Abstract
Karst river systems (KRSs) are essential to regional biogeochemical cycling and are defined by their distinct geomorphological and hydrological features. Understanding the factors controlling litter decomposition and associated nutrient fluxes within these systems is essential for predicting ecosystem responses to environmental changes. While extensive research is underway on litter decomposition and nutrient dynamics, studies on the interactions between species richness and elevation across seasons in KRSs remain limited. This study investigates the effects of elevation (800 m, 110 m, and 60 m) and seasonality (spring and autumn) on foliage decomposition and associated nutrient fluxes in KRSs along the Li River in China. We examined the foliage decomposition of three species: Taxodium distichum (Linn.) Rich., Taxodium ascendens Brongn., and Salix matsudana Koidz. It included seven litter treatments in single-species and mixed-species litter bags (3 species in all single-, 2- and 3-species combinations). A total of 336 decomposition bags were used to measure leaf mass loss and nutrient release during two six-month periods at three elevations in the study area in 2023. Results revealed that seasonal changes significantly influenced initial leaf elemental concentrations, with spring samples showing the most pronounced effects. Elevation had more effect on mass loss than litter treatments, with distinct nutrient release patterns observed across different elevations. Among single species, S. matsudana exhibited the highest mass loss and nutrient release at lower elevations during spring, while T. ascendens showed the lowest rates in autumn at high elevations. Mixed-species treatments displayed different decomposition patterns, with mass loss and nutrient release following the sequence: T. distichum × T. ascendens × S. matsudana < T. ascendens × S. matsudana < T. distichum × S. matsudana < T. distichum × T. ascendens. Nutrient release in mixed species showed significant interactions with elevation and season, displaying both positive and negative non-additive effects. Correlation analysis indicated stronger relationships between nutrient release and mass loss in S. matsudana than in T. distichum and T. ascendens. This study underscores the intricate interactions between biotic and abiotic factors in KRSs. It highlights the importance of considering elevation and seasonal dynamics in ecological restoration efforts in KRSs.
1 Introduction
Karst landscapes cover approximately 12% of the continents, and nearly 25% of the global population depends on them for water (Chen and Arif, 2025; Ford et al., 2019). Karst terrains are highly diverse because of nutrient influxes (Green et al., 2019; Jiao et al., 2024; Liu et al., 2021). This diversity can lead to excessive algae and aquatic plant growth downstream, resulting in environmental pollution (Oleksy et al., 2020). There is an increasing awareness that karst landscapes play a contributing role in controlling biogeochemical transformations in ecosystem functioning under seasonal changes (Chen et al., 2024a; Wang et al., 2025). Factors such as seasonal variations, elevation differences, and diverse vegetation—ranging from coniferous to broad-leaved species—interact with litter decomposition and associated nutrient fluxes at different rates (Geng et al., 2024; Ma et al., 2024). These interactions significantly influence nutrient dynamics in karst river systems (KRSs), also contributing to changes in litter nutrient cycling and decomposition processes in karst regions (Yang et al., 2020, Yang et al., 2022b). Despite their critical importance, the impacts of elevation and season on litter decomposition in KRSs remain underexplored (Chen et al., 2024a). This complexity emphasizes the need for focused studies to better capture variability in litter decomposition and associated nutrient fluxes (Mora et al., 2021). While recent research has advanced our understanding of how hydrological processes influence nutrient dynamics in KRSs (Dai et al., 2021; Wang et al., 2021; Yi et al., 2021), the complex ecological processes involved in regional biogeochemical cycling are still not fully understood.
Litter decomposition plays a vital role in nutrient cycling within karst ecosystems, while surface waters in karst landscapes drive significant biogeochemical transformations (Li et al., 2021). Nutrient dynamics in the KRS can also be influenced by vegetation characteristics than other factors (Ni and Li, 2020). Aquatic vegetation, including bottom-dwelling algae and floating plants, can thrive under these conditions, affecting nutrient storage and transformation (Li et al., 2023). However, our understanding of their specific impact on the nutrient flux budget along streams remains limited. Defense mechanisms shape leaf tissue structure among vegetation types (Salgado-Luarte et al., 2023). Leaf decomposition rates are strongly influenced by their chemical composition, particularly the concentrations of nitrogen (N), phosphorus (P), lignin, and cellulose, as well as the carbon-to-nitrogen (C/N) ratio (Bertrand et al., 2006; Talbot and Treseder, 2012). Specifically, high concentrations of lignin or tannins typically inhibit decomposition, while lower C/N ratios accelerate leaf decomposition (He et al., 2019). Generally, broad-leaved trees decompose quicker than pine trees since they have a lower N concentration and a higher C/N ratio (Prescott et al., 2017). Studies indicate that elevated N concentrations promote microbial growth on leaf surfaces (Wu et al., 2023), which accelerates reproduction and enhances decomposition.
Forest ecosystem studies have shown that interactions in mixed litter decomposition change at different stages (Putra et al., 2023; Rahman et al., 2023). In the early stages, nutrients are transferred, and microbes help the process along, leading to synergistic interactions (Lyu et al., 2019). As compounds that are difficult to break down accumulate over time, antagonistic effects are likely to become more prevalent (Liu et al., 2020; Yang et al., 2022a). Translocating nutrients from high-quality to low-quality leaves enhances nutrient subsidy and relieves nutrient limitations for microorganisms through the decomposition of low-quality leaf litter (Zeng et al., 2021). For instance, this facilitates the decomposition of mixed coniferous and broad-leaved litter, demonstrating its positive impact on the ecological environment overall (Niu et al., 2020). Studies have shown that decomposition rates in the KRS are similar to those found in forest ecosystems at different elevations and seasons (Barbosa et al., 2017; Geng et al., 2024). However, when researchers used litter mixtures for decomposition, they observed non-additive but modest effects at different sampling times (Liu et al., 2020). This interaction highlights the need for comprehensive assessments of both physical (e.g., hydrological pathways, soil structure) and chemical (e.g., nutrient concentrations, biogeochemical transformations) scales to clarify the unique hydrogeological characteristics of KRSs and their complex biogeochemical processes affect regional water resources and ecological environments (Li et al., 2021).
Improved hydrologic characterization of KRSs can provide valuable insights into nutrient movement under varying conditions (Dai et al., 2021). The season and elevation affect litter decomposition and nutrient dynamics (Chen et al., 2024b). Elevation affects environmental conditions such as temperature, humidity, and vegetation types, which influence decomposition. Higher elevations usually have lower temperatures, potentially slowing down decomposition (Yang et al., 2022b). During storm events, rapid water flow at higher elevations reduces water nutrient concentrations compared to low elevation areas where nutrients accumulate due to overland flow and macropore drainage, connecting high soil nutrient levels to the stream network (Ford et al., 2019; Wang et al., 2018). Karst channels are particularly effective at retaining nutrient fluxes, even during rapid flow periods (Oehler et al., 2018; Zhao et al., 2021). This effect is largely due to temporary sediment deposits that absorb or trap nutrients, such as P (Li et al., 2021; Ni and Li, 2020). Seasonality has a pronounced effect on nutrient concentrations in karst systems, with peaks in summer and troughs during autumn and winter, driven by environmental changes (Wang et al., 2018; Yue et al., 2020; Zhao et al., 2021). In addition, climate change may accelerate phase changes in freshwater because it increases terrestrial nutrient cycling (Arif et al., 2024; Naz et al., 2024). It is predicted that ecosystem degradation can increase river transport of P, N, and suspended particles but also reduce biodiversity under these circumstances of rapid human-induced climate change (Finlay et al., 2013; Nessel et al., 2021). Recent studies indicate that these thin, fine-particle layers can impact nutrient reduction and temporary storage in stream channels (Drummond et al., 2022; Field et al., 2023), yet the role of elevation changes in streambed composition in gently sloping areas remains poorly understood. Comprehending these complex interactions is essential for fully grasping biogeochemical processes in KRSs and their implications for species and regional water resources.
KRSs in Guangxi, China, exhibit large-scale limestone landscape types, distinct hydrologic processes, and strong decomposition and nutrient transfer processes (Wang et al., 2023). It is well known for its unique landscapes and multiple ecosystem services (Liao et al., 2018; Zhang et al., 2018). The area covers approximately 2.5 million km² (Jiang et al., 2018). This is connected by surface and groundwater systems, which carry nutrients and organic matter. The alkaline nature of karst waters affects microbial activity and decomposition rates, whereas fast-flow dynamics lead to resource recycling (Wang et al., 2019). The research concluded that these systems have rapid decomposition rates and nutrient dynamics (Chen et al., 2024a). It is necessary to conduct further research in these areas to clarify the modulating factors involved in litter decomposition and nutrient flux (Wang et al., 2023). It is essential for better environmental management and protection (Li et al., 2024b). This area contains many different types of broad-leaved trees and pine habitats. Notable species attracting ecotourism include Taxodium distichum (Linn.) Rich., Taxodium ascendens Brongn., and Salix matsudana Koidz. Therefore, we hypothesized: (1) Elevation and seasonal variations significantly influence foliage decomposition and nutrient release in KRSs. (2) Mixed-species litter decomposes faster than single-species treatments, and certain species combinations can produce non-additive effects, influenced by both species richness and elevation. (3) The relationship between nutrient release and mass loss varies across species, with some exhibiting a stronger correlation, particularly in response to seasonal changes. This study aims to assess the effects of litter mixing, elevation, and season on litter decomposition and associated nutrient concentrations at Karts River sites in China. The main objectives are to (1) investigate how leaf species decomposition varies across elevations in KRSs. (2) Examine the season change (spring and autumn) of nutrient dynamics within single and mixed species and determine if these patterns change with elevation. (3) Explore how elevation and season change interact to influence nutrient cycling in KRSs. Such insights could aid conservation efforts by providing a more profound understanding of ecosystem dynamics in karst landscapes.
