Lane, 1984; Sala et al., 2007), with parental bulb weight significantly impacting plant biomass in shaded environments, as evident by the production of significantly more leaves in plants originated from bigger bulbs than from smaller ones (Verdaguer et al., 2010). However, although fitting nicely the results, there are several lines of evidence that do not completely support this hypothesis and make our findings difficult to interpret. First, it is difficult to disentangle the effects of ploidy level from those related with evolutionary changes. Although, bulb size of the offspring of 4x sterile double-flowered form and the 5x S-morph were similar, the larger bulbs in the 5x individuals might be driven by ploidy, similarly to the pattern observed in the flowering structures (results herein) and to the patterns observed in other polyploid complexes (Levin, 2002). Second, 5x S-morph individuals showed that parent bulb weight has a small overall effect on O. pes-caprae plant biomass (Sala et al., 2007; Verdaguer et al., 2010). In general, bulbs emerged successfully and vigorously regardless of their sizes (Vilà et al., 2006; Verdaguer et al., 2010), still, parent bulb size might be particularly important for plant emergence and initial development, depending on the conditions where the plant is growing (Vilà et al., 2006; Sala et al., 2007; Verdaguer et al., 2010). Regardless of the effects in early stages, bulb size was not determinant for the development of adult plants and subsequent offspring production, possibly because further plant growth might become independent of this storage organ once the plant starts to photosynthesize. Finally, the production of bulbs in O. pes-caprae was shown to be plastic and highly dependent of nutrient availability (Sala et al., 2007). If bulb weight has no fitness advantage, then producing many small bulbs would be advantageous, especially when mate limitation is strong and no allocation trade-off between sexual and asexual investment is observed (see below). In this context, sexual invasive forms have a higher dispersal potential, through both sexual and asexual means than the other invasive forms and might become widespread in the future.
Bermuda Buttercup Invasion History: What Have We Learned So Far?
The Bermuda buttercup is a classic example in biological invasions, known as a strictly asexual form that successfully spread in Mediterranean climate regions of the world (5x S-morph; Baker, 1965). However, the origin of this invasive form is still unclear and the colonization history revealed to be more complex and dynamic than previously envisaged. Native populations are composed of the 4x cytotype, with the 5x S-morph being extremely rare (Michael, 1964; te Beest et al., 2012; Signorini et al., 2013; Ferrero et al., 2015). Contrarily, the 5x S-morph dominates all Mediterranean climate regions, except in Australia where both asexual (monomorphic 5x S-morph populations) and sexual populations (4x trimorphic populations) have been reported (Symon, 1961; Michael, 1964; Ferrero, personal observations). The most accepted hypothesis is that the 5x S-morph has been originated from 4x individuals in the introduced range and subsequently introduced in several areas of the world (Krejčíková et al., 2013; Signorini et al., 2013; Ferrero et al., 2015; most probably multiple times in the Mediterranean basin, Signorini et al., 2011), including South Africa, where it was recently reported in a new semi-natural location for the first time (Ferrero et al., 2015). The combination of several factors, including strong heteromorphic incompatibility system, lack of compatible mates and odd ploidy, constrained the production of dispersal units mostly to asexual means, and consequently the successful spread of this form in introduced ranges became dependent on bulb production (Baker, 1965; Ornduff, 1987). Our results strongly support this hypothesis showing a clear selection toward clonality through a significantly increase in the number of bulbs as well as in their size (the latter driven or not exclusively by the ploidy level) in comparison with natives. Producing more bulbs would be selectively advantageous since it increases the number of propagules, while larger bulbs may confer significant advantages under stressful environments, allowing faster plant emergence and providing more reserves, which will translate into larger plants (Vilà and Gimeno, 2006; Sala et al., 2007; Verdaguer et al., 2010; Tavares, 2014).
