 Michael W. Taylor
Michael W. Taylor Céline A. M. Ferré1
Céline A. M. Ferré1 J. Allan Feurtado
J. Allan Feurtado Aaron D. Beattie
Aaron D. Beattie- 1Department of Plant Sciences, College of Agriculture and Bioresources, University of Saskatchewan, Saskatoon, SK, Canada
- 2Aquatic and Crop Resource Development, National Research Council of Canada, Saskatoon, SK, Canada
Lodging caused by stem buckling or root anchorage failure results in significant economic losses each year globally due to increased disease load, downgrading of quality, and yield loss. To increase lodging resistance in western Canadian barley, a study was undertaken to identify traits associated to lodging score. Mechanical and architectural traits encompassing whole plant, stem and root features of 13 spring barley genotypes, representing a diversity of classes, height, and lodging resistance, were evaluated at six field locations over three years. Correlation analysis identified plant height, internode length, crown root angle and root system solidity as traits with the largest influence on lodging (r = 0.37, 0.27, -0.32 and 0.62, respectively). A structural equation model (SEM) was created to further evaluate which traits had direct or indirect influence on barley lodging. The best fitting SEM included nine traits that captured aspects of the whole plant, stem and root system. Plant height (effect estimate = 0.46) and root system solidity (0.14) showed a direct influence on lodging, while root angle had both direct (-0.21) and indirect (via height) influence on lodging. Stem strength, stem outer diameter, and stem volume had indirect effects on lodging through height, while root network area, convex area and total root length had indirect effects on lodging through root system solidity. The three traits that directly influenced lodging in both correlation analysis and SEM, plant height, root angle, and root system solidity, displayed moderate to high heritability (0.85, 0.78 and 0.57, respectively), thus making them suitable for breeding selections. Collectively, this study confirmed the role of plant height and root angle in lodging, identified root system solidity as a novel trait associated to barley lodging resistance, and revealed root imaging as a new screening tool to help breeders select for, and improve, lodging resistance in the absence of observable field lodging.
1 Introduction
Barley (Hordeum vulgare L.) is the 4th most grown field crop globally, with 148 million tons of annual production (United States Department of Agriculture, 2025) and an annual export trade value of $9.5 billion USD over the past decade (World Bank, 2025). In Canada, barley is the 3rd largest crop by area, with an average of 3.0 million hectares seeded over the past five years (Statistics Canada, 2025a). It is estimated that barley contributes over $14.4 billion CND annually to the Canadian GDP through farm-gate sales, exports of malt and barley, taxation on alcohol, and employment creation (LMC International, 2023).
Lodging is a detrimental phenomenon that occurs globally throughout all barley-growing regions with yield losses estimated between 3-40% depending on the growing season, growth stage at which lodging occurs and cultivar (Dyson et al., 1984; Laidig et al., 2022; Stanca et al., 1979). This can result in significant financial losses to the industry due to the combination of yield loss, increased disease load, harvest difficulty, and negative impacts on grain quality (Berry et al., 2004). For example, annual losses in the UK due to lodging across all cereal crops was estimated at ₤60-200 million (Baker et al., 2014). In Canada, there was an average price difference of $55 per ton between malt and feed barley price over the past decade (Statistics Canada, 2025b). Thus, downgrading barley from malt to feed due to increased disease load or decreased quality as a result of lodging would result in a loss of $200 per hectare.
Berry et al. (2006) described lodging as “buckling in any part of the stem (stem lodging) or failure of the root-soil anchorage system (root lodging)” which results from the interaction of environmental, agronomic, and genetic factors. The primary environmental factors typically associated with lodging are wind and precipitation, but disease, insects, and soil composition are also contributing elements (Bayer Crop Science, 2021; Gavloski, 2019; Lynch, 1995; Ma and Yamaji, 2006; Niu et al., 2016; Sterling et al., 2003). For example, Niu et al. (2016) observed that sustained wind speeds of 14.9 m/s produced 20% lodging in wheat, while wind gusts of 26.5 m/s could cause 80% lodging. When heavy rain was introduced a wind speed of only 8.4 m/s was required to cause severe lodging (Niu et al., 2016). Following rain events, the resulting increase in soil moisture can impact lodging by affecting root anchorage strength. An increase in soil moisture from 17% to 26% decreased soil shear strength and the peak load that wheat roots could resist before root anchorage failure occurred (Easson et al., 1993).
Agronomic practices such as fertilizer rates, seeding rates and the use of plant growth regulators also influence lodging. Higher nitrogen rates cause an increase in the shoot to root ratio which leads to greater lodging. At low nitrogen application rates (0.09 g nitrogen per plant) the shoot to root weight ratio was observed to be 2:3, but increased to 3:1 at higher nitrogen rates (0.46 g nitrogen per plant) (Grossman and Rice, 2012). O’Donovan et al. (2011) observed that barley lodging was more prevalent as nitrogen increased across five application rates ranging from 0-120 kg/Ha. Increasing soil potassium has been documented to reduce lodging in corn (Melis and Farina, 1984), oat (Casserly, 1956), rice (Liao et al., 2023) and wheat (Beaton and Sekhon, 1985), possibly by increasing stem wall strength which has been observed in some studies (Li et al., 2022; Melis and Farina, 1984). Seeding rates influence competition among plants and lower seeding rates have been reported to limit competition which leads to increased stem strength, decreased height and less lodging (Hobbs et al., 1998). Higher seeding rates in barley were shown to increase height (Tidemann et al., 2020) and when combined with later seeding dates caused an increase in lodging rate (O’Donovan et al., 2012).
Plant growth regulators (PGRs) are another option to manage lodging in cereal crops through reduction of plant height and increasing stem diameter and strength (Jung, 1984). PGRs, such as chlormequat chloride and trinexapac-ethyl, inhibit gibberellin synthesis which reduces cell elongation and ultimately internode length. Despite the benefits of PGRs, issues associated with variety- and environment-specific efficacy (Rajala and Peltonen-Sainio, 2002; Tidemann et al., 2020; Wiersma et al., 1986), delayed maturity (Ma and Smith, 1992a) and decreased grain weight (Ma and Smith, 1992b; Tidemann et al., 2020) are undesirable attributes associated with their use.
There are several important plant traits with a strong connection to lodging resistance, including height, stem strength and root plate spread (Berry et al., 2006) that can be targeted by plant breeders. Plant height impacts lodging in two critical ways. Firstly, a tall plant is exposed to greater wind force which increases the tendency for stem failure or uprooting (Berry et al., 2006), and secondly, the leverage created by the weight of the head, which can cause stem bending, is greater for a tall plant (Kristensen et al., 2016; Kuczyńska et al., 2013). Stem strength represents the ability of the stem (or internode) to resist forces, such as wind, and is influenced by both physical properties and composition. For example, a large basal internode diameter and thicker stem walls have been associated with higher stem strength in wheat (Yao et al., 2011). Similarly, the STRONGCULM2 (SCM2) gene in rice was demonstrated to increase stem wall thickness and stem diameter which imparted better lodging resistance (Ookawa et al., 2010). With regards to chemical composition, increased expression of enzymes involved in lignin synthesis as well as increased lignin deposition have been associated with stronger lodging resistance (through increased stem strength) in wheat and barley (Begović et al., 2018; Zheng et al., 2017).
Root characteristics that influence lodging are less understood due to the difficulty of studying below-ground traits, however, traits like root plate spread, angle, and volume have been shown to increase anchorage strength and subsequently prevent lodging. For example, Wu and Ma (2018) identified that root volume was positively associated with root anchorage strength in canola. Crook and Ennos (1994) indicated that root anchorage strength (and thus lodging resistance) was dependent upon the root angle and strength of the crown roots. Similarly, root plate spread in spring and winter wheat was correlated with root anchorage across multiple environments around the world (Berry et al., 2003c; Dreccer et al., 2020; Piñera-Chavez et al., 2016a). Root growth angle and volume have also been reported in maize to be important for root-lodging resistance (Hebert et al., 1992; Zhang et al., 2022).
Within the context of the traits listed above, the development of lodging models for wheat and barley have been foundational for lodging research and identified both stem and root lodging as important determinants for overall standability (Baker et al., 2014; Berry et al., 2006, 2007). In these models, wheat (or barley) is represented by two masses, the ear and root crown, connected through a weightless stem (Baker et al., 2014). In the model for barley, Berry et al. (2006) demonstrated that stem diameter of the middle internodes had the largest effect on stem lodging risk, while changes to ear area, drag coefficient, plant height, shoot natural frequency, and stem strength had moderate influence. For wheat, stem radius was identified as most important, followed by stem failure stress and ear area. However, in wheat, failure of the stem has been modeled and demonstrated experimentally to occur close to the soil surface (Berry et al., 2003a, c). For root lodging, anchorage failure in barley has been shown to be like wheat, where the most important traits were spread of the root plate and tiller number with structural rooting depth and ear area contributing less (Berry et al., 2003c, 2006).
More recently, structural equation modeling has been applied to evaluate and quantify the causal relationships among traits influencing lodging-related traits such as stem strength. In the structural equation model (SEM) developed by Li et al. (2023), flag leaf net photosynthetic rate, stem chemical components (lignin, cellulose, soluble sugars), filling degree, and stem wall thickness were identified as key components of stem breaking strength in wheat. Structural equation modeling is a multivariate analysis method that allows for the examination of hypothetical models that describe large, intercorrelated and complex systems (Lamb et al., 2011) by analyzing the direct and indirect influences of numerous variables relevant to the system (Kline, 2015). Hypothetical models are constructed to portray causal relationships among relevant variables based on a priori knowledge obtained from past observations and/or relevant literature. The covariance matrix calculated among the variables in the hypothetical model is then assessed against the actual covariance matrix derived from experimental data to determine how well the model represents the data. Through an iterative process of adding or removing variables, the initial hypothetical model is altered and reassessed against the experimental data until a statistically significant, good fitting model is obtained (Lamb et al., 2011). While SEMs and correlation analysis both describe the relationship between variables, the multivariate aspect of SEM provides additional insight as to how a group of causal traits interact to influence the primary trait of interest.
The most common and simple lodging assessment method is by visual scoring of field plots (Berry et al., 2004). While this is traditionally done by eye, the use of unoccupied aerial vehicles (UAV) carrying multispectral cameras and other sensors has now been shown to accurately assess lodging (Chauhan et al., 2019; Sakamoto et al., 2010; Wilke et al., 2019). Visual or UAV lodging assessment is only possible when suitable environmental conditions (e.g. storm events) and management techniques (e.g. higher nitrogen application) promote lodging, however, it is often the case that lodging does not occur. Various methodologies have been used to assess lodging resistance in the absence of lodging by testing the mechanical or bending resistance at the whole plant or canopy level, or by assessing the strength of individual stems (Zhang et al., 2016; Erndwein et al., 2020). Berry et al. (2003b) designed an H-shaped push-bar device that measures the force required to displace a collection of stems from the vertical position. Other instruments which have been developed include the ‘Stalker’, a T-bar device which continuously records the resistance force of a group of plants when pushed from the vertical position, and a prostrate tester which measures the force required to push a collection of stems to 45 degrees from the vertical (Heuschele et al., 2019; Xiao et al., 2015).