2 Materials and methods
2.1 Study area
This study encompasses three experimental stations within KRSs along the Li River in Guilin, China (Supplementary Figure S1). It begins in the Mao’er Mountains (25°43’38.64”N, 110°24’14.40”E) in Xing’an county. It winds its way south for 164 km, passing through Guilin (25°16’12.00”N, 110°18’00.00”E) and Yangshuo (24°46’50.88”N, 110°29’47.76”E). The subtropical monsoon weather in this region significantly influences the hydrological dynamics and seasonal flow patterns of the river (Wang et al., 2023). This results in dramatic shifts from March to September. Floods tend to occur during heavy rain, such as in 2009, when river levels rose extensively (Li et al., 2019). The Li River basin flow rate ranges from close to 300 m³ per second in the wet season to less than 50 m³ per second in the dry season (Dong et al., 2024). This decreases significantly during the dry season, but water levels fluctuate widely throughout the year. The river conditions, including water characteristics, during the incubation periods are detailed in Supplementary Table S1. Yangshuo experiences water level fluctuations that mirror seasonal rainfall patterns and upstream water management practices. During the wet season, water levels can soar 2–5 m higher than during dry seasons, leading to flooding of low-lying areas. In contrast, levels drop dramatically during the dry season, exposing the riverbed. Water levels fluctuate from 1.5 m in the dry season to 6 m at peak flood levels, impacting navigation and other ecosystems. This contributes to the distinct ecological and hydrological features of the river. For example, it flows between 800 and 60 m above sea level (ASL) from the Mao’er Mountains to Yangshuo. The dramatic limestone peaks in this karst landscape create unique microclimates that influence local vegetation and soil types.
Human influences such as ecotourism activities and subtropical monsoon conditions also affect this ecosystem. These conditions are characterized by high humidity and significant annual rainfall (Zhang et al., 2018). The Li River basin boasts a diverse range of ecosystems, from subtropical forests to riparian plant communities. Its boundaries are home to bamboo, camphor trees, various shrubs, grasses, and other species (Chen et al., 2024a). Soil types and hydrological conditions are closely linked to vegetation patterns. Riverine vegetation along riverbanks plays a crucial role in maintaining ecological equilibrium, providing habitat for wildlife, and stabilizing riverbanks. However, deforestation, land conversion to agriculture, and tourism contribute to habitat loss and reduced biodiversity (Li et al., 2024b). Conservation efforts focus on protecting native vegetation, restoring degraded habitats through reforestation, and employing sustainable land management practices for maximum ecological benefit. In this study, flooding-tolerant tree species planted along the Li River were selected, including Taxodium distichum (Linn.) Rich., Taxodium ascendens Brongn., and Salix matsudana Koidz. These coniferous and broad-leaved tree species can survive in the wet season and germinate after the water level recedes but also increase the landscape value of the riverbank and enhance the beauty of the place.
2.2 Experimental design and sampling
In late March (spring) and early September (autumn) of 2023, T. distichum, T. ascendens, and S. matsudana plants with similar growth characteristics were selected from the riparian zone at three elevations of the Li River basin. Only well-developed, mature branches free of pests and diseases were chosen. Healthy, fresh leaves were collected from branches at different heights and orientations. Leaves from the same tree species were then thoroughly mixed and placed in plastic bags, which were subsequently numbered. After collection, a portable electronic balance was utilized to accurately measure 15.00 g of fresh leaves from the same tree species, as a single-species group, including two pines (T. distichum and T. ascendens) and one broadleaf species (S. matsudana). In pairwise mixed combinations, each species weighs 7.50 g in equal proportions mixed. This includes three foliage treatments: T. distichum × T. ascendens, T. distichum × S. matsudana, and T. ascendens × S. matsudana. When the three species are combined as T. distichum × T. ascendens × S. matsudana, each species weighs 5 grams. We packed each type of sample in the size of 20 cm × 20 cm, and the mesh size was 0.65 mm litter bags. This study implemented seven foliage treatments, with 168 samples collected at each sampling time, resulting in a total of 336 samples (7 litter treatments × 8 samples × 3 elevations × 2 seasons). This allowed for six-month decomposition periods under seven different foliage treatments. The study was based on naturally controlled experiments, seasonal changes (late March–spring and early September–autumn), and random flooding times at heights between 800 m ASL in the Mao’er Mountains (site 1), 110 m ASL in Guilin City (site 2), and 60 m ASL in Yangshuo (site 3).
Decomposition bags were placed on March 29, 2023, and September 2, 2023, respectively, as part of ecosystem restoration efforts. We cleared any debris from the sediment surface prior to placement. Moreover, we secured the decomposition bags with plastic nets, bamboo sticks, and wire to prevent animal damage and water-induced erosion (Liu et al., 2019). In addition, we collected 5 samples for each litter treatment and packed them into the marked envelope for the determination of initial leaf dry weight and leaf traits. We then transferred these bags back to the laboratory, labeled them, and dried them until they reached an even weight distribution. Before crushing and sieving, we recorded their initial dry weights to determine their elemental concentration.
After the 180-day decomposition duration, we stored the samples in an icebox and returned them to the laboratory for further processing. We carefully examined the sediment and debris attached to the decomposition bag, as well as the plant roots and benthic animals entering it. We then stored some samples in a -80°C refrigerator for further analysis (Slade and Riutta, 2012). Furthermore, we cleaned the decomposition residue with pure water and then dried it in an oven at 60°C to a constant weight. Samples collected at both intervals were subjected to two distinct seasonal treatments, including natural flooding and three elevational environments. We kept any changes to environmental variables natural; these were not considered within the scope of this study.
2.3 Laboratory analyses
The remaining dry mass of the sample was accurately weighed using a balance with a precision of 1/10,000 (Shanghai Shunyu Hengping Scientific Instrument Co., Ltd., FA2004) (Feijó Delgado et al., 2013). To determine the nutrient concentration in the leaves, the sample was finely ground with a ball mill (Leech MM400, Ball Mill, Germany) and sifted through a 65-mesh (0.3-mm) sieve. The sample was then dried again and weighed according to measurement index requirements (Fletcher et al., 2021). C and N concentrations were analyzed using a German ZNS-O element analyzer (ZNS-O-Vario EL cube; Heraeus Elementar, Hanau, Germany). Total P concentration was digested using a microwave digester (SpeedWave WS-4) and subsequently measured with an inductively coupled plasma spectrometer (ICP-AES).
2.4 Calculation methods for leaf-dry mass loss and nutrient release during mixed leaf species decomposition
The leaf dry mass loss (%) and nutrient release (%) were calculated using the following methods (Liu et al., 2017; Vogado et al., 2020; Yang et al., 2020):
ML is dry mass loss (%). NL is the nutrient release (%). M0 refers to the initial dry weight of the sample (g), C0 is its initial nutrient concentration of the sample (g·kg-1), Mt is the dry weight of the sample after decomposition time (g), and Ct is the nutrient concentration of the sample at decomposition time (g·kg-1) (Liu et al., 2017; Smart et al., 2017).
MLe is the expected mass loss of mixed species, and MLi is the mass loss of species i. Mi is the weight of mixed species i; ML0 is the actual mass loss of mixed species. At the same time, the expected mass loss of mixed leaves was calculated according to the average mass loss of individual species, and the mixing effect of leaf decomposition was calculated combined with its actual mass loss to explore whether the mass loss of mixed decomposition could be predicted by its component species. Non-additive effects in mixed leaf litter decomposition exist if the mixed decomposition rate and nutrient release are different from the expected results based on the decomposition rate of each individual leaf species. This effect can be manifested as a synergistic effect, that is, mixed decomposition is faster than single decomposition (p<0.05). It can also be manifested as an antagonistic effect, that is, mixed decomposition is slower than expected (Zhang et al., 2016).
2.5 Statistical analyses
To assess differences in initial element concentration and ratio across different litter treatments and elevations for each sample batch, we used an independent sample T-test. A one-way ANOVA was performed to analyze variations in initial element concentration and ratio at the litter treatment level between seasonal sample batches at different elevations (Pan et al., 2013). A two-way ANOVA was conducted to examine the effects of litter treatment, elevation, and their interactions on dry mass loss and nutrient release across seasonal periods (Wang et al., 2018). Additionally, a one-way ANOVA was used to determine whether mass loss and nutrient release in mixed-species treatments significantly differed from expected values. A mixed effect greater than 0 indicates a promoting effect, or a positive additive effect (p < 0.05), whereas a mixed effect less than 0 suggests an antagonistic effect (Zhang et al., 2016). Furthermore, a Pearson correlation test was conducted to analyze the relationship between nutrient release and mass loss of pine and broadleaf species in river ecosystems (Zheng et al., 2024). All data analyses were performed using SPSS 22.0 (IBM, Chicago, USA) and Origin Pro 2023 (OriginLab Corp., USA).