However, the story does not end here. In the western Mediterranean region, invasive populations seem to be changing very rapidly (Castro et al., 2007, 2013; Costa et al., 2014, in press). Molecular studies have shown an invasion punctuated by multiple introductions of other floral morphs comprised of the tetraploid ploidy level (Ferrero et al., 2015), and field surveys detected a reacquisition of sexual reproduction in this region (Castro et al., 2013; Costa et al., 2014, in press). Although the introduction or incipient occurrence of M- and L-morphs would constitute a source of compatible mates, these individuals are still under a scenario of strong mate limitation due to the dominance of the predominantly asexual 5x S-morph. Thus, they will be subjected to similar strong selective pressures toward uniparental reproduction. Our results clearly support that these new individuals have also diverged from native populations and present an inversion toward uniparental reproduction via asexual reproduction (results herein), but also via changes in the strength of the incompatibility system (Costa et al., unpublished results). Additionally, our results show that these sexual forms have superior reproductive fitness in comparison with the 5x S-morph and the 4x sterile double-flowered form. So, how can we explain the distribution patterns in the western Mediterranean basin? Based on the reproductive traits, the current distribution patterns can only be explained under a scenario of different introduction timings, first with the introduction and spread of the 5x individuals and more recently with the introduction of 4x sexual plants that are starting to become more dominant than previously documented. Given the superior reproductive fitness of the 4x sexual individuals, they are expected to become more dominant in the future. Still, other life-history transition stages and ecological responses, including bulb viability and emergence, competitive ability, resistance to herbivory and response to soil disturbance, need to be addressed in future studies in order to fully characterize the fitness of each form.
The successful spread of the sterile double-flowered form in south–west Iberian Peninsula is particularly intriguing. This form had the lowest dispersal potential among invasive forms and therefore it is likely under a competitive disadvantage with other floral forms. Recent molecular studies have shown a close relationship between these individuals and native plants, supporting the occurrence of several multiple introductions. These multiple introductions might have provided a sufficiently high number of propagules to mediate a successful invasion process. Although species traits are extremely important, several studies have shown that propagule pressure is also a determinant factor for successful invasion (Novak and Mack, 2005; Colautti et al., 2006; Dlugosch and Parker, 2008; Simberloff, 2009). Interestingly, propagule pressure was also shown to be important in colonization by O. pes-caprae along altitudinal gradients within invaded areas (Ross et al., 2008). Additionally, besides ecological and life history traits, human mediated dispersal (e.g., in earlier stages as ornamental plant, and currently through soil movements in agriculture, horticulture and gardening, or through land translocations during road constructions; Michael, 1964; Signorini et al., 2011; Castro et al., 2013) might have also promoted the dispersal of this invasive form, as well as the others (Pyšek and Richardson, 2007).
Trade-offs between Sexual and Asexual Strategies
No trade-off between the production of sexual and asexual structures has been detected in O. pes-caprae. These observations agree with studies in other species (Vallejo-Marín et al., 2010; Van Drunen and Dorken, 2012) and with previous experiments with O. pes-caprae (Vilà and Gimeno, 2006; Verdaguer et al., 2010). This lack of a trade-off might be explained by the particular developmental processes of the plant as the production of flowering structures and bulbs are asynchronous in O. pes-caprae likely reducing the competition for resources between both reproductive processes. In the first half of the plants’ life cycle, most of the energy is redirected to growth and flowering, and only afterward, when the aboveground part of the plant starts to senesce, energy is directed to the production of underground structures, namely to the production of bulbs (Pütz, 1994; Verdaguer et al., 2010). Indeed, we observed the opposed pattern, with a positive correlation between bulb and flower biomass. This could simply be a reflection of plant size rather than resource management strategies.
Conclusion
Different sexual and asexual reproductive traits were quantified between native and invasive populations, as well as among different forms within invasive populations. Different reproductive strategies and ecological and genetic contexts created by long-distance dispersal seem to generate divergent selective pressures in several sexual and asexual reproductive traits. The introduction process seems to have promoted clonal reproduction and this is most probably the major trait driving the invasion success of O. pes-caprae; however, invasive sexual forms have increased dispersal potential and additional means to produce dispersal units and promote heterozygosity. Consequently, invasive sexual forms are expected to be in competitive advantage in relation to the predominately asexual and obligated asexual plants, and thus could become widespread in the invaded range in the future. Historical processes, with the introduction of the predominantly asexual 5x S-morph first and more recently of the 4x sexual morphs, were probably important in establishing the current distributional patterns of the different forms in the western Mediterranean basin. This study shows that invasion processes can be incredibly complex and dynamic, while the interaction between ecological and genetic constraints determined by the invasion process might result in different reproductive strategies which in turn determine the success of invasive populations.
Author Contributions
SC, VF, LN, and JL designed the experiment; SC, VF, JC, and JL conducted field collections; MC, with the collaboration of all the authors, conducted the common garden experiment; SC analyzed the data with the other authors participating in the discussion of the results; SC and MC, with contribution of all the authors, wrote the manuscript.
Funding
This research was supported by FEDER funds through the COMPETE Program and by Portuguese Foundation for Science and Technology (FCT) funds in the ambit of the project PTDC/BIA-BIC/110824/2009, CRUP Acções Integradas Luso-Espanholas 2010 with the project E10/10, MCI-Programa de Internacionalización de la I+D (PT2009-0068) of the Spanish DGICYT (CGL2009-10466 and CGL2013-45941) and the Xunta de Galicia (INCITE09-3103009PR and R2014/036). FCT also supported the work of SC (BPD/41200/2007, Starting Grant IF/01267/2013), MC (PTDC/BIA-BIC/110824/2009; SFRH/BD/89617/2012) and JC (SFRH/BD/89910/2012), and Xunta de Galicia supported the work of VF (I2C Program).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Authors are thankful to Western Cape Nature Conservation Board and Department of Environment and Nature Conservation, Northern Cape, for providing authorizations to collect and undertake scientific research (no. AAA005-00218-0028). The authors are also thankful to Ruben Heleno for critical reading and English review.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2016.00874
References
Baker, G. H., and Stebbins, J. L. (1965). The Genetics of Colonizing Species. New York, NY: Academic Press.
Baker, H. G. (1955). Self-compability and establishment after “long-distance” dispersal. Evolution 9, 347–348. doi: 10.2307/2405656
Baker, H. G. (1965). “Characteristics and modes of origin of weeds,” in The Genetics of Colonizing Species, eds H. G. Baker and G. L. Stebbins (New York, NY: Academic Press), 147–172.
Barrett, S. C. H. (2002). The evolution of plant sexual diversity. Nat. Rev. Genet. 3, 274–284. doi: 10.1038/nrg776
Barrett, S. C. H. (2011). “Why reproductive systems matter for the invasion biology of plants,” in Fifty Years of Invasion Ecology: The Legacy of Charles Elton, ed. D. M. Richardson (Oxford: Oxfrod University Press), 195–210.
Barrett, S. C. H. (2015). Influences of clonality on plant sexual reproduction. Proc. Natl. Acad. Sci. U.S.A. 112, 8859–8866. doi: 10.1073/pnas.1501712112
Barrett, S. C. H., Colautti, R. I., and Eckert, C. G. (2008). Plant reproductive systems and evolution during biological invasion. Mol. Ecol. 17, 373–383. doi: 10.1111/j.1365-294X.2007.03503.x
Barrett, S. C. H., and Husband, B. C. (1990). “The genetics of plant migration and colonization,” in Plant Population Genetics, Breeding, and Genetic Resources, eds A. H. D. Brown, M. T. Clegg, A. L. Kahler, and B. S. Weir (Sunderland, MA: Sinauer & Associates), 254–277.
Beckmann, M., Erfmeier, A., and Bruelheide, H. (2009). A comparison of native and invasive populations of three clonal plant species in Germany and New Zealand. J. Biogeogr. 36, 865–878. doi: 10.1111/j.1365-2699.2008.02048.x
Bengtsson, B. O., and Ceplitis, A. (2000). The balance between sexual and asexual reproduction in plants living in variable environments. J. Evol. Biol. 13, 415–422. doi: 10.1046/j.1420-9101.2000.00187.x
Bolker, B. M., Brooks, M. E., Clark, C. J., Geange, S. W., Poulsen, J. R., Stevens, M. H. H., et al. (2009). Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127–135. doi: 10.1016/j.tree.2008.10.008
Brown, J. S., and Eckert, C. G. (2005). Evolutionary increase in sexual and clonal reproductive capacity during biological invasion in an aquatic plant Butomus umbellatus (Butomaceae). Am. J. Bot. 92, 495–502. doi: 10.3732/ajb.92.3.495
Castro, S., Ferrero, V., Costa, J., Sousa, A. J., Navarro, L., and Loureiro, J. (2013). Reproductive strategy of the invasive Oxalis pes-caprae: distribution patterns of flower morphs, ploidy levels and sexual reproduction. Biol. Invasions 15, 1863–1875. doi: 10.1007/s10530-013-0414-2
Castro, S., Loureiro, J., Santos, C., Ater, M., Ayensa, G., and Navarro, L. (2007). Distribution of flower morphs, ploidy level and sexual reproduction of the invasive weed Oxalis pes-caprae in the Western area of the Mediterranean region. Ann. Bot. 99, 507–517. doi: 10.1093/aob/mcl273
Castro-Díez, P., Valle, G., González-Muñoz, N., and Alonso, Á (2014). Can the life-history strategy explain the success of the exotic trees Ailanthus altissima and Robinia pseudoacacia in Iberian floodplain forests? PLoS ONE 9:e100254. doi: 10.1371/journal.pone.0100254
Charpentier, A. (2002). Consequences of clonal growth for plant mating. Evol. Ecol. 15, 521–530. doi: 10.1023/A:1016057503722
Colautti, R. I., Grigorovich, I. A., and MacIsaac, H. J. (2006). Propagule pressure: a null model for biological invasions. Biol. Invasions 8, 1023–1037. doi: 10.1007/s10530-005-3735-y
Costa, J., Ferrero, V., Castro, M., Jorge, A., Afonso, A., Loureiro, J., et al. (in press). Pollen flow between flowers of the same morph in invasive populations of Oxalis pes-caprae L. in the Western Mediterranean region. Plant Biosyst. doi: 10.1080/11263504.2014.991363
Costa, J., Ferrero, V., Loureiro, J., Castro, M., Navarro, L., and Castro, S. (2014). Sexual reproduction in the invasive pentaploid short-styled Oxalis pes-caprae allows the production of viable offspring. Plant Biol. 16, 208–214. doi: 10.1111/plb.12010
Crawford, K. M., and Whitney, K. D. (2010). Population genetic diversity influences colonization success. Mol. Ecol. 19, 1253–1263. doi: 10.1111/j.1365-294X.2010.04550.x
Dlugosch, K. M., and Parker, I. M. (2008). Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Mol. Ecol. 17, 431–449. doi: 10.1111/j.1365-294X.2007.03538.x
Dorken, M. E., and Eckert, C. G. (2001). Severely reduced sexual reproduction in northern populations of a clonal plant, Decodon verticillatus (Lythraceae). J. Ecol. 89, 339–350. doi: 10.1046/j.1365-2745.2001.00558.x
Ducellier, L. (1914). Note sur la vegétátion de l’Oxalis cernua Thunb. en Algérie. Rev. Gen. Bot. 25:217.
Eckert, C. G. (2002). The loss of sex in clonal plants. Evol. Ecol. 15, 501–520. doi: 10.1023/A:1016005519651
Elton, C. S. (1958). The Ecology of Invasions by Animals and Plants. Chicago, IL: University of Chicago Press.
Ferrero, V., Barrett, S. C. H., Castro, S., Caldeirinha, P., Navarro, L., Loureiro, J., et al. (2015). Invasion genetics of the Bermuda buttercup (Oxalis pes-caprae): complex intercontinental patterns of genetic diversity, polyploidy and heterostyly characterize both native and introduced populations. Mol. Ecol. 24, 2143–2155. doi: 10.1111/mec.13056
Fox, J., and Weisberg, S. (2015). An {R} Companion to Applied Regression, 2nd Edn. Thousand Oaks CA: Sage.
Galil, J. (1968). Vegetative dispersal in Oxalis cernua. Am. J. Bot. 55, 68–73. doi: 10.2307/2440494
Geber, M. A., Watson, M. A., and Furnish, R. (1992). Genetic differences in clonal demography in Eichhornia crassipes. J. Ecol. 80, 329–341. doi: 10.2307/2261015
Goodwillie, C., Kalisz, S., and Eckert, C. G. (2005). The evolutionary enigma of mixed mating systems in plants: occurrence, theoretical explanations, and empirical evidence. Ann. Rev. Ecol. Evol. Syst. 36, 47–79. doi: 10.1146/annurev.ecolsys.36.091704.175539
Haddadchi, A., Gross, C. L., and Fatemi, M. (2013). The expansion of sterile Arundo donax (Poaceae) in southeastern Australia is accompanied by genotypic variation. Aquat. Bot. 104, 153–161. doi: 10.1016/j.aquabot.2012.07.006
Handel, S. N. (1985). The intrusion of clonal growth patterns on plant breeding systems. Am. Nat. 125, 367–384. doi: 10.1086/284348
Hayes, K. R., and Barry, S. C. (2008). Are there any consistent predictors of invasion success? Biol. Invasions 10, 483–506. doi: 10.1007/s10530-007-9146-5
Henriques, J. A. (1920). Patologia vegetal e entomologia agrícola: Oxalis cernua. Gazeta das aldeias 25:146.
Herben, T., Šerá, B., and Klimešová, J. (2015). Clonal growth and sexual reproduction: tradeoffs and environmental constraints. Oikos 124, 469–476. doi: 10.1111/oik.01692
Hildebrand, F. (1887). Experimente über die geschlechtliche Fortpflanzungsweise der Oxalisarten. Bot. Ztg. 45, 33–40.
Hollingsworth, M. L., and Bailey, J. P. (2000). Evidence for massive clonal growth in the invasive weed Fallopia japonica (Japanese Knotweed). Bot. J. Linn. Soc. 133, 463–472. doi: 10.1111/j.1095-8339.2000.tb01589.x
Hothorn, T., Bretz, F., and Westfall, P. (2008). Simultaneous inference in general parametric models. Biom. J. 50, 346–363. doi: 10.1002/bimj.200810425
Knoche, H. (1922). Flora Balearica. Étude phytogéographique sur les iles Baléares. 2. Montpellier: Roumégous & Déhen.
Krejčíková, J., Sudová, R., Oberlander, K. C., Dreyer, L. L., and Suda, J. (2013). Cytogeography of Oxalis pes-caprae in its native range: where are the pentaploids? Biol. invasions 15, 1189–1194. doi: 10.1007/s10530-012-0370-2
Lane, D. (1984). Factors affecting the development of populations of Oxalis pes-caprae L. Weed Res. 24, 219–225. doi: 10.1111/j.1365-3180.1984.tb00591.x
Lavergne, S., and Molofsky, J. (2007). Increased genetic variation and evolutionary potential drive the success of an invasive grass. Proc. Natl. Acad. Sci. U.S.A. 104, 3883–3888. doi: 10.1073/pnas.0607324104
Levin, D. A. (2002). The Role of Chromosomal Change in Plant Evolution. Oxford: Oxford University Press.
Liu, F., Chen, J. M., and Wang, Q. F. (2009). Trade-offs between sexual and asexual reproduction in a monoecious species Sagittaria pygmaea (Alismataceae): the effect of different nutrient levels. Plant Syst. Evol. 277, 61–65. doi: 10.1007/s00606-008-0103-2
Lui, K., Thompson, F. L., and Eckert, C. G. (2005). Causes and consequences of extreme variation in reproductive strategy and vegetative growth among invasive populations of a clonal aquatic plant, Butomus umbellatus L. (Butomaceae). Biol. Invasions 7, 427–444. doi: 10.1007/s10530-004-4063-3
Marco, A., Lavergne, S., Dutoit, T., and Bertaudiere-Montes, V. (2010). From the backyard to the backcountry: how ecological and biological traits explain the escape of garden plants into Mediterranean old fields. Biol. Invasions 12, 761–779. doi: 10.1007/s10530-009-9479-3
Michael, P. W. (1964). The identity and origin of varieties of Oxalis pes-caprae L. naturalizad in Australia. Trans. R. Soc. S. Aust. 88, 167–173.
Moravcová, L., Pyšek, P., Jarošík, V., and Pergl, J. (2015). Getting the right traits: reproductive and dispersal characteristics predict the invasiveness of herbaceous plant species. PLoS ONE 10:e0123634. doi: 10.1371/journal.pone.0123634
Mulligan, G. A., and Findlay, J. N. (1970). Reproductive systems and colonization in Canadian weeds. Can. J. Bot. 48, 859–860. doi: 10.1139/b70-119
Ness, R. W., Wright, S. I., and Barrett, S. C. H. (2010). Mating-system variation, demographic history and patterns of nucleotide diversity in the tristylous plant Eichhornia paniculata. Genetics 184, 381–392. doi: 10.1534/genetics.109.110130
Novak, S. J., and Mack, R. N. (2005). “Genetic bottlenecks in alien plant species: influences of mating systems and introduction dynamics,” in Species Invasions: Insights into Ecology, Evolution, and Biogeography, eds D. F. Sax, J. J. Stachowicz, and S. D. Gaines (Sunderland, MA: Sinauer Associates), 201–228.
Oduor, A. M. (2013). Evolutionary responses of native plant species to invasive plants: a review. New Phytol. 200, 986–992. doi: 10.1111/nph.12429
Ornduff, R. (1987). Reproductive systems and chromossome races of Oxalis pes-caprae L. and their bearing on the genesis of a noxious weed. Ann. Mo. Bot. Gard. 74, 79–84. doi: 10.2307/2399264
Pannell, J. R., Auld, J. R., Brandvain, Y., Burd, M., Busch, J. W., Cheptou, P. O., et al. (2015). The scope of Baker’s law. New Phytol. 208, 656–667. doi: 10.1111/nph.13539
Petanidou, T., Godfree, R. C., Song, D. S., Kantsa, A., Dupont, Y. L., and Waser, N. M. (2012). Self-compatibility and plant invasiveness: comparing species in native and invasive ranges. Perspect. Plant Ecol. Evol. Syst. 14, 3–12. doi: 10.1016/j.ppees.2011.08.003
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., and R Core Team. (2015). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-122. Available at: http://CRAN.R-project.org/package=nlme
Prentis, P. J., Wilson, J. R. U., Dormontt, E. E., Richardson, D. M., and Lowe, A. J. (2008). Adaptive evolution in invasive species. Trends Plant Sci. 13, 288–294. doi: 10.1016/j.tplants.2008.03.004
Pütz, N. (1994). Vegetative spreading of Oxalis pes-caprae (Oxalidaceae). Plant Syst. Evol. 191, 57–67. doi: 10.1007/BF00985342
Pyšek, P., Jarošík, V., Hulme, P. E., Pergl, J., Hejda, M., Schaffner, U., et al. (2012). A global assessment of invasive plant impacts on resident species, communities and ecosystems: the interaction of impact measures, invading species’ traits and environment. Global Change Biol. 18, 1725–1737. doi: 10.1111/j.1365-2486.2011.02636.x
Pyšek, P., and Richardson, D. M. (2007). “Traits associated with invasiveness in alien plants: where do we stand?,” in Biological Invasions, Ecological Studies, ed. W. Nentwig (Berlin: Springer-Verlag), 99–126.
R Core Team (2014). R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing.
Rambuda, T. D., and Johnson, S. D. (2004). Breeding systems of invasive alien plants in South Africa: does Baker’s rule apply? Divers. Distrib. 10, 409–416. doi: 10.1111/j.1366-9516.2004.00100.x
Rollins, L. A., Moles, A. T., Lam, S., Buitenwerf, R., Buswell, J. M., Brandenburger, C. R., et al. (2013). High genetic diversity is not essential for successful introduction. Ecol. Evol. 3, 4501–4517. doi: 10.1002/ece3.824
Ronsheim, M. L., and Bever, J. D. (2000). Genetic variation and evolutionary trade-offs for sexual and asexual reproductive modes in Allium vineale (Liliaceae). Am. J. Bot. 87, 1769–1777. doi: 10.2307/2656827
Ross, L. C., Lambdon, P. W., and Hulme, P. E. (2008). Disentangling the roles of climate, propagule pressure and land use on the current and potential elevational distribution of the invasive weed Oxalis pes-caprae L. on Crete. Perspect. Plant Ecol. Evol. Syst. 10, 251–258. doi: 10.1016/j.ppees.2008.06.001
Sadras, V. O. (2007). Evolutionary aspects of the trade-off between seed size and number in crops. Field Crops Res. 100, 125–138. doi: 10.1016/j.fcr.2006.07.004
Sakai, A. K., Allendorf, F. W., Holt, J. S., Lodge, M., Molofsky, J., With, K. A., et al. (2001). The population biology of invasive species. Annu. Rev. Ecol. Syst. 32, 305–332. doi: 10.1146/annurev.ecolsys.32.081501.114037
Sala, A., Verdaguer, D., and Vilà, M. (2007). Sensitivity of the invasive geophyte Oxalis pes-caprae to nutrient availability and competition. Ann. Bot. 99, 637–645. doi: 10.1093/aob/mcl289
Sánchez-Pedraja, O. (2015). “Oxalis L,” in Flora Iberica, Vol. IX, eds S. Castroviejo, C. Aedo, M. Laínz, F. Muñoz Garmendia, G. Nieto Feliner, J. Paiva, et al. (Madrid: Real Jardín Botánico, CSIC).
Savinykh, N. P. (2003). Biomorphology of Veronicas of Russia and neighbouring states. Wulfenia 10, 73–102.
Signorini, M. A., Della Giovampaola, E., Ongaro, L., Vivona, L., Bruschi, P., and Foggi, B. (2011). Introduction and spread of the exotic invasive species Oxalis pes-caprae L. in Italy and the Mediterranean area of Europe. An attempt at historical reconstruction. Boll. Mus. Ist. Biol. Univ. Genova 73:138.
Signorini, M. A., Giovampaola, E. D., Bruschi, P., Foggi, B., and Tani, C. (2013). Karyological investigations on the South African invasive Oxalis pes-caprae L. (Oxalidaceae) in native and invaded areas, with special focus on Italy. Plant Biosyst. 147, 298–305. doi: 10.1080/11263504.2013.772924
Silander, J. A. Jr. (1985). “Microevolution in clonal plants,” in Population Biology and Evolution of Clonal Plants, eds J. B. C. Jackson, L. W. Buss, and R. E. Cook (New Haven, CT: Yale University Press), 107–152.
Silvertown, J. (2008). The evolutionary maintenance of sexual reproduction: evidence from the ecological distribution of asexual reproduction in clonal plants. Int. J. Plant Sci. 169, 157–168. doi: 10.1086/523357
Simberloff, D. (2009). The role of propagule pressure in biological invasions. Ann. Rev. Ecol. Evol. Syst. 40, 81–102. doi: 10.1146/annurev.ecolsys.110308.120304
Smith, C. C., and Fretwell, S. D. (1974). Genetic improvement of field crops. New York, NY: Marcel Dekker Inc.
Somme, L., Mayer, C., Raspe, O., and Jacquemart, A. L. (2014). Influence of spatial distribution and size of clones on the realized outcrossing rate of the marsh cinquefoil (Comarum palustre). Ann. Bot. 113, 477–487. doi: 10.1093/aob/mct280
Souza, L., Bunn, W. A., Simberloff, D., Lawton, R. M., and Sanders, N. J. (2011). Biotic and abiotic influences on native and exotic richness relationship across spatial scales: favourable environments for native species are highly invasible. Funct. Ecol. 25, 1106–1112. doi: 10.1111/j.1365-2435.2011.01857.x
Stebbins, G. L. (1957). Self-fertilization and population variability in the higher plants. Am. Nat. 91, 337–354. doi: 10.1086/281999
Symon, D. E. (1961). The species of Oxalis established in South Australia. Trans. R. Soc. S. Aust. 84, 71–77.
Tavares, D. (2014). Evolution of Invasiveness: The Case Study of the Invasive Oxalis pes-caprae in the Mediterranean Basin. Master thesis, University of Coimbra, Coimbra.
te Beest, M., Le Roux, J. J., Richardson, D. M., Brysting, A. K., Suda, J., Kubešová, M., et al. (2012). The more the better? The role of polyploidy in facilitating plant invasions. Ann. Bot. 109, 19–45. doi: 10.1093/aob/mcr277
Thompson, F. L., and Eckert, C. G. (2004). Trade-offs between sexual and clonal reproduction in an aquatic plant: experimental manipulations versus phenotypic correlations. J. Evol. Biol. 17, 581–592. doi: 10.1111/j.1420-9101.2004.00701.x
Turketti, S. S. (2010). A Study of Tristyly in South African Oxalis. Ph.D. thesis, Stellenbosch University, Stellenbosch.
Vallejo-Marín, M., Dorken, M. E., and Barrett, S. C. H. (2010). The ecological and evolutionary consequences of clonality for plant mating. Ann. Rev. Ecol. Evol. Syst. 41, 193–213. doi: 10.1073/pnas.1501712112
Van Drunen, W. E., and Dorken, M. E. (2012). Trade-offs between clonal and sexual reproduction in Sagittaria latifolia (Alismataceae) scale up to affect the fitness of entire clones. New Phytol. 196, 606–616. doi: 10.1111/j.1469-8137.2012.04260.x
Van Drunen, W. E., van Kleunen, M., and Dorken, M. E. (2015). Consequences of clonality for sexual fitness: clonal expansion enhances fitness under spatially restricted dispersal. Proc. Natl. Acad. Sci. U.S.A. 112, 8929–8936. doi: 10.1073/pnas.1501720112
van Kleunen, M., Fischer, M., and Schmid, B. (2002). Experimental life-history evolution: selection on the allocation to sexual reproduction and its plasticity in a clonal plant. Evolution 56, 2168–2177. doi: 10.1111/j.0014-3820.2002.tb00141.x
van Kleunen, M., Weber, E., and Fischer, M. (2010). A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 13, 235–245. doi: 10.1111/j.1461-0248.2009.01418.x
van Kleunen, M. E., Manning, J. C., Pasqualetto, V., and Johnson, S. D. (2008). Phylogenetically independent associations between autonomous self-fertilization and plant invasiveness. Am. Nat. 171, 195–201. doi: 10.1086/525057
Vasconcelos, M. T., and Moreira, I. (1976). “Weeds of apple and pear orchards,” in Proceedings of the II Simposio Nacional de Herbologia, Vol. 2, Oeiras, 41–44.
Verdaguer, D., Sala, A., and Vilà, M. (2010). Effect of enviromental factors and bulb mass on the invasive geophyte Oxalis pes-caprae development. Acta Oecol. 36, 92–99. doi: 10.1016/j.actao.2009.10.006
Vilà, M., Bartolomeus, I., Gimeno, I., Traveset, A., and Moragues, E. (2006). Demography of the invasive geophyte Oxalis pes-caprae across a Mediterranean island. Ann. Bot. 97, 1055–1062.
Vilà, M., and Gimeno, I. (2006). Potential for higher invasiveness of the alien Oxalis pes-caprae on islands than on the mainland. Plant Ecol. 183, 47–53. doi: 10.1007/s11258-005-9005-3
Ward, M., Johnson, S. D., and Zalucki, M. P. (2012). Modes of reproduction in three invasive milkweeds are consistent with Baker’s Rule. Biol. Invasions 14, 1237–1250. doi: 10.1007/s10530-011-0152-2
Weigel, D., and Meyerowitz, E. M. (1994). The ABCs of floral homeotic genes. Cell 78, 203–209. doi: 10.1016/0092-8674(94)90291-7
Wisz, M. S., Pottier, J., Kissling, W. D., Pellissier, L., Lenoir, J., Damgaard, C. F., et al. (2013). The role of biotic interactions in shaping distributions and realised assemblages of species: implications for species distribution modelling. Biol. Rev. 88, 15–30. doi: 10.1111/j.1469-185X.2012.00235.x
Young, D. (1968). “Oxalis L.,” in Flora europaea, Vol. 2, eds T. Tutin, V. Heywood, N. Burges, D. Moore, D. Valentine, S. Walters, et al. (Cambridge, MA: Cambridge University Press), 192–193.
Keywords: clonality, evolution of reproduction, Mediterranean regions, pentaploid, polyploidy, reproductive strategy, sexual and asexual reproduction, tristyly
Citation: Castro S, Castro M, Ferrero V, Costa J, Tavares D, Navarro L and Loureiro J (2016) Invasion Fosters Change: Independent Evolutionary Shifts in Reproductive Traits after Oxalis pes-caprae L. Introduction. Front. Plant Sci. 7:874. doi: 10.3389/fpls.2016.00874
Received: 13 January 2016; Accepted: 03 June 2016;
Published: 24 June 2016.
Edited by:
Peter Alpert, University of Massachusetts Amherst, USAReviewed by:
Gerhard Karrer, University of Natural Resources and Life Sciences Vienna, AustriaMukhtar Ahmed, Washington State University, USA
Copyright © 2016 Castro, Castro, Ferrero, Costa, Tavares, Navarro and Loureiro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sílvia Castro, c2Nhc3Ryb0Bib3QudWMucHQ=
†These authors have contributed equally to this work.