 Pietro Gramazio1,2†
Pietro Gramazio1,2† Ruth Jaén-Molina3†
Ruth Jaén-Molina3† Santiago Vilanova2
Santiago Vilanova2 Jaime Prohens2
Jaime Prohens2 Águedo Marrero3
Águedo Marrero3 Juli Caujapé-Castells3*Gregory J. Anderson4
Juli Caujapé-Castells3*Gregory J. Anderson4- 1Faculty of Life and Environmental Sciences, University of Tsukuba, Tsukuba, Japan
- 2Instituto de Conservación y Mejora de la Agrodiversidad Valenciana, Universitat Politècnica de València, Valencia, Spain
- 3Jardín Botánico Canario “Viera y Clavijo” – Unidad Asociada al CSIC, Cabildo de Gran Canaria, Las Palmas de Gran Canaria, Spain
- 4Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs, CT, United States
Islands provide unique opportunities to integrated research approaches to study evolution and conservation because boundaries are circumscribed, geological ages are often precise, and many taxa are greatly imperiled. We combined morphological and hybridization studies with high-throughput genotyping platforms to streamline relationships in the endangered monophyletic and highly diverse lineage of Solanum in the Canarian archipelago, where three endemic taxa are currently recognized. Inter-taxa hybridizations were performed, and morphological expression was assessed with a common-garden approach. Using the eggplant Single Primer Enrichment Technology (SPET) platform with 5,093 probes, 74 individuals of three endemic taxa (Solanum lidii, S. vespertilio subsp. vespertilio, and S. vespertilio subsp. doramae) were sampled for SNPs. While morphological and breeding studies showed clear distinctions and some continuous variation, inter-taxon hybrids were fertile and heterotic for vigor traits. SPET genotyping revealed 1,421 high-quality SNPs and supported four, not three, distinct taxonomic entities associated with post-emergence geological, ecological and geographic factors of the islands. Given the lack of barriers to hybridization among all the taxa and their molecular differences, great care must be taken in population management. Conservation strategies must take account of the sexual and breeding systems and genotypic distribution among populations to successfully conserve and restore threatened/endangered island taxa, as exemplified by Solanum on the Canary Islands.
Introduction
Islands have long been recognized as biological laboratories because they provide particularly insightful understanding into the evolution, ecology, and conservation of plant species (Baldwin and Warren, 2010; Caujapé-Castells et al., 2010; Warren et al., 2015; Weigelt et al., 2015; Nogales et al., 2016; Stuessy et al., 2017a; Price et al., 2018). Their geologic ages can also be determined fairly precisely, thereby making it possible to infer approximate colonization times associated with diversification events, including, sometimes, the order of colonization (Curto et al., 2017). When coupled with phylogenies thoroughly sampled geographically and genetically, such methodological advances have led to an improved understanding of tempos and patterns of speciation (Mairal et al., 2015). Furthermore, when the biology of the taxa is well studied, including reproductive systems, the inference of drivers of evolution is possible, and conservation and restoration programs can be developed for populations that are often vulnerable and small (Crawford et al., 2019).
Notably, a number of molecular-based studies have revealed sharp genetic discontinuities within and between islands in both narrowly distributed and widespread Canarian endemic plant lineages (Rumeu et al., 2014; Jaén-Molina et al., 2015; Puppo et al., 2015; García-Verdugo et al., 2017). These results emphasize the value of comprehensive sampling of populations and genomes for a thorough understanding of evolution prior to the implementation of the necessary components of an effective conservation strategy (Caujapé-Castells et al., unpublished; Díaz-Bertrana and Saturno, 2017).
Solanum lidii Sunding and S. vespertilio Aiton are endangered endemic Solanaceae from the Canary Islands that are notable for their suite of reproductive features [(e.g., andromonoecious, self-compatible, heterandrous, sometimes four-parted, always zygomorphic, and sometimes enantiostylous (Anderson et al., 2015)]. Furthermore, molecular genetic studies of these species (Prohens et al., 2007; Caujapé-Castells et al., unpublished) revealed unexpected patterns of variation that have promoted a significant investment in their conservation, particularly on Gran Canaria (Mesa-Coello et al., 2013).
Solanum lidii (hereafter referred to as SL) is endemic to Gran Canaria, where it grows in ruderal shrublands on disturbed sites and rocky areas as inland cliffs (Rodríguez-Delgado et al., 2004) at between 350 and 725 m. In 2004, the total size of SL in the wild was estimated at only 93 individuals, distributed among six isolated and severely fragmented subpopulations in five locations. The area around Temisas, in the southeast of Gran Canaria (Figure 1) is the largest but includes only 32 individuals (Rodríguez-Delgado et al., 2004). Intrusive human development, changes in land use, collapse of terrain (or landslides) and herbivory (often by goats) are the main threats to the continued existence of this species (Rodríguez-Delgado et al., 2004). SL is listed under the category “Critically Endangered” by the IUCN criterion [CR B2ab (ii, iii)]; C2a(i) in the Spanish Red List 2008 (Moreno, 2008), and as a species in danger of extinction on the Spanish national and regional catalogs of threatened species1. Among the proposed measures to reduce extinction risks for SL are the construction of fences and the local elimination of goat grazing.
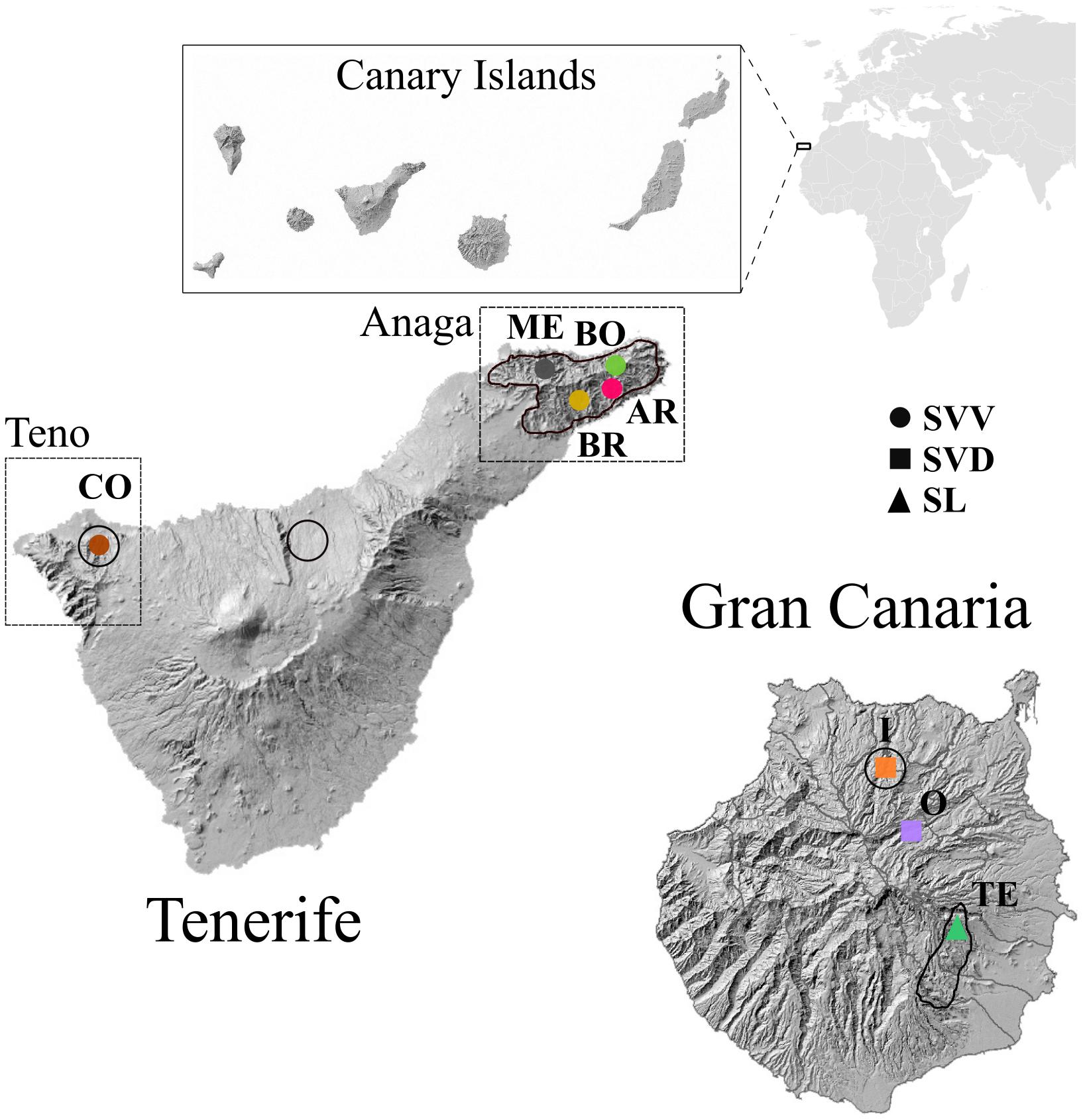
Figure 1. Wild distribution of the three Canarian endemic Solanum taxa in Tenerife and Gran Canaria highlighted in the map with a black line, following Rodríguez and Garzón (2019) and https://www.biodiversidadcanarias.es/. Populations from the same taxa are represented with the same shape but different colors, following codes as in Table 1.

Table 1. Samples and population details of the Solanum taxa studied: S. vespertilio subsp. vespertilio (SVV), S. vespertilio subsp. doramae (SVD), and S. lidii (SL).
Solanum vespertilio includes two subspecies, S. vespertilio subsp. vespertilio (hereafter SVV) and S. vespertilio subsp. doramae (hereafter SVD) Marrero Rodr. & Gonzalez-Martin. SVV is restricted to Tenerife, the largest of the seven main islands in the Canarian archipelago, where the greatest concentration of individuals occurs in two areas: the palaeo-islands of Teno (in the northwest; a dryer habitat) and Anaga (in the northeast; a moister habitat) (Figure 1). While only one population is found in Teno, the Anaga area includes at least five populations scattered throughout several ravines. The main threats to this subspecies are competing exotic species, goat herbivory and parasitism. It is included as “Vulnerable to changes in its habitat” in the Canarian Catalogue of Endangered Species (2001) and considered as “Critically endangered” by the IUCN [CR B2ab (iii, v)] in the Spanish Red List 2008 (Moreno, 2008). Proposed conservation measures include local elimination of goat grazing and supplemental planting within the current distribution, and in other similar habitats.
SVD is a very rare, and possibly extinct in the wild, endemic of Gran Canaria. It was cited first from just two localities by Webb and Berthel (1845); Kunkel (1975a) considered it extinct. However, a population of around 15 individuals was rediscovered in “Barranco de Azuaje” (Marrero, 1986). Based on morphological and ecological differences, the Gran Canaria individuals were recognized as a new subspecies: S. vespertilio subsp. doramae (Marrero and González Martín, 1998). Since 1987, four censuses (Marrero and González Martín, 1998 and SEGAs 2002, 2005, and 2006) have reported the decline of SVD, first to six and later to only 2–3 mature plants (Santana and Naranjo, 2002; Docoito-Díaz and Herrera, 2005; Cabrera et al., 2006). The decline to near-extinction of SVD was mainly due to agriculture, including grazing, and to the competition with introduced exotics (e.g., Agave americana and various Opuntia, Bañares et al., 2003). In 2000, SVD was included under the category “endangered” in the National Catalogue of Threatened Species and the following year, in the Canarian Catalogue of Endangered Species. In 2008, the Canarian Government approved a recovery plan for this subspecies.
In the most recent molecular study of the relationships among these three taxa, Caujapé-Castells et al. (unpublished data) assessed five cpDNA sequences (matK, rbcL, trnT-L, trnK-rps16, and trnF-L) and ISSR markers. That study, conducted on limited population sampling (five individuals of each of SVV and SVD and three SL), aimed to quantify the genetic differentiation among the three taxa to assess the potential for reintroduction of the threatened SVD in its natural habitats on Gran Canaria. Despite the relatively high levels of polymorphism detected with both molecular techniques, the barcode sequences (matK and rbcL) did not reveal distinction between the two subspecies of S. vespertilio, and only one position in trnL-F discriminated S. lidii from S. vespertilio. These results reinforced the close phylogenetic relationship among the taxa highlighted in the AFLP analysis by Prohens et al. (2007) but also indicated the need for future multi-disciplinary studies that included molecular approaches with greater discriminatory power.