Overall, the identification and continued refinement of screening methods which can be used in the absence of visual lodging will help breeders select for, and improve, lodging resistance. The present study focused on evaluating a variety of field and lab-based methods that measured mechanical and architectural features of barley to determine which plant, stem, or root traits were most impactful, as assessed through correlation analysis and SEM, to lodging resistance in western Canada. The ultimate goal of this study was to identify the best methods and traits which can be used for barley lodging selections in a breeding program.
2 Materials and methods
2.1 Plant materials
Thirteen barley genotypes representing North American and European germplasm that are adapted to western Canada were used in this study (Table 1). The genotypes represent a diversity of spring barley classes (i.e. malt, feed, and forage) that vary in plant height and lodging resistance.
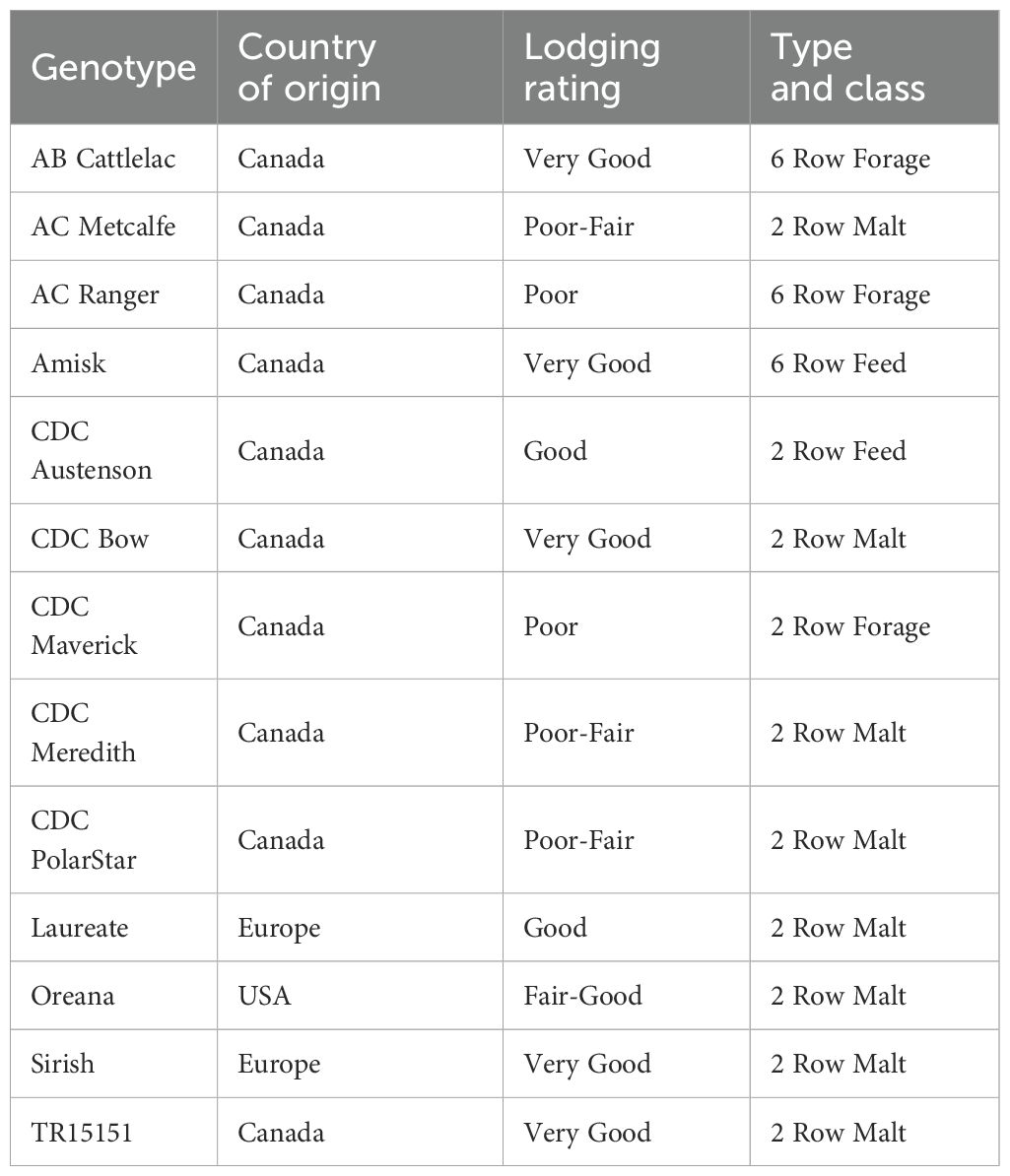
Table 1. Description of the barley genotypes used in the study.
2.2 Field trial design
Field trials were completed in the summers of 2020-23 as randomized complete block designs with three replications. The 2020 trials were grown at Saskatoon, SK (irrigated; 52°08’N, 106°36’W; 486m above sea level; dark brown, fine texture clay-clay loam soil) and Waldheim, SK (52°37’N, 106°39’W; 549m above sea level; black medium texture loam soil), 2021 trials were grown at Saskatoon, SK, 2022 trials were grown at Saskatoon, Kernen Crop Research Farm (KCRF; 52°09’N, 106°31’W, 486m above sea level, orthic dark brown, clay-clay loam soil) and Rosthern, SK (52°39’N, 106°20’W, 505m above sea level, black, medium texture loam soil), and 2023 trials were grown in Saskatoon, KCRF, and Rosthern. All plots were seeded at a rate of 300 plants/m2 and a depth of 3.8 cm. Plots were 1.22 m x 3.66 m (4.47 m2). The seed was treated with 325 ml/100 kg seed of Raxil Pro (active ingredients: tebucanzole, prothioconazole and metalaxyl; Bayer Crop Sciences). Fertilizer was applied during seeding using a 32-23-0 blend at 56 kg/Ha (18 kg/Ha N and 14 kg/Ha P) via granular fertilizer, application rates may change based on lab soil results. At the Saskatoon, KCRF and Waldheim locations, the field was chem-fallowed the prior year, at Rosthern the prior year was sown to canola. Plots were sprayed at the Zadoks stage 13 stage with Axial Extreme IPAK (active ingredients: pinoxaden, fluroxypyr, pyrasulfotole and bromoxynil; Syngenta) at 1.24 l/hectare. Fungicides were not applied.
2.3 Trait data collection
A total of 18 plant, stem and root traits were measured from 2021, 2022 and 2023 Saskatoon, 2022 and 2023 KCRF, and 2022 Rosthern sites. All traits were measured at the late milk to early dough stage of development (Zadoks stages 77-83). A description of all traits is provided in Supplementary Table 1.
2.3.1 Plant traits
Visual lodging scores were recorded at sites with differential lodging among the genotypes where a score of 1 indicated no lodging and a score of 9 indicated that the whole plot was completely lodged. Plant height (mm) was measured from soil level to the top of the head not including the awns. Force (i.e., plant bending resistance) measurements were collected with the Stalker (Heuschele et al., 2020) on two sections of the middle row of each plot. The Stalker was placed on the ground parallel to the middle row of a plot and the horizontal load-detecting bar was adjusted to half the plot’s average height. The load-detecting bar was then slowly pushed from 0° to 45° from vertical and both the force (N) and angle were recorded by the onboard computer. The number of plants contacted by the load-detecting bar was recorded and force per plant was calculated using the maximum force recorded by the Stalker.
2.3.2 Stem traits
Five or six stems from the Stalker-tested sections were harvested and the second internode was cut and used for subsequent measurements. The length of the 2nd internode (mm) was measured and then stem strength was measured on the internode using a custom three-point bend testing machine (designed by Dr. Scott Noble, University of Saskatchewan). The stem was placed across two fulcra which were 10 cm apart with an anvil placed above the middle of the stem. The anvil was moved at a rate of 55 mm/min until the internode failed. The peak applied force (N) was recorded and used to calculate stem strength (N·mm). Internode stem strength was calculated according to Equation 2.1.
where F is the maximum bending force (N), that is, the force exerted at the time of breaking; and L is the length of the second internode (mm) (Gordon, 1978; Wu and Ma, 2018).
After the three-point test was completed a 1-mm cross-section was cut from the internode beside the failure point. Images of the cross-sections were taken using an inverted 18MP USB 3.0 Real-Time Live Video Microscope Digital Camera connected to an Amscope stereo microscope in 2021. In subsequent years a 3MP USB 2.0 High-speed Color CMOS C-Mount Microscope Camera was used. Images were processed using custom software (developed by R. Peters, University of Saskatchewan) to extract the minimum stem wall thickness (mm), maximum inner stem diameter (mm) and maximum outer stem diameter (mm). Stem volume (mm3) was calculated using the inner and outer stem diameters to obtain the radius, and internode length to obtain the length.
2.3.3 Root traits
The root systems of the five or six plants sampled for the internode stem measurements were excavated through shoveling. The soil volume excavated around each plant was approximately 20 cm in diameter and 25 cm in depth. Roots were placed into water-filled tubs for 15-30 minutes to remove adhering soil followed by measurement (using a ruler and protractor) of the root systems to determine their maximum root angle (degrees) and root plate spread (mm). Root angle was measured as the greatest angle between opposing crown roots. The root plate spread (or simply root spread) was the distance between the tips of the roots used to measure crown root angles (Berry, 1998).
Field excavated root systems were taken to a lab and imaged with a Nikon D850 DLSR camera. Images were imported into ImageJ2 (Rueden et al., 2017) where they were cropped and the contrast adjusted to improve differentiation between roots and the background medium. Images were then exported into RhizoVision Explorer (Seethepalli et al., 2021) to extract eight root traits: root diameter (mm), root number, total root length (mm), root system depth (mm), root system width (mm) network area (mm2), convex area (mm2), and solidity (mm3). The root trait solidity describes the ratio obtained by dividing the total root area (i.e. network area) by the area captured within a convex polygon (i.e. convex area) that encompasses the outer dimensions of the root system (Figure 1).
