 Alma Balestrazzi1*
Alma Balestrazzi1* Cinzia Calvio1
Cinzia Calvio1 Anca Macovei1
Anca Macovei1 Andrea Pagano1
Andrea Pagano1 Patrick Laux2
Patrick Laux2 Hassane Moutahir2Loїc Rajjou3
Hassane Moutahir2Loїc Rajjou3 Eleni Tani4Dimosthenis Chachalis5Christos Katsis6Lamiae Ghaouti7Said Gmouh8Sanaa Majid9Amine Elleuch10
Eleni Tani4Dimosthenis Chachalis5Christos Katsis6Lamiae Ghaouti7Said Gmouh8Sanaa Majid9Amine Elleuch10 Moez Hanin10Bassem Khemakhem10Hanen El Abed10
Moez Hanin10Bassem Khemakhem10Hanen El Abed10 Joao Nunes11Susana Araújo11†
Joao Nunes11Susana Araújo11† Aziz Benhamrouche12Mohand Bersi12
Aziz Benhamrouche12Mohand Bersi12- 1Department of Biology and Biotechnology “L. Spallanzani”, University of Pavia, Pavia, Italy
- 2Institute of Meteorology and Climate Research (IMK-IFU), Karlsruhe Institute of Technology, Garmisch-Partenkirchen, Germany
- 3Université Paris-Saclay, National Research Institute for Agriculture, Food and the Environment (INRAE), AgroParisTech, Institut Jean-Pierre Bourgin (IJPB), Versailles, France
- 4Laboratory of Plant Breeding and Biometry, Department of Crop Science, Agricultural University of Athens, Athens, Greece
- 5Department of Pesticides’ Control and Phytopharmacy, Benaki Phytopathological Institute, Athens, Greece
- 6Agroland S.A., Sofades, Greece
- 7Department of Plant Production, Protection and Biotechnology, Hassan II Institute of Agronomy and Veterinary Medicine, Rabat, Morocco
- 8Laboratory Laboratory of Engineering and Materials (LIMAT), Faculty of Sciences Ben M’sick, University Hassan II of Casablanca, Casablanca, Morocco
- 9Laboratory GeMEV, Faculty of Sciences Aïn Chock, University Hassan II of Casablanca, Casablanca, Morocco
- 10Higher Institute of Biotechnology, University of Sfax, Sfax, Tunisia
- 11Center Bio R&D Unit, Association BLC3–Technology and Innovation Campus, Oliveira do Hospital, Portugal
- 12Institute of Architecture and Earth Science, University Ferhat Abbas-Setif 1, Setif, Algeria
In developing countries, orphan legumes stand at the forefront in the struggle against climate change. Their high nutrient value is crucial in malnutrition and chronic diseases prevention. However, as the ‘orphan’ definition suggests, their seed systems are still underestimated and seed production is scanty. Seed priming is an effective, sustainable strategy to boost seed quality in orphan legumes for which up-to-date guidelines are required to guarantee reliable and reproducible results. How far are we along this path? What do we expect from seed priming? This brings to other relevant questions. What is the socio-economic relevance of orphan legumes in the Mediterranean Basin? How to potentiate a broader cultivation in specific regions? The case study of the BENEFIT-Med (Boosting technologies of orphan legumes towards resilient farming systems) project, developed by multidisciplinary research networks, envisions a roadmap for producing new knowledge and innovative technologies to improve seed productivity through priming, with the long-term objective of promoting sustainability and food security for/in the climate-sensitive regions. This review highlights the existing drawbacks that must be overcome before orphan legumes could reach the state of ‘climate-ready crops’. Only by the integration of knowledge in seed biology, technology and agronomy, the barrier existing between research bench and local agricultural fields may be overcome, generating high-impact technical innovations for orphan legumes. We intend to provide a powerful message to encourage future research in line with the United Nations Agenda 2030 for Sustainable Development.
1 Introduction
In the climate-vulnerable regions, local populations are exposed to high mortality rate due to malnutrition exacerbated by climate change. The complex interactions between climate-change and food systems, and their domino effect on human health, represent a major challenge that requires urgent and enduring actions (Hertel et al., 2021; Zurek et al., 2022). Mitigation strategies have been designed to speed up the transition towards sustainable, climate-resilient food systems, e.g. those reported by the Intergovernmental Panel on Climate Change: demand-side changes, protection of ecosystems, improved farm management, and decarbonizing supply chain (IPCC, 2022). Mitigation strategies involve multi-level players, such as political, financial, industrial, and scientific actors, together with the civil society (IPCC, 2022; Global Panel on Agriculture and Food Systems for Nutrition, 2023).
Major crops show poor genetic diversity and high vulnerability to extreme climate events, implying their replacement by a new generation of ‘climate-ready’ crops (Bourke et al., 2021; Xiong et al., 2022). This goal can be achieved by valorizing local minor crops (orphan crops or neglected, underutilized species-NUS) that currently have very limited impact on the market but are yet essential for small farmers’ survival in developing countries (Tadele, 2019; Popoola et al., 2022). According to FAO (2019), these species “have lost relevance over the last 500 years due to societal, agronomic, or biological factors”. Orphan crops include legumes, appreciated for their drought tolerance and capacity to improve soil fertility (Popoola et al., 2022), on top of their nutritional and medicinal features (Ogbole et al., 2023). These species and their seed systems are still undervalued, despite the crucial role they play in supporting human and animal nutrition, and favoring the growth of other crops (Cullis and Kunert, 2017; Popoola et al., 2022). This work provides a multidisciplinary and collaborative vision for the creation of new solutions allowing to cope with the challenges related to orphan legumes, with a particular focus on seed quality issues.
2 Orphan legumes as ‘climate-ready’ crops
Major crops are not able to adapt to climate change. Conversely, orphan legumes are ‘climate-smart’ plants with outstanding agronomic features, able to support sustainable livelihood, even under harsh environments (Popoola et al., 2022). Orphan legumes did not undergo the intensive breeding selection experienced by major crops, thus retaining a huge genetic variability and unique assortments of stress resilience traits. These enormous genetic resources are exposed to erosion, linked to their underutilization, reducing the chances of their future exploitation (Popoola et al., 2022; Kumar et al., 2023). Orphan legumes are poorly represented in germplasm collections and lack formal seed supply systems. They are usually identified based on their local relevance, adaptation to marginal areas, agroecological niches. Their cultivation and use are still relying on local knowledge. The limited market potential and lack of functional value chains hamper full valorization. To revert this status, several actions are needed at different levels: policies should encourage basic/applied research on these species, breeding programs should be established to obtain improved hybrids, agrifood industries should diversify their investments including orphan legumes, and an increased consumers’ awareness should stimulate the process from the basis, laying at the intersection between supply and demand (Cullis et al., 2019; Borelli et al., 2020; Elolu et al., 2023).
Orphan legumes are native to Africa, Asia, South America, North America, South Pacific, and Australia, although some species are classified as orphan in one location and as conventional crops in other areas (Talabi et al., 2022). Institutions involved in the identification and the proper management of orphan legumes are shown in Table 1. A list of underutilized crops (Future Smart Food) was compiled by the African Orphan Crops Consortium (AOCC) and the Agriculture Organization of the United Nations Regional Office for Asia and the Pacific (FAO/RAP), according to high nutritional profile, climate, resilience, local availability, and economic viability.
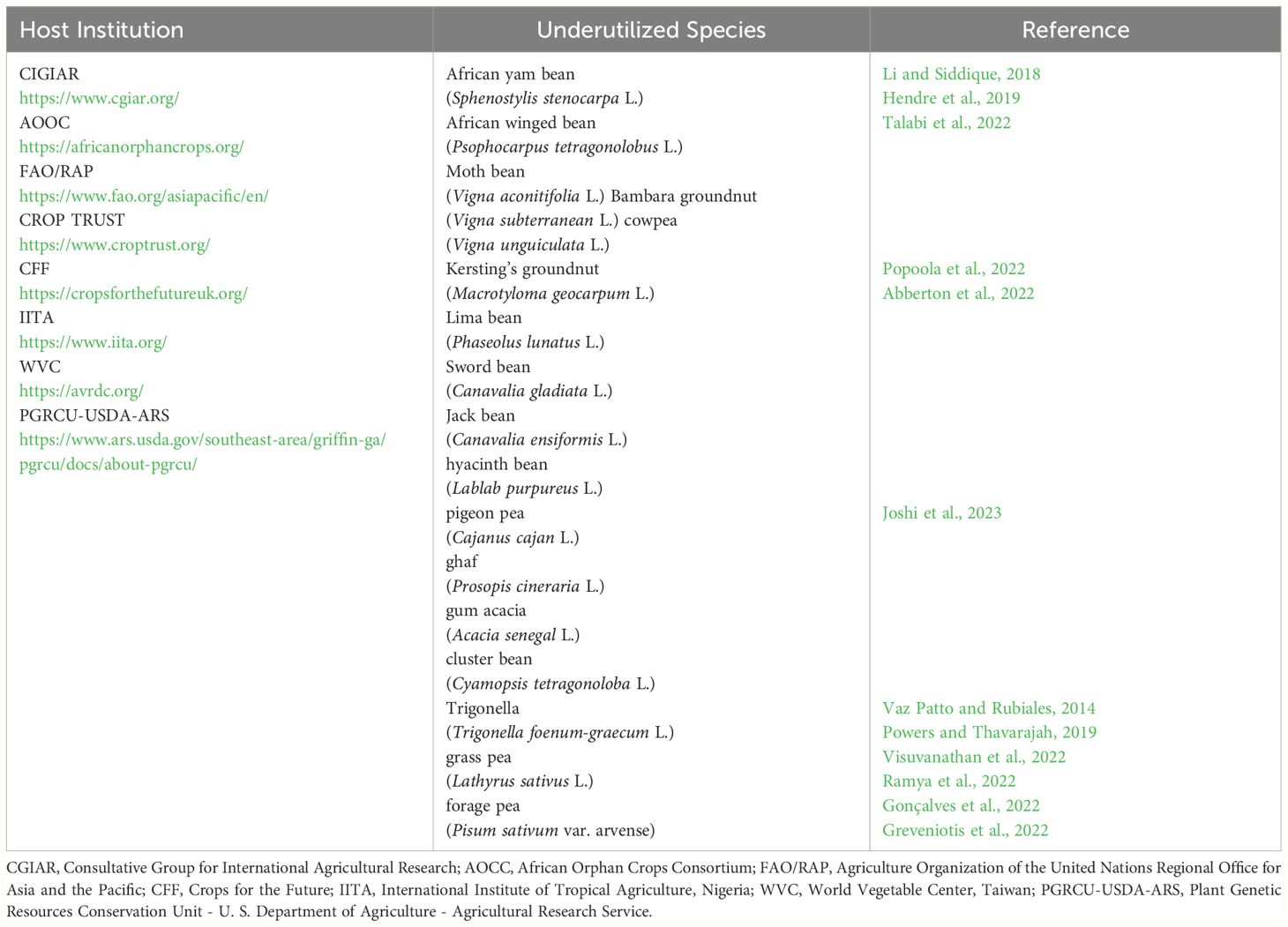
Table 1 List of International Istitutions that currently host germplasm collections of orphan legumes.
