 Hendrik Poorter1,2*†
Hendrik Poorter1,2*† Grégoire M. Hummel3†
Grégoire M. Hummel3† Kerstin A. Nagel1†
Kerstin A. Nagel1† Fabio Fiorani1†
Fabio Fiorani1† Philipp von Gillhaussen4†Olivia Virnich1
Philipp von Gillhaussen4†Olivia Virnich1 Ulrich Schurr1†
Ulrich Schurr1† Johannes A. Postma1†
Johannes A. Postma1† Rick van de Zedde5†
Rick van de Zedde5† Anika Wiese-Klinkenberg1,6†
Anika Wiese-Klinkenberg1,6†- 1Plant Sciences (IBG-2), Forschungszentrum Jülich GmbH, Jülich, Germany
- 2Department of Natural Sciences, Macquarie University, North Ryde, NSW, Australia
- 3Phenospex, Heerlen, Netherlands
- 4International Plant Phenotyping Network e.V. (IPPN), Jülich, Germany
- 5Plant Sciences Group, Wageningen University & Research, Wageningen, Netherlands
- 6Bioinformatics (IBG-4), Forschungszentrum Jülich GmbH, Jülich, Germany
Automated high-throughput plant phenotyping (HTPP) enables non-invasive, fast and standardized evaluations of a large number of plants for size, development, and certain physiological variables. Many research groups recognize the potential of HTPP and have made significant investments in HTPP infrastructure, or are considering doing so. To make optimal use of limited resources, it is important to plan and use these facilities prudently and to interpret the results carefully. Here we present a number of points that users should consider before purchasing, building or utilizing such equipment. They relate to (1) the financial and time investment for acquisition, operation, and maintenance, (2) the constraints associated with such machines in terms of flexibility and growth conditions, (3) the pros and cons of frequent non-destructive measurements, (4) the level of information provided by proxy traits, and (5) the utilization of calibration curves. Using data from an Arabidopsis experiment, we demonstrate how diurnal changes in leaf angle can impact plant size estimates from top-view cameras, causing deviations of more than 20% over the day. Growth analysis data from another rosette species showed that there was a curvilinear relationship between total and projected leaf area. Neglecting this curvilinearity resulted in linear calibration curves that, although having a high r2 (> 0.92), also exhibited large relative errors. Another important consideration we discussed is the frequency at which calibration curves need to be generated and whether different treatments, seasons, or genotypes require distinct calibration curves. In conclusion, HTPP systems have become a valuable addition to the toolbox of plant biologists, provided that these systems are tailored to the research questions of interest, and users are aware of both the possible pitfalls and potential involved.
1 Introduction
For decades, plant growth has been studied by taking non-destructive measurements such as plant height, along with destructive measurements such as plant dry mass. Typically, these assessments involved serial measurements over time, or comparisons of plants or plots at a ‘final’ harvest (Evans, 1972). Usually, 3-8 plants per treatment were harvested, or 2-30 genotypes compared, in a process that could easily take a full day or more for a single person. However, In the past 15 years, there has been a significant shift towards high-throughput plant phenotyping (HTPP). Fully automated systems now screen up to hundreds of genotypes and thousands of individual plants or field plots using non-destructive sensors, with the collected data automatically processed and stored for later use. (Furbank and Tester, 2011; Fiorani and Schurr, 2013; Tardieu et al., 2017; Lorence and Jimenez, 2022). In controlled conditions, automated phenotyping is often achieved by bringing individual plants to sensors (Yang et al., 2014; Al-Tamimi et al., 2016), or by moving sensors to or over the plants (Granier et al., 2006; Nagel et al., 2012). In the field, sensors are also brought to plants, either through mobile vehicles (White and Conley, 2013; Deery et al., 2014) or via drones or other aerial platforms that fly over field trials (Vargas et al., 2020; Roth et al., 2022). In all of these cases, automation has enabled a significant increase in the number of individual plants or plots that can be processed daily, often by an order of magnitude.
An important driver for the development of HTPP systems has been the rapid progress in the field of molecular biology. The extensive expansion and utilization of molecular tools at continuously decreasing costs have enabled thorough genotypic characterization of many plant species, cultivars, and genotypes. Phenotypic characterization, however, still lags behind, first because it is a time-consuming and often still manual measurement process, and second because plants of the same genotype can exhibit a range of different phenotypes, depending on environmental conditions (Furbank and Tester, 2011; Zavafer et al., 2023). This phenotyping bottleneck is particularly pronounced when studying traits that are controlled by multiple genes. In such cases, top-down approaches like Quantitative Trait Loci (QTL) analysis or Genome-Wide Association Studies (GWAS), are necessary to identify regions of the genome that are determining these traits (Gibson, 2018). However, conducting these analyses requires phenotyping hundreds of different genotypes, preferably all grown simultaneously in a common environment. Such analyses greatly benefit from the automation and standardization provided by HTPP systems (reviewed in Xiao et al., 2022).
Various technological advancements have facilitated the development of automated HTPP systems. A wide range of non-destructive sensors, including digital RGB cameras, hyperspectral, thermal, and fluorescence cameras, laser scanners, and LIDARs (Liu et al, 2020) enable repeated measurements of individual plants over time. This provides a higher resolution for capturing time-related phenotypic changes compared to experimental designs where new plants need to be harvested destructively for each time point (Poorter and Garnier, 1996; Walter et al., 2007). The use of automated gantries and transportation systems as well as drones allow us to minimize the distance between sensors and plants. Another significant factor is the increased computational power and improved algorithms that enable efficient image processing, with or without machine learning (Walter et al., 2015; Dobrescu et al., 2020).
As mentioned above, HTPP systems encompass a diverse range of approaches, all centered around non-destructive measurements. In the years ahead, more and more of these systems will be built to effectively screen a large number of plants for their size, growth trajectory, and other traits. However, like any complex equipment, the use of these platforms also presents challenges and limitations that may be overlooked by those who have not yet utilized them. Therefore, before investing in the purchase, construction, and utilization of these platforms, it is crucial to consider potential issues that may arise, and how they can be addressed. In the Results and Discussion section, we share insights gained from our experience in developing and deploying HTPP systems over the past 15 years. While our focus is primarily on systems operating in (semi-)controlled environments, several of the issues discussed will apply to field phenotyping as well.
2 Materials and methods
Part of the discussion that follows will address the relationship between non-destructive measurements, such as projected leaf area, and variables that require destructive analysis, including total leaf area, shoot biomass, and total plant biomass. We illustrate this part of the discussion with data from two experiments. In the first experiment, Arabidopsis thaliana (Col-0) plants were cultivated in a growth room using a rack equipped with neon tubes (Fluora, Osram, Munich). The plants were grown in soil (ED73, Einheitserde, Uetersen, Germany) in a cultivation tray with adjacent 80 ml cells. After incubating the seeds in the dark at 4°C, they germinated in the tray. The plants were then subjected to a 12-hour day length, a photosynthetic photon flux density (PPFD) of 40-50 μmol m-2 s-1, a day/night temperature of 23/20°C and a relative humidity (RH) of ca. 55%. Plants were watered from below when the top-soil was fully dry. Thirty-nine days after sowing, the plants were imaged six times during the day, with 2-hour intervals. Screening and image analysis were performed following Walter et al. (2007). The data from this experiment can be found in Supplementary Data Sheet A1.
The second experiment involved studying Plantago major, another rosette-forming species, throughout a significant part of its growth cycle and included a large number of replicates per harvest (n=12). The plants were grown hydroponically under two different CO2 concentrations (350 and 700 µmol mol-1), in growth chambers with a 12-hour day length, a PPFD of 230-270 μmol m-2 s-1, a day/night temperature of 20/18°C and a day/night RH of 60/90%. Plants from both treatments were monitored weekly over a 7-week growth period. Projected leaf area (PLA) was determined by capturing slides with an analogue camera placed at a height of 2 meters. The slides were projected, plant outlines traced on papers, and those were subsequently digitized semi-manually using a digitizer (model 9874a, Hewlett Packard, Stanford, CA, USA). Total leaf area per plant (TLA) was determined using a leaf area meter (LiCor 3100, Li-Cor Inc, Lincoln, NE, USA) equipped with a conveyor belt system. The determination of the total dry mass of shoots and roots was done manually, with plant size varying >300-fold throughout the course of the experiment. The data for this experiment is sourced from a previously published study by Poorter et al. (1988) and can be found in Supplementary Data Sheet 2.
Data were analyzed using R version 4.1 (R Core Team, 2022). Various calibration curves were established by employing the function lm, with or without a ln-transformation of the relevant variables, and with or without a quadratic term for the explanatory variable.
3 Results and discussion
Before proceeding with the design and implementation of a high-throughput plant phenotyping (HTPP) system, it is essential to consider the following questions. Figure 1 shows a schematic overview of the relevant aspects to be considered.
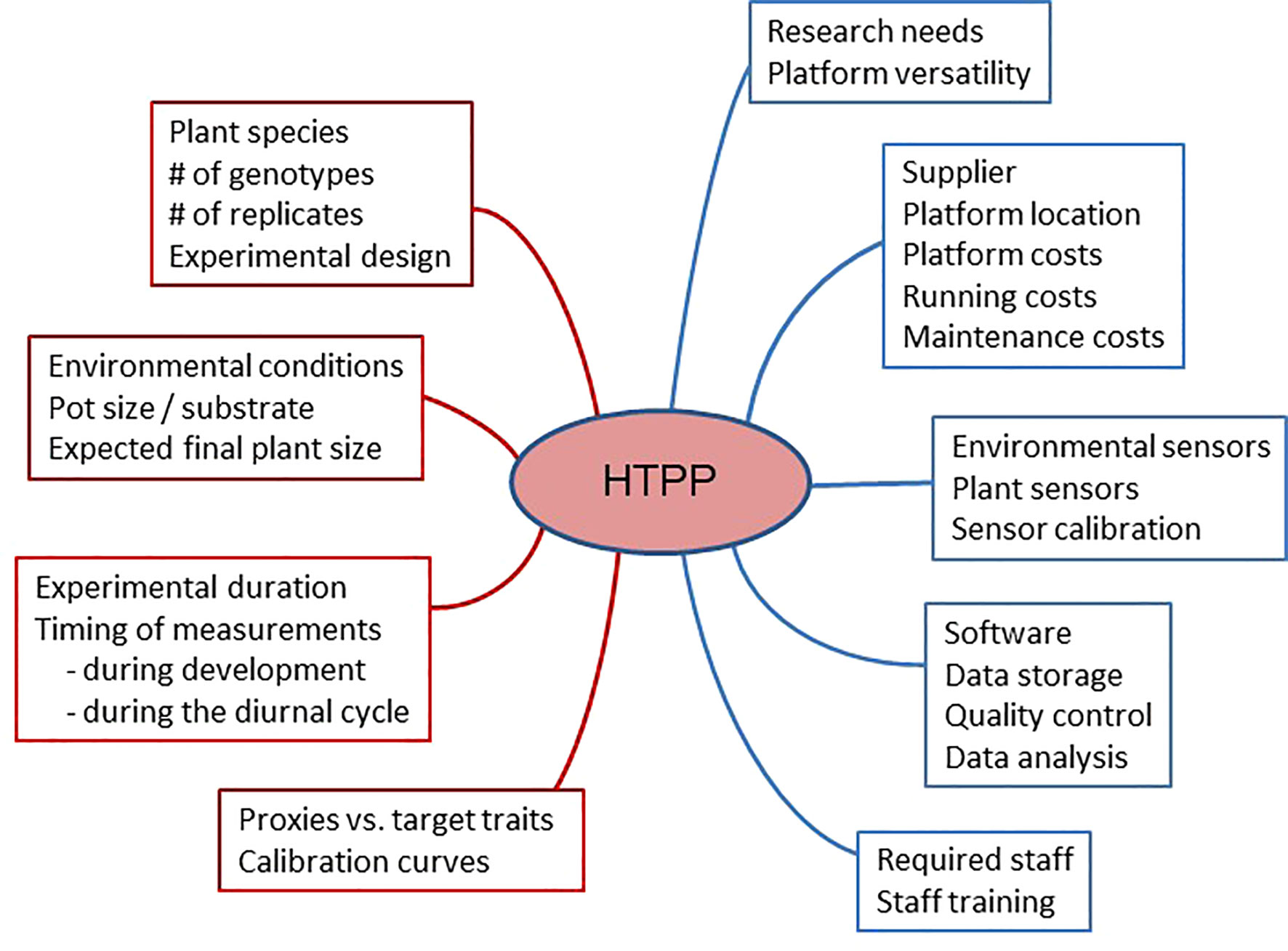
Figure 1 A schematic diagram indicating different aspects of high-throughput plant phenotyping (HTPP) systems that should be considered before purchasing such a system (in blue) and using it (in brown).