Single nucleotide polymorphisms (SNPs) are among the most abundant and easy to automate molecular markers (Thomson, 2014; Kim et al., 2016). As a result of the tremendous advances in next-generation sequencing (NGS) platforms and the decrease in sequencing costs, it has become easier to identify hundreds of thousands or even million SNPs in a short time and at a reasonable cost (Scheben et al., 2017). Recently, the first SNP high-throughput genotyping platform (specifically designed for eggplant and its wild relatives, Barchi et al., 2019a) was developed, derived from the resequencing of seven S. melongena and one S. incanum accessions (Gramazio et al., 2019). The eggplant SPET platform is based on 5,093 target SNPs distributed throughout the whole eggplant genome. Barchi et al. (2019a) pointed out the suitability and transferability of the eggplant SPET platform to closely related species as a result of the discovery of novel SNPs in the surrounding sequenced regions of the target probes. Particularly relevant for this study, the threatened taxa at issue are closely related to the eggplant group (Prohens et al., 2007).
Traditional and new molecular tools have been revealed as powerful and indispensable aids to conservation efforts, particularly with endangered species (Corlett, 2017). Herein, we combine data from traditional studies of morphological expression in common garden experiments with experimental plant hybridizations, and the latest DNA technology (via high-throughput SNP genotyping using the eggplant SPET platform), to substantiate conservation strategies for rare and endangered island plants. We believe our multi-faceted approach will help clarify the possible genetic origin and stability of the morphological characters separating these taxa at a level not previously possible (Prohens et al., 2007; Särkinen et al., 2013). The main aims of this study were to (i) study the morphological differences of the Canarian endemics S. lidii and S. vespertilio s.l., and their crossability, (ii) provide new data to assess their genetic relationships and (iii) suggest effective conservation strategies for these endangered taxa, that include the extinct-in-the wild S. vespertilio ssp. doramae.
Materials and Methods
Plant Material
A total of 74 individuals from the three Solanum taxa were sampled (Table 1). Leaves were collected during 2018 and dried and conserved in silica gel for subsequent analysis. The bags with dried leaves were deposited in the DNA Bank of the Canarian Flora at Jardín Botánico Canario “Viera y Clavijo” – Unidad Asociada al CSIC of the Cabildo de Gran Canaria (JBCVCSIC hereafter) and vouchers for each taxon and population were collected and deposited at the LPA herbarium at JBCVCSIC. Samples were collected in eight localities that encompass their main wild areas of distribution in Gran Canaria and Tenerife, save for SVD, for which currently only cultivated populations are known. For SL, we sampled only one of the five known localities because this species is morphologically so obviously distinct from both S. vespertilio subspecies that larger samples were not needed.
Morphological Analyses, Hand-Manipulated Interspecific Crosses, and Viability Assessments
The plants used for the scoring or measurement of characters noted as key differences among the species (Table 2) depended on the character and depth of analysis. For instance, overall features scored for Table 2 included both field-collected data (e.g., seed/fruit) and greenhouse samples (see Figure 2). Leaf features, in particular, the length, width and length/width ratio, have been important in distinguishing the subspecies of S. vespertilio. Thus, leaf variation was studied more intensively in larger populations of plants grown from wild-collected seed (or the JBCVCSIC seedbank for SVD) in “common gardens” (research greenhouses at the University of Connecticut), and in hand-generated hybrids between all possible pairwise combinations of SVV, SVD, and SL. Measurements were taken on the topmost mature leaves on mature plants (i.e., several months to year-old plants), and included length of the blade from tip to the lowest point on the basal lobes of the blades (i.e., not including the petioles); the width was taken at the widest point. The length/width ratio was then calculated.
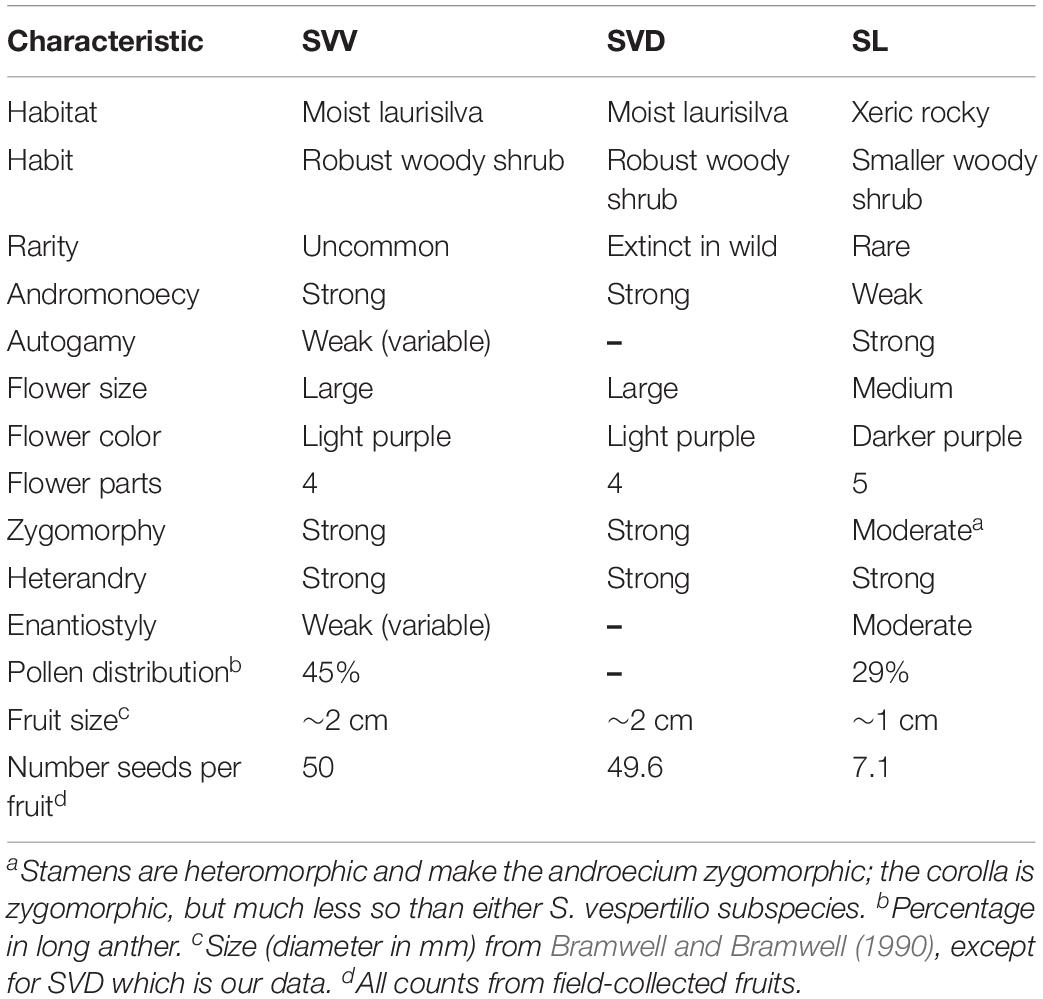
Table 2. Characterization of the three Solanum Canarian taxa.

Figure 2. Flower structure (drawings and photos), leaves, fruits, and plants of the Solanum taxa endemic from the Canary Islands: S. vespertilio spp. vespertilio (SVV), S. vespertilio spp. doramae (SVD), and S. lidii (SL). Photos by G. Anderson, Á. Marrero, R. Jaén-Molina, and Óscar Saturno.
Artificial hybrids were generated between all three possible pairwise combinations: SVV with SVD, SVV with SL, and SVD with SL (female parents listed first in each case; see details of crossing techniques in Anderson and Levine, 1982). Seeds from successful crosses were harvested from ∼4-month-old fruits and grown for 2–4 years in the research greenhouses. We used at least 16 successful artificial hybrid “families” (i.e., the multiple seeds grown from a particular hybrid combination). For each hybrid combination, there were usually several “duplicate families” (i.e., different plants of the same taxon involved in generating that hybrid combination). The vouchers representing the parental taxa used, and hybrids generated, are in CONN (The University of Connecticut herbarium collections). We used a total of 16 hybrid families including 67 genets (separate genetic individuals) within them, from which measurements were taken (Table 3). For the parents, we used 15 genets of SVV, 8 genets of SVD and 13 genets of SL. Each parental genet was a distinct individual, but they may have been derived from seeds grown from the same fruit. Two-tailed Student’s t-tests were used to test for significant differences between various pairs in Table 3. Artificial hybrids were evaluated for overall growth and vigor, and then specifically for fertility by via pollen viability assayed via staining pollen grains with aniline blue in lactophenol (Anderson and Symon, 1989).
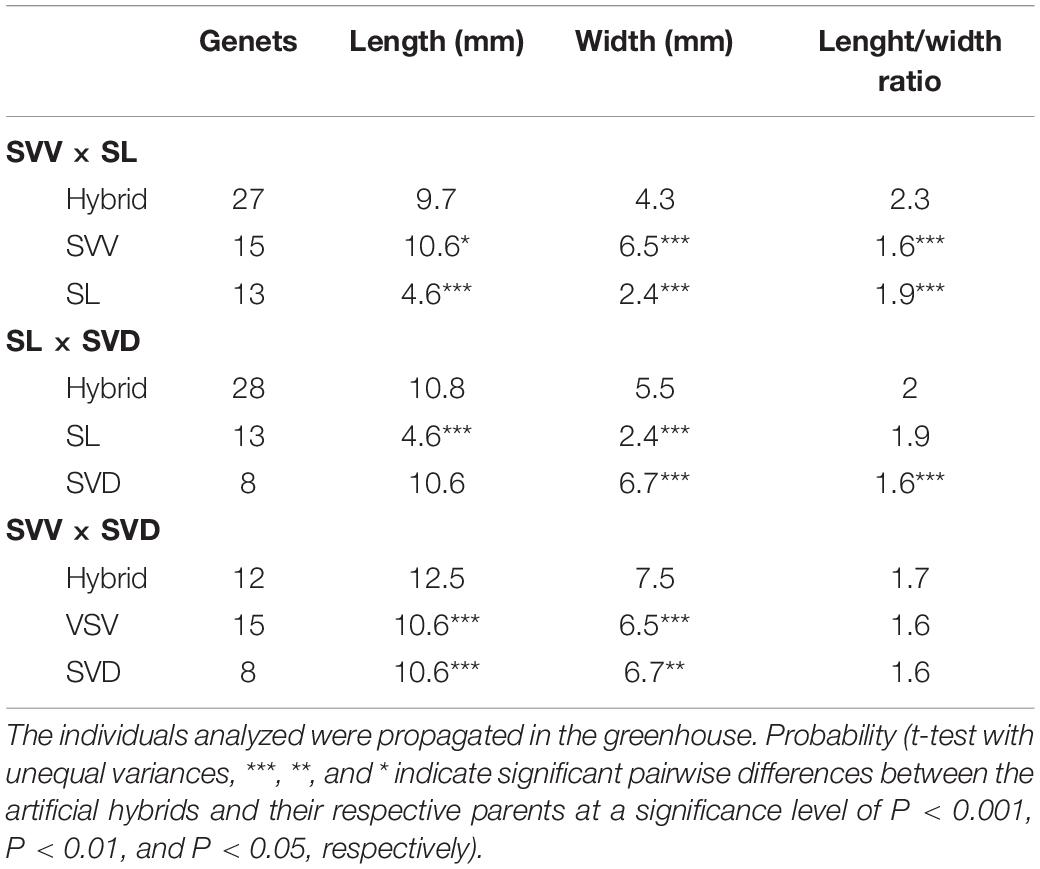
Table 3. Leaf morphology characterization of the three Solanum Canarian taxa.
DNA Extraction, Library Preparation, and Sequencing
Genomic DNA was extracted from freeze-dried leaves using an in-house DNA extraction protocol developed for NGS applications (Alonso et al., 2018). DNA quality and integrity were evaluated using a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, United States) checking 260/280 and 260/230 nm ratios and by agarose electrophoresis. DNA concentration was estimated with Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, United States). Samples were diluted and sent to IGA Technology Services (IGATech, Udine, Italy) for genotyping using the eggplant SPET platform, where genomic libraries were constructed and sequenced using an Illumina NextSeq 500 sequencer in single-end mode (150 bp). The eggplant SPET platform was designed using the SNP dataset (over 10.9 million SNPs) generated from the whole-genome resequencing of seven S. melongena and one S. incanum accessions (Gramazio et al., 2019) and selecting the 5,093 most reliable SNPs, 3,372 of them in CDSs and 1,721 in introns and UTRs regions, by applying stringent filter criteria and running pilot tests (Barchi et al., 2019a).
Reads Processing, Mapping, and SNP Calling
After sequencing, the base calling and demultiplexing steps were performed using a standard Illumina pipeline, while read quality check and adapter trimming were accomplished using ERNE (Del Fabbro et al., 2013) and Cutadapt (Martin, 2011) software, respectively. Subsequently, reads were mapped to the reference genome “67/3” (Barchi et al., 2019b) using BWA-MEM aligner (Li, 2013) with default parameters and only uniquely aligned reads (i.e., reads with a mapping quality > 10) were selected). Variant calling was performed following the Germline short variant discovery (SNPs + Indels) workflow2 using the GATK 4.0 software (Depristo et al., 2011). Specifically, the variants were called per-sample by the HaplotypeCaller (Poplin et al., 2017) to generate an intermediate GVCF (Genomic Variant Call Format) file for each sample. The GVCFs files from multiple samples were consolidated into a GenomicsDB datastore to improve scalability and gathered together to the GenotypeGVCFs tool for a cohort-wide joint-called genotyping analysis. Reduction of the number of false positives was performed with the SelectVariants and VariantFiltration tools to apply stringent filter criteria (QD < 2.0, MQ < 40.0 and MQRankSum < −12.5). To select only highly reliable SNPs, we applied stringent criteria to the VCF file using VCFtools (version 0.1.15) (Danecek et al., 2011) with the following parameters: min-meanDP 30, minDP 30, max-missing 0.95 and maf 0.02. After checking the reliability and efficiency of the SPET technique (i.e., no polymorphisms between the two technical replicates of a sample), we decided to remove duplicates for the subsequent analyses.
Molecular Data Analysis
To analyze and manipulate the high-throughput genotype data, the VCF file was uploaded to R (Ihaka and Gentleman, 1996) using the vcfR package (Knaus and Grünwald, 2017) and the data transformed to genlight and genind objects using the function vcfR2genlight of the R package Adegenet (Jombart, 2008). Basic statistics were estimated using the VCFtools software (Danecek et al., 2011) and the function basic.stats of the R package hierFstat (Goudet, 2005) based on the analysis of gene diversity proposed by Nei (1973). Confidence intervals for population−specific Fis were estimated using the boot.ppfis function in hierfstat by performing 10,000 bootstrap replicates over loci. Pairwise Fst values among populations were calculated for populations with four or more individuals (n ≥ 4) (Willing et al., 2012). To clarify the complex relationships among the accessions from the three endemic Canarian Solanum taxa and provide insight into their population structure, we carried out complementary analyses. First, a hierarchical clustering based on pairwise similarities was performed to assess the genetic relationships among the 74 accessions. A consensus UPGMA (unweighted pair group method with arithmetic mean) dendrogram based on Nei (1972, 1978) genetic distance was obtained using the function about of the R package poppr (Kamvar et al., 2014). Branches of the dendrogram were tested with bootstrap support for Bruvo’s distance (Bruvo et al., 2004) with 1,000 replications. The dendrogram was graphically plotted using the function plot.phylo of the R package ape (Paradis and Schliep, 2018). To confirm the reliability of the UPGMA dendrogram, a SVDQuartets analysis was performed in PAUP∗ program (Swofford, 2003). The SVDquartets method (Chifman and Kubatko, 2014, 2015) is a coalescent-aware phylogenetic method based on nucleotide sequence data that can identify the correct species tree in the face of deep coalescence leading to incomplete lineage sorting (ILS). ILS can lead to artifacts in standard methods/models when there are short branches anywhere in the tree and effective population sizes are large enough that sister lineages do not coalesce before (looking backwards in time) their common ancestor. To infer the population structure and detect additional sub-populations, a fastSTRUCTURE analysis, which is based on a variational Bayesian approach, was performed (Raj et al., 2014). To convert the data from the VCF file to the fastSTRUCTURE format, the software PGDSpider (Lischer and Excoffier, 2012) was used. A principal component analysis (PCA) was performed using the function glPCA of the package Adegenet (Jombart, 2008), and represented graphically using the package ggplot2 (Wickham, 2016).
Molecular Data Access
The sequence data have been deposited into the NCBI Short Read Archive under the Bioproject identifier PRJNA556343. Raw reads of each accession are deposited under the accession numbers from SRX6649376 to SRX6649471. VCF files with the corresponding variants identified are available upon request to the corresponding author.
Results
Analysis of Morphology and Pollen Stainability
Both subspecies of S. vespertilio are distinct from S. lidii in terms of leaf length and width, but less distinct in terms of the length/width ratio (Table 3). Analyses of the artificial hybrids resulting from hand-generated crosses between SL and either parental subspecies shows that they are mostly distinct as well, with some features (e.g., leaf length and width) similar to one species or the other, but with some hybrids manifesting intermediate characters or heterosis. The hybrids between the two subspecies of S. vespertilio are also distinct in leaf length and width, but not in the length/width ratio. All three taxa and all the hybrid combinations were, on average, fully fertile, with pollen staining percentages averaging from 80 to 95%. Thus, none of the taxa, nor the artificial hybrids between them, manifested sterility.
High-Throughput Sequencing and Mapping
After the sequencing, cleaning and trimming steps, over 5 billion bp were generated in the 74 accessions used in this study, which correspond to over 33 million clean reads of 150 bp (Supplementary Data S1). Due to the different quality of the plant material (some samples were stored for a longer time than others), the number of sequenced reads per sample varied considerably among the accessions with an average of 446,472 clean reads per sample (range from 124,841 of SL_TE_9 and to 1,801,500 of SVV_CO_2). Of those reads, on average, 95.3% were mapped onto the eggplant reference genome “67/3” (Barchi et al., 2019b). The average of the mean accession coverage considering the target regions in each accession was 78.8 reads, ranging from 22.0 for sample (SL_TE_9) to 316.8 reads for sample (SVV_CO_2); thus, deep and reliable large enough for most of the bases sequenced to be mapped.
SNP Calling and Population Structure Statistics
A total of 6,504 SNPs were identified using the 5,093 probes of the eggplant SPET platform (Table 4). The distribution of the SNPs identified along the chromosomes was uneven and ranged from 336 of chromosome 11–951 of chromosome 1 with an average of 542 SNPs per chromosome. Surprisingly, there were only 284 targeted SNPs identified (i.e., SNPs selected for the SPET platform), around 5.6% of the total set, while there were 6,220 untargeted SNPs (i.e., novel SNPs identified in the surroundings of the targeted SNPs), almost 22-fold more than the targeted SNPs. On the other hand, even though the average of the mean probe coverage was 86.9 reads, some of the probes showed low coverage (Supplementary Data S2), so that stringent criteria were applied to retain only the most reliable SNPs. After filtering, a total of 1,421 high-quality SNPs were selected and used for the subsequent analyses. Only 34 SNPs were targeted SNPs (0.67% of the SPET set), while there were 1,387 novel SNPs, over 40-fold greater than the targeted SNPs (Table 4).
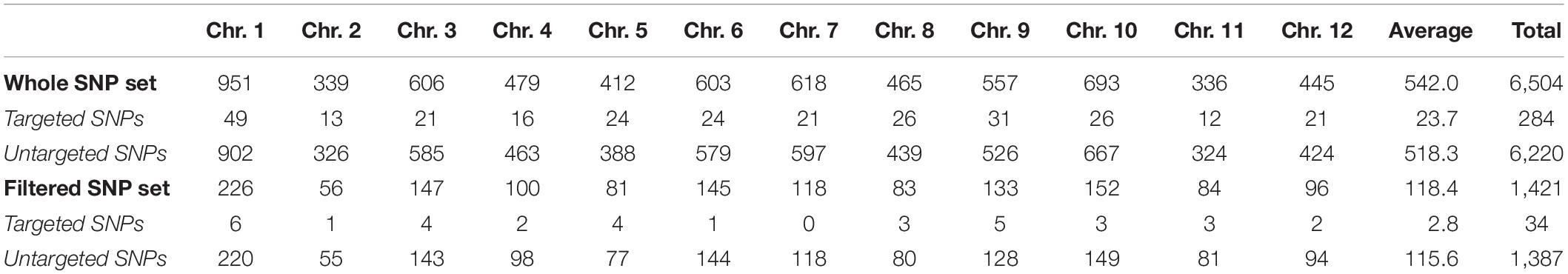
Table 4. SNP distribution per chromosome identified among the 74 accessions from the three Solanum Canarian taxa.
Most of the SNPs identified were biallelic and only 13 presented more than two alleles. The average of the reference allele frequency (i.e., the same as the reference genome) was 0.782 with a wide range from 0.014 to 0.993 (Supplementary Data S3). Overall, the number of heterozygous SNPs was low, as reflected in the low average value for the observed heterozygosity per locus (Ho; 0.113), in the average within-population gene diversity per locus (Hs; 0.110) and in the average total population gene diversity per locus (Ht; 0.198), respectively. As well, the average inbreeding coefficient (Fis) per locus was 0.085. However, the average fixation index per locus (Fst) was 0.331, indicating moderate to a high level of differentiation due to the members of some populations carrying unique alleles compared to the rest. On the other hand, the average gene diversity (Dst) per locus among subpopulations was low (Dst = 0.088), and ranged from 0 to 0.403, similar to the average measure of population differentiation (Dest) per locus (Dest = 0.116), which ranged from 0 to 0.510. Population SL_TE showed the highest average observed heterozygosity (Ho) per locus with a value of 0.230, followed by populations SVV_BR (0.134), SVV_AR (0.128), and SVV_ME (0.121); the lowest value was observed for SVD_O (0.039) (Table 5 and Supplementary Data S4). A similar pattern was found for gene diversity, where SL_TE showed the highest average value per locus (0.208) and population SVD_O the lowest (0.022). For the average Fis, populations SVV_BO and SVV_ME showed the only average positive values (0.170 and 0.090, respectively), while population SVD_O presented the most negative average values with −0.798. Individual values for all the accessions for Ho, He, and Fis are reported in Supplementary Data S5. The estimated Fst values between pairwise populations displayed low values among SVV populations, ranging from 0.106 between SVV_ME and SVV_BR to 0.296 between SVV_AR and SVV_CO (Table 6). The values comparing the geographically most separate SVV_CO (from Teno, on Tenerife) with other SVV populations were consistently the highest. Fst values between SVD_O and SVV populations were even higher, apart from the comparison with SVV_BO (0.109). Not surprisingly, the SL_TE pairwise values were generally uniformly higher among all the comparisons, interestingly the highest for the comparison with SVV_CO (0.431).