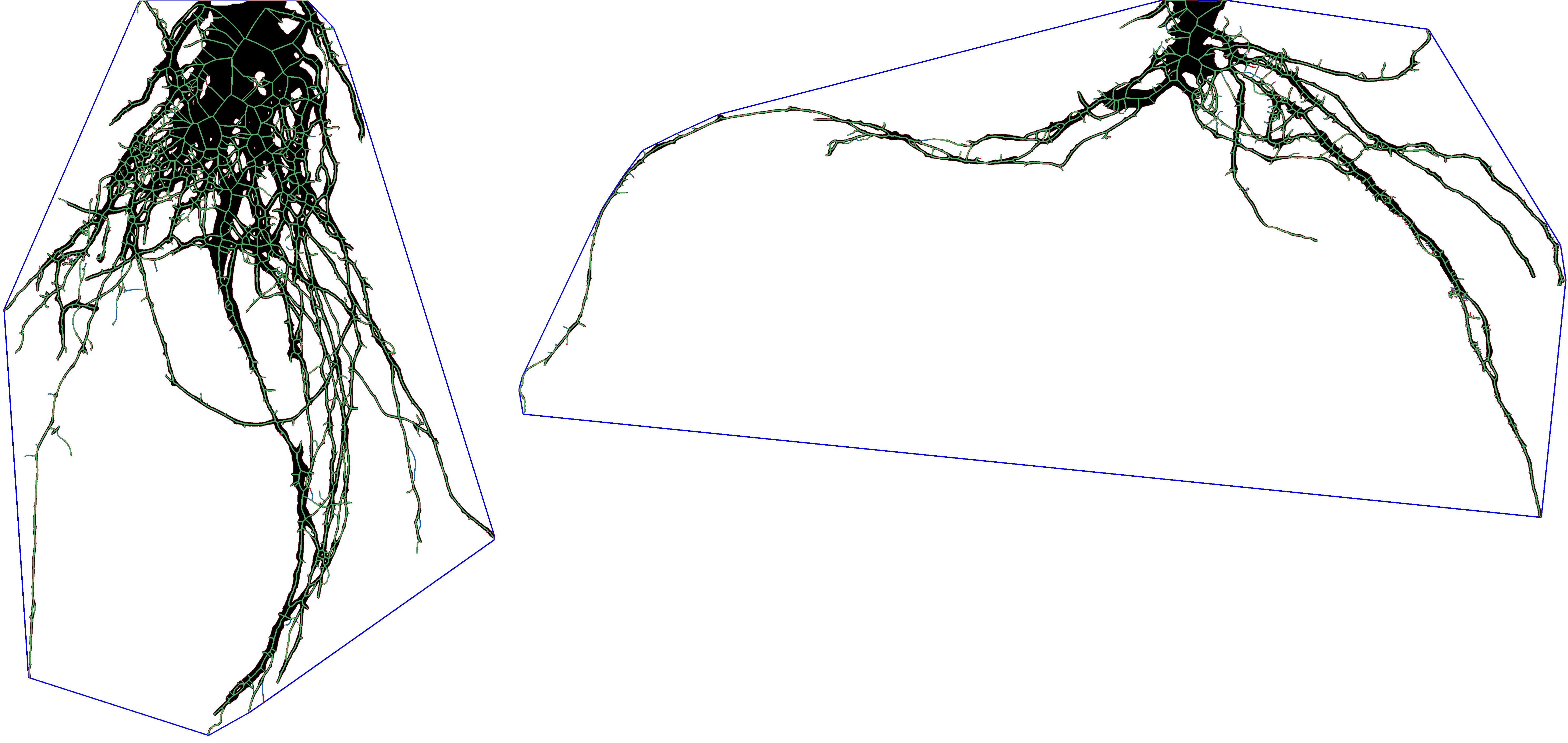
Figure 1. A post-processed root system image obtained from RhizoVision Explorer used to calculate root system solidity. The blue line represents the convex hull polygon that encompasses the root system, while black and green pixels represent the roots. Dividing the pixelated root area (i.e. network area) by the area encompassed by the polygon (i.e. convex area) allows the calculation of solidity. The image on the left represents a root system with high solidity while the root system on the right is one with low solidity.
2.4 Statistical analysis
The effect of genotype, site-year, genotype by site-year interaction and block (i.e. replication) on each of the 18 traits were analyzed as a RCBD using a linear mixed effect model in Minitab (v. 21.2, Minitab LLC). Genotype was treated as a fixed effect while site-year, genotype by site-year interaction and blocks nested within site-year were treated as random effects. The assumptions of homogeneous variances and normal distribution of residuals were evaluated using plots of predicted values versus residuals and the Shapiro-Wilk test, respectively. A total of six site-years were used in this analysis. Significant site-year and/or genotype by site-year interactions were observed for almost all traits (Supplementary Table 2) so adjusted means were calculated for each genotype by site-year. Adjusted means for significant genotypic effects were separated using Tukey’s HSD post hoc test at a 0.05 significance level.
Four site-years produced differential lodging among the 13 genotypes from which a single adjusted mean for lodging was calculated for each genotype (using the linear mixed model described above) and used for correlation analysis to the 18 traits. A single lodging value for each genotype was used because these trials were not the same as those used to measure the 18 plant, stem and root traits and using a single value based on multiple site-years provides a more accurate indication of a genotype’s lodging resistance. As such, Pearson correlation coefficients were calculated by incorporating the single lodging value associated with each genotype into each of the six site-year datasets that contained adjusted means for each genotype for the 18 plant, stem and root traits (i.e. the lodging data was included six times in the overall dataset used for the correlation analysis). Correlation heat plots were produced with the packages ggplot2 (Wickham, 2016) and ggcorrplot (Kassambara, 2019) using the R statistical computing language (R Core Team, 2021) in the RStudio development environment (Posit Team, 2025).
Broad-sense heritability was calculated for all traits according to Equation 2.2 (Cooper et al., 1994).
where H is the broad sense heritability, σg2 is the genotypic variance, σe2 is the error variance, is the genotype by site-year interaction, ne is the number of site-years and nr is the number of replications.
A structural equation model (SEM) was created with the ‘sem’ function in the ‘lavaan’ R package (Rosseel et al., 2022) to quantify the direct and indirect influence of the traits measured in this study on lodging. Structural equation models require a sample size greater than 150, thus individual plot data from the six site-years was used to meet sample-size requirements. The initial model was built based on inclusion of variables identified in this study with significant (P<0.05) correlation to lodging and additional variables shown to be relevant to lodging in other studies. Traits were added or removed in an iterative process to produce the best-fitting model. According to Xia and Yang (2018), the indices of a good fitting SEM are as follows: a chi-square P-value of >0.05, a comparative fit index (CFI) of >0.95 and a standardized root mean square residual (SRMR) of <0.08. According to Cohen (1988), estimate effects (Est) with a value of <0.1 indicate small effects, 0.3 are moderate effects and >0.5 are large effects on the variable by the predictor variable. Effect estimates were calculated by regression according to Equation 2.3.
where Y is the predicted value, X is the predictor variable, e is the error term and b is the path coefficient. The path coefficient is calculated according to Equation 2.4.
where cov(X,Y) is the covariance between the variable being predicted and the predictor variable and var(X) is the variance of the predictor variable (Rosseel et al., 2022). Indirect path coefficients are calculated by multiplying the path coefficients for all steps between traits (Baron and Kenny, 1986).
3 Results
3.1 Variation and heritability of plant, stem and root traits
Analysis of variance demonstrated that significant genotypic effects were observed for all plant, stem and root traits (P = 0.04 to <0.01) (Supplementary Table 2). Summary trait data for each genotype is provided in Table 2 and Supplementary Table 3. The greatest range in trait values, expressed as a proportion calculated between the largest and smallest mean among the 13 genotypes, was observed for stem strength. A ratio of 2.90 was observed for this trait with Laureate showing the lowest value at 28 N·mm and AB Cattlelac having the highest value at 82 N·mm (Table 2). Other traits with a large range in values among the 13 genotypes included root spread (ratio = 2.00; Sirish = 46 mm, Amisk = 92 mm), stem volume (ratio = 1.94; Laureate = 2104 mm3, AB Cattlelac = 4092 mm3), force per plant (ratio = 1.89; Laureate = 0.08, CDC Meredith = 0.16) and convex area (ratio = 1.87; Sirish = 4575 mm2, Amisk = 8571 mm2) (Table 2). Root diameter showed the least range in values with a ratio of 1.15 (AC Metcalfe = 0.62 mm, Ac Ranger = 0.71 mm) (Table 2). Traits with similarly low trait value ranges included internode length (ratio = 1.23; Laureate and Oreana = 100, Amisk = 123), stem inner diameter (ratio = 1.27; TR15151 = 4.12 mm, AB Cattlelac = 5.23 mm), stem outer diameter (ratio = 1.28; Laureate = 6.73 mm, AB Cattlelac = 8.65 mm) and root system depth (ratio = 1.30; CDC Maverick = 90 mm, Amisk = 118 mm) (Table 2).
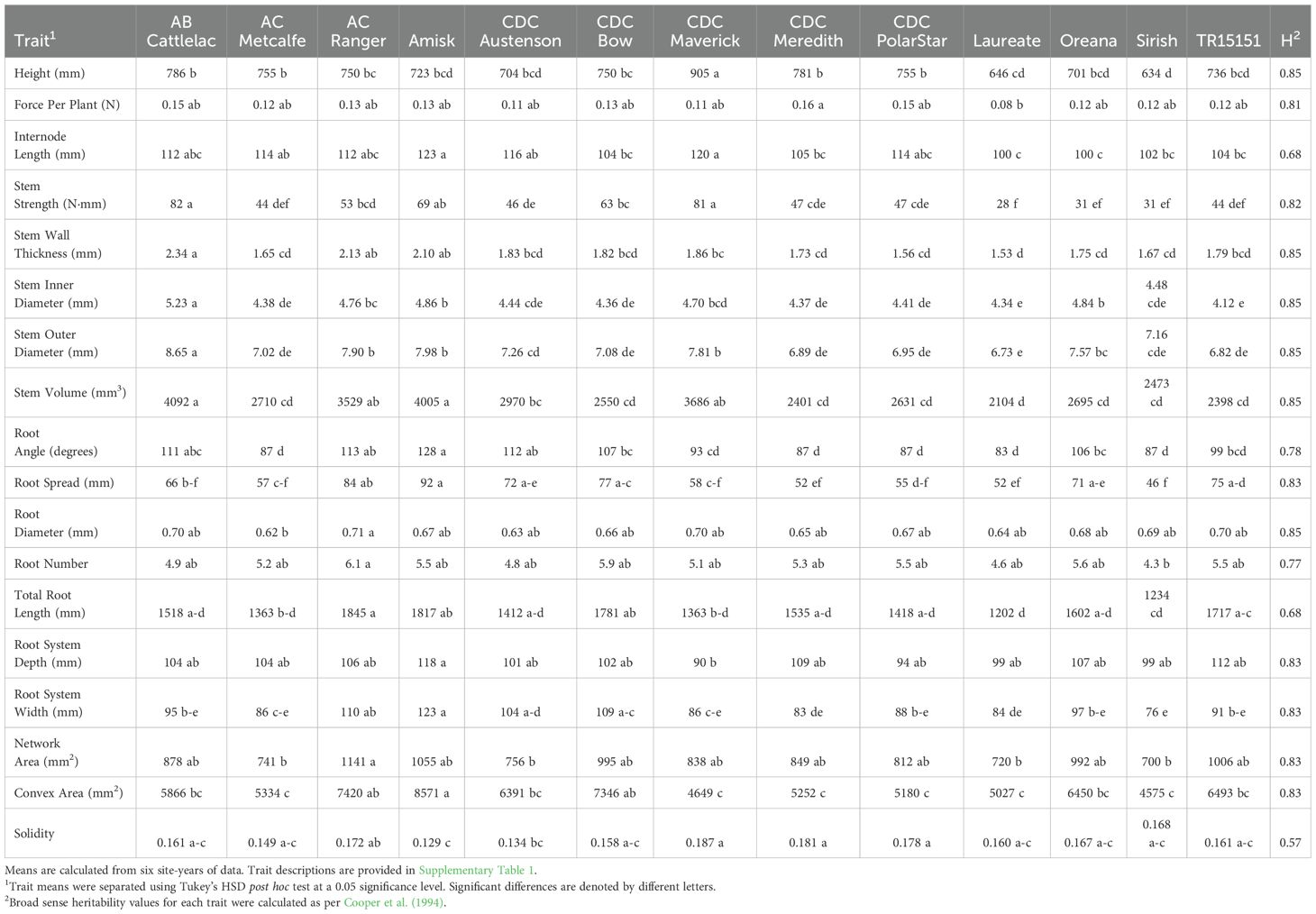
Table 2. Summary of adjusted means for the 18 plant, stem and root traits measured on the 13 barley genotypes used in the study.
