 Aditya Banerjee
Aditya Banerjee Shabir H. Wani
Shabir H. Wani Aryadeep Roychoudhury
Aryadeep Roychoudhury- 1Post Graduate Department of Biotechnology, St. Xavier's College-Autonomous, Kolkata, India
- 2Mountain Research Centre for Field Crops, Sher-e-Kashmir University of Agricultural Sciences and Technology of Kashmir, Srinagar, India
- 3Department of Plant Soil and Microbial Sciences, Michigan State University, East Lansing, MI, United States
Cold Stress in Plants
Higher plants are sedentary organisms which inevitably endure a variety of environmental stresses throughout the life cycle. Abiotic stresses can be atmospheric like cold, heat and UV irradiation; or can also be edaphic like salinity, drought, and heavy metal toxicity (Wani and Gosal, 2011; Surekha et al., 2015). Of all these, cold stress is regarded as a major environmental factor which limits agricultural expansion and crop productivity in hilly terrains (Sanghera et al., 2011). Non-freezing low temperatures deteriorate plant growth physiology by inducing chilling injuries like photosynthesis-associated damages, chlorosis, unregulated apoptosis, loss of membrane fluidity and ultimately wilting (Wani et al., 2016). Depending on the extent of sensitivity among plants, cold stress has been sub-divided into two types. Chilling stress is characterized by 0–15°C, whereas temperatures below 0°C cause freezing stress (Wani et al., 2013). By virtue of cold acclimation and associated alterations at the molecular and biochemical levels, temperate climatic plants exhibit greater ranges of cold tolerance compared to their tropical and sub-tropical counterparts (Yamaguchi-Shinozaki and Shinozaki, 2006). Deciphering the epigenomic landscape in plants exposed to cold conditions is a rapidly developing field (Hu et al., 2011). Intricate research focussing on epigenetic processes during cold stress has led to the identification of molecular targets which can be genetically manipulated to generate cold tolerant lines.
Vernalization is a floral regulatory process preventing precocious flowering during autumn or winter. It gradually promotes flowering competence after prolonged exposures to cold conditions in a species-dependent manner (Kim et al., 2009). Physiologically, vernalization is a “memory response” which is correlated with epigenetic regulation, as observed in the model plant Arabidopsis thaliana (Song et al., 2012).
Epigenetic Control of Plants during Cold Acclimation
Histone Modifications during Cold Acclimation
The expression of epigenetic regulators varies under cold conditions (Banerjee and Roychoudhury, 2017a). Up regulated expression of the histone deacetylases (HDACs) was observed in Zea mays during cold acclimation. This resulted in the global deacetylation at the lysine residues on the histone subunits H3 and H4 (Hu et al., 2011). Cold-dependent alternative splicing was observed in the histone demethylase, Jumonji C domain-containing gene, JMJC5 in Medicago truncatula (Shen et al., 2016). Sequencing of the products from the four alternatively spliced RNA isoforms revealed the presence of three premature termination codon-containing variants and a full length protein. Under cold stress, the protein variant having 3′-alternative splice site at the second intron exhibited less sensitivity than the one containing the splice site at the first intron. All the variants were however sensitive to nonsense-mediated decay (Shen et al., 2016). Selective non-silencing of the heterochromatin tandem repeats was reported in Z. mays exposed to cold. These plants accumulated non-canonical histone subunit H3, acetylated on the ninth lysine residue (H3K9ac) along with reduced DNA methylation and dimethylation at H3K9 (H3K9me2) in the unsilenced stretches (Hu et al., 2012). Pan (2013) reviewed that the thermosensory responses in Arabidopsis were modulated via the interaction of histone variants like H2A.Z with the nucleosomes.
Phosphorylation of the N-terminal tails of histone H3 are post-translational nucleosomal signals manipulating phytocellular stochastics (Nowak and Corces, 2000). Two closely associated Ser/Thr kinases (At3g03940 and At5g18190) crucially phosphorylate histone H3-Thr 3 (H3T3ph) in the pericentromeric/knob regions in Arabidopsis to maintain heterochromatin organization and chromosome functions (Wang et al., 2015). Arabidopsis plants exposed to cold stress-like osmotic conditions exhibited increased H3T3ph and H3K4me3 at the genomic level with decreased histone H3 occupancy at the pericentromeric/knob regions (Wang et al., 2015). High H3T3ph levels correlated with the transcriptionally active loci of stress-responsive genes (Bej and Basak, 2017).