3.1 What is the specific research need?
HTPP systems are designed based on principles of automation and standardization. However, several critical aspects must be considered and discussed in order to effectively design and implement these systems. These include the required scale of the experiments, the selection of sensors that will be installed, the necessary software infrastructure, and the expected return on investment in terms of financial and human resources. These aspects can only be fruitfully discussed if guided by the question what purpose the system should serve and what goal(s) one would like to achieve through this platform. This is even more important in the case where multiple research groups with diverse interests are involved, as there is a risk that the intended platform becomes a compromise that in the end does not satisfy any.
3.2 What are the costs of investment and maintenance for a phenotyping platform?
HTPP systems involve a complex combination of logistics and technology, particularly when plants need to be moved to sensors. Such machines require the use of conveyor belts, gantries, or mobile robots to transport plants through growth chambers or glasshouses to an imaging station. At the imaging station, plants are generally subjected to controlled conditions, such as a consistent light spectrum and intensity, allowing for a consistent acquisition of images or other sensor-based data. Additionally, plants in pots or carriers can be automatically weighed and watered. Measurement equipment must be linked to large-scale data storage facilities and databases, in order to store and retrieve the collected measurement information and other metadata. Clearly, expert knowledge of automation, logistics, error control, fine mechanics, non-destructive sensing, image analysis, and database management is required to build and operate these systems properly. Companies specializing in these different fields offer to construct customized machines on-site. The costs of these systems can be substantial. Prices typically range from approximately €60,000 to €120,000 for small systems with a lower degree of automation and throughput, €350,000 – €500,000 for fully automated sensor-to-plant or plant-to-sensor systems, and up to €3,000,000 for high-end systems with extensive optimization and automation of the workflow. The costs vary depending on factors such as the types and number of sensors used, system size, quality of the product and service provided by the supplier. In addition, these custom-built facilities require expert maintenance, including sensor calibration, replacement of worn-out parts and preventive check-ups. Yearly servicing contracts often amount to 5-10% of the acquisition price and are necessary for the duration of system usage.
Given the above considerations, certain laboratories opt to construct HTPP systems themselves, either utilizing off-the-shelf products or with the support of robotics companies (Bagley et al., 2020). Building customized platforms can allow for increased flexibility (as discussed in section 3.4) and better fit the specific requirements of the involved labs. However, it is important to acknowledge that the path to a fully functional machine is challenging and time-consuming. While the direct hardware costs may be lower than those of commercial HTPP systems, the substantial human resources required for the development and maintenance of such automated systems should not be underestimated (Reynolds et al., 2019).
There are two additional points to consider regarding personnel investment. Firstly, it is advisable to involve expert users, project managers, and engineers throughout the planning and building phase. This ensures that any necessary compromises that almost inevitably have to be made during the whole process do not strongly constrain the desired goals of the end-users. Secondly, operating these systems can be complex, particularly when troubleshooting errors, as there are numerous variables to consider within the platform software, platform mechanics and network communications. Therefore, it is recommended to have an operational team led by an expert who is responsible for the platform. This lead expert can receive training from the supplier or the constructors, and subsequently train all other individuals who will be utilizing this platform. Clearly, such a person requires both technological as well as plant biological expertise to effectively manage and operate the HTPP system.
3.3 What other logistical and infrastructural adjustments are necessary?
An HTPP platform allows researchers to significantly scale up their experiments, often by an order of magnitude. This implies that also an order of magnitude more plants are processed, more containers have to be filled, more consumables are used, more electricity and irrigation are required, and additional cleaning has to be done at the end of the experiment. While these aspects are not part of the phenotyping platform per se, they contribute to a significant workload. As a result, there is often a need for subsequent adjustments to the workflow’s ergonomics, such as automating tasks like pot filling, seed sowing (Jahnke et al., 2016) or plant transplantation. Together with the sampling for further physiological characterization and disinfection or disposal of used components, they bring about additional planning and investments on top of those for the platform itself.
Another aspect to consider is the data infrastructure. Given the massive amount of data generated by HTPP systems, effective storage and accessibility of data for both short-term and long-term use are essential. Such utilization of data requires a good documentation of data and metadata, as described in the ‘Minimal Information About Plant Phenotyping Experiments’ (MIAPPE) guidelines (Krajewski et al., 2015; Papoutsoglou et al., 2020). Also, data and metadata should be stored in accordance with the FAIR principle (findable, accessible, interoperable, and reusable; Wilkinson et al., 2016) as in GnpIS (Pommier et al., 2019) and as currently established in research data management infrastructures like DataPLANT (www.nfdi4plants.de).
Remote support by the different suppliers requires a safe, secure, and reliable access policy. Moreover, in the analysis of data from such platforms, there is a growing emphasis on integrating genotypic and phenotypic data, as well as incorporating detailed environmental characterization of the experiments. Bringing all these data together often requires a considerable a-posteriori effort from trained personnel, which also needs to be budgeted in advance to make full use of the potential of these HTPP systems (Reynolds et al., 2019).
3.4 How flexible can or should the system be?
Automation can replace a considerable amount of tedious repetitive manual labor, especially when routine operations are customized to the research question and species of interest. However, this increased automation often comes at the cost of reduced flexibility. For instance, a system that is perfectly fit for small rosette plants like Arabidopsis thaliana, may not be as suitable for a large species with different architecture, such as Zea mays. Increased flexibility of the system can be achieved by planning for modular components that can be interchanged depending on the prevailing research question. However, increased flexibility to accommodate the needs of a variety of researchers also implies that the system becomes increasingly complicated, with a higher likelihood of failures and more efforts to solve these problems. A ‘jack-of-all-trades’ system will hardly provide the same level of detail and raw data resolution across the entire range of plant sizes and architectures as a dedicated high-resolution system designed specifically for either small or big plants, due to limitations imposed by the optical properties of the sensor and system layout. In those cases, a potential solution could be to have two smaller and targeted systems, rather than relying on a one-size-fits-all solution.
Another aspect of flexibility is the expected lifespan of system components. Technological advancements occur rapidly, and new sensors, for example, will generally be more powerful and informative than their predecessors. However, owners of commercially-acquired phenotyping systems often face problems in that the software to run the whole system is proprietary to the company, and therefore not accessible for further development. In such cases, it is complicated or impossible to integrate new sensors or apply other modifications to existing HTPP systems. New buyers are advised to discuss with their suppliers what support they can get in that respect. Self-builders are suggested to set up their software interfaces as flexible as possible, to easier adjust their system when new sensors or updates become available.
3.5 What constraints does the system impose on growth conditions?
Plants are strongly influenced by their environment and different genotypes or species may show varying degrees of genotype x environment interactions for many of their phenotypic traits. This becomes particularly critical, because in most (semi-)controlled environments we impose abiotic conditions that significantly deviate from the natural conditions plants experience outdoors (Poorter et al., 2016; Chiang et al., 2020). Considerations about the location of the HTPP system (growth chamber, glasshouse, or field) and the range of environmental conditions provided to the plants are therefore an integral part of the design process.
Compared to traditional experiments, the use of high-throughput phenotyping systems often introduces additional constraints, that can impact the growth environment of the plants and, consequently, the outcomes of experiments. For instance, in a typical plant-to-sensor system, the combined weight of plants plus pots is limited by the strength of the conveyor belt used, as well as the scales used for gravimetric measurements. This limitation results in experiments being confined to plants in relatively small pots with substrates of low specific mass, which clearly affects plant growth and experimental outcomes (Passioura, 2002; Poorter et al., 2012). Transportation may also have other consequences. Tall plants, such as Zea mays, may topple over if the conveyor belts move too quickly. During transport, leaves of sensitive species (e.g. Brassica rapa, Hordeum vulgare) can get damaged, causing them to droop downwards along the pot. To mitigate damage, researchers may choose to place suitable support structures next to or around each plant, except in cases where wilting would be a phenotypic trait of interest. Apart from direct damage, one would also expect thigmo-morphogenetic responses to occur as a reaction to the mechanical perturbations during transport of plants, such as thicker and shorter stems (Anten et al., 2005). However, plant height was not negatively affected in the experiment of Brien et al. (2013), and neither was shoot biomass or leaf area in various experiments (Table 1).
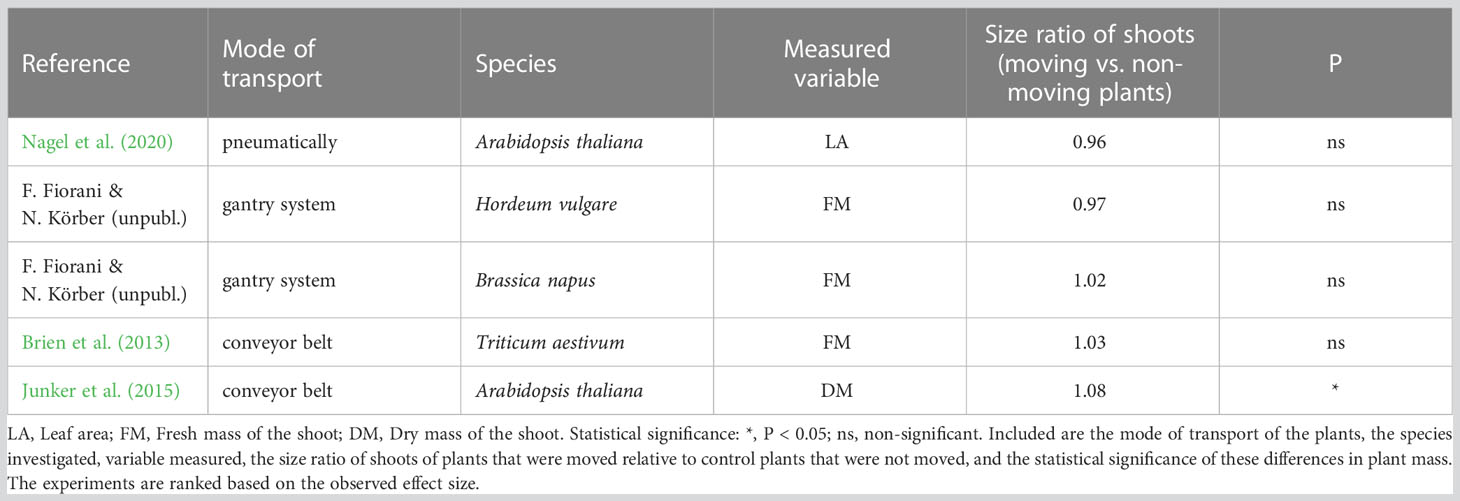
Table 1 Effect of plant-to-sensor transport on shoot biomass, as based on various experiments carried out in glasshouses.
Fixed distances between cameras/sensors and plants may restrict the range of plant sizes that can be investigated, thereby limiting the developmental stages that can be studied in such HTPP systems to young vegetative plants. A last example pertains to the watering of the plants. Well-watered containers, especially with peat substrate, often show algal growth on the soil surface, which hampers the non-destructive derivation of plant size through image analysis (Figure 2A). To avoid this complication, researchers may opt to cover the top of the pots with a plastic sheet of contrasting color (Junker et al., 2015). Alternatively, for small rosette species like Arabidopsis, reducing the volume and/or frequency of watering can keep the topsoil dry for a longer duration. Although this suppresses algal growth, it also has unknown consequences for the growth rate and phenotype of the plants. It is recommended to consider a-priori whether the additional constraints imposed by an HTPP system are acceptable within the scope of the research question.