3 Results
3.1 Seasonal changes affect the initial element concentration of single and mixed litter treatments at different elevations
The initial element concentrations and their ratios varied significantly across litter treatments at all three elevations (Table 1). Notably, the standard error variance was relatively lower at the lower elevation (60 m) compared to the upper elevations (800 m) and middle elevations (110 m). Single-type foliage exhibited higher initial C and C/N ratios than mixtures. Conversely, mixed-type foliage demonstrated higher initial TN and N/P ratios compared to single-type foliage.
Table 1
| Litter treatment | Elevation (m) | TC (mg·g-1) | TN (mg·g-1) | TP (mg·g-1) | C/N | C/P | N/P |
|---|---|---|---|---|---|---|---|
| Td | 800 | 478.64 ± 1.74c | 16.30 ± 0.21c | 1.85 ± 0.09c | 29.39 ± 0.42a | 261.78 ± 13.51a | 8.90 ± 0.36a |
| 110 | 507.58 ± 1.29b | 20.80 ± 0.92a | 2.24 ± 0.08b | 24.57 ± 0.97b | 227.95 ± 8.05b | 9.40 ± 0.76a | |
| 60 | 519.42 ± 0.90a | 18.14 ± 0.28b | 2.57 ± 0.04a | 28.66 ± 0.43a | 202.40 ± 3.26b | 7.07 ± 0.21b | |
| Ta | 800 | 499.86 ± 1.87a | 14.22 ± 0.49c | 2.01 ± 0.13a | 35.31 ± 1.14a | 254.01 ± 17.72a | 7.19 ± 0.40a |
| 110 | 484.46 ± 0.78c | 15.32 ± 0.31b | 1.95 ± 0.05a | 31.68 ± 0.68b | 248.81 ± 6.51a | 7.87 ± 0.27a | |
| 60 | 494.50 ± 1.02b | 18.54 ± 0.21a | 2.01 ± 0.37a | 26.69 ± 0.31c | 203.77 ± 14.15b | 7.63 ± 0.51a | |
| Sm | 800 | 447.96 ± 1.31a | 31.20 ± 0.26a | 1.76 ± 0.03b | 14.36 ± 0.11a | 254.31 ± 4.44a | 17.71 ± 0.34a |
| 110 | 447.14 ± 1.34a | 30.88 ± 0.32a | 2.13 ± 0.09a | 14.49 ± 0.13a | 211.21 ± 9.37b | 14.59 ± 0.71b | |
| 60 | 446.66 ± 1.01a | 30.02 ± 0.57a | 2.00 ± 0.05a | 14.90 ± 0.26a | 224.32 ± 5.1b | 15.08 ± 0.44b | |
| Td × Ta | 800 | 498.69 ± 2.56b | 17.35 ± 0.38b | 2.02 ± 0.09b | 28.79 ± 0.57a | 248.54 ± 9.52a | 8.64 ± 0.32a |
| 110 | 493.31 ± 3.01b | 18.33 ± 0.3ab | 2.26 ± 0.05a | 26.94 ± 0.57b | 218.3 ± 4.61b | 8.11 ± 0.17a | |
| 60 | 507.91± 0.33a | 18.72 ± 0.27a | 2.31 ± 0.05a | 27.15 ± 0.39b | 220.45 ± 4.86b | 8.13 ± 0.29a | |
| Td × Sm | 800 | 465.4 ± 1.24c | 23.9 ± 0.54b | 1.86 ± 0.04b | 19.51 ± 0.46a | 250.63 ± 5.42a | 12.86 ± 0.3a |
| 110 | 474.4 ± 2.02b | 25.18 ± 0.18a | 1.98 ± 0.08b | 18.84 ± 0.1a | 241.31 ± 9.76a | 12.81 ± 0.56a | |
| 60 | 480.4 ± 0.51a | 24.74 ± 0.17ab | 2.2 ± 0.02a | 19.42 ± 0.15a | 218.72 ± 1.75b | 11.26 ± 0.13b | |
| Ta × Sm | 800 | 473.06 ± 4.16a | 26.31± 0.74a | 1.87 ± 0.04b | 18.03 ± 0.46a | 253 ± 7.14a | 14.07 ± 0.5a |
| 110 | 461.44 ± 2.54b | 25.99 ± 0.42a | 1.98 ± 0.09b | 17.77 ± 0.25a | 235.51 ± 11.23ab | 13.24 ± 0.54a | |
| 60 | 465.8 ± 1.02ab | 25.76 ± 0.49a | 2.2 ± 0.05a | 18.11 ± 0.38a | 212 ± 4.91b | 11.73 ± 0.42b | |
| Td × Ta × Sm | 800 | 476.03 ± 2.23b | 25.94 ± 0.33a | 2.1 ± 0.05a | 18.37 ± 0.31c | 226.97 ± 5.84a | 12.36 ± 0.27a |
| 110 | 481.43 ± 1.71ab | 24.89 ± 0.45a | 2.01 ± 0.03a | 19.37 ± 0.35b | 240.1 ± 3.46a | 12.42 ± 0.35a | |
| 60 | 483.79 ± 1.54a | 23.71 ± 0.25b | 2.07 ± 0.04a | 20.41 ± 0.19a | 233.63 ± 4.6a | 11.45 ± 0.23b |
The initial element concentration and ratio (mean ± S.E.) of each litter treatment at different altitudes in river ecosystems.
Means with different lowercase letters represent significant differences at p < 0.05 according to the independent sample t-test. The abbreviations in the table are Taxodium distichum (Td), Taxodium ascendens (Ta), Salix matsudana (Sm), total (T), carbon (C), nitrogen (N), and phosphorus (P). Sample size was n = 336.
The interactions among litter treatments, elevation, and the combination of litter treatments and elevation significantly influenced the initial element content of all leaf types in both spring and autumn. However, there were some exceptions: in spring, litter treatments did not significantly affect C/N ratio, and in autumn, elevation did not substantially influence C, N concentrations, C/N, and N/P ratios, nor did the interaction between leaf types and elevation contribute to C/P ratios (Table 2). The effects were remarkably pronounced in spring, as samples collected during this season exhibited more substantial responses in terms of litter decomposition and nutrient release patterns. Additionally, litter treatments were more influential when comparing elevation factors.
Table 2
| Index | Factor type | F value | |
|---|---|---|---|
| Spring | Autumn | ||
| Carbon (C) concentration (mg·g-1) | Litter treatment | 336.371*** | 73.982*** |
| Elevation | 39.415*** | 2.334 | |
| Litter treatment × Elevation | 23.546*** | 5.687*** | |
| Nitrogen (N) concentration (mg·g-1) | Litter treatment | 462.321*** | 82.968*** |
| Elevation | 8.127** | 1.118 | |
| Litter treatment × Elevation | 11.110*** | 7.693*** | |
| Phosphorus (P) concentration (mg·g-1) | Litter treatment | 3.113* | 28.529*** |
| Elevation | 12.214*** | 8.113* | |
| Litter treatment × Elevation | 2.216* | 2.457* | |
| C/N ratio | Litter treatment | 479.918*** | 122.657*** |
| Elevation | 18.217*** | 0.967 | |
| Litter treatment × Elevation | 17.368*** | 14.784*** | |
| C/P ratio | Litter treatment | 0.429 | 38.981*** |
| Elevation | 28.428*** | 6.314* | |
| Litter treatment × Elevation | 3.229** | 1.312 | |
| N/P ratio | Litter treatment | 161.362*** | 36.985*** |
| Elevation | 19.528*** | 4.718 | |
| Litter treatment × Elevation | 4.213*** | 4.319** | |
Effect of litter treatments and elevation on the initial element concentrations and ratios of leaf species during different seasons in river ecosystems.
According to the two-way ANOVA, the statistical differences are significant at p < 0.001***, P < 0.01**, and P < 0.05*. Sample size was n = 336.
The mass loss of all samples was notably affected by various litter treatments and elevations (Table 3). Elevation exerted a more pronounced influence compared to litter treatment. Furthermore, this effect was significantly more pronounced during spring, as indicated by higher effect values. In addition, when considering litter treatment and elevation together in both seasons, statistical tests did not yield significant results.
Table 3
| Index | Factor type | F value | |
|---|---|---|---|
| Spring | Autumn | ||
| Mass loss (%) | Litter treatments | 32.312*** | 16.867*** |
| Elevation | 34.986*** | 19.793*** | |
| Litter treatments × Elevation | 0.331 | 1.124 | |
Effect of litter treatments and elevation on mass-loss rates during different seasons in river ecosystems.
According to the two-way ANOVA, the *** denotes statistically significant differences at p < 0.001. Sample size was n = 336.
The release rates of C, N, and P in all samples were notably influenced by different litter treatments (Table 4). The patterns mirrored those of elevation factors, except for the N release rate in spring, which did not show significance. Likewise, the joint effect of litter treatments and elevation on these nutrient concentrations was largely significant, apart from C and N release in autumn.