Significant differential lodging (P< 0.01; Supplementary Table 2) was observed among the 13 genotypes at the 2020 Waldheim and 2020, 2022 and 2023 Saskatoon sites, with AB Cattlelac, Amisk, CDC Austenson, CDC Bow, Laureate, Oreana, Sirish and TR15151 generally displaying lodging resistance while AC Metcalfe, AC Ranger, CDC Maverick, CDC Meredith and CDC PolarStar were often susceptible to lodging (Table 3).
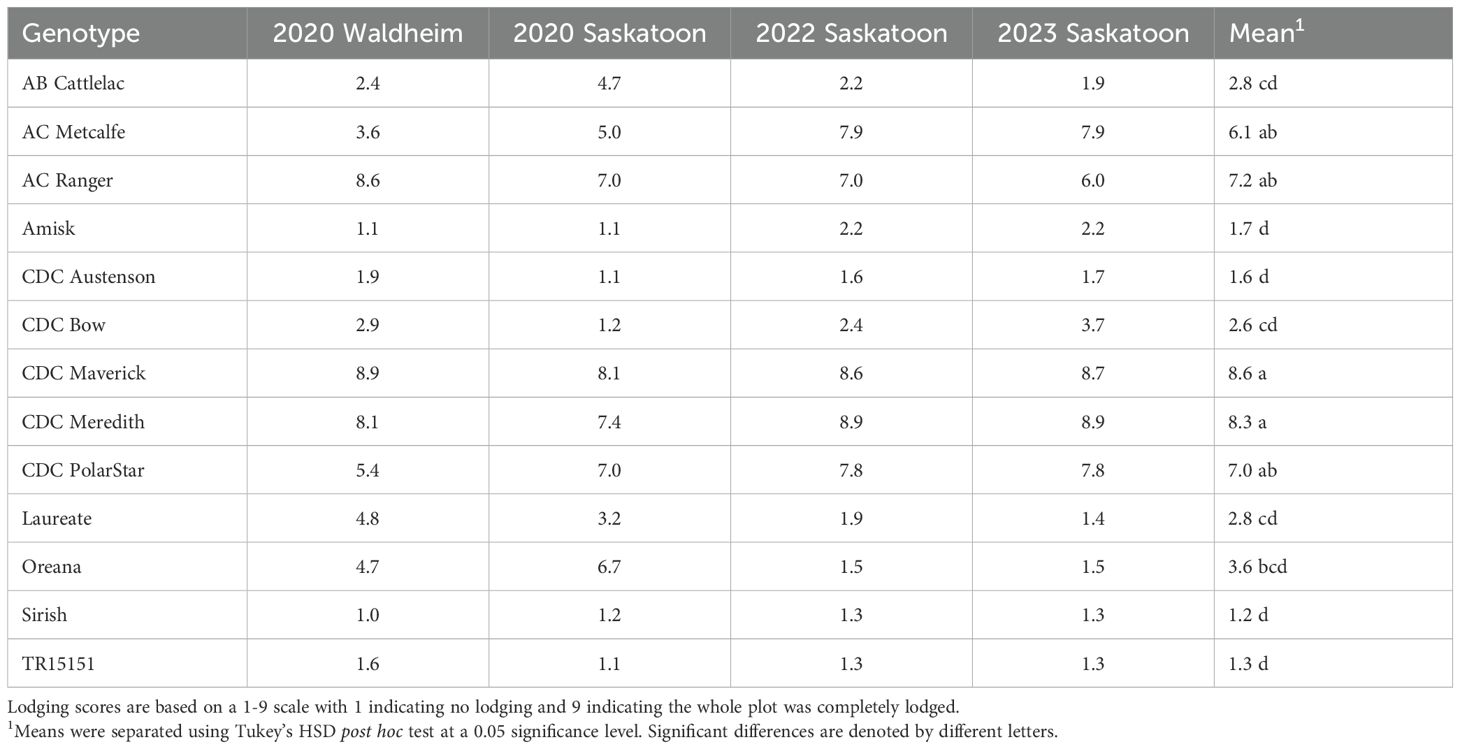
Table 3. Summary of lodging scores obtained for the 13 genotypes from four site-years that displayed significant differential lodging.
Significant site-year and/or genotype by site-year interaction effects were observed for most traits (P = 0.05 to <0.01) except for internode length, root number, total root length, root system depth and solidity (Supplementary Table 3). Summary trait data for each site-year is provided in Supplementary Table S4.
Broad sense heritability estimates were moderate to high for all 18 plant, stem and root traits (Table 2). Among the plant traits, force per plant showed a heritability of 0.81 while plant height was determined to have a heritability of 0.85. Stem traits had heritability values that ranged from 0.68 for internode length to 0.85 for the stem size traits (stem wall thickness, inner diameter, outer diameter and volume). Finally, root traits displayed heritability values from 0.57 for solidity to 0.85 for root diameter.
3.2 Plant, stem and root trait correlations to lodging
Among the plant and stem traits, both height (r = 0.37) and internode length (r = 0.27) demonstrated significant (P< 0.05) positive correlations to lodging (Figure 2). Given that internode length is one component of height, it was interesting that the correlation between height and internode length (r = 0.38) was not stronger. The stem size traits (stem wall thickness, inner diameter, outer diameter and volume) were highly and positively correlated to one another (r = 0.72-0.96), with the correlation between stem inner diameter and stem thickness being moderate (r = 0.54). All stem size traits showed positive correlations to stem strength (r = 0.38-0.66), with stem wall thickness and stem volume both showing the highest influence on stem strength (r = 0.66) and stem inner diameter showing the lowest influence (r = 0.38). Similar correlation strengths were observed between the stem size traits and internode length (r = 0.36-0.78), with stem volume displaying the highest correlation and stem wall thickness the lowest. Following this pattern of correlations, stem strength showed a positive correlation to internode length (r = 0.58). Stem size traits all displayed weaker correlation to height than to internode length (r = 0.28-0.41) with stem wall thickness having a non-significant correlation. Similarly, stem strength was not significantly correlated to height.
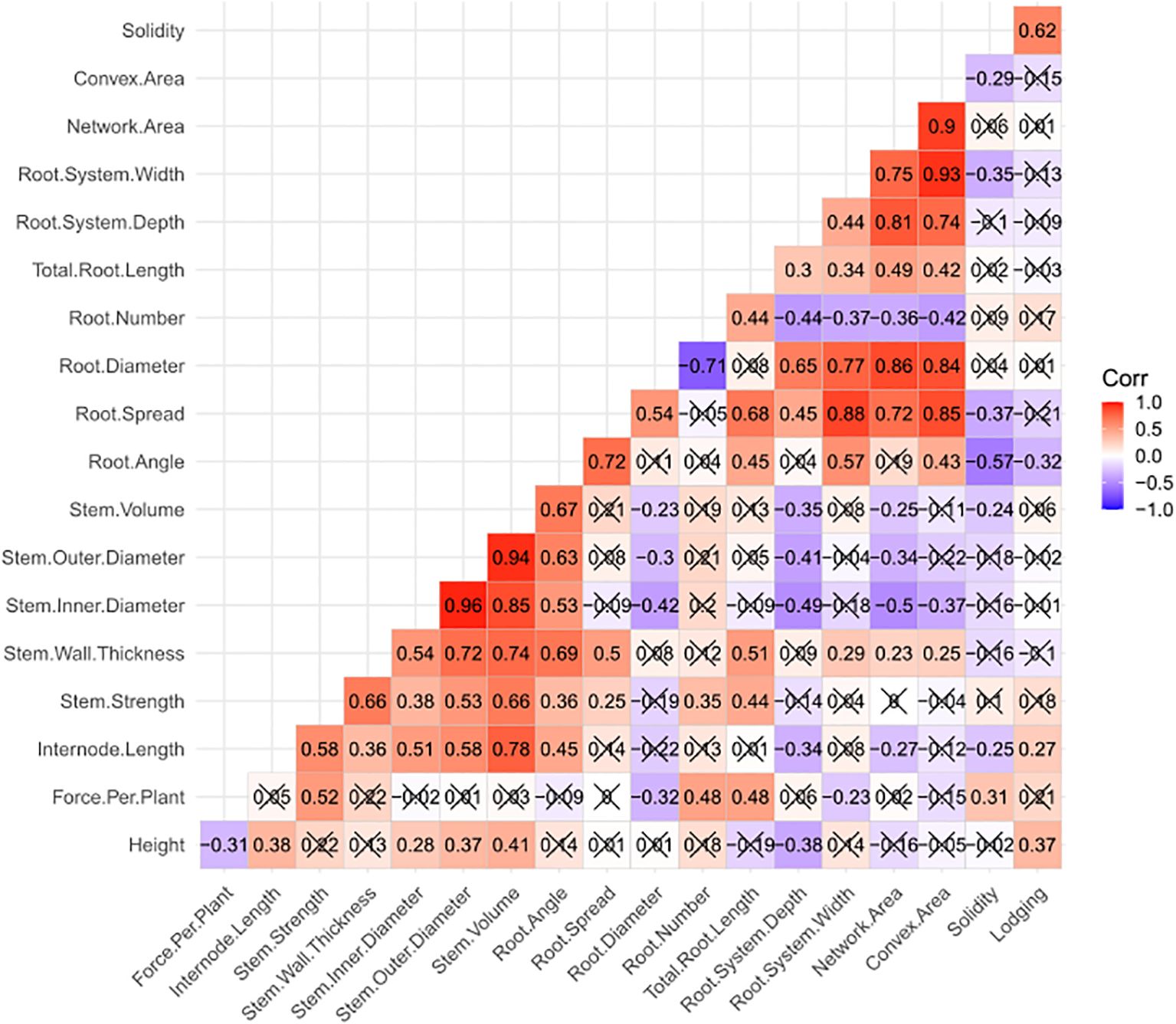
Figure 2. Heatmap displaying Pearson correlation coefficients among 18 plant, stem and root traits measured in the study. Lodging data is based on four site-years, while all other trait data is based on six site-years. Correlations that are not significant (P > 0.05) are crossed out.