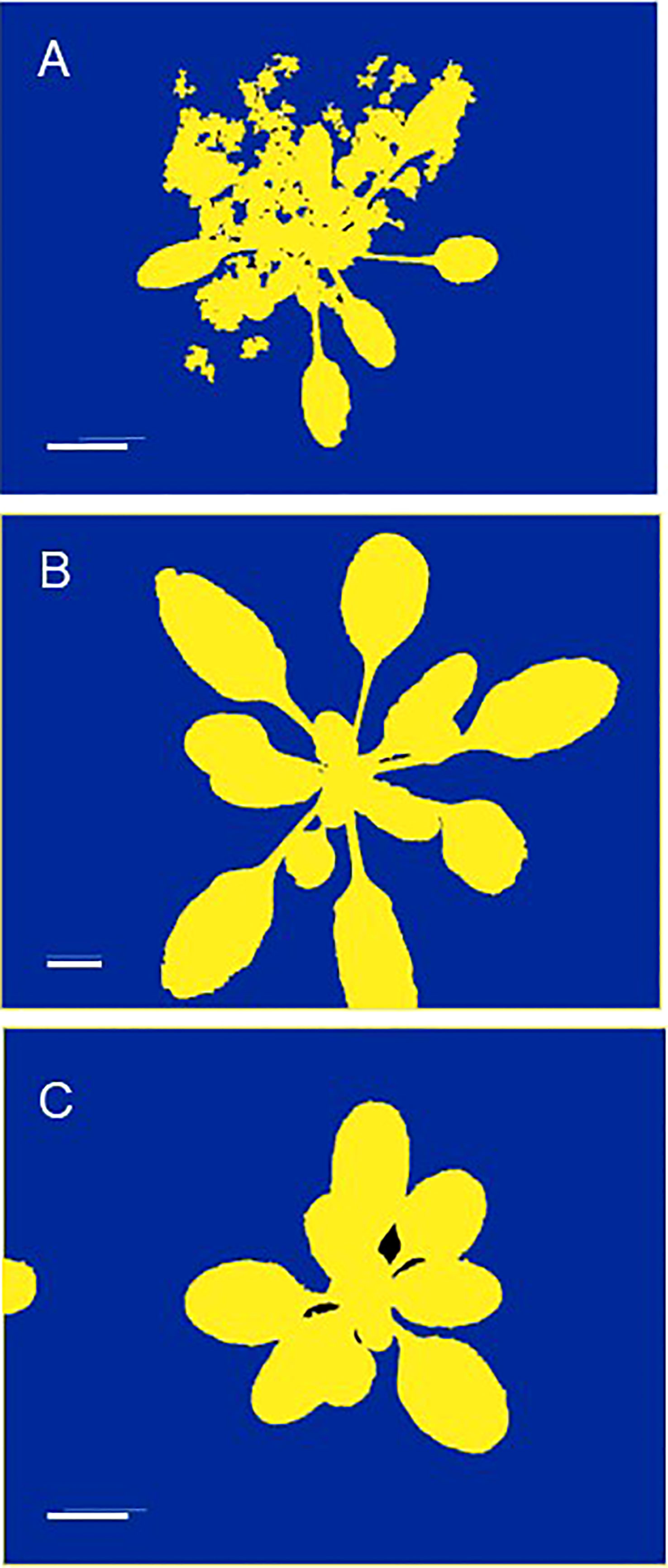
Figure 2 Examples where unsupervised automated high-throughput phenotyping may lead to incorrect results. (A) Algal growth resulting in a mask that is too broad and yields an overestimation of projected leaf area (PLA). (B) A plant grown out of the image acquisition area, resulting in an underestimation of PLA. (C) A neighboring plant growing into the image acquisition area, resulting in an overestimation of PLA. All pictures depict Arabidopsis thaliana plants and are masked images, used for measuring PLA by counting the number of green pixels. These images are for illustrative purposes only. The scale indicates a length of 1 cm.
3.6 When and how often should the plants be measured?
Once an operational HTPP system is in place, attention can shift to the performance of experiments. Given their typical large scale, careful consideration of experimental design is essential. For a comprehensive discussion on this topic, the reader is referred to Thompson et al. (2022). One notable advantage of high-throughput phenotyping systems is that individual plants (or microplots) can be measured frequently and non-destructively. This allows for the repeated measurements of the same plants, enabling the tracking of their growth and development over time. Such analyses can yield valuable insights, with a good example discussed in Box 1. However, in cases where the research question focuses on identifying the best-performing genotype at the end of the experiment, repeated measurements may be unnecessary. In those cases, researchers could also opt for one final destructive harvest, which might be more simple, cheaper, and more informative, as also illustrated in Box 1. A lower measurement frequency may also be advantageous if the measurements have the potential to interfere with plant growth. This is particularly relevant when plants are taken out of their growth environment for longer-duration measurements, such as magnetic resonance imaging (MRI) or computer tomography (CT) scans. A higher measurement frequency then provides better insight into plant development, but may also have a stronger negative effect on plant growth.
Box 1. Example of the trade-off between measuring with higher frequency or higher precision.
Here we provide two examples of experiments focusing on root distribution. The first one followed root development non-destructively over time (Nabel et al., 2018). Seedlings of a woody shrub (Sida hermaphrodita) were grown for 90 days in rhizotrons of 36 x 75 x 2.6 cm in size, where one side consists of transparent acrylic glass. The researchers placed digestate, which is a residue remaining after anaerobic digestion of biomass to methane, at a specific location in the rhizobox. This location is indicated by the brown circles in Figures 3A–C. Root growth of the part of the root system close to the transparent side of the rhizobox was monitored by regularly capturing images (Nagel et al., 2012). These images showed that plants strongly avoided the digestate patch in the first 60 days of the experiment (Figures 3A, B). However, roots strongly proliferated into this patch of nutrients later in time (Figure 3C). In this case, the timing proved to be an essential aspect of how these plants reacted to the treatment. Although these analyses of root distribution are still challenging for computers and often need human supervision, the effort in this case proved worthwhile for understanding the timing of root responses.
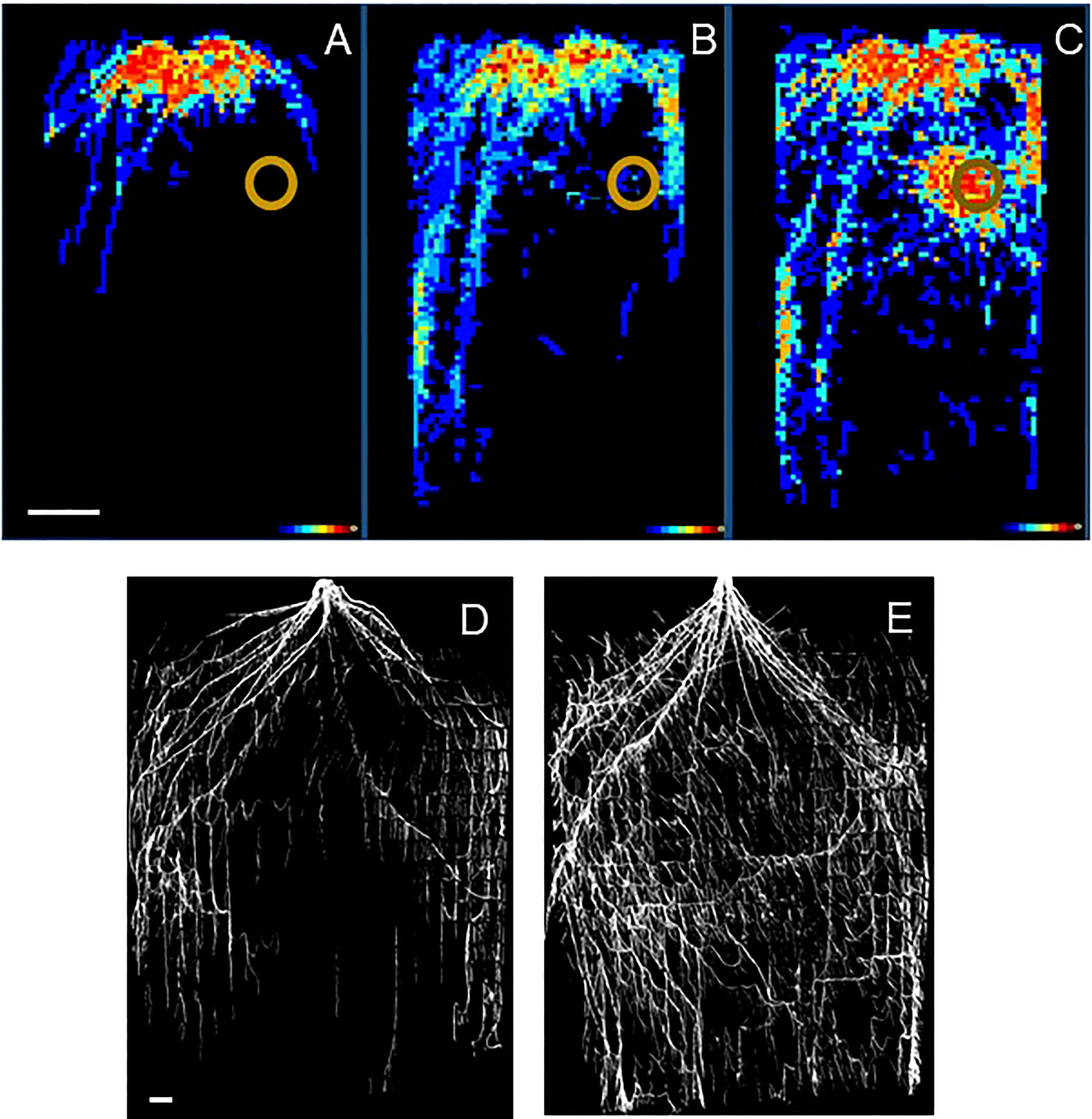
Figure 3 (A–C) Root distribution of Sida hermaphrodita plants in rhizotron boxes with a localized depot of digestate, indicated by the brown circle. (D) Root distribution in a rhizobox at the end of an experiment after the soil has been removed. Figures (A–C) are adapted from Nabel et al. (2018) and show in false colors ranging from dark blue to bright red the number of rhizotrons (out of 10 in total) where roots were observed for each x-y location in the rhizotron. (A) 30 days, (B) 60 days and (C) 90 days after the start of the experiment. Figures (D, E) show the root systems of two Sorghum bicolor genotypes at the 12th-leaf stage (ca. 6-8 weeks after germination) in an experiment similar to that described in Singh et al. (2010). The white bars indicate a length of 10 cm. Picture credits Figures (A–C): Moritz Nabel, Forschungszentrum Jülich, Germany. (D, E): Vijaya Singh, University of Queensland, Australia.
An alternative approach was followed by Singh et al. (2010; Figure 3D). They grew two Sorghum genotypes in rhizotrons measuring 120 x 240 x 10 cm. When it comes to selecting, for example, the best-performing genotypes of a panel of genotypes, an evaluation at the end of the experiment could be as informative as a complete analysis over time. If this is the case, researchers may also opt for alternative and cheaper set-ups, such as rhizoboxes that are not integrated into automated HTPP systems. Those rhizoboxes could have larger dimensions and allow a wider range of root substrates. By pushing a pinboard into one of the sides before washing away the soil with water, the distribution of a whole root system can be characterized, rather than only those roots close to the acrylic glass (Singh et al., 2010; Figures 3D, E).
The dilemma faced by researchers in these cases is whether it is more informative to have estimated data over the course of the experiment for only a portion of the root system, as provided by automated rhizotron systems, or to have more precise data capturing the entire root system but only at the end of the experiment.
A second point to consider is the time of day when plants will be measured. When using a drone to fly over an experimental field, it can capture images of many microplots simultaneously. Acquiring images or other data for these plots using sensors mounted on vehicles will take longer. Depending on the type of measurements taken, it may require minutes or more per microplot. With plant-to-sensor systems within a glasshouse, where it takes minutes to move a plant through the imaging station and measure its characteristics, phenotyping all plants in one experiment could take a full day. During that period, large changes in environmental variables, such as light intensity and temperature, may occur, likely resulting in large variation in physiological variables such as stomatal conductance and photosynthesis as well. One morphological trait that can exhibit considerable diurnal variation is leaf angle (Rosa and Forseth, 1996), which has obvious implications for the projected leaf area (PLA; Dobrescu et al., 2017) as used in many HTPP systems. For example, during the diurnal part of the diel cycle, Arabidopsis plants may increase their leaf area and biomass by 20% (Wiese et al., 2007). However, in a similar experiment, PLA values were found to decrease by 18-35% during the light period (Figures 4A, B, 5), due to upward movements of leaves and petioles. Consequently, when conducting consecutive 2D measurements of plants day after day, it is important to measure them at the same time of day, to avoid bias caused by the diurnal rhythm of leaf movements. Moreover, different genotypes or treatments should be blocked into the same time window, to ensure that no confounding effects occur. One fast alternative approach is to perform a 3D laser scan (Dornbusch et al., 2012). Other options include using multiple imaging stations, or employing a gantry system where sensors are brought to the plants, enabling parallel measurements of many plants in short time.
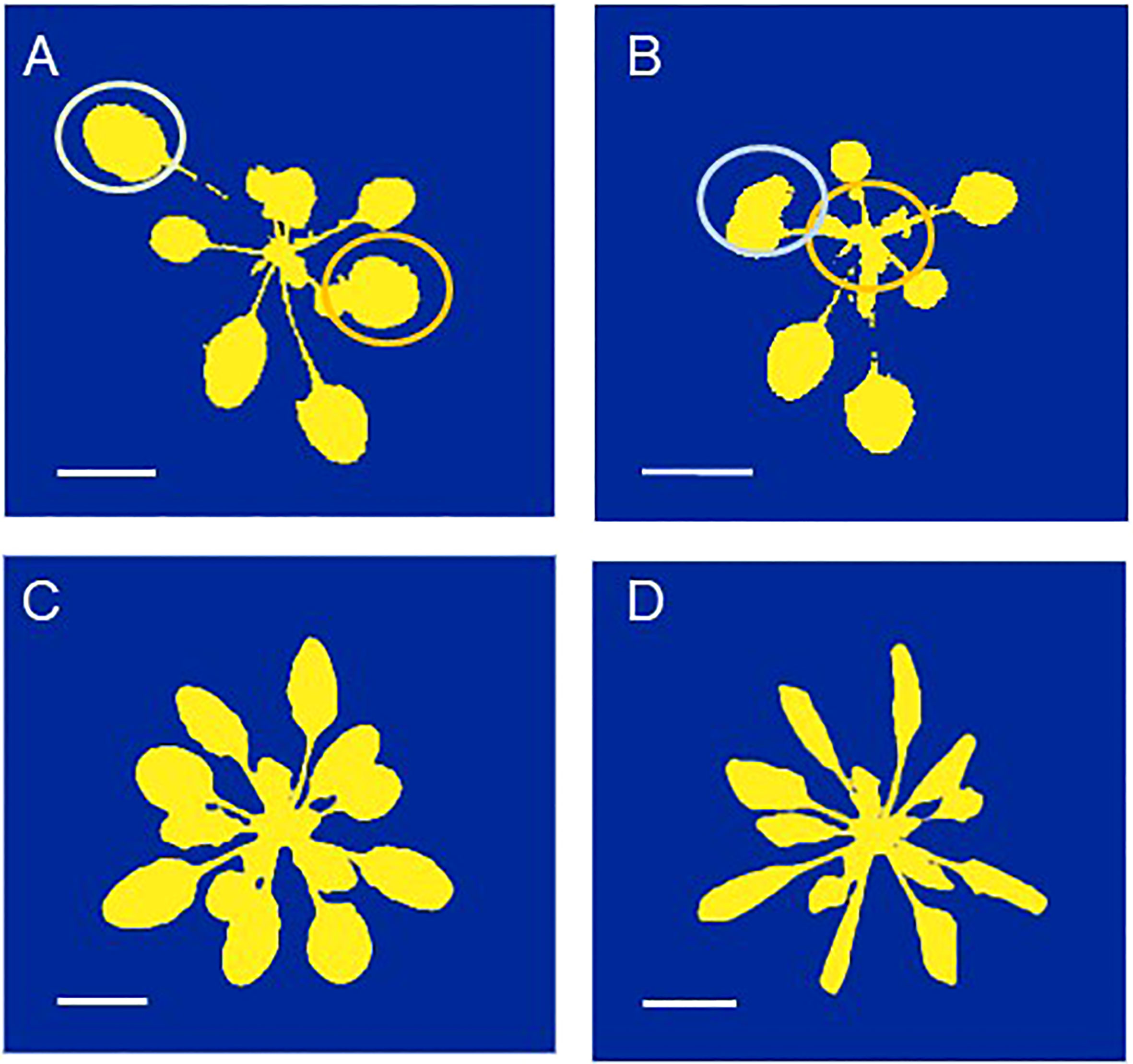
Figure 4 (A, B) The same Arabidopsis thaliana plant photographed (A) in the morning, with a low leaf angle and (B) at the end of the day with a much higher leaf angle. The yellow and orange circles indicate the positions of the youngest full-grown leaves which exhibit the largest change in leaf angle. (C, D). The same plant photographed (C) before and (D) after the onset of water stress, which resulted in leaf wilting. These images are for illustrational purposes only. The scale indicates a length of 1 cm.
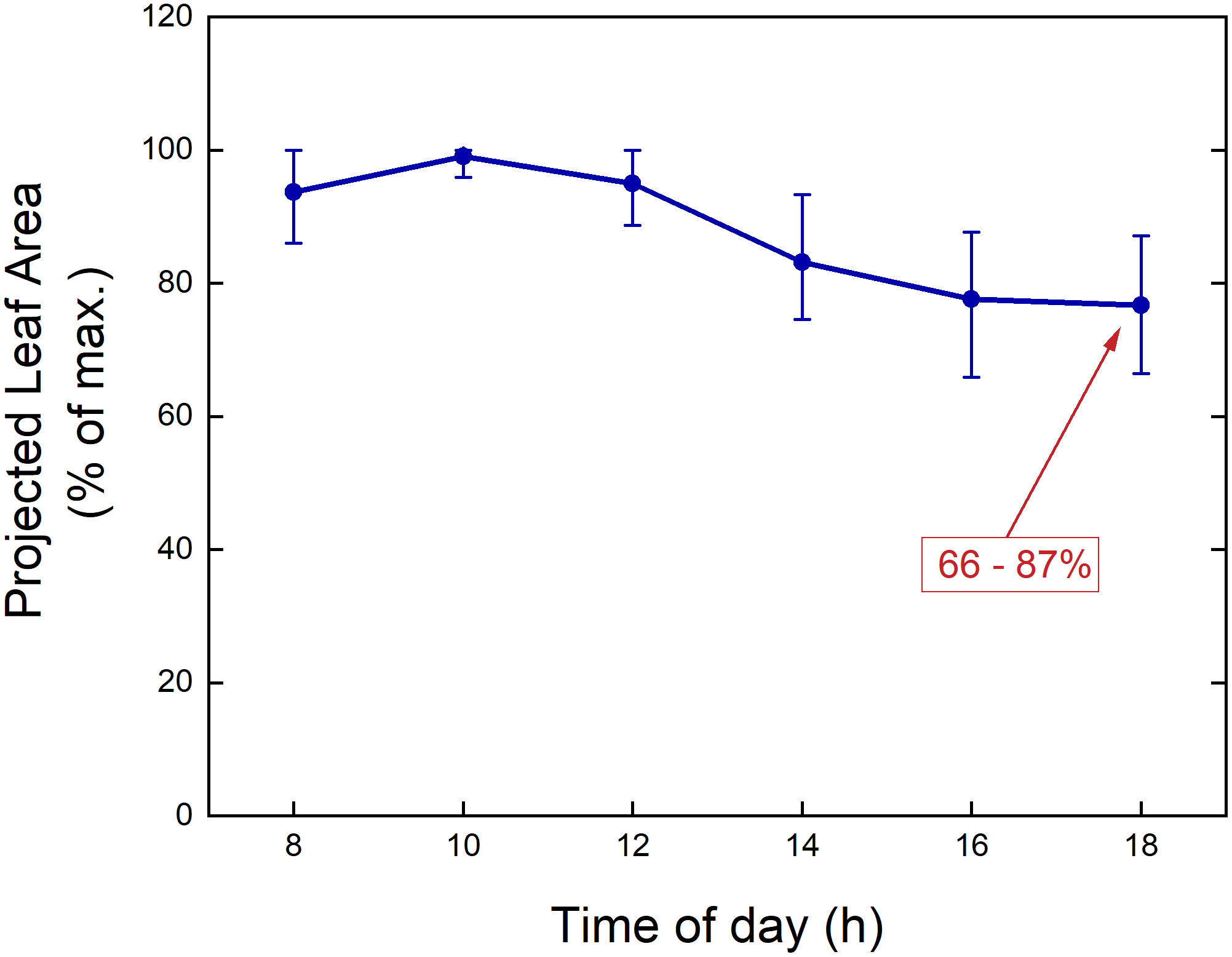
Figure 5 Measured Projected Leaf Area (PLA) over a day for Arabidopsis thaliana plants grown in a growth chamber. The values for each plant are normalized to the maximum value measured during the diurnal part of the day-night cycle. The dots indicate mean values, and the ‘error bars’ represent the 5% and 95% percentiles (n = 44).
In some cases, the research question requires a high frequency of measurements on the same plants. Examples are physiological responses of plants following the application of a compound, exposure to a pathogen, or exposure to abiotic stress (Jansen et al., 2009; Mahlein et al., 2019). In some species, capturing images at higher frequencies and analyzing them in almost real-time can be used to detect the onset of potentially undesired drought stress during the experiment (Figures 4C, D; Eberius and Lima-Guerra, 2009; Janni et al., 2019).
3.7 How adequate is the quality control and data handling?
The amount of information collected from a single experiment can be substantial, especially when imaging of any kind is involved. The question is how well we, as experimenters, can handle this vast quantity of data (Eberius and Lima-Guerra, 2009; Tardieu et al., 2017). Based on our own experience, we have observed several issues that can arise during an experiment. Sensors, especially environmental sensors that are deployed throughout the year, may have not been calibrated for a long time or show failures of various kinds during the experimental period. With larger amounts of plants, problems might more easily go undetected. For example, some plants could topple over or do not receive adequate watering. In image analysis systems, leaves from neighboring plants might appear in pictures taken, photos might not cover the full plant, or masking might not function properly (Figures 2B, C). These errors are easily noticed if they occur frequently, but in the midst of hundreds of plants, thousands of pictures, and tens of thousands of other collected data points, such errors can easily go unnoticed. Thorough data inspection and double-checking for mistakes are therefore crucial, but can be cumbersome without the assistance of digital tools. Dedicated software programs for data visualization and targeted image retrieval, such as Azure, iRods, PHIS, Fairdom, Zegami, or similar solutions, enable fast selection of images of specific plants over time, aiding in the identification of potential outlier data. Automated quality control procedures should routinely flag instances where parts of leaves are outside the picture, or leaves of neighboring plants are distorting the results. Graphical analysis of data distribution, time courses, or dose-response curves can provide insights into potential issues with specific plants or entire groups of plants (Xu et al., 2015). This is particularly useful in ongoing experiments, when possible problems can be detected and solved by data analysis at an early stage. Real-time reporting, along with easy and user-friendly (remote) access to visualizations and resulting analyses, facilitates early detection and problem-solving.
3.8 How informative are the selected proxies for the actual variables of interest?
For decades, scientists have relied on spectrophotometric measurements to asses enzyme activity, wet digestion or pyrolysis for leaf nitrogen determination, infra-red gas analyzers to determine photosynthesis, and manual harvesting to measure plant biomass. However, these conventional measurements all require significant manual effort, and are therefore not suitable for high-throughput phenotyping. Efforts have been made to automate such measurements (e.g. Gibon et al., 2004; Gomez et al., 2010), but challenges remain in automating processes such as grinding and weighing, particularly under low-temperature conditions to prevent chemical degradation (Hall et al., 2022). In search of alternatives, scientists have explored measurements that are easier to perform, yet still provide valuable information. For example, leaf nitrogen content can be estimated non-destructively using multispectral analysis (Ye et al., 2020), photosynthesis can be assessed through fluorescence, and biomass by counting green pixels in plant images.
Are these proxies informative enough? The chlorophyll fluorescence parameter (Fv/Fm), for instance, is often measured, but in many cases variation in Fv/Fm does not reflect variation in the actual rate of photosynthesis (Poorter et al., 2019). The electron transport rate offers a better approximation, but still does not capture the true rate of C-fixation (Kalaji et al., 2014). Similarly, estimating digital biomass based on the number of green pixels in an image can provide an indication of plant size, but does not give the actual biomass, or information on biomass allocation to leaves, stems, and roots. This lack of information hampers comparisons across experiments, platforms, and the published literature. However, advancements in machine learning techniques now enable the segmentation of 2D or 3D images into leaves and stems (Golbach et al., 2016; Jin et al., 2019; Shi et al., 2019), so far only for smaller and/or specific species. Moreover, combined shoot and root phenotyping is feasible in rhizotron or agar-based platforms (Nagel et al., 2012; Nagel et al., 2020). These developments hold the potential to extract more comprehensive information from these photographs or raw sensor data.
In all cases, users must maintain a critical approach to their data, particularly when changes in plant morphology occur. A clear example is observed in drought-stressed plants where a loss in turgor can result in leaf rolling or wilting, leading to a noticeable decrease in projected leaf area (PLA; Figures 4C, D), while actual dry biomass is little affected. Even when the variable of interest is directly provided by sensors, it is wise to verify the definitions used. For one researcher, plant height may be the distance from the shoot basis to the highest plant part, for another it could be the distance from the basis to the apical meristem. In some non-destructive systems, a more statistical approach is taken, where this variable is defined as the average value of the pixels (or voxels) from the 80th till the 90th percentile with respect to height (Kjaer and Ottosen, 2015). Ideally, these definitions are included in the meta-data provided by the HTPP system.
3.9 Are non-destructive measurements sufficient?
The phenotype of plants is multifaceted, composed of hundreds of variables related to anatomy, morphology, chemical composition, carbon and water economy, growth, as well as reproduction (Lambers and Oliveira, 2019; Poorter et al., 2022; Zafaver et al., 2023). While some of these variables can be estimated non-destructively, the majority of plant traits require destructive sampling or harvesting. Consequently, HTPP systems, which are primarily non-destructive by nature, can only cover a subset of the phenotypic traits that researchers would ideally like to measure. However, by bringing sensors to the plant, additional measurements may become feasible. For example, continuous monitoring of transpiration over a plant’s lifespan in real time can be achieved by placing plants on a balance (Tardieu et al., 2017; Dalal et al., 2020). Nevertheless, many other traits can only be measured through destructive sampling or harvesting, which necessitates additional planning and manpower.
A highly promising advancement is the development of robots capable of approaching a plant and taking a leaf punch from a specific leaf blade (Alenyà et al., 2013; Foix et al., 2018). By promptly storing these samples in liquid nitrogen, a broad array of relevant biochemical analyses can be conducted, including the assessment of key metabolites and RNA expression levels (Hall et al., 2022).
3.10 What calibration curve is required?
In certain cases, well-calibrated phenotyping equipment can directly provide data on physiologically relevant variables of interest. For instance, measurements such as leaf temperature or Fv/Fm yield output that is readily biologically interpretable and can be easily related to published work in the literature. In other cases, however, additional calibration is required to transform acquired data (e.g., number of green pixels) into biological meaningful variables (e.g., shoot dry mass). In such cases, a common procedure involves periodic measurements of a subset of plants, initially through non-destructive imaging, and subsequently destructively by determining leaf area, shoot dry mass or other traits of interest. An example is shown in Figure 6, where the projected leaf area of Plantago major plants grown under different [CO2] levels was assessed non-destructively through imaging, followed by destructive measurement of total leaf area and shoot biomass (see Material & Methods).