Cold stress up regulates several downstream transcription factors (TFs) and their target genes (Wani et al., 2016). Histone occupancy-dependent and –independent decrease of H3K27me3 was observed in Cold Regulated 15A (COR15A) and Galactinol Synthase 3 in Arabidopsis. H3K27me3 decrease permitted the de-repression of these cold responsive genes and ensured systemic acclimatization to low temperatures (Kwon et al., 2009; Banerjee and Roychoudhury, 2016). Gene activating histone acetylation was noted in cold responsive genes like Drought Responsive Element Binding 1 (DREB1) and COR413 in Z. mays exposed to cold (Hu et al., 2011; Banerjee and Roychoudhury, 2017b). Low temperatures induced similar histone acetylation in the Oryza sativa DREB1b promoter (Roy et al., 2014). The RING finger E3 ligase, HIGH EXPRESSION OF OSMOTICALLY RESPONSIVE GENE 1 (HOS1) negatively regulates crucial cold responsive genes like C-repeat binding factors (CBFs)/ DREB1s, CORs, Responsive to Dehydration genes (RDs), cold induced genes (KINs), and the cold induced TF, ICE1 during cold stress (Dong et al., 2006). An interesting epigenetic-floral regulation has been observed in Arabidopsis where HOS1 dissociated HDAC6 from the Flowering Locus C (FLC) chromatin to maintain FLC expression (Jung et al., 2013; Figure 1).
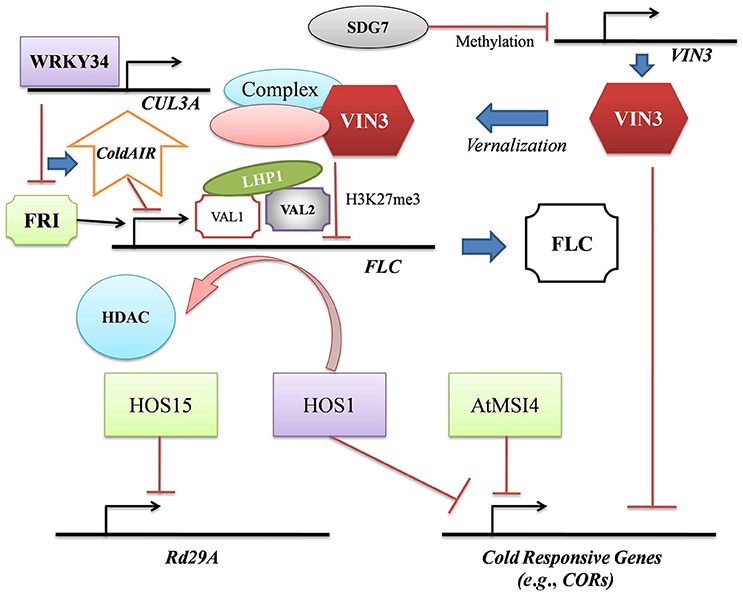
Figure 1. The epigenetic regulation during cold stress highlights that the intricate crosstalks between cold acclimation and vernalization cascades are mediated by VIN3 and HOS1. During vernalization, reduced abundance of SDG7 reverses VIN3 suppression. Functional VIN3-complex simultaneously inhibits the expression of CORs (required for cold acclimation) and FLC via repressive histone methylation and promotes vernalization. The epigenetic regulation of FLC is multivariate. LHP1 associates with VAL1 and VAL2 to inhibit FLC transcription. WRKY34 promotes the expression of CUL3A, encoding an ubiquitin E3 ligase. CUL3A proteasomally degrades FRI, the positive regulator of FLC. The lncRNA, ColdAIR triggers post-transcriptional gene silencing of FLC and positively regulates vernalization. The second crosstalk mediator, HOS1 promotes FLC expression via HDAC dissociation and also inhibits the transcription of CORs. HOS1 thus acts as a negative regulator of vernalization as well as cold acclimation. HOS15 generally improves cold stress tolerance but inhibits the expression of drought-inducible Rd29A. MSI4 acts as a negative regulator of cold acclimation.
Low temperatures induced the accumulation of a WD40-repeat protein, MSI1 (a subunit of the chromatin assembly factor 1) and the 600 kDa polycomb complex (MEDEA, FERTILIZATION-INDEPENDENT ENDOSPERM and FERTILIZATION-INDEPENDENT SEED 2). The MSI-like protein, MSI4/FVE negatively regulated COR genes during cold stress in Arabidopsis (Dhar et al., 2014; Figure 1). Recent studies have identified distinct RNA recognition motif and PWWP domains in AtMSI4 which regulate the assembly of nucleoprotein complexes and determine the pleiotropic functions in response to stress (Kenzior and Folk, 2015). Significant sequence homology exists between the cold induced WD40-repeat protein, HOS15 and the human transducin-beta like protein 1 (TBL1) (Zhu et al., 2008). TBL1 constitutes the SMRT/N-CoR repressor complex which interacts with HDACs. Thus, deacetylation at H4 is supposedly regulated by HOS15 to improve cold tolerance in Arabidopsis. The hos15 mutant lines exhibited cold hypersensitivity with increased expression of stress-inducible genes like Rd29A (Kim et al., 2015) (Figure 1). Both the abscisic acid-dependent and –independent pathways up regulate Rd29A expression under drought (Roychoudhury and Banerjee, 2015). Establishment of evidential correlation between HOS15 and H4 deacetylation during Rd29A expression can open a new avenue in plant cold tolerance research.
DNA Methylation during Cold Acclimation
Investigating the spatio-temporal variability in cold-induced DNA methylation is essential since it ensures proper chromatin packaging and conditional gene expression (Banerjee and Roychoudhury, 2017a). Methylation-sensitive amplified fragment-length polymorphism markers detected the changes in cytosine methylation in the alpine subnival plant, Chorispora bungeana exposed to 4°C chilling and −4°C freezing stress. Rapid alterations in cytosine methylation occurred throughout the periods of chilling and freezing (Song et al., 2015). Comparative methylome analysis in Populus simonii grown under cold, osmotic, heat and salt stresses identified condition-dependent variable cytosine methylation patterns and 1,376 stress-specific differentially methylated regions (SDMRs) (Song et al., 2016). Epigenetic regulation was observed in the SDMR162 region consisting of non-coding RNAs like microRNA 396e (miR396e) and long-non-coding RNA 00268512 (lncRNA00268512). It was proposed that lncRNA00268512 controlled miR396e transcription by acting as a target mimic. Signature methylation marks regulated miR6445a stability in P. simonii exposed to cold, salt, osmotic and heat stresses (Song et al., 2016). Non-coding RNAs are also known to play crucial roles in designing the epigenomic landscape in plants exposed to multiple abiotic stresses (Banerjee et al., 2016; Banerjee and Roychoudhury, 2017c). lncRNAs contain conserved stress-specific elements involved in the binding of TFs, phytochrome-interacting factor 4 (PIF4) and PIF5 (Di et al., 2014). Among the 245 poly A+ and 58 poly A- lncRNAs, a salt-responsive UUC motif and a cold-responsive AU-rich stem-loop were annotated (Di et al., 2014). Characterization of such cold stress responsive cis acting elements is crucial for deciphering the unidentified transient interactions occurring between the non-coding RNAs and their target genes.
Vernalization in the Context of Epigenetics
Vernalization in the Arabidopsis ecotypes is regulated by FLC, Vernalization 1 (VRN1), VRN2 and Vernalization Insensitive 3 (VIN3). Cold stress induces the expression of VIN3, a homeodomain finger containing gene encoding protein. VIN3 associates with an unidentified complex and triggers repressive H3K27me3 and H3K9me in the FLC chromatin (Kim and Sung, 2017; Figure 1). The complex also initiates loss of H3 acetylation and H3K4 methylation, thus suppressing FLC expression and promoting vernalization. It appears that the VIN3 and FLC families actually co-evolved to ensure proper epigenetic regulation of vernalization (Kim and Sung, 2013). In another instance, the H3K27 methyltransferase, SET DOMAIN GROUP 7 (SDG7) negatively regulated the temporal expression of VIN3 in Arabidopsis under control conditions (Figuer 1). Abiding by the FLC-VIN3 regulatory model, sdg7 Arabidopsis mutants exhibited partial vernalization even without cold exposure (Lee et al., 2015). The polycomb group (PcG) proteins along with two homologous trans-acting epigenome readers, VIVIPAROUS 1/ABA-INSENSITIVE 3-LIKE 1 (VAL1) and VAL2 stably repress FLC during prolonged cold treatment (Yuan et al., 2016). The epigenome readers recognized the cis-acting Cold Memory Elements and H3K27me3 to directly associate with LIKE HETEROCHROMATIN PROTEIN 1. This increased H3K27me3 in the FLC nucleation region leading to its repression during vernalization (Yuan et al., 2016; Figure 1).
The scaffold protein, FRIGIDA (FRI) recruits epigenetic factors to regulate vernalization-related floral gene expression in winter annual accessions of Arabidopsis (Hu et al., 2014). It was reported that the TF, WRKY34 promotes the expression of the ubiquitin E3 ligase, CULLIN 3A which proteasomally degrades FRI and triggers the accumulation of lncRNA, Cold Induced Long Antisense Intragenic RNA (ColdAIR) in the late phases of vernalization. High ColdAIR levels reduce H3K4me3 in FLC and facilitates flowering after vernalization (Hu et al., 2014; Figure 1).
Trans-generational repression of FLC during vernalization is maintained as a cellular memory via the mitotic inheritance of the repressive histone marks in the chromatin (Achrem et al., 2012). However, repeated brief cold exposures due to local weather disturbances degrade the central floral regulator, CONSTANS (CO) and delay flowering in Arabidopsis. The E3-ubiquitin ligase HOS1 together with its target, CO serve as the molecular integrator of the photoperiodic and cold stress pathways (Jung et al., 2012). Research in temperate cereal crops (Triticum spp., Hordeum vulgare, Avena sativa, Secale cereale etc.) has verified the increase in gene activating H3K9me3 and decrease in gene suppressing H3K27me3 in VRN1 during vernalization (Oliver et al., 2009). Such variations in H3K methylation status signify vernalization memory in cereals (Banerjee and Roychoudhury, 2017c). However, increased H3K27me3 and decreased H3K4me3 levels in VRN2 and Flowering Locus T1 (FT1) were observed in both the vernalized and non-vernalized seedlings. This depicts that unlike in VRN1, cold treatment does not modify the chromatin of these genes (Achrem et al., 2012). Vernalization in cereal crops is supposedly mediated by DNA demethylation. The PcG proteins recognizing H3K27me3 sites inhibit VRN2 and FT expression in the germinating caryopses but refrain from regulating VRN1 expression in the shoot apices (Li and Liu, 2010). Such spatial regulation of epigenetic factors complicates the systemic overview of cold treatment in plants. Vernalized Arabidopsis seedlings exposed to 30°C, directly after cold treatment exhibited low H3K27me3 at the FLC locus (Bouche et al., 2015). Thus cold shock memory imprinted in the plant epigenome is ushered by the condition-dependent reversible alteration of chromatin architecture.
Future Perspectives
Epigenetic regulation under cold stress is a manifestation of multiple crosstalks (Figure 1) and plant intelligence. Another crosstalk among the genes involved in flowering, vernalization and generation of cold tolerance can be hypothesized in Triticum monococcum where VRN1 initiates the regulatory cascade to down-regulate cold acclimation genes like CORs and CBFs (Galiba et al., 2009). Thus, it is quintessential to design an epigenomic blueprint acting as a global database of chromatin alterations occurring at low temperatures. Molecular characterization of the epigenetic stress memory is crucially required to integrate plant behavioral responses with intelligence. Stress conditions induce the production of volatile organic carbons which often act as inter-species communicating molecules (Ueda et al., 2012). The epigenetic regulation of such metabolite synthesis during cold stress remains to be elucidated. Genome-wide association studies followed by epiallele mining and subsequent annotations on next generation sequencing platforms should be undertaken to unravel the intricate epigenomic landscape especially in staple crops exposed to cold stress. Understanding the spatio-temporal expression of such epialleles, their interactions with small RNAs during cold stress and stringent mapping of candidate quantitative trait loci can identify potential molecular targets which can be engineered to generate tolerant phenotypes. These can then be recommended for field trials.