3 How would seed quality impact the productivity of orphan legumes under the challenges of climate change?
3.1 Seed systems: a variegated landscape moving towards climate-resilience
Enhanced seed vigor is the first step towards the valorization of any crop and, in particular, of those missing an official seed supply system. Seed systems include all the activities for variety development (Louwaars and Manicad, 2022). They are classified into categories that often co-exist in the same territory, resulting in a variegated landscape (Figure 1). Community-based seed systems rely on experienced farmers who take care of seeds provided by other farmers and share the improved products (Gonsalves, 2013). Besides a special care in ensuring a wide agrobiodiversity, the current challenges for community-based seed systems refer to a more inclusive approach aimed at engaging increased numbers of local farmers, the organization of wide and frequent participatory activities, as well as the development of strategic partnerships with public/private sectors. Community-based seed systems are the core of local enterprises (Vernooy et al., 2022). In formal seed system, activities are strictly regulated. Seed producers are registered and possess a certification under the control of public institutions. Certified seeds are also produced in collaboration with private seed companies (Kansiime et al., 2021). Small farmers still rely on informal seed system in which seeds are acquired through personal exchanges, gifts, local markets. However, informally traded seeds are at risk in terms of genetic purity, viability and productivity. Informal systems, the main source of seeds in climate-vulnerable regions, do not provide deal with ‘breeder seed’ (genetically and physically pure seeds propagated under strict control) and ‘basic seed’ (produced by registered growers, under the control of certification agencies) (Vernooy et al., 2022). Finally, semi-formal seed systems integrate the formal and informal systems. They are represented by farmers who manage all the value chain steps, reaching enhanced seed quality with the support of technical and financial partners (ICRISAT, 2008). In response to adverse environments, the different seed systems provide complementary characteristics (Figure 1), and seeds can be moved from the informal to the formal context (or vice versa) to buffer such changes (Louwaars and Manicad, 2022). However, this is not sufficient to mitigate the impact of severe climate change and ensure food security. The resilience of this variegated landscape should be reinforced not only in terms of scientific/technical skills, required for proper management of germplasm collections, but also by supporting socio-economic and political strategies. The goal is the design of political roadmaps to protect and provide local farmers with the most adapted varieties, in a rapid and efficient manner (Louwaars and Manicad, 2022).
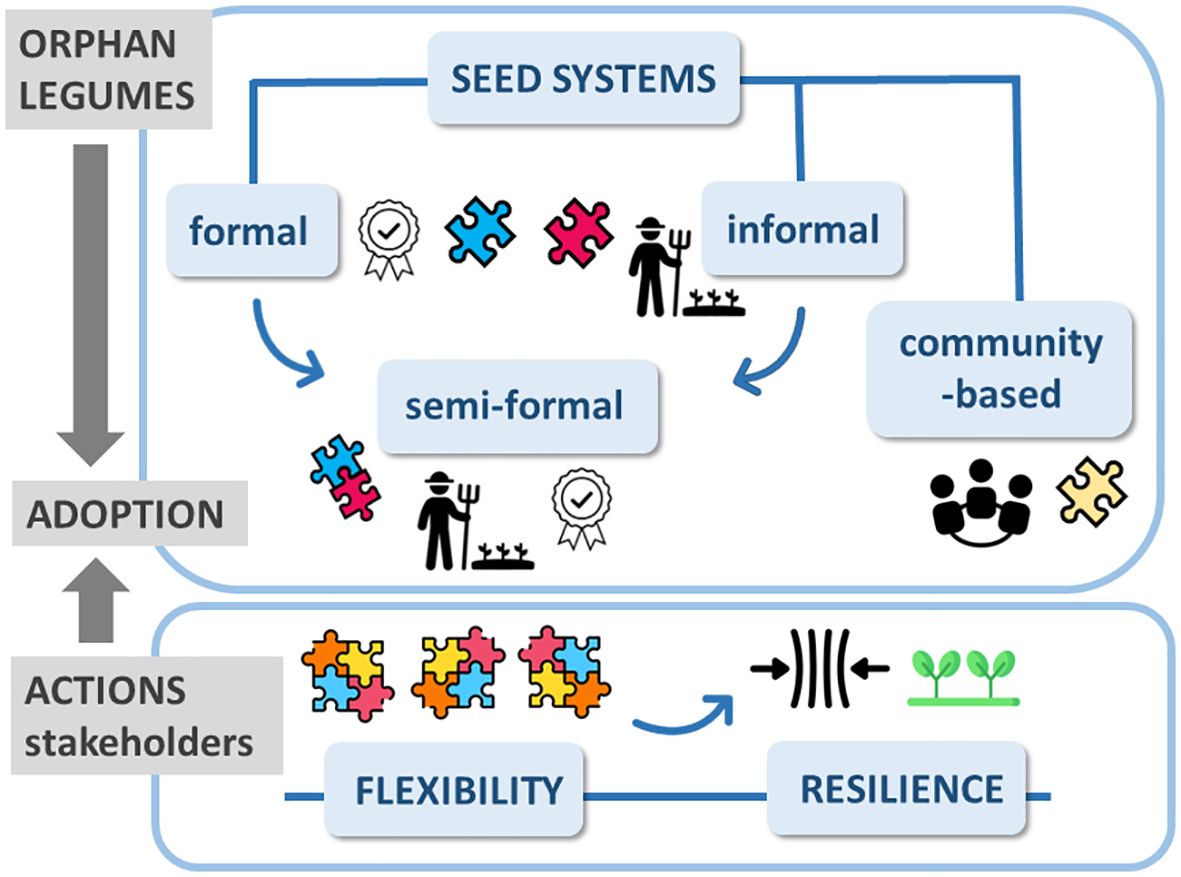
Figure 1 Seed systems are crucial vehicles that allow farmers to acquire good quality seeds of new varieties, playing an essential role in the process of orphan legumes adoption in the climate-vulnerable regions of Mediterranean area. They are part of a variegated puzzle, since different types of seed systems currently exist: formal, semi-formal, informal, and community-based seed systems. Actions should be taken in order to promote remodelling and integration of the different seed systems, based on local agronomic, technical, socio-economic, and cultural needs. Stakeholders, including policy makers, are committed to provide such flexibility, as a key factor towards better climate resilience of local agroecosystems.
3.2 Seed traits for climate-resilience
For the correct management/selection of the seeds to be used in challenging conditions, a fundamental asset is the identification of the most relevant seed traits in terms of climate resilience. Detailed agronomic and genetic information is not generally available for orphan legumes. Thus, it is imperative to link phenotype- and genotype-based information to be able to exploit these traits for future breeding strategies (Chapman et al., 2022). The identification of valuable genetic characteristics relies on the identification of quantitative trait loci (QTLs), development of codominant molecular markers and linkage maps (Oluwole et al., 2021). In Kersting’s groundnut, genome wide association studies (GWAS) identified SNP (single nucleotide polymorphism)-based marker-trait associations related to seed size/weight (Akohoue et al., 2020). GWAS performed on cowpea and castor bean accessions retrieved candidate genes associated with seed size, involved in endosperm/embryo development, and cell elongation (Lo et al., 2019; Xu et al., 2021). Increase in seed size was reported to be associated with enhanced drought tolerance, making this trait very promising in future breeding strategies (Kaydan and Yagmur, 2008; Gholami et al., 2009; Mut and Akay, 2010; Čanak et al., 2020; Martínez-López et al., 2020; Christie et al., 2022). There is also increasing evidence for a cross-talk between drought stress-mediated signaling and endogenous molecular pathways can control the final seeds size (Lv et al., 2019). It has been reported that seed size contributes to drought stress tolerance in camelina (Camelina sativa L.), however the response varies depending on biotype and intensity of the imposed stress (Čanak et al., 2020). These findings highlight the need to assess the plant performance under field conditions. Conversely, a larger seed size entails higher management costs by smallholder farmers and increased risk for seed damaging during storage and transport (Ojiewo et al., 2018).
Genes controlling seed dispersal and facilitating harvesting, as well as genes related to seed size may contribute to reduce farmers’ time and efforts. Seed dispersal dynamics allow plant adaptation to climate change by facilitating the migration of species at risk (Fricke et al., 2022). The mutualistic interactions with animals acting as vehicles of dispersal is increasingly affected by the disruptive impact of climate change, and this contributes to further threaten the viability of ecosystems (Fricke et al., 2022). Seed dispersal is facing rapid changes in response to climate change. This trait, resulting from a complex, polygenic genetic background, is still poorly investigated in both crop and model plants (Saastamoinen et al., 2018). In the specific case of crop wild relatives, source of unexplored gene pools for resilience, improved knowledge on this and other seed-related traits is required to better manage long-term conservation (Brǿnnvik and von Wettberg, 2019). Seed yield losses caused by pod dehiscence in forage legumes has been targeted during domestication (Liu et al., 2019; Parker et al., 2021). A better understanding of the mechanisms controlling such a process can help improving the production of forage legumes, particularly those highly resilient species, so far neglected by breeders (Parker et al., 2021; Guo et al., 2022).
