 Ying Ma
Ying Ma Helena Freitas
Helena Freitas Maria Celeste Dias
Maria Celeste Dias- Centre for Functional Ecology, Department of Life Sciences, University of Coimbra, Calçada Martim de Freitas, Coimbra, Portugal
Global climate change-induced abiotic stresses (e.g., drought, salinity, extreme temperatures, heavy metals, and UV radiation) have destabilized the fragile agroecosystems and impaired plant performance and thereby reducing crop productivity and quality. Biostimulants, as a promising and eco-friendly approach, are widely used to address environmental concerns and fulfill the need for developing sustainable/modern agriculture. Current knowledge revealed that plant and animal derived stimulants (e.g., seaweeds and phytoextracts, humic substances, and protein hydrolysate) as well as microbial stimulants (e.g., plant beneficial bacteria or fungi) have great potential to elicit plant tolerance to various abiotic stresses and thus enhancing plant growth and performance-related parameters (such as root growth/diameter, flowering, nutrient use efficiency/translocation, soil water holding capacity, and microbial activity). However, to successfully implement biostimulant-based agriculture in the field under changing climate, the understanding of agricultural functions and action mechanism of biostimulants coping with various abiotic stresses at physicochemical, metabolic, and molecular levels is needed. Therefore, this review attempts to unravel the underlying mechanisms of action mediated by diverse biostimulants in relation to abiotic stress alleviation as well as to discuss the current challenges in their commercialization and implementation in agriculture under changing climate conditions.
Introduction
Rapid climate change has exacerbated various biotic and abiotic stresses on agricultural plants/crops, causing a change in plants’ physiological, biochemical, cellular, and molecular mechanisms (Ma et al., 2020). The acceleration of global warming has devastating effects on plant growth and crop yield, as well as nutritional quality, threatening sustainable crop production worldwide (Saddiq et al., 2021). During the decades of the industrial revolution, humankind has relied on the massive use of chemical inputs to stimulate crop production, therefore continuously unbalancing its environment (Delitte et al., 2021). The intensive anthropogenic activities and rapid industrialization in combination with current climate change impacts have led to a reduction in the quality of arable land and environmental pollution (e.g., over-accumulation of heavy metals in the soils) (Oleńska et al., 2020). Under natural climate/field/agricultural conditions, plants usually face a combination of different environmental stresses such as drought, salinity, extreme temperatures, heavy metals, and UV radiation, contributing to the augmented joint stress severity (Ma et al., 2020; Srivastava et al., 2021). To adapt to these stresses, not all species have the ability to evolve their plastic responses and grow in such harsh environments (Shah et al., 2021). It was estimated by the U.S. National Climate Assessment that environmental stresses account for the losses in crop yield of up to 50%. In this regard, various agrochemical approaches have been developed to alleviate the adverse environmental effects and synthetic fertilizers (e.g., nano-silicon, FeO nanoparticles) and pesticides have been perceived as key agricultural inputs (Haghighi and Pessarakli, 2013; Manzoor et al., 2021; Silva et al., 2022). Nevertheless, while the imbalanced application of these agrichemicals enhances crop productivity, they also damage the environment and have impact on human health. There is an increasing demand for alternatives to traditional/conventional agrichemicals.
Lately, the application of biostimulants has been considered a potentially novel approach to stimulate plant growth and crop productivity under both stress and control conditions, representing a momentous/significant scientific breakthrough toward safeguarding future food production in a sustainable manner (Rai et al., 2021). Although numerous studies have demonstrated the promising potential, functions, and possible challenges of biostimulants to mitigate abiotic stresses and improve quality and yield (Van Oosten et al., 2017; Zaid et al., 2020), the underlying mechanisms of biostimulant-plant interactions at the cellular, metabolic and molecular levels to overcome stress adversities are still elusive (Nephali et al., 2020). In the light of recent advances in the field of plant-biostimulant interactions, this review attempts to unravel the underlying mechanisms of action mediated by diverse biostimulants in relation to abiotic stress alleviation as well as to discuss the current challenges in their commercialization and implementation in agriculture under changing climate conditions.
Impact of abiotic stresses on plant performance
Global climate change scenarios have contributed to increasing the intensity of abiotic stresses, such as drought, salinity, heat, and high UV radiation, which lead to extensive losses in agriculture (Waqas et al., 2019). Moreover, climate change projections and crop yield models predict a worsening of losses in main agricultural crops, such as wheat, rice, and maize, in the next years, with serious impacts on food security (Tigchelaar et al., 2018). Therefore, the development of sustainable strategies to reduce the impact of abiotic stresses on plant performance is of high priority. Among the various strategies to mitigate the negative effects of abiotic stresses on plants, the use of eco-friendly compounds, such as microbial and non-microbial biostimulants, is one of the most promising (Rouphael et al., 2020; Ali et al., 2021).
As sessile organisms, plants developed sophisticated perception mechanisms, as well as signaling and acclimation strategies to cope with harsh environmental conditions, but at the cost of decreased growth and yield (Zandalinas et al., 2020). These mechanisms include increased production of antioxidant metabolites (e.g., flavonoids, proline, and enzymes) that help in the control of reactive oxygen species (ROS) to avoid oxidative stress and changes in the levels of phytohormones (e.g., ABA and JA) (Zandalinas et al., 2020; Ma et al., 2020; Dias et al., 2022; Dias et al., 2018). Hence, drought and salinity represent a major threat to plant productivity (Daliakopoulos et al., 2016; de Oliveira et al., 2022). The first morphophysiological responses of plants to drought and salinity are very similar, inducing water stress, leading to cellular dehydration and a reduction in water potential, hampering cell expansion and wall synthesis, growth and shoot development (Ma et al., 2020). Moreover, drought and salinity reduce transpiration and nutrient uptake (Ahluwalia et al., 2021). With the extent of drought, root elongation is continuous for groundwater search, while under salinity ionic stress also occurs, and roots started to accumulate a high quantity of ions, principally Na2+ (Ma et al., 2020). Photosynthesis impairment occurs, together with reductions in pigment synthesis and enzyme activities. Under these conditions, the light absorbed in excess and not used for photosynthesis can lead to oxidative stress (Ma et al., 2020). Some species have mechanisms of salt exclusion, and others concentrate salt in vacuoles (Khan et al., 2020), leading to lower negative effects on metabolic processes, such as photosynthesis. Also, some metabolites such as carbohydrates, amino acids, and nitrogen play a vital role in protecting plants against stresses by acting as osmolytes and/or ROS scavengers (Sharma et al., 2019).
Moreover, frequent and severe heat waves have been already occurring and affecting the phenology, productivity, and yield of many plants (Dusenge et al., 2019). Heat stress can increase membrane permeability, degradation of proteins and decrease in their synthesis, inactivation of enzymes, reduction of photosynthesis, and inhibition of pigment synthesis (Jagadish et al., 2021; Zhao et al., 2021). These effects can lead to a slowdown in growth and an increase in ROS production (Vargas et al., 2021).
In addition, plants are usually subjected to a simultaneous combination of different abiotic stresses, triggering specific responses (Savvides et al., 2016). Most of the studies reported higher negative impacts on plant performance when some abiotic stresses (drought and heat, salinity and heat) are combined, compared to each abiotic stress applied individually (Escobar-Bravo et al., 2017; Zandalinas et al., 2018; Zandalinas et al., 2020). For instance, in Arabidopsis, salinity combined with heat induces higher impacts on growth, chlorophyll content and Na+/K+ than these stresses applied separately (Suzuki et al., 2016). Dissimilarly, in tomato plants, heat combined with salinity improved photosynthesis, water and osmotic potential, and decreased protein oxidation and H2O2 levels when compared to the effects of salinity stress alone (Rivero et al., 2014). Plants seem to integrate simultaneously two different systemic signals generated when exposed to a combination of stresses, and the way plants perceive the stresses that trigger these signals strongly influences the intensity and efficiency of responses (ROS signals, changes in transcript abundance, hormonal, and stomatal responses) (Zandalinas et al., 2020). Moreover, these responses must be fast and well-coordinated between the distinct parts of the plants (leaves, roots, or stem) (Zandalinas et al., 2020).
Biostimulants: Definition, type,and role in agriculture
Biostimulants are organic or inorganic compounds and/or microorganisms, that when applied to plants, stimulate several processes, leading to improved growth and productivity, and tolerance to stresses (Ali et al., 2021; Franzoni et al., 2022). They are available as soluble powder, granular form, or liquid and can be applied as foliar sprays or/and in soil near the root zone. Biostimulants are considered environmentally friendly agronomic tools, and the market for these compounds is increasing at an annual growth rate of 12% possibly reaching 5.6 billion dollars in 2026 (Bulgari et al., 2017; Lau et al., 2022). Several factors have influenced the increase in the biostimulant market, namely the changes in agricultural and environmental policies allied with climate change, pushed a necessity for more sustainable alternatives to synthetic chemicals (Lau et al., 2022). Biostimulants display low or null toxicity and do not accumulate in long term (Sangiorgio et al., 2020). There are several available types of biostimulants, and they can be classified based on the source of raw material into six major groups, such as seaweed, plant extracts, protein hydrolysates, humic substances, inorganic compounds and microorganisms (Franzoni et al., 2021).
Seaweed and plant extracts
Biostimulants based on seaweeds are the most studied and commercialized (Cristiano et al., 2018). This group includes macroscopic and multicellular marine algae from several taxonomic groups, such as red, green, and brown algae (Bulgari et al., 2019). Seaweeds are used in agriculture as fertilizers since ancient times due to their positive effects on crops (Franzoni et al., 2022). These algae contain hormones (e.g., auxins and cytokinins), polyphenols (e.g., phloroglucinol and eckol), polysaccharides (e.g., fucoidan, laminaran, carrageenan and alginates) and kahydrin, a derivative of vitamin K1, that are growth promoters and activators of plant defense system (Ertani et al., 2018; Baltazar et al., 2021). Most of the products used in agriculture are based on the red (Lithothamnium calcareum, Kappaphycus alvarezii, Jania rubens, and Gracilaria edulis), green (Ulva lactuca) and brown (Ascophyllum nodosum, Ecklonia maxima, Durvillaea potatorum and Laminaria) algae (Van Oosten et al., 2017; Baltazar et al., 2021).
Some biostimulants can be obtained from extracts of different plant structures, leaves, fruits, stems, and flowers (Franzoni et al., 2022). These extracts are rich in bioactive compounds that can activate some physiological processes in plants, and thereby improve their performance (Zulfiqar et al., 2020). For example, root extracts of carrots and licorice have been shown to improve the growth of cowpea, beans, pear tree, and fennel. Also, leaf extracts from borage, aloe, garlic, and green tea improved growth and several physiological processes in various horticultural crops (such as lettuce, tomato, eggplants, guar, and olive trees) and ornamental plants (e.g., dwarf umbrella tree) (Ali et al., 2018; Elzaawely et al., 2018; Moreno-Hernández et al., 2020; Zulfiqar et al., 2020). Seagrass aqueous extracts mitigate the negative effects of salt stress in tomato plants, improving the activity of antioxidant enzymes (Vinoth et al., 2017). Moringa oleifera leaf extract is also used to increase plant growth in several crops, such as pumpkin, peas, and common bean (Del Buono, 2021). This beneficial action is associated with the high levels of minerals, carbohydrates, protein, amino acids, hormones, and antioxidant enzymes (Zulfiqar et al., 2020; Del Buono, 2021).
Protein hydrolysates, humic substances and inorganic compounds
Hydrolyzed proteins are a mixture of amino acids, polypeptides, and oligopeptides obtained by chemical, thermal and enzymatic hydrolyses of proteins from different matrices, like animal or plant (e.g., plant-derived biomass or animal epithelial and connective tissues) (Cristofano et al., 2021). Protein hydrolysates derived from by-products of agricultural and industrial activities are gaining popularity as they represent a sustainable and eco-friendly solution to many wastes and contribute to a circular economy (Colla et al., 2017). Most of the commercialized protein hydrolysate biostimulants are from animal-derived proteins, and they have a higher nitrogen content (9 to 16% DM) and a slower release rate (Cristiano et al., 2018). The amino acids proline and glycine are the most abundant components in protein hydrolysate biostimulants based on collagen, while the glutamic acid is more abundant in biostimulants derived from plant sources (Baroccio et al., 2017).
Another kind of biostimulants are the humic substances (HS) that comprise humic and fulvic acids, and they are the main components of lignites, soil and peat (Van Oosten et al., 2017). Humic substances are supramolecular structures with hydrophilic (-OH and -COOH groups) and hydrophobic portions, resulting from the association of small molecules by the decomposition processes of animals, plants, or from the metabolic activity of soil microbes (Franzoni et al., 2022; Popa et al., 2022).
Some inorganic compounds such as silicon, selenium, cobalt, sodium, and aluminum can promote plant growth and induce stress responses, but they are not essential for all plants (Du Jardin, 2015). They can be found in plants and soils in different forms, as inorganic salts and insoluble, like the amorphous silica (Franzoni et al., 2022).
Microorganisms
Microorganisms-based biostimulants are mainly comprised of bacteria, fungi, and arbuscular mycorrhizal fungi (AMF) that can be isolated from soils, plants, and organic materials (Baltazar et al., 2021; Del Buono, 2021). These biostimulants can be applied directly to the soils or seeds (Franzoni et al., 2021). These biostimulants interact directly with the plants, establishing a mutual symbiotic association or indirectly by improving nutrient availability to plants (Franzoni et al., 2021).
The plant growth-promoting bacteria (PGPB) include rhizobacteria, which colonize the rhizosphere, and free-living bacteria from the soil (Baltazar et al., 2021). The plant growth-promoting rhizobacteria (PGPR) are the most important group and include the genera Azotobacter, Arthrobacter, Azospirillum, Acinetobacter, Bacillus, Bradyrhizobium, Enterobacter, Pseudomonas, Rhodococcus, Rhizobium, Streptomyces, and Ochrobactrum (Rai et al., 2021). AMF can establish a symbiotic relationship with plants and are the most commonly used biostimulants (Du Jardin, 2015). This kind of biostimulant includes the Rhizophagus spp., Septoglycus viscosum, Claroideoglomus etunicatum and Claroideoglomus claroideum (Rai et al., 2021). Besides AMF, other fungal-based biostimulants are very used in sustainable agriculture production, such as the endophytic fungi Trichoderma spp. (Ascomycota) and Sebacinales (Basidiomycota) (Du Jardin, 2015).
Mechanism of action of biostimulants for mitigation of abiotic stresses and plant growth
Although several studies indicated that they act as priming agents (Shukla et al., 2019), the action modes of microbial and non-microbial biostimulants in plants are not well known yet. Priming, or stress hardening, triggers several molecular and physiological defense mechanisms that increase plants’ ability to defend themselves when exposed to various stresses. These molecular and cellular changes induced by these priming agents in plants are stored as primed memory (Nephali et al., 2020) and once primed, the plants will defend faster and stronger to cope with subsequent stresses.
Biostimulants activate and regulate several defense mechanisms through different action modes (Rai et al., 2021). The biostimulants can be applied directly in the leaves (foliar application) or/and in the soil near the root system. The biostimulants based on proteins and amino acids (e.g., proline and glycine) can penetrate the leaf tissues directly and enter into the cells. The protein hydrolysates-based biostimulants enter the plant cell via diffusion processes through membrane pores, along with energy costs (Yakhin et al., 2017). Whereas in the microorganism-based biostimulants, the hyphae penetrate the tissues and establish symbiotic or mycorrhizal associations. When biostimulants reach the leaves and/or roots they are translocated and distributed to the other parts of the plant (Rai et al., 2021). Within the plant, the mechanisms of action differ according to the type of biostimulants (nature and characteristics of the biostimulants).
Molecular, metabolic and physiological mechanisms
Several studies demonstrated that plant priming with microbial and/or non-microbial biostimulants results in enhanced plant defense response to stresses such as increased antioxidant enzymes activities, accumulation of polyphenols and osmolytes (Shukla et al., 2019; Vaseva et al., 2022). Table 1 represents an overview of the molecular, metabolic, and physiological mechanisms underlying biostimulants-induced abiotic stress alleviation.
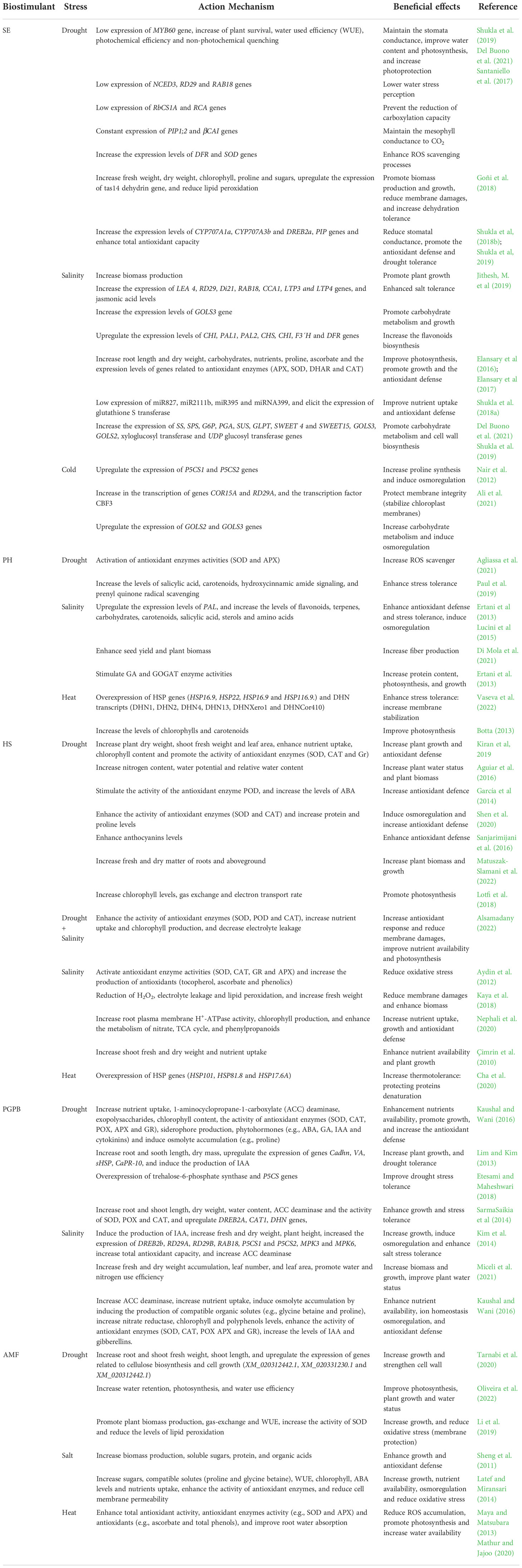
Table 1 Action mechanisms and beneficial effects of biostimulants (SE, seaweed; PH, protein hydrolysates; HS, humic substances; PGPB, plant growth promoting bacteria and AMF, arbuscular mycorrhizal fungi) in plants under drought, salinity, cold and heat stress.
Seaweed
The seaweed extracts, as one of the most used biostimulants, can be used to increase plant tolerance to various abiotic stresses (Shukla et al., 2019; Ali et al., 2021). Plant treatment with seaweed-based biostimulants can act in central metabolic pathways, involved in photosynthesis, activating the defense system, and increasing ROS scavenging levels (Shukla et al., 2019). Moreover, biostimulants based on A. nodosum promote growth, increasing Arabidopsis length, tomato plants fresh and dry weight, and seashore paspalum root length and dry weight (Elansary et al., 2017; Goñi et al., 2018; Shukla et al., 2018a; Jithesh et al., 2019). Concerning photosynthesis, several improvements achieved under stress conditions were related to seaweed priming. For instance, seaweed biostimulants based on A. nodosum are described to regulate stomatal conductance in Arabidopsis by modulating the expression of MYB60, NCED3, RK2, aquaporins (PIP1;2) and bCA1 genes and increase dehydration protection by activating the expression of several late embryogenesis abundant proteins (LEA, from group 2) and dehydrins genes (such as RAB18) under drought and salinity stress (Santaniello et al., 2017; Shukla et al., 2019). These changes induced by these biostimulants enable the plants to preserve water status and avoid dehydration, increase photosynthesis and water use efficiency (WUE), and protect proteins and enzymes under drought and salt stress conditions (Del Buono, 2021). Seaweed extracts of A. nodosum modulate the expression of genes involved in the biosynthesis of abscisic acid (ABA) (an increase of NCED3) in Glycine max L., resulting in less partial stomatal closure and higher WUE, and also in the catabolism of ABA by regulating the expression of CYP707A1a and CYP707A3b under drought stress (Shukla et al., 2018b). Moreover, other ABA-dependent genes, like Di-21, and lipid transfer protein (LTP3 and LTP4) are positively regulated by seaweed biostimulants extracts of A. nodosum, thus enhancing Arabidopsis tolerance to oxidative stress under salt conditions (Jithesh et al., 2019). Other phytohormones, like jasmonic acid, with an important role in plant stress defense, are also positively regulated by seaweed extracts (Jithesh et al., 2019).
Moreover, extracts from A. nodosum act directly at the carbon assimilation levels by adjusting the expression of RbCS1A and RCA genes, which are related to RuBisCO activation (Santaniello et al., 2017; González-Morales et al., 2021), protect the photosystem II through the regulation of the FIB1a expression, promote the biosynthesis carbohydrates such as sucrose, raffinose and starch (increase the expression of raffinose synthase (RS), GOLS2 and GOLS3, LHCB4, SS, SPS, G6P, PGA, SUS, GLPT and SWEET 4 and SWEET15), sugar alcohols (e.g. trehalose and myo-inositol), amino acids (e.g. proline and isoleucine), pigments and proteins, therefore increasing salinity, drought and low-temperature stress tolerance in Arabidopsis, tomato plants, wheat and sweet pepper (Goñi et al., 2018; Ali et al., 2021). The increased chlorophyll contents are linked to the upregulation of choline monooxygenase (CMO) and betaine aldehyde dehydrogenase (BADH), and also to the activation of protective mechanisms that reduce pigments degradation (Ali et al., 2021). Seaweed biostimulants based on A. nodosum positively regulate the expression of P5CS1 and P5CS2 genes linked to the biosynthesis of proline, and negatively regulate the expression of other genes related to proline catabolism in Arabidopssis (Rai et al., 2021), increasing plant tolerance to several types of abiotic stresses. Seaweed extracts (A. nodosum, K. alvarezii, E. maxima and Laminaria spp.) also improve nutrient uptake in maize, cottonwood, and mustard plants under drought and salinity stress (Shukla et al., 2019; Ali et al., 2021). Transcriptomic analysis indicated that A. nodosum extracts could increase S and P uptake under salt stress by modulating the expression of miR827, miR2111b, miR395 and miRNA399 in A. thaliana (Shukla et al., 2018a).
The mechanism of ROS scavenging activated by the seaweed involves both enzymatic antioxidants and non-enzymatic antioxidants. Seaweed biostimulants based on A. nodosum increase the activity of the antioxidant enzymes (e.g., CAT, SOD, and APX), and the production of antioxidants (e.g., ascorbate), leading to lower ROS accumulation and less membrane damage in plants (e.g., Phaseolus vulgaris and Paspalum vaginatum) under drought and salt stress (Elansary et al., 2016; Elansary et al., 2017; Shukla et al., 2019). Moreover, they down-regulate the expression levels of the GROWTH-REGULATING FACTOR7 (GRF7) gene resulting in an overexpression of the genes DREB2a and RD29 and interact with several transcription factors like COR47, COR15A, CCA1, NF-YA, AGF2 and LHY1 that provide drought stress tolerance in G. max (Shukla et al., 2018b). A. nodosum extracts alleviate the oxidative stress levels under salinity by eliciting the expression of glutathione S transferase, regulating the ath-miR398 that modulates the expression of the copper/zinc SOD (CSD1) gene (Shukla et al., 2018a) and stimulating the phenylpropanoid pathway and flavonoids biosynthesis, by increasing the expression of ammonia lyase 1 and 2 (PAL1 and PAL2), chalcone synthase (CHS) and isomerase (CHI), flavonoid 3´-hydroxylase (F3´H) and dihydroflavonol 4-reductase (DFR) genes in Arabidopsis (Jithesh et al., 2019). The glutathione S transferase also participates in the transport of flavonoids to vacuoles by ABC transporters, whose expression is induced by seaweeds (Jithesh et al., 2019).
Protein hydrolysates
The mode of action of PH involves activation of the antioxidant system and improvement of photosynthesis (Lucini et al., 2015; Elansary et al., 2017). Plant-derived PH (alfa-alfa derived protein hydrolysate and LISIVEG®) upregulate the expression levels of phenylalanine ammonia lyase (PAL) and induce the production of defense-related metabolites such as flavonoids, terpenes, carbohydrates, sterols, and amino acids to increase salt stress tolerance in lettuce and maize plants (Ertani et al., 2013; Lucini et al., 2015). Under drought conditions, plant-derived PH (GHI_16_VHL and Trainer®) improved tolerance to ROS through a coordinated action of salicylic acid, carotenoids, hydroxycinnamic amide signaling, and prenyl quinone radical scavenging (Paul et al., 2019) and/or by the activation of antioxidant enzyme (e.g., SOD and APX) in Solanum lycopersicum and Capsicum annuum (Agliassa et al., 2021). PH (LISIVEG® and GHI_16_VHL) also promote osmoregulation to increase tolerance to drought and salinity in C. annuum and Lactuca satuca (Lucini et al., 2015; Agliassa et al., 2021). Moreover, these plant-derived biostimulants (alfa-alfa derived protein hydrolysate and Kaishi®) act at the photosynthesis and growth level by increasing protein production, glutamine synthetase (GS) and glutamate synthase (GOGAT) activities to improve salinity tolerance (Ertani et al., 2013), and protein degradation protection and starch maintenance for thermotolerance in Zea mays (Vaseva et al., 2022). Under salinity stress, the animal-derived PH Stressal is reported to lower chloride uptake and translocation to aerial parts, therefore reducing leaf necrosis symptoms in Diospyros kaki (Visconti et al., 2015). Colla et al. (2015) and Carillo et al. (2019) reported for maize and spinate that the protective effect on the photosynthetic apparatus under drought conditions was due to an auxin-like activity effect of the PH (Trainer®). Also, a plant-derived PH (Terra-Sorb® Foliar) improve the photosynthetic efficiency under heat conditions, by upregulating the levels of chlorophylls and carotenoids in L. sativa and Lolium perenne (Botta, 2013). PH (Kaishi®)) induced the overexpression of several heat shock proteins (HSP), like HSP16.9 and HSP22 in leaves and HSP16.9 and HSP116.9 in roots, providing higher heat tolerance to maize through membrane stabilization (Vaseva et al., 2022). Furthermore, PH upregulate dehydrins (DHN) transcripts (DHN2, DHN4, DHN13, DHNXero1, and DHNCor410) in leaves and in roots (DHN1 and DHN4) under heat stress, thus increasing protection from protein denaturation and membrane fluidity (Vaseva et al., 2022).
The positive effects of these kinds of biostimulants under salinity stress are also visible in the production, boosting seed yield and plant residual biomass of Cannabis sativa, which is relevant in fiber production (Di Mola et al., 2021).
Humic substances
Concerning the HS, they trigger several molecular processes that lead to physiological responses in plants, conferring tolerance to several types of stresses. HS can induce overexpression of several families of HSP, such as HSP101, HSP81.8, and HSP17.6A, increasing Arabidopsis thermotolerance by helping in repairing processes of denatured proteins and in maintaining protein homeostasis (Cha et al., 2020). Moreover, HS (e.g., extracted from leonardites and vermicompost) stimulate nutrient uptake (N, P, K, Ca, Fe, Mg, S, Mn, and Cu), root plasma membrane H+-ATPase activity, chlorophyll production, and enhance the metabolism of nitrate, TCA cycle, and phenylpropanoids in several species (e.g., lettuce, bean, garden cress, maize and sweet pepper) under salt stress (Çimrin et al., 2010; Aydin et al., 2012; Kaya et al., 2018; Nephali et al., 2020). The application of HS (K-humate-Proxin 85) increases salinity tolerance by decreasing oxidative stress through the reduction of the levels of ROS (e.g., H2O2), MDA and cell membrane permeability in beans and maize plants (Aydin et al., 2012; Kaya et al., 2018). These benefits lead to maize biomass increase and salt stress tolerance (Kaya et al., 2018). Under drought and salinity conditions, nutrient uptake is also improved, and Mg increase promotes chlorophyll production, leading to higher photosynthesis in Vigna radiata (Alsamadany, 2022). The upregulation of drought stress tolerance genes, DREB2A, bZIP17 and HsfA6a is also modulated by HS application (Alsamadany, 2022). Moreover, HS (e.g., derived from vermicompost) prevent drought and salinity-induced oxidative stress by enhancing antioxidant enzymes activities (SOD, CAT, Gr, and APX), and osmoprotectants (proline) that reduce ROS levels in several species (e.g., Oryza sativa, Saccharum officinarum, Cucumis melo, Setaria italica and Vigna radiata) (García et al., 2014; Aguiar et al., 2016; Kıran et al., 2019; Shen et al., 2020; Alsamadany, 2022). HA exerted a drought stress-protective effect in O. sativa through a signaling mechanism independent of ABA, regulating the expression of tonoplast aquaporin (TIPs) genes, putatively triggered by chemical and physical interactions between HA and the root system (García et al., 2014). These biostimulants substances can also promote the plant water status by increasing the relative water content and water potential, as well as increase N content leading to higher biomass accumulation in S. officinarum and C. melo plants under drought conditions (Aguiar et al., 2016; Kıran et al., 2019). Matuszak-Slamani et al. (2022) also reported that HS (e.g., derived from vermicompost) increases several other morphological features in soybean, like fresh and plant dry matter of root and aboveground, and plant length under drought conditions. Furthermore, HS improve photosynthesis under water deficit conditions by stimulating gas exchange, electron transport rate, and chlorophyll content in Brassica napus (Lotfi et al., 2018), and promote vegetative and generative yield, as well as anthocyanins accumulation in Hibiscus sabdariffa (Sanjarimijani et al., 2016). Signaling pathways of several hormones such as ABA, gibberellins and auxins, and stress-responsive genes are positively modulated by HS, therefore alleviating salinity, drought, and heavy metal adverse effects in maize (Canellas et al., 2020).
Microorganisms
The beneficial effects of several bacterial and fungal strains on plant growth under stress conditions have been studied (Etesami and Maheshwari, 2018; Leontidou et al., 2020). Arbuscular mycorrhizal fungi (AMF) and plant growth-promoting bacteria (PGPB) are the most common microbial biostimulants that act in stimulating plant performance (Miceli et al., 2021).
Several PGPB, such as Azospirillum sp., Achromobacter sp., Aeromonas sp., Acetobacter sp., Bacillus sp., Bradyrhizobium sp., Chryseobacterium sp., Flavobacterium sp., Sinorhizobium sp. and Pseudomonas sp. promote plant growth and development under stress conditions, leading to higher crop yield (e.g., Arabidopsis thaliana, Pinus halepensis, Cucumis sativus, Oryza sativa and Phyllobacterium brassicacearum) (Kaushal and Wani, 2016). PGPB are able to enhance plant tolerance to drought and salinity through various mechanisms including: a) synthesis of the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase that promote plant growth by cleaving plant-produced ACC, and thereby lowering the ethylene level in plants; b) enhancement of availability of nutrients (e.g. N, P, K, Fe, Mn, Zn, Cu and B) by solubilization and mineralization (e.g., N fixation, P solubilization, and siderophores production); c) osmolyte accumulation by inducing the production of soluble sugars and compatible organic solutes (e.g., proline and glycine betaine); d) production of phytohormones (e.g., ABA, GA, IAA and cytokinins); e) production of volatile organic compounds by modulating the expression of genes related to cell wall structure, synthesis of choline and glycine betaine, and stomata closure; f) decrease oxidative stress through the scavenging of ROS mediated by antioxidant enzymes (SOD, CAT, APX and Gr) and antioxidant metabolites (ascorbate and phenolic compounds); and g) maintenance of ion homeostasis by increasing K+ and sustaining K+/N+ ratio (Hayat et al., 2010; Kim et al., 2014; Kaushal and Wani, 2016; Ertani et al., 2018; Leontidou et al., 2020; Miceli et al., 2021). Microarray analysis indicated that Rhizobium tropici and Paenibacillus polymyxa induce the overexpression of the trehalose-6-phosphate synthase (TPS) gene involved in P. vulgaris drought tolerance (Ertani et al., 2018). Also, Bacillus strains upregulate the expression of the P5CS genes involved in proline biosynthesis, leading to osmotic tolerance in maize and cucumber plants (Ertani et al., 2018). Pseudomonas aeruginosa, Paenibacillus polymyxa, and Bacillus licheniformi activate the expression of the genes ERD15, sHSP, CaPR-10, VA and Cadhn, conferring drought tolerance to pepper plants (Lim and Kim, 2013).
Plant symbiotic association with several AMF strains (e.g., Glomus spp, Rhizophagus spp, and Funneliformis spp) are described to improve several physiological processes in plants (e.g., wheat, barley, maize, soybean, strawberry, onion and olive tree), therefore increasing plant tolerance to stresses (Begum et al., 2019; Oliveira et al., 2022). The mechanisms by which AMF can enhance plant tolerance to drought and salinity involve a) oxidative stress mitigation through the upregulation of antioxidant enzymes (SOD and CAT) and metabolites (glutathione), and downregulation of lipoxygenase; b) improvement of nutrient availability (e.g. Mg2+ and N) through the facilitation of uptake and modifications of the root morphology; e) stomatal regulation by controlling ABA metabolism; f) increase of photosynthesis, chlorophyll production, leaf water relation and photosynthate translocation leading to better growth and plant water status; g) osmolyte accumulation by inducing the production of compatible organic solutes (e.g., proline and betaine); and h) adjust the levels of growth regulators (cytokinins) (Sheng et al., 2011; Latef and Miransari, 2014; Li et al., 2019; Begum et al., 2019; Diagne et al., 2020; Oliveira et al., 2022). The AMF Funneliformis mosseae modulate the expression of XM_020312442.1, XM_020331230.1, XM_020312442.1 genes related to cellulose biosynthetic process and cell growth, promoting drought tolerance in Triticum aestivum via strengthening cell wall and membrane (Tarnabi et al., 2020). Another AMF, like Funneliformis (Glomus spp.), can increase plant thermotolerance, through the enhancement of the total antioxidant activity, antioxidant enzymes activity (e.g., SOD and APX) and antioxidants (e.g., ascorbate and total phenols) that reduce ROS accumulation, and by helping Cyclamen persicum and Z. mays plants to improve water absorption to ensure higher photosynthetic capacity (Maya and Matsubara, 2013; Mathur and Jajoo, 2020).
Evidently, microorganisms play an essential role in environmental stress tolerance/resistance. However, designing and developing efficient commercial formulations of microbial inoculants has been the key factor determining the successful implementation of these beneficial microbes in agriculture (e.g., microbes could survive in the formulation products) (Bashan et al., 2014; Ma et al., 2020). Many bacterial genera such as Azotobacter, Azospirillum, Rhizobium, Bacillus, and Pseudomonas isolated from alkaline, saline, arid and acidic soils are found to have great potential to adapt to such adverse conditions and mitigate plant abiotic stress responses (in species like maize, wheat, pea, fava bean, chickpea, cotton, sorghum, potatoes, lettuce and sweet pepper), which can be served as biostimulants (Van Oosten et al., 2014). For instance, inoculation of certain microbial biostimulants via soil/seedling drench and seed coating is able to change plant cell wall composition by producing exopolysaccharides to form a protective biofilm on the root surface and accumulate high concentrations of solutes by synthesizing phytohormones (Egamberdieva et al., 2017), therefore increasing water retention and/or tolerance to salinity-induced ionic and osmotic stress in chickpea plants. The beneficial and protective functions of biostimulants on plants under diverse environmental stresses such as drought, salinity, and extreme temperatures are of vital importance for crops to survive in such environments. However, the development and selection of biostimulants should consider the physiological, biochemical, metabolic, and genetic mechanisms to cope with various stresses.
Implementation of biostimulants in agriculture under climatic stress
Biostimulants were initially used in France, Italy, and Spain, and were later extended to other EU countries, America, Australia, etc., to improve agricultural production and quality worldwide, particularly under climate change-induced abiotic stress (Li et al., 2022). In Europe, biostimulants have been used in many types of plants and crops, such as vegetable crops (e.g., Allium cepa, Capsicum annuum, Brassica oleracea, Solanum tuberosum, Cucumis sativus, Allium sativum, Solanum lycopersicum, Cucurbita pepo, Daucus carota, L. sativa, and Solanum melongena), fruit crops (e.g., Olea europaea, Prunus sp., Fragaria ananassa, Cucumis melo, Citrullus lanatus, Vitis vinifera, Citrus sinensis and Pyrus communis), grain crops (e.g., Hordeum vulgare, Triticum aestivum, Oryza sativa, and Zea mays), oil crops (e.g., Brassica napus and Glycine max), horticultural crops (e.g., flowers, nurseries, lawns), and ornamental plants. Biostimulants have been widely used at all stages of production of the above-mentioned crops under various climatic stresses including as seed treatments, as foliar sprays during growth, as well as on harvested products (Khan et al., 2020; El Boukhari et al., 2021; Sorrentino et al., 2021; Jacomassi et al., 2022).
Biostimulants have been shown to affect multiple metabolic processes in crops, such as respiration, photosynthesis, ion transport, redox reactions, DNA synthesis, etc. (Rai et al., 2021). Numerous studies and practices have shown that biostimulants are capable of improving the ability of crops to absorb and utilize mineral nutrients, the utilization rate of fertilizers and crop health, plant resistance to various stresses, and ultimately achieve enhanced crop yield and quality products. Table 2 presents the categorized biostimulants and their action mechanism and implementation in crop performance.
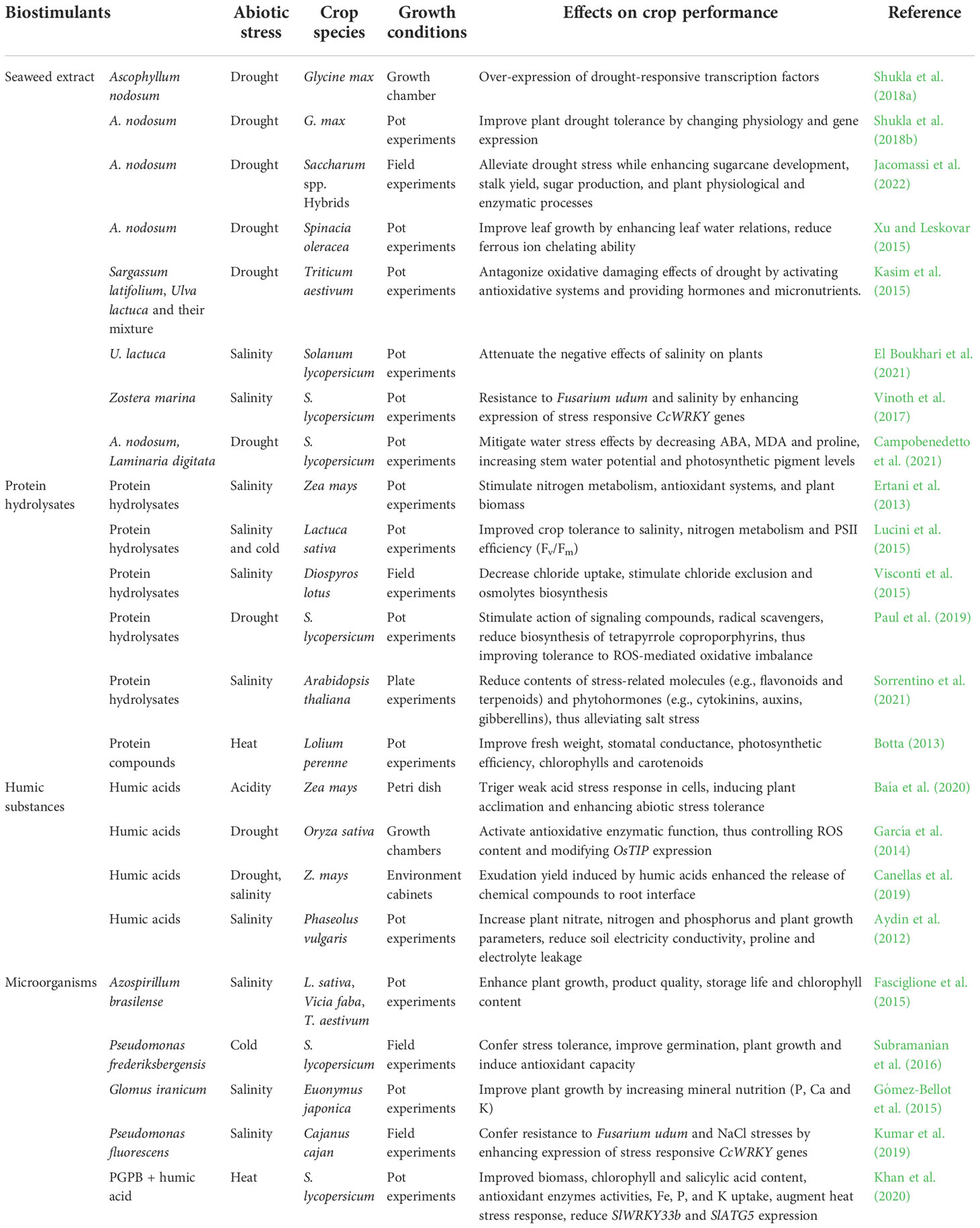
Table 2 From lab to field implementation of biostimulants in crop production.
Conclusion and future prospective
The current research on biostimulants has received unprecedented attention in both academia and industry. As mentioned above, biostimulants have been globally used in food, vegetables, fruit trees, flowers, nurseries, etc., to achieve high agricultural productivity and reduce the consequences/adverse effects of climate change and agrochemicals. The rapid development of research and application of biostimulants contributes significantly to sustainable agricultural practices. Although the efficiency of biostimulants has been widely recognized and the prospects for development are promising, there are still many problems to be solved, as follows:
1. Biostimulant product specification and standardization. There are a wide variety of biostimulants, and the market products are mixed and uneven. As they are different from traditional chemical pesticides and fertilizers, it is urgent for relevant national departments and industries to formulate relevant laws, regulations, policies, regulations, and technical standards/frameworks or guidelines. Moreover, a sustainable production system of biostimulants must be ensured. Despite the natural origin of most of the biostimulants (e.g., by-products of different value chains, natural and cheap resources) the production process must respect the circular economy and sustainability concept.
2. Lack of efficient production technology. Although all types of biostimulant products are currently produced and sold worldwide, most of them are primary products, and only a few high-end products are in the market as there is a lack of preparation technology for high-quality products (such as high purity and high activity) or the failure to achieve industrial application/implementation. Superior scientific research units and enterprises should speed up technological innovation in this field, deepen industry-university-research unit cooperation, and solve technical problems for further efficient production. For instance, more research on efficient techniques to improve and enlarge the shelf-life of most biostimulants is crucial.
3. The mechanism of action is still unclear, particularly under stress conditions. Due to the relatively complex composition of biostimulants, this feature determines that the target of its action mechanism is not very clear. Thus, the research on its action mechanism can be a long and complicated process. Some of the benefits demonstrated in plants growing under optimal conditions are not verified under stress conditions. Selecting a single compound with strong activity and clear structure from specific biostimulants to study the mechanism of action under specific environmental conditions (e.g., drought, salinity and extreme temperatures) could be a better model for in-depth research in this area and increase the knowledge of the biostimulants effects under climate change scenarios.
4. The application technologies are still largely unrealized. The concept of biostimulants comes from practical applications, however, there are also problems with specific application technologies. Through field trials, technology popularization, and application, it is an important guarantee for its industrial application to clarify the use of various biostimulants for a variety of crops and diverse conditions in different regions considering climate change scenarios. This requires normative experiments and regular summaries from scientific researchers and related practitioners.
All the mentioned problems will be gradually solved under the background of public needs and incentives, national requirements, and the attention of the industry. Recently, the research on biostimulants has become a hot spot in the field of plant protection and the use of biostimulants as an alternative to pesticides and fertilizers plays an important role in developing sustainable agriculture. In addition, all future improvements in the application of biostimulants for modern agriculture, e.g., the development of new products and/or upgrades, production, marketing and distribution of commercial biostimulants, could implement a sustainable strategy to improve plant tolerance/resistance against such environmental limitations, which is of great importance to secure and optimize global agricultural production under climate change scenarios.
Author contributions
YM developed the ideas and wrote and revised the manuscript. MD wrote and revised the manuscript. HF was the project sponsor, and revised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work is carried out at the R&D Unit Center for Functional Ecology - Science for People and the Planet (CFE), with reference UIDB/04004/2020, financed by FCT/MCTES through national funds (PIDDAC). The FCT supported the research contracts of YM (SFRH/BPD/76028/2011) and MC Dias (SFRH/BPD/100865/2014).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abd El-Salam, M. H., El-Shibiny, S. (2017). Preparation, properties, and uses of enzymatic milk protein hydrolysates. Crit. Rev. Food Sci. Nutr. 57 (6), 1119–1132. doi: 10.1080/10408398.2014.899200
Agliassa, C., Mannino, G., Molino, D., Cavalletto, S., Contartese, V., Bertea, C. M. (2021). Francesca Secchi a new protein hydrolysate-based biostimulant applied by fertigation promotes relief from drought stress in Capsicum annuum l. Plant Physiol. Biochem. 166, 1076–1086. doi: 10.1016/j.plaphy.2021.07.015
Aguiar, N. O., Medici, L. O., Olivares, F. L., Dobbss, L. B., Torres-Netto, A., Silva, S. F., et al. (2016). Metabolic profile and antioxidant responses during drought stress recovery in sugarcane treated with humic acids and endophytic diazotrophic bacteria. Ann. Appl. Biol. 168, 203–213. doi: 10.1111/aab.12256
Ahluwalia, O., Singh, P. C., Bhatia, R. (2021). A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resources Environ. Sustainability 5, 100032. doi: 10.1016/j.resenv.2021.100032
Ahmed, I. M., Dai, H., Zheng, W., Cao, F., Zhang, G., Sun, D., et al. (2013). Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiol. Biochem. 63, 49–60. doi: 10.1016/j.plaphy.2012.11.004
Ali, O., Ramsubhag, A., Jayaraman, J. (2018). Ascophyllum nodosum (Linnaeus) le jolis seaweed extract improves seed germination in tomato and sweet pepper under NaCl-induced salt stress. Trop. Agric. 95, 141–148.
Ali, O., Ramsubhag, A., Jayaraman, J. (2021). Biostimulant properties of seaweed extracts in plants: implications towards sustainable crop production. Plants 10, 531. doi: 10.3390/plants10030531
Alsamadany, H. (2022). Physiological, biochemical and molecular evaluation of mungbean genotypes for agronomical yield under drought and salinity stresses in the presence of humic acid. Saudi J. Biol. Sci. 9, 103385. doi: 10.1016/j.sjbs.2022.103385
Aydin, A., Kant, C., Turan, M. (2012). Humic acid application alleviates salinity stress of bean (Phaseolus vulgaris l.) plants decreasing membrane leakage. Afr. J. Agric. Res. 7, 1073–1086. doi: 10.5897/AJAR10.274
Baía, D. C., Olivares, F. L., Zandonadi, D. B., Soares, C. P., Spaccini, R., Canellas, L. P., et al. (2020). Humic acids trigger the weak acids stress response in maize seedlings. Chem. Biol. Technol. Agric. 7, 31. doi: 10.1186/s40538-020-00193-5
Baltazar, M., Correia, S., Guinan, K. J., Sujeeth, N., Bragança, R., Gonçalves, B. (2021). Recent Advances in the Molecular Effects of Biostimulants in Plants: An Overview. Biomolecules 11, 1096. doi: 10.3390/biom11081096
Baroccio, F., Barilaro, N., Tolomei, P. (2017). Classification of biostimulants origin using amino acids composition of hydrolyzed proteins. J. Hortic. Sci. Res. 1, 30–35. doi: 10.36959/745/395
Bashan, Y., de-Bashan, L. E., Prabhu, S. R., Herbandez, J. P. (2014). Advances in plant growth-promoting bacterial inoculant technology: formulations and practical perspective(1998–2013). Plant Soil 378, 1–33. doi: 10.1007/s11104-013-1956-x
Begum, N., Qin, C., Ahanger, M. A., Raza, S., Khan, M. I., Ashraf, M., et al. (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01068
Botta, A. (2013). Enhancing plant tolerance to temperature stress with amino acids: an approach to their mode of action. Acta Hortic. 1009, 29–35. doi: 10.17660/ActaHortic.2013.1009.1
Bulgari, R., Franzoni, G., Ferrante, A. (2017). Biostimulants Application in Horticultural Crops under Abiotic Stress Conditions. Agronomy 9, 306. doi: 10.3390/agronomy9060306
Campobenedetto, C., Mannino, G., Agliassa, C., Acquadro, A., Contartese, V., Garabello, C., et al. (2021). Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants 9, 1308. doi: 10.3390/plants9101308
Canellas, L. P., Canellas, N. O. A., Soares, T. S., Olivares, F. L. (2019). Humic Acids Interfere with Nutrient Sensing in Plants Owing to the Differential Expression of TOR. J. Plant Growth Regul. 38, 216–224. doi: 10.1007/s00344-018-9835-6
Canellas, L. P., Canellas, N. A. O., da S Irineu, L. E. S., Olivares, F. L., Piccolo, A. (2020). Plant chemical priming by humic acids. Chem. Biol. Technol. Agric. 7, 12. doi: 10.1186/s40538-020-00178-4
Carillo, P., Colla, G., Fusco, G. M., Dell’Aversana, E., El-Nakhel, C., Giordano, M., et al. (2019). Morphological and physiological responses induced by protein hydrolysate-based biostimulant and nitrogen rates in greenhouse spinach. Agronomy 8, 450. doi: 10.3390/agronomy9080450
Cha, J.-Y., Kang, S.-H., Ali, I., Lee, S. C., Ji, M. G., Jeong, S. Y., et al. (2020). Humic acid enhances heat stress tolerance via transcriptional activation of Heat-Shock proteins in Arabidopsis. Sci. Rep. 10, 15042. doi: 10.1038/s41598-020-71701-8
Çimrin, K. M., Türkmen, Ö., Turan, M., Tuncer, B. (2010). Phosphorus and humic acid application alleviate salinity stress of pepper seedling. Afr. J. Biotechnol. 9, 5845–5851.
Colla, G., Hoagland, L., Ruzzi, M., Cardarelli, M., Bonini, P., Canaguier, R., et al. (2017). Biostimulant action of protein hydrolysates: unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02202
Colla, G., Rouphael, Y., Di Mattia, E., El-Nakhel, C., Cardarelli, M. (2015). Co-Inoculation of glomus intraradices and trichoderma atroviride acts as a biostimulant to promote growth, yield and nutrient uptake of vegetable crops. J. Sci. Food Agric. 95, 1706–1715. doi: 10.1002/jsfa.6875
Cristiano, G., Pallozzi, E., Conversa, G., Tufarelli, V., De Lucia, B. (2018). Effects of an animal-derived biostimulant on the growth and physiological parameters of potted snapdragon (Antirrhinum majus l.). Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00861
Cristofano, F., El-Nakhel, C., Rouphael, Y. (2021). Biostimulant substances for sustainable agriculture: Origin, operating mechanisms and effects on Cucurbits, leafy greens, and nightshade vegetables species. Biomolecules 11, 1103. doi: 10.3390/biom11081103
Daliakopoulos, I. N., Tsanis, I. K., Koutroulis, A., Kourgialas, N. N., Varouchakis, A. E., Karatzas, G. P., et al. (2016). The threat of soil salinity: A European scale review. Sci. Total Environ. 573, 727–739. doi: 10.1016/j.scitotenv.2016.08.177
Del Buono, D. (2021). Can biostimulants be used to mitigate the effect of anthropogenic climate change on agriculture? It is time to respond. Sci. Total Environ. 751, 141763. doi: 10.1016/j.scitotenv.2020.141763
Delitte, M., Caulier, S., Bragard, C., Desoignies, N. (2021). Plant microbiota beyond farming practices: a review. Front. Sustain. Food Syst. 5. doi: 10.3389/fsufs.2021.624203
de Oliveira, J. M. P. F., Santos, C., Araújo, M., Oliveira, M., Dias, M. C. (2022). High-salinity activates photoprotective mechanisms in Quercus suber via accumulation of carbohydrates and involvement of non-enzymatic and enzymatic antioxidant pathways. New Forests 53, 285–300. doi: 10.1007/s11056-021-09856-z
Di Mola, I., Conti, S., Cozzolino, E., Melchionna, G., Ottaiano, L., Testa, A., et al. (2021). Plant-based protein hydrolysate improves salinity tolerance in Hemp: agronomical and physiological aspects. Agronomy 11, 342. doi: 10.3390/agronomy11020342
Diagne, N., Ngom, M., Djighaly, P. I., Fall, D., Hocher, V., Svistoonoff, S. (2020). Roles of arbuscular mycorrhizal fungi on plant growth and performance: importance in biotic and abiotic stressed regulation. Diversity 12, 370. doi: 10.3390/d12100370
Dias, M. C., Pinto, D. C. G. A., Correia, C., Moutinho-Pereira, J., Oliveira, H., Freitas, H., et al. (2018). UV-B radiation modulates physiology and lipophilic metabolite profile in Olea europaea. J. Plant Physiol. 222, 39–50. doi: 10.1016/j.jplph.2018.01.004
Dias, M. C., Santos, C., Araújo, M., Barros, P. M., Oliveira, M., de Oliveira, J. M. P. F. (2022). Quercus suber roots activate antioxidant and membrane protective prcesses in response to high salinity. Plants 11, 557. doi: 10.3390/plants11040557
Du Jardin, P. (2015). Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hort. 196, 3–14. doi: 10.1016/j.scienta.2015.09.021
Dusenge, M. E., Duarte, A. G., Way, D. A. (2019). Plant carbon metabolism and climate change: Elevated CO2 and temperature impacts on photo-synthesis, photorespiration and respiration. New Phytol. 221, 32–49. doi: 10.1111/nph.15283
Egamberdieva, D., Wirth, S. J., Shurigin, V. V., Hashem, A., Abd-Allah, E. F. (2017). Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum l.) and induce suppression of root rot caused by fusarium solani under salt stress. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.01887
Elansary, H. O., Skalicka-Woźniak, K., King, I. W. (2016). Enhancing stress growth traits as well as phytochemical and antioxidant contents of spiraea and pittosporum under seaweed extract treatments. Plant Physiol. Biochem. 105, 310–320. doi: 10.1016/j.plaphy.2016.05.024
Elansary, H. O., Yessoufou, K., Abdel-Hamid, A. M. E., El-Esawi, M. A., Ali, H. M., Elshikh, M. S. (2017). Seaweed extracts enhance salam turfgrass performance during prolonged irrigation intervals and saline shock. Front. Plant Sci. 12. doi: 10.3389/fpls.2017.00830
El Boukhari, M. E. M., Barakate, M., Choumani, N., Bouhia, Y., Lyamlouli, K. (2021). Ulva lactuca extract and fractions as seed priming agents mitigate salinity stress in tomato seedlings. Plants 10 (6), 1104. doi: 10.3390/plants10061104
Elzaawely, A. A., Ahmed, M. E., Maswada, H. F., Al-Araby, A. A., Xuan, T. D. (2018). Growth traits, physiological parameters and hormonal status of snap bean (Phaseolus vulgaris l.) sprayed with garlic cloves extract. Arch. Agron. Soil Sci. 64, 1068–1082. doi: 10.1080/03650340.2017.1410543
Ertani, A., Francioso, O., Tinti, A., Schiavon, M., Pizzeghello, D., Nardi, S. (2018). Evaluation of seaweed extracts from Laminaria and Ascophyllum nodosum spp. as biostimulants in Zea mays l. using a combination of chemical, biochemical and morphological approaches. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00428
Ertani, A., Schiavon, M., Muscolo, A., Nardi, S. (2013). Alfalfa plant-derived biostimulant stimulate short-term growth of salt stressed Zea mays l. plants. Plant Soil 364, 145–158. doi: 10.1007/s11104-012-1335-z
Escobar-Bravo, R., Klinkhamer, P. G. L., Leiss, K. A. (2017). Interactive effects of UV-b light with abiotic factors on plant growth and chemistry, and their consequences for defense against arthropod herbivores. Front. Plant Sci. 8, 278. doi: 10.3389/fpls.2017.00278
Etesami, H., Maheshwari, D. K. (2018). Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol Environ. Saf. 156, 225–246. doi: 10.1016/j.ecoenv.2018.03.013
Fasciglione, G., Casanovas, E. M., Quillehauquy, V., Yommi, A. K., Goñi, M. G., Roura, S. I., et al. (2015). Azospirillum inoculation effects on growth, product quality and storage life of lettuce plants grown under salt stress. Sci. Hortic. 195, 154–162. doi: 10.1016/j.scienta.2015.09.015
Franzoni, G., Cocetta, G., Prinsi, B., Ferrante, A., Espen, L. (2022). Biostimulants on crops: Their impact under abiotic stress conditions. Horticulturae 8, 189. doi: 10.3390/horticulturae8030189
Franzoni, G., Bulgari, R., Ferrante, A. (2021). Maceration time affects the efficacy of borage extracts as potential biostimulant on rocket salad. Agronomy 11 (11), 2182. doi: 10.3390/agronomy11112182
García, A. C., Santos, L. A., Guridi-Izquierdo, F., Rumjanek, V. M., Castro, R. N., Santos, F. S., et al. (2014). Potentialities of vermicompost humic acids to alleviate water stress in rice plants (Oryza sativa l.). J. Geochem Explor. 136, 48–54. doi: 10.1016/j.gexplo.2013.10.005
Gómez-Bellot, M. J., Ortuño, M. F., Nortes, P. A., Vicente-Sánchez, J., Bañón, S., Sánchez-Blanco, M. J. (2015). Mycorrhizal euonymus plants and reclaimed water: biomass, water status and nutritional responses. Sci. Hortic. 186, 61–69. doi: 10.1016/j.scienta.2015.02.022
Goñi, G., Quille, P., O'Connell, S. (2018). Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Bio. 126, 63–73. doi: 10.1016/j.plaphy.2018.02.024
González-Morales, S., Solís-Gaona, S., Valdés-Caballero, M. V., Juárez-Maldonado, A., Loredo-Treviño, A., Benavides-Mendoza, A. (2021). Transcriptomics of Biostimulation of Plants Under Abiotic Stress. Front. Genet. 12, 583888. doi: 10.3389/fgene.2021.583888
Haghighi, M., Pessarakli, M. (2013). Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hortic. 161, 111–117. doi: 10.1016/j.scienta.2013.06.034
Hayat, R., Ali, S., Amara, U., Khalid, R., Ahmed, I. (2010). Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 60, 579–598. doi: 10.1007/s13213-010-0117-1
Jacomassi, L. M., Viveiros, J. O., Oliveira, M. P., Momesso, L., de Siqueira, G. F., Crusciol, C. A. C. (2022). A seaweed extract-based biostimulant mitigates drought stress in sugarcane. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.865291
Jagadish, K. S. V., Way, D. A., Sharkey, T. D. (2021). Plant heat stress: Concepts directing future research. Plant Cell Environ. 44, 1992–2005. doi: 10.1111/pce.14050
Jithesh, ,M.N., Shukla, P. S., Kant, P., Joshi, J., Critchley, A. T., Prithiviraj, B. (2019). Physiological and transcriptomics analyses reveal that Ascophyllum nodosum extracts induce salinity tolerance in arabidopsis by regulating the expression of stress responsive genes. J. Plant Growth Regul. 38, 463–478. doi: 10.1007/s00344-018-9861-4
Kasim, W. A., Hamada, E. A. M., Shams El-Din, N. G., Eskander, S. K. (2015). Influence of seaweed extracts on the growth, some metabolic activities and yield of wheat grown under drought stress. Int. J. Agron. Agric. Res. 7, 173–189.
Kaushal, M., Wani, S. P. (2016). Plant-growth-promoting rhizobacteria: drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 66, 35–42. doi: 10.1007/s13213-015-1112-3
Kaya, C., Akram, N. A., Ashraf, M., Sonmez, O. (2018). Exogenous application of humic acid mitigates salinity stress in maize (Zea mays l.) plants by improving some key physico-biochemical attributes. Cereal Res. Comm 46 (1), 67–78. doi: 10.1556/0806.45.2017.064
Khan, M. A., Asaf, S., Khan, A. L., Jan, R., Kang, S.-M., Kim, K.-M., et al. (2020). Extending thermotolerance to tomato seedlings by inoculation with SA1 isolate of Bacillus cereus and comparison with exogenous humic acid application. PloS One 15 (4), e0232228. doi: 10.1371/journal.pone.0232228
Kim, K., Jang, Y.-J., Lee, S.-M., Oh, B.-T., Chae, J.-C., Lee, K.-J. (2014). Alleviation of salt stress by Sp. EJ01 in tomato and Is accompanied by up-regulation of conserved salinity responsive factors in plants. Mol. Cells 37 (2), 109–117. doi: 10.14348/molcells.2014.2239
Kıran, S., Furtana, G. B., Talhouni, M., Ellialtıoğlu, S. S. (2019). Drought stress mitigation with humic acid in two Cucumis melo l. genotypes differ in their drought tolerance. Bragantia Campinas. 78 (4), 490–497. doi: 10.1590/1678-4499.20190057
Kumar, G., Bajpai, R., Sarkar, A., Mishra, R. K., Gupta, V. K., Singh, H. B., et al. (2019). Identification, characterization and expression profiles of fusarium udum stress-responsive WRKY transcription factors in cajanus cajan under the influence of NaCl stress and. Pseudomonas fluorescens OKC. Sci. Rep. 9, 14344. doi: 10.1038/s41598-019-50696-x
Latef, A. A., Miransari, M. (2014). “The role of arbuscular mycorrhizal fungi in alleviation of salt stress,” in Use of microbes for the alleviation of soil stresses (New York, USA: Springer. Science+Business Media), 23–38. doi: 10.1007/978-1-4939-0721-2_2
Lau, S. E., Teo, W. F. A., Teoh, E. Y., Tan, B. C. (2022). Microbiome engineering and plant biostimulants for sustainable crop improvement and mitigation of biotic and abiotic stresses. Discov. Food 2, 9. doi: 10.1007/s44187-022-00009-5
Leontidou, K., Genitsaris, S., Papadopoulou1, A., Kamou, N., Bosmali, I., Matsi, T., et al. (2020). Plant growth promoting rhizobacteria isolated from halophytes and drought-tolerant plants: genomic characterisation and exploration of phyto-benefcial traits. Sci. Rep. 10, 14857. doi: 10.1038/s41598-020-71652-0
Li, J., Meng, B., Chai, H., Yang, X., Song, W., Li, S., et al. (2019). Arbuscular mycorrhizal fungi alleviate drought stress in C3 (Leymus chinensis) and C4 (Hemarthria altissima) grasses via altering antioxidant enzyme activities and photosynthesis. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00499
Lim, J. H., Kim, S. D. (2013). Induction of drought stress resistance by multi-functional PGPR bacillus licheniformis K11 in pepper. Plant Pathol. J. 29, 201–208. doi: 10.5423/PPJ.SI.02.2013.0021
Li, J., Van Gerrewey, T., Geelen, D. (2022). A meta-analysis of biostimulant yield effectiveness in field trials. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.836702
Lotfi, R., Kalaji, H. M., Valizadeh, G. R., Khalilvand Behrozyar, E., Hemati, A., Gharavi-kochebagh, P., et al. (2018). Effects of humic acid on photosynthetic efficiency of rapeseed plants growing under different watering conditions. Photosynthetica 56 (3), 962–970. doi: 10.1007/s11099-017-0745-9
Lucini, L., Rouphael, Y., Cardarelli, M., Canaguier, R., Kumar, P., Colla, G. (2015). The effect of a plant-derived biostimulant on metabolic profiling and crop performance of lettuce grown under saline conditions. Sci. Hortic. 182, 124–133. doi: 10.1016/j.scienta.2014.11.022
Ma, Y., Dias, M. C., Freitas, H. (2020). Drought and salinity stress responses and microbe-induced tolerance in plants. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.591911
Manzoor, S., Rahman, M. H. U., Haider, G., Ghafoor, I., Ahmad, S., Afzal, M., et al. (2021). Biochar and slow release nitrogen fertilizer improve growth, yield, NUE, fiber quality of cotton and reduce nitrogen losses under arid climatic conditions. Environ. Sci. pollut. Res. 11, 1–14. doi: 10.1007/s11356-021-16576-6
Mathur, S., Jajoo, A. (2020). Arbuscular mycorrhizal fungi protects maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crop Prod. 143, 111934. doi: 10.1016/j.indcrop.2019.111934
Matuszak-Slamani, R., Bejger, R., Włodarczyk, M., Kulpa, D., Sienkiewicz, M., Gołebiowska, D., et al. (2022). Effect of humic acids on soybean seedling growth under polyethylene-Glycol-6000-Induced drought stress. Agronomy 12, 1109. doi: 10.3390/agronomy12051109
Maya, M. A., Matsubara, Y.-I. (2013). Influence of arbuscular mycorrhiza on the growth and antioxidative activity in cyclamen under heat stress. Mycorrhiza 23, 381–90. doi: 10.1007/s00572-013-0477-z
Miceli, A., Moncada, A., Vetrano, F. (2021). Use of microbial biostimulants to increase the salinity tolerance of vegetable transplants. Agronomy 11, 1143. doi: 10.3390/agronomy11061143
Moreno-Hernández, J. M., Benítez-García, I., Mazorra-Manzano, M. A., Ramírez-Suárez, J. C., Sánchez, E. (2020). Strategies for production, characterization and application of protein-based biostimulants in agriculture: A review. Chil. J. Agric. Res. 80, 274–289. doi: 10.4067/S0718-58392020000200274
Nair, P., Kandasamy, S., Zhang, J., Ji, X., Kirby, C., Benkel, B., et al. (2012). Transcriptional and metabolomic analysis of Ascophyllum nodosum mediated freezing tolerance in arabidopsis thaliana. BMC Genom. 13, 1–23. doi: 10.1186/1471-2164-13-643
Nephali, L., Piater, L. A., Dubery, I. A., Patterson, V., Huyser, J., Burgess, K., et al. (2020). Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 10, 505. doi: 10.3390/metabo10120505
Oleńska, E., Małek, W., Wójcik, M., Swiecicka, I., Thijs, S., Vangronsveld, J. (2020). Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: a methodical review. Sci. Total Environ. 743, e14068. doi: 10.1016/j.scitotenv.2020.140682
Oliveira, T. C., Cabral, J. S. R., Santana, L. R., Tavares, G. G., Santos, L. D. S., Paim, T. P., et al. (2022). The arbuscular mycorrhizal fungus Rhizophagus clarus improves physiological tolerance to drought stress in soybean plants. Sci. Rep. 12, 9044. doi: 10.1038/s41598-022-13059-7
Paul, K., Sorrentino, M., Lucini, L., Rouphael, Y., Cardarelli, M., Bonini, P., et al. (2019). A combined phenotypic and metabolomic approach for elucidating the biostimulant action of a plant-derived protein hydrolysate on tomato grown under limited water availability. Front. Plant Sci. 10, 1–18. doi: 10.3389/fpls.2019.00493
Popa, D. G., Lupu, C., Constantinescu-Aruxandei, D., Oancea, F. (2022). Humic substances as microalgal biostimulants — implications for microalgal biotechnology. Mar. Drugs 20, 327. doi: 10.3390/md20050327
Rai, N., Rai, S. P., Sarma, B. K. (2021). Prospects for abiotic stress tolerance in crops utilizing phyto- and bio-stimulants. Front. Sustain. Food Syst. 5, 54853. doi: 10.3389/fsufs.2021.754853
Rivero, R. M., Mestre, T. C., Mittler, R., Rubio, F., Garcia-Sanchez, F., Martinez, V. (2014). The combined effect of salinity and heat reveals a specific physiological, biochemical and molecular response in tomato plants. Plant Cell Environ. 37, 1059–1073. doi: 10.1111/pce.12199
Rouphael, Y., Colla, G. (2020). Editorial: Biostimulants in Agriculture. Front. Plant Sci. 11, 40. doi: 10.3389/fpls.2020.00040
Saddiq, M. S., Afzal, I., Iqbal, S., Hafeez, M. B., Raza, A. (2021). Low sodium content in leaves improves grain yield and physiological performance of wheat genotypes in saline-sodic soil. Trop. Agric. Res. / Pesqu. Agropec. Trop. 51, e67663. doi: 10.1590/1983-40632021v5167663
Sangiorgio, D., Cellini, A., Donati, I., Pastore, C., Onofrietti, C., Spinelli, F. (2020). Facing climate change: Application of microbial biostimulants to mitigate stress in horticultural crops. Agronomy 10 (6), 794. doi: 10.3390/agronomy10060794
Sanjarimijani, M., Sirousmehr, A., Fakheri, B. (2016). The effects of drought stress and humic acid on morphological traits, yield and anthocyanin of roselle (Hibiscus sabdariffa l.). Agroecology 8, 346–358.
Santaniello, A., Scartazza, A., Gresta, F., Loreti, E., Biasone, A., Di Tommaso, D., et al. (2017). Ascophyllum nodosum seaweed extract alleviates drought stress in arabidopsis by affecting photosynthetic performance and related gene expression. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01362
Sarma, R. K., Saikia, R. (2014). Alleviation of drought stress in mung bean by strain Pseudomonas aeruginosa GGRJ21. Plant Soil 377, 111–126. doi: 10.1007/s11104-013-1981-9
Savvides, A., Ali, S., Tester, M., Fotopoulos, V. (2016). Chemical priming of plants against multiple abiotic stresses: mission possible? Trends Plant Sci. 21, 329–340. doi: 10.1016/j.tplants.2015.11.003
Shah, A., Nazari, M., Antar, M., Msimbira, L. A., Naamala, J., Lyu, D., et al. (2021). PGPR in agriculture: a sustainable approach to increasing climate change resilience. Front. Sustain. Food Syst. 5. doi: 10.3389/fsufs.2021.667546
Sharma, A., Shahzad, B., Kumar, V., Kohli, S. K., Sidhu, G. P. S., Bali, A. S., et al. (2019). Phytohormones regulate accumulation of osmolytes under abiotic stress. Biomolecules 9, 285. doi: 10.3390/biom9070285
Sheng, M., Tang, M., Zhang, F., Huang, Y. (2011). Influence of arbuscular mycorrhiza on organic solutes in maize leaves under salt stress. Mycorrhiza 21, 423–430. doi: 10.1007/s00572-010-0353-z
Shen, J., Guo, M., Wang, Y., Yuan, X., Wen, Y., Song, X., et al. (2020). Humic acid improves the physiological and photosynthetic characteristics of millet seedlings under drought stress. Plant Sig. Behav. 15, 1774212. doi: 10.1080/15592324.2020.1774212
Shukla, P. S., Borza, T., Critchley, A. T., Hiltz, D., Norrie, J., Prithiviraj, B. (2018a). Ascophyllum nodosum extract mitigates salinity stress in Arabidopsis thaliana by modulating the expression of miRNA involved in stress tolerance and nutrient acquisition. PloS One 13, e0206221. doi: 10.1371/journal.pone.0206221
Shukla, P. S., Mantin, E. G., Adil, M., Bajpai, S., Critchley, A. T., Prithiviraj, B. (2019). Ascophyllum nodosum-based biostimulants: Sustainable applications in agriculture for the stimulation of plant growth, stress tolerance, and disease management. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00655
Shukla, P. S., Shotton, K., Norman, E., Neily, W., Critchley, A. T., Prithiviraj, B. (2018b). Seaweed extract improve drought tolerance of soybean by regulating stress-response genes. AoB Plants 10 (1), plx051. doi: 10.1093/aobpla/plx051
Silva, S., Dias, M. C., Silva, A. M. S. (2022). Titanium and zinc based nanomaterials in agriculture: a promising approach to deal with (a)biotic stresses? Toxics 10, 172. doi: 10.3390/toxics10040172
Srivastava, A. K., Suresh Kumar, J., Suprasanna, P. (2021). Seed ‘primeomics': plants memorize their germination under stress. Biol. Rev. 96, 1723–1743. doi: 10.1111/brv.12722
Sorrentino, M., Panzarová, K., Spyroglou, I., Spíchal, L., Buffagni, V., Ganugi, P., et al. (2022). Integration of Phenomics and Metabolomics Datasets Reveals Different Mode of Action of Biostimulants Based on Protein Hydrolysates in Lactuca sativa L. and Solanum lycopersicum L. Under Salinity. Front. Plant Sci. 12, 808711. doi: 10.3389/fpls.2021.808711
Subramanian, P., Kim, K., Krishnamoorthy, R., Mageswari, A., Selvakumar, G., Sa, T. (2016). Cold stress tolerance in psychrotolerant soil bacteria and their conferred chilling resistance in tomato (Solanum lycopersicum mill.) under low temperatures. PloS One 11, e0161592. doi: 10.1371/journal.pone.0161592
Suzuki, N., Bassil, E., Hamilton, J. S., Inupakutika, M. A., Zandalinas, S. I., Tripathy, D., et al. (2016). ABA is required for plant acclimation to a combination of salt and heat stress. PloS One 11 (1), e0147625. doi: 10.1371/journal.pone.0147625
Tarnabi, Z. M., Iranbakhsh, A., Mehregan, I., Ahmadvand, R. (2020). Impact of arbuscular mycorrhizal fungi (AMF) on gene expression of some cell wall and membrane elements of wheat (Triticum aestivum l.) under water deficit using transcriptome analysis. Physiol. Mol. Biol. Plants 26 (1), 143–162. doi: 10.1007/s12298-019-00727-8
Tigchelaar, M., Battisti, D. S., Naylor, R. L., Ray, D. K. (2018). Future warming increases probability of globally synchronized maize production shocks. Proc. Natl. Acad. Sci. 115, 6644–6649. doi: 10.1073/pnas.1718031115
Van Oosten, M. J., Pepe, O., De Pascale, S., Silletti, S., Maggio, A. (2017). The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 4, 1–12. doi: 10.1186/s40538-017-0089-5
Van Oosten, M. J., Pepe, O., De Pascale, S., Silletti, S., MAggio, A. (2014). The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 4, 5. doi: 10.1186/s40538-017-0089-5
Vargas, Y., Mayor-Duran, V. M., Buendia, H. F., Ruiz-Guzman, H., Raatz, B. (2021). Physiological and genetic characterization of heat stress effects in a common bean RIL population. PloS One 16, e0249859. doi: 10.1371/journal.pone.0249859
Vaseva, I. I., Simova-Stoilova, L., Kostadinova, A., Yuperlieva-Mateeva, B., Karakicheva, T., Vassileva, V. (2022). Heat-Stress-Mitigating effects of a protein-Hydrolysate-Based biostimulant are linked to changes in protease, DHN, and HSP gene expression in maize. Agronomy 12, 1127. doi: 10.3390/agronomy12051127
Vinoth, S., Sundari, Gurusaravanan, P., Sivakumar, S., Siva, G., Kumar, G. P., et al. (2017). Evaluation of seagrass liquid extract on salt stress alleviation in tomato plants. Asian J. Plant Sci. 16, 172–183. doi: 10.3923/ajps.2017.172.183
Visconti, F., de Paz, J. M., Bonet, L., Jordà, M., Quinones, A., Intrigliolo, D. S. (2015). Effects of a commercial calcium protein hydrolysate on the salt tolerance of diospyros kaki l. cv. “Rojo brillante” grafted on Diospyros lotus l. Sci. Hortic. 185, 129–138. doi: 10.1016/j.scienta.2015.01.028
Waqas, M. A., Kaya, C., Riaz, A., Farooq, M., Nawaz, I., Wilkes, A., et al. (2019). Potential mechanisms of abiotic stress tolerance in crop plants induced by thiourea. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01336
Xu, C., Leskovar, D. I. (2015). Effects of a. nodosum seaweed extracts on spinach growth, physiology and nutrition value under drought stress. Sci. Hortic. 12 (183), 39–47. doi: 10.1016/j.scienta.2014.12.004
Yakhin, O. I., Lubyanov, A. A., Yakhin, I. A., Brown, P. H. (2017). Biostimulants in plant science: A global perspective. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.02049
Zaid, A., Mohammad, F., Fariduddin, Q. (2020). Plant growth regulators improve growth, photosynthesis, mineral nutrient and antioxidant system under cadmium stress in menthol mint (Mentha arvensis l.). Physiol. Mol. Biol. Plants 26, 25–39. doi: 10.1007/s12298-019-00715-y
Zandalinas, S. I., Fichmana, Y., Devireddy, A. R., Senguptac, S., Azad, R. K., Mittler, R. (2020). Systemic signaling during abiotic stress combination in plants. Biol. Sci. 117 (24), 13810–13820. doi: 10.1073/pnas.2005077117
Zandalinas, S. I., Mittlerb, R., Balfagóna, D., Arbonaa, V., Gómez-Cadenasa, A. (2018). Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 162 (1), 2–12. doi: 10.1111/ppl.12540
Zhao, J., Lu, Z., Wang, L., Jin, B. (2021). Plant responses to heat stress: Physiology, transcription, noncoding RNAs, and epigenetics. Int. J. Mol. Sci. 22, 117. doi: 10.3390/ijms22010117
Keywords: phytostimulants, plant beneficial microbes, abiotic stresses, climate change, modern agriculture
Citation: Ma Y, Freitas H and Dias MC (2022) Strategies and prospects for biostimulants to alleviate abiotic stress in plants. Front. Plant Sci. 13:1024243. doi: 10.3389/fpls.2022.1024243
Received: 21 August 2022; Accepted: 28 November 2022;
Published: 22 December 2022.
Edited by:
Vicent Arbona, University of Jaume I, SpainReviewed by:
Rosalba Mireya Hernández-Herrera, University of Guadalajara, MexicoArchana Singh, University of Delhi, India
Lin Xu, Ghent University, Belgium
Copyright © 2022 Ma, Freitas and Dias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helena Freitas, aGZyZWl0YXNAdWMucHQ=; Maria Celeste Dias, Y2VsZXN0ZS5kaWFzQHVjLnB0