Author Contributions
AR and SW suggested the title of the opinion article. AB designed the article, drafted the entire manuscript and arranged the references. AR incorporated all the necessary modifications. All the authors checked and confirmed the final version of the manuscript.
Funding
SW is thankful to University Grants Commission, New Delhi India for Raman Post Doctoral Fellowship grant. AB is grateful to University Grants Commission, Government of India, for providing research fellowship. The Council of Scientific and Industrial Research (CSIR), Government of India, is also acknowledged.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Achrem, M., Skuza, L., Kalinka, A., Szucko, I., Filip, E., et al. (2012). Role of epigenetic mechanisms in plant response to low temperature. Acta Biol. Cracoviensia Ser. Bot. 54, 7–15. doi: 10.2478/v10182-012-0014-y
Banerjee, A., and Roychoudhury, A. (2016). Group II late embryogenesis abundant (LEA) proteins: structural and functional aspects in plant abiotic stress. Plant Growth Regul. 79, 1–17. doi: 10.1007/s10725-015-0113-3
Banerjee, A., and Roychoudhury, A. (2017a). Epigenetic regulation during salinity and drought stress in plants: histone modifications and DNA methylation. Plant Gene. doi: 10.1016/j.plgene.2017.05.011. [Epub ahead of print].
Banerjee, A., and Roychoudhury, A. (2017b). Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma 254, 3–16. doi: 10.1007/s00709-015-0920-4
Banerjee, A., and Roychoudhury, A. (2017c). The gymnastics of epigenomics in rice. Plant Cell Rep. doi: 10.1007/s00299-017-2192-2. [Epub ahead of print].
Banerjee, A., Roychoudhury, A., and Krishnamoorthi, S. (2016). Emerging techniques to decipher microRNAs (miRNAs) and their regulatory role in conferring abiotic stress tolerance of plants. Plant Biotechnol. Rep. 10, 185–205. doi: 10.1007/s11816-016-0401-z
Bej, S., and Basak, J. (2017). “Abiotic stress induced epigenetic modifications in plants: How much do we know?” in Plant Epigenetics, eds N. Rajewsky and S. Jurga, J. Barciszewski (Germany: Springer International Publishing AG), 493–512.
Bouche, F., Detry, N., and Perilleux, C. (2015). Heat can erase marks of vernalization in Arabidopsis. Plant Signal. Behav. 10:e990799. doi: 10.4161/15592324.2014.990799
Dhar, M. K., Vishal, P., Sharma, R., and Kaul, S. (2014). Epigenetic dynamics: role of epimarks and underlying machinery in plants exposed to abiotic stress. Int. J. Genomics 2014:187146. doi: 10.1155/2014/187146
Di, C., Yuan, J., Wu, Y., Li, J., Lin, H., et al. (2014). Characterization of stress-responsive lncRNAs in Arabidopsis thaliana by integrating expression, epigenetic and structural features. Plant J. 80, 848–861. doi: 10.1111/tpj.12679
Dong, C. H., Agarwal, M., Zhang, Y., Xie, Q., and Zhu, J. K. (2006).The negative regulator of plant cold responses, HOS1, is a RING E3 ligase that mediates the ubiquitination and degradation of ICE1. Proc. Natl. Acad. Sci. U.S.A. 103, 8281–8286. doi: 10.1073/pnas.0602874103
Galiba, B., Vagujfalvi, A., Li, C. H., Soltesz, A., and Dubcovsky, J. (2009). Regulatory genes involved in the determination of frost tolerance in temperate cereals. Plant Sci. 176, 12–19. doi: 10.1016/j.plantsci.2008.09.016
Hu, X., Kong, X., Wang, C., Ma, L., Zhao, J., et al. (2014). Proteasome-mediated degradation of FRIGIDA modulates flowering time in Arabidopsis during vernalization. Plant Cell 26, 4763–4781. doi: 10.1105/tpc.114.132738
Hu, Y., Zhang, L., Zhao, L., Li, J., He, S., Zhou, K., et al. (2011). Trichostatin a selectively suppresses the cold-induced transcription of the ZmDREB1 gene in maize. PLoS ONE 6:e22132. doi: 10.1371/journal.pone.0022132