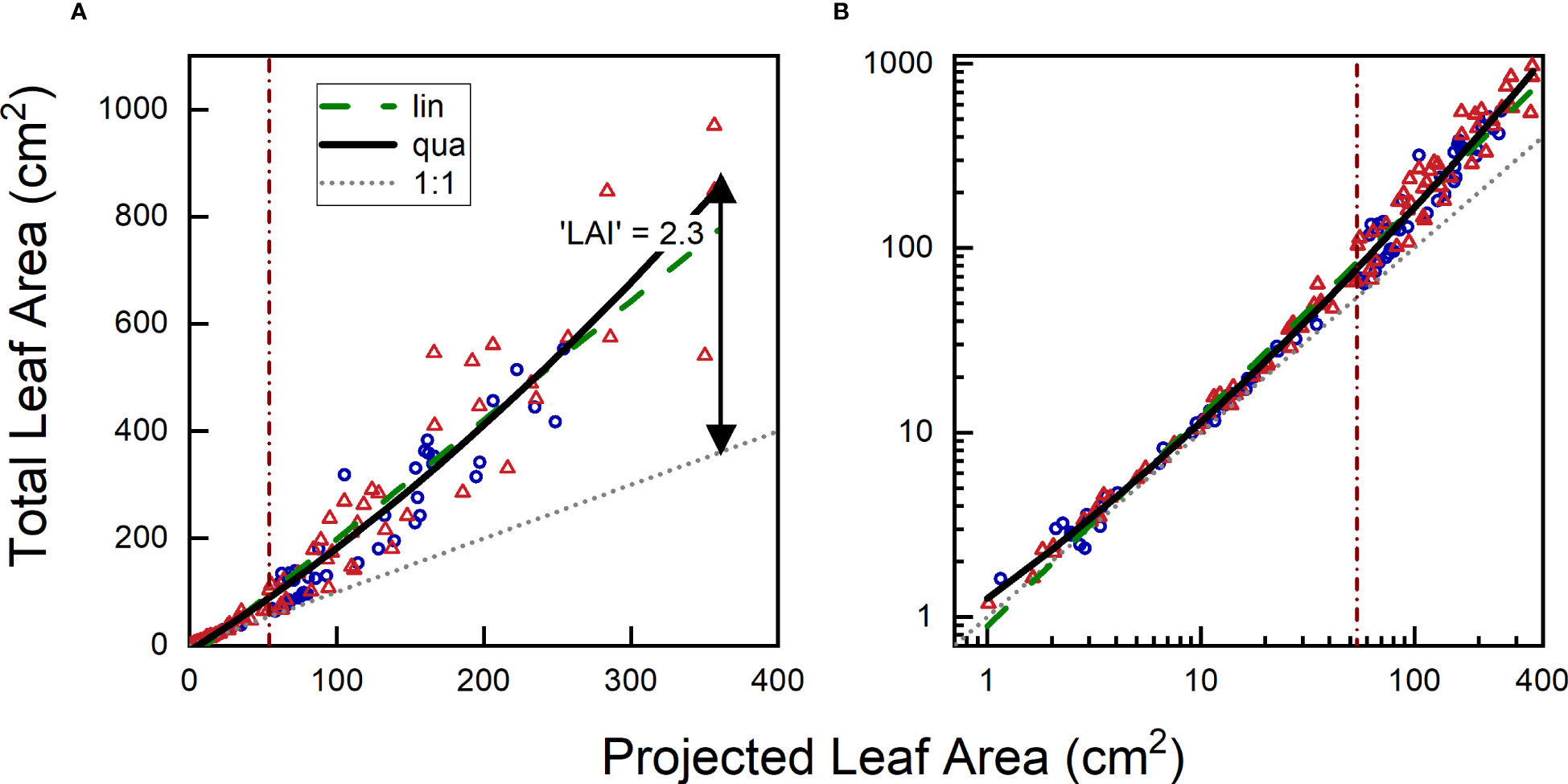
Figure 6 Relationship between projected leaf area and total leaf area of Plantago major plants harvested over an 8-week period and plotted on (A) linear or (B) logarithmic scales. Blue circles represent plants grown at low [CO2] (350 µL L-1), and orange triangles representplants grown at high [CO2] (700 µL L-1). The grey dotted line represents the 1:1 relationship between TLA (Total Leaf Area) and PLA (Projected Leaf Area), the green dashed line represents the linear fit through the data, and the black continuous line shows the quadratic fit. The vertical brown line indicates where the divide is between the 50% smaller and 50% larger plants, based on PLA.
The first step in establishing a calibration curve involves plotting the variable of interest against the measured variable. We illustrate this process with a graph that depicts the relationship between total leaf area (TLA) and projected leaf area (PLA). The graph demonstrates that for small plants (< 30 cm2 in this case, for plants up to 4 weeks old), TLA and PLA exhibit largely similar values. However, in larger plants, TLA increases at a faster rate than PLA, as newly-grown leaves will inevitably overlap partially or even fully with older leaves. In the experiment presented here, the TLA at the final harvest was approximately 2.3 times larger than the corresponding PLA, with no clear difference between plants of the two treatments.
The second step in constructing the calibration curve involves computing a regression line. Taken over both treatments, a linear regression yielded highly significant results (P < 0.001). Based on the calculated r2, we found that variation in PLA accounted for 92% of the variation in TLA. While this initial outcome may appear very satisfactory, further examination showed that the regression line underestimated TLA at very small and high TLA values, while overestimating TLA at intermediate PLA values. Given the gradual increase in leaf overlap with plant size, a curved relationship appears to be a more appropriate model. Subsequent analysis with a second-order polynomial confirmed the high significance (P<0.001) of the quadratic term, resulting in a slightly improved r2 (Table 2).
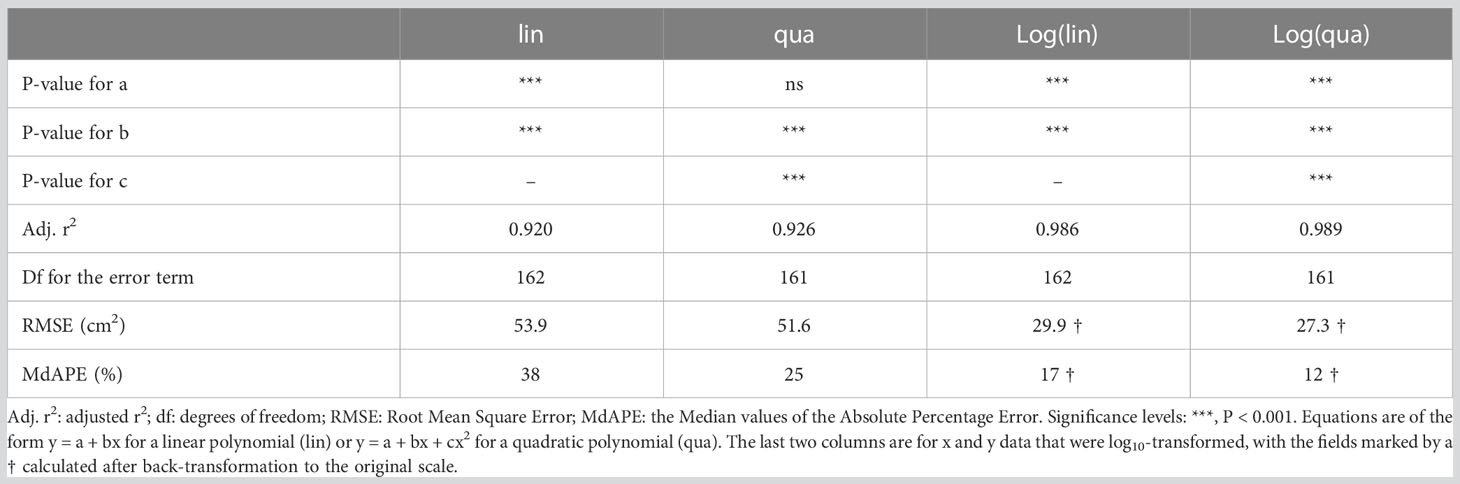
Table 2 Characterization of different calibration curves for estimating total leaf area (TLA) from projected leaf area (PLA).
Although many users are satisfied with the aforementioned correlative approach and the high r2 values (e.g. Nagel et al., 2012; Vadez et al., 2015; Banerjee et al., 2020), certain aspects warrant further inspection. For instance, the growth of young plants often follows an exponential pattern, characterized by smaller absolute size increases in small plants, and larger increases as plants grow bigger. As a consequence, in the experiment we are discussing with weekly harvests, the first half of the calibration curve is determined by 82% of the observations, while the remaining 18% contribute to the second half. To achieve a more balanced distribution, we could log-transform both PLA and TLA. In our experiment, this log-transformation resulted in the first half of the curve containing 32% of the data, with the remaining 68% in the second half (Figure 6B). Although the distribution is still not perfectly equal, it improved considerably compared to the non-transformed dataset. Performing a linear regression on the log-transformed data yielded a highly significant fit, with an r2 of 0.986. However, it is important to acknowledge the biological phenomenon of overlapping leaves. Incorporating a quadratic term into the equation further improved the fit, resulting in an r2 value of 0.989. Clearly, it pays to analyze which function is most appropriate, and whether log-transformation of the data and/or non-linear fits can provide a more robust basis for the calibration curve than a standard linear regression on non-transformed data.
3.11 How accurate is the calibration curve?
The coefficient of determination (r2) is a convenient parameter to describe the goodness of fit of a statistical relationship between variables, and because it (generally) scales between 0 and 1, it allows easy comparisons across various experiments (Chicco et al., 2021). The calibration curves discussed in the previous section all exhibit relatively high r2 values (Table 2), but does this r2 truly represent the desired accuracy? Different expressions of plant size, such as leaf number, leaf area, and total biomass, are generally well correlated. How well the calibration curve works depends partly on the appropriateness of the chosen proxy trait. For instance: a top-view picture might provide more information for a rosette plant, while side views or views from different angles could be more informative for species with a single stem. Furthermore, it is important to note that r2 provides information on the total variation in the y-variable that can be explained by the total variation in the x-variable. Hence, in monotonically increasing relationships like the one depicted in Figure 6, the larger the span in size in both x and y, the higher the r2 will be. If we restrict the calculation to plants with a PLA >100 cm2 instead of considering all data, the r2 value decreases from 0.92 to 0.76. Consequently, a high r2 for a calibration curve like the one shown in Figure 6, indicates that we are able to effectively distinguish between small and large plants. However, if a researcher’s prime interest lies in understanding the variation in final size across genotypes, relying solely on the r2 of the full calibration curve may provide a somewhat misleading sense of accuracy.
An alternative measure to assess the goodness of fit is the root mean square error (RMSE), which quantifies the average distance in the y-direction between the observed data points and the fitted line. It gives more weight to points that are further away from the line, compared to those that are closer (Hodson, 2022). The advantage of RMSE is that, all else being equal, it is not influenced by the total variation range in x and y, as is the case with r2. Additionally, RMSE provides an absolute error in the units of the Y-axis. If the residuals follow a normal distribution, it informs the researcher that there is a 68% probability that the estimated total leaf area (TLA) deviates by less than the RMSE from the true TLA. However, RMSE may not be suitable for calibration curves that cover a wider range of plant sizes, as the error is not equal for plants of all sizes. For instance, an RMSE of 5 cm2 may represent a minor deviation for a plant of 1000 cm2, but a huge variation for a plant with a leaf area of 1 cm2.
To assess the accuracy of the estimates, we calculated for each plant the absolute percentage error. This involved determining the absolute difference between the actual TLA, and the TLA value estimated from the calibration curve, normalized to the actual TLA measured. These values ranged from nearly 0%, indicating a highly accurate estimate, to over 1000% in the specific case of very young plants where TLA was fitted with a straight line across all plant sizes (Figure 7A). In the last case, the actual TLA value was 1 cm2, whereas the estimated TLA value was calculated to be -10 cm2. This illustrates that even an r2 value exceeding 0.90 does not necessarily guarantee accurate estimates for every individual plant. The median absolute percentage error (MdAPE) serves as a useful summary descriptor for non-normally distributed data. For the linear calibration curve, MdAPE was approximately 37% (Table 2; Figure 7A), indicating that the accuracy fell short of our expectations. However, when utilizing a quadratic fit with log-log transformed values, the MdAPE decreased to 11%, signifying a substantial improvement in accuracy. Additionally, in the quadratic fit, MdAPE values were lower for smaller compared to larger ones (Figure 7B), which is logical given the increased leaf overlap in larger plants.
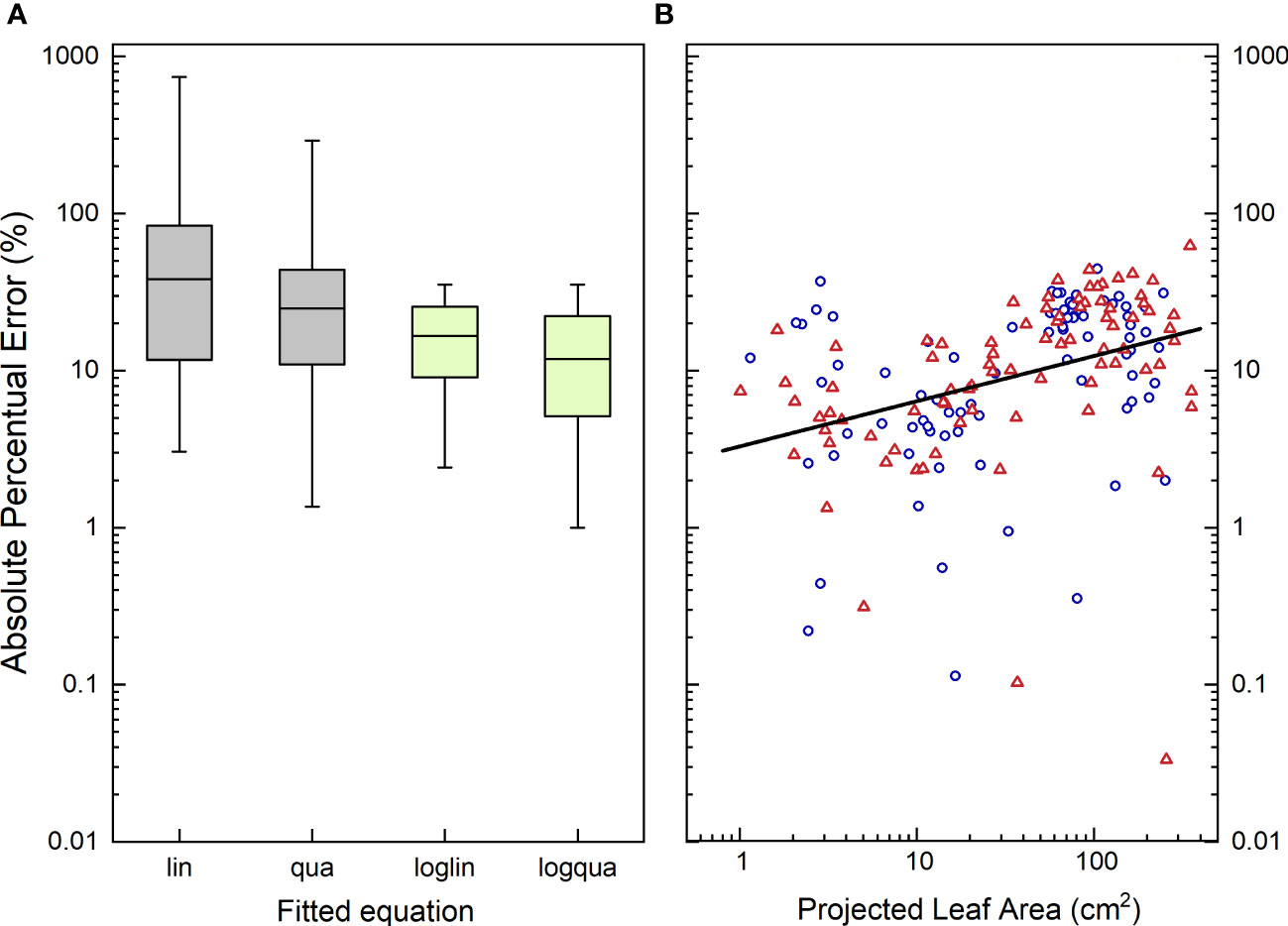
Figure 7 (A) Boxplot characterizing the distribution of the Absolute Percentage Error (APE) in the estimate of Total Leaf Area (TLA) from the measurements of Projected Leaf Area (PLA), using four different calibration curves. (B) Absolute Percentage Error in the estimate of Total Leaf Area using a log-quadratic calibration curve plotted against Projected Leaf Area. In (A), boxplots indicate the 5th, 25th, 50th, 75th and 95th percentile of the APE values, taken over all plants and treatments. lin, linear regression; qua, quadratic regression; loglin, linear regression through the log10-transormed value of TLA and PLA; logqua, quadratic regression through the log10-transformed values. In (B), blue circles represent plants grown at low [CO2] (350 µL L-1), and orange triangles represent high [CO2]-grown plants (700 µL L-1). The regression line passes through all points, and is significantly (P<0.001) different from zero, with an adjusted r2 of 0.15.
The mean absolute percentage error (MAPE) is increasingly utilized in the field of high-throughput plant phenotyping (e.g. Paproki et al., 2012; Paulus, 2019; Rossi et al., 2022). However, given the log-normal distribution of these data, we advocate for the use of the median absolute percentage error (MdAPE) as a more informative measure of the general accuracy. By employing the MdAPE, we aim to capture a representative summary of the actual accuracy, rather than relying solely on the r2 of a calibration curve.
3.12 How many calibration curves are required?
So far, we have considered a common calibration curve for both low and high CO2 plants. For the relationship between projected leaf area (PLA) and total leaf area (TLA), this approach may seem reasonable as long as the treatment does not influence leaf angle or any other aspect of leaf display. However, what would happen if we aim to use PLA to estimate shoot biomass (Figure 8A)? The relationship between leaf dry mass and leaf area is known to shift, as plants exposed to elevated CO2 almost invariably exhibit higher leaf mass per area (LMA; Poorter et al., 2022). Using quadratic polynomials on log-transformed PLA and shoot dry mass, we indeed found different curves (Figure 8B). They indicated that for a given PLA, elevated CO2 plants were 20-30% heavier, although not for the smallest or largest plants. These findings align with the LMA data, which also showed LMA averages to be 20-30% heavier, except for the first and last harvest (cf. Figure 8B in Poorter et al., 1988). However, statistically, this did not show up as a significant effect, neither for CO2 as a main factor nor for the interaction of CO2 with the linear and quadratic components of PLA.
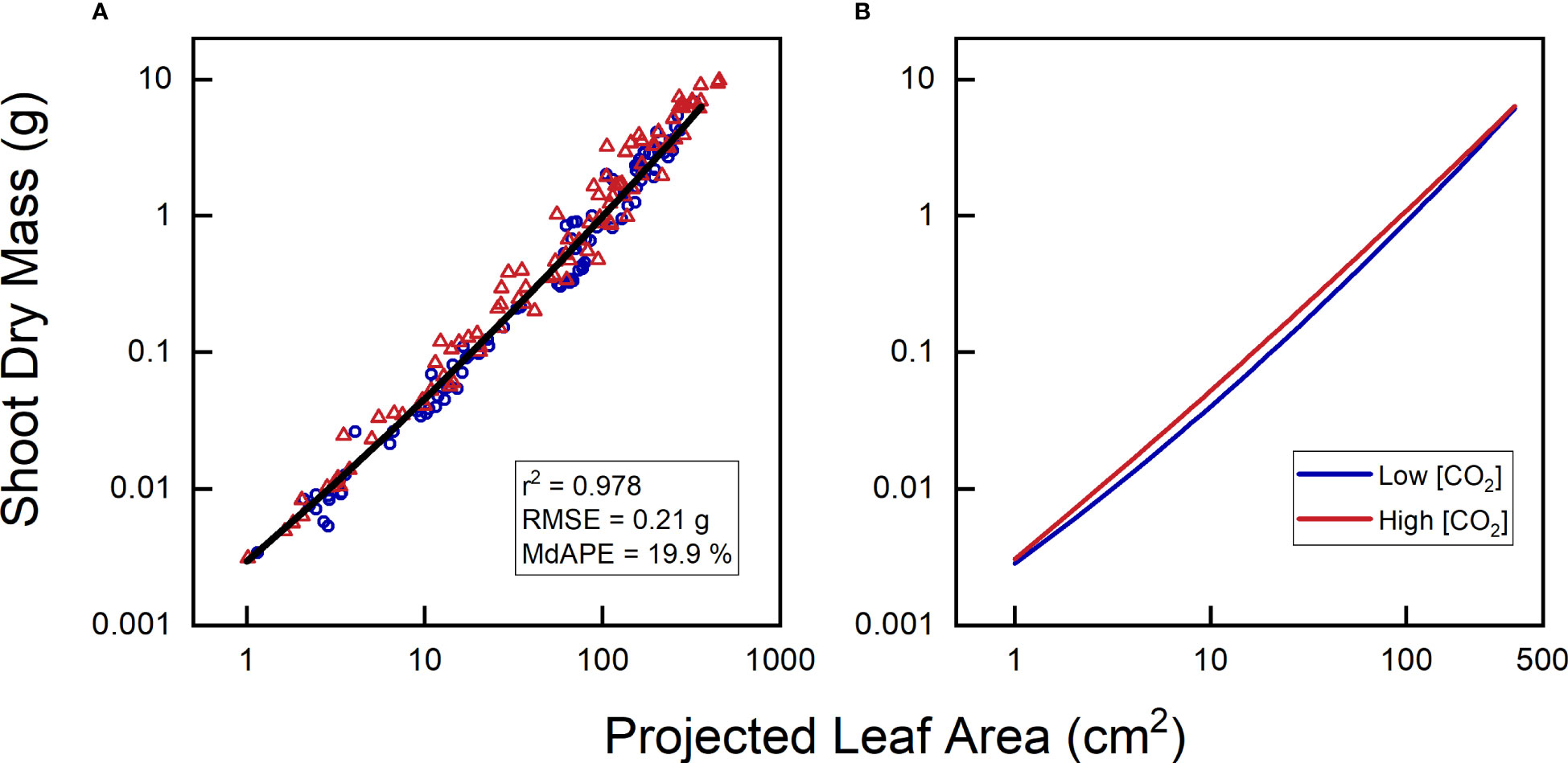
Figure 8 Relationship between projected leaf area and shoot dry mass of Plantago major, grown at low {CO2] (350 µL L-1) and high [CO2] (700 µL L-1). (A) Combined data for plants of both treatments; (B) calibration curves separately calculated for plants from each [CO2]. All curves exhibited highly significant linear and quadratic components (P < 0.001) and r2 > 0.978.
Having different calibration curves for different treatments, or maybe for different genotypes, can be quite inconvenient, as it requires more manual harvesting, partially nullifying the intended savings in human effort. What could we do about that? A well-cited method paper by Golzarian et al. (2011) discussed the phenotyping aspects of calibration curves, using the example of an experiment where plants were exposed to salinity or to control conditions. The study found that the two calibration curves were significantly different, with salt-stressed plants exhibiting higher biomass estimates for a given number of pixels compared to control plants. That would fit with the general understanding that salt stress increases leaf mass per area (LMA; Poorter et al., 2009; Kamanga et al., 2023). Golzarian et al. (2011), however, reasoned that salt-stressed plants were smaller, and considered them to be of ‘younger’ age. By adding the factor age to the equation, a single calibration function could be achieved for plants of both treatments. However, this approach mixes plant ontogeny with the direct effects of salinity, and is likely not broadly valid, especially when multiple treatments with varying salinity concentrations would be involved. Consequently, if the treatment of interest affects LMA, leaf angle, or other relevant morphological parameters, different calibration curves for different treatments may indeed be unavoidable.
Another important question to consider is the validity of a calibration curve that has been developed for a particular species, and whether it can be applied to other experiments involving the same species. In growth chambers, where light and temperature stay fixed to the same level, the transferability of a calibration curve seems more likely compared to glasshouses or field settings, where seasonal variation in light and temperature may strongly impact both LMA and stem thickness (and consequently stem mass per projected area). To use HTPP systems more effectively, calibration curves deserve more attention than they got so far. As is custom in many other laboratory methods, it might be good to regularly validate the measuring pipeline with a couple of reference samples that are measured using destructive methods.
3.13 How many replicates per genotype or treatment?
Plants grown singly in pots may show quite some variability in plant mass or other traits, which can negatively impact the statistical power to detect differences between genotypes or treatments (Poorter and Garnier, 1996). When planning the size of an HTPP system, it is important to consider not only the number of species or genotypes to be tested, but also the number of replicates per genotype that will be required. Genome-wide association studies (GWAS) or Quantitative Trait Loci (QTL) experiments often benefit more from including additional genotypes rather than increasing the number of replicates per genotype (Zou et al., 2006). If treatments are compared across many genotypes, the large number of plants grown in HTPP systems will provide sufficient statistical power for general conclusions. However, when researchers are also interested in testing specific differences between individual genotypes, the number of replicates becomes more critical. This is particularly true when a calibration curve is used to estimate the values of the trait of interest. Calibration curves with low r2 and high MdAPE introduce additional variability on top of the inherent variation that will already be present among plants. In the case of the CO2 experiment, a t-test conducted at the final harvest revealed that the actual shoot dry mass for the two treatments was only marginally significant (P = 0.10). However, the difference was far further from significance when the shoot dry mass estimates based on PLA values were used (P = 0.24). In situations where HTPP system users are interested in specific contrasts, the utilization of calibration curves implies that they may need to include more replicates than in traditional experiments to achieve the same level of statistical power.