Two root traits were also observed to have significant correlations to lodging, with root angle having a negative correlation (r = -0.32) and solidity having a positive correlation (r = 0.62) (Figure 2). There was also a negative correlation between root angle and solidity (r = -0.57). This was the strongest correlation with solidity, with root spread, root system width and convex area showing similar, but lower negative correlations (r = -0.29 to -0.37). It was interesting to note that network area (a component of solidity along with convex area) and root system depth were not significantly correlated to solidity. Convex area and network area were very strongly and positively correlated (r = 0.90) and shared similar strength of correlation with root number (r = -0.42 and -0.36, respectively), root diameter (r = 0.84 and 0.86, respectively), root system width (r = 0.93 and 0.75), root system depth (r = 0.74 and 0.81, respectively), total root length (r = 0.42 and 0.49, respectively) and root spread (r = 0.85 and 0.72, respectively). Root angle and root spread were strongly and positively correlated to one another (r = 0.72), and both were positively correlated to root system width (r = 0.57 and 0.88, respectively) and total root length (r = 0.45 and 0.68, respectively). In addition, root spread was positively correlated with both root system depth (r = 0.45) and root diameter (r = 0.54). Root system width and depth showed a moderate positive correlation to one another (r = 0.44), and both similarly influenced root characteristics including total root length (r = 0.34 and 0.30, respectively), root diameter (r = 0.77 and 0.65, respectively), and root number (r = -0.37 and -0.44, respectively). Finally, root number was moderately and positively correlated to total root length (r = 0.44) and negatively correlated to root diameter (r = -0.71).
Correlations were also observed between root traits and above-ground traits, however, these will not be mentioned as the basis of such correlations are less clear, perhaps reflecting an underlying genetic basis, such as pleiotropy. However, significant correlations involving the four traits directly correlated to lodging (i.e. height, internode length, root angle and solidity) are worth noting. Root angle was observed to have moderate to strong positive correlations with internode length (r = 0.45), stem strength (r = 0.36) and the four stem size traits (r = 0.53-0.69). Solidity was also significantly correlated to internode length (r = -0.25) and stem volume (r = -0.24), but also with force per plant (r = 0.31). Height and internode length both showed a negative correlation with root system depth (r = -0.38 and -0.34, respectively), while internode length was also negatively correlated to root network area (r = -0.27).
3.3 A structural equation model for lodging
To build a structural equation model representing trait relationships to lodging, an initial model which hypothesized the causal relationships (i.e. paths) amongst variables and in relation to lodging was built (Figure 3). The inclusion of, and relationships between, variables included in the initial model was based on both first-hand knowledge and relevant literature. Internode length was considered a component of height so a path from internode length to height was included. Stem strength was included as a path to internode length and as a direct path lodging because of its strong correlation to internode length in this study and its importance to lodging resistance based on past studies (Berry et al., 2006). Stem size traits were included as direct paths to stem strength due to their strong correlations found in this study and based on their connection to stem strength in past research (Ookawa et al., 2010; Yao et al., 2011). Root network area and convex area were included as direct paths to solidity since these are the two component traits used to calculate solidity, the first describing the physical size of the roots and the second describing the outer shape of the root system within the soil. Root length and root diameter were included as direct paths to network area since these two traits were positively correlated to network area and collectively would relate to the physical size of the root system. A path from root angle to convex area was included due to their positive correlation and the idea that root angle is key to setting the overall outer shape of the root system. Finally, despite no direction correlation to lodging, force per plant was included as a direct path to lodging based on past literature indicating the connection between these two traits (Berry et al., 2003b). A direct path to force per plant from both solidity and internode strength was included since both root lodging (as influenced by solidity) and stem lodging (as influenced by stem strength) can be captured in the force per plant measurement (Berry et al., 2003b). An iterative process followed where the initial model was assessed by calculating and comparing the covariance matrix (known as the model-implied covariance matrix) associated with the variables in the initial model to the actual covariance matrix calculated for the variables using the measured data. The model-implied covariance matrix associated with the initial model, or subsequent models created by removing or adding paths in the initial model, were compared and when it was not significantly different (i.e. P > 0.05) from the actual covariance matrix, then the model was considered to accurately represent the data.
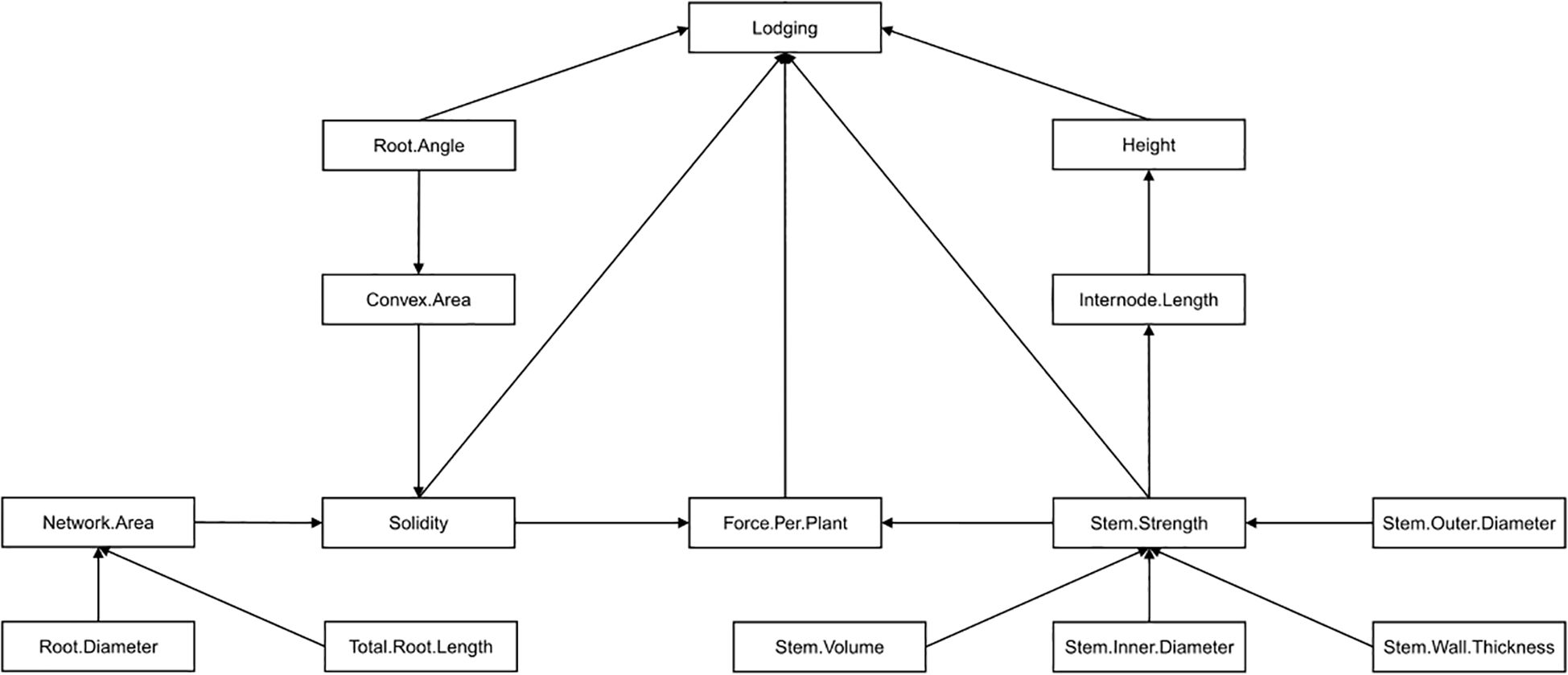
Figure 3. The initial hypothesized structural equation model describing the influence of 14 plant, stem and root traits on barley lodging. Traits were included in the model based on significant (P< 0.05) correlations to lodging observed in this study, significant correlations to one another observed in this study, or identified as relevant to lodging in other studies.
The iterative process of removing and including different sets of traits (paths) ultimately assessed 10 different models, where a best-fitting final model was identified with P = 0.10, CFI = 0.990, and SRMR = 0.04 (Figure 4; Table 4). Direct paths to lodging, consistent with observed correlations (Figure 2) were identified for height, root angle and solidity (r = 0.46, -0.21, 0.14, respectively). As with the correlation analysis, force per plant did not produce a significant path to lodging and therefore it was removed from the model, along with paths from solidity and internode strength to force per plant. The inclusion of an internode length to height path did not permit the creation of a good fitting model, but once removed the subsequent path from stem strength to height improved model fit. However, the direct path from stem strength to lodging was not significant and was removed from the final model. Among the four stem size traits included in the initial model, it was determined that stem outer diameter and stem volume were sufficient to capture the influence of these traits on stem strength, ultimately contributing to the final model. The inclusion of stem outer diameter and stem volume in the model is consistent with their correlation to stem strength observed in this study. A path from convex area to solidity remained in the final model which was consistent with the correlation observed between these two traits. Interestingly, the network area to solidity path was also significant in the final model which differed from the correlation analysis in which no significant correlation was observed between these traits. Between the total root length and root diameter paths to network area that were included in the initial model, only total root length remained as a significant path which was consistent with the correlation observed between these traits. The path from root angle to convex area proved to be not significant, but it was determined that a path from root angle to height was an important component to creating a good fitting model. This differed from the correlation analysis in which no significant correlation was observed between root angle and height.