Traits that influence the seed nutritional content, the cooking time, and processability have been also identified (Dawson et al., 2019). Climate-resilient crops must be able to provide healthy foods enriched with essential vitamins and minerals (iron, zinc), folate, and fiber. This is typical of neglected species producing highly nutritious seed grains, proxy of food security. In-depth research is necessary, to disclose the properties of such orphan crops and speed up the diversification of healthy diets for the global population (Zenda et al., 2021). Such efforts include the identification of major QTLs, genes and metabolic pathways that shapes the nutritional profile of seeds. This is the case of QTLs controlling iron and zinc concentration in chickpea seeds (Upadhyaya et al., 2016), oil and protein content in soybean (Huang et al., 2020), oil content, protein content, and fatty acids (linoleic and oleic acids) in groundnut (Roorkiwal et al., 2021). Cooking times are influenced by storage, environment, and genotype (Cichy et al., 2019). There is poor knowledge concerning the genetic variation of this trait. Studies carried out using fast and slow cooking bean (Phaseolus vulgaris L.) genotypes with different seed types highlighted significant correlations with seed coat thickness and cotyledon cell wall (Bassett et al., 2021). Pasting and cooking properties were assessed in a germplasm collection of orphan legumes that included grass pea (Santos et al., 2018). Multiple seed traits, e.g. size, shape, color, surface influenced physico-chemical parameters as viscosity, hydration capacity and cooking time. A deeper understanding the genetic bases ruling variability across species/accessions could provide useful indicators for the screening of germplasm cooking properties (Santos et al., 2018).
Seed processability is related to the presence of components affecting seed protein digestibility. Such inhibitors, known as antinutrients or antinutritional factors can be removed using dedicated processing technologies, either traditional (cooking, milling, extrusion, germination, fermentation) or innovative technologies (e.g., high-pressure processing, ultrasound, irradiation, microwave). The seed response to these treatments is influenced by genetic factors, such as the qualitative and quantitative profiles of trypsin inhibitors, phytates, tannins, and lectins (Ohanenye et al., 2022). For orphan legumes, the genetic characterization of the most valuable traits and the subsequent breeders’ work are part of a long-term innovation program that must be supported by seed technologists. Once the breeding program has started, several generations are necessary to reach the status of certified seeds. Seed quality changes depending on the seed system, crop species/variety, and location of production.
3.3 Seed priming as a tool to mitigate the impact of climate change
Pre-sowing techniques (seed priming) are easily implementable tools, immediately available and applicable to crops, used to increase seed vigor (Pagano et al., 2023a). Seed imbibition is performed under controlled conditions (time, temperature, light/dark) with water, or in solutions containing priming agents, followed by dehydration. Controlled imbibition must be stopped before radicle protrusion occurs, otherwise, seeds will lose desiccation tolerance (Pagano et al., 2023a). Seed priming acts on specific components of the pre-germinative metabolism, a complex array of molecular pathways underlying the seed ability to scavenge the cytotoxic reactive oxygen species and repair DNA damage (Rajjou et al., 2012; Pagano et al., 2023a). The benefits provided by seed priming include improved germination performance and enhanced plant stress tolerance. Seed priming can be performed using water (hydropriming, matrix priming) or a wide range of priming agents that include salts (halopriming), PEG (polyethylene glycol; osmopriming), phytohormones (hormopriming), antioxidant and bioactive molecules (chemical priming), beneficial microorganisms (biopriming) and nanoparticles (nanopriming) (for a more comprehensive description, see review by Pagano et al., 2023a).
Seed priming improves drought tolerance in a wide range of cereal and legume crops (Saha et al., 2022) and several priming agents can effectively promote the seed antioxidant response and osmotic regulation, as mannose (Hameed and Iqbal, 2014), SiO2, KNO3, salycilic acid (Zheng et al., 2016; Tabassum et al., 2018; Ali et al., 2021), polyethylene glycol (Tounekti et al., 2020). Biopriming mediated by plant growth promoting bacteria can also mitigate drought stress (Pravisya and Jayaram, 2015; Shaffique et al., 2023; Srivastava et al., 2024). Soil salinity was reported to currently affect more than 30% of global irrigated land, being NaCl the most abundant salt in the soil. Seed germination is impaired under salt stress due to unfavorable osmotic potential and ion-mediated toxicity (Khan et al., 2022). Seed priming can alleviate the deleterious effects of salinity, as reported for rice seeds treated with CaCl2-based priming (Al-Tamimi et al., 2016). Seed priming applied to wheat using glycine betaine, vitamin B12, sodium nitroprusside, jasmonate, CaCl2, and KCl enhances chlorophyll content, stabilizes membranes and triggers antioxidant enzymes activity, mitigating salt stress (Sadeghi and Robati, 2015; Keshavarz and Moghadam, 2017).
Heat stress affects the plant physiological and metabolic processes due to protein denaturation, altered membrane integrity, and enzyme inactivation. Hydropriming, osmopriming with CaCl2, and hormopriming with salicylic acid were able to support germination of garden pea cultivars under heat stress, improving seed quality and plant development (Tamindžić et al., 2023). Priming with nitrate salts improves nitrate reductase and amylase activity, nitrogen, amino acid and chlorophyll content in leaves, increases proline and sugar content, antioxidant metabolism, under heat stress (Kumar et al., 2021; Tamindžić et al., 2023). Brassinosteroids used as seed priming agents significantly improved the growth and physiological parameters of Brassica juncea L. triggering enhanced antioxidant defense under heat stress (Neha et al., 2022). Cytokinins were used as priming agent to assess mitigation protocols against heat stress in wheat (Jaiswal et al., 2022). Field studies showed that plant survival under stress requires proteomic plasticity mediated by the ubiquitin-proteasome system. The effects of flooding stress on seed germination in rice (Oryza sativa L.) can be alleviated using seed priming with selenium (Se) (Hu et al., 2022). Selenium could limit malondialdehyde content, increase starch hydrolysis efficiency, enhance antioxidant activities, leading to improved biomass production. However, Se concentrations should be carefully selected in order to avoid the risk of seedling death during long-term flooding (Hu et al., 2022). These treatments also induce molecular mechanisms related to stress memory, accounting for trans-generational effects (Louis et al., 2023). Seed priming has an enormous potential to improve seed quality in fragile agroecosystems and fasten the route to food security, promoting drought tolerance and multiple stress resilience. Primed seeds are crucial for successful stand establishment and uniformity when farmers are forced to change the timing of sowing, due to environmental constrains (Singh et al., 2020; Devika et al., 2021; Liu et al., 2022). Improving seed quality support breeders’ work during the selection of ‘climate-smart’ varieties (Pagano et al., 2023a). However, the complexity of seed physiology, genetic background, and the influence of environment result in variability when the technique is applied. Current seed priming technologies would benefit from increased flexibility and efficacy, generated by integrating knowledge in seed biology/technology and agronomy of orphan legumes, moving from bench to field.
3.4 From bench to field: ‘on-farm’ seed priming
Seed priming is part of the Seed Treatment Market (estimated size in 2022: > 12.4 billion USD) (Global Market Insights, 2023). Seed companies own patented priming protocols, tailored for major crops but village farmers do not benefit from these resources. ‘On-farm’ seed priming, managed directly by farmers, differs from industrial strategies (Harris et al., 2005; Carrillo-Reche et al., 2018; Devika et al., 2021). In most cases, farmers know that soaking seeds in water before sowing improves plant establishment but they are often unaware of safe limits and drawbacks that might derive from inappropriate treatments. When dealing with fragile ecosystems, the choice of the most suitable priming approach should be carefully handled to maximize benefits. To fully exploit seed priming advantages, specific training with procedures tailored on seed lot/variety should be provided by means of participatory trials with local farmers, seed technologists and researchers (Harris et al., 2005) (Figure 2).
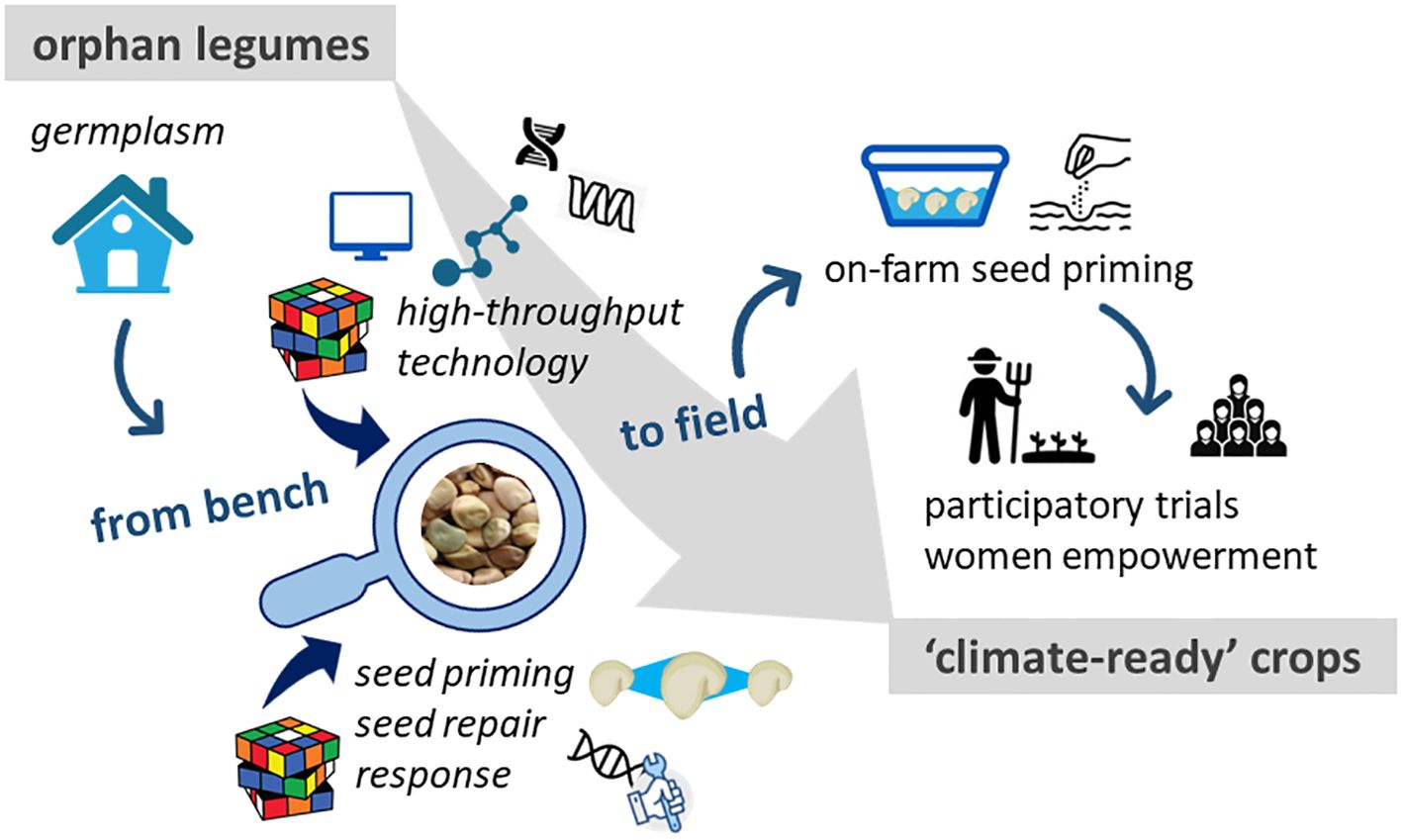
Figure 2 How can orphan legumes reach the status of ‘climate-ready crops’? To what extent can seed quality promote this transition? The ‘seed quality’ perspective, in terms of scientific, technological, and agronomic contributions is depicted. This avenue moves from bench to field as follows. The potential ‘climate-ready’ germplasm available deserves in-depth characterization to assess its real value. This requires the contribution of i) basic research on the molecular mechanisms (e.g. antioxidant players and DNA damage response) underlying seed quality, ii) applied research leading to advanced seed technology (e.g. more effective seed priming protocols), iii) knowledge/technology transfer to farmers and other stakeholders through training, participatory work, with focus on women empowerment.
4 Strategies to enhance seed quality in orphan legumes
Valorization of orphan legumes implies the intensification of their value chains, taking a series of actions ranging from geographic/climatic studies to socio-economic support measures. Multi-actor and cross-disciplinary research networks can bring innovation in orphan legumes production, building novel models in terms of profitability, resilience and environmental sustainability. A recent search carried out in PubMed using the keyword “orphan legume” (https://pubmed.ncbi.nlm.nih.gov/?term=orphan%20legume&timeline=expandedshowed) highlighted a stationary phase in the scientific production in the 1990-2004 period, followed by a progressive increase in the number of articles dealing with this topic, mostly focused on breeding whereas issues of related value chains and socioeconomic impact are still poor investigated. EU-funded, multidisciplinary projects such as CROPDIVA (https://www.cropdiva.eu/), ForEVA (https://www.ecpgr.org/working-groups/grain-legumes/foreva), LEGUMINOSE (https://www.leguminose.eu/), and LEGU-MED (https://www.era-learn.eu/network-information/networks/prima/section-2-call-2019-multi-topic/legumes-in-biodiversity-based-farming-systems-in-mediterranean-basin) are currently promoting the use of underutilized legume species, and the cross-talk between such established research networks will be a powerful driver of more extensive and effective knowledge transfer as well as deeper dialogue with end users.
The case study hereby described is the BENEFIT-Med (Boosting technologies of orphan legumes towards resilient farming systems; https://www.benefit-med.eu/) project, funded by the PRIMA Foundation (https://prima-med.org/), whose main goal is to develop an innovative, and manageable seed priming-based technology for the sustainable production of highly resilient orphan legumes in the Mediterranean Basin.
4.1 The BENEFIT-Med project: filling the gap of knowledge
Sustainable and resilient agroecosystems are made of agronomic, economic, social and ecological components. The synergic combination of technical and scientific expertise, multi-actors’ networks, and participatory work are required to design such complex structures (Westengen et al., 2023). Trigonella, grass pea, and forage pea are target species selected for valorization by the multidisciplinary BENEFIT-Med consortium, featuring the synergic interaction of eleven partners from eight countries along the Mediterranean Basin (Figure 3A). ‘On-farm’ seed priming protocols have been developed at lab scale, and tested under controlled conditions whereas the impact on agronomic performance is currently under evaluation in open-field trials, at different locations in Southern Europe and North Africa (Pagano et al., 2023b). The extreme climate conditions occurred in 2023 allowed to carry out ‘stress-tests’ to assess the benefits of seed priming but at the same time forced the asynchronous experimentation among partner countries. When the output data will be collected, the performance of accession-tailored protocols will be evaluated. The participatory work with local farmers, planned in the near future (2024) will play a crucial role to further optimize treatments and to train farmers about their rationale and effective use of the strategies developed within the BENEFIT-Med consortium. Such tailored seed technologies will be integrated into plant breeding studies to speed up the characterization of pre-breeding materials (Weissmann et al., 2023).
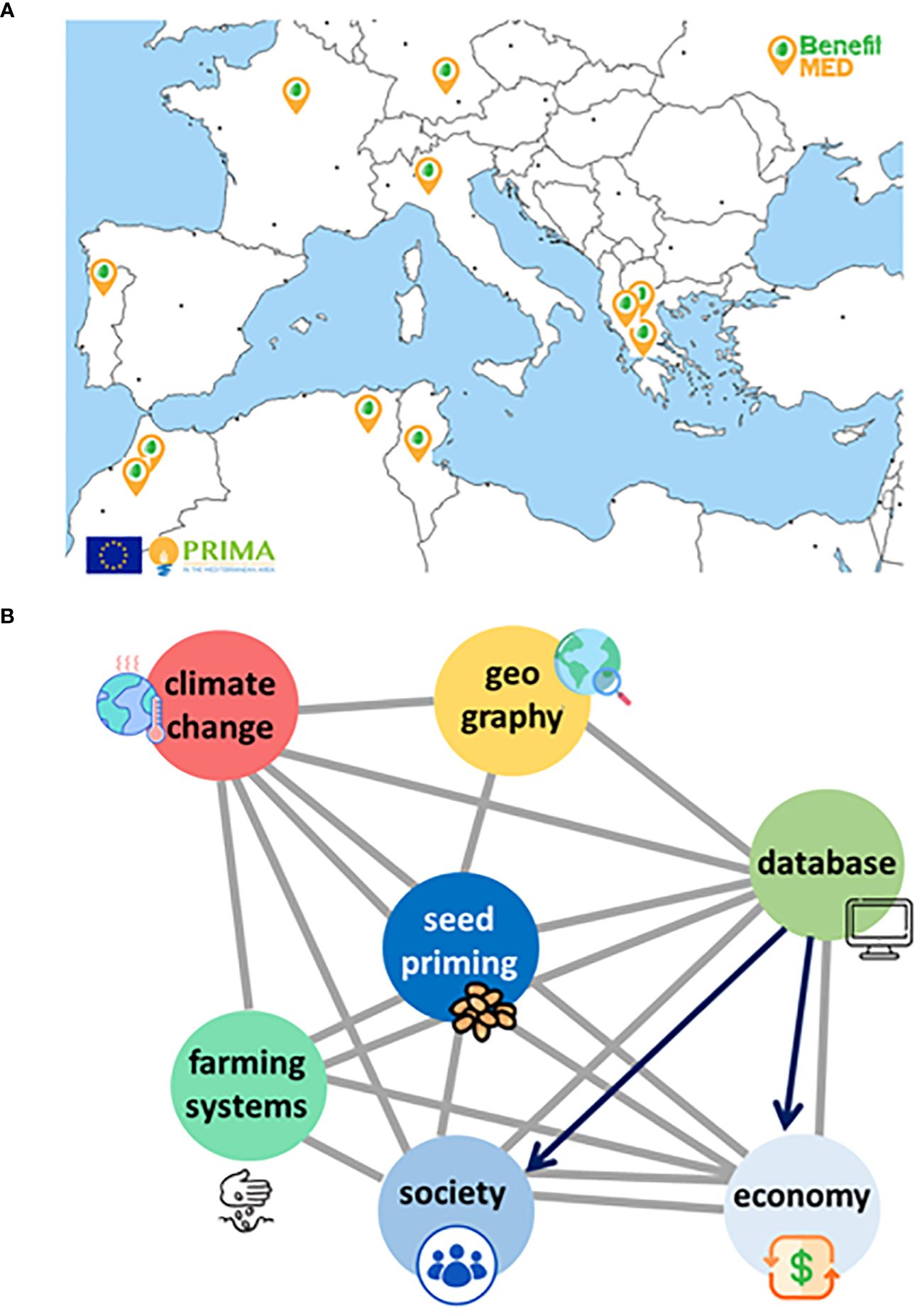
Figure 3 (A) The multidisciplinary BENEFIT-Med Consortium across the Mediterranean Basin includes eleven Partners distributed in eight countries: Germany, France, Portugal, Italy, and Greece from the EU side; Morocco, Algeria, and Tunisia from North Africa. (B) The ‘seed priming interactome’ represents the multidisciplinary effort and multitarget action of the BENEFIT-Med Consortium, envisaged as a mitigation strategy to boost orphan legumes adoption. Such strategy, relying on dedicated seed technology, requires cross-talk at different levels: i) climate change is targeted through a multidisciplinary approach (e.g. climate science, agronomy), ii) geography is included as mapping the climate vulnerable sites of the Mediterranean area is an essential step for future actions, iii) databases (BENEFIT-Med Hub-Med database) are crucial, as knowledge source and repository, iv) farming systems as the target of seed quality-based innovation, recipients of the benefits brought by orphan legumes, v) socio-economic development of vulnerable regions supported by dedicated actions (dissemination and training, involvement of policy-makers).
Exploring the tip of the ‘iceberg’ represented by the biodiversity of orphan legumes and transferring such biodiversity into their value chains are crucial. The knowledge of local communities should be preserved and valorized, as a carrier of resilient seed systems (Louwaars and Manicad, 2022; Westengen et al., 2023). It should be also underlined that mining for molecular mechanisms underlying seed quality is an urgent task, being an example of translational research that can accelerate the environmental adaptation of orphan legumes and related agroecosystems (Pagano et al., 2023a). Although seed priming is a main driver in the BENEFIT-Med model, this technology will lose its potential if not integrated into a much wider picture accounting for essential interactors, namely geography, climate, existing farming systems (Figure 3B). In BENEFIT-Med, climatologists are also investigating adaptation scenarios to unveil the impact of climate change, estimate site vulnerability and develop adaptation strategies able to support stakeholders and decision-makers (Cradock-Henry et al., 2021).
4.2 Vulnerability mapping: an interdisciplinary approach
The rural areas of the Mediterranean region are included in the list of most worldwide vulnerable targets (Ali et al., 2022). In order to support orphan legumes integration into Mediterranean food systems, a deeper understanding of consumption barriers and production constraints is required. Without these efforts, orphan legumes will face extinction because of ‘anthropogenic negligence’ (Talabi et al., 2022). A detailed mapping of vulnerable sites, reflecting the severity of barriers, should be developed. Vulnerability mapping requires an interdisciplinary approach embracing geography, environmental sciences, economics and political sciences, agronomy and climatology. Social aspects integrated with climate exposure metrics will allow to assess vulnerability drivers and adaptation indicators (de Sherbinin et al., 2023; Ngcamu, 2023; Goufa et al., 2024).
Climate-vulnerable sites are mapped based on socio-cultural barriers resulting from the different sensitivity and adaptation response of individuals and specific social/cultural groups. Economic barriers impair seed production and seed systems development, compromising market competitiveness. The challenge lies in promoting financial investments in order to strengthen the role of orphan legumes in the sustainable agrosystems of the Mediterranean region. Economic failure is the result of weak/missing supportive policies (political barriers), agriculture-related policies and policy frameworks (institutional barriers) should be revised to expand the use of orphan legumes e.g., by improving services to rural populations, adding these species in the national export strategy, or investing in the maintenance of traditional knowledge (Aktürk and Dastgerdi, 2021). Regular cross-talks with stakeholders are absolutely required to stimulate continuous participatory work and mitigate conflicting interests (Aktürk and Dastgerdi, 2021). Furthermore, awareness on climate-change impact and the potential of orphan legumes as climate-smart crops, as well as the nutritional value of traditional dishes, can be raised through education. Initiatives such as the CGIAR Generation Challenge Program (https://www.generationcp.org/) have contributed to build a new generation of breeders who vehicle cultural changes. Multiple socioeconomic factors (gender, class, ethnicity, age, disability) contribute to women vulnerability in rural areas (Abu-Rabia-Queder and Morris, 2018; FAO, 2023). Prolonged and high workload limits the access of young girls, in charge of water collection, to school. This compromises the possibility to bridge the gender gap in education, among the Sustainable Developmental Goals (SDGs) targets (UNICEF, 2020; Hamlet et al., 2021). Furthermore, climate-driven migration disrupts communities, charging women with heavier responsibilities and economic insecurity (Nwoke and Ibe, 2014; FAO, 2023).
Vulnerability mapping across the Mediterranean basin will face these problems, attempting to provide an in-depth description of the socioeconomic and climate scenarios essential for understanding the dynamics involving human/climatic factors and for the prediction of future expected impacts. The multidisciplinary character of the BENEFIT-Med team will lead to a dedicated vulnerability mapping of the Greater Mediterranean Region (GMR), as a starting point to develop more effective roadmaps focused on the mitigation effects resulting from the productive introduction of target species.
4.3 The Mediterranean region: a climate-change hot-spot
The Mediterranean region is considered a climate-change hot-spot of the 21st century (Giorgi, 2006). In this area, global warming rates are occurring at a greater pace and temperature extremes are expected to be reached (Lionello and Scarascia, 2018). Global air temperature increases by 1.5 and 2.0°C might lead to an increase of maximum air temperatures in the order of 2.2 and 3.0°C, respectively. Projected warming rates are connected to increased heat wave intensities and more frequent heat waves (Perkins-Kirkpatrick and Gibson, 2017), leading to reduced soil moisture and increased water stress for crops during all seasons, but with larger reduction rates during winter and spring, the main planting season for orphan legumes in the GMR (Samaniego et al., 2018). The evolution of new farming systems across the GMR, based on reliable climate information, is inevitable. One of the major challenges to overcome is the identification of the impact of climate change and variability on existing and new farming systems.
A suite of regional climate projections already exists from the Coordinated Regional Climate Downscaling Experiment (CORDEX) initiative (https://cordex.org/). Through global partnerships in regional climate modelling, various regional climate models (RCMs) are driven by large-scale general circulation models (GCMs) to derive regional-scale climate information under different emission scenarios. However, these climate projections are associated with large uncertainties as well as systematic biases, which should be reduced before applying them to climate change impact assessments (Laux et al., 2021). For the quantification of devastating extreme events in the GMR, such as the joint occurrence of severe drought and heat events, more sophisticated (multivariate) methods for bias correction (BC) are necessary (Adeyeri et al., 2023). Based on BC approaches, the co-variability of the most critical climate variables can be retained (Dieng et al., 2022; Olschewski et al., 2023), allowing the mapping of crop suitability and vulnerability to climate risk under different climate change scenarios. Crop suitability mapping is based on the growing thresholds for crops in relation to climate conditions. Within the BENEFIT-Med project, the concept of climatic crop suitability will be extended by edaphic, hydrological, and topographic conditions. Suitable threshold parameters for specific orphan legumes will be derived from field experimentations and finally upscaled to the GMR. Crop vulnerability, in this context, will be analyzed through the characterization of current and future patterns in soil temperature and soil moisture. In fact, soil temperature and soil moisture, besides other factors, affect seed germination and govern stand establishment (Elias et al., 2019).
4.4 Organization of the available knowledge
The current knowledge on orphan legumes is still fragmented, dispersed across repositories, from which data on few species can be retrieved. This is the case of Moringa oleifera L. and Vigna subterranea L., whose genomes are accessible at ORCAE-AOCC (Online Resource for Community Annotation of Eukaryotes - African Orphan Crops Consortium) (https://bioinformatics.psb.ugent.be/orcae/aocc/), a public genome annotation resource dedicated to underutilized crops, designed to facilitate stakeholders’ efforts (Yssel et al., 2019). Ideally, a globally accessible database that collects all the information on orphan legumes should be assembled and elaborated for working at multiple levels: as interface for interactions with public authorities, for dissemination/scientific/technical activities. A data model spanning the main components of the crop value chain in the food system has been suggested by Nizar et al. (2021). Such initiatives should be expanded to orphan legumes, merging the knowledge collected within past and current research projects.
5 Conclusions
This review highlights the drawbacks that must be overcome before orphan legumes could reach the state of ‘climate-ready crops’. The technical/social interfaces at which multiple actors play - still inadequate - roles are discussed. From the technical side, engagement of orphan legumes in climate-resilience breeding has become a priority and breeders can benefit from the current multidisciplinary approaches and high-throughput technologies to address the need of high-quality seeds. As for seed priming, an ideal roadmap should move from basic research to the translational and agricultural research domains, fishing new hallmarks of the seed response from the complex molecular pathways that showcase the pre-germinative metabolism before their validation in applied purposes. Seed systems should be reinforced by balancing the increased potential for seed quality and profitability offered by formal systems and an easier access to seeds should be ensured by community-based seed systems.
Author contributions
AlB: Writing – review & editing, Writing – original draft, Conceptualization. CC: Writing – review & editing. AM: Writing – review & editing. AP: Writing – review & editing. PL: Writing – review & editing. HM: Writing – review & editing. LR: Writing – review & editing. ET: Writing – review & editing. DC: Writing – review & editing. CK: Writing – review & editing. LG: Writing – review & editing. SG: Writing – review & editing. SM: Writing – review & editing. AE: Writing – review & editing. MH: Writing – review & editing. BK: Writing – review & editing. HE: Writing – review & editing. JN: Writing – review & editing. SA: Writing – review & editing. AzB: Writing – review & editing. MB: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by PRIMA (Partnership for Research and Innovation in the Mediterranean Area) Programme-H2020, Call Multi-Topic 2021, Research & Innovation Activities-RIA, Project “Boosting technologies of orphan legumes towards resilient farming systems in the Greater Mediterranean Region: from bench to open field (BENEFIT-Med) ID: 1726 (2022-2025). AP has been awarded a Post-Doctoral Fellowship in the frame of the BENEFIT-Med project.
Conflict of interest
Author CK was employed by the company Agroland S.A.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Cullis, C., Lawlor, D. W., Chimwamurombe, P., Bbebe, N., Kunert, K., Vorster, J. (2019). Development of marama bean, an orphan legume, as a crop. Food Energy Sec. 8, e00164. doi: 10.1002/fes3.164
Chapman, M. A., He, Y., Zhou, M. (2022). Beyond a reference genome: pangenomes and population genomics of underutilized and orphan crops for future food and nutrition security. New Phytol. 234, 1583–1597. doi: 10.1111/nph.18021
Abberton, M., Paliwal, R., Faloye, B., Marimagne, T., Moriam, A., Oyatomi, O. (2022). Indigenous african orphan legumes: potential for food and nutrition security in SSA. Front. Sust. Food Syst. 6. doi: 10.3389/fsufs.2022.708124
Abu-Rabia-Queder, S., Morris, A. (2018). Women in drylands: barriers and benefits for sustainable livelihoods. J. Arid Environ. 149, 1–3. doi: 10.1016/j.jaridenv.2017.11.009
Adeyeri, O. E., Zhou, W., Laux, P., Ndehedehe, C. E., Wang, X., Usman, M., et al. (2023). Multivariate drought monitoring, propagation, and projection using bias-corrected general circulation models. Earth's Future 11, e2022EF003303. doi: 10.1029/2022EF003303
Akohoue, F., Achigan-Dako, E. G., Sneller, C., Van Deynze, A., Sibiya, J. (2020). Genetic diversity, SNP-trait associations, and genomic selection accuracy in a west African collection of Kersting’s groundnut [Macrotyloma geocarpum (harms) Maréchal & Baudet]. PloS One 15, e0234769. doi: 10.1371/journal.pone.0234769
Aktürk, G., Dastgerdi, A. S. (2021). Cultural landscapes under the threat of climate change: a systematic study of barriers to resilience. Sustainability 13, 9974. doi: 10.3390/su13179974
Ali, L. G., Nulit, R., Ibrahim, M. H., Yien, C. Y. S. (2021). Efficacy of KNO3, SiO2 and SA priming for improving emergence, seedling growth and antioxidant enzymes of rice (Oryza sativa), under drought. Sci. Rep. 11, 1–11. doi: 10.1038/s41598-021-83434-3
Al-Tamimi, N., Brien, C., Oakey, H., Berger, B., Saade, S., Ho, Y. S., et al. (2016). Salinity tolerance loci revealed in rice using high-throughput non-invasive phenotyping. Nat. Commun. 7, 1–11. doi: 10.1038/ncomms13342
Bassett, A., Hooper, S., Cichy, K. (2021). Genetic variability of cooking time in dry beans (Phaseolus vulgaris L.) related to seed coat thickness and the cotyledon cell wall. Food Res. Int. 141, 109886. doi: 10.1016/j.foodres.2020.109886
Borelli, T., Hunter, D., Padulosi, S., Amaya, N., Meldrum, G., Moura de Oliveira Beltrame, D., et al. (2020). Local solutions for sustainable food systems: the contribution of orphan crops and wild edible species. Agronomy 10, 231. doi: 10.3390/agronomy10020231
Bourke, P. M., Evers, J. B., Bijma, P., van Apeldoorn, D. F., Smulders, M. J. M., Kuyper, T. W., et al. (2021). Breeding beyond monoculture: putting the “intercrop“ into crops. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.734167
Brǿnnvik, H., von Wettberg, E. J. (2019). Bird dispersal as a pre-adaptation for domestication in legumes: insights for neo-domestication. Front. Plant Sci. 10, 1293. doi: 10.3389/fpls.2019.01293
Čanak, P., Jeromela, A. M., Vujošević, B., Kiprovski, B., Mitrović, B., Alberghini, B., et al. (2020). Is drought stress tolerance affected by biotypes and seed size in the emerging oilseed crop Camelina? Agronomy 10, 1856. doi: 10.3390/agronomy10121856
Carrillo-Reche, J., Vallejo-Marin, M., Quilliam, R. S. (2018). Quantifying the potential of ‘on-farm’ seed priming to increase crop performance in developing countries. A meta-analysis. Agr. Sust. Dev. 38, 64. doi: 10.1007/s13593-018-0536-0
Christie, K., Natalie, R., Pierson, N. R., Lowry, D. B., Holeski, L. M. (2022). Local adaptation of seed and seedling traits along a natural aridity gradient may both predict and constrain adaptive responses to climate change. Am. J. Bot. 109, 1529–1544. doi: 10.1002/ajb2.16070
Cichy, K. A., Wiesinger, J. A., Berry, M., Nchimbi-Msolla, S., Fourie, D., Porch, T. G., et al. (2019). The role of genotype and production environment in determining the cooking time of dry beans (Phaseolus vulgaris L.). Legume Sci. 1, e13. doi: 10.1002/leg3.13
Cradock-Henry, N. A., Diprose, G., Frame, B. (2021). Towards local-parallel scenarios for climate change impacts, adaptation and vulnerability. Climate Risk Manage. 34, 100372. doi: 10.1016/j.crm.2021.100372
Cullis, C., Kunert, K. J. (2017). Unlocking the potential of orphan legumes. J. Exp. Bot. 68, 1895–1903. doi: 10.1093/jxb/erw437
Dawson, I. K., Powell, W., Hendre, P., Bančič, J., Hickey, J. M., Kindt, R., et al. (2019). The role of genetics in mainstreaming the production of new and orphan crops to diversify food systems and support human nutrition. New Phytol. 224, 37–54. doi: 10.1111/nph.15895
de Sherbinin, A., Bukvic, A., Rohat, G., Gall, M., McCusker, B., Preston, B., et al. (2023). Climate vulnerability mapping: a systematic review and future prospects. WIREs Climate Change 10, e600. doi: 10.1002/wcc.600
Devika, O. S., Singh, S., Sarkar, D., Barnwal, P., Suman, J., Rakshit, A. (2021). Seed priming: a potential supplement in integrated resource management under fragile intensive ecosystems. Front. Sust. Food Syst. 5. doi: 10.3389/fsufs.2021.654001
Dieng, D., Cannon, A. J., Laux, P., Hald, C., Adeyeri, O., Rahimi, J., et al. (2022). Multivariate bias-correction of high-resolution regional climate change simulations for West Africa: Performance and climate change implications. J Geophysical Res: Atmospheres 127, e2021JD034836. doi: 10.1029/2021JD034836
Elias, E. H., Flynn, R., Idowu, O. J., Reyes, J., Sanogo, S., Schutte, B. J., et al. (2019). Crop vulnerability to weather and climate risk: analysis of interacting systems and adaptation efficacy for sustainable crop production. Sustainability 11, 6619. doi: 10.3390/su11236619
Elolu, S., Byarugaba, R., Opiyo, A. M., Nakimbugwe, D., Mithöfer, D., Huyskens-Keil, S. (2023). Improving nutrition-sensitive value chains of African indigenous vegetables: current trends in postharvest management and processing. Front. Sust. Food Syst. 7. doi: 10.3389/fsufs.2023.1118021
Guo, M. W., Zhu, L., Li, H. Y., Liu, W. P., Wu, Z. N., Wang, C. H., et al. (2022)Mechanism of pod shattering in the forage legume Medicago ruthenica. Plant Physiol. Biochem. 185, 260–267. doi: 10.1016/j.plaphy.2022.06.013
FAO. (2019). The State of the World’s Biodiversity for Food and Agriculture. Eds. Bélanger, J., Pilling, D. (Rome: FAO Commission on Genetic Resources for Food and Agriculture Assessments), 572. Available at: http://www.fao.org/3/CA3129EN/CA3129EN.pdf. pp.
Fricke, E. C., Ordonez, A., Rogers, H. S., Svenning, J.-C. (2022). The effects of defaunation on plants’ capacity to track climate change. Science 375, 210–214. doi: 10.1126/science.abk3510
Gholami, A., Sharafi, S., Sharafi, A., Ghasemi, S. (2009). Germination of different seed size of pinto bean cultivars as affected by salinity and drought stress. J. Food Agric. Environ. 7, 555–558.
Global Market Insights (GMI). (2023). Seed treatment market - By method (chemical seed treatment, biological seed treatment, physical seed treatment) by crop (corn, wheat, barley, oats, soybean, canola, sunflower), by formulation, by equipment, by function and forecast 2023-2032. Report ID: GMI5673. Available online at: https://www.gminsights.com/industry-analysis/seed-treatment-market.
Global Panel on Agriculture and Food Systems for Nutrition. (2023). Food Systems and Diets: a Handbook of essential policies. Available online at: https://www.glopan.org/handbook/.
Gonçalves, L., Rubiales, D., Bronze, M. R., Vaz Patto, M. C. (2022). Grass Pea (Lathyrus sativus L.) - a sustainable and resilient answer to climate challenges. Agronomy 12, 1324. doi: 10.3390/agronomy12061324
Gonsalves, J. (2013). A new relevance and better prospects for wider uptake of social learning within CGIAR. CCAFS Working Paper no. 37 (Copenhagen, Denmark: CGIAR Research Program on Climate Change, Agriculture and Food Security (CCAFS). Available at: https://hdl.handle.net/10568/36033.
Goufa, M., Makeroufas, E., Gerakari, M., Sarri, E., Ragkos, A., Bebeli, P. J., et al. (2024). Understanding the potential to increase adoption of orphan crops: the case of Lathyrus spp. cultivation in Greece. Agronomy 14, 108. doi: 10.3390/agronomy14010108
Greveniotis, V., Bouloumpasi, E., Zotis, S., Korkovelos, A., Ipsilandis, C. G. (2022). Stability, the last frontier: forage yield dynamics of peas under two cultivation systems. Plants 11, 892. doi: 10.3390/plants11070892
Hameed, A., Iqbal, N. (2014). Chemo-priming with mannose, mannitol and H2O2 mitigate drought stress in wheat. Cereal Res. Commun. 42, 450–462. doi: 10.1556/CRC.2013.0066
Hamlet, L. C., Chakrabarti, S., Kaminsky, J. (2021). Reduced water collection time improves learning achievement among primary school children in India. Water Res. 203, 117527. doi: 10.1016/j.watres.2021.117527
Harris, D., Breese, W. A., Rao, J.V.D.K.K. (2005). The improvement of crop yield in marginal environments using “on-farm“ seed priming: nodulation, nitrogen fixation and disease resisitance. Aust. J. Agr. Res. 56, 1211–1218. doi: 10.1071/AR05079
Hendre, P. S., Muthemba, S., Kariba, R., Muchugi, A., Fu, Y., Chang, Y., et al. (2019). African Orphan Crops Consortium (AOCC): status of developing genomic resources for African orphan crops. Planta 250, 989–1003. doi: 10.1007/s00425-019-03156-9
Hertel, T., Elouafi, I., Tanticharoen, M., Ewert, F. (2021). Diversification for enhanced food systems. Nat. Food 2, 832–834. doi: 10.1038/s43016-021-00403-9
Hu, F. Q., Jiang, S. C., Wang, Z., Hu, K., Xie, Y. M., Zhou, L., et al. (2022). Seed priming with selenium: effects on germination, seedling growth, biochemical attributes, and grain yield in rice growing under flooding conditions. Plant Direct. 6, e378. doi: 10.1002/pld3.378
Huang, J., Ma, Q., Cai, Z., Xia, Q., Li, S., Jia, J., et al. (2020). Identification and mapping of stable QTLs for seed oil and protein content in soybean [Glycine max (L.) Merr.]. J. Agric. Food Chem. 68, 6448–6460. doi: 10.1021/acs.jafc.0c01271
ICRISAT - International Crops Research Institute for the Semi-Arid Tropics. (2008). Community-based seed production system (CBSP) in Nigeria: an opening for complementing national supply of quality seeds. Available online at: https://www.fao.org/plant-treaty/tools/toolbox-for-sustainable-use/details/en/c/1369802/.
IPCC (Intergovernmental Panel on Climate Change). (2022). Climate Change 2022: Mitigation of Climate Change (Geneva: Contribution of Working Group III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change). Available at: https://www.ipcc.ch/report/sixth-assessment-report-working-group-3/.
Jaiswal, P., Sahi, A. N., Barthakur, S. (2022). Cytokinin seed priming mediated induction of terminal heat stress tolerance and expression profiling of SKP1 transcripts, a component of ubiquitin proteasome system in bread wheat (Triticum aestivum L.): a transgenerational analysis. Plant Growth Regul. 98, 259–280. doi: 10.1007/s10725-022-00861-6
Joshi, T., Sehgal, H., Mandal, S. K., Joshi, M., Deepa, P. R., Sharma, P. K. (2023). Nutraceutical and flavor profiles in underutilized desert legumes of India: gene editing strategies towards sustainable food development. J. Plant Biochem. Biotech. 29, 1–8. doi: 10.1007/s13562-023-00836-2
Kansiime, M. K., Bundi, M., Nicodemus, J., Ochieng, J., Marandu, D., Njau, S. S., et al. (2021). Assessing sustainability factors of farmer seed production: a case of the good seed initiative project in Tanzania. Agr. Food Sec. 10, 1–10. doi: 10.1186/s40066-021-00289-7
Kaydan, D., Yagmur, M. (2008). Germination, seedling growth and relative water content of shoot in different seed sizes of triticale under osmotic stress of water and NaCl. Afr. J. Biotechnol. 7, 2862–2868. ISSN 1684–5315. doi: 10.5897/AJB08.512
Keshavarz, H., Moghadam, R. S. G. (2017). Seed priming with cobalamin (vitamin B12) provides significant protection against salinity stress in the common bean. Rhizosphere 3, 143–149. doi: 10.1016/j.rhisph.2017.04.010
Khan, M. O., Irfan, M., Muhammad, A., Ullah, I., Nawaz, S., Khalil, M. K., et al. (2022). A practical and economical strategy to mitigate salinity stress through seed priming. Front. Environ. Sci. 10, 991977. doi: 10.3389/fenvs.2022.991977
Kumar, V., Dwivedi, P., Kumar, P., Singh, B. N., Kumar Pandey, D., Kumar, V., et al. (2021). Mitigation of heat stress responses in crops using nitrate primed seeds. South Afr. J. Bot. 140, 25–36. doi: 10.1016/j.sajb.2021.03.024
Kumar, B., Singh, A. K., Bahuguna, R. N., Pareek, A., Singla-Pareek, S. L. (2023). Orphan crops: a genetic treasure trove for hunting stress tolerance genes. Food Energy Sec. 12, e436. doi: 10.1002/fes3.436
Laux, P., Rotter, R. P., Webber, H., Dieng, D., Rahimi, J., Wei, J., et al. (2021). To bias correct or not to bias correct? An agricultural impact modelers’ perspective on regional climate model data. Agr. For. Met. 304-305, 108406. doi: 10.1016/j.agrformet.2021.108406
Li, X., Siddique, K. H. M. (2018). Future Smart Food: rediscovering hidden treasures of neglected and underutilized species for Zero Hunger in Asia Vol. 242 (Bangkok: FAO). doi: 10.18356/23b5f7ab-en
Lionello, P., Scarascia, L. (2018). The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Change 18, 1481–1493. doi: 10.1007/s10113-018-1290-1
Liu, X., Quan, W., Bartels, D. (2022). Stress memory responses and seed priming correlate with drought tolerance in plants: an overview. Planta 255, 45. doi: 10.1007/s00425-022-03828-z
Liu, X., Zhang, H., Qin, S., Wang, Q., Yang, X., Wang, K. (2019). Optical fiber amplifier for quantitative and sensitive point-of-care testing of myoglobin and miRNA-141. Bios. Bioelectr. 129, 87–92. doi: 10.1016/j.bios.2018.12.056
Lo, S., Muñoz-Amatriaín, M., Hokin, S. A., Cisse, N., Roberts, P. A., Farmer, A. D., et al. (2019). A genome-wide association and meta-analysis reveal regions associated with seed size in cowpea [Vigna unguiculata (L.) Walp]. Theor. Appl. Gen. 132, 3079–3087. doi: 10.1007/s00122-019-03407-z
Louis, N., Dhankher, O. P., Puthur, J. T. (2023). Seed priming can enhance and retain stress tolerance in ensuing generations by inducing epigenetic changes and trans-generational memory. Physiol. Plant 175, e13881. doi: 10.1111/ppl.13881
Louwaars, N. P., Manicad, G. (2022). Seed systems resilience - An overview. Seeds 1, 340–356. doi: 10.3390/seeds1040028
Lv, M.-J., Wan, W., Yu, F., Meng, L.-S. (2019). New insight into the molecular mechanisms undelying seed size control under drought stress. J. Agr. Food Chem. 67, 9697–9704. doi: 10.1021/acs.jafc.9b02497
Martínez-López, M., Tinoco-Ojanguren, C., Martorell, C. (2020). Drought tolerance increases with seed size in a semiarid grassland from southern Mexico. Plant Ecol. 221, 989–1003. doi: 10.1007/s11258-020-01056-7
Mut, Z., Akay, H. (2010). Effect of seed size and drought stress on germination and seedling growth of naked oat (Avena sativa L.). Bulg. J. Agric. Sci. 16, 459–467.
Neha, T., Mohapatra, S., Sirhindi, G., Dogra, V. (2022). Seed priming with brassinolides improves growth and reinforces antioxidative defenses under normal and heat stress conditions in seedlings of Brassica juncea. Physiol. Plant 174, e13814. doi: 10.1111/ppl.13814
Ngcamu, B. S. (2023). Climate change effects on vulnerable populations in the Global South: a systematic review. Natural Hazards 118, 977–991. doi: 10.1007/s11069-023-06070-2
Nizar, N. M. M., Jahanshiri, E., Tharmandram, A. S., Salama, A., Sinin, S. S. M., Abdullah, N. J., et al. (2021). Underutilised crops database for supporting agricultural diversification. Comp. Electr. Agr. 180, 105920. doi: 10.1016/j.compag.2020.105920
Nwoke, E. A., Ibe, S. N. O. (2014). Climate change impact on the health of African women and adaptation strategies. Int. J. Global Warm Climate Change 1, 15–23.
Ogbole, O. O., Akin-Ajani, O. D., Ajala, T. O., Ogunniyi, Q. A., Fettke, J., Odeku, O. A. (2023). Nutritional and pharmacological potentials of orphan legumes: Subfamily faboideae. Heliyon 9, e15493. doi: 10.1016/j.heliyon.2023.e15493
Ohanenye, I. C., Ekezie, F.-G. C., Sarteshnizi, R. A., Boachie, R. T., Emenike, C. U., Sun, X., et al. (2022). Legume seed protein digestibility as influenced by traditional and emerging physical processing technologies. Foods 11, 2299. doi: 10.3390/foods11152299
Ojiewo, C. O., Rubyogo, J. C., Wesonga, J. M., Bishaw, Z., Gelalcha, S. W., Abang, M. M. (2018). Mainstreaming efficient legume seed systems in Eastern Africa: challenges, opportunities and contributions towards improved livelihoods (Rome: FAO), 72. ppLicense: CC BY-NC-SA 3.0 IGO.
Olschewski, P., Dieng, M. D. B., Moutahir, H., Böker, B., Haas, E., Kunstmann, H., et al. (2023). Amplified potential for vegetation stress under climate change-induced intensifying compound extreme events in the Greater Mediterranean Region. Nat. Hazards Earth Syst. Sci. Discuss. 24, 1099–1134. doi: 10.5194/nhess-2023-129
Oluwole, O. O., Olomitutu, O. E., Paliwal, R., Oyatomi, O. A., Abberton, M. T., Obembe, O. O. (2021). Preliminary assessment of the association between DArT-SEQ SNP and some nutritional traits in African yam bean. Trop. J. Nat. Prod. Res. 4, 877–879. doi: 10.26538/tjnpr/v1i4.5
Pagano, A., Bontà, V., Calvio, C., Macovei, A., Pinna, F., Bedotto, N., et al. (2023b). “From bench to field: scaling up seed priming technology on orphan legumes of the Mediterranean Area,” in 14th Biennal ISSS Conference - Challenges of Seed Science in a Changing Word, Book of Abstract. 168. Available at: https://isss2023.sciencesconf.org/.