Hu, Y., Zhang, L., He, S., Huang, M., Tan, J., Zhao, L., et al. (2012). Cold stress selectively unsilences tandem repeats in heterochromatin associated with accumulation of H3K9ac. Plant Cell Environ. 35, 2130–2142. doi: 10.1111/j.1365-3040.2012.02541.x
Jung, J. H., Park, J. H., Lee, S., To, T. K., Kim, J. M., Seki, M., et al. (2013). The cold signaling attenuator high expression of osmotically responsive gene 1 activates flowering locus C transcription via chromatin remodeling under short-term cold stress in Arabidopsis. Plant Cell 25, 4378–4390. doi: 10.1105/tpc.113.118364
Jung, J.-H., Seo, P. J., and Park, C.-M. (2012). The E3 ubiquitin ligase HOS1 regulates Arabidopsis flowering by mediating constans degradation under cold stress. J. Biol. Chem. 287, 43277–43287. doi: 10.1074/jbc.M112.394338
Kenzior, A., and Folk, W. R. (2015). Arabidopsis thaliana MSI4/FVE associates with members of a novel family of plant specific PWWP/RRM domain proteins. Plant Mol. Biol. 87, 329–339. doi: 10.1007/s11103-014-0280-z
Kim, D. H., Doyle, M. R., Sung, S., and Amasino, R. M. (2009). Vernalization: winter and the timing of flowering in plants. Annu. Rev. Cell Dev. Biol. 25, 277–299. doi: 10.1146/annurev.cellbio.042308.113411
Kim, D. H., and Sung, S. (2013). Coordination of the vernalization response through a VIN3 and FLC gene family regulatory network in Arabidopsis. Plant Cell 25, 454–469. doi: 10.1105/tpc.112.104760
Kim, D. H., and Sung, S. (2017). Accelerated vernalization response by an altered PHD-finger protein in Arabidopsis. Plant Signal. Behav. 12:e1308619. doi: 10.1080/15592324.2017.1308619
Kim, J.-M., Sasaki, T., Ueda, M., Sako, K., and Seki, M. (2015). Chromatin changes in response to drought, salinity, heat, and cold stresses in plants. Front. Plant Sci. 6:114. doi: 10.3389/fpls.2015.00114
Kwon, C. S., Lee, D., Choi, G., and Chung, W. I. (2009). Histone occupancy-dependent and –independent removal of H3K27 trimethylation at cold-responsive genes in Arabidopsis. Plant J. 60, 112–121. doi: 10.1111/j.1365-313X.2009.03938.x
Lee, J., Yun, J. Y., Zhao, W., Shen, W. H., and Amasino, R. M. (2015). A methyltransferase required for proper timing of the vernalization response in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 112, 2269–2274. doi: 10.1073/pnas.1423585112
Li, X., and Liu, Y. (2010). The conversion of spring wheat into winter wheat and vice versa: false claim or Lamarckian inheritance. J. Biosci. 35, 321–325. doi: 10.1007/s12038-010-0035-1
Nowak, S. J., and Corces, V. G. (2000). Phosphorylation of histone H3 correlates with transcriptionally active loci. Genes Dev. 14, 3003–3013. doi: 10.1101/gad.848800
Oliver, S. N., Finnegan, E. J., Dennis, E. S., Peacock, W. J., and Trevaskis, B. (2009). Vernalization induced flowering in cereals is associated with changes in histone methylation at the VERNALIZATION 1 gene. Proc. Natl. Acad. Sci. U.S.A. 106, 8386–8391. doi: 10.1073/pnas.0903566106
Pan, L. N. (2013). Epigenetic regulation of abiotic stress response in plants to improve the stress tolerance. Yi Chuan 35, 745–751. doi: 10.3724/SP.J.1005.2013.00745
Roy, D., Paul, A., Roy, A., Ghosh, R., Ganguly, P., and Chaudhuri, S. (2014). Differential acetylation of histone H3 at the regulatory region of OsDREB1b facilitates chromatin remodeling and transcription activation during cold stress. PLoS ONE 9:e100343. doi: 10.1371/journal.pone.0100343
Roychoudhury, A., and Banerjee, A. (2015). Trancriptome analysis of abiotic stress response in plants. Transcriptomics 3:2. doi: 10.4172/2329-8936.1000e115
Sanghera, G. S., Wani, S. H., Hussain, W., and Singh, N. B. (2011). Engineering cold stress tolerance in crop plants. Curr. Genomics 12, 30–43. doi: 10.2174/138920211794520178
Shen, Y., Wu, X., Liu, D., Song, S., Liu, D., and Wang, H. (2016). Cold-dependent alternative splicing of a Jumonji C domain-containing gene MtJMJC5 in Medicago truncatula. Biochem. Biophys. Res. Commun. 474, 271–276. doi: 10.1016/j.bbrc.2016.04.062
Song, J., Angel, A., Howard, M., and Dean, C. (2012). Vernalization—a cold induced epigenetic switch. J. Cell Sci. 125, 3723–3731. doi: 10.1242/jcs.084764
Song, Y., Ci, D., Tian, M., and Zhang, D. (2016). Stable methylation of a non-coding RNA gene regulates gene expression in response to abiotic stress in Populus simonii. J. Exp. Bot. 67, 1477–1492. doi: 10.1093/jxb/erv543
Song, Y., Liu, L., Feng, Y., Wei, Y., Yue, X., et al. (2015). Chilling- and freezing-induced alterations in cytosine methylation and its association with the cold tolerance of an alpine subnival plant, Chorispora bungeana. PLoS ONE 10:e0135485. doi: 10.1371/journal.pone.0135485
Surekha, C., Aruna, L., Hossain, M. A., Wani, S. H., and Neelapu, N. R. R. (2015). “Present status and future prospects of transgenic approaches for salt tolerance in plants/crop plants,” in Managing Salt Tolerance in Plants: Molecular and Genomic Perspectives, eds S. H. Wani and M. A. Hossain (New York, NY: CRC Press), 329–352.
Ueda, H., Kikuta, Y., and Matsuda, K. (2012). Plant communication. Plant Signal. Behav. 7, 222–226. doi: 10.4161/psb.18765
Wang, Z., Casas-Mollano, J. A., Xu, J., Riethoven, J.-J. M., Zhang, C., and Cerutti, H. (2015). Osmotic stress induces phosphorylation of histone at threonine 3 in pericentriomeric regions of Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A. 112, 8487–8492. doi: 10.1073/pnas.1423325112
Wani, S. H., and Gosal, S. S. (2011). Introduction of OsglyII gene into Oryza sativa for increasing salinity tolerance. Biol. Plant. 55, 536–540. doi: 10.1007/s10535-011-0120-9
Wani, S. H., Sah, S. K., Sanghera, G., Hussain, W., and Singh, N. B. (2016). “Genetic engineering for cold stress tolerance in crop plants,” in Advances in Genome Science, Vol. 4, ed Atta-ur-Rahman (London, UK: Bentham Science), 173–201.
Wani, S. H., Singh, N. B., Haribhushan, A., and Mir, J. I. (2013). Compatible solute engineering in plants for abiotic stress tolerance-Role of glycine betaine. Curr. Gen. 14, 157–165. doi: 10.2174/1389202911314030001
Yamaguchi-Shinozaki, K., and Shinozaki, K. (2006). Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. Annu. Rev. Plant Biol. 57, 781–803. doi: 10.1146/annurev.arplant.57.032905.105444
Yuan, W., Luo, X., Li, Z., Yang, W., and Wang, Y. (2016). A cis cold memory element and a trans epigenome reader mediate polycomb silencing of FLC by vernalization in Arabidopsis. Nat. Genet. 48, 1527–1534. doi: 10.1038/ng.3712
Keywords: cold stress, DNA methylation, histone modifications, vernalization genes, cold acclimation, crosstalks
Citation: Banerjee A, Wani SH and Roychoudhury A (2017) Epigenetic Control of Plant Cold Responses. Front. Plant Sci. 8:1643. doi: 10.3389/fpls.2017.01643
Received: 08 August 2017; Accepted: 07 September 2017;
Published: 21 September 2017.
Edited by:
Avinash Mishra, Central Salt & Marine Chemicals Research Institute (CSIR), IndiaReviewed by:
Qing-lin Liu, Sichuan Agricultural University, ChinaSanthi Sri Parankusam, International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), India
Copyright © 2017 Banerjee, Wani and Roychoudhury. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Shabir H. Wani, c2hhYmlyaHVzc2FpbndhbmlAZ21haWwuY29t
Aryadeep Roychoudhury, YXJ5YWRlZXAucmNAZ21haWwuY29t