Table 4
| Index | Factor type | F value | |
|---|---|---|---|
| Spring | Autumn | ||
| Carbon release rate (%) | Litter treatments | 6.127*** | 16.416*** |
| Elevation | 47.213*** | 45.129*** | |
| Litter treatments × Elevation | 5.301** | 2.018 | |
| Nitrogen release rate (%) | Litter treatments | 20.224*** | 39.108*** |
| Elevation | 2.332 | 30.238*** | |
| Litter treatments × Elevation | 3.225* | 2.027 | |
| Phosphorus release rate (%) | Litter treatments | 42.416*** | 11.109*** |
| Elevation | 64.968*** | 7.692** | |
| Litter treatments × Elevation | 10.318*** | 3.227* | |
Effect of litter treatments and elevation on nutrient release from leaves during different seasons in river ecosystems.
According to the two-way ANOVA, the statistical differences are significant at p < 0.001***, P < 0.01**, and P < 0.05*. Sample size was n = 336.
The release rates of C, N, and P from all litter treatments exhibit significant correlations with their rates of dry mass loss (Figure 1). Particularly, the correlation strengths for C and N were notably stronger in comparison to P. Furthermore, this correlation is more pronounced in broad-leaf tree species as opposed to pine trees. Additionally, these correlations are more prominent in mixed-species environments compared to monocultures.
Figure 1

Heat map of Pearson’s correlation of leaf species nutrient release and mass loss of litter treatments in river ecosystems. **The correlation is significant at the 0.01 level (two-tailed). Abbreviations in the figure are Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). Sample size was n = 336.
3.2 Mass loss of single and mixed litter treatment at different elevations under seasonal changes
The mass loss of single species was the largest (90.19 ± 0.77%) in S. matsudana during spring at 60 m elevation and the lowest (29.82 ± 10%) in T. ascendens during autumn at 800 m elevation (Figure 2). However, this pattern was inconsistent across different seasons and elevations. The mass loss ratios were substantially higher in spring compared to autumn. Moreover, the mass loss of mixed species was the largest (80.89 ± 4.88%) in T. distichum × T. ascendens × S. matsudana during spring at 110 m elevation and the lowest (35.91 ± 3.18%) in T. distichum × T. ascendens during autumn at 800 m elevation (Figure 2). This pattern followed a trend: T. distichum × T. ascendens × S. matsudana < T. ascendens × S. matsudana < T. distichum × S. matsudana < T. distichum × T. ascendens at different seasons and elevations. Furthermore, this pattern followed the distinctive trend of 60 m < 110 m < 800 m elevations. The mass loss was substantially higher in spring compared to autumn. Comparing the observed and expected mass loss of mixed species showed that non-additive effects of mass loss were more obvious in T. ascendens × S. matsudana during autumn at 110 m (9.65 ± 6.14%) and 800 m (8.81 ± 4.75%) elevations. These non-additive effects became negative in certain observations: T. ascendens × S. matsudana at 800 m (-0.22 ± 3.02%) and 60 m (-1.44 ± 2.37%) elevations, and T. distichum × T. ascendens × S. matsudana at 60 m elevation (-1.78 ± 1.15%) during spring. Additionally, standard error ranges were notable in T. ascendens × S. matsudana arrangements. However, the limited arrangements showed statistical differences, and most showed insignificant differences (Figure 2).
Figure 2

Grouped bars of mass loss of single litter treatments (a), mixed litter treatments (b), and comparison between the observed and expected value of mass loss of mixed litter treatments (c) in river ecosystems. Means with different lowercase letters represent significant differences at P < 0.05 according to the two-way ANOVA (a, b). According to the independent sample t-test, statistical differences are significant at p < 0.05* (c). Positive values indicate an observed value that exceeds its expected value, and vice versa (c). The vertical whisker represents the standard error (black color). Abbreviations in the figure are Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). The Y-axis displays three elevations: 800, 110, and 60 m. Sample size was n = 336.
The comparison of the non-additive effects on mass loss of mixed litter treatments of pine and broadleaf species for spring and autumn is shown in Figure 3. The coniferous mixture T. distichum × T. ascendens showed synergistic effects across all sampling seasons and elevations. The results indicated that the mixed T. distichum × T. ascendens of two conifer species promoted the decomposition of single leaf T. distichum and T. ascendens. The coniferous and broad-leaved mixture T. distichum × S. matsudana showed non-additive effects across all sampling seasons and elevations. Similarly, the coniferous and broad-leaved mixture T. ascendens × S. matsudana promoted the decomposition of the single leaf of T. ascendens, influencing the mass loss with sampling season and elevation. The antagonism of the three mixed T. distichum × T. ascendens × S. matsudana occurred, while a synergistic effect occurred in the decomposition across all sampling seasons and elevations.
Figure 3

The non-additive effects of mixed litter treatments on the mass loss of leaf species in spring (a-d) and autumn (e-h) in river ecosystems. Abbreviations used are Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). The abscissa represents the expected mass loss of mixed samples, and the ordinate is their measured mass loss. A circle represents 60 m elevation, a square represents 110 m elevation, and a triangle represents 800 m elevation. The hollow shape of a circle, triangle, or square indicates a significant non-addition. Sample size was n = 192.
3.3 Nutrient release of single and mixed litter treatments at different elevations under seasonal changes
All foliar types significantly affect the release of C, N, and P, while different litter treatments, elevations, and their interactions significantly affect their release rates (Figure 4). The C release rate of single species was the largest (94.60 ± 0.58%) in S. matsudana during spring at 110 m elevation and the lowest (46.01 ± 3.02%) in T. ascendens during autumn at 110 m elevation (Figure 4). The pattern of C release followed the same pattern across S. matsudana, T. ascendens, and T. distichum: 110 m < 60 m < 800 m. However, this pattern was inconsistent across different seasons and elevations. The C release rate was substantially higher in spring compared to autumn. The N release rate of single species followed the same pattern as C and was the largest (93.27 ± 0.86%) in S. matsudana during spring at 110 m elevation. It was lowest (15.30 ± 3.44%) in the same T. ascendens during autumn at 800 m elevation (Figure 4). The pattern of N release followed the same patterns across S. matsudana, T. ascendens, and T. distichum: 110 m < 60 m < 800 m. The N release rate was considerably higher in spring compared to autumn. The P release rate of single species followed the same pattern as that of C and N. It was the largest (87.62 ± 1.02%) in S. matsudana during spring at 60 m elevation. However, it was lowest (9.20 ± 4.47%) in T. distichum but not in T. ascendens during autumn at 800 m elevation (Figure 4). The pattern of P release followed the patterns of S. matsudana and T. distichum: 60 m < 110 m < 800 m, and this pattern was reversed for T. ascendens. Importantly, the P release rate was considerably higher in spring compared to autumn.
Figure 4

The bar graph groups the rates of element concentration loss for carbon (a), nitrogen (b), and phosphorus (c) in single litter treatment in river ecosystems. Means with different lowercase letters indicate significant differences at p < 0.05, according to the two-way ANOVA; abbreviations include Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). The vertical whisker represents the standard error (black color). The Y axis displays three elevations: 800, 110, and 60 m, respectively. Sample size was n = 144.
The C release rate of mixed species was the largest (90.20 ± 1.65%) in T. distichum × S. matsudana during spring at 110 m elevation and the lowest (52.56 ± 1.95%) in T. distichum × T. ascendens × S. matsudana during autumn at 800 m elevation (Figure 5). The pattern of C release followed the same pattern across all mixture combinations: 110 m < 60 m < 800 m. However, this pattern was inconsistent across different seasons and elevations. The C release rate was significantly higher in spring compared to autumn. The N release rate of mixed species followed the same pattern as C and was the largest (87.42 ± 1.64%) in T. distichum × S. matsudana during spring at 110 m elevation. It was lowest (38.56 ± 2.82%) in T. distichum × T. ascendens during autumn at 800 m elevation (Figure 5). The pattern of N release followed dissimilar patterns across mixture combinations: 110 m < 800 m < 60 m. The N release rate was considerably higher in spring than autumn. The P release rate of mixed species showed a distinctive pattern different from C and N. It was the largest (82.67 ± 0.55%) in T. distichum × T. ascendens × S. matsudana during spring at 60 m elevation and the lowest (42.87 ± 4.46%) in T. distichum × S. matsudana during autumn at 800 m elevation (Figure 5). A dissimilar pattern was observed for P release, with 110 m < 60 m < 800 m, differing for T. distichum × S. matsudana. Importantly, the P release rate was substantially higher in spring compared to autumn.