3.14 How often do you need to repeat an experiment?
Despite the robotized and computerized systems, HTPP experiments often push limits and compare large amounts of genotypes in a standardized manner. In as far as these experiments have a background in (eco)physiological approaches, with single plants growing in pots under controlled conditions, data from a single experiment is often considered sufficient for publication. However, in agriculture it is generally regarded as the gold standard to repeat an experiment in multiple years or locations, before any importance is attached to the results.
Possibly, HTPP in controlled conditions can be seen as the initial, important step in a two-phase approach. During this first step, a wide range of genotypes or species can be tested, either in their own right or in combination with specific treatment factors. Without being overly concerned about genotypic effects on calibration curves (as mentioned in point 12), this step can be used to identify the worst-performing and best-performing genotypes, simply based on green pixel counts or similar proxy traits (‘forward phenomics’ sensu Mir et al., 2019; cf. Merlot et al., 2002). The most extreme and interesting genotypes, for example those carrying contrasting alleles for important QTLs can then be further investigated in a targeted experiment. This subsequent phase would involve non-destructive phenotyping complemented by more labor-intensive physiological analyses such as gas exchange and chemical characterization on the one hand, and destructive harvesting of both shoot and root biomass on the other hand (Poudyal et al., 2018). Such a two-step approach would also be helpful in screening a wide range of germplasm for contrasting genotypes.
4 Conclusions and outlook
In this paper, we discussed a number of issues relevant to consider during the design and implementation phases of high-throughput plant phenotyping (HTPP) systems. A crucial aspect of an HTPP platform is its alignment with the specific research questions of interest and its careful design, both from the hardware and the workflow perspective. Different treatments applied to the shoot environment (such as light, CO2, temperature) are often more complicated to implement, as this requires various growth environments all integrated into one HTPP system, or replicated HTPP systems, which is feasible but expensive. Treatments that can be applied to separate pots in the same location (such as drought, nutrients, salinity) are relatively easier to implement and amenable to computerized control. Additionally, efforts must be made to effectively address unforeseen problems and errors.
Regardless of the treatment approach, it is important to acknowledge that investment costs and maintenance requirements for most phenotyping systems are substantial. This is in part because the systems build now are often highly customized. If over time researchers will settle for more standardized systems and sensors, platform costs and time spend on complications will hopefully decrease.
Obtaining meaningful information from HTPP experiments requires to carefully consider the selection of meaningful proxy traits that enable us to answer the research questions at hand. Attention should also be paid to well-designed and regularly validated calibration curves, if necessary. An alternative strategy is to use HTPP systems as a good opportunity for prescreening. Such a prescreening would then be followed by an experiment focusing on a limited number of the most interesting genotypes. It allows to measure not only the proxy variables easily acquired by the phenotyping system (‘soft’ traits), but also the physiological ‘hard-to-get’ traits that provide valuable insights and a more comprehensive understanding of observed differences in plant performance.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
HP and PvG initiated the idea, HP, OV and AWK collected data/images. HP wrote the ms. All authors contributed their experiences and viewpoints on phenotyping and commented on the ms. All authors contributed to the article and approved the submitted version.
Funding
This paper was partially funded by the Helmholtz Association of Germany and the European Union Grant agreement No. 101059784 (CROPINNO). Open access was partly funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) – 491111487.
Acknowledgments
We thank Erik van Oosterom, Vijaya Singh and Nicolai Jablonowski for providing illustrative pictures for Box 1. Trevor Garnett and Ewaut Kissel commented on previous versions of this ms. We also appreciated the constructive comments of the reviewers. Open access was enabled by the DEAL project agreement. ChatGPT was used to check the language of the written text.
Conflict of interest
One of the authors (GMH) is co-founder of a company in high-throughput phenotyping and contributed valuable experience to this paper.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2023.1233794/full#supplementary-material
Abbreviations
HTPP, High-Throughput Plant Phenotyping; MAPE, Mean Absolute Percentage Error; MdAPE, Median Absolute Percentage Error; RMSE, Root Mean Square Error; PLA, Projected Leaf Area; TLA, Total Leaf Area.
References
Alenyà, G., Dellen, B., Foix, S., Torras, C. (2013). Robotized plant probing: Leaf segmentation utilizing time-of-flight data. IEEE Robotics Autom. Magaz. 20, 50–59. doi: 10.1109/MRA.2012.2230118
Al-Tamimi, N., Brien, C., Oakey, H., Berger, B., Saade, S., Ho, Y. S., et al. (2016). Salinity tolerance loci revealed in rice using high-throughput non-invasive phenotyping. Nat. Commun. 7, 13342. doi: 10.1038/ncomms13342
Anten, N. P., Casado-Garcia, R., Nagashima, H. (2005). Effects of mechanical stress and plant density on mechanical characteristics, growth, and lifetime reproduction of tobacco plants. Amer. Nat. 166, 650–660. doi: 10.1086/497442
Bagley, S. A., Atkinson, J. A., Hunt, H., Wilson, M. H., Pridmore, T. P., Wells, D. M. (2020). Low-cost automated vectors and modular environmental sensors for plant phenotyping. Sensors 20, 3319. doi: 10.3390/s20113319
Banerjee, B. P., Joshi, S., Thoday-Kennedy, E., Pasam, R. K., Tibbits, J., Hayden, M., et al. (2020). High-throughput phenotyping using digital and hyperspectral imaging-derived biomarkers for genotypic nitrogen response. J. Exp. Bot. 71, 4604–4615. doi: 10.1093/jxb/eraa143
Brien, C. J., Berger, B., Rabie, H., Tester, M. (2013). Accounting for variation in designing greenhouse experiments with special reference to greenhouses containing plants on conveyor systems. Plant Meth. 9, 1–22. doi: 10.1186/1746-4811-9-5
Chiang, C., Bånkestad, D., Hoch, G. (2020). Reaching natural growth: The significance of light and temperature fluctuations in plant performance in indoor growth facilities. Plants 9, 1312. doi: 10.3390/plants9101312
Chicco, D., Warrens, M. J., Jurman, G. (2021). The coefficient of determination R-squared is more informative than SMAPE, MAE, MAPE, MSE and RMSE in regression analysis evaluation. PeerJ. Comp. Sci. 7, e623. doi: 10.7717/peerj-cs.623
Dalal, A., Shenhar, I., Bourstein, R., Mayo, A., Grunwald, Y., Averbuch, N., et al. (2020). A telemetric, gravimetric platform for real-time physiological phenotyping of plant–environment interactions. J. Visual. Experim. 162, e61280. doi: 10.3791/61280
Deery, D., Jimenez-Berni, J., Jones, H., Sirault, X., Furbank, R. (2014). Proximal remote sensing buggies and potential applications for field-based phenotyping. Agronomy 4, 349–379. doi: 10.3390/agronomy4030349
Dobrescu, A., Giuffrida, M. V., Tsaftaris, S. A. (2020). Doing more with less: a multitask deep learning approach in plant phenotyping. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00141
Dobrescu, A., Scorza, L. C., Tsaftaris, S. A., McCormick, A. J. (2017). A “Do-It-Yourself” phenotyping system: measuring growth and morphology throughout the diel cycle in rosette shaped plants. Plant Meth. 13, 1–12. doi: 10.1186/s13007-017-0247-6
Dornbusch, T., Lorrain, S., Kuznetsov, D., Fortier, A., Liechti, R., Xenarios, I., et al. (2012). Measuring the diurnal pattern of leaf hyponasty and growth in Arabidopsis–a novel phenotyping approach using laser scanning. Funct. Plant Biol. 39, 860–869. doi: 10.1071/FP12018
Eberius, M., Lima-Guerra, J. (2009). “High-throughput plant phenotyping–data acquisition, transformation, and analysis,” in Bioinformatics: tools and applications eds Edwards, D., Stajich, J., Hansen, D., (New York, NY: Springer), 259–278. doi: 10.1007/978-0-387-92738-1_13
Evans, G. C. (1972). The quantitative analysis of plant growth. Berkeley and Los Angeles: Univ. California Press.
Fiorani, F., Schurr, U. (2013). Future scenarios for plant phenotyping. Annu. Rev. Plant Biol. 64, 267–291. doi: 10.1146/annurev-arplant-050312-120137
Foix, S., Alenyà, G., Torras, C. (2018). Task-driven active sensing framework applied to leaf probing. Comp. Electr. Agric. 147, 166–175. doi: 10.1016/j.compag.2018.01.020
Furbank, R. T., Tester, M. (2011). Phenomics–technologies to relieve the phenotyping bottleneck. Trends Plant Sci. 16, 635–644. doi: 10.1016/j.tplants.2011.09.005
Gibon, Y., Blaesing, O. E., Hannemann, J., Carillo, P., Hohne, M., Hendriks, J. H., et al. (2004). A robot-based platform to measure multiple enzyme activities in Arabidopsis using a set of cycling assays: comparison of changes of enzyme activities and transcript levels during diurnal cycles and in prolonged darkness. Plant Cell 16, 3304–3325. doi: 10.1105/tpc.104.025973
Gibson, G. (2018). Population genetics and GWAS: a primer. PloS Biol. 16, e2005485. doi: 10.1371/journal.pbio.2005485
Golbach, F., Kootstra, G., Damjanovic, S., Otten, G., van de Zedde, R. (2016). Validation of plant part measurements using a 3D reconstruction method suitable for high-throughput seedling phenotyping. Mach. Vision Appl. 27, 663–680. doi: 10.1007/s00138-015-0727-5
Golzarian, M. R., Frick, R. A., Rajendran, K., Berger, B., Roy, S., Tester, M., et al. (2011). Accurate inference of shoot biomass from high-throughput images of cereal plants. Plant Meth. 7, 1–11. doi: 10.1186/1746-4811-7-2
Gomez, L. D., Whitehead, C., Barakate, A., Halpin, C., McQueen-Mason, S. J. (2010). Automated saccharification assay for determination of digestibility in plant materials. Biotechn. Biofuels 3, 1–12. doi: 10.1186/1754-6834-3-23
Granier, C., Aguirrezabal, L., Chenu, K., Cookson, S. J., Dauzat, M., Hamard, P., et al. (2006). PHENOPSIS, an automated platform for reproducible phenotyping of plant responses to soil water deficit in Arabidopsis thaliana permitted the identification of an accession with low sensitivity to soil water deficit. New Phytol. 169, 623–635. doi: 10.1111/j.1469-8137.2005.01609.x
Hall, R. D., D’Auria, J. C., Ferreira, A. C. S., Gibon, Y., Kruszka, D., Mishra, P., et al. (2022). High-throughput plant phenotyping: a role for metabolomics? Trends Plant Sci. 27, 549–563. doi: 10.1016/j.tplants.2022.02.001
Hodson, T. O. (2022). Root-mean-square error (RMSE) or mean absolute error (MAE): when to use them or not. Geosci. Model. Dev. 15, 5481–5487. doi: 10.5194/gmd-15-5481-2022
Jahnke, S., Roussel, J., Hombach, T., Kochs, J., Fischbach, A., Huber, G., et al. (2016). Pheno seeder-a robot system for automated handling and phenotyping of individual seeds. Plant Physiol. 172, 1358–1370. doi: 10.1104/pp.16.01122
Janni, M., Coppede, N., Bettelli, M., Briglia, N., Petrozza, A., Summerer, S., et al. (2019). In vivo phenotyping for the early detection of drought stress in tomato. Plant Phenom. 2019, 6168209. doi: 10.34133/2019/6168209
Jansen, M., Gilmer, F., Biskup, B., Nagel, K. A., Rascher, U., Fischbach, A., et al. (2009). Simultaneous phenotyping of leaf growth and chlorophyll fluorescence via GROWSCREEN FLUORO allows detection of stress tolerance in Arabidopsis thaliana and other rosette plants. Funct. Plant Biol. 36, 902–914. doi: 10.1071/FP09095
Jin, S., Su, Y., Gao, S., Wu, F., Ma, Q., Xu, K., et al. (2019). Separating the structural components of maize for field phenotyping using terrestrial LiDAR data and deep convolutional neural networks. IEEE T. Geosci. Remote Sens. 58, 2644–2658. doi: 10.1109/TGRS.2019.2953092
Junker, A., Muraya, M. M., Weigelt-Fischer, K., Arana-Ceballos, F., Klukas, C., Melchinger, A. E., et al. (2015). Optimizing experimental procedures for quantitative evaluation of crop plant performance in high throughput phenotyping systems. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00770
Kalaji, H. M., Schansker, G., Ladle, R. J., Goltsev, V., Bosa, K., Allakhverdiev, S. I., et al. (2014). Frequently asked questions about in vivo chlorophyll fluorescence: practical issues. Photosynth. Res. 122, 121–158. doi: 10.1007/s11120-014-0024-6
Kamanga, R. M., Kopa, F., Kamala, F. D., Sefasi, A., Ndakidemi, P. A. (2023). Screening and evaluation of salinity stress tolerance in local Malawian tomato cultivars. Plant Physiol. Rep. 28, 259–271. doi: 10.1007/s40502-023-00718-8
Kjaer, K. H., Ottosen, C. O. (2015). 3D laser triangulation for plant phenotyping in challenging environments. Sensors 15, 13533–13547. doi: 10.3390/s150613533
Krajewski, P., Chen, D., Ćwiek, H., van Dijk, A. D., Fiorani, F., Kersey, P., et al. (2015). Towards recommendations for metadata and data handling in plant phenotyping. J. Exp. Bot. 66, 5417–5427. doi: 10.1093/jxb/erv271
Liu, H., Bruning, B., Garnett, T., Berger, B. (2020). The performances of hyperspectral sensors for proximal sensing of nitrogen levels in wheat. Sensors 20, 4550. doi: 10.3390/s20164550
Lorence, A., Jimenez, K. M. (2022). High-throughput plant phenotyping: methods and protocols (Methods in molecular biology vol. 2539) (New York: Humana Press), ISBN: ISBN 978-1-0716-2537-8.
Mahlein, A. K., Kuska, M. T., Thomas, S., Wahabzada, M., Behmann, J., Rascher, U., et al. (2019). Quantitative and qualitative phenotyping of disease resistance of crops by hyperspectral sensors: seamless interlocking of phytopathology, sensors, and machine learning is needed! Curr. Opin. Plant Biol. 50, 156–162. doi: 10.1016/j.pbi.2019.06.007
Merlot, S., Mustilli, A. C., Genty, B., North, H., Lefebvre, V., Sotta, B., et al. (2002). Use of infrared thermal imaging to isolate Arabidopsis mutants defective in stomatal regulation. Plant J. 30, 601–609. doi: 10.1046/j.1365-313X.2002.01322.x
Mir, R. R., Reynolds, M., Pinto, F., Khan, M. A., Bhat, M. A. (2019). High-throughput phenotyping for crop improvement in the genomics era. Plant Sci. 282, 60–72. doi: 10.1016/j.plantsci.2019.01.007
Nabel, M., Schrey, S. D., Poorter, H., Koller, R., Nagel, K. A., Temperton, V. M., et al. (2018). Coming late for dinner: Localized digestate depot fertilization for extensive cultivation of marginal soil with Sida hermaphrodita. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01095
Nagel, K. A., Lenz, H., Kastenholz, B., Gilmer, F., Averesch, A., Putz, A., et al. (2020). The platform GrowScreen-Agar enables identification of phenotypic diversity in root and shoot growth traits of agar grown plants. Plant Meth. 16, 89. doi: 10.1186/s13007-020-00631-3
Nagel, K. A., Putz, A., Gilmer, F., Heinz, K., Fischbach, A., Pfeifer, J., et al. (2012). GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 39, 891–904. doi: 10.1071/FP12023
Papoutsoglou, E. A., Faria, D., Arend, D., Arnaud, E., Athanasiadis, I. N., Chaves, I., et al. (2020). Enabling reusability of plant phenomic datasets with MIAPPE 1.1. New Phytol. 227, 260–273. doi: 10.1111/nph.16544
Paproki, A., Sirault, X., Berry, S., Furbank, R., Fripp, J. (2012). A novel mesh processing-based technique for 3D plant analysis. BMC Plant Biol. 12, 1–13. doi: 10.1186/1471-2229-12-63
Passioura, J. B. (2002). Soil conditions and plant growth. Plant Cell Environ. 25, 311–318. doi: 10.1046/j.0016-8025.2001.00802.x
Paulus, S. (2019). Measuring crops in 3D: using geometry for plant phenotyping. Plant Meth. 15, 1–13. doi: 10.1186/s13007-019-0490-0
Pommier, C., Michotey, C., Cornut, G., Roumet, P., Duchêne, E., Flores, R., et al. (2019). Applying FAIR principles to plant phenotypic data management in GnpIS. Plant Phenom. 2019, 1671403. doi: 10.34133/2019/1671403
Poorter, H., Bühler, J., van Dusschoten, D., Climent, J., Postma, J. A. (2012). Pot size matters: a meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 39, 839–850. doi: 10.1071/FP12049
Poorter, H., Fiorani, F., Pieruschka, R., Wojciechowski, T., van der Putten, W. H., Kleyer, M., et al. (2016). Pampered inside, pestered outside? Differences and similarities between plants growing in controlled conditions and in the field. New Phytol. 212, 838–855. doi: 10.1111/nph.14243
Poorter, H., Garnier, E. (1996). Plant growth analysis: an evaluation of experimental design and computational methods. J. Exp. Bot. 47, 1343–1351. doi: 10.1093/jxb/47.12.1969
Poorter, H., Knopf, O., Wright, I. J., Temme, A. A., Hogewoning, S. W., Graf, A., et al. (2022). A meta-analysis of responses of C3 plants to atmospheric CO2: dose–response curves for 85 traits ranging from the molecular to the whole-plant level. New Phytol. 233, 1560–1596. doi: 10.1111/nph.17802
Poorter, H., Niinemets, Ü., Ntagkas, N., Siebenkäs, A., Mäenpää, M., Matsubara, S., et al. (2019). A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 223, 1073–1105. doi: 10.1111/nph.15754
Poorter, H., Niinemets, Ü., Poorter, L., Wright, I. J., Villar, R. (2009). Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytol. 182, 565–588. doi: 10.1111/j.1469-8137.2009.02830.x
Poorter, H., Pot, S., Lambers, H. (1988). The effect of an elevated atmospheric CO2 concentration on growth, photosynthesis and respiration of Plantago major. Physiol. Plant 73, 553–559. doi: 10.1111/j.1399-3054.1988.tb05440.x
Poudyal, D., Rosenqvist, E., Ottosen, C. O. (2018). Phenotyping from lab to field–tomato lines screened for heat stress using Fv/Fm maintain high fruit yield during thermal stress in the field. Funct. Plant Biol. 46, 44–55. doi: 10.1071/FP17317
R Core Team (2022). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Reynolds, D., Baret, F., Welcker, C., Bostrom, A., Ball, J., Cellini, F., et al. (2019). What is cost-efficient phenotyping? Optimizing costs for different scenarios. Plant Sci. 282, 14–22. doi: 10.1016/j.plantsci.2018.06.015
Rosa, L. M., Forseth, I. N. (1996). Diurnal patterns of soybean leaf inclination angles and azimuthal orientation under different levels of ultraviolet-B radiation. Agric. For. Meteor. 78, 107–119. doi: 10.1016/0168-1923(95)02249-X
Rossi, R., Costafreda-Aumedes, S., Leolini, L., Leolini, C., Bindi, M., Moriondo, M. (2022). Implementation of an algorithm for automated phenotyping through plant 3D-modeling: A practical application on the early detection of water stress. Comput. Electron. Agric. 197, 106937. doi: 10.1016/j.compag.2022.106937
Roth, L., Barendregt, C., Bétrix, C. A., Hund, A., Walter, A. (2022). High-throughput field phenotyping of soybean: Spotting an ideotype. Remote Sens. Environ. 269, 112797. doi: 10.1016/j.rse.2021.112797
Shi, W., van de Zedde, R., Jiang, H., Kootstra, G. (2019). Plant-part segmentation using deep learning and multi-view vision. Biosyst. Eng. 187, 81–95. doi: 10.1016/j.biosystemseng.2019.08.014
Singh, V., van Oosterom, E. J., Jordan, D. R., Messina, C. D., Cooper, M., Hammer, G. L. (2010). Morphological and architectural development of root systems in sorghum and maize. Plant Soil 333, 287–299. doi: 10.1007/s11104-010-0343-0
Tardieu, F., Cabrera-Bosquet, L., Pridmore, T., Bennett, M. (2017). Plant phenomics, from sensors to knowledge. Curr. Biol. 27, R770–R783. doi: 10.1016/j.cub.2017.05.055
Thompson, A., Kantar, M., Rainey, K. (2022). “Designing experiments for physiological phenomics,” in High-throughput plant phenotyping: methods and protocols (New York, NY: Springer US), 159–170.
Vadez, V., Kholová, J., Hummel, G., Zhokhavets, U., Gupta, S. K., Hash, C. T. (2015). LeasyScan: a novel concept combining 3D imaging and lysimetry for high-throughput phenotyping of traits controlling plant water budget. J. Exp. Bot. 66, 5581–5593. doi: 10.1093/jxb/erv251
Vargas, J. Q., Bendig, J., Mac Arthur, A., Burkart, A., Julitta, T., Maseyk, K., et al. (2020). Unmanned aerial systems (UAS)-based methods for solar induced chlorophyll fluorescence (SIF) retrieval with non-imaging spectrometers: state of the art. Remote Sens. 12, 1624. doi: 10.3390/rs12101624
Walter, A., Liebisch, F., Hund, A. (2015). Plant phenotyping: from bean weighing to image analysis. Plant Meth. 11, 1–11. doi: 10.1186/s13007-015-0056-8
Walter, A., Scharr, H., Gilmer, F., Zierer, R., Nagel, K. A., Ernst, M., et al. (2007). Dynamics of seedling growth acclimation towards altered light conditions can be quantified via GROWSCREEN: a setup and procedure designed for rapid optical phenotyping of different plant species. New Phytol. 174, 447–455. doi: 10.1111/j.1469-8137.2007.02002.x
White, J. W., Conley, M. M. (2013). A flexible, low-cost cart for proximal sensing. Crop Sci. 53, 1646–1649. doi: 10.2135/cropsci2013.01.0054
Wiese, A., Christ, M. M., Virnich, O., Schurr, U., Walter, A. (2007). Spatio-temporal leaf growth patterns of Arabidopsis thaliana and evidence for sugar control of the diel leaf growth cycle. New Phytol. 174, 752–761. doi: 10.1111/j.1469-8137.2007.02053.x
Wilkinson, M. D., Dumontier, M., Aalbersberg, I. J., Appleton, G., Axton, M., Baak, A., et al. (2016). The FAIR Guiding Principles for scientific data management and stewardship. Sci. Data 3, 1–9. doi: 10.1038/sdata.2016.18
Xiao, Q., Bai, X., Zhang, C., He, Y. (2022). Advanced high-throughput plant phenotyping techniques for genome-wide association studies: A review. J. Adv. Res. 35, 215–230. doi: 10.1016/j.jare.2021.05.002
Xu, L., Cruz, J. A., Savage, L. J., Kramer, D. M., Chen, J. (2015). Plant photosynthesis phenomics data quality control. Bioinformatics 31, 1796–1804. doi: 10.1093/bioinformatics/btu854
Yang, W., Guo, Z., Huang, C., Duan, L., Chen, G., Jiang, N., et al. (2014). Combining high-throughput phenotyping and genome-wide association studies to reveal natural genetic variation in rice. Nat. Commun. 5, 5087. doi: 10.1038/ncomms6087
Ye, X., Abe, S., Zhang, S. (2020). Estimation and mapping of nitrogen content in apple trees at leaf and canopy levels using hyperspectral imaging. Precis. Agric. 21, 198–225. doi: 10.1007/s11119-019-09661-x
Zavafer, A., Bates, H., Mancilla, C., Ralph, P. J. (2023). Phenomics: conceptualization and importance for plant physiology. Trends Plant Sci. 28, 1004-1013. doi: 10.1016/j.tplants.2023.03.023
Keywords: calibration curve, digital biomass, high-throughput plant phenotyping, leaf mass per area, sensors
Citation: Poorter H, Hummel GM, Nagel KA, Fiorani F, von Gillhaussen P, Virnich O, Schurr U, Postma JA, van de Zedde R and Wiese-Klinkenberg A (2023) Pitfalls and potential of high-throughput plant phenotyping platforms. Front. Plant Sci. 14:1233794. doi: 10.3389/fpls.2023.1233794
Received: 02 June 2023; Accepted: 01 August 2023;
Published: 23 August 2023.
Edited by:
Hanwei Mei, Shanghai Agrobiological Gene Center, ChinaReviewed by:
Joaquim Miguel Costa, University of Lisbon, PortugalDimitrios Fanourakis, Technological Educational Institute of Crete, Greece
Copyright © 2023 Poorter, Hummel, Nagel, Fiorani, von Gillhaussen, Virnich, Schurr, Postma, van de Zedde and Wiese-Klinkenberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hendrik Poorter, aC5wb29ydGVyQGZ6LWp1ZWxpY2guZGU=
†ORCID: Hendrik Poorter, orcid.org/0000-0001-9900-2433
Grégoire M. Hummel, orcid.org/0009-0002-3639-0461
Kerstin A. Nagel, orcid.org/0000-0003-3025-0388
Fabio Fiorani, orcid.org/0000-0001-8775-1541
Philipp von Gillhaussen, orcid.org/0009-0001-7466-696X
Ulrich Schurr, orcid.org/0000-0003-0369-8777
Johannes A. Postma, orcid.org/0000-0002-5222-6648
Rick van de Zedde, orcid.org/0000-0002-8394-4538
Anika Wiese-Klinkenberg, orcid.org/0000-0001-9188-809X