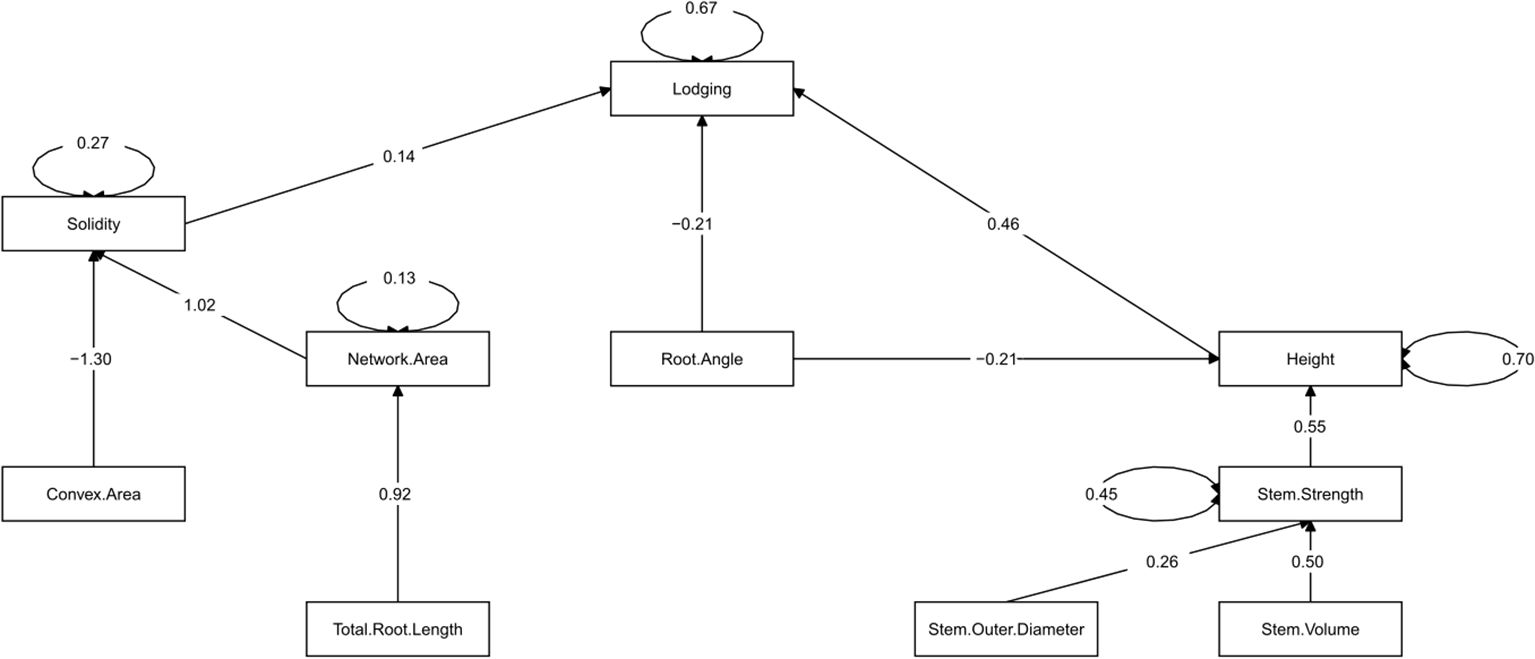
Figure 4. The final fitted structural equation model describing the influence of nine plant, stem and root traits on barley lodging. Causal relationships are indicated by path arrows with the predictor trait indicated at the tail of the arrow and the outcome trait at the head of the arrow. The number associated with each path arrow indicates the standardized effect estimate of the predictor trait on the outcome trait. Only significant paths (P< 0.05) are displayed. A circle created by two arrows indicates the unaccounted variation associated with the outcome trait.
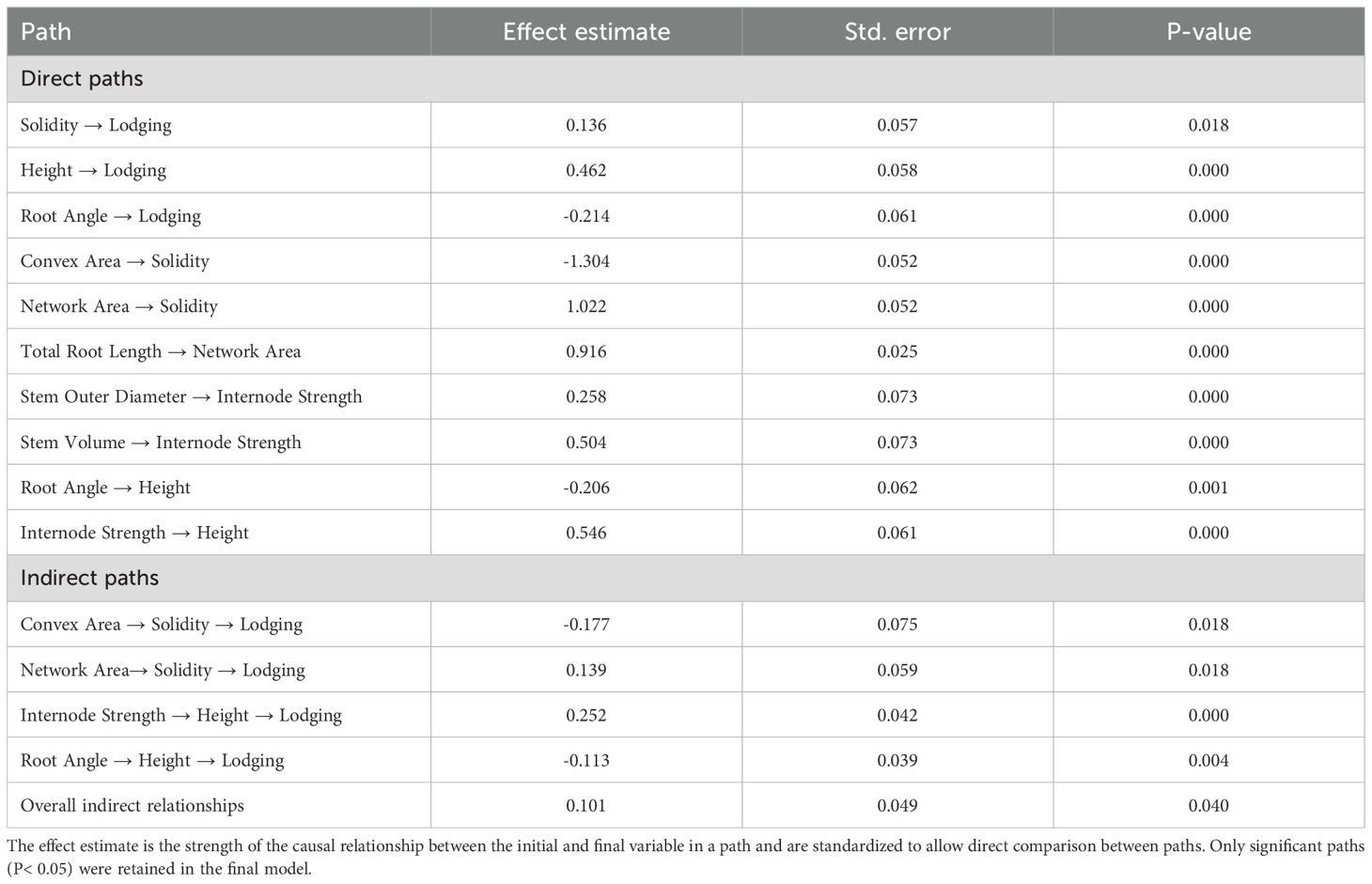
Table 4. Summary of the direct and indirect paths associated with the nine traits used in the final fitted structural equation model to describe barley lodging.
4 Discussion
Crop lodging resulting from either stem breakage or displaced root systems leads to challenging harvest conditions, reduced yield, lower grain quality and increased disease, thus diminishing profit margins for growers (Bayer Crop Science, 2021; Berry et al., 2006; Clarkson, 1981; Nakajima et al., 2008; Niu et al., 2016; Sterling et al., 2003). As such, lodging resistance is a key trait for improvement by plant breeders. Visual evaluation of lodging, either by eye or with the aid of UAVs, is the most common method to assess lodging, but is reliant on lodging to be present in a particular field environment which is frequently not the case. Methodologies to assess lodging risk in the absence of natural lodging have focused on: (1) the development of mechanical tools to assess the forces (resistance) required to push over stems, and (2) the identification of shoot and root traits which are most influential to lodging resistance (Berry et al., 2006; Erndwein et al., 2020). In this study, evaluation of a diverse set of 13 barley genotypes for 18 above and below-ground traits revealed traits that were significantly correlated to lodging and important in a SEM for lodging. The most relevant included the above-ground trait, plant height, and the below-ground traits, crown root angle and root system solidity. To move forward, a consideration of how these traits relate to lodging, in the context of the literature, is important for the ultimate success of defining traits and methods for generating cultivars with improved resistance.
4.1 Height is the primary above-ground driver of barley lodging resistance
Plant height, a well-documented factor related to lodging, exhibited a positive correlation with lodging (r = 0.37) with shorter plants being more lodging resistant. This result was consistent with Berry et al. (2006) who showed a 12% increase in lodging for every 25% increase in height. In cereals such as wheat and rice, introduction of the semi-dwarf trait during the ‘Green-Revolution’ not only promoted significant increases in harvest index but also increased lodging resistance by decreasing shoot (stem) leverage forces (Berry et al., 2004; Hedden, 2003). In barley, the introduction of semi-dwarf mutations to decrease lodging through reduced height is also generally associated with more robust culm (stem) phenotypes (Braumann et al., 2018; Kristensen et al., 2016). However, in wheat, it has been demonstrated that reducing height below an optimum of approximately 70-90 cm can reduce grain yield due to sub-optimal source-sink balance and light distribution to the canopy (Reynolds et al., 2009; Slafer et al., 2023). In barley, dwarf genotypes (as compared to semi-dwarf) often exhibit later maturity, increased disease susceptibility, and reduced malt quality (Mickelson and Rasmusson, 1994). Madić et al. (2016) reported that an optimal height for spring malting barley in Serbia has been reached (at about 80 cm) with winter malt barley heights of between 90-100 cm having the potential for further reductions to promote lodging resistance. In this study, genotypes with a lodging score less than four were on average 71 cm, whereas when the lodging score was greater than 4, the average height was 79 cm. The high heritability associated with plant height, along with its ease of measurement, make it a useful and suitable trait for early generation selection in breeding programs. It should also be noted that for forage barley, height can be an important trait driving dry matter (Gill et al., 2013), thus other traits would be important for reducing lodging risk if height compromises forage yields.