Figure 5
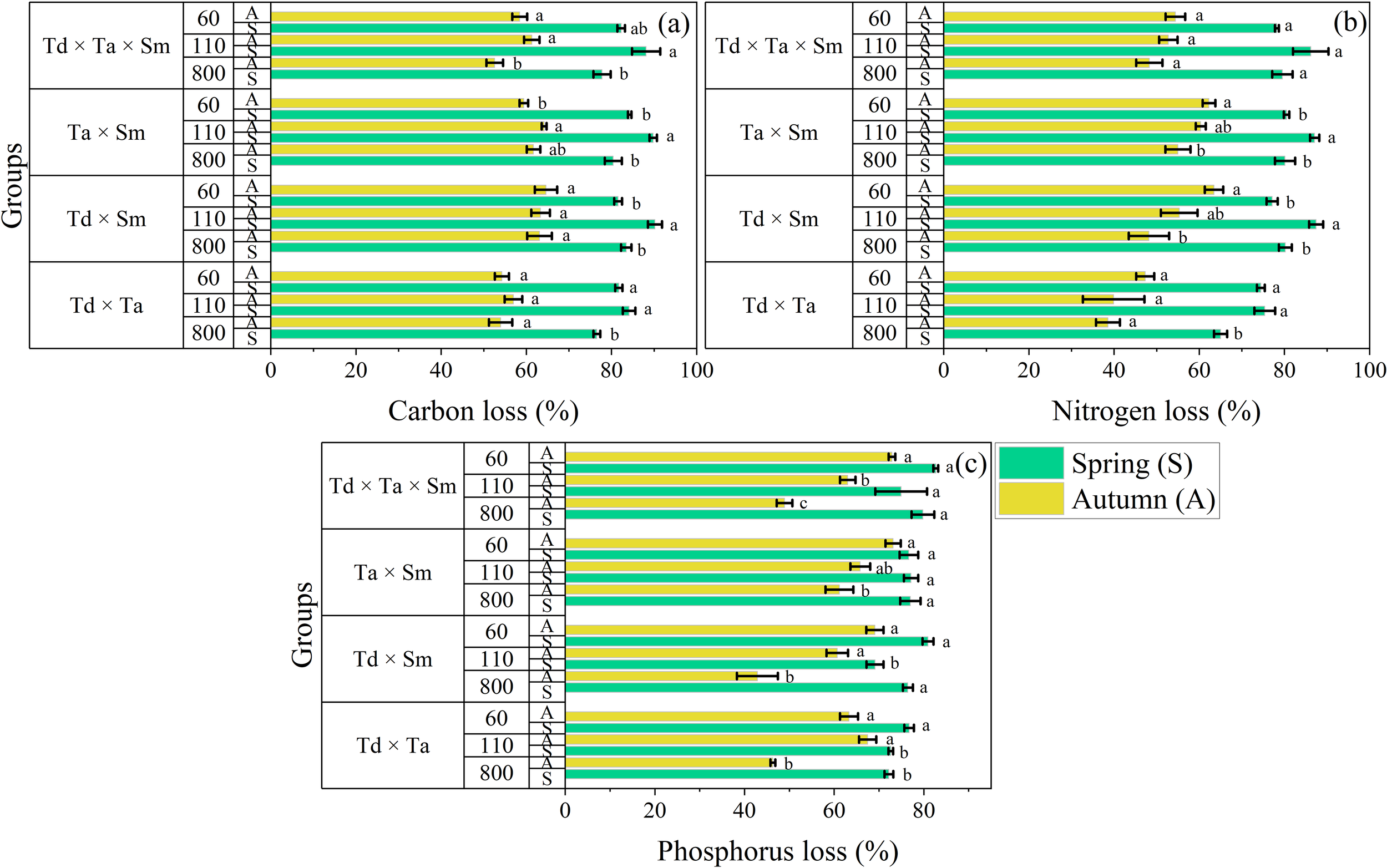
The bar graph groups the rates of element concentration loss for carbon (a), nitrogen (b), and phosphorus (c) in mixed litter treatment in river ecosystems. Means with different lowercase letters indicate significant differences at p < 0.05, according to the two-way ANOVA; abbreviations include Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). The vertical whisker represents the standard error (black color). The Y axis displays three elevations: 800, 110, and 60 m, respectively. Sample size was n = 192.
3.4 Effects of different treatment of leaf mixing on nutrient release at different elevations under seasonal changes
The non-additive effects of C, N, and P release from all litter treatments at different elevations and seasons are shown in Figure 6. The C release rate of each mixed type of elevation and season has a non-additive effect, but the effect varies with sampling season and elevation (Figure 6). The non-additive effects of C release in mixed species were the largest (10.12 ± 4.37%) in T. ascendens × S. matsudana during autumn at 110 m elevation and the lowest (-6.22 ± 1.07%) in T. distichum × S. matsudana during spring at 60 m elevation (Figure 6). The pattern of non-additive effects of C release followed dissimilar patterns across all mixture combinations and mostly had positive values. However, this pattern also had some negative values, mostly from T. distichum × T. ascendens × S. matsudana. Moreover, limited combinations showed statistical differences at p<0.05. The non-additive effects of N release in mixed species followed a different pattern from C and was the largest (22.81 ± 3.74%) in T. ascendens × S. matsudana during autumn at 800 m elevation. However, it was lowest (-4.26 ± 1.49%) in T. distichum × S. matsudana during spring at 60 m elevation (Figure 6). The pattern of non-additive effects of N release followed distinctive patterns across mixture combinations: T. ascendens × S. matsudana < T. distichum × T. ascendens × S. matsudana < T. distichum × T. ascendens < T. distichum × S. matsudana. This pattern mostly had positive values, and most combinations showed statistical differences at p<0.05. The non-additive effects of P release in mixed species followed a distinctive pattern different from C and N. It was the largest (23.79 ± 4.31%) in T. distichum × T. ascendens during autumn at 800 m elevation and the lowest (-5.06 ± 1.94%) in T. distichum × S. matsudana during spring at 110 m elevation (Figure 6). The pattern of non-additive effects of P release followed dissimilar patterns, mostly positive except for T. distichum × S. matsudana, which were overwhelmingly negative. Most combinations showed statistical differences at p<0.05.
Figure 6

The bar graph groups the non-additive effects of carbon (a), nitrogen (b), and phosphorus (c) on mixed litter treatment in river ecosystems. According to the independent sample t-test, statistical differences are significant at p < 0.05*, p < 0.01**, and p < 0.001***. Abbreviations in the figure are Taxodium distichum (Td), Taxodium ascendens (Ta), and Salix matsudana (Sm). The vertical whisker represents the standard error (black color). The Y-axis displays three elevations: 800, 110, and 60 m. Sample size was n = 192.
4 Discussion
This study investigates the influence of elevation, season, and litter treatments on litter decomposition and associated nutrient fluxes. In aquatic systems, litter decomposition is closely linked to both physical and chemical properties (Tables 1–4), as well as environmental factors such as elevation, temperature, humidity, and water characteristics—including pH, dissolved oxygen, water temperature, and flow rate (Chen et al., 2024a; Zheng et al., 2024). Biological factors, such as benthic invertebrates and microorganisms, also play a crucial role (Li et al., 2024a; Putra et al., 2023). Physical properties of leaves—like texture, thickness, hardness, wax presence, and cuticle thickness—significantly influence decomposition rates (Rahman et al., 2023). Research indicates that leaves with a softer texture and lower leathery degree decompose more rapidly than those with tougher textures and greater rigidity (Grootemaat et al., 2015; Hoeber et al., 2020), regardless of whether they are in flowing or still-water environments. Leaves with lower toughness break down faster. In our study, S. matsudana, which has softer and thinner leaves, exhibited higher decomposition rates than T. distichum and T. ascendens. The thicker cuticles of the latter species hinder fungal invasion and water absorption, slowing decomposition. This pattern was consistent across all sampling stages and elevations (Figure 2), likely due to differences in leaf tissue structure and resistance to physical fragmentation (Salgado-Luarte et al., 2023).
The chemical composition of leaves also plays a crucial role in decomposition rates, with factors such as N, P, lignin, cellulose, and the C/N ratio being key determinants (He et al., 2019; Talbot and Treseder, 2012). High lignin or tannin content generally slows down decomposition, whereas a low C/N ratio accelerates the process (Bertrand et al., 2006). In our study, S. matsudana, which had a higher N concentration and lower C/N ratio, exhibited significantly greater dry mass loss than T. distichum and T. ascendens, both with lower N concentrations. Higher N availability promotes microbial growth on leaf surfaces, leading to increased microbial reproduction and, consequently, faster decomposition (Wu et al., 2023). Previous research on mixed-leaf decomposition in forest ecosystems suggests that approximately 67% of mixed litter exhibit diverse effects during decomposition (Chapman and Koch, 2007; Zhang et al., 2017). This occurs because high-quality leaves (those with higher nutrient content) transfer nutrients to lower-quality leaves, enhancing their nutritional availability and mitigating nutrient limitations for microorganisms. Specifically, about 64.3% of coniferous and broadleaf mixtures accelerate decomposition, highlighting their overall positive effect (Qiu et al., 2023). Our findings, spanning different elevations and seasons, aligning with those observed in KRSs, demonstrating non-additive effects in mixed decomposition across all sampling periods, though the magnitude of these effects varies (Figure 3).
Among the litter treatments studied, T. distichum and T. ascendens have smaller, thicker cuticles and more rigid textures, which may slow the leaching rate during the initial stage of decomposition (Chen et al., 2024a). In contrast, mixed-leaf treatments, such as coniferous-broadleaf mixtures (T. distichum × S. matsudana and T. ascendens × S. matsudana), exhibited higher N concentrations and lower C/N ratios compared to coniferous mixtures (T. distichum × T. ascendens). High-quality litter, characterized by elevated N concentrations, attracts more decomposers, who may transfer nutrients to adjacent leaves through leaching or mycelial networks, accelerating decomposition (Lyu et al., 2019). Our results indicate that dry mass loss in coniferous-broadleaf mixtures was consistently greater than in coniferous mixtures across different seasons and elevations (Figure 4). This suggests that similar processes observed in forest ecosystems also occur in aquatic environments but with faster leaching and higher decomposition rates in flowing-water zones. Environmental factors and decomposition cycles further influence leaf decomposition (Putra et al., 2023). In KRSs, leaves at lower elevations experience prolonged flooding periods, which enhance decomposition. For example, during summer, the mass loss of all leaf types at 60 m was significantly higher than at other elevations. This confirms that extended flooding accelerates decomposition. Additionally, stable water temperatures, continuous water supply, and rapid nutrient cycling in aquatic environments support this process. Flowing water enhances decomposer activity, promoting bacterial and fungal colonization (Li et al., 2024a). Temperature also plays a crucial role, as higher water temperatures stimulate microbial activity, further accelerating decomposition (Calero Preciado et al., 2021).
The dynamics of element release during leaf decomposition in KRSs are influenced by both leaf characteristics and aquatic environmental factors (Zhang et al., 2017). Our results revealed distinct patterns of element release across different seasons and elevations, with notable species-specific variations (Figure 6). Decomposition-driven release of C, N, and P plays a crucial role in ecosystem biogeochemical cycles. Changes in hydrological patterns within KRSs directly impact leaf decomposition, influencing material circulation and interactions between terrestrial and aquatic ecosystems (Chen and Arif, 2025; Ford et al., 2019). Nutrient fluxes generally increase over time, particularly at lower elevations during the summer, driven by leaf matrix quality, environmental conditions, and the specific elements involved. Biodiversity plays a key role in nutrient dynamics during mixed-leaf decomposition, as species interactions create mixing effects. High-nutrient-content leaves supply nutrients to lower-nutrient-content leaves, slowing nutrient release and mitigating microbial nutrient limitations (Du et al., 2020). This nutrient redistribution facilitates decomposition and enhances nutrient release. Our study found that nutrient flux release rates were higher in coniferous-broadleaf mixtures than in coniferous mixtures, suggesting that mixed-leaf treatments create a more favorable environment for decomposers. This accelerated both decomposition and nutrient release, highlighting the ecological significance of species diversity in nutrient cycling.
Spring and summer rainstorms, which lead to increased discharge, are closely associated with rapid increases in the nutrient flux released from leaves (Oehler et al., 2018; Zhao et al., 2021). Summer conditions promote decomposition faster than autumn. As autumn temperatures drop and precipitation increases at higher elevations, leaf nutrient flux declines. Furthermore, significant rainfall events marking the end of the dry period can elevate surface water nutrient concentrations (Fong et al., 2020). In contrast, autumn brings only minor increases in river water nutrient concentrations within KRSs. Our measurements confirm previous studies indicate that large discharge events caused by heavy summer precipitation quickly flush away nutrients from surface soils, resulting in rapid nutrient export (Skidmore et al., 2023). Elevated discharge is linked to higher nutrient concentrations. This suggests that these short-lived flow events account for significant portions of the annual nutrient flux in KRSs. This finding aligns with other river studies (Oehler et al., 2018). Nutrient flux concentrations vary with elevation, and organic matter composition changes based on litter treatment and season. Lower nutrient flux was observed during the autumn season, while higher values occurred during the spring season at lower elevations. During spring, surface soils and vegetation contribute substantially to higher nutrient flux values. Numerous watershed studies have established that spring typically hosts an abundance of nutrient sources from surface soils and vegetation (Li et al., 2021; Ni and Li, 2020; Yi et al., 2021). This phenomenon contributes to greater nutrient flux than at any other time of year. Thus, we conclude that during the high-flow spring—and during some large summer rainstorms with rapid, shallow flows—KRSs along the Li River experience increased nutrient flux from surface soil and plant litter, which tends to be high in C but low in N. In contrast, in-situ production during low-flow conditions in autumn significantly reduces flux ratios.
Future research could focus on the long-term implications of climate change on KRSs, particularly the effects of changes in temperature and precipitation (Lian et al., 2020). Additionally, it is crucial to assess the impact of vegetation and land use changes on biogeochemical outcomes within KRSs (Chi and He, 2023). Added field data to hydrological and ecological models could help make better predictions about biogeochemical dynamics in KRSs, which would lead to better conservation and management plans (Johnson et al., 2021).
5 Conclusion
This study highlights the critical roles elevation and seasonality play in shaping leaf decomposition and nutrient release within the Li River karst systems. The results revealed significant interactions between litter treatments, elevation, and season. These interactions were notably higher in spring than in autumn. At lower elevations, decomposition and nutrient release were more pronounced, highlighting the significant influence of elevation on temperature and moisture variations. Distinct patterns emerged among litter treatments, with species like Salix matsudana exhibiting the highest decomposition rates at lower elevations in spring. In contrast, Taxodium ascendens showed the lowest rates at higher elevations in autumn. Mixed-leaf treatments displayed both synergistic and antagonistic effects, depending on species and environmental context. Importantly, non-additive effects on nutrient release and mass loss were observed in mixed-species treatments, reflecting complex interspecies interactions that fluctuate with elevation and season. These findings provide valuable insights into ecological restoration strategies in karst systems, advocating for approaches that consider both spatial and temporal dynamics. Effective management must integrate knowledge of how elevation and seasonality influence decomposition to preserve and enhance ecosystem health. Future research can explore the long-term impacts of these patterns and their interactions with broader climatic and anthropogenic changes, aiming to develop sustainable conservation and restoration practices for karst river systems.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
ZC: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MA: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. MT: Data curation, Formal analysis, Investigation, Methodology, Software, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the 2024 Ministry of Education Humanities and Social Sciences Research Project for Western and Border Regions (No. 24XJC630001); Research Projects of the Scientific Research Team of the School of Tourism Ecology and Environment at Guilin Tourism University in China (No. STKY-2024001); 2024 Annual Young and Middle-aged Teachers’ Basic Research Capability Improvement Project in Guangxi Universities (2024KY0847); the 2024 Guangxi Higher Education Undergraduate Teaching Reform Engineering Project (2024JGB406); and Environmental Protection and Ecological Restoration Cross-teaching Organization of Guilin Tourism University.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. During the preparation of this work, the author(s) used Wordtune and Grammarly in order to fix grammatical errors and enhance understanding of context and meaning. After using this tool or service, the author(s) reviewed and edited the content as needed and took full responsibility for the content of the publication.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1543011/full#supplementary-material
References
1
Arif M. Petrosillo I. Changxiao L. (2024). Effects of changing riparian topography on the decline of ecological indicators along the drawdown zones of long rivers in China. Front. For. Glob. Change7. doi: 10.3389/ffgc.2024.1293330
2
Barbosa R. I. de Castilho C. V. de Oliveira Perdiz R. Damasco G. Rodrigues R. Fearnside P. M. (2017). Decomposition rates of coarse woody debris in undisturbed Amazonian seasonally flooded and unflooded forests in the Rio Negro-Rio Branco Basin in Roraima, Brazil. For. Ecol. Manage.397, 1–9. doi: 10.1016/j.foreco.2017.04.026
3
Bertrand I. Chabbert B. Kurek B. Recous S. (2006). Can the biochemical features and histology of wheat residues explain their decomposition in soil? Plant Soil281, 291–307. doi: 10.1007/s11104-005-4628-7
4
Calero Preciado C. Boxall J. Soria-Carrasco V. Martínez S. Douterelo I. (2021). Implications of climate change: how does increased water temperature influence biofilm and water quality of chlorinated drinking water distribution systems? Front. Microbiol.12. doi: 10.3389/fmicb.2021.658927
5
Chapman S. K. Koch G. W. (2007). What type of diversity yields synergy during mixed litter decomposition in a natural forest ecosystem? Plant Soil299, 153–162. doi: 10.1007/s11104-007-9372-8
6
Chen Z. Arif M. (2025). The dynamics of bacterial communities during leaf decomposition of various species combinations in riparian forest buffers in China. For. Ecol. Manage.584, 122607. doi: 10.1016/j.foreco.2025.122607
7
Chen Z. Jiang Z. Li Q. Tan Y. Yan P. Arif M. (2024a). Examining the stoichiometry of C:N:P:K in the dynamics of foliar-litter-soil within dominant tree species across different altitudes in southern China. Glob. Ecol. Conserv.51, e02885. doi: 10.1016/j.gecco.2024.e02885
8
Chen Z. Li Q. Jiang Z. Yan P. Arif M. (2024b). Leaf functional traits of Daphniphyllum macropodum across different altitudes in Mao’er Mountain in Southern China. Front. For. Glob. Change7. doi: 10.3389/ffgc.2024.1359361
9
Chi Y. He C. (2023). Impact of land use change on the spatial and temporal evolution of ecosystem service values in south China karst areas. Forests14, 893. doi: 10.3390/f14050893
10
Dai Y. Lang Y. Wang T. Han X. Wang L. Zhong J. (2021). Modelling the sources and transport of ammonium nitrogen with the SPARROW model: a case study in a karst basin. J. Hydrol.592, 125763. doi: 10.1016/j.jhydrol.2020.125763
11
Dong Q. Y. Zhang H. Y. Xu J. X. Dai J. F. (2024). Precipitation and runoff mutation analysis in different water periods in Lijiang River Basin. Water Sav. Irrig.9), 74–81. doi: 10.12396/jsgg.2024093
12
Drummond J. D. Bernal S. Meredith W. Schumer R. Martí E. (2022). Stream hydrology controls the longitudinal bioreactive footprint of urban-sourced fine particles. Environ. Sci. Tech.56, 9083–9091. doi: 10.1021/acs.est.2c00876
13
Du N. Li W. Qiu L. Zhang Y. Wei X. Zhang X. (2020). Mass loss and nutrient release during the decomposition of sixteen types of plant litter with contrasting quality under three precipitation regimes. Ecol. Evol.10, 3367–3382. doi: 10.1002/ece3.6129
14
Feijó Delgado F. Cermak N. Hecht V. C. Son S. Li Y. Knudsen S. M. et al . (2013). Intracellular water exchange for measuring the dry mass, water mass and changes in chemical composition of living cells. PloS One8, e67590. doi: 10.1371/journal.pone.0067590
15
Field H. R. Sawyer A. H. Welch S. A. Benefiel R. K. Mathie D. M. Hood J. M. et al . (2023). Importance of dense aquatic vegetation in seasonal phosphate and particle transport in an agricultural headwater stream. Water Resour. Res.59, e2022WR033782. doi: 10.1029/2022WR033782
16
Finlay J. C. Small G. E. Sterner R. W. (2013). Human influences on nitrogen removal in lakes. Science342, 247–250. doi: 10.1126/science.1242575
17
Fletcher N. D. Manard B. T. Bostick D. A. Bostick W. D. Metzger S. C. Ticknor B. W. et al . (2021). Determination of phosphorus and sulfur in uranium ore concentrates by triple quadrupole inductively coupled plasma mass spectrometry. Talanta221, 121573. doi: 10.1016/j.talanta.2020.121573
18
Fong C. R. Gaynus C. J. Carpenter R. C. (2020). Extreme rainfall events pulse substantial nutrients and sediments from terrestrial to nearshore coastal communities: a case study from French Polynesia. Sci. Rep.10, 2955. doi: 10.1038/s41598-020-59807-5
19
Ford W. I. Husic A. Fogle A. Taraba J. (2019). Long-term assessment of nutrient flow pathway dynamics and in-stream fate in a temperate karst agroecosystem watershed. Hydrol. Process.33, 1610–1628. doi: 10.1002/hyp.13427
20
Geng Q. Arif M. Yin F. Chen Y. Gao J. Liu J. et al . (2024). Characteristics of forest understory herbaceous vegetation and its influencing factors in biodiversity hotspots in China. Ecol. Indic.167, 112634. doi: 10.1016/j.ecolind.2024.112634
21
Green S. M. Dungait J. A. J. Tu C. Buss H. L. Sanderson N. Hawkes S. J. et al . (2019). Soil functions and ecosystem services research in the Chinese karst Critical Zone. Chem. Geol.527, 119107. doi: 10.1016/j.chemgeo.2019.03.018
22
Grootemaat S. Wright I. J. van Bodegom P. M. Cornelissen J. H. C. Cornwell W. K. (2015). Burn or rot: leaf traits explain why flammability and decomposability are decoupled across species. Funct. Ecol.29, 1486–1497. doi: 10.1111/1365-2435.12449
23
He M. Zhao R. Tian Q. Huang L. Wang X. Liu F. (2019). Predominant effects of litter chemistry on lignin degradation in the early stage of leaf litter decomposition. Plant Soil442, 453–469. doi: 10.1007/s11104-019-04207-6
24
Hoeber S. Fransson P. Weih M. Manzoni S. (2020). Leaf litter quality coupled to Salix variety drives litter decomposition more than stand diversity or climate. Plant Soil453, 313–328. doi: 10.1007/s11104-020-04606-0
25
Jiang H. Liu W. Zhao T. Sun H. Xu Z. (2018). Water geochemistry of rivers draining karst-dominated regions, Guangxi province, South China: implications for chemical weathering and role of sulfuric acid. J. Asian Earth Sci.163, 152–162. doi: 10.1016/j.jseaes.2018.05.017
26
Jiao M. Yan J. Zhao Y. Xia T. Shen K. He Y. (2024). Dominance of rock exposure and soil depth in leaf trait networks outweighs soil quality in karst limestone and dolomite habitats. For. Ecosyst.11, 100220. doi: 10.1016/j.fecs.2024.100220
27
Johnson B. Gomez M. Munch S. B. (2021). Leveraging spatial information to forecast nonlinear ecological dynamics. Methods Ecol. Evol.12, 266–279. doi: 10.1111/2041-210X.13511
28
Li J. Arif M. Changxiao L. (2024a). Rare methanotrophs adapt to broader environmental gradients than abundant methanotrophs in the riparian zone of the Three Gorges Reservoir. Land Degrad. Dev.35, 249–263. doi: 10.1002/ldr.4913
29
Li J. Arif M. Dongdong D. Xin H. Qianwen G. Fan Y. et al . (2023). The diversity of plant communities in different habitats can lead to distinct methanotrophic communities. Rhizosphere26, 100690. doi: 10.1016/j.rhisph.2023.100690
30
Li C. Gao X. Wang W. Zhang X. Zhang X. Jiang C. et al . (2021). Hydro-biogeochemical processes of surface water leakage into groundwater in large scale karst water system: A case study at Jinci, northern China. J. Hydrol.596, 125691. doi: 10.1016/j.jhydrol.2020.125691
31
Li P. Guo Z. Furtado K. Chen H. Li J. Milton S. et al . (2019). Prediction of heavy precipitation in the eastern China flooding events of 2016: added value of convection-permitting simulations. Q. J. R. Meteor. Soc145, 3300–3319. doi: 10.1002/qj.3621
32
Li Q. Wang X. Chen Z. Arif M. (2024b). Assessing the conjunction of environmental sustainability and tourism development along Chinese waterways. Ecol. Indic.166, 112281. doi: 10.1016/j.ecolind.2024.112281
33
Lian J. Chen H. Wang F. Nie Y. Wang K. (2020). Separating the relative contributions of climate change and ecological restoration to runoff change in a mesoscale karst basin. Catena194, 104705. doi: 10.1016/j.catena.2020.104705
34
Liao C. Yue Y. Wang K. Fensholt R. Tong X. Brandt M. (2018). Ecological restoration enhances ecosystem health in the karst regions of southwest China. Ecol. Indic.90, 416–425. doi: 10.1016/j.ecolind.2018.03.036
35
Liu S. Fan R. Yang X. Zhang Z. Zhang X. Liang A. (2019). Decomposition of maize stover varies with maize type and stover management strategies: a microcosm study on a Black soil (Mollisol) in northeast China. J. Environ. Manage.234, 226–236. doi: 10.1016/j.jenvman.2019.01.008
36
Liu C. Huang Y. Wu F. Liu W. Ning Y. Huang Z. et al . (2021). Plant adaptability in karst regions. J. Plant Res.134, 889–906. doi: 10.1007/s10265-021-01330-3
37
Liu J. Liu X. Song Q. Compson Z. G. LeRoy C. J. Luan F. et al . (2020). Synergistic effects: a common theme in mixed-species litter decomposition. New Phytol.227, 757–765. doi: 10.1111/nph.16556
38
Liu G. Sun J. Tian K. Xiao D. Yuan X. (2017). Long-term responses of leaf litter decomposition to temperature, litter quality and litter mixing in plateau wetlands. Freshw. Biol.62, 178–190. doi: 10.1111/fwb.12860
39
Lyu M. Nie Y. Giardina C. P. Vadeboncoeur M. A. Ren Y. Fu Z. et al . (2019). Litter quality and site characteristics interact to affect the response of priming effect to temperature in subtropical forests. Funct. Ecol.33, 2226–2238. doi: 10.1111/1365-2435.13428
40
Ma L. Li L. Fang W. Dong Z. Liu Y. Wang C. et al . (2024). Genetic variability and population divergence of Rhododendron platypodum Diels in China in the context of conservation. Front. For. Glob. Change.7. doi: 10.3389/ffgc.2024.1320995
41
Mora J. L. Molina–Clerencia M. Girona–García A. Martí–Dalmau C. Badía–Villas D. (2021). Factors controlling the buildup of humus and particulate organic matter in European beech and Scots pine stands at their southernmost distribution limits (Moncayo Massif, Spain). Geoderma401, 115211. doi: 10.1016/j.geoderma.2021.115211
42
Naz F. Arif M. Xue T. Changxiao L. (2024). Seasonal dynamics of soil ecosystems in the riparian zones of the Three Gorges Reservoir, China. Glob. Ecol. Conserv.54, e03174. doi: 10.1016/j.gecco.2024.e03174
43
Nessel M. P. Konnovitch T. Romero G. Q. González A. L. (2021). Nitrogen and phosphorus enrichment cause declines in invertebrate populations: a global meta-analysis. Biol. Rev.96, 2617–2637. doi: 10.1111/brv.12771
44
Ni M. Li S. (2020). Spectroscopic indices trace spatiotemporal variability of dissolved organic matter in a river system with Karst characteristic. J. Hydrol.590, 125570. doi: 10.1016/j.jhydrol.2020.125570
45
Niu X. Y. Sun X. M. Chen D. S. Zhang S. G. (2020). Mixing litter from Larix kaempferi (lamb.) Carr. and broad-leaved trees enhances decomposition by different mechanisms in temperate and subtropical alpine regions of China. Plant Soil452, 43–60. doi: 10.1007/s11104-020-04527-y
46
Oehler T. Eiche E. Putra D. Adyasari D. Hennig H. Mallast U. et al . (2018). Seasonal variability of land-ocean groundwater nutrient fluxes from a tropical karstic region (southern Java, Indonesia). J. Hydrol.565, 662–671. doi: 10.1016/j.jhydrol.2018.08.077
47
Oleksy I. A. Baron J. S. Beck W. S. (2020). Nutrients and warming alter mountain lake benthic algal structure and function. Freshw. Sci.40, 88–102. doi: 10.1086/713068
48
Pan S. Liu C. Zhang W. Xu S. Wang N. Li Y. et al . (2013). The scaling relationships between leaf mass and leaf area of vascular plant species change with altitude. PloS One8, e76872. doi: 10.1371/journal.pone.0076872
49
Prescott C. E. Corrao K. Reid A. M. Zukswert J. M. Addo-Danso S. D. (2017). Changes in mass, carbon, nitrogen, and phosphorus in logs decomposing for 30 years in three Rocky Mountain coniferous forests. Can. Can. J. For. Res.47, 1418–1423. doi: 10.1139/cjfr-2017-0001
50
Putra P. S. Achmad A. Yamada T. Ngakan P. O. (2023). Seasonal decomposition rates of broadleaf and conifer wood litter in far eastern tropical forest communities. Int. J. For. Res.2023, 9677809. doi: 10.1155/2023/9677809
51
Qiu L. Xiao T. Bai T. Mo X. Huang J. Deng W. et al . (2023). Seasonal dynamics and influencing factors of litterfall production and carbon input in typical forest community types in Lushan Mountain, China. Forests14, 341. doi: 10.3390/f14020341
52
Rahman N. E. B. Smith S. W. Lam W. N. Chong K. Y. Chua M. S. E. Teo P. Y. et al . (2023). Leaf decomposition and flammability are largely decoupled across species in a tropical swamp forest despite sharing some predictive leaf functional traits. New Phytol.238, 598–611. doi: 10.1111/nph.18742
53
Salgado-Luarte C. González-Teuber M. Madriaza K. Gianoli E. (2023). Trade-off between plant resistance and tolerance to herbivory: mechanical defenses outweigh chemical defenses. Ecology104, e3860. doi: 10.1002/ecy.3860
54
Skidmore M. Andarge T. Foltz J. (2023). The impact of extreme precipitation on nutrient runoff. J. Agric. Appl. Econ. Assoc.2, 769–785. doi: 10.1002/jaa2.90
55
Slade E. M. Riutta T. (2012). Interacting effects of leaf litter species and macrofauna on decomposition in different litter environments. Basic Appl. Ecol.13, 423–431. doi: 10.1016/j.baae.2012.06.008
56
Smart S. M. Glanville H. C. Blanes M. Mercado L. M. Emmett B. A. Jones D. L. et al . (2017). Leaf dry matter content is better at predicting above-ground net primary production than specific leaf area. Funct. Ecol.31, 1336–1344. doi: 10.1111/1365-2435.12832
57
Talbot J. M. Treseder K. K. (2012). Interactions among lignin, cellulose, and nitrogen drive litter chemistry–decay relationships. Ecology93, 345–354. doi: 10.1890/11-0843.1
58
Vogado N. O. Winter K. Ubierna N. Farquhar G. D. Cernusak L. A. (2020). Directional change in leaf dry matter δ 13C during leaf development is widespread in C3 plants. Ann. Bot.126, 981–990. doi: 10.1093/aob/mcaa114
59
Wang S. Liping Y. Arif M. (2025). Evolutionary analysis of ecological-production-living space-carrying capacity in tourism-centric traditional villages in Guangxi, China. J. Environ. Manage.375, 124182. doi: 10.1016/j.jenvman.2025.124182
60
Wang A. Wang X. Tognetti R. Lei J.-P. Pan H.-L. Liu X.-L. et al . (2018). Elevation alters carbon and nutrient concentrations and stoichiometry in Quercus aquifolioides in southwestern China. Sci. Total Environ.622, 1463–1475. doi: 10.1016/j.scitotenv.2017.12.070
61
Wang L. Wu X. Guo J. Zhou J. He L. (2023). Spatial–temporal pattern of vegetation carbon sequestration and its response to rocky desertification control measures in a karst area, in Guangxi Province, China. Land Degrad. Dev.34, 665–681. doi: 10.1002/ldr.4485
62
Wang K. Zhang C. Chen H. Yue Y. Zhang W. Zhang M. et al . (2019). Karst landscapes of China: patterns, ecosystem processes and services. Landsc. Ecol.34, 2743–2763. doi: 10.1007/s10980-019-00912-w
63
Wang N. Zhang M. Zhao N. Feng F. Zhao M. (2021). Season-dependence of soil extracellular enzyme activities in a Pinus koraiensis forest on Changbai Mountain. J. Fores. Res.32, 1713–1722. doi: 10.1007/s11676-020-01213-8
64
Wu D. Ma X. Meng Y. Cai R. Zhang X. Liu L. et al . (2023). Impact of nitrogen application and crop stage on epiphytic microbial communities on silage maize leaf surfaces. PeerJ11, e16386. doi: 10.7717/peerj.16386
65
Yang Y. Berhe A. A. Barnes M. E. Moreland K. C. Tian Z. Kelly A. E. et al . (2022b). Climate warming alters nutrient storage in seasonally dry forests: insights from a 2,300 m elevation gradient. Global Biogeochem. Cycles36, e2022GB007429. doi: 10.1029/2022GB007429
66
Yang B. He Y. Chen W. (2020). A simple method for estimation of leaf dry matter content in fresh leaves using leaf scattering albedo. Glob. Ecol. Conserv.23, e01201. doi: 10.1016/j.gecco.2020.e01201
67
Yang K. Zhu J. Zhang W. Zhang Q. Lu D. Zhang Y. et al . (2022a). Litter decomposition and nutrient release from monospecific and mixed litters: comparisons of litter quality, fauna and decomposition site effects. J. Ecol.110, 1673–1686. doi: 10.1111/1365-2745.13902
68
Yi Y. Zhong J. Bao H. Mostofa K. M. G. Xu S. Xiao H.-Y. et al . (2021). The impacts of reservoirs on the sources and transport of riverine organic carbon in the karst area: a multi-tracer study. Water Res.194, 116933. doi: 10.1016/j.watres.2021.116933
69
Yue F.-J. Li S.-L. Waldron S. Wang Z.-J. Oliver D. M. Chen X. et al . (2020). Rainfall and conduit drainage combine to accelerate nitrate loss from a karst agroecosystem: Insights from sta ble isotope tracing and high-frequency nitrate sensing. Water Res.186, 116388. doi: 10.1016/j.watres.2020.116388
70
Zeng Q. Chen Z. Tan W. (2021). Plant litter quality regulates soil eco-enzymatic stoichiometry and microbial nutrient limitation in a citrus orchard. Plant Soil466, 179–191. doi: 10.1007/s11104-021-05025-5
71
Zhang X. Liu Z. Zhu B. Bing Y. Luc N. T. Du L. et al . (2016). Impacts of mixed litter decomposition from Robinia pseudoacacia and other tree species on C loss and nutrient release in the Loess Plateau of China. J. For. Res.27, 525–532. doi: 10.1007/s11676-015-0175-0
72
Zhang X.-H. Sun X.-X. Mao R. (2017). Effects of litter evenness, nitrogen enrichment and temperature on short-term litter decomposition in freshwater marshes of Northeast China. Wetlands37, 145–152. doi: 10.1007/s13157-016-0855-3
73
Zhang M. Wang K. Liu H. Zhang C. Yue Y. Qi X. (2018). Effect of ecological engineering projects on ecosystem services in a karst region: a case study of northwest Guangxi, China. J. Clean. Prod.183, 831–842. doi: 10.1016/j.jclepro.2018.02.102
74
Zhao H. Jiang Y. Xiao Q. Zhang C. Behzad H. M. (2021). Coupled carbon-nitrogen cycling controls the transformation of dissolved inorganic carbon into dissolved organic carbon in karst aquatic systems. J. Hydrol.592, 125764. doi: 10.1016/j.jhydrol.2020.125764
75
Zheng J. Arif M. Cao W. Li C. (2024). Dam inundation modulates the effect of plant diversity on soil multifunctionality in the riparian zone of the Three Gorges Reservoir. Land Degrad. Dev.35, 5584–5595. doi: 10.1002/ldr.5317
Summary
Keywords
single species, mixed species, leaf decomposition, river ecosystems, karst ecosystems, nutrients
Citation
Chen Z, Arif M and Tian M (2025) Elevation and seasonality modulate the leaf decomposition rates and nutrient flux of diverse species and species richness in karst river systems across China. Front. Plant Sci. 16:1543011. doi: 10.3389/fpls.2025.1543011
Received
10 December 2024
Accepted
16 May 2025
Published
04 June 2025
Volume
16 - 2025
Edited by
Raul Antonio Sperotto, Federal University of Pelotas, Brazil
Reviewed by
Mir Muhammad Nizamani, Shantou University, China
Youzheng Zhang, Ministry of Natural Resources, China
Updates
Copyright
© 2025 Chen, Arif and Tian.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Muhammad Arif, muhammadarif@gltu.edu.cn; arifgreenexpert@outlook.com
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.