Pagano, A., Macovei, A., Balestrazzi, A. (2023a). Molecular dynamics of seed priming at the crossroads between basic and applied research. Plant Cell Rep. 42, 657–688. doi: 10.1007/s00299-023-02988-w
Parker, T. A., Berny Mier y Teran, J. C., Palkovic, A., Jernestedt, J., Gepts, P. (2021). Pod indehiscence is a domestication and aridity resilience trait in common bean. New Phytol. 225, 558–570. doi: 10.1111/nph.16164
Perkins-Kirkpatrick, S. E., Gibson, P. B. (2017). Changes in regional heatwave characteristics as a function of increasing global temperature. Sci. Rep. 7, 12256. doi: 10.1038/s41598-017-12520-2
Popoola, J. O., Aworunse, O. S., Ojuederie, O. B., Adewale, B. D., Ajani, O. C., Oyatomi, O. A., et al. (2022). The exploitation of orphan legumes for food, income, and nutrition security in Sub-Saharan Africa. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.782140
Powers, S. E., Thavarajah, D. (2019). Checking agriculture’s pulse: field pea (Pisum Sativum L.), sustainability, and phosphorus use efficiency. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01489
Pravisya, P., Jayaram, K. M. (2015). Priming of Abelmoschus esculentus (L.) Moench (okra) seeds with liquid phosphobacterium: an approach to mitigate drought stress. Soc Trop. Plant Res. 2, 276–281. ISSN (E): 2349-1183.
Rajjou, L., Duval, M., Gallardo, K., Catusse, J., Bally, J., Job, C., et al. (2012). Seed germination and vigor. Annu. Rev. Plant Biol. 63, 507–533. doi: 10.1146/annurev-arplant-042811-05550
Ramya, K. R., Tripathi, K., Pandey, A., Barpete, S., Gore, P. G., Raina, A. P., et al. (2022). Rediscovering the potential of multifaceted orphan legume grasspea - a sustainable resource with high nutritional values. Front. Nutr. 8. doi: 10.3389/fnut.2021.826208
Roorkiwal, M., Pandey, S., Thavarajah, D., Hemalatha, R., Varshney, R. K. (2021). Molecular mechanisms and biochemical pathways for micronutrient acquisition and storage in legumes to support biofortification for nutritional security. Front. Plant Sci. 12, 682842. doi: 10.3389/fpls.2021.682842
Saastamoinen, M., Bocedi, G., Cote, J., Legrand, D., Guillaume, F., Wheat, et al. (2018). Genetics of dispersal. Biol. Rev. Cambridge Phil. Soc 93, 574–599. doi: 10.1111/brv.12356
Sadeghi, H., Robati, Z. (2015). Response of Cichorium intybus L. to eight seed priming methods under osmotic stress conditions. Biocatal. Agric. Biotechnol. 4, 443–448. doi: 10.1016/j.bcab.2015.08.003
Saha, D., Choyal, P., Mishra, U. N., Dey, P., Bose, B., Prathibha, M. D., et al. (2022). Drought stress responses and inducing tolerance by seed priming approach in plants. Plant Stress 4, 100066. doi: 10.1016/j.stress.2022.100066
Samaniego, L., Thober, S., Kumar, R., Wanders, N., Rakovec, O., Pan, M., et al. (2018). Anthropogenic warming exacerbates European soil moisture droughts. Nat. Climate Change 8, 421–426. doi: 10.1038/s41558-018-0138-5
Santos, C. S., Carbas, B., Castanho, A., Bronze, M. R., Serrano, C., Vasconcelos, M. W., et al. (2018). Relationship between seed traits and pasting and cooking behaviour in a pulse germplasm collection. Crop Past. Sci. 69, 892–903. doi: 10.1071/CP18205
Shaffique, S., Imran, M., Kang, S. M., Khan, M. A., Asaf, S., Kim, W. C., et al. (2023). Seed bio-priming of wheat with a novel bacterial strain to modulate drought stress in Daegu, South Korea. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1118941
Singh, V. K., Singh, R., Tripathi, S., Devi, R. S., Srivastava, P., Singh, P., et al. (2020). “Seed priming: state of the art and new perspectives in the era of climate change,” in Climate change and soil interactions. Eds. Prasad, M. N. P., Pietrzykowski, M. (Elsevier), 143–170.
Srivastava, S., Tyagi, R., Sharma, S. (2024). Seed biopriming as a promising approach for stress tolerance and enhancement of crop productivity: a review. J. Sci. Food Agr. 104, 1244–1257. doi: 10.1002/jsfa.13048
Tabassum, T., Farooq, M., Ahmad, R., Zohaib, A., Wahid, A., Shahid, M. (2018). Terminal drought and seed priming improves drought tolerance in wheat. Physiol. Mol. Biol. Plants 24, 845–856. doi: 10.1007/s12298-018-0547-y
Tadele, Z. (2019). Orphan crops: their importance and the urgency of improvement. Planta 250, 677–694. doi: 10.1007/s00425-019-03210-6
Talabi, A. O., Vikram, P., Thushar, S., Rahman, H., Ahmadzai, H., Nhamo, N., et al. (2022). Orphan crops: a best fit for dietary enrichment and diversification in highly deteriorated marginal environments. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.839704
Tamindžić, G., Azizbekian, S., Miljaković, D., Turan, J., Nikolić, Z., Ignjatov, M., et al. (2023). Comprehensive metal-based nanopriming for improving seed germination and initial growth of field pea (Pisum sativum L.). Agronomy. 13, 2932. doi: 10.3390/agronomy13122932
Tounekti, T., Mahdhi, M., Zarraq, A. F., Khemira, H. (2020). Priming improves germination and seed reserve utilization, growth, antioxidant responses and membrane stability at early seedling stage of Saudi sorghum varieties under drought stress. Notulae Bot. Horti Agrobot. Cluj-Napoca 48, 938–953. doi: 10.15835/nbha48211841
UNICEF. (2020). Gender and education (New York: United Nations). Available at: https://data.unicef.org/topic/gender/gender-disparities-in-education.
Upadhyaya, H. D., Bajaj, D., Das, S., Kumar, V., Gowda, C. L. L., Sharma, S., et al. (2016). Genetic dissection of seed-iron and zinc concentrations in chickpea. Sci. Rep. 6, 24050. doi: 10.1038/srep24050
Vaz Patto, M. C., Rubiales, D. (2014). Lathyrus diversity: available resources with relevance to crop improvement - L. sativus and L. cicera as case studies. Ann. Bot. 113, 895–908. doi: 10.1093/aob/mcu024
Vernooy, R., Rana, J., Otieno, G., Mbozi, H., Shrestha, P. (2022). Farmer-led seed production: community seed banks enter the national seed market. Seeds 1, 164–180. doi: 10.3390/seeds1030015
Visuvanathan, T., Than, L. T. L., Stanslas, J., Chew, S. Y., Vellasamy, S. (2022). Revisiting Trigonella foenum-graecum L.: pharmacology and therapeutic potentialities. PLants 11, 1450. doi: 10.3390/plants11111450
Weissmann, E. A., Yadav, R. N., Seth, R., Udaya Bhaskar, K. (2023). “Principles of variety maintenance for quality seed production,” in Seed Science and Technology. Eds. Dadlani, M., Yadava, D. K. (Springer, Singapore). doi: 10.1007/978-981-19-5888-5_8
Westengen, O. T., Dalle, S. P., Mulesa, T. H. (2023). Navigating toward resilient and inclusive seeds systems. Proc. Nat. Acad. Sci. U.S.A. 120, e2218777120. doi: 10.1073/pnas.2218777120
Xiong, W., Reynolds, M., Xu, Y. (2022). Climate change challenges plant breeding. Curr. Opin. Plant Biol. 70, 102308. doi: 10.1016/j.pbi.2022.102308
Xu, W., Wu, D. I., Yang, T., Sun, C., Wang, Z., Han, B., et al. (2021). Genomic insights into the origin, domestication and genetic basis of agronomic traits of castor bean. Genome Biol. 22, 113. doi: 10.1186/s13059-021-02333-y
Yssel, A. E., Kao, S.-M., Van de Peer, Y., Sterck, L. (2019). ORCAE-AOCC: a centralized portal for the annotation of African orphan crop Genomes. Genes 10, 950. doi: 10.3390/genes10120950
Zenda, T., Liu, S., Dong, A., Li, J., Wang, Y., Liu, X., et al. (2021). Omics-facilitated crop improvement for climate resilience and superior nutritive value. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.774994
Zheng, M., Tao, Y., Hussain, S., Jiang, Q., Peng, S. (2016). Seed priming in dry direct-seeded rice: consequences for emergence, seedling growth and associated metabolic events under drought stress. Plant Growth Regul. 78, 167–178. doi: 10.1007/s10725-015-0083-5
Keywords: climate change, Greater Mediterranean Region, resilience, food security, multi-actor approach, underutilized legumes, seed priming and seed system, vulnerability mapping
Citation: Balestrazzi A, Calvio C, Macovei A, Pagano A, Laux P, Moutahir H, Rajjou L, Tani E, Chachalis D, Katsis C, Ghaouti L, Gmouh S, Majid S, Elleuch A, Hanin M, Khemakhem B, El Abed H, Nunes J, Araújo S, Benhamrouche A and Bersi M (2024) Seed quality as a proxy of climate-ready orphan legumes: the need for a multidisciplinary and multi-actor vision. Front. Plant Sci. 15:1388866. doi: 10.3389/fpls.2024.1388866
Received: 20 February 2024; Accepted: 12 July 2024;
Published: 01 August 2024.
Edited by:
Leo Sabatino, University of Palermo, ItalyReviewed by:
Dr. Debasis Mitra, Graphic Era University, IndiaMohd. Kamran Khan, Selçuk University, Türkiye
Copyright © 2024 Balestrazzi, Calvio, Macovei, Pagano, Laux, Moutahir, Rajjou, Tani, Chachalis, Katsis, Ghaouti, Gmouh, Majid, Elleuch, Hanin, Khemakhem, El Abed, Nunes, Araújo, Benhamrouche and Bersi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alma Balestrazzi, YWxtYS5iYWxlc3RyYXp6aUB1bmlwdi5pdA==
†Present address: Susana Araújo, Mountain Environment and Ecosystem Management, MORE - Collaborative Laboratory Mountains of Research (MORE), Bragança, Portugal