Stem strength has been regarded as an important trait to improve lodging resistance in wheat, barley, and canola (Berry et al., 2006; Hai et al., 2005; Wu and Ma, 2018). However, in the current study there was no direct correlation between stem strength and lodging resistance, suggesting that the stem strength variation present in western Canadian barley is sufficient to resist lodging and stem-based failure within the study’s environmental conditions and agronomic inputs. This is consistent with the observation that selections for shorter, more rigid stems which have occurred in breeding programs (over decades) have resulted in less stem-based lodging (Crook and Ennos, 1993) and placed more emphasis on the root system as the source of lodging (Crook and Ennos, 1994). The importance of stem strength, at least in the context of the genotypes used in this study, is to support taller plants, as seen by the strong positive path in the SEM. The positive correlation between these traits differs from reported relationships of shorter plants having stronger stems (Niu et al., 2012), a relationship which is also the basis of PGR effectiveness (Kamran et al., 2018a, 2018; Niu et al., 2022). The inclusion of tall forage barley genotypes in this study (i.e. CDC Maverick and AB Cattlelac) in which tallness is intentionally bred into these varieties to improve biomass is likely the reason for the positive correlation observed in this study as these two genotypes were the tallest and also had the highest stem strengths. Additionally, the observation that AB Cattlelac showed good lodging tolerance while CDC Maverick was the poorest further highlights that stem strength is not a predictive trait for lodging tolerance in this set of germplasm. The SEM also showed that both stem outer diameter and stem volume had positive paths towards stem strength. The importance of both stem diameter and cell wall thickness (which is a component of the stem volume calculation in this study) has been noted previously in wheat and rice (Ookawa et al., 2010; Piñera-Chavez et al., 2016a; Yao et al., 2011), thus indicating that these relationships also hold true in barley.
Root lodging is usually reported as the predominant form of lodging, especially in conditions of high soil moisture, but stem lodging can still be prevalent especially in conditions of high soil nitrogen (Crook and Ennos, 1994; Berry et al., 2003a; Wu et al., 2019). The priority for trait selections for stem versus root lodging resistance can vary for breeding programs depending on the production environment, agronomic inputs (e.g. PGRs), and breeding germplasm available. For example, in the Yellow-Huai River region of China, an initial wheat breeding target of reducing plant height transitioned to focus on improving stem strength (Zhang et al., 2020). The importance of trait selections to improve barley stem strength have recently been documented (Gianinetti and Baronchelli, 2024; Jaff and Jarvis, 2021). In comparison to wheat, it has been shown that barley cultivars typically have lower stem failure moment and flexural rigidity (Berry et al., 2006; Gianinetti and Baronchelli, 2024). Thus, there is perhaps further potential to improve stem strength by improving traits like stem diameter as has been accomplished in other cereals (Niu et al., 2022), although priority traits for western Canadian environments would target root anchorage improvement as discussed below.
The force required to push over a collection of stems has been investigated in relation to lodging resistance. For example, Berry et al. (2003b) demonstrated that reduced stem and root lodging in winter wheat was correlated to a greater amount of force required to push over a collection of stems. In addition, Mangin et al. (2022) showed that agronomic practices like lower plant density, split nitrogen treatments, and plant growth regulator application could also increase stem strength (pushing force), thus supplementing traits like stem wall thickness (Kashiwagi et al., 2008) or root anchorage traits like root spread, angle, length and volume (Berry et al., 2006; Wu and Ma, 2018; Zhang et al., 2022) that are correlated with pushing resistance. However, Heuschele et al. (2020) reported the opposite relationship between pushing force and lodging, with lower pushing force values associated with lodging resistant barley, oat and wheat varieties. In the current study, force per plant was correlated with familiar traits such as height, stem strength, root length and solidity (which captures features of the root system similar to volume), but neither the correlation analysis nor the SEM found that force per plant was relevant to lodging. One explanation for the lack of relationship between force per plant and lodging would be that once stem strength reaches the threshold to protect plants from stem failure, then flexibility becomes a more important trait that would allow barley plants to flex and recover (as opposed to acting as a rigid object against wind-imposed forces). As noted by Wu and Ma (2018), plants with strong, less flexible stems that can prevent stem buckling may become susceptible to root anchorage failure as the leverage exerted on a plant by the wind is translated to the root system. Greater flexibility of barley stems, in comparison to wheat stems, has been noted by Berry et al. (2006). Stems of barley landraces with thinner diameters have been shown to flex and curve without any stem failure with the spike eventually touching the ground (Jaff, 2019). As the current study was not able to measure shoot flexibility or elasticity, this would be a future point of focus.
4.2 Crown root angle and solidity are key below-ground traits impacting barley lodging resistance
Crown root angle emerged as an important trait associated with barley lodging in the current study. This is the first report to demonstrate a direct relationship between root angle and lodging in barley. In corn, increased brace root angle was also observed to enhance lodging resistance (Brune et al., 2017; Shao et al., 2021; Sparks, 2022; Zhang et al., 2022). Root angle has also been shown to be relevant to wheat root anchorage strength (Crook and Ennos, 1994), with root anchorage strength reported to be an important component of lodging resistance in wheat, barley and canola (Berry et al., 2006; Piñera-Chavez et al., 2016b; Wu and Ma, 2018). Crown root angle was a strong driver of root plate spread in the current study (r = 0.72) and previous studies have observed a correlation between greater root plate spread and lodging resistance in wheat and corn (Berry et al., 2003c; Hostetler et al., 2022). Despite the strong correlation between crown root angle and root plate spread, no correlation between root plate spread and lodging was observed. The importance of root angle to lodging was also confirmed in the SEM, with a direct negative path between the two traits being necessary for the final model. It was also interesting to note that root angle displayed an indirect path to lodging, via a negative path to height. This path was required to produce a good fitting model and indicated that narrower root systems were associated with taller plants. This relationship may hint at an underlying genetic basis, for example, Bai et al. (2013) identified overlapping QTL in wheat that controlled plant height, as well as root volume and root length. Identifying two paths to lodging from root angle emphasizes the importance of this trait to lodging resistance. Notably, root angles were highly heritable not only in this study, but also in sorghum and corn (Lopez et al., 2017; Wang et al., 2021). Thus, root angle would be a suitable selection target for lodging resistance at early stages of breeding programs, potentially by using controlled environments facilities or greenhouses. For example, wheat seedling root traits, which can act as surrogate predictors of important adult root traits, have been assessed using growth pouches (Richard et al., 2015).
Root system solidity, which describes the ratio between network area (i.e. total root tissue) and convex area (i.e. smallest polygon containing the root system), exhibited correlations to several root systems traits such as root angle, root spread, root system width and as expected, convex area. Such correlations between various root traits and solidity have been observed previously in wheat, soybean, strawberries, Arabidopsis, and alfalfa (Cockerton et al., 2020; Deja-Muylle et al., 2022; Falk et al., 2020; Mattupalli et al., 2019; Shao et al., 2021). However, the most relevant and novel correlation that solidity displayed was to lodging (r = 0.62), a finding that no prior studies have described.
Solidity was also an important component of the SEM. A direct positive path from solidity to lodging was observed, while strong paths from network area and convex area to solidity were important contributors to the model. The finding that solidity represents a central trait describing root system architecture and the distribution of biomass across soil space in both the correlation analysis and SEM was not surprising. Solidity has been described as a measure of soil exploration efficiency comparing compact versus exploratory architectures and reflects the trade-off between foraging in-place behavior and space exploration (Bontpart et al., 2020; Mattupalli et al., 2019). In the present study, cultivars with a wider root angle showed correlations to both convex area and solidity (r = 0.43 and -0.57, respectively), indicting that root angle plays an important role in setting the overall outer dimensions of the root system (or soil volume the root system occupies). Wider root angle promoted lower solidity and a more exploratory architecture with increased root plate spread, hence why lower solidity is related to lodging resistance. The strong paths from convex area and network area to solidity captured by the SEM reflect the traits used to calculate solidity, while the strong path from total root length to network area connect two traits related to the size of a root system. Although solidity has not been identified previously as being related to lodging, total root length (Wu and Ma, 2018) and root volume (Wu and Ma, 2018; Zhang et al., 2022) have been associated with increased lodging resistance in maize and canola.
The correlation between solidity and lodging, the inclusion of solidity in the SEM, and the moderate heritability of solidity positions this trait as a promising candidate for barley breeders to use for selections. Moderate to high heritability values for solidity have been reported in corn and Brachypodium (Ingram et al., 2012; Shao et al., 2021). Findings in corn and rice have demonstrated that solidity can remain relatively constant over time as a means of controlling root density and maintaining the ratio of root mass to root system size and the space it occupies (Topp et al., 2013; Zurek et al., 2015). However, it should be noted that differences in soil fertility (e.g., phosphorus or nitrogen) have been shown to impact solidity in sorghum and canola and thus can influence heritability (Parra-Londono et al., 2018; Qin et al., 2019). Thus, future assessments of solidity across different soil fertility levels are warranted.
4.3 Summary
Among the 18 traits evaluated in this study, plant height, root angle and solidity were consistently observed in both correlation analysis and SEMs to influence lodging. Importantly, this study is the first to establish in barley that root angle is an important trait relevant to lodging resistance, and is the first report that root system solidity is also a key lodging resistance trait. The high heritability of both plant height and root angle indicates they could be incorporated into earlier generation selection strategies by barley breeders while solidity, with its moderate heritability and somewhat lower throughput, could be utilized at later generations when lower numbers of genotypes are being assessed. Predictive modelling for root traits using simple regression or machine learning algorithms could be used to rigorously assess potential parental germplasm prior to crosses and expansion into large-scale field trials within a breeding program. Exploring the possibility of rapidly assessing root traits, such as angle or solidity, in a high-throughput indoor environment is a future goal towards increasing both time and cost efficiencies for lodging selections. It was also revealed that stem-related traits, such as stem strength, diameter or wall thickness, and plant bending resistance (i.e. force per plant), which have been shown to influence lodging in other barley growing regions or in other crops, demonstrated no significant predictive value for barley lodging resistance in western Canadian environments.
This study confirmed previous assertions that decreased height and increased root spread, demonstrated here through increased root angle, contribute to lodging resistance and lends credence to the use of root crown imaging to assess the relationship of root architectural features with complex phenomenon such as lodging. Collectively, this study offers breeders avenues to increase lodging resistance especially in the absence of visible lodging in field trials during germplasm development.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
MWT: Data curation, Formal analysis, Investigation, Visualization, Writing – original draft, Writing – review & editing. CAMF: Data curation, Investigation, Writing – review & editing. SY: Investigation, Methodology, Writing – review & editing. XY: Investigation, Methodology, Writing – review & editing. JAF: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing, Writing – original draft. ADB: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – review & editing, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Funding was provided through an Agriculture Development Fund project funded by the Government of Saskatchewan (20190282), Western Grains Research Foundation (VarD2011), Saskatchewan Barley Development Commission (SaskBarley, 5078), Alberta Barley (R2-0155), and Saskatchewan Cattlemen's Association (2019-124) with in-kind contributions from the National Research Council of Canada and University of Saskatchewan. MT was, in part, supported with graduate scholarships from SaskBarley from 2020-2022.
