 Kumud Saini1
Kumud Saini1 Hamada AbdElgawad1,2
Hamada AbdElgawad1,2 Marios N. Markakis1,3
Marios N. Markakis1,3 Sébastjen Schoenaers1
Sébastjen Schoenaers1 Han Asard1
Han Asard1 Els Prinsen1
Els Prinsen1 Gerrit T. S. Beemster1
Gerrit T. S. Beemster1 Kris Vissenberg1,4*
Kris Vissenberg1,4*- 1Integrated Molecular Plant Physiology Research, University of Antwerp, Antwerp, Belgium
- 2Department of Botany, Faculty of Science, Beni-Suef University, Beni Suef, Egypt
- 3Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark
- 4Plant and Biochemistry and Biotechnology Lab, Department of Agriculture, School of Agriculture, Food and Nutrition, Technological Educational Institute of Crete: University of Applied Sciences, Heraklion, Greece
Under normal and stress conditions plant growth require a complex interplay between phytohormones and reactive oxygen species (ROS). However, details of the nature of this crosstalk remain elusive. Here, we demonstrate that PINOID (PID), a serine threonine kinase of the AGC kinase family, perturbs auxin homeostasis, which in turn modulates rosette growth and induces stress responses in Arabidopsis plants. Arabidopsis mutants and transgenic plants with altered PID expression were used to study the effect on auxin levels and stress-related responses. In the leaves of plants with ectopic PID expression an accumulation of auxin, oxidative burst and disruption of hormonal balance was apparent. Furthermore, PID overexpression led to the accumulation of antioxidant metabolites, while pid knockout mutants showed only moderate changes in stress-related metabolites. These physiological changes in the plants overexpressing PID modulated their response toward external drought and osmotic stress treatments when compared to the wild type. Based on the morphological, transcriptome, and metabolite results, we propose that perturbations in the auxin hormone levels caused by PID overexpression, along with other hormones and ROS downstream, cause antioxidant accumulation and modify growth and stress responses in Arabidopsis. Our data provide further proof for a strong correlation between auxin and stress biology.
Introduction
The growth regulator auxin plays a role in many aspects of growth and development in plants (Teale et al., 2006; Benjamins and Scheres, 2008; Enders and Strader, 2015). There is increasing evidence for the involvement of auxin metabolism, transport and signaling in stress responses as well (Shibasaki et al., 2009; Shen et al., 2010; Zhang et al., 2012; Kazan, 2013; Remy et al., 2013). Morphogenesis and stress adaptive responses are closely linked to cellular hormonal homeostasis, including auxin. On one hand, mutants overproducing auxin or with altered auxin distribution show developmental defects such as inhibition of shoot growth, cotyledon and leaf epinasty, a longer hypocotyl and excessive lateral root growth (Boerjan et al., 1995; King et al., 1995; Delarue et al., 1998; Zhao et al., 2001). On the other hand, several stress conditions are also known to modify auxin homeostasis and response. Auxin homeostasis could be perturbed by stress-induced changes in the concentration of phenolics compounds such as quercetin and kaempferol (auxin transport inhibitor affecting cycling of auxin efflux carriers), and changes in apoplastic pH affecting auxin uptake and distribution (Potters et al., 2007, 2009). Cadmium treated, phosphorus-, or sulfur-starved plants showed altered auxin metabolism (Potters et al., 2007). Water deficit conditions upregulated Gretchen Hagen 3 (GH3) genes, suppressed YUC genes and lowered indole-3-acetic acid (IAA) levels in rice (Zhang et al., 2009; Du et al., 2013). Hyper-osmolarity promoted PIN FORMED1 (PIN1) and PIN2 internalization while hypo-osmolarity showed opposite effect in Arabidopsis roots (Zwiewka et al., 2015). Salt stress is also shown to affect free and conjugated IAA levels in the developing xylems in poplar, resulting in negative effects on the expansion of xylem vessels (Junghans et al., 2006). Studies on rice and sorghum suggest that auxin response factors (ARFs) and several other early auxin-responsive genes also function in abiotic stress responses (Jain and Khurana, 2009; Wang et al., 2010). Alternatively, stress also induced changes in H2O2 levels that are co-factors for peroxidases to catalyze the oxidative degradation of IAA (Gazarian et al., 1998; Tognetti et al., 2012). Apoplastic reactive oxygen species (ROS) transiently decreased auxin signaling, modified auxin homeostasis and altered leaf morphology in ozone (O3) treated plants (Blomster et al., 2011).
Interestingly, many studies have reported an increase in abiotic stress tolerance in transgenic plants, and in conditions with altered auxin levels (Shi et al., 2014; Naser and Shani, 2016). For instance, elevated auxin levels in transgenic lines overexpressing YUCCA7 (a gene involved in the auxin biosynthesis pathway) increased resistance to drought stress in Arabidopsis (Lee et al., 2012). Similarly, overexpression of YUCCA6 led to enhanced resistance to water stress in potato (Kim et al., 2013). Furthermore, ectopic expression of H2O2-inducible UGT74E2 led to an increase in free indole-3-butyric acid (IBA, another form of active auxin) and IBA-glucose, and also increased the tolerance to drought stress (Tognetti et al., 2010). Interestingly, Park et al. (2007a), on the other hand, showed that overexpression of the GH3 gene lowered IAA levels, reduced growth and enhanced the resistance to abiotic stresses. These studies clearly demonstrate a link between auxin physiology and response to environmental stress factors. Clearly, the relation between auxin and stress adaptations is complex. The specific role of auxin in the induction of stress responses also remains elusive.
The kinase PINOID (PID) regulates PIN localizations on the cellular membranes and thus regulates polar auxin transport (PAT; Friml et al., 2004; Kleine-Vehn et al., 2009). PID is an early auxin inducible gene and a member of the AGCVIII protein kinase family (Benjamins et al., 2001). AGC kinases are named after three classes of animal proteins involved in a receptor-mediated growth factor signal transduction: protein kinase A (PKA), cyclic GMP-dependent protein kinases (PKG) and protein kinase C (PKC). There are 37 such kinases in Arabidopsis, 23 of which belong to the AGCVIII group (Galván-Ampudia and Offringa, 2007). AGC kinases regulate cell growth, morphogenesis and cell immunity in animals, where mutations and malfunctioning kinases can lead to diseases such as cancer (Pearce et al., 2010). In plants as well, apart from supporting normal development, AGC kinases also participate in stress signaling and in regulation of plant immunity (Devarenne et al., 2006; Hirt et al., 2011; Garcia et al., 2012). For example, the AGC kinase, oxidative signal-inducible 1 (OXI1), is essential for H2O2 -mediated oxidative stress signaling, defense against the oomycete pathogen Hyaloperonospora parasitica, and immunity against Pseudomonas syringae (Rentel et al., 2004; Petersen et al., 2009).
We have performed a detailed growth analysis on two pid knockouts (pid-14 and SALK_009478) and two PID overexpression lines (PIDOE), 35S::PID10 (addressed here after as P10) and 35S::PID21 (here after P21), and discussed their contrasting effect on auxin homeostasis and rosette phenotypes (Saini et al., 2017). PIDOE lines showed severely reduced rosette growth and high auxin levels in the first pair of leaves. RNA sequencing of the leaves of PIDOE lines pointed toward induction of stress-related responses. Therefore, we here investigate the effect of altered PID expression on the perturbation of auxin homeostasis and on the induction of stress responses in whole rosettes in Arabidopsis. We also investigate the effect of such perturbations on the modulation of whole plant responses toward osmotic and drought stress conditions.
Materials and Methods
Plant Material and Growth Conditions of Arabidopsis
Arabidopsis thaliana Col-0 ecotype seeds were grown in half strength Murashige and Skoog (MS) medium including vitamins (Duchefa, Netherlands), at pH 5.8 containing 1% sugar, 0.5 g/L MES buffer and 0.7% agar. Seeds were sterilized briefly with 70% ethanol followed by 6% bleach for 5 to 10 min and finally rinsed in water. Plants were kept at 21°C in 16/8 h light period under a light intensity of 70–90 μmol m-2s-1. The knockout T-DNA insertion line SALK_009478 was obtained from NASC. The pid-14 mutant is SALK_049736 as reported by Haga et al. (2014). The two PIDOE lines, P10 and P21, were developed by Benjamins et al. (2001) by cloning the PID cDNA in sense orientation behind the strong Cauliflower Mosaic Virus 35S promoter (35S::PID) and introducing this construct into Arabidopsis thaliana ecotype Columbia (Col).
Drought Stress Application
Seeds (stratified in cold and dark) were sown in separate pots containing an equal amount of soil (Tref substrate). Pots were well watered initially to assist germination. To avoid heterogeneity, trays with pots were rotated 90° on a daily basis. Watering was subsequently withheld 4–5 days after germination. Three conditions were generated, where soil relative water content (RWC) was maintained by weighing pots daily and adjusted to desired values by controlled watering: control pots with 70% RWC, mild stress pots with 45% RWC and severe stress pots at 40% RWC. The experiment continued until 25 days after stratification (DAS). Rosette pictures were taken for measurements with a Canon EOS 40D digital camera equipped with a Canon EF-S 60mm f/2.8 USM macro lens.
For in vitro osmotic stress, seeds were sown on half strength MS medium containing different concentrations of mannitol (Duchefa, Netherlands) and Sorbitol (Duchefa, Netherlands) and allowed to grow until 25 DAS. Digital rosette pictures were analyzed using ImageJ1.
Hormone Measurements
To measure concentrations of IAA, abscisic acid (ABA), gibberellins, salicylic acid, cytokinins and jasmonic acid, whole rosettes were harvested at various time points between 7 to 22 DAS. The samples for three replicates were obtained from multiple plates in each experiment. Dissected samples were collected, frozen in liquid nitrogen, and ground using a MagNA Lyser (Roche) with 2 mm glass beads. The detailed hormone extraction procedures and measurements can be found in the Supplementary Material and Methods section.
qPCR Analysis
Quantitative PCR using SyBr green was performed at 9 and 22 DAS using the first pair of leaves. RNA was isolated using Purelink Plant RNA reagent (Ambion Life Technologies) and quantified with a nanodrop NZ 1000 (Thermo Scientific). An average of 1 μg of RNA was used for first strand cDNA synthesis using RQ1 RNase-Free DNase treatment and the GoScriptTM Reverse Transcription System (Promega). Actin 8 primers and gene-specific primers spanning the intron region were used at 55°C as annealing temperature (Supplementary Table 1). A SyBr green assay for qPCR was accomplished according to the developer’s protocol using ROX SYBR Mastermix blue dTTP (Takyon) and a Step one plus Thermocycler (Life technologies). This experiment was done in three independent biological and three technical repetitions. The results were analyzed as ΔΔCT comparison with the StepOnePlusTM Real-Time PCR System (Life TechnologiesTM) software with a confidence level set at 95%.
RNA Sequencing
18 RNA samples (three genotypes, three replicates, two time points), originating from the first pair of leaves at 9 DAS and 16 DAS from wild type (WT) and both 35S::PID lines were sequenced using an IlluminaTM platform. Prior to library preparation the RNA quality and integrity was assessed according to IlluminaTM guidelines. Library preparation was done using the TruSeq® Stranded mRNA sample preparation 96-reaction kit (IlluminaTM) following the low sample protocol according to manufacturer’s recommendations. In brief, approximately 2.5 μg of total RNA was diluted and purified using RNA purification beads targeting the poly-A tail of the mRNA and this was subsequently fragmented by means of the enzymes provided in the kit. After the cDNA synthesis adenylation of 3′ ends and ligation of the adaptors were performed. Adaptors were ligated in 12-plex formations, allowing the pooling of 12 samples. Subsequently, the library was quantified using PicoGreen® dye (Life TechnologiesTM) as described in the manufacturer’s protocol. In order to accurately quantify the concentration in nM of the sample, the Kapa SYBR® FAST universal qPCR kit (Kapa BiosystemsTM) for IlluminaTM sequencing was used to quantify the number of the amplifiable molecules in the sample and the Bioanalyzer® machine (Agilent TechnologiesTM) to determine the average fragment size of the sample. These measurements allowed optimizing the flow cell clustering and proceeding with the Run. The sample was 50 bp pair-end sequenced in one lane of an IlluminaTM HiSeq1500 sequencer.
RNA Sequencing Data Analysis
Resulting sequence data was preliminary analyzed by CLC Genomics Workbench v.6 using the Arabidopsis thaliana (Col-0 TAIR10) sequence database2 as reference genome. The RNA-Seq analysis was carried out for sequence reads obtained from the three genotypes. Throughout the analysis with CLC default settings were used. Briefly, after the trimming of the sequences they were mapped against the reference genome with the default settings. The expression values were calculated based on “reads per kilo base of exon model per million mapped reads” (RPKM) values (Mortazavi et al., 2008). The RNA-seq data was grouped accordingly and two group comparisons (unpaired) were performed. The expression values were normalized by scaling to the default setting of 10 million reads. Moderated t statistics for pairwise contrasts were calculated using the Baggerly’s test (Baggerly et al., 2003). Genes with no counts in all three replicates for at least one of the genotype/time combinations were discarded as not detectable above the background. Statistical analysis for the effect of genotype and time was conducted by a two-way ANOVA on the MeV software (Multi Experiment Viewer 4.9.0). The Baggerly’s p values were corrected for multiple testing for each contrast separately by means of false discovery rate (FDR; Benjamini and Hochberg, 1995) for significant genes based on ANOVA. FDR corrected p-value < 0.05 and log2 of fold change > 0.75 was used as a cutoff. All significantly induced or repressed genes with known functions were classified into groups based on gene ontology information obtained from the TAIR Database3 using MapMan (Thimm et al., 2004) and overrepresented functions and gene enrichment studies were carried out in Cytoscape using the BiNGO plugin (Maere et al., 2005).
Metabolite Analyses
Malondialdehyde (MDA) and H2O2 were determined by the thiobarbituric acid–MDA (TBA–MDA) assay (Hodges et al., 1999) and xylenol orange-based FOX1 method (Jiang et al., 1990), respectively, from a 100 mg frozen sample. Antioxidants were extracted by homogenizing 100 mg sample in 1.4 ml 80% ethanol (v/v). Total antioxidant capacity was measured after mixing the tissue extract with ferric reducing/antioxidant power (FRAP) assay reagent (0.3 M acetate buffer, pH 3.6, 0.01 mM TPTZ in 0.04 mM Hal, 0.02 M FeCl3⋅6H2O) at 600 nm using a microplate reader (Synergy Mx, Biotek Instruments Inc., Winooski, VT, United States) (Benzie and Strain, 1996). Trolox was used as standard.
Enzyme Assays
Glycolate oxidase was measured according to Feierabend and Beevers (1972). GO was measured by the formation of a glyoxylate complex with phenylhydrazine (𝜀324 = 17 mM-1 cm-1). NADPH oxidase (oxidative stress-related enzyme) was assayed according to Sarath et al. (2007) measuring NADPH-dependent superoxide generation as the reduction rate of NBT into monoformazan (𝜀530 = 12.8 mM-1 cm-1). Enzymes were measured in extracts obtained from 100 mg of frozen tissue, in 1 mL of extraction buffer: 50 mM MES/KOH (pH 6.0) containing 0.04 M KCl, 2 mM CaCl2 and homogenized with a MagNA Lyser (Roche, Vilvoorde, Belgium). Catalase activity was assayed by observing the H2O2 decomposition rate (𝜀240 = 39.4 M-1cm-1) (Aebi, 1984). SOD activity was determined according to Dhindsa et al. (1982) by measuring the inhibition of NBT reduction at 560 nm. SOD and CAT measurements were scaled down for semi-high throughput measurement using a micro-plate reader (Synergy Mx, Biotek Instruments Inc., Winooski, VT, United States). Glutathione S-transferase (GST) activity was determined according to Habig et al. (1974). Enzyme activity was calculated by measuring the conjugation of GSH with excess 1-chloro-2,4-dinitrobenzene (CDNB) in 50 mM potassium phosphate buffer at 340 nm. The activity of glutaredoxin enzymes was measured according to Holmgren and Aslund (1995). The assay contains tris-HCl buffer (100 mM, pH 8.0), 100 μg/mL bovine serum albumin, 1 mM GSH, 6 μg/mL yeast glutathione reductase, 0.1 M Tris-Hcl, 2 mM EDTA, 0.4 mM NADPH and 0.7 mM of 2-hydroxy-ethyl-disulfide (HED). The peroxiredoxin activity was performed by measuring the decrease in H2O2 concentration in the reaction mixture contained 100 mM K2HPO4 buffer (pH 7.0), 0.3 to 3 mM Prx, 10 mM DTT, and 100 uM H2O2 (Horling et al., 2003). The reaction was stopped with 12,5% trichloroacetic acid (TCA). To quantify the reduction in H2O2 content, 10 mM Fe(NH4)2(SO4)2 and 2.5 mM KSCN were added and the absorbance was measured at 480 nm. Ferredoxin enzyme activity measured in a 50 mM HEPES-KOH (pH 8.0) contains 5 mM MgCl2 0.3 mM NADP+, 3 mM glucose 6-phosphate, 1 unit/mL glucose 6-phosphate dehydrogenase and 1 mM potassium ferricyanide. The decrease in NADP+ was measured at 420 nm.
Superoxide Detection in Leaves
Superoxide radical detection was performed using the nitroblue tetrazolium (NBT) staining method (Ramel et al., 2009). NBT reacts with O2- to form a dark blue insoluble formazan compound. Arabidopsis plants were infiltrated in 0.1% (w/v) NBT solution in PBS buffer (0.2 g KCl + 0.2 g KH2PO4 + 1.15 g NaH2PO4 + 8 g NaCl in 1000 ml of H2O; pH 7.1) containing 10 mM of sodium azide (NaN3), for 30 min under vacuum. After 4 h of incubation they were cleared and pictured under a Nikon AZ-100 macroscope equipped with a Nikon DS-Ri1 digital camera.
Statistical Analysis
Rosette and auxin measurements were analyzed by t-test using the R statistical package4. Conditions of normality of distribution and homogeneity of variance were checked and met. Metabolite and enzyme data were analyzed using SPSS (SPSS Inc, Chicago, IL, United States). Conditions of normality of distribution and homogeneity of variance were not examined due to the small sample size. One-way analyses of variance (ANOVA) were performed on the original data to evaluate the differences between genotypes and time (days). Significant differences between means were determined with the Duncan test (P < 0.05).
Accession Number
The Arabidopsis Genome Initiative locus identifier for the PID gene is AT2G34650. Raw RNA sequencing data were deposited in NCBI’s Gene Expression Omnibus as GEO accession no. GSE82086.
Results
Overexpression of PID Leads to Smaller Rosettes and Elevated Auxin Pools
In an attempt to deepen our knowledge on auxin-regulated leaf growth and development, we chose to analyze two PIDOE lines, P10 and P21, which clearly had smaller rosettes compared to WT at 22 DAS (days after stratification; Figure 1A and Saini et al., 2017). To quantify the PID transcript levels between WT and PIDOE lines and to understand the differences in the phenotype between P10 and P21, we performed qPCR. Relative quantification of PID transcript levels revealed that P21 had a two-fold higher PID overexpression level than P10 at 9 DAS, whereas at 22 DAS this was inversed and PID overexpression in P21 was four-fold lower compared to P10 (Figure 1B). The severity of the phenotype and PID transcript abundance differences between P10 and P21 suggests that PID affects rosette growth in a dose-dependent manner (Figures 1A,B). Since PID affects auxin transport and thus its distribution (Christensen et al., 2000; Raftopoulou, 2004; Kleine-Vehn et al., 2009), we measured the total pool of IAA (free IAA and IAA-conjugates). IAA levels were strongly elevated in the rosettes of both PIDOE lines compared to the WT from 7 to 22 DAS (Figure 1C). Significantly higher IAA levels in P10 and P21 from 12 DAS onward reflect the difference in expression levels and that a certain threshold of PID abundance is required to sustain these high IAA levels (Figures 1B,C).
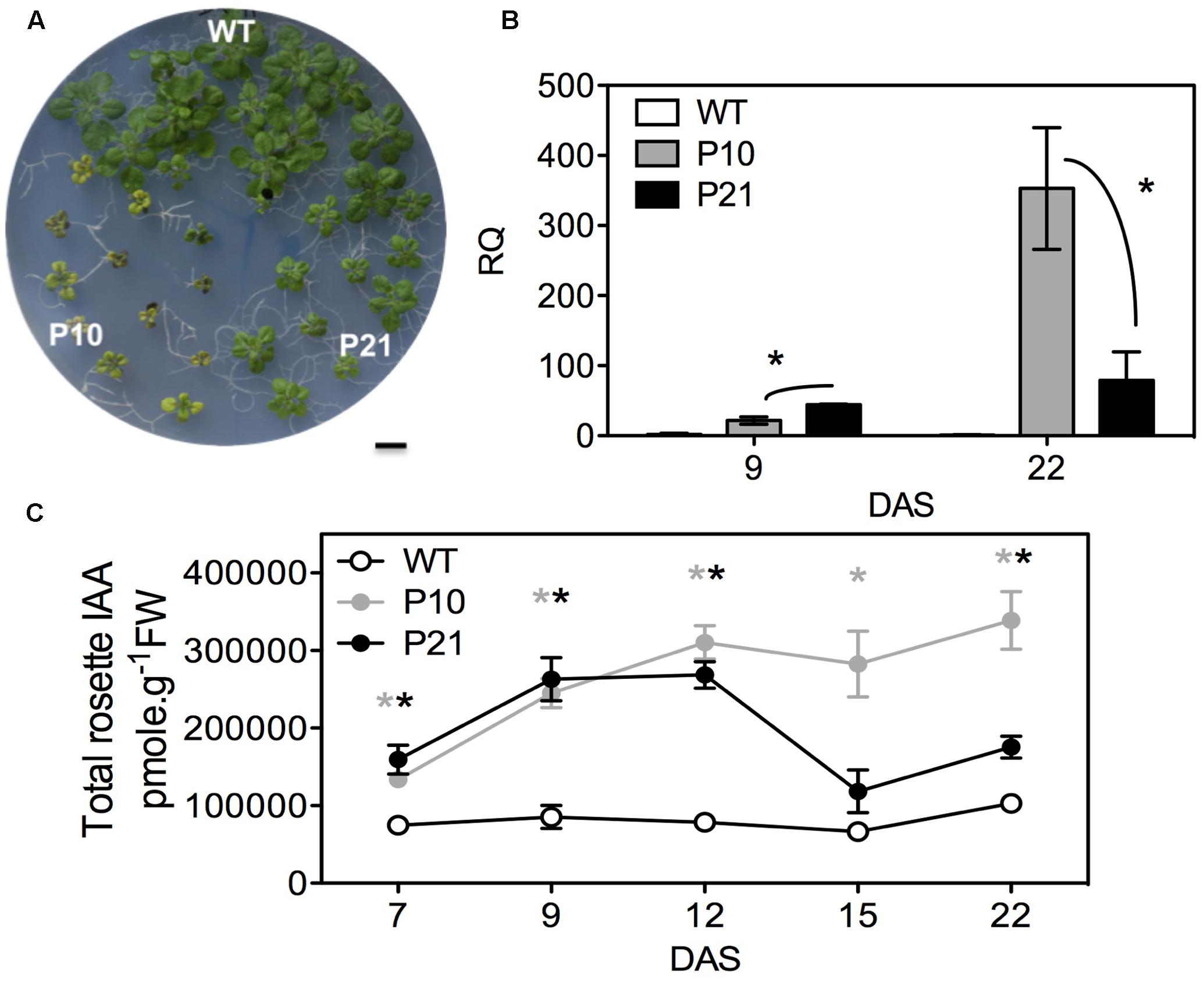
FIGURE 1. Effect of PID overexpression on rosette phenotype and auxin pool in the whole rosettes. Rosette phenotypes of WT and two PIDOE lines, P10 and P21, at 22 DAS (A). Relative quantification (RQ) of PID transcript in the leaves of WT, P10, and P21 at two different time points, n = 3; Asterisks mark the significant differences between the PID overexpression (PIDOE) lines (B). Total auxin (indole-3-acetic acid; IAA) measurements in the rosettes (C). The averages and standard error of relative expression are based on three biological replicates. Gray and black asterisks represent significant differences toward WT, for P10 and P21, respectively. DAS, days after stratification. Error bars: ±SE (t-test P < 0.05). Scale bar = 10 mm.
Transcriptome Data Point to the Induction of Stress in PID Overexpression Lines
To gain further insight into PID function, we checked the expression levels of the AT2G34650 gene (PID) under various conditions in Arabidopsis using perturbation as condition search tool in Genevestigator. Publicly available microarray data revealed that PID is differentially expressed in response to several abiotic stress conditions (Figure 2A). We performed RNA sequencing on the first pair of leaves of PIDOE lines and WT at 9 and 16 DAS to understand the small-rosette phenotype (Supplementary Data 1). In total there were 3805 genes that expressed differentially in at least one genotype and one time point (FDR < 0.05 and log2 fold change > 0.75; Supplementary Data 2). Consistent with the link to abiotic stress suggested by Genevestigator, gene enrichment analysis of the 3805 differentially expressed genes (DEGs), using BiNGO (Maere et al., 2005), showed a significant overrepresentation of ABA signaling, abiotic and biotic stress-related genes, stimulus to chemical and stress, including response to water, water deprivation and osmotic stress (Figure 2B and Supplementary Data 3). Moreover, a MapMan (Thimm et al., 2004) plot of the DEGs also showed that many stress and redox-related genes were among induced or repressed genes in P10 and P21 (Figure 2C). The MapMan analysis also revealed that similar categories were changed in P10 and P21, but at a more pronounced level (more genes and higher fold changes) in P10. This is consistent with the PID expression level of P10 being higher than in P21 at 16 DAS when the transcriptome analysis was performed (Figure 1B).
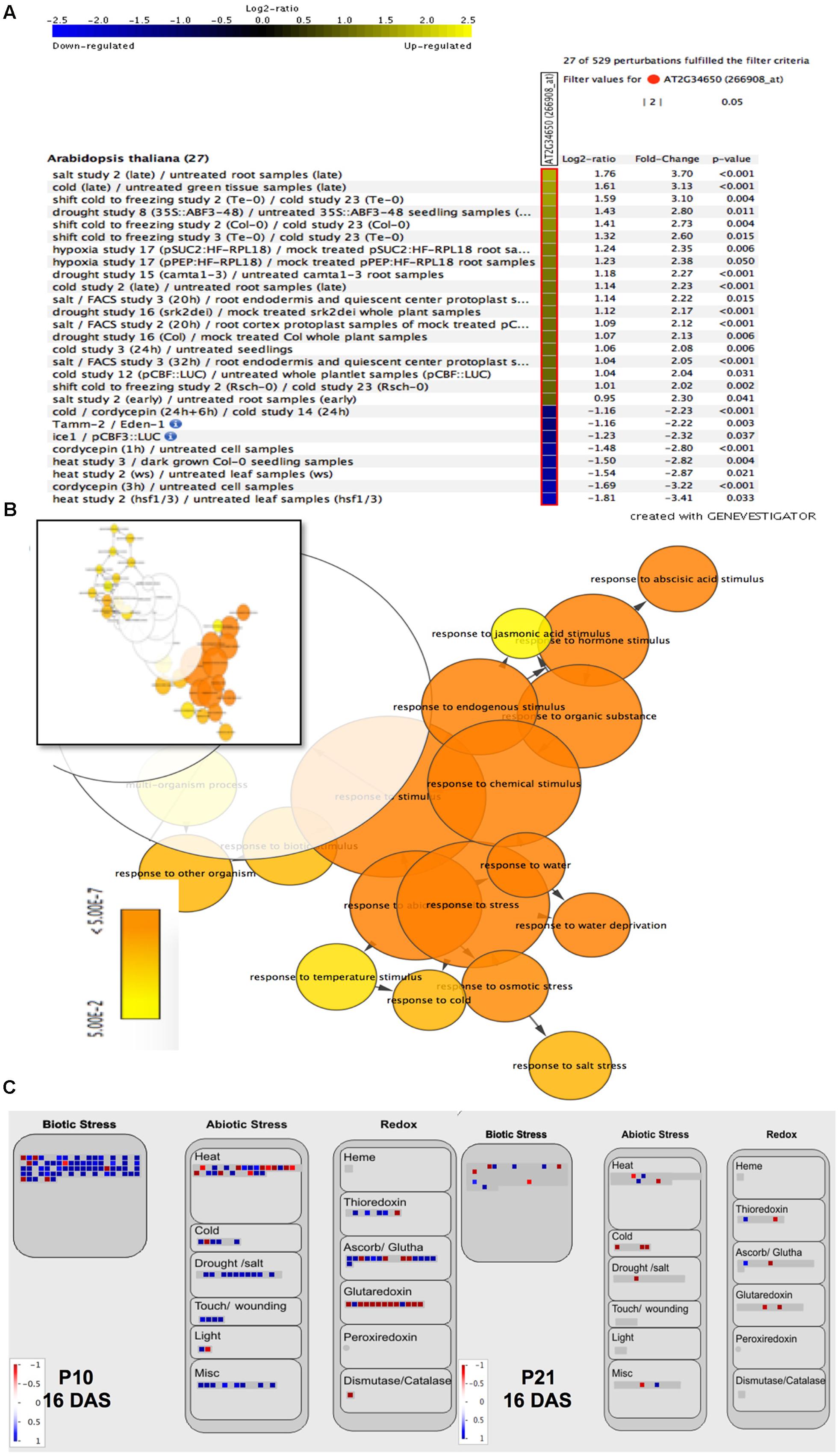
FIGURE 2. Effect of abiotic stress on PID expression levels and of PID overexpression on stress-related genes. Genevestigator analysis shows up- (yellow) and downregulation (blue) of PID under various abiotic stress conditions (A). Gene enrichment analysis of differentially expressed genes in PID overexpression lines vs. WT, visualized via the Cytoscape plugin BiNGO. The inset shows a representation of all 3805 genes, while the major overrepresented genes (orange part of the inset) are magnified (B). MapMan analysis of up- (blue squares) and downregulated (red squares) genes in P10 and P21 at 16 DAS compared to WT, shows the upregulation of biotic and abiotic stress-related genes and changes in redox status (C).
Multiple Hormonal Responses Are Modulated in PID Overexpression Plants
Physiological and molecular mechanisms underlying plant growth and stress responses are complex and likely involve feedback mechanism and crosstalk with other hormones. Consistently, several auxin metabolism, transport and signaling-related genes were affected by PID overexpression that point at a perturbed auxin homeostasis and signaling. These include GH3, IAA-leucine conjugate hydrolase (ILR1), indole-3-butyric acid response (IBR 1 and 2), PINs, auxin/indole-3-acetic acid (Aux/IAAs), small auxin up RNAs (SAURs), Lateral Organ Boundaries-Domain (LBDs), and auxin responsive factors (ARFs) (Supplementary Data 2). Auxin is known to modulate the metabolism of other hormones (Nemhauser et al., 2006). In our data, besides auxin, other hormone signaling components were also modulated in the overexpression lines (Supplementary Figure 1). The expression level of signature genes of several hormones (Nemhauser et al., 2006) were significantly affected in at least one of the PIDOE lines on one time point: ABA-induced carotenoid dioxygenase (NCED3) and abscisic acid responsive elements-binding factor (AREB1/ABF2, AREB2/ABF4, and ABF3), gibberellic acid (GA) inducible, RGL1, and jasmonic acid induced, lipoxygenases LOX1 and LOX2 were all upregulated, whereas cytokinin-induced response regulators, ARR4 and UGT76C2, were downregulated (see Supplementary Table 2 for full list). Thus, these data suggest that the accumulation of IAA in the PIDOE lines leads to changes in signaling components of other hormones as well. Consistently, there was an overrepresentation of stress-related genes that are reported to function in ABA-dependent stress responses. This includes ABA insensitive (ABI1 and ABI2), AREB1/ABF2 and AREB2/ABF4 and ABF3 (Murata et al., 2001; Abe et al., 2003; Fujita et al., 2005; Yoshida et al., 2010). Multiple genes involved in stress regulation that are auxin inducible, such as RAB18, RD22, RD29A, RD29B (Shi et al., 2014; Supplementary Table 2), were also differentially expressed in our data, suggesting upregulation of hormone-related stress responses in PIDOE lines.
To study whether indeed the levels of other hormones were affected in the PIDOE lines, we quantified ABA, GA, cytokinins, salicylic acid, and jasmonic acid in the rosettes (Figure 3). Both PIDOE lines showed elevated ABA levels at 9 and 16 DAS compared to the WT, while P10 also maintained significantly high ABA levels at 22 DAS (Figure 3A). Thus ABA concentration varied in parallel with PID expression levels and the auxin accumulation. PIDOE lines had slightly increased GA4 levels at 16 DAS, while there was a clear increase in cytokinins levels at 16 and 22 DAS. Salicylic acid and GA19 levels were not significantly changed between the genotypes (Figures 3B–E), whereas other GAs were either absent or below the detection limit. Also jasmonic acid concentrations were below the detection limit (1.41 pmol/g).
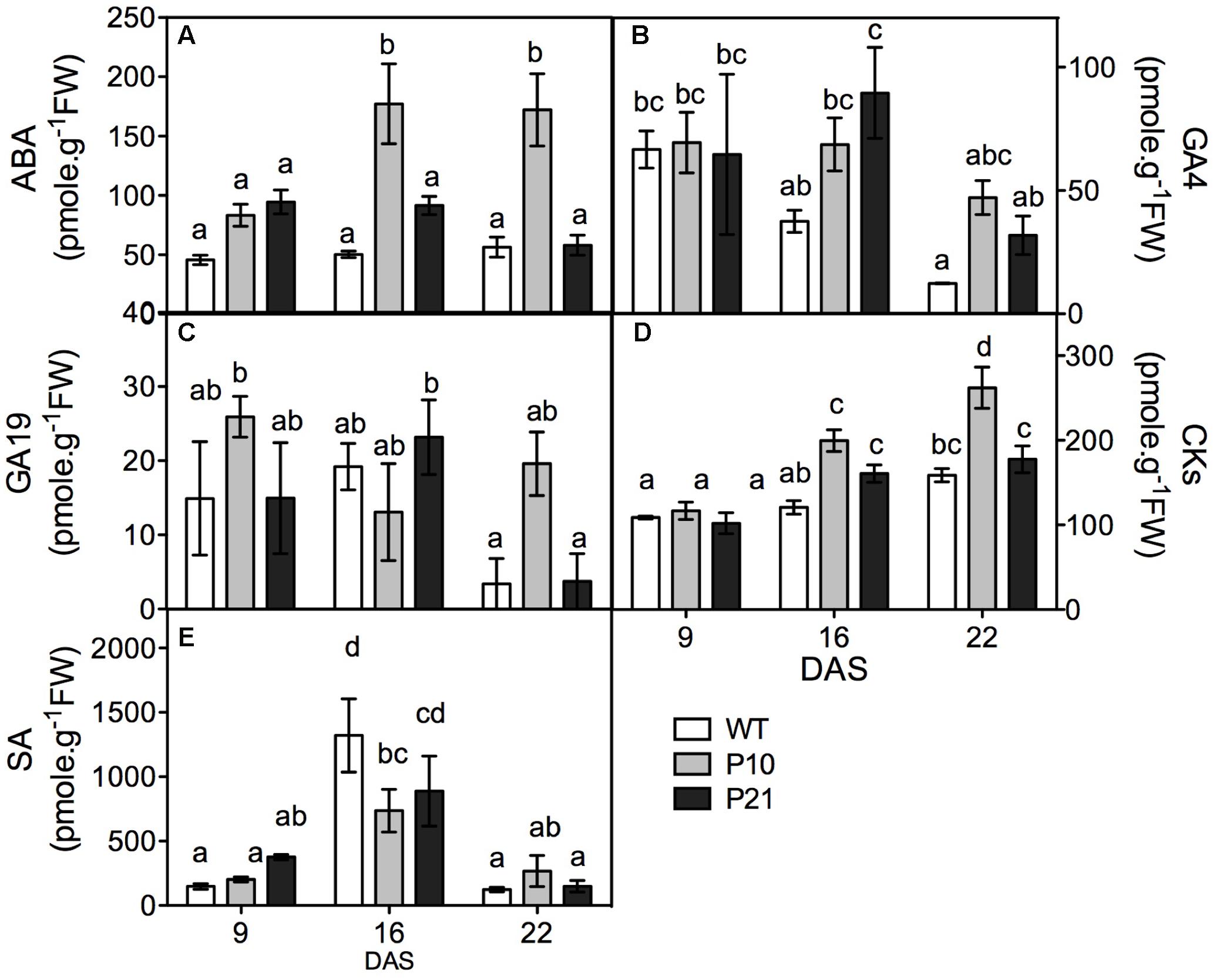
FIGURE 3. Hormone quantification in the rosettes of WT and PID overexpression lines at different time points. Abscisic acid (ABA; A). Gibberellic acid (GA4 and GA19; B,C). Cytokinins (CKs; D) and salicylic acid (SA; E). Different letters in the graph represent significant differences between genotypes and time. (Duncan test; P < 0.05; n = 3; Error bars: ±SE).
PID Overexpression Lines Show Changes in the Activity of Stress Markers
To verify the transcriptome data and study the relation between stress and PID, we tested the effect of altered PID expression on cellular stress markers using both pid knockouts and overexpression mutants. Being a common response to most adverse abiotic conditions, we looked at the parameters related to oxidative stress (Polle and Rennenberg, 1993). Photorespiration, NADPH oxidase, apoplastic peroxidases, acyl-CoA oxidase, and mitochondrial electron transport are among important ROS sources and some have also been involved in responses related to IAA (Elstner, 1982; Wingler et al., 2000; Gill and Tuteja, 2010; Sandalio et al., 2016). We measured the parameters that were also differentially expressed in our transcriptome data (Supplementary Table 3). Our global transcript data suggest an upregulation of photorespiration (Supplementary Table 3), and both PIDOE lines showed elevated GO activities at 16 DAS and P10 at 22 DAS (Figure 4A). The elevation in GO activity was closely linked to PID expression levels (Figure 1B). Compared to the WT, NADPH oxidase activity was increased in both the PIDOE lines at 16 and 22 DAS (Figure 4B). Unlike in the PIDOE lines, the activities of both enzymes were largely unchanged in pid mutants relative to the WT. Oxidative damage to the cell membranes can be measured in terms of MDA levels. Our results show increased MDA levels in the overexpression lines, but these were unchanged in pid mutants (Figure 4C), clearly suggesting lipid peroxidation in the membranes of PIDOE lines, unlike in the membranes of the WT and pid knockouts. H2O2, quantified using the xylenol orange-based FOX1 method (Jiang et al., 1990), showed a gradual increase from 16 to 22 DAS to six-fold higher levels in P10 at 22 DAS, while the mutants and P21 showed no differences compared to the WT (Figure 4D). Rosettes and leaves from all the three genotypes were stained for superoxides and both P10 and P21 showed high staining based on the reaction between NBT and O2-, leading to the formation of the blue colored formazan compound. The staining was most prominent at 9 DAS and particularly enriched in the top-half of the leaves of PIDOE lines, while it was spread across the leaf blade in the WT suggesting a relation between auxin accumulation (i.e., top leaf blade in PIDOE lines) and superoxide radicals (Saini et al., 2017; Supplementary Figure 2).
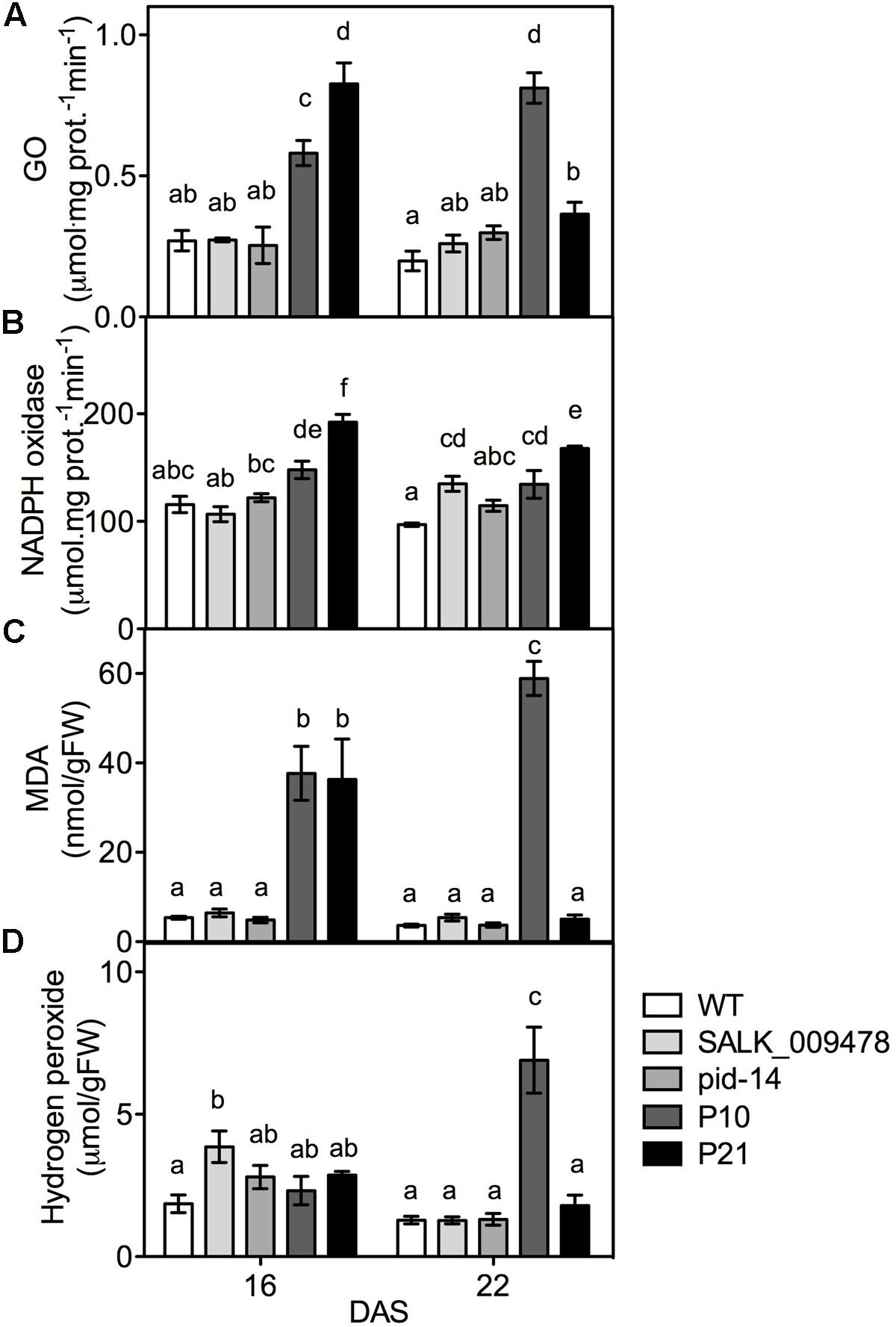
FIGURE 4. Quantification of oxidative stress indicators in rosettes of WT and lines with altered PID expression. Glycolate oxidase; GO activity (A), NADPH oxidase activity (B), malondialdehyde; MDA (C), and hydrogen peroxide abundance (D) in the rosettes of WT, pid knockouts and PIDOE lines at 16 and 22 DAS. Different letters in the graphs (a–f) represent significant differences between genotypes and time (Duncan test; P < 0.05; n = 3; Error bars: ±SE).
PID Overexpression Lines Have High Cellular Antioxidant Levels
Given the dual role of ROS as signaling molecules and molecules causing cellular damage, plants strictly regulate ROS levels in various cellular compartments by means of various antioxidant systems (Noctor and Foyer, 1998; Arora et al., 2002; Neill et al., 2002; Foreman et al., 2003; Foyer and Noctor, 2005). Therefore, we determined the concentrations of some chemical antioxidants and enzymatic antioxidants in pid mutants, PIDOE lines and the WT. The transcripts related to phenylpropanoid and flavonoid biosynthesis pathways showed higher abundance in PIDOE lines compared to the WT in the transcriptome data (Supplementary Figure 3 and Data 2). Since the results could indicate higher concentrations of the end products, we measured the concentrations of flavonoids and anthocyanins along with the total antioxidant capacity in all the genotypes. Indeed, the total antioxidant capacity, reflecting overall changes in the radical scavenging capacity, was higher in P21 lines at 16 DAS and in both lines at 22 DAS compared to the WT (Figure 5A). In addition to the enhanced total antioxidant capacity, also the levels of one or several groups of antioxidant molecules increased gradually (over time, from 16 to 22 DAS) and considerably in at least one genotype and time point in PIDOE lines. This is particularly clear for flavonoids, anthocyanins, and polyphenols, more so in P10 than P21 (Figures 5B–D), which is again in congruence with comparatively higher auxin levels, PID transcript and ROS accumulation in P10 (Figures 1, 4D). Contrary to PIDOE lines, antioxidants levels were unchanged in pid knockout mutants compared to the WT, with exception of increased flavonoids in pid-14 at 16 DAS. At the level of enzymatic antioxidants, with the exception of catalase (CAT), a large number of enzymes showed moderate changes in pid knockouts and overexpression lines [superoxide dismutase (SOD), peroxidases (POX), glutaredoxins, ferredoxins, and peroxiredoxins), while levels of GSTs, enzymes involved in detoxification of xenobiotic compounds (Marrs, 1996), were unchanged in all the genotypes (Supplementary Figure 4). These results demonstrate an altered antioxidant status as a consequence of altered PID expression.
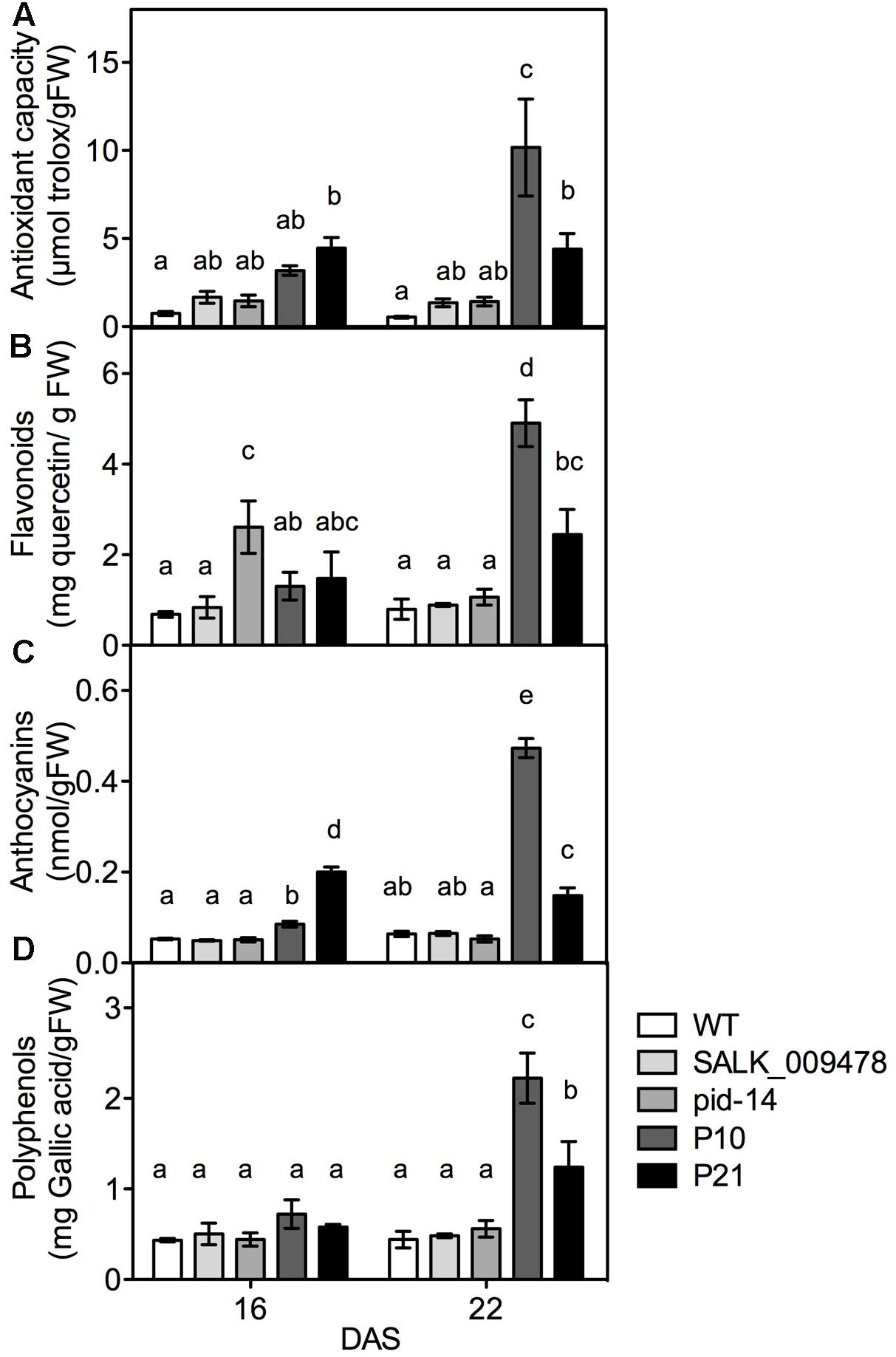
FIGURE 5. Quantification of molecular antioxidants and related enzymes in rosettes of WT and PID overexpression lines. Total antioxidant capacity; TAC (A), flavonoids (B), anthocyanins (C), and polyphenols (D) in WT and PIDOE lines at 16 and 22 DAS. Different letters in the graph represent significant differences between the genotypes and time (Duncan test; P < 0.05; n = 3; Error bars: ±SE).
PID Overexpression Lines Show Modulated Response to External Stresses
Many reports showed that modulation of auxin and antioxidant levels confer resistance or tolerance to drought or osmotic stress (Tognetti et al., 2010; Lee et al., 2012; Espinoza et al., 2013; Kim et al., 2013; Cha et al., 2014; Nakabayashi et al., 2014; Shi et al., 2014; Avramova et al., 2015, 2017; Islam et al., 2015). To investigate if the PID-mediated auxin responses in stress regulation affected whole plant responses to abiotic stresses, we subjected PIDOE lines to osmotic and drought stress. Mannitol and sorbitol were used to lower the water potential in the media (Verslues et al., 2006; Claeys et al., 2014). Rosette area measurements were made at 25 DAS when the first pair of leaves have reached maturity (Beemster et al., 2005). Increasing concentrations of sorbitol or mannitol reduced rosette growth in WT (Figures 6A–D). All the lines showed growth reduction although both PIDOE lines, especially P10, showed less impact of the water deficit.
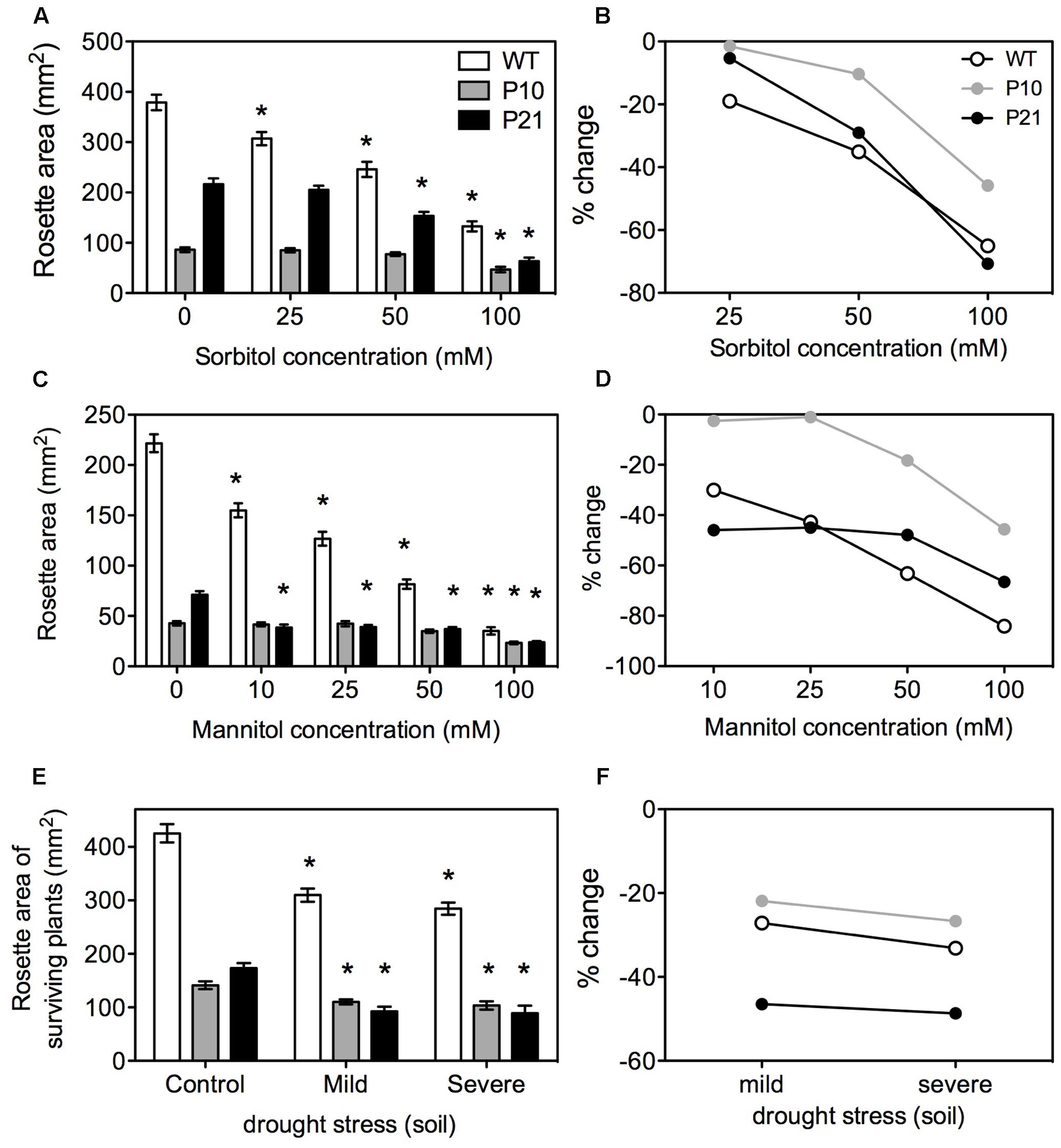
FIGURE 6. Effect of osmotic and drought stress on rosettes of WT and PID overexpression lines. Rosette area under increasing concentrations of sorbitol at 25 DAS; Error bars: ±SE (A) and corresponding percent changes; Error bars represent error of propagation (B). Rosette area under increasing concentrations of mannitol at 25 DAS; Error bars: ±SE (C) and corresponding percent changes; Error bars represent error of propagation (D). Rosette area measurements under soil drought stress at 25 DAS (Error bars: ±SE) with percent changes (E,F). Asterisks represent statistical significant differences with the respective controls (n = 40–50; P < 0.05).
To further confirm the in vitro responses to low water potentials, plants were subjected to different water availabilities in soil by altered watering regimes. WT, P10, and P21 showed a growth reduction under mild (45% RWC) and severe (40% RWC) drought stress. Growth reductions in P10 were similar to WT and stronger in P21 (Figures 6E,F). After 4 weeks, WT plants had a 100% survival rate (i.e., number of plants surviving at 25 DAS), while the survival in P10 and P21 was reduced (90 and 80% in mild stress; 55 and 35% in severe stress, respectively).
Discussion
PID Overexpression Perturbs the Homeostasis of Auxin and Other Hormones, Inducing Stress Responses in Arabidopsis Rosettes
Previously, we elaborated on the fact that PID overexpression causes reduced growth in Arabidopsis leaves due to elevated auxin levels. On the other hand, pid knockouts displayed similar rosette growth and total auxin levels as the WT (Bennett et al., 1995; Christensen et al., 2000; Saini et al., 2017). In order to get a better understanding of the molecular changes induced by the altered PID expression levels we performed RNA sequencing on WT and PIDOE lines. Our transcriptome data suggested differential expression of genes related to auxin metabolism and signaling in PIDOE lines (Supplementary Figure 1 and Data 2). Since PID is a regulator of PAT (Benjamins et al., 2001) and controls the subcellular localization of PIN proteins (Friml et al., 2004), the most likely explanation for the increased auxin levels is due to defective transport from the young leaves, which are generally considered as a site of auxin production (Ljung et al., 2001; Saini et al., 2017).
Several studies using genetic mutants with modified auxin metabolism or signaling have demonstrated a role for auxin in abiotic stress. Such studies have, so far, mostly assessed the plant’s tolerance toward abiotic stresses. Here, we uniquely show that PID, a regulator of auxin transport, when overexpressed in Arabidopsis perturbs auxin homeostasis, which consequently leads to induction of stress responses in the rosettes (measured in terms of severity of phenotype, ROS, antioxidants accumulation and upregulation of genes known to be involved in stress and redox signaling in plants; Supplementary Data 2, 3). It is evident from public transcriptome data that PID is differentially expressed in stress conditions, although up to four-fold only (Figure 2), suggesting that PID might play a role in stress responses. However, a role for PID as a regulator of PIN polarity and thus, auxin transport, is well established and we believe that it is the auxin accumulation in the leaves that causes the reduction in rosette growth, induction of stress responses and modification of plant response toward drought treatment. Our study does strongly point to a relationship between PID-modulated auxin homeostasis and signaling, and the observed stress responses.
PIDOE plants also showed changes in levels of other hormones and their responsive genes, as shown by PageMan, hormone quantification and expression changes in our transcriptome data (Figure 3, Supplementary Figure 1, and Data 2). The involvement of multiple hormones suggests involvement of complex and interlinked hormonal regulatory pathways in growth and stress responses in Arabidopsis leaves, that is also frequently evidenced across literature (Mouchel et al., 2006; Nemhauser et al., 2006; Chapman and Estelle, 2009; Harrison, 2012; Lanza et al., 2012; Šimášková et al., 2015). Several reports indeed suggest that auxin conjugates could be involved in stress responses (Tognetti et al., 2010; Ludwig-Müller, 2011; Kinoshita et al., 2012). In fact a stress responsive GH3 gene, WES1, is known to modulate the cross talk between auxin-SA and auxin-ABA (Park et al., 2007a,b; Zhang et al., 2007). Upregulation of MYB96, a molecular link in the ABA-auxin crosstalk in stress conditions, along with other genes involved in the MYB96 regulated pathways such as RD-22 (drought stress inducing gene) and GH3 suggests the involvement of auxin-ABA interactive responses in the PIDOE lines (Seo et al., 2009). However, since PID is only known to directly affect auxin, we primarily focus on auxin and speculate that other hormones act downstream of auxin or in concert with auxin to induce stress responses.
Like hormones, ROS are signaling molecules that co-regulate growth and development in plants and coordinate responses to environment cues (Neill et al., 2002; Mittler et al., 2004; Jaspers and Kangasjärvi, 2010). Plants respond to environmental stresses by adopting various developmental modulations that include altered growth and development (collectively known as stress induced morphogenic responses; SIMR), reduced metabolism and increased antioxidant accumulation. In fact, plant growth is influenced by a controlled balance between ROS and hormones where auxin and ROS are seen as the key players in stress adaptive responses (Potters et al., 2007, 2009; Tognetti et al., 2012). Evidences showing frequent crosstalk between ROS signaling and hormonal networks are not uncommon. Interplay between ROS and phytohormones is evidenced in abiotic stress adaptation (Tognetti et al., 2010), induction of plant defense responses (Mühlenbock et al., 2008), programmed cell death (Kuriyama and Fukuda, 2002; Gechev et al., 2004), growth and developmental aspects such as cell cycle and cell elongation (Hirt, 2000; D’Haeze et al., 2003; Teale et al., 2006) and regulation of stomatal aperture (Neill et al., 2002). Auxin induced changes in ROS levels are shown to facilitate root gravitropism (Joo et al., 2001), stomatal opening (Song et al., 2006) and cell elongation (Schopfer, 2001). Several sources of ROS, i.e., NADPH oxidases, photo respiratory enzymes, apoplastic peroxidases, acyl-CoA oxidase, and mitochondrial electron transport, are also involved in IAA regulatory networks (Sandalio et al., 2016). PIDOE lines had elevated ROS levels that can possibly be explained by increased photorespiration, but also by increased NADPH oxidase activity (Peer et al., 2013). Auxin overproducing/accumulating mutants are known to have reduced rosette growth and epinastic leaves (Boerjan et al., 1995; Delarue et al., 1998; Zhao et al., 2001). Leaf epinasty is primarily controlled by auxin and involves ROS-auxin interplay as well (Sandalio et al., 2016). PIDOE lines also show enhanced auxin and ROS and downward leaf curling (Christensen et al., 2000; this study). We believe that the high auxin levels in PIDOE lines could be causing high ROS production, resulting in growth retardation and distinct morphological phenotypes in the leaves (Figure 7).
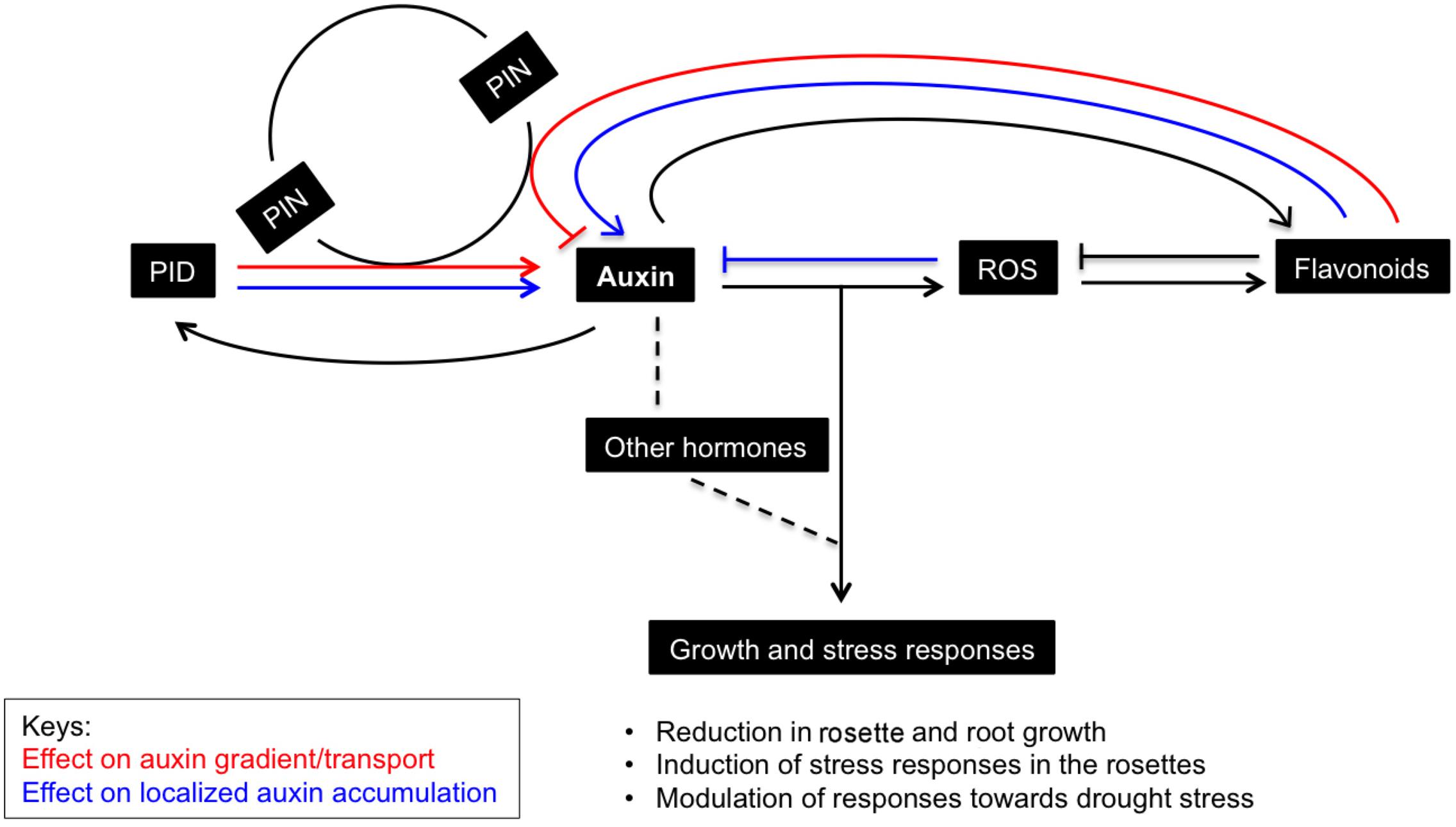
FIGURE 7. Schematic representation of the effects of PID-modulated auxin homeostasis and signaling on rosette growth and stress responses. PID, an early auxin responsive gene and a regular of PIN polarity and auxin transport, causes accumulation of auxin in the rosettes when it is ectopically expressed. The perturbation in auxin homeostasis and signaling affects levels of other interacting hormones and leads to ROS generation, which is known to promote oxidative degradation of auxins. Consequently, flavonoids accumulate to curb ROS and inhibit auxin transport, and supposedly further promote auxin retention in the leaves. This results in growth retardation and increased sensitivity toward drought stress in Arabidopsis plants. Factors affecting the auxin gradient and localized auxin accumulation (here in leaves) are marked in red and blue, respectively.
Apart from growth reduction, plants adapt to abiotic stress by mechanisms such as stomatal closure, accumulation of osmolytes, antioxidants, and the induction of LEA proteins (Verslues et al., 2006). Similarly, in response to induced stress conditions due to PIDOE, various antioxidants appeared to buffer ROS accumulation in plants. The increase in overall antioxidant capacity is probably caused by increase in flavonoids, anthocyanins, and other polyphenols. Consistently, most of the genes related to flavonoids/anthocyanin biosynthesis were also upregulated in the PIDOE lines (Supplementary Figure 3 and Data 2). Flavonoids and anthocyanins are plant secondary metabolites produced via the phenylpropanoid pathway that affect several developmental processes including protection against UV, ROS, etc. (Taylor and Grotewold, 2005; Grotewold, 2006). Flavonoids are known to alter auxin transport and promote localized auxin accumulation in a tissue-specific manner (Peer et al., 2004; Peer and Murphy, 2006, 2007; Kuhn et al., 2011; Buer et al., 2013). Kuhn et al. (2017) showed that flavonol (a subgroup of flavonoids) accumulation in the rol1-2 mutant, which is deficient in rhamnose synthase, affected shoot development and altered export of naphthalene-1-acetic (NAA), but not of IAA. Many reports suggest that flavonoids affect auxin transport directly or indirectly by modifying vesicular trafficking and PIN cycling, other auxin efflux proteins like ATP-binding cassette transporter superfamily (ABCB), or by modifying activities of PAT regulators, or protein phosphatase 2A (PP2A) and its antagonist PID (Peer et al., 2004; Terasaka et al., 2005; Geisler and Murphy, 2006; Kuhn et al., 2017). Clearly flavonoids are modulators of PAT, however, anthocyanins seem to have little or no effect on auxin transport (reviewed in Peer and Murphy, 2007). Interestingly, auxin accumulation is shown to promote flavonoid accumulation, presumably to quench ROS signal generated during auxin catabolism (Peer and Murphy, 2007; Peer et al., 2011). Similarly, enhanced flavonoid synthesis in auxin accumulating PIDOE lines could also help to scavenge elevated ROS molecules. Another interesting assumption is that gradual accumulation of flavonoids in the leaves of PIDOE lines further promotes localized auxin accumulation in the leaves, which is evident in both P10 and P21 (Figures 1C, 5B, 7).
In addition to increases in molecular antioxidants, also the activity of a relatively large range of antioxidant enzymes is, at least moderately, modified. The decreased activity of enzymatic antioxidants such as peroxidases, peroxiredoxins could also be seen as contributory factor in ROS accumulation in PIDOE lines. The comparison of changes at the level of transcripts and enzyme activity is difficult to establish or rather inconclusive as most of redox-related genes were downregulated (Supplementary Table 3). The observation that a relatively large number of antioxidant enzymes and molecules change (mostly increase), suggest that a wide range of cellular redox processes are affected in the PIDOE lines, rather than any specific process. The fact that changes in specific metabolites (anthocyanins) and enzymes (GO) are closely related to PID overexpression levels, indicates a positive correlation between them. We believe that the differences between two overexpression lines itself are because of the differences in PID expression levels and thus auxin levels and the fact that even transient changes in auxin can hugely impact cellular and molecular events in the plant (Nemhauser et al., 2006; Paponov et al., 2008; Saini et al., 2017). These data clearly indicate the presence of enhanced (oxidative) stress responses, in PIDOE lines compared to pid knockouts, as shown by measurements of ROS, ROS producing and detoxifying enzymes, MDA and antioxidant molecules. To summarize, it is well established that environmental stresses impact cellular ROS levels, antioxidant concentration and their redox state (Mittler, 2002; Apel and Hirt, 2004; Hong-bo et al., 2008). Here, we demonstrate that in the absence of external stress, ROS and antioxidants levels change in response to cellular auxin perturbations, and modulate growth and stress adaptive responses.
PID Overexpression Does Not Confer Tolerance to Water Stress
There are many reports providing links between abiotic stress, antioxidant abundance and auxin (Jung and Park, 2011; Hasanuzzaman et al., 2013; Rahman, 2013; Avramova et al., 2015, 2017). For example, activation of YUCCA7 elevates IAA levels and enhances resistance to drought in Arabidopsis (Lee et al., 2012). However, as a result of the overexpression of this gene, growth was reduced and plants had narrow and curled leaves. Similarly, overexpression of OsPIN3t increases water tolerance in rice (Zhang et al., 2012). Phot1 (Wan et al., 2012), a close homolog of PINOID belonging to the same AGC kinase family (Galván-Ampudia and Offringa, 2007), is also shown to improve drought tolerance in Arabidopsis seedlings (Galen et al., 2007). Similarly, overaccumulation of flavonoids is shown to enhance tolerance to drought and osmotic stress (Nakabayashi et al., 2014), and anthocyanins are also suggested to have an osmoprotectant role among others, such as ROS scavenging (Hughes et al., 2010, 2013).
In our results, all three genotypes responded to osmotic stress by reducing their rosette growth (Figure 6). The differences in growth reduction between PIDOE and WT could be due to already smaller size of non-stressed PIDOE rosettes that allowed a lower degree of growth reduction when subjected to osmotic stress conditions. Both PIDOE lines showed lower survival than the WT during drought stress assays in the soil. We believe that the increased IAA levels, due to PID overexpression, already induce stress conditions in the rosettes and that the water deficit treatment is additive, thereby leading to higher lethality. Another explanation could be that PIDOE lines have much shorter roots and reduced numbers of lateral roots (Benjamins et al., 2001), making their quest for water in the soil very difficult, which could have resulted in poor performance under drought stress. Evidently, the shoot/root ratio and stress responses are closely related (Woo et al., 2007; Werner et al., 2010). This suggests that PID-induced auxin alterations induce the observed stress responses and clearly do not confer advantage in water deficit conditions.
To conclude, various reports show that plant AGC kinases participate in the response to biotic and abiotic stresses. We demonstrated that ectopic expression of PID causes changes in auxin levels and its response, and consequentially ROS is generated and other hormones are affected which results in the generation of growth inhibitory stress responses. Additionally, in contrast to several previous reports, our study uniquely shows that despite their high auxin and antioxidant levels, plants are impaired in their tolerance to osmotic and drought stress and show a low survival under severe drought conditions. For future work it will be interesting to know how direct the relation between PID and stress responses is and whether auxin accumulation in leaves is necessary or sufficient to cause the observed stress responses.
Author Contributions
KS planned and performed most of the experiments, analyzed the data and wrote the article with contribution from HAs, EP, GB, and KV; HAb and SS provided assistance with data analysis; MM performed next generation sequencing; EP, GB, and KV conceived the project and KV supervised the research.
Funding
This work was supported by a concerted research activity (GOA) research grant, ‘A Systems Biology Approach of Leaf Morphogenesis’ from the research council of the University of Antwerp. This work was supported by the Research Foundation Flanders (FWO-Vlaanderen; G.0656.13N, 1.5.091.11.N.00).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The 35S::PID10 and 35S::PID21 mutants were kindly provided by Eva Benkova (IST Austria). We sincerely thank Sevgi Öden and Tim Willems (Biology Department, UA) for the hormone measurements.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2017.01308/full#supplementary-material
Footnotes
- ^ http://rsbweb.nih.gov/ij/
- ^ http://www.arabidopsis.org/
- ^ www.arabidopsis.org
- ^ https://www.r-project.org/
References
Aebi, H. (1984). Oxygen radicals in biological systems. Methods Enzymol. 105, 121–126. doi: 10.1016/S0076-6879(84)05016-3
Abe, H., Urao, T., Ito, T., Seki, M., and Shinozaki, K. (2003). Transcriptional activators in abscisic acid signaling. Society 15, 63–78. doi: 10.1105/tpc.006130.salt
Apel, K., and Hirt, H. (2004). Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 55, 373–399. doi: 10.1146/annurev.arplant.55.031903.141701
Arora, A., Sairam, R. K., and Srivastava, G. C. (2002). Oxidative stress and oxidative system in plants. Curr. Sci. 82, 1227–1238.
Avramova, V., AbdElgawad, H., Vasileva, I., Petrova, A. S., Holek, A., Mariën, J., et al. (2017). High antioxidant activity facilitates maintenance of cell division in leaves of drought tolerant maize hybrids. Front. Plant Sci 8:84. doi: 10.3389/fpls.2017.00084
Avramova, V., AbdElgawad, H., Zhang, Z., Fotschki, B., Casadevall, R., Vergauwen, L., et al. (2015). Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone. Plant Physiol. 169, 1382–1396. doi: 10.1104/pp.15.00276
Baggerly, K. A., Deng, L., Morris, J. S., and Aldaz, C. M. (2003). Differential expression in SAGE: accounting for normal between-library variation. Bioinformatics 19, 1477–1483. doi: 10.1093/bioinformatics/btg173
Beemster, G. T. S., De Veylder, L., Vercruysse, S., West, G., Rombaut, D., Van Hummelen, P., et al. (2005). Genome-wide analysis of gene expression profiles associated with cell cycle transitions in growing organs of Arabidopsis. Plant Physiol. 138, 734–743. doi: 10.1104/pp.104.053884
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300. doi: 10.2307/2346101
Benjamins, R., Quint, A., Weijers, D., Hooykaas, P., and Offringa, R. (2001). The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport. Development 128, 4057–4067.
Benjamins, R., and Scheres, B. (2008). Auxin: the looping star in plant development. Annu. Rev. Plant Biol. 59, 443–465. doi: 10.1146/annurev.arplant.58.032806.103805
Bennett, R. M. S., Alvarez, J., Gerd, B., and Smyth, D. R. (1995). Morphogenesis in pinoid mutants of Arabidopsis thaliana. Plant J. 8, 505–520.
Benzie, I. F. F., and Strain, J. J. (1996). The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal. Biochem. 239, 70–76. doi: 10.1006/abio.1996.0292
Blomster, T., Salojärvi, J., Sipari, N., Brosche, M., Ahlfors, R., Keinänen, M., et al. (2011). Apoplastic reactive oxygen species transiently decrease auxin signaling and cause stress-induced morphogenic. Plant Physiol. 157, 1866–1883. doi: 10.1104/pp.111.181883
Boerjan, W., Cervera, M. T., Delarue, M., Beeckman, T., Dewitte, W., Bellini, C., et al. (1995). Superroot, a recessive mutation in Arabidopsis, confers auxin overproduction. Plant Cell 7, 1405–1419. doi: 10.1105/tpc.7.9.1405
Buer, C. S., Kordbacheh, F., Truong, T. T., Hocart, C. H., and Djordjevic, M. A. (2013). Alteration of flavonoid accumulation patterns in transparent testa mutants disturbs auxin transport, gravity responses, and imparts long-term effects on root and shoot architecture. Planta 238, 171–189. doi: 10.1007/s00425-013-1883-3
Cha, J. Y., Kim, J. Y., Jung, I. J., Kim, M. R., Melencion, A., Alam, S. S., et al. (2014). NADPH-dependent thioredoxin reductase A (NTRA) confers elevated tolerance to oxidative stress and drought. Plant Physiol. Biochem. 80, 184–191. doi: 10.1016/j.plaphy.2014.04.008
Chapman, E. J., and Estelle, M. (2009). Cytokinin and auxin intersection in root meristems. Genome Biol. 10:210. doi: 10.1186/gb-2009-10-2-210
Christensen, S. K., Dagenais, N., Chory, J., and Weigel, D. (2000). Regulation of auxin response by the protein kinase PINOID. Cell 100, 469–478.
Claeys, H., Van Landeghem, S., Dubois, M., Maleux, K., and Inzé, D. (2014). What is stress? Dose-response effects in commonly used in vitro stress assays. Plant Physiol. 165, 519–527. doi: 10.1104/pp.113.234641
Delarue, M., Prinsen, E., Van Onckelen, H., Caboche, M., and Bellini, C. (1998). Sur2 mutations of Arabidopsis thaliana define a new locus involved in the control of auxin homeostasis. Plant J. 14, 603–611. doi: 10.1046/j.1365-313X.1998.00163.x
Devarenne, T. P., Ekengren, S. K., Pedley, K. F., and Martin, G. B. (2006). Adi3 is a Pdk1-interacting AGC kinase that negatively regulates plant cell death. EMBO J. 25, 255–265. doi: 10.1038/sj.emboj.7600910
D’Haeze, W., De Rycke, R., Mathis, R., Goormachtig, S., Pagnotta, S., Verplancke, C., et al. (2003). Reactive oxygen species and ethylene play a positive role in lateral root base nodulation of a semiaquatic legume. Proc. Natl. Acad. Sci. U.S.A. 100, 11789–11794. doi: 10.1073/pnas.1333899100
Dhindsa, R. S., Plumb-Dhindsa, P. L., and Reid, D. M. (1982). Leaf senescence and lipid peroxidation: effects of some phytohormones, and scavengers of free radicals and singlet oxygen. Physiol. Plant. 56, 453–457. doi: 10.1111/j.1399-3054.1982.tb04539.x
Du, H., Liu, H., and Xiong, L. (2013). Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 4:397. doi: 10.3389/fpls.2013.00397
Enders, T. A., and Strader, L. C. (2015). Auxin activity: past, present, and future. Am. J. Bot. 102, 180–196. doi: 10.3732/ajb.1400285
Espinoza, A., San Martín, A., López-Climent, M., Ruiz-Lara, S., Gómez-Cadenas, A., and Casaretto, J. A. (2013). Engineered drought-induced biosynthesis of α-tocopherol alleviates stress-induced leaf damage in tobacco. J. Plant Physiol. 170, 1285–1294. doi: 10.1016/j.jplph.2013.04.004
Feierabend, J., and Beevers, H. (1972). Developmental studies on microbodies in wheat leaves. Plant Physiol. 49, 28–32.
Foreman, J., Demidchik, V., Bothwell, J. H. F., Mylona, P., Miedema, H., Torres, M. A., et al. (2003). Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 422, 442–446. doi: 10.1038/nature01485
Foyer, C. H., and Noctor, G. (2005). Oxidant and antioxidant signalling in plants: a re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 28, 1056–1071. doi: 10.1111/j.1365-3040.2005.01327.x
Friml, J., Yang, X., Michniewicz, M., Weijers, D., Quint, A., Tietz, O., et al. (2004). A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux. Science 306, 862–865. doi: 10.1126/science.1100618
Fujita, Y., Fujita, M., Satoh, R., Maruyama, K., Parvez, M. M., Seki, M., et al. (2005). AREB1 is a transcription activator of novel ABRE-dependent ABA signaling that enhances drought stress tolerance in Arabidopsis. Plant Cell 17, 3470–3488. doi: 10.1105/tpc.105.035659
Galen, C., Rabenold, J. J., and Liscum, E. (2007). Functional ecology of a blue light photoreceptor: effects of phototropin-1 on root growth enhance drought tolerance in Arabidopsis thaliana. New Phytol. 173, 91–99. doi: 10.1111/j.1469-8137.2006.01893.x
Galván-Ampudia, C. S., and Offringa, R. (2007). Plant evolution: AGC kinases tell the auxin tale. Trends Plant Sci. 12, 541–547. doi: 10.1016/j.tplants.2007.10.004
Garcia, A. V., Al-Yousif, M., and Hirt, H. (2012). Role of AGC kinases in plant growth and stress responses. Cell. Mol. Life Sci. 69, 3259–3267. doi: 10.1007/s00018-012-1093-3
Gazarian, I. G., Lagrimini, L. M., Mellon, F. A., Naldrett, M. J., Ashby, G. A., and Thorneley, R. N. F. (1998). Identification of skatolyl hydroperoxide and its role in the peroxidase- catalysed oxidation of indol-3-yl acetic acid. Biochem. J. 333, 223–232.
Gechev, T. S., Gadjev, I. Z., and Hille, J. (2004). An extensive microarray analysis of AAL-toxin-induced cell death in Arabidopsis thaliana brings new insights into the complexity of programmed cell death in plants. Cell. Mol. Life Sci. 61, 1185–1197. doi: 10.1007/s00018-004-4067-2
Geisler, M., and Murphy, A. S. (2006). The ABC of auxin transport: the role of p-glycoproteins in plant development. FEBS Lett. 580, 1094–1102. doi: 10.1016/j.febslet.2005.11.054
Gill, S. S., and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
Grotewold, E. (ed.). (2006). The Science of Flavonoids. Berlin: Springer. doi: 10.1007/978-0-387-28822-2
Habig, W. H., Pabst, M. J., and Jakoby, W. B. (1974). Glutathione S-Transferases. J. Biol. Chem. 249, 7130–7139.
Haga, K., Hayashi, K.-I., and Sakai, T. (2014). PINOID AGC kinases are necessary for phytochrome-mediated enhancement of hypocotyl phototropism in Arabidopsis. Plant Physiol. 166, 1535–1545. doi: 10.1104/pp.114.244434
Harrison, M. A. (2012). “Cross-talk between phytohormone signaling pathways under both optimal and stressful environmental conditions,” in Phytohormones and Abiotic Stress Tolerance in Plants, eds N. A. Khan, R. Nazar, N. Iqbal, and N. A. Anjum (Berlin: Springer Science & Business Media), 1–49. doi: 10.1007/978-3-642-25829-9_2
Hasanuzzaman, M., Nahar, K., Gill, S. S., and Fujita, M. (2013). “Drought stress responses in plants, oxidative stress, and antioxidant defense,” in Climate Change and Plant Abiotic Stress Tolerance, eds N. Tuteja and S. S. Gill (Weinheim: Wiley-VCH Verlag GmbH & Co), 209–250. doi: 10.1002/9783527675265.ch09
Hirt, H. (2000). Connecting oxidative stress, auxin, and cell cycle regulation through a plant mitogen-activated protein kinase pathway. Proc. Natl. Acad. Sci.U.S.A. 97, 2405–2407. doi: 10.1073/pnas.97.6.2405
Hirt, H., Garcia, A. V., and Oelmüller, R. (2011). AGC kinases in plant development and defense. Plant Signal. Behav. 6, 1030–1033. doi: 10.4161/psb.6.7.15580
Hodges, D. M., DeLong, J. M., Forney, C. F., and Prange, R. K. (1999). Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 207, 604–611. doi: 10.1007/s004250050524
Holmgren, A., and Aslund, F. (1995). Biothiols Part B: Glutathione and Thioredoxin: Thiols in Signal Transduction and Gene Regulation. Amsterdam: Elsevier. doi: 10.1016/0076-6879(95)52031-7
Hong-bo, S., Li-ye, C., Ming-an, S., Abdul, J. C., and Hong-mei, M. (2008). Higher plant antioxidants and redox signaling under environmental stresses. C. R. Biol. 331, 433–441. doi: 10.1016/j.crvi.2008.03.011
Horling, F., Lamkemeyer, P., König, J., Finkemeier, I., Kandlbinder, A., Baier, M., et al. (2003). Dependent regulation of expression of the peroxiredoxin gene family in Arabidopsis. Plant Physiol. 131, 317–325. doi: 10.1104/pp.010017
Hughes, N. M., Carpenter, K. L., and Cannon, J. G. (2013). Estimating contribution of anthocyanin pigments to osmotic adjustment during winter leaf reddening. J. Plant Physiol. 170, 230–233. doi: 10.1016/j.jplph.2012.09.006
Hughes, N. M., Reinhardt, K., Feild, T. S., Gerardi, A. R., and Smith, W. K. (2010). Association between winter anthocyanin production and drought stress in angiosperm evergreen species. J. Exp. Bot. 61, 1699–1709. doi: 10.1093/jxb/erq042
Islam, T., Manna, M., and Reddy, M. K. (2015). Glutathione peroxidase of Pennisetum glaucum (PgGPx) is a functional Cd2+ dependent peroxiredoxin that enhances tolerance against salinity and drought stress. PLoS ONE 10:e0143344. doi: 10.1371/journal.pone.0143344
Jain, M., and Khurana, J. P. (2009). Transcript profiling reveals diverse roles of auxin-responsive genes during reproductive development and abiotic stress in rice. FEBS J. 276, 3148–3162. doi: 10.1111/j.1742-4658.2009.07033.x
Jaspers, P., and Kangasjärvi, J. (2010). Reactive oxygen species in abiotic stress signaling. Physiol. Plant. 138, 405–413. doi: 10.1111/j.1399-3054.2009.01321.x
Jiang, Z. Y., Woollard, A. C., and Wolff, S. P. (1990). Hydrogen peroxide production during experimental protein glycation. FEBS Lett. 268, 69–71. doi: 10.1016/0014-5793(90)80974-N
Joo, J. H., Bae, Y. S., and Lee, J. S. (2001). Role of auxin-induced reactive oxygen species in root gravitropism. Plant Physiol. 126, 1055–1056.
Jung, J.-H., and Park, C.-M. (2011). Auxin modulation of salt stress signaling in Arabidopsis seed germination. Plant Signal. Behav. 6, 1198–1200. doi: 10.4161/psb.6.8.15792
Junghans, U., Polle, A., Düchting, P., Weiler, E., Kuhlman, B., Gruber, F., et al. (2006). Adaptation to high salinity in poplar involves changes in xylem anatomy and auxin physiology. Plant Cell Environ. 29, 1519–1531. doi: 10.1111/j.1365-3040.2006.01529.x
Kazan, K. (2013). Auxin and the integration of environmental signals into plant root development. Ann. Bot. 112, 1655–1665. doi: 10.1093/aob/mct229
Kim, J. I., Baek, D., Park, H. C., Chun, H. J., Oh, D. H., Lee, M. K., et al. (2013). Overexpression of arabidopsis YUCCA6 in potato results in high-auxin developmental phenotypes and enhance. Mol. Plant 6, 337–349. doi: 10.1093/mp/sss100
King, J. J., Stimart, D. P., Fisher, R. H., and Bleecker, A. B. (1995). A mutation altering auxin homeostasis and plant morphology in Arabidopsis. Plant Cell 7, 2023–2037.
Kinoshita, N., Wang, H., Kasahara, H., Liu, J., MacPherson, C., Machida, Y., et al. (2012). IAA-Ala resistant3, an evolutionarily conserved target of miR167, mediates Arabidopsis root architecture changes during high osmotic stress. Plant Cell 24, 3590–3602. doi: 10.1105/tpc.112.097006
Kleine-Vehn, J., Huang, F., Naramoto, S., Zhang, J., Michniewicz, M., Offringa, R., et al. (2009). PIN auxin efflux carrier polarity is regulated by PINOID kinase-mediated recruitment into GNOM-independent trafficking in Arabidopsis. Plant Cell 21, 3839–3849. doi: 10.1105/tpc.109.071639
Kuhn, B. M., Geisler, M., Bigler, L., and Ringli, C. (2011). Flavonols accumulate asymmetrically and affect auxin transport in Arabidopsis. Plant Physiol. 156, 585–595. doi: 10.1104/pp.111.175976
Kuhn, B. M., Nodzyński, T., Errafi, S., Bucher, R., Gupta, S., Aryal, B., et al. (2017). Flavonol-induced changes in PIN2 polarity and auxin transport in the Arabidopsis thaliana rol1-2 mutant require phosphatase activity. Sci. Rep. 7:41906. doi: 10.1038/srep41906
Kuriyama, H., and Fukuda, H. (2002). Developmental programmed cell death in plants. Curr. Opin. Plant Biol. 5, 568–573. doi: 10.1016/S1369-5266(02)00305-9
Lanza, M., Garcia-Ponce, B., Castrillo, G., Catarecha, P., Sauer, M., Rodriguez-Serrano, M., et al. (2012). Role of actin cytoskeleton in brassinosteroid signaling and in its integration with the auxin response in plants. Dev. Cell 22, 1275–1285. doi: 10.1016/j.devcel.2012.04.008
Lee, M., Jung, J.-H., Han, D.-Y., Seo, P. J., Park, W. J., and Park, C.-M. (2012). Activation of a flavin monooxygenase gene YUCCA7 enhances drought resistance in Arabidopsis. Planta 235, 923–938. doi: 10.1007/s00425-011-1552-3
Ljung, K., Bhalerao, R. P., and Sandberg, G. (2001). Sites and homeostatic control of auxin biosynthesis in Arabidopsis during vegetative growth. Plant J. 28, 465–474. doi: 10.1046/j.1365-313X.2001.01173.x
Ludwig-Müller, J. (2011). Auxin conjugates: their role for plant development and in the evolution of land plants. J. Exp. Bot. 62, 1757–1773. doi: 10.1093/jxb/erq412
Maere, S., Heymans, K., and Kuiper, M. (2005). BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21, 3448–3449. doi: 10.1093/bioinformatics/bti551
Marrs, K. A. (1996). The functions and regulation of glutathione S-transeferases in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 47, 127–158.
Mittler, R. (2002). Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 7, 405–410. doi: 10.1016/S1360-1385(02)02312-9
Mittler, R., Vanderauwera, S., Gollery, M., and Van Breusegem, F. (2004). Reactive oxygen gene network of plants. Trends Plant Sci. 9, 490–498. doi: 10.1016/j.tplants.2004.08.009
Mortazavi, A., Williams, B. A., McCue, K., Schaeffer, L., and Wold, B. (2008). Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 5, 621–628. doi: 10.1038/nmeth.1226
Mouchel, C. F., Osmont, K. S., and Hardtke, C. S. (2006). BRX mediates feedback between brassinosteroid levels and auxin signalling in root growth. Nature 443, 458–461. doi: 10.1038/nature05130
Mühlenbock, P., Szechynska-Hebda, M., Plaszczyca, M., Baudo, M., Mateo, A., Mullineaux, P. M., et al. (2008). Chloroplast signaling and lesion simulating disease1 regulate crosstalk between light acclimation and immunity in Arabidopsis. Plant Cell 20, 2339–2356. doi: 10.1105/tpc.108.059618
Murata, Y., Pei, Z., Mori, I. C., and Schroeder, J. (2001). Abscisic acid activation of plasma membrane Ca2+ channels in guard cells requires cytosolic NAD(P)H and is differentially disrupted upstream and downstream of reactive oxygen species production in abi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell 13, 2513–2523. doi: 10.1105/tpc.010210.2514
Nakabayashi, R., Yonekura-Sakakibara, K., Urano, K., Suzuki, M., Yamada, Y., Nishizawa, T., et al. (2014). Enhancement of oxidative and drought tolerance in Arabidopsis by overaccumulation of antioxidant flavonoids. Plant J. 77, 367–379. doi: 10.1111/tpj.12388
Naser, V., and Shani, E. (2016). Auxin response under osmotic stress. Plant Mol. Biol. 91, 661–672. doi: 10.1007/s11103-016-0476-5
Neill, S., Desikan, R., and Hancock, J. (2002). Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 5, 388–395. doi: 10.1016/S1369-5266(02)00282-0
Nemhauser, J. L., Hong, F., and Chory, J. (2006). Different plant hormones regulate similar processes through largely nonoverlapping transcriptional responses. Cell 126, 467–475. doi: 10.1016/j.cell.2006.05.050
Noctor, G., and Foyer, C. H. (1998). Ascorbate and glutathione: keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 49, 249–279. doi: 10.1146/annurev.arplant.49.1.249
Paponov, I. A., Paponov, M., Teale, W., Menges, M., Chakrabortee, S., Murray, J. A. H., et al. (2008). Comprehensive transcriptome analysis of auxin responses in Arabidopsis. Mol. Plant 1, 321–337. doi: 10.1093/mp/ssm021
Park, J.-E., Park, J.-Y., Kim, Y.-S., Staswick, P. E., Jeon, J., Yun, J., et al. (2007a). GH3-mediated Auxin homeostasis links growth regulation with stress adaptation response in Arabidopsis. J. Biol. Chem. 282, 10036–10046. doi: 10.1074/jbc.M610524200
Park, J. E., Seo, P. J., Lee, A. K., Jung, J. H., Kim, Y. S., and Park, C. M. (2007b). An Arabidopsis GH3 gene, encoding an auxin-conjugating enzyme, mediates phytochrome B-regulated light signals in hypocotyl growth. Plant Cell Physiol. 48, 1236–1241. doi: 10.1093/pcp/pcm086
Pearce, L. R., Komander, D., and Alessi, D. R. (2010). The nuts and bolts of AGC protein kinases. Nat. Rev. Mol. Cell Biol. 11, 9–22. doi: 10.1038/nrm2822
Peer, W. A., Bandyopadhyay, A., Blakeslee, J. J., Makam, S. N., Chen, R. J., Masson, P. H., et al. (2004). Variation in expression and protein localization of the PIN family of auxin efflux facilitator proteins in flavonoid mutants with altered auxin transport in Arabidopsis thaliana. Plant Cell 16, 1898–1911. doi: 10.1105/tpc.021501
Peer, W. A., Blakeslee, J. J., Yang, H., and Murphy, A. S. (2011). Seven things we think we know about auxin transport. Mol. Plant 4, 487–504. doi: 10.1093/mp/ssr034
Peer, W. A., Cheng, Y., and Murphy, A. S. (2013). Evidence of oxidative attenuation of auxin signalling. J. Exp. Bot. 64, 2629–2639. doi: 10.1093/jxb/ert152
Peer, W. A., and Murphy, A. S. (2006). “Flavonoids as signal molecules,” in The Science of Flavonoids, ed. E. Grotewold (Berlin: Springer), 239–268.
Peer, W. A., and Murphy, A. S. (2007). Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci. 12, 556–563. doi: 10.1016/j.tplants.2007.10.003
Petersen, L. N., Ingle, R. A., Knight, M. R., and Denby, K. J. (2009). OXI1 protein kinase is required for plant immunity against Pseudomonas syringae in Arabidopsis. J. Exp. Bot. 60, 3727–3735. doi: 10.1093/jxb/erp219
Polle, A., and Rennenberg, H. (1993). “Significance of antioxidants in plant adaptation to environmental stress,” in Plant Adaptations to Environmental Stresses, eds L. Fowden, T. Mansfield, and J. Stoddart (London: Chapman & Hall), 263–273.
Potters, G., Pasternak, T. P., Guisez, Y., and Jansen, M. A. K. (2009). Different stresses, similar morphogenic responses: integrating a plethora of pathways. Plant Cell Environ 32, 158–169. doi: 10.1111/j.1365-3040.2008.01908.x
Potters, G., Pasternak, T. P., Guisez, Y., Palme, K. J., and Jansen, M. A. K. (2007). Stress-induced morphogenic responses: growing out of trouble? Trends Plant Sci. 12, 98–105. doi: 10.1016/j.tplants.2007.01.004
Raftopoulou, M. (2004). PINOID pinpoints auxin. Nat. Cell Biol. 6, 1149–1149. doi: 10.1038/ncb1204-1149
Rahman, A. (2013). Auxin: a regulator of cold stress response. Physiol. Plant 147, 28–35. doi: 10.1111/j.1399-3054.2012.01617.x
Ramel, F., Sulmon, C., Bogard, M., Couée, I., and Gouesbet, G. (2009). Differential patterns of reactive oxygen species and antioxidative mechanisms during atrazine injury and sucrose-induced tolerance in Arabidopsis thaliana plantlets. BMC Plant Biol. 9:28. doi: 10.1186/1471-2229-9-28
Remy, E., Cabrito, T. R., Baster, P., Batista, R. A., Teixeira, M. C., Friml, J., et al. (2013). A major facilitator superfamily transporter plays a dual role in polar auxin transport and drought stress tolerance in Arabidopsis. Plant Cell 25, 901–926. doi: 10.1105/tpc.113.110353
Rentel, M. C., Lecourieux, D., Ouaked, F., Usher, S. L., Petersen, L., Okamoto, H., et al. (2004). OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature 427, 858–861. doi: 10.1038/nature02353
Saini, K., Markakis, M. N., Zdanio, M., Balcerowicz, D. M., Beeckman, T., De Veylder, L., et al. (2017). Alteration in auxin homeostasis and signaling by overexpression of PINOID kinase causes leaf growth defects in Arabidopsis thaliana. Front. Plant Sci. 8:1009. doi: 10.3389/FPLS.2017.01009
Sandalio, L. M., Rodríguez-Serrano, M., and Romero-Puertas, M. C. (2016). Leaf epinasty and auxin: a biochemical and molecular overview. Plant Sci. 253, 187–193. doi: 10.1016/j.plantsci.2016.10.002
Sarath, G., Hou, G., Baird, L. M., and Mitchell, R. B. (2007). Reactive oxygen species, ABA and nitric oxide interactions on the germination of warm-season C4-grasses. Planta 226, 697–708. doi: 10.1007/s00425-007-0517-z
Schopfer, P. (2001). Hydroxyl radical-induced cell-wall loosening in vitro and in vivo: implications for the control of elongation growth. Plant J. 28, 679–688. doi: 10.1046/j.1365-313x.2001.01187.x
Seo, P. J., Xiang, F., Qiao, M., Park, J.-Y., Lee, Y. N., Kim, S.-G., et al. (2009). The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol. 151, 275–289. doi: 10.1104/pp.109.144220
Shen, C., Bai, Y., Wang, S., Zhang, S., Wu, Y., Chen, M., et al. (2010). Expression profile of PIN, AUX/LAX and PGP auxin transporter gene families in Sorghum bicolor under phytohormone and abiotic stress. FEBS J. 277, 2954–2969. doi: 10.1111/j.1742-4658.2010.07706.x
Shi, H., Chen, L., Ye, T., Liu, X., Ding, K., and Chan, Z. (2014). Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 82, 209–217. doi: 10.1016/j.plaphy.2014.06.008
Shibasaki, K., Uemura, M., Tsurumi, S., and Rahman, A. (2009). Auxin response in Arabidopsis under cold stress: underlying molecular mechanisms. Plant Cell 21, 3823–3838. doi: 10.1105/tpc.109.069906
Šimášková, M., O’Brien, J. A., Khan, M., Van Noorden, G., Ötvös, K., Vieten, A., et al. (2015). Cytokinin response factors regulate PIN-FORMED auxin transporters. Nat. Commun. 6:8717. doi: 10.1038/ncomms9717
Song, X.-G., She, X.-P., He, J.-M., Huang, C., and Song, T. (2006). Cytokinin- and auxin-induced stomatal opening involves a decrease in levels of hydrogen peroxide in guard cells of Vicia faba. Funct. Plant Biol. 33, 573. doi: 10.1071/FP05232
Taylor, L. P., and Grotewold, E. (2005). Flavonoids as developmental regulators. Curr. Opin. Plant Biol. 8, 317–323. doi: 10.1016/j.pbi.2005.03.005
Teale, W. D., Paponov, I. A., and Palme, K. (2006). Auxin in action: signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 7, 847–859. doi: 10.1038/nrm2020
Terasaka, K., Blakeslee, J. J., Titapiwatanakun, B., Peer, W. A., Bandyopadhyay, A., Makam, S. N., et al. (2005). PGP4, an ATP binding cassette P-glycoprotein, catalyzes auxin transport in Arabidopsis thaliana roots. Plant Cell 17, 2922–2939. doi: 10.1105/tpc.105.035816
Thimm, O., Bläsing, O., Gibon, Y., Nagel, A., Meyer, S., Krüger, P., et al. (2004). Mapman: a user-driven tool to display genomics data sets onto diagrams of metabolic pathways and other biological processes. Plant J. 37, 914–939. doi: 10.1111/j.1365-313X.2004.02016.x
Tognetti, V. B., Mühlenbock, P., and van Breusegem, F. (2012). Stress homeostasis - the redox and auxin perspective. Plant Cell Environ. 35, 321–333. doi: 10.1111/j.1365-3040.2011.02324.x
Tognetti, V. B., Van Aken, O., Morreel, K., Vandenbroucke, K., van de Cotte, B., De Clercq, I., et al. (2010). Perturbation of indole-3-Butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell 22, 2660–2679. doi: 10.1105/tpc.109.071316
Verslues, P. E., Agarwal, M., Katiyar-Agarwal, S., Zhu, J., and Zhu, J.-K. (2006). Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 45, 523–539. doi: 10.1111/j.1365-313X.2005.02593.x
Wan, Y., Jasik, J., Wang, L., Hao, H., Volkmann, D., Menzel, D., et al. (2012). The signal transducer NPH3 integrates the phototropin1 photosensor with PIN2-based polar auxin transport in arabidopsis root phototropism. Plant Cell 24, 551–565. doi: 10.1105/tpc.111.094284
Wang, S., Bai, Y., Shen, C., Wu, Y., Zhang, S., Jiang, D., et al. (2010). Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genomics 10, 533–546. doi: 10.1007/s10142-010-0174-3
Werner, T., Nehnevajova, E., Kollmer, I., Novak, O., Strnad, M., Kramer, U., et al. (2010). Root-specific reduction of cytokinin causes enhanced root growth, drought tolerance, and leaf mineral enrichment in Arabidopsis and tobacco. Plant Cell 22, 3905–3920. doi: 10.1105/tpc.109.072694
Wingler, A., Lea, P. J., Quick, W. P., and Leegood, R. C. (2000). Photorespiration: metabolic pathways and their role in stress protection. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 355, 1517–1529. doi: 10.1098/rstb.2000.0712
Woo, Y. M., Park, H. J., Su’udi, M., Yang, J. I., Park, J. J., Back, K., et al. (2007). Constitutively wilted 1, a member of the rice YUCCA gene family, is required for maintaining water homeostasis and an appropriate root to shoot ratio. Plant Mol. Biol. 65, 125–136. doi: 10.1007/s11103-007-9203-6
Yoshida, T., Fujita, Y., Sayama, H., Kidokoro, S., Maruyama, K., Mizoi, J., et al. (2010). AREB1, AREB2, and ABF3 are master transcription factors that cooperatively regulate ABRE-dependent ABA signaling involved in drought stress tolerance and require ABA for full activation. Plant J. 61, 672–685. doi: 10.1111/j.1365-313X.2009.04092.x
Zhang, Q., Li, J., Zhang, W., Yan, S., Wang, R., Zhao, J., et al. (2012). The putative auxin efflux carrier OsPIN3t is involved in the drought stress response and drought tolerance. Plant J. 72, 805–816. doi: 10.1111/j.1365-313X.2012.05121.x
Zhang, S.-W., Li, C.-H., Cao, J., Zhang, Y.-C., Zhang, S.-Q., Xia, Y.-F., et al. (2009). Altered architecture and enhanced drought tolerance in rice via the down-regulation of indole-3-acetic acid by TLD1/OsGH3.13 activation. Plant Physiol. 151, 1889–1901. doi: 10.1104/pp.109.146803
Zhang, Z., Li, Q., Li, Z., Staswick, P. E., Wang, M., Zhu, Y., et al. (2007). Dual regulation role of GH3.5 in salicylic acid and auxin signaling during Arabidopsis-Pseudomonas syringae interaction. Plant Physiol. 145, 450–464. doi: 10.1104/pp.107.106021
Zhao, Y., Christensen, S. K., Fankhauser, C., Cashman, J. R., Cohen, J. D., Weigel, D., et al. (2001). A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 291, 306–309. doi: 10.1126/science.291.5502.306
Keywords: auxin, PINOID (PID), reactive oxygen species (ROS), flavonoids, drought stress, osmotic stress
Citation: Saini K, AbdElgawad H, Markakis MN, Schoenaers S, Asard H, Prinsen E, Beemster GTS and Vissenberg K (2017) Perturbation of Auxin Homeostasis and Signaling by PINOID Overexpression Induces Stress Responses in Arabidopsis. Front. Plant Sci. 8:1308. doi: 10.3389/fpls.2017.01308
Received: 07 April 2017; Accepted: 12 July 2017;
Published: 02 August 2017.
Edited by:
Vicent Arbona, Jaume I University, SpainReviewed by:
Luisa M. Sandalio, Consejo Superior de Investigaciones Científicas (CSIC), SpainChristine Helen Foyer, University of Leeds, United Kingdom
Copyright © 2017 Saini, AbdElgawad, Markakis, Schoenaers, Asard, Prinsen, Beemster and Vissenberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kris Vissenberg, a3Jpcy52aXNzZW5iZXJnQHVhbnR3ZXJwZW4uYmU=