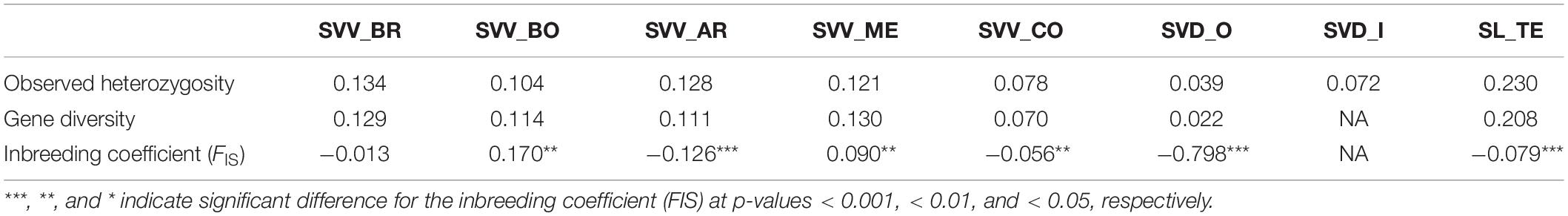
Table 5. Population structure statistics for eight populations from the three Solanum Canarian taxa. Population codes as in Table 1.
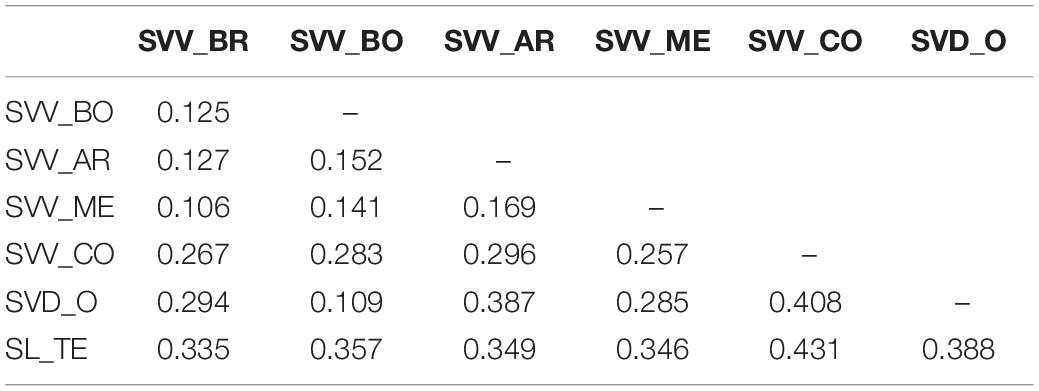
Table 6. Pairwise Fst estimates between Solanum vespertilio subsp. vespertilio, S. vespertilio subsp. doramae, and S. lidii populations with number of individuals (n ≥ 4).
Private and Discriminant SNPs
One of the main aims of this study, apart from establishing genetic relationships among the accessions assessed, was to provide genetic tools to easily discriminate the three taxa. As expected, given the larger morphological distinction, the largest set of private SNPs (i.e., SNPs specific to a particular taxon or population) was found for SL with 55 SNPs that discriminate it from the two subspecies of S. vespertilio (Supplementary Data S6). Private SNPs for SL were found to be distributed across all chromosomes except in chromosome 5, although the number of SNPs in chromosomes carrying private SNPs varied considerably, ranging from chromosome 1 with 17 SNPs to chromosome 11 with just one SNP. However, not all the private SNP loci provide the same discriminatory power because some of them shared SNP alleles with some accessions of the two subspecies of S. vespertilio. The most informative and reliable private SNPs, i.e., those represented by the same homozygous alleles for all the individuals of SL (e.g., A/A), and different homozygous alleles for the rest of the accessions of the two subspecies of S. vespertilio (e.g., C/C), were selected to create a subset called “Core private SNPs.” This subset presented 36 SNPs with five SNPs on chromosomes 3, 7, 10, and 12, four SNPs on chromosomes 1 and 6, three SNPs on chromosomes 4, two SNPs in 2 and 8 and just one SNP on chromosome 11, while chromosomes 5 and 9 did not bear any SNPs. Some of the private SNPs in the Supplementary Data S6, like other SNPs in the full set of SNPs, can be considered as potential haplotypes because they are physically very close and probably are tightly linked and inherited together. However, all the analyses were conducted using SNPs as a single information unit and not as haplotypes. For the two subspecies of S. vespertilio, only one private SNP was found for SVV and none for SVD. However, for SVD, when the accession (SVD_I_1) is removed from the analyses, seven private SNPs were identified between SVV and SVD, although none of them was specific and universal and therefore did not meet the requirement for being included in the “Core private SNPs” subset.
Population Structure and Genetic Relationships Analysis
The UPGMA-based dendrogram obtained for 74 accessions revealed a sister basal clustering of all SL individuals, although the relationships among them were not always well supported by bootstrap values (Figure 3). Interestingly, the SVD accession from Moya (SVD_I_1) was placed outside of and as basal sister to the rest of the accessions of SVD, and close to the SL accessions. The fastSTRUCTURE analysis (Figure 3) also points to the connection of this SVD to SL, in that it is the only SVD that shows a genetic admixture with SL. The remainder of the SVD accessions (from Teror, SVD_O1-4) nested in the middle of the SVV accessions from the northeastern populations on Tenerife (the Anaga peninsula). The SVV samples from Teno clustered together in a separate branch from all the accessions of SVV collected in Anaga. This Teno group is as distinct from the rest of the subspecies SVV as the single Moya accession of SVD (SVD_I_1) is from all the other SVV (and SVD) accessions. Except for a few samples that were intermingled in other groups, the four populations from Anaga constituted distinct individual clusters.
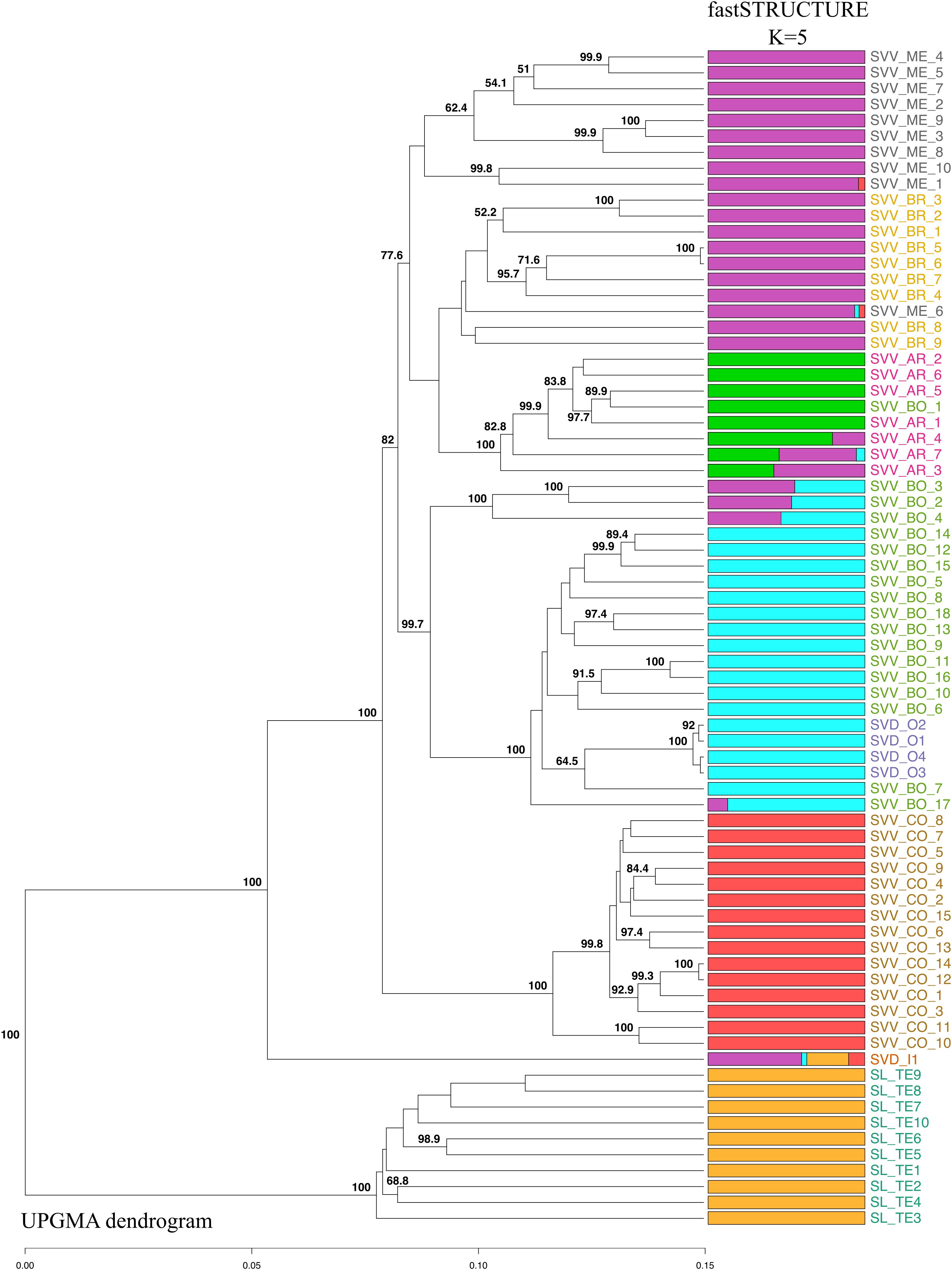
Figure 3. Genetic population structure of the 74 accessions from the three Canarian endemic Solanum taxa according to an UPGMA-based dendrogram and a fastSTRUCTURE (K = 5) analysis with 1,421 SNPs. Bootstrap values greater than 50%, based on 1,000 replications, are indicated in percentage at the corresponding nodes. Each accession is represented by a horizontal bar that is partitioned into colored segments corresponding the estimated membership of an accession to one of the five specific inferred clusters. Population codes as in Table 1 and colors as in Figure 1.
Although there were some minor rearrangements in branch disposition, the two different methods for estimating species trees (UPGMA and SVDquartets), gave the same topology of accessions, confirming the results of the UPGMA analysis that we show herein, and suggesting that there are no significant artifactual groupings due to deep coalescence (Supplementary Data S7). The fastSTRUCTURE analysis suggested that the optimal number of genetic clusters was obtained at K = 5. Only two clusters (red and yellow) consisted mainly of the individuals from a single population (or species) (Figure 3). In fact, individuals from SL and those from Teno SVV_CO showed perfect intrapopulation uniformity with apparently no indications of hybridization or migration from other groups. This pattern, where all individuals of one population were linked in one part of the diagram, was also observed for SVD_O and SVV_BR, although, in this instance, they shared clusters with other populations SVV_BO in the case of SVD_O and SVV_ME for SVV_BR. This pattern suggests that some event led to a sharing of alleles and genetic structure. However, while in the case of SVV_ME for SVV_BR geographical proximity might have facilitated the contact, in the case of SVV_BO (from Tenerife) and SVD_O (from Gran Canaria) the two populations are from islands separated by 80 km. On the other hand, SVV_BO and SVV_AR showed less uniformity with fragments from three different clusters. Overall, the multivariate PCA analysis bolstered the results of the dendrograms, although some groups were nested (Figure 4A). The first principal coordinate, which explained 30% of the genetic variation, clearly separated the SL accessions and the SVD_I_1 accession from the rest, while the second coordinate, which accounted for 12% of the variation, separated the SVV Teno accessions from the rest of SVV accessions from Tenerife (notably also including all the SVD accessions except the single SVD Moya accession (SVD_I_1). To enhance resolution among the SVV groups, an additional PCA analysis was performed on data that did not include the SL accessions (Figure 4B). The first axis, which accounted for 20% of the genetic variation, divided the accessions from Teno from the rest of the S. vespertilio populations, while the second axis, constituting 17% of the genetic variation, strongly separated the SVD_O (Teror) accessions from the SVD_I_1 (Moya) accession. Furthermore, this second coordinate separated almost all the SVV accessions from Lomo de Las Bodegas SVV_BO from the rest of the Anaga SVV populations SVV_BR, SVV_AR and SVV_ME. The latter showed a strong admixture pattern.
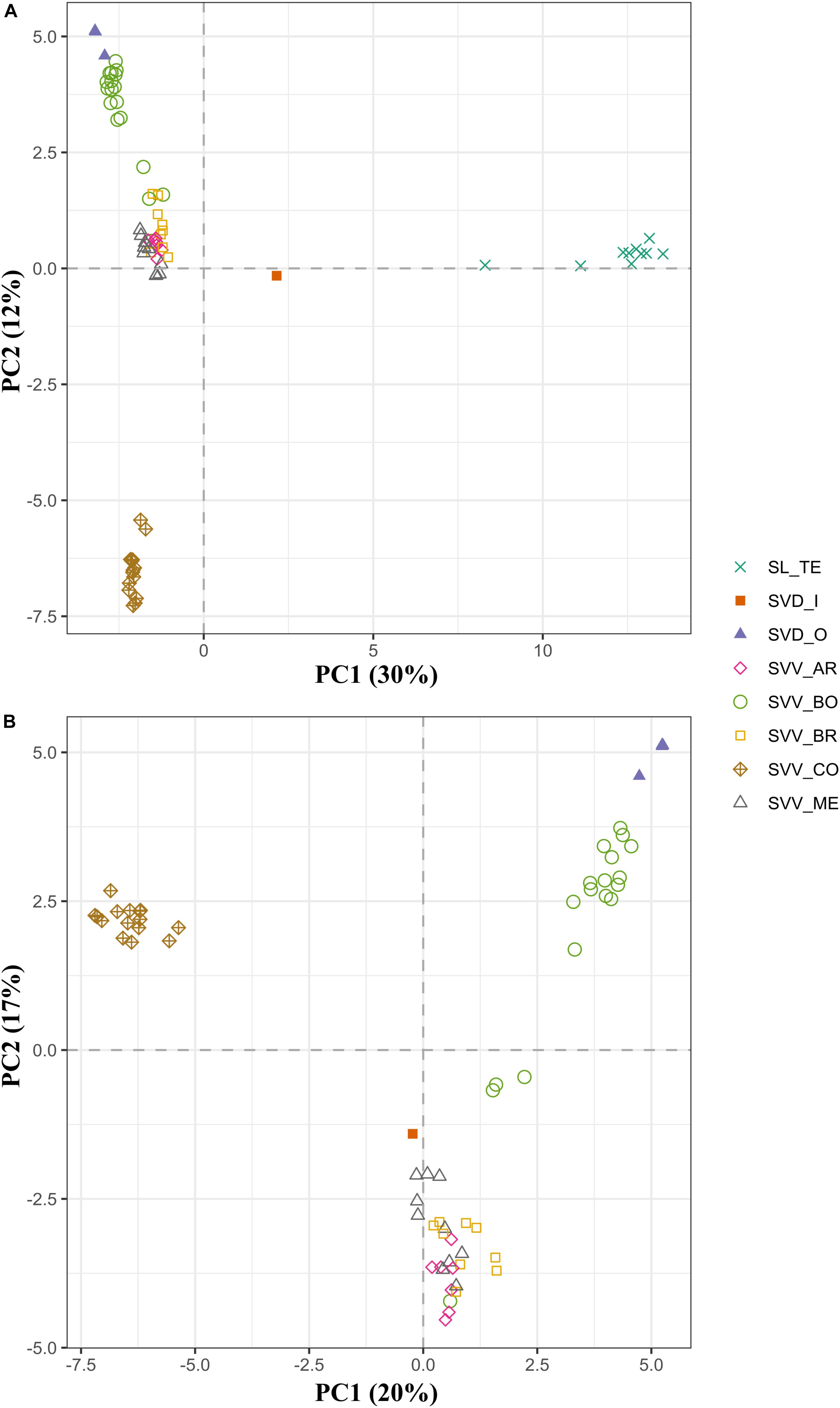
Figure 4. Principal coordinates analysis (PCA) similarities based on the whole set of SNPs (1,421) for the 74 accessions from the three endemic Solanum taxa in the Canary Islands (A) and on partitioned data without S. lidii accessions (B). The first and second principal coordinates are displayed. Population codes as in Table 1 and colors as in Figure 1.
Discussion
High-Throughput Genotyping SPET Platform
The eggplant SPET platform has been reliably applied to species closely related to eggplant (Barchi et al., 2019a), and this investigation shows that it is transferable to S. lidii and S. vespertilio, two rare Canary Island endemics much more distantly related to the eggplant. Therefore, this study provides additional proof of the reliability of this technology as well as of its wide cross-species transferability. This new genotyping solution combines the benefits of (a) complexity reduction (e.g., high-throughput and cost-effective), (b) high-density DNA arrays (e.g., targeted and reproducible), and (c) discovery of novel polymorphisms in addition to the target SNPs in regions of interest, thus allowing high-transferability across species (Scaglione et al., 2019). The number of SNPs identified in this investigation (6,504) was considerably higher than the number of the probes designed (5,093). Furthermore, only 284 polymorphisms were targeted SNPs versus the 6,220 novel SNPs detected in the 74 accessions. Using single-end sequencing of 150 bp, after removing 40 bp of the probe, up to 110 bp per read have been mapped onto the reference genome (Barchi et al., 2019b); this allowed us to screen the flanking regions of the targeted SNP for novel and private SNPs. In addition, this strategy allows the detection of potential haplotypes among populations as well as SNPs in linkage disequilibrium (Scaglione et al., 2019). An additional benefit of the high reproducibility of SPET is that it could allow data to be merged from independent studies that use the SPET platform (with the same configuration of probes) to facilitate much broader-based phylogenetic meta-analyses. Ultimately, novel analytical technologies like SPET (Barchi et al., 2019a; Scaglione et al., 2019), provide great strength in a range of studies, including those like this where our goal is to understand island colonization patterns, evolution and suggest conservation strategies for rare taxa.
Relationships Among Taxa and Tentative Hypotheses on Their Evolution on Islands
Solanum lidii and S. vespertilio are clearly separated by morphology and molecular data. Interestingly, the hybrids between them are intermediate and fertile. Särkinen et al. (2013) synthesis has given high support to that the species S. lidii and S. vespertilio are derived from common ancestors of the African Solanum subgenus Leptostemon taxa. Also, Särkinen et al. (2013) suggest that the common ancestor of the Canarian Solanum taxa split from African lineages about 1.68 Myr (though there is a wide standard error associated with the age estimates, see Supplementary Material). Assuming that this estimate represents the stem age of Solanum in the Canaries (i.e., the colonization of the archipelago by the ancestor of the current endemics), it would mean that these taxa are recent endemics on Gran Canaria and Tenerife, in spite of the fact that these two large islands are among the older in the archipelago (they emerged 15 and 12 Myr respectively; Van Den Bogaard, 2013).
The date of first colonization of the Canaries by Solanum is unknown, but it may have been even later, closer to the suggested date at which S. lidii and S. vespertilio became distinct lineages (i.e., 0.93 Myr; Särkinen et al., 2013). From present data, we cannot be sure whether that common ancestor migrated to the Canaries or it split in Africa yielding the ancestral forms of S. lidii and S. vespertilio prior to colonization. The cladogram (Figure 3) also cannot answer the question of whether the common ancestor was more similar to what we now recognize as S. lidii or S. vespertilio. However, the ecology and habitat preference of the proposed extant closest relatives (African species: S. capense, S. tomentosum, and S. humile; Särkinen et al., 2013) suggest a hypothesis: the continental African taxa occupy similar habitats to that of S. lidii that are drier, often disturbed, open woodland, sandy or rocky places (Vorontsova and Knapp, 2016)3. If the common ancestor to the Canarian taxa was similar, then the descendants of S. lidii could have grown first in these more xeric habitats on both Gran Canaria and Tenerife, and then radiated into the more mesic-moist scrub land yielding the selection that gave the larger-leaved, more robust, less-tomentose vespertilio-like plants. Similar patterns are reported for the colonization of Hawaii by Baldwin and Warren (2010). However, the lack of any evidence for S. lidii on Tenerife at any time argues against it occurring there.
Solanum vespertilio may have split with S. lidii from the common African ancestor and adapted independently to the more mesic habitats on both islands. If the split from the common ancestor that yielded the two taxa occurred after Canarian colonization, this would suggest that S. vespertilio occurred first on Gran Canaria and then underwent short-distance dispersal (∼80 km) to Tenerife, where it proliferated in the more common moist habitats. In this scenario, S. vespertilio subsp. doramae (today virtually extinct-in-the-wild) can be construed as one remnant of that split and further radiation into what are today increasingly uncommon undisturbed moist habitats on Gran Canaria. The position in the tree (Figure 3) of SVD_I (a descendant of individuals of SVD collected from the wild in Moya) supports this hypothesis, in that it is intermediate between S. lidii and the other SVV and SVD accessions. The molecular profile of the other SVD accessions firmly embeds them within the Tenerife SVV accessions. Consistent with data from morphological analyses and hybridization studies, these genotypically and morphologically indistinct SVD accessions are compatible with the conclusion that there may have been contemporary natural hybridization between typical S. vespertilio subsp. vespertilio and the Gran Canarian S. vespertilio subsp. doramae. Alternatively, the original samples may have already undergone rounds of hybridization (see the “conservation implications” section below).
The SVV accessions are distributed in Anaga and Teno massifs (Figure 1), with estimated geological ages of respectively 3.9–4.9 Myr and 5.6–6.2 Myr, and therefore much older than the central massif of Tenerife (Carracedo and Day, 2002). At least 13 endemic plant species share with SVV this disjunct distribution between the West (Teno) and East (Anaga) of Tenerife (Moya et al., 2004; Trusty et al., 2005; Mairal et al., 2015). The geographical gap between these two ends of Tenerife is clearly reflected in the tree (Figure 3) and in the PCA (Figure 4). This pattern of genotype distribution is similar to those reported in phylogeographic investigations of the Canarian flora and has been detected (to different extents) in either widespread or more narrowly distributed Canarian plant endemics belonging to disparate lineages, like Canarina canariensis (Campanulaceae, Mairal et al., 2015), Dorycnium broussonetii (Fabaceae, Jaén-Molina et al., 2015), Ruta pinnata (Rutaceae, Soto and Jaén-Molina, unpublished data), or Navaea phoenicea (García et al., 2009).
Not all the lineages with an Anaga-Teno distribution are old. Northern Tenerife has been subjected to a number of geologic events such as volcanic landslides and avalanches in different periods of time from 2.2 Ma (Tigaiga landslide) to 170 ka (formation of Las Cañadas Caldera) (Ancochea et al., 1990; Watts and Masson, 1995; Ablay and Hürlimann, 2000) that likely have caused recent disruptions of gene flow in taxa that formerly constituted a continuum of populations. For instance, the populations of Bystropogon (Trusty et al., 2005), and some populations of Micromeria (Puppo et al., 2015) or Canarina canariensis (Mairal et al., 2015), presently distributed in Teno and Anaga may be the remnants of a larger ancestral distribution area affected by these volcanic events. In the case of Navaea phoenicea (Malvaceae), a palaeo-endemic with a similar distribution to SVV, González De (2016) finds modest levels of gene flow mediated by pollinators among Anaga and Teno. Our genetic data in Solanum do not provide support for current gene flow between these widely separated areas, so that the separation between the distribution areas in Teno and Anaga might well be more ancient than the reported upheavals.
These putative barriers to gene flow may also mean that the two geographical elements of S. vespertilio on Tenerife may constitute separate subspecies, as genetically differentiated as S. vespertilio on Gran Canaria (SVD) is from SVV on Tenerife. The fact that the individuals from Teno show perfect intrapopulation uniformity with no indications of hybridization or migration from other groups in the fastSTRUCTURE analysis (Figure 3) provides further support of the possibility that this population originated from a founder event independent of the SVV in Anaga.
The pattern of relationships suggested by the SNP data (Figure 3; i.e., the nesting of the samples of SVD_O from Gran Canaria within the SVV Anaga populations) implies a closer relationship between the SVV from the Teno region of Tenerife and SVD from Gran Canaria. In the context of the currently accepted taxonomy of S. vespertilio, our results and the geographical closeness between Anaga and Gran Canaria suggest that dispersal from Anaga to Gran Canaria could have occurred much later than the interruption of gene flow between the populations of SVV from Anaga and Teno. The facts that (a) the area of Anaga has been ecologically (if not geologically) stable historically, and (b) it reportedly acted as a source of genetic diversity for other regions in the archipelago (Mairal et al., 2015), provide support to this first hypothesis. Our morphological data support as well the hypothesis that more recent natural hybridization between introduced SVV and some of the Gran Canarian SVD.
Taxonomic Implications
The experimental analyses of a large sample of common garden-grown plants and the molecular data confirm that the two species, S. lidii and S. vespertilio, are clearly distinct. The common garden studies show that the subspecies of S. vespertilio are morphologically very similar. Interestingly, the results of the SNP genotypic analyses suggest great populational variation, and the possibility of a complex colonization pattern involving SVV, SVD, and SL. Combined with the experimental hybridization studies and morphology, these data bolster the hypothesis of a recent hybridization between SVV and SVD. As noted, hybrids between the subspecies are easy to obtain in the greenhouse and they are fertile. Morphologically, the subspecies are, as is often the case, not very distinct: the morphology of the hybrids is either intermediate or a mixture of greater similarity to one parent than to the other. However, the high fertility of hybrids between the unquestionably distinct S. lidii and S. vespertilio also suggests that all three of the currently recognized Canarian native Solanum taxa are genetically very closely related. Thus, although infertility studies of hybrids can sometimes help evaluate species distinction (e.g., Anderson, 1977, 1979), in other instances the combination of genomes in hybrids is not inhibited by pre- or post-zygotic barriers, and species boundaries are presumably more ecologically or geographically based (Whalen and Anderson, 1981; Plazas et al., 2016). Furthermore, although there may be morphological and ecological differences between taxa, these do not necessarily imply more wholesale genome differences that might be manifest in F1 hybrid sterility.
In this case, artificial hybridization tells us that all three taxa are quite similar for a number of key genotypic characters, i.e., hybrids are not separated by genetic factors that are integral to the production of reproductive cells. Thus, not surprisingly, the subspecies are also inter-compatible, yielding hybrids with high fertility, an outcome not unprecedented in Angiosperms, not even in the genus Solanum (Plazas et al., 2016).
The lack of a suite of consistently expressed morphological characters distinguishing the subspecies is problematic (Table 2). However, the geographic and genotypic distinction of one of the few remaining representatives of S. vespertilio ssp. doramae (SVD_I_1 from Moya), and the nesting of other accessions of SVD fully interspersed within the collections of SVV (Figure 3) suggests that many of the presumed SVD samples (for instance those planted in Osorio) may constitute recent hybrids between SVV and SVD, raising important points for conservation programs that are considered further in the section below.
As noted above, the SNP genotypic data highlight a very distinct genotypic population of S. vespertilio from Teno (Bco. Los Cochinos, Los Silos), a more arid region in the northwest of Tenerife. The Teno samples of SVV are notably different from all other Tenerife accessions. Thus, these data perhaps reflect the geological and geographical distinction of the Teno area as a paleo-island (Carracedo and Day, 2002). A thorough morphological analysis is being carried out to confirm whether there are sufficient and consistent morphological characters to justify a new taxon within S. vespertilio (Marrero et al., in prep.).
Conservation Implications
The rareness and vulnerability of all the taxa considered, especially the two from Gran Canaria, make this study extremely important for designing effective conservation strategies and restoration programs that are based on the full understanding of the relationships, genotypic diversity, ecological requirements and reproductive biology (including pollination biology) of the most threatened taxa. For instance, there is an interesting paradox in the context of mating systems (Neal and Anderson, 2005) as applied to conservation. All three currently recognized taxa are capable of autogamy, though at a relatively low rate. Interestingly, the entity with the highest rate of autogamous fruit set in experimental settings (S. lidii), has the lowest fixation index (Fis), implying low self-breeding, in spite of starkly declining population sizes.
Our results show average Fis values (range between 0.301 and 0.587) for the larger and more widespread S. vespertilio (SVV) populations. These values make sense in terms of the reproductive biology and population biology of SVV as we understand it. However, they do not for S. lidii (SL), where Fis is negative (-0.204), indicating an excess of heterozygotes. This is uncommon and could be caused by either: (a) previous hybridization between genetically very different SL populations that have not yet reached Hardy-Weinberg equilibrium, or (b) natural selection for highly heterozygous individuals. However, regardless of the underlying cause, these perspectives are important for effective conservation design. Hybridization of SVV, SVD, or SL with other Solanum species occurring in the Canary islands, including the cultivated S. melongena L., that might endanger the genetic integrity of these Canarian endemics, is unlikely given the different habitats and phylogenetic distance with other Solanum species occurring in the Canary islands (Vorontsova and Knapp, 2016), the small size of the endemic populations, low success after artificial hybridization with cultivated eggplant (Kouassi et al., 2016), and the potential sterility of interspecific hybrids. Low Fst values for the comparisons of SVV with SVD indicates low genetic differentiation between these two subspecies, in spite of their distribution on separate islands. These lower values may suggest genetic flow between SVV and SVD, which may endanger the genetic integrity of the highly vulnerable SVD. To the contrary, the higher Fst values for comparisons of the geographically distinct SVV from Teno suggests that this entity has been less subject to hybridization with other SVV, SVD, or SL populations. Thus, the proposed subspecies (Marrero, in prep.) is likely to be more stable. In this way, for the distinctive Gran Canarian genotypes (SVD) to be maintained, conservation plans must limit the growth of introduced SVV near SVD. Indeed, the nesting of the presumably “pure” SVD individuals sampled at Finca de Osorio (Gran Canaria) within the SVV samples from Las Bodegas (Tenerife) (see Figure 3) indicates that these SVD samples may, in fact, be the result of hybridization with SVV (hybrids may have been easily achieved given the lack of intrinsic breeding barriers, and the cultivation at Osorio of several collections of SVV from Tenerife). Another explanation that cannot be ruled out would be that the plants introduced in Osorio SVD_O were originally subsp. vespertilio from Tenerife and may have been misidentified as the morphologically similar subsp. doramae. These issues highlight the necessity of careful identification and tracking of material introduced into conservation nurseries, and the essential value of molecular tools to achieve this objective, especially in cases of close morphological similarity. Given the results of the molecular analysis, and the forthcoming treatment of the subspecific ranks of S. vespertilio (Marrero et al., in prep.), we suggest that the SVD_I source, which is the most differentiated from SVV, might be considered for conservation-restoration efforts. Similarly, independently of whether the two areas of distribution of S. vespertilio subsp. vespertilio on the palaeo-islands of Tenerife (Anaga and Teno) represent distinct subspecies or not, their clear genetic differentiation indicates that in situ conservation efforts should avoid genetic mixing between them because the populations could be the result of two different evolutionary histories. At the minimum, the ravine in Teno (“Los Cochinos”) harbors a genotypically and ecologically distinct population that makes it a biodiversity micro-reserve, a refugium perhaps, of Solanum diversity (similar to conclusions by others, e.g., Mairal et al., 2015; Puppo et al., 2016; Soto et al., in prep.). Thus, new populations should be established with local genetic stock, under strict genetic traceability, and in ecologically similar nearby areas (Marrero-Gómez et al., 2010). And, obviously, irrespective of plant genotypes, removing competing invasive plants and eliminating herbivores like feral goats (Campbell and Donlan, 2005) or rabbits (Stuessy et al., 2017b, c) from islands are powerful conservation tools to prevent extinction and allow recovery of ecosystems.
Finally, as with most considerations of biodiversity, it is difficult to assess how future climate scenarios may affect the endemic populations we studied. However the current cloud forests of the Canaries generated by trade winds, crucial for the water balance in many of the natural habitats for S. vespertilio in particular (Kunkel, 1975b; Bramwell and Bramwell, 1990; Marrero-Gómez et al., 2010), have been predicted to decline in the forthcoming decades due to likely climate change (Sperling et al., 2004; Otto et al., 2017; Mairal et al., 2018). Given these vulnerabilities, the seed stocks of these Solanum kept at the seed bank of the JBCVCSIC should be reinforced to ensure success in future restoration efforts.
Conclusion
A broad combination of approaches employed in this study of the endemic Solanum of the Canary Islands indicates that complex morphological and molecular variation is distributed among these taxa. Although they are still capable of sexual reproduction, suitable habitats are diminishing and populations are seriously declining. To successfully protect and restore native populations of these Solanum and other oceanic island endemics (Stuessy et al., 2017b), the biology of the plants and the distribution of diversity among them must be fully understood.
Our results showed that the three taxa display clear morphological differences, but they are fully cross-compatible, which may endanger their genetic integrity. The novel SNPs and SPET technology we utilized provides a powerful tool that, together with other multidisciplinary data generated, build the comprehensive foundation necessary for the successful ex situ management and in situ restoration of the Canarian Solanum and other island taxa. In this context, despite the small population sizes, we have found low levels for the inbreeding coefficient and a low genetic differentiation between SVV and SVD, suggesting that genetic flow that may compromise the genetic integrity of SVD may have taken place. Our results on relationships and genetic differentiation also suggest that an additional taxonomic entity, in addition to SVV and SVD, may exist within S. vespertilio.
Importantly, the different complementary approaches that we applied provide solid foundations for the conservation strategies of S. vespertilio and S. lidii. We consider that dialogue among scientists and various stakeholders is necessary to implement recovery plans that reduce direct threats such as land development, feral herbivores, invasive species, and genetic contamination on these endangered Canarian endemisms.
Data Availability Statement
The datasets generated for this study can be found in the sequence data have been deposited into the NCBI Short Read Archive under the Bioproject identifier PRJNA556343. Raw reads of each accession are deposited under the accession numbers from SRX6649376 to SRX6649471.
Author Contributions
PG and RJ-M: data curation, project administration, and writing – original draft preparation. PG, RJ-M, SV, and GA: formal analysis. PG, RJ-M, SV, JP, ÁM, JC-C, and GA: methodology, writing – review and editing, and data visualization. JP and JC-C: resources. JP, JC-C, and GA: supervision and research design and funding management.
Funding
This work was supported by funds from project DEMIURGO to the Jardín Botánico Canario “Viera y Clavijo” – Unidad Asociada al CSIC of the Cabildo de Gran Canaria. Consumables and genotyping were funded by the Jardín Botánico Canario “Viera y Clavijo” – Unidad Asociada al CSIC. Participation of authors from Universitat Politècnica de València has been partially funded by the European Union’s Horizon 2020 Research and Innovation Programme under the Grant Agreement No. 677379 (G2P-SOL project: Linking genetic resources, genomes, and phenotypes of solanaceous crops). PG was grateful to Universitat Politècnica de València and to Japan Society for the Promotion of Science for their respective Postdoctoral Grants [PAID-10-18 and FY2019-P19105 JSPS Postdoctoral Fellowship for Research in Japan (Standard)].
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Paul Lewis generously provided an additional SVDQuartets tree as an aid to our analyses. We thank the Cabildos of Tenerife and Gran Canaria (Canary Islands) for providing permits for collecting samples of the three Solanum taxa. We thank Óscar Saturno, Arnoldo Santos Guerra, and Conchi Santiago who assisted collecting specimens in their natural populations. We thank Rafael Almeida for preserving and cultivating seeds and maintaining two plants of vespertilio subsp. doramae. We thank Miguel Ángel Cabrera from Gobierno de Canarias for the information provided, especially about the population of Barranco de Azuaje. The Department of Ecology and Evolutionary Biology provided support for GA. Clinton Morse, Matt Opel, and Adam Histen provided excellent greenhouse facilities for plant culture at the University of Connecticut. Two students, Catherine Menousek and Lauren M. Albert assisted GA in data gathering, and Mona Anderson and Dan Crawford in fieldwork. Catherine Menousek provided the floral drawings that are part of Figure 2.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2020.00757/full#supplementary-material
DATA S1 | Statistics of sequencing and mapping onto the reference genome “67/3” (Barchi et al., 2019b).
DATA S2 | Mean probe coverage of the high-quality reads using the eggplant SPET platform (Barchi et al., 2019a).
DATA S3 | SNP and population structure statistics.
DATA S4 | Population structure statistics for the eight populations from the three Solanum Canarian taxa.
DATA S5 | Population structure statistics for the accessions from the three Solanum Canarian taxa used in this study.
DATA S6 | Set of private SNPs and core private SNPs for SL, SVV, and SVD.
DATA S7 | SVDquartets dendrogram.
Footnotes
- ^ https://www.miteco.gob.es/es/biodiversidad/legislacion/leg-espanola-conservacion-especies.aspx
- ^ https://software.broadinstitute.org/gatk/best-practices/workflow?id=11145
- ^ http://solanaceaesource.org/
References
Ablay, G., and Hürlimann, M. (2000). Evolution of the north flank of Tenerife by recurrent giant landslides. J. Volcanol. Geotherm. Res. 103, 135–159. doi: 10.1016/S0377-0273(00)00220-1
Alonso, D., Gramazio, P., Plazas, M., Villanueva, G., García-Fortea, E., Díez, M. J., et al. (2018). “Protocolo innovador para la obtención de ADN genómico vegetal de alta calidad,” in Proceedings of the Congress I Congreso de Jóvenes Investigadores en Ciencias Agroalimentarias; 2018 Dec 20, Almeria, 282–283.
Ancochea, E., Fuster, J., Ibarrola, E., Cendrero, A., Coello, J., Hernan, F., et al. (1990). Volcanic evolution of the island of Tenerife (Canary Islands) in the light of new K-Ar data. J. Volcanol. Geotherm. Res. 44, 231–249. doi: 10.1016/0377-0273(90)90019-C
Anderson, G. J. (1977). The variation and evolution of selected species of Solanum section Basarthrum (Solanaceae) II. Brittonia 29, 116–128. doi: 10.2307/2805892
Anderson, G. J. (1979). “Systematic and evolutionary consideration consideration of species of Solanum, section Basarthrum,” in The Biology and Taxonomy of the Solanaceae, eds J. G. Hawkes, R. N. Lester, and A. D. Skelding (Cambridge, MA: Academic Press), 549–562.
Anderson, G. J., Bernardello, G., and Santos-Guerra, A. (2015). Reproductive biology of Solanum vespertilio (Solanaceae), a zygomorphic, heterantherous, enantiostylous, and andromonoecious rare Canary Islands endemic. Plant Syst. Evol. 301, 1191–1206. doi: 10.1007/s00606-014-1143-4
Anderson, G. J., and Levine, D. A. (1982). Three taxa constitute the sexes of a single dioecious species of Solanum. Taxon 31, 667–672. doi: 10.2307/1219682
Anderson, G. J., and Symon, D. E. (1989). Functional dioecy and andromonoecy in Solanum. Evolution 43, 204–219. doi: 10.2307/2409175
Baldwin, B. G., and Warren, L. W. (2010). Hawaiian angiosperm radiations of North American origin. Ann. Bot. 105, 849–879. doi: 10.1093/aob/mcq052
Bañares, A., Blanca, G., Güemes, J., Moreno, J. C., and Ortiz, S. (2003). Atlas y Libro Rojo de la Flora Vascular Amenazada de España. Madrid: Dirección General de Conservación de la Naturaleza, 1072.
Barchi, L., Acquadro, A., Alonso, D., Aprea, G., Bassolino, L., Demurtas, O. C., et al. (2019a). Single Primer Enrichment Technology (SPET) for high-throughput genotyping in tomato and eggplant germplasm. Front. Plant Sci. 10:1005. doi: 10.3389/fpls.2019.01005
Barchi, L., Pietrella, M., Venturini, L., Minio, A., Toppino, L., Acquadro, A., et al. (2019b). A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci. Rep. 9, 1–13. doi: 10.1038/s41598-019-47985-w
Bramwell, D., and Bramwell, Z. I. (1990). Flores silvestres de las Islas Canarias. Madrid: Rueda Madrid.
Bruvo, R., Michiels, N. K., D’Souza, T. G., and Schulenburg, H. (2004). A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 13, 2101–2106. doi: 10.1111/j.1365-294X.2004.02209.x
Cabrera, M. A., Abreu, S., and Ojeda, E. (2006). Visita a la Población Natural Para la Redacción del Plan de Recuperación de la Especie Vegetal Rejalgadera de Doramas (“Solanum Vespertilio Doramae”). Canary Islands: Regional Ministry of the Environment and Territorial Organisation of Canary Islands.
Campbell, K., and Donlan, C. J. (2005). Feral goat eradications on islands. Conserv. Biol. 19, 1362–1374. doi: 10.1111/j.1523-1739.2005.00228.x
Caujapé-Castells, J., Tye, A., Crawford, D. J., Santos-Guerra, A., Sakai, A., Beaver, K., et al. (2010). Conservation of oceanic island floras: Present and future global challenges. Perspect. Plant Ecol. Evol. Syst. 12, 107–129. doi: 10.1016/j.ppees.2009.10.001
Chifman, J., and Kubatko, L. (2014). Quartet inference from SNP data under the coalescent model. Bioinformatics 30, 3317–3324. doi: 10.1093/bioinformatics/btu530
Chifman, J., and Kubatko, L. (2015). Identifiability of the unrooted species tree topology under the coalescent model with time-reversible substitution processes, site-specific rate variation, and invariable sites. J. Theor. Biol. 374, 35–47. doi: 10.1016/j.jtbi.2015.03.006
Corlett, R. T. (2017). A bigger toolbox: biotechnology in biodiversity conservation. Trends Biotechnol. 35, 55–65. doi: 10.1016/j.tibtech.2016.06.009
Crawford, D. J., Moura, M., Borges, S. L., Mort, M. E., Kerbs, B., Schaefer, H., et al. (2019). The transition to selfing in Azorean Tolpis (Asteraceae). Plant Syst. Evol. 305, 305–317. doi: 10.1007/s00606-019-01573-7
Curto, M., Puppo, P., Kratschmer, S., and Meimberg, H. (2017). Genetic diversity and differentiation patterns in Micromeria from the Canary Islands are congruent with multiple colonization dynamics and the establishment of species syngameons. BMC Evol. Biol. 17:198. doi: 10.1186/s12862-017-1031-y
Danecek, P., Auton, A., Abecasis, G., Albers, C. A., Banks, E., DePristo, M. A., et al. (2011). The variant call format and VCFtools. Bioinformatics 27, 2156–2158. doi: 10.1093/bioinformatics/btr330
Del Fabbro, C., Scalabrin, S., Morgante, M., and Giorgi, F. M. (2013). An extensive evaluation of read trimming effects on illumina NGS data analysis. PLoS One 8:e85024. doi: 10.1371/journal.pone.0085024
Depristo, M. A., Banks, E., Poplin, R., Garimella, K. V., Maguire, J. R., Hartl, C., et al. (2011). A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 43, 491–501. doi: 10.1038/ng.806
Díaz-Bertrana, M., and Saturno, O. (2017). Servicio de Realización de Acciones Referentes a los Planes de Recuperación Aprobados Para Tres Especies de Plantas Amenazadas en Gran Canaria: La Flor de Mayo Leñosa (Pericallis Hadrosoma), Rejalgadera de Doramas (Solanum Vespertilio subsp. Doramae), Escobilla de Guayadeque (Kunkeliella Canariensis) y Evaluar las Acciones Realizadas Para la Yerbamuda de Jinámar (Lotus Kunkelii). Canary Islands: Regional Ministry of the Environment and Territorial Organisation of Canary Islands.
Docoito-Díaz, J. R., and Herrera, B. (2005). Solanum Vespertilio subsp. Doramae Marrero Rodr. & Glez. Martín. Seguimiento de Poblaciones de Especies Amenazadas. Canary Islands: Regional Ministry of the Environment and Territorial Organisation of Canary Islands.
García, P. E., Schönswetter, P., Fuertes Aguilar, J., Nieto Feliner, G., and Schneeweiss, G. M. (2009). Five molecular markers reveal extensive morphological homoplasy and reticulate evolution in the Malva alliance (Malvaceae). Mol. Phylogenet. Evol. 50, 226–239. doi: 10.1016/j.ympev.2008.10.015
García-Verdugo, C., Mairal, M., Monroy, P., Sajeva, M., and Caujapé-Castells, J. (2017). The loss of dispersal on islands hypothesis revisited: Implementing phylogeography to investigate evolution of dispersal traits in Periploca (Apocynaceae). J. Biogeogr. 44, 2595–2606. doi: 10.1111/jbi.13050
González De, F. C. A. (2016). Islas Dentro de Islas: Biología y Conservación del Paleoendemismo Macaronésico Navaea Phoenicea (Vent.) Webb & Bertel. Ph.D. Thesis, Madrid, SP, Autonomous University of Madrid.
Goudet, J. (2005). HIERFSTAT, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 5, 184–186. doi: 10.1111/j.1471-8286.2004.00828.x
Gramazio, P., Yan, H., Hasing, T., Vilanova, S., Prohens, J., and Bombarely, A. (2019). Whole-genome resequencing of seven eggplant (Solanum melongena) and one wild relative (S. incanum) accessions provides new insights and breeding tools for eggplant enhancement. Front. Plant Sci. 10:1220. doi: 10.3389/fpls.2019.01220
Ihaka, R., and Gentleman, R. (1996). R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 5, 299–314. doi: 10.1080/10618600.1996.10474713
Jaén-Molina, R., Marrero, Á, Medina, F. M., Mesa-Coello, R., and Caujapé-Castells, J. (2015). “Detection of possible cryptic taxa in Dorycnium sect. canaria for the Canary Islands,” in Proceedings of the congress Floramac 2015Mar 24-27, (Canary Islands: Las Palmas de Gran Canaria).
Jombart, T. (2008). adegenet: a R package for the multivariate analysis of genetic markers. Bioinformatics 24, 1403–1405. doi: 10.1093/bioinformatics/btn129
Kamvar, Z. N., Tabima, J. F., and Grünwald, N. J. (2014). Poppr: an R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2:e281. doi: 10.7717/peerj.281
Kim, C., Guo, H., Kong, W., Chandnani, R., Shuang, L. S., and Paterson, A. H. (2016). Application of genotyping by sequencing technology to a variety of crop breeding programs. Plant Sci. 242, 14–22. doi: 10.1016/j.plantsci.2015.04.016
Knaus, B. J., and Grünwald, N. J. (2017). VCFR: a package to manipulate and visualize variant call format data in R. Mol. Ecol. Resour. 17, 44–53. doi: 10.1111/1755-0998.12549
Kouassi, B., Prohens, J., Gramazio, P., Kouassi, A. B., Vilanova, S., Galán-Ávila, A., et al. (2016). Development of backcross generations and new interspecific hybrid combinations for introgression breeding in eggplant (Solanum melongena). Sci. Hort. 213, 199–207. doi: 10.1016/j.scienta.2016.10.039
Kunkel, G. (1975a). Endemismos Canarios. Inventario de las Plantas Vasculares Endémicas de la Provincia de Las Palmas. Canary Islands: Spanish Ministry of Agriculture monograph.
Kunkel, G. (1975b). Flora y Vegetación. Inventario de los Recursos naturales renovables de la Provincia de Las Palmas de Gran Canaria. Canary Islands: Spanish Ministry of Agriculture monograph.
Li, H. (2013). Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM. arXiv [Preprint]. arXiv:1303.3997.
Lischer, H. E. L., and Excoffier, L. (2012). PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 28, 298–299. doi: 10.1093/bioinformatics/btr642
Mairal, M., Caujapé-Castells, J., Pellissier, L., Jaén-Molina, R., and Álvarez, N. (2018). A tale of two forests: ongoing aridification drives population decline and genetic diversity loss at continental scale in Afro-Macaronesian evergreen-forest archipelago. Ann. Bot. 122, 1005–1017. doi: 10.1093/aob/mcy107
Mairal, M., Sanmartín, I., Aldasoro, J. J., Culshaw, V., Manolopoulou, I., and Alarcõn, M. (2015). Palaeo-islands as refugia and sources of genetic diversity within volcanic archipelagos: The case of the widespread endemic Canarina canariensis (Campanulaceae). Mol. Ecol. 24, 3944–3963. doi: 10.1111/mec.13282
Marrero, Á (1986). Sobre plantas relícticas de Gran Canaria: comentarios corólogicos-ecológicos. Bot. Macaronésica 12, 51–62.
Marrero, Á, and González Martín, M. (1998). Solanum vespertilio subsp. doramae Á. Marrero & M. González Martín, subsp. nov. (Solanaceae) de Gran Canaria (Islas Canarias). Anal. Jard. Bot. Madrid 56, 388–390.
Marrero-Gómez, M. C., Mesa-Coello, R., Acevedo-Rodríguez, A., and Oval de la Rosa, J. P. (2010). “Solanum vespertilio Aiton subsp. vespertilio,” in Atlas y Libro Rojo de la Flora Vascular Amenazada de España, eds Á Bañares, G. Blanca, J. Güemes, J. C. Moreno, and S. Ortiz (Canary Islands: Ministry of Agriculture, Fisheries and Food), 506–507.
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 17, 10–12. doi: 10.14806/ej.17.1.200
Mesa-Coello, R., Paredes-Gil, R., and Díaz-Bertrana, M. (2013). Seguimiento de poblaciones de especies amenazadas: Solanum lidii Sunding. Canary Islands: Canary Islands Government.
Moreno, J. C. (2008). De la Flora Vascular Española. Canary Islands: Spanish Ministry of the Environment, and Rural and Marine Affairs.
Moya, Ó, Contreras-Díaz, H. G., Oromí, P., and Juan, C. (2004). Genetic structure, phylogeography and demography of two ground-beetle species endemic to the Tenerife laurel forest (Canary Islands). Mol. Ecol. 13, 3153–3167. doi: 10.1111/j.1365-294X.2004.02316.x
Neal, P. R., and Anderson, G. J. (2005). Are “mating systems” “breeding systems” of inconsistent and confusing terminology in plant reproductive biology? or is it the other way around? Pl. Syst. Evol. 250, 173–185. doi: 10.1007/s00606-004-0229-9
Nei, M. (1973). Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. U.S.A. 70, 3321–3323. doi: 10.1073/pnas.70.12.3321
Nei, M. (1978). Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89, 583–590.
Nogales, M., Heleno, R., Rumeu, B., González-Castro, A., Traveset, A., Vargas, P., et al. (2016). Seed-dispersal networks on the Canaries and the Galápagos archipelagos: interaction modules as biogeographical entities. Glob. Ecol. Biogeogr. 25, 912–922. doi: 10.1111/geb.12315
Otto, R., Garzón-Machado, V., del Arco, M., Fernández-Lugo, S., de Nascimento, L., Oromí, P., et al. (2017). Unpaid extinction debts for endemic plants and invertebrates as a legacy of habitat loss on oceanic islands. Divers. Distrib. 23, 1031–1041. doi: 10.1111/ddi.12590
Paradis, E., and Schliep, K. (2018). Ape 5.0: An environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35, 526–528. doi: 10.1093/bioinformatics/bty633
Plazas, M., Vilanova, S., Gramazio, P., Rodríguez-Burruezo, A., Fita, A., Herraiz, F. J., et al. (2016). Interspecific hybridization between eggplant and wild relatives from different genepools. J. Am. Soc. Hortic. Sci. 141, 34–44. doi: 10.21273/jashs.141.1.34
Poplin, R., Ruano-Rubio, V., DePristo, M., Fennell, T. J., Carneiro, M. O., Van der Auwera, G. A., et al. (2017). Scaling accurate genetic variant discovery to tens of thousands of samples. BioRxiv [Preprint]. BioRxiv: 201178.
Price, J. P., Otto, R., Menezes de, S. M., Kueffer, C., Schaefer, H., Caujapé-Castells, J., et al. (2018). Colonization and diversification shape species–area relationships in three Macaronesian archipelagos. J. Biogeogr. 45, 2027–2039. doi: 10.1111/jbi.13396
Prohens, J., Anderson, G. J., Herraiz, F. J., Bernardello, G., Santos-Guerra, A., Crawford, D., et al. (2007). Genetic diversity and conservation of two endangered eggplant relatives (Solanum vespertilio Aiton and Solanum lidii Sunding) endemic to the Canary Islands. Genet. Resour. Crop Evol. 54, 451–464. doi: 10.1007/s10722-006-9174-5
Puppo, P., Curto, M., Gusmão-Guedes, J., Cochofel, J., Pérez de Paz, P. L., Bräuchler, C., et al. (2015). Molecular phylogenetics of Micromeria (Lamiaceae) in the Canary Islands, diversification and inter-island colonization patterns inferred from nuclear genes. Mol. Phylogenet. Evol. 89, 160–170. doi: 10.1016/j.ympev.2015.04.017
Puppo, P., Curto, M., and Meimberg, H. (2016). Genetic structure of Micromeria (Lamiaceae) in Tenerife, the imprint of geological history and hybridization on within-island diversification. Ecol. Evol. 6, 3443–3460. doi: 10.1002/ece3.2094
Raj, A., Stephens, M., and Pritchard, J. K. (2014). FastSTRUCTURE: Variational inference of population structure in large SNP data sets. Genetics 197, 573–589. doi: 10.1534/genetics.114.164350
Rodríguez N. M. L., and Garzón M. V. (2019). Predictions about the distribution of Solanum vespertilio Aiton ssp. vespertilio (Solanaceae), Tenerife, Canary Islands. Vieraea 46, 203–230. doi: 10.31939/vieraea.2019.46.tomo01.10
Rodríguez-Delgado, O., García-Gallo, A., and Cruz-Trujillo, G. M. (2004). “Solanum lidii Sunding,” in Atlas y Libro Rojo de la Flora Vascular Amenazada en España, eds A. Bañares, G. Blanca, J. Güemes, J. C. Moreno, and S. Ortiz (Canary Islands: Spanish Ministry of the Environment, and Rural and Marine Affairs).
Rumeu, B., Vargas, P., Jaén-Molina, R., Nogales, M., and Caujapé-Castells, J. (2014). Phylogeography and genetic structure of the threatened Canarian Juniperus cedrus (Cupressaceae). Bot. J. Linn. Soc. 175, 376–394. doi: 10.1111/boj.12172
Santana, I., and Naranjo, M. (2002). Seguimiento de Poblaciones de Especies Amenazadas. Solanum Vespertilio Doramae. Canary Islands: Regional Ministry of the Environment and Territorial Organisation of Canary Islands.
Särkinen, T., Bohs, L., Olmstead, R. G., and Knapp, S. (2013). A phylogenetic framework for evolutionary study of the nightshades (Solanaceae): a dated 1000-tip tree. BMC Evol. Biol. 13:214. doi: 10.1186/1471-2148-13-214
Scaglione, D., Pinosio, S., Marroni, F., Di Centa, E., Fornasiero, A., Magris, G., et al. (2019). Single primer enrichment technology as a tool for massive genotyping: a benchmark on black poplar and maize. Ann. Bot. 124:mcz054. doi: 10.1093/aob/mcz054
Scheben, A., Batley, J., and Edwards, D. (2017). Genotyping-by-sequencing approaches to characterize crop genomes: choosing the right tool for the right application. Plant Biotechnol. J. 15, 149–161. doi: 10.1111/pbi.12645
Sperling, F. N., Washington, R., and Whittaker, R. J. (2004). Future climate change of the subtropical North Atlantic: implications for the cloud forests of tenerife. Clim. Change 65, 103–123. doi: 10.1023/B:CLIM.0000037488.33377.bf
Stuessy, T., Crawford, D., López-Sepúlveda, P., Baeza, C., and Ruiz, E. (2017a). Plants of Oceanic Islands: Evolution, Biogeography, and Conservation of the Flora of the Juan Fernandez (Robinson Crusoe) Archipelago. Cambridge: Cambridge University Press.
Stuessy, T. F., Baeza, C. M., López-Sepúlveda, P., Rodríguez, R., and Swenson, U. (2017b). “Conservation of native and endemic species,” in Plants of Oceanic Islands: Evolution, Biogeography, and Conservation of the Flora of the Juan Fernandez (Robinson Crusoe) Archipelago, eds T. Stuessy, D. Crawford, P. López-Sepúlveda, C. Baeza, and E. Ruiz (Cambridge: Cambridge University Press), 117–133.
Stuessy, T. F., Marticorena, C., Swenson, U., Greimler, J., and López-Sepúlveda, P. (2017c). “Impacts of Vegetation,” in Plants of Oceanic Islands: Evolution, Biogeography, and Conservation of the Flora of the Juan Fernandez (Robinson Crusoe) Archipelago, eds T. Stuessy, D. Crawford, P. López-Sepúlveda, C. Baeza, and E. Ruiz (Cambridge: Cambridge University Press), 335–353.
Swofford, D. L. (2003). PAUP∗. Phylogenetic Analysis Using Parsimony (∗ and Other Methods). Sunderland: Sinauer Associates.
Thomson, M. J. (2014). High-throughput SNP genotyping to accelerate crop improvement. Plant Breed. Biotechnol. 2, 195–212. doi: 10.9787/pbb.2014.2.3.195
Trusty, J. L., Olmstead, R. G., Santos-Guerra, A., Sá-Fontinha, S., and Francisco-Ortega, J. (2005). Molecular phylogenetics of the Macaronesian-endemic genus Bystropogon (Lamiaceae): Palaeo-islands, ecological shifts and interisland colonizations. Mol. Ecol. 14, 1177–1189. doi: 10.1111/j.1365-294X.2005.02487.x
Van Den Bogaard, P. (2013). The origin of the Canary Island Seamount Province-New ages of old seamounts. Sci. Rep. 3:2107. doi: 10.1038/srep02107
Vorontsova, M. S., and Knapp, S. (2016). A Revision of the “Spiny Solanums,” Solanum subgenus Leptostemonum (Solanaceae), in Africa and Madagascar. Syst. Bot. Monogr. 99, 1–432.
Warren, B. H., Simberloff, D., Ricklefs, R. E., Aguilée, R., Condamine, F. L., Gravel, D., et al. (2015). Islands as model systems in ecology and evolution: Prospects fifty years after MacArthur-Wilson. Ecol. Lett. 18, 200–217. doi: 10.1111/ele.12398
Watts, A. B., and Masson, D. G. (1995). A giant landslide on the north flank of Tenerife, Canary Islands. J. Geophys. Res. 100, 24487–24498. doi: 10.1029/95jb02630
Weigelt, P., Daniel, K. W., Kisel, Y., Fritz, S. A., Karger, D. N., Kessler, M., et al. (2015). Global patterns and drivers of phylogenetic structure in island floras. Sci. Rep. 5:12213. doi: 10.1038/srep12213
Whalen, M. D., and Anderson, G. J. (1981). Distribution of gametophytic self-incompatibility and infrageneric classification in Solanum. Taxon 30, 761–767. doi: 10.2307/1220077
Keywords: conservation, endangered endemics, reproductive biology, SNPs, Solanaceae, Solanum, Canary Islands, SPET
Citation: Gramazio P, Jaén-Molina R, Vilanova S, Prohens J, Marrero Á, Caujapé-Castells J and Anderson GJ (2020) Fostering Conservation via an Integrated Use of Conventional Approaches and High-Throughput SPET Genotyping: A Case Study Using the Endangered Canarian Endemics Solanum lidii and S. vespertilio (Solanaceae). Front. Plant Sci. 11:757. doi: 10.3389/fpls.2020.00757
Received: 17 March 2020; Accepted: 12 May 2020;
Published: 10 July 2020.
Edited by:
Marcial Escudero, University of Seville, SpainReviewed by:
Ezio Portis, University of Turin, ItalyMaria Manuel Romeiras, Linking Landscape, Environment, Agriculture and Food, Higher Institute of Agronomy, University of Lisbon, Portugal
Copyright © 2020 Gramazio, Jaén-Molina, Vilanova, Prohens, Marrero, Caujapé-Castells and Anderson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juli Caujapé-Castells, anVsaWNhdWphcGVAZ21haWwuY29t
†These authors have contributed equally to this work