Acknowledgments
We would like to thank the University of Saskatchewan’s Barley and Oat field crew for their years of commitment in collecting field data. We are also very grateful to the laboratory staff at the National Research Council of Canada (NRC) for their help in collection of root data and training of graduate students and the plant growth facility at the NRC for contributions to plant care. We would lastly like to thank the academic committee of Michael Taylor for their hard work on the thesis on which this paper is based.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1547207/full#supplementary-material
References
Bai, C., Liang, Y., and Hawkesford, M. J. (2013). Identification of QTLs associated with seedling root traits and their correlation with plant height in wheat. J. Exp. Bot. 64, 1745–1753. doi: 10.1093/jxb/ert041
Baker, C. J., Sterling, M., and Berry, P. (2014). A generalised model of crop lodging. J. Theor. Biol. 363, 1–12. doi: 10.1016/j.jtbi.2014.07.032
Baron, M. and Kenny, D. (1986). The moderator-mediator variable distinction in social psychological research: Conceptual, strategic, and statistical considerations. J. Pers. Soc Pschol. 51, 1173–1182. doi: 10.1037/0022-3514.51.6.1173
Bayer Crop Science (2021). True eyespot. Available online at: https://cropscience.bayer.co.uk/threats/diseases/wheat-diseases/true-eyespot/ (Accessed March 18, 2021).
Beaton, J. D. and Sekhon, G. S. (1985). “Potassium nutrition of wheat and other small grains,” in Potassium in Agriculture. Ed. Munson, R. D. (ASA, CSSA, SSSA, Madison, WI), 701–752. doi: 10.2134/1985.potassium
Begović, L., Abičić, I., Lalić, A., Lepeduš, H., Cesar, V., and Leljak-Levanić, D. (2018). Lignin synthesis and accumulation in barley cultivars differing in their resistance to lodging. Plant Physiol. Biochem. 133, 142–148. doi: 10.1016/j.plaphy.2018.10.036
Berry, P. (1998). Predicting lodging in winter wheat. University of Nottingham, England. Available at: https://eprints.nottingham.ac.uk/13494/1/266918.pdf (Accessed May 07, 2025).
Berry, P. M., Spink, J. H., Gay, A. P., and Craigon, J. (2003a). A comparison of stem and root lodging risks among winter wheat cultivars. J. Agric. Sci. 141, 191–202. doi: 10.1017/S002185960300354X
Berry, P. M., Spink, J., Sterling, M., and Pickett, A. A. (2003b). Methods for rapidly measuring the lodging resistance of wheat cultivars. J. Agron. Crop Sci. 189, 390–401. doi: 10.1046/j.0931-2250.2003.00062.x
Berry, P. M., Sterling, M., Baker, C. J., Spink, J., and Sparkes, D. L. (2003c). A calibrated model of wheat lodging compared with field measurements. Agric. For. Meteorol. 119, 167–180. doi: 10.1016/S0168-1923(03)00139-4
Berry, P. M., Sterling, M., and Mooney, S. J. (2006). Development of a model of lodging for barley. J. Agron. Crop Sci. 192, 151–158. doi: 10.1111/j.1439-037X.2006.00194.x
Berry, P. M., Sterling, M., Spink, J. H., Baker, C. J., Sylvester-Bradly, R., Mooney, S. J., et al. (2004). Understanding and reducing lodging in cereals. Adv. Agron. 84, 217–271. doi: 10.1016/S0065-2113(04)84005-7
Berry, P. M., Sylvester-Bradley, R., and Berry, S. (2007). Ideotype design for lodging-resistant wheat. Euphytica 154, 165–179. doi: 10.1007/s10681-006-9284-3
Bontpart, T., Concha, C., Giuffrida, M. V., Robertson, I., Admkie, K., Degefu, T., et al. (2020). Affordable and robust phenotyping framework to analyse root system architecture of soil-grown plants. Plant J. 103, 2330–2343. doi: 10.1111/tpj.14877
Braumann, I., Urban, W., Preuẞ, A., Dockter, C., Zakhrabekova, S., and Hansson, M. (2018). Semi-dwarf barley (Hordeum vulgare L.) brh2 and ari-l mutants are deficient in a U-box E3 ubiquitin ligase. Plant Growth Regul. 86, 223–234. doi: 10.1007/s10725-018-0423-3
Brune, P. F., Baumgarten, A., McKay, S. J., Technow, F., and Podhiny, J. J. (2017). A biomechanical model for maize root lodging. Plant Soil 422, 397–408. doi: 10.1007/s11104-017-3457-9
Casserly, L. M. (1956). The effect of nitrogen, phosporous and potassium on lodging in oats. Can. J. Plant Sci. 37, 245–251. doi: 10.4141/cjps57-029
Chauhan, S., Darvishzadeh, R., Lu, Y., Stroppiana, D., Boschetti, M., Pepe, M., et al. (2019). Wheat lodging assessment using multispectral UAV data. Int. Arch. Photogramm. Remote Sens. Spatial Inf. Sci. 42, 235–240. doi: 10.5194/isprs-archives-XLII-2-W13-235-2019
Clarkson, J. (1981). Relationship between eyespot severity and yield loss in winter wheat. Plant Path. 30, 125–131. doi: 10.1111/j.1365-3059.1981.tb01243.x
Cockerton, H. M., Li, B., Stavridou, E., Johnson, A., Karlström, A., Armitage, A. D., et al. (2020). Genetic and phenotypic associations between root architecture, arbuscular mycorrhizal fungi colonization and low phosphate tolerance in strawberry (Fragaria × ananassa). BMC Plant Biol. 20, 154. doi: 10.1186/s12870-020-02347-x
Cohen, J. (1988). Statistical power analysis for the behavioral sciences. 2nd Ed. (Hillsdale, NJ: Lawrence Erlbaum Associates).
Cooper, M., DeLacy, I. H., and Basford, K. E. (1994). “Relationships among analytical methods used to analyse genotypic adaptation in multi-environment trials,” in Plant Adaptation and Crop Improvement. Eds. Cooper, M. and Hammer, G. L. (CAB International, Wallingford, UK), 193–224.
Crook, M. J. and Ennos, A. R. (1993). The mechanics of root lodging in winter wheat, Triticum aestivum L. J. Exp. Bot. 44, 1219–1224. doi: 10.1093/jxb/44.7.1219
Crook, M. J. and Ennos, A. R. (1994). Stem and root characteristics associated with lodging resistance in four winter wheat cultivars. J. Agric. Sci. 123, 167–174. doi: 10.1017/S0021859600068428
Deja-Muylle, A., Opdenacker, D., Parizot, B., Motte, H., Lobet, G., Storme, V., et al. (2022). Genetic variability of Arabidopsis thaliana mature root system architecture and genome-wide association study. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.814110
Dreccer, M. F., Condon, A. G., Macdonald, B., Rebetzke, G. J., Awasi, M.-A., Borgognone, M. G., et al. (2020). Genotypic variation for lodging tolerance in spring wheat: wider and deeper root plates, a feature of low lodging, high yielding germplasm. Field Crops Res. 258, 107942. doi: 10.1016/j.fcr.2020.107942
Dyson, D. H., Goettel, C., Helm, J. H., and Stewart, W. M. (1984). Barley yield losses due to lodging in central Alberta. Barley Newsl. 28, 59–60.
Easson, D. L., White, E. M., and Pickles, S. J. (1993). The effects of weather, seed rate and cultivar on lodging and yield in winter wheat. J. Agric. Sci. 121, 145–156. doi: 10.1017/S0021859600077005
Erndwein, L., Cook, D. D., Robertson, D. J., and Sparks, E. E. (2020). Field-based mechanical phenotyping of cereal crops to assess lodging resistance. Appl. Plant Sci. 8, e11382. doi: 10.1002/aps3.11382
Falk, K. G., Jubery, T. Z., Mirnezami, S. V., Parmley, K. A., Sarkar, S., Singh, A., et al. (2020). Computer vision and machine learning enabled soybean root phenotyping pipeline. Plant Methods 16, 5. doi: 10.1186/s13007-019-0550-5
Gavloski, J. (2019). Cutworms in field crops. Province of Manitoba, Agriculture. Available online at: http://www.manitoba.ca/agriculture/crops/insects/print,cutworms-field-crops.html (Accessed November 27, 2023).
Gianinetti, A. and Baronchelli, M. (2024). The pulvinus is the weak point for stem lodging resistance in ripe barley. Plants 13, 3172. doi: 10.3390/plants13223172
Gill, K. S., Omokanye, A. T., Pettyjohn, J. P., and Elsen, M. (2013). Evaluation of forage type barley varieties for forage yield and nutritive value in the Peace Region of Alberta. J. Agric. Sci. 5, 24–36. doi: 10.5539/jas.v5n2p24
Grossman, J. D. and Rice, K. J. (2012). Evolution of root plasticity responses to variation in soil nutrient distribution and concentration. Evol. Appl. 5, 850–857. doi: 10.1111/j.1752-4571.2012.00263.x
Hai, L., Guo, H., Xiao, S., Jiang, G., Zhang, X., Yan, C., et al. (2005). Quantitative trait loci (QTL) of stem strength and related traits in a doubled-haploid population of wheat (Triticum aestivum L.). Euphytica. 141, 1–9. doi: 10.1007/s10681-005-4713-2
Hebert, Y., Barriere, Y., and Bertholeau, J. C. (1992). Root lodging resistance in forage maize: genetic variability of root system and aerial part. Maydica 37, 173–183.
Hedden, P. (2003). The genes of the green revolution. Trends Genet. 19, 5–9. doi: 10.1016/S0168-9525(02)00009-4
Heuschele, D. J., Garcia, T. A., Ortiz, J. B., Smith, K. P., and Marchetto, P. (2020). Cereal stem stress: In situ biomechanical characterization of stem elasticity. Appl. Sci. 10, 7965. doi: 10.3390/app10227965
Heuschele, D. J., Wiersma, J., Reynolds, L., Mangin, A., Lawley, Y., and Marchetto, P. (2019). The Stalker: An open-source force meter for rapid stalk strength phenotyping. HardwareX 6, E000067. doi: 10.1016/j.ohx.2019.e00067
Hobbs, P. R., Sayre, K. D., and Ortiz-Monasterio, J. I. (1998). Increasing Wheat Yields Sustainably Through Agronomic Means (Mexico: CIMMYT). doi: 10.22004/ag.econ.232958
Hostetler, A. N., Erndwein, L., Reneau, J. W., Stager, A., Tanner, H. G., Cook, D., et al. (2022). Multiple brace root phenotypes promote anchorage and limit root lodging in maize. Plant Cell Environ. 45, 1573–1583. doi: 10.1111/pce.14289
Ingram, P. A., Zhu, J., Shariff, A., Davis, I. W., Benfey, P. N., and Elich, T. (2012). High-throughput imaging and analysis of root system architecture in Brachypodium distachyon under differential nutrient availability. Philos. Trans. R. Soc Lond. B. Biol. Sci. 367, 1559–1569. doi: 10.1098/rstb.2011.0241
Jaff, D. (2019). Straw stiffness in landraces of two-row barley (Hordeum vulgare L.). Iraqi J. Agric. Sci. 50, 1597–1604. doi: 10.36103/ijas.v50i6.849
Jaff, D. and Jarvis, M. C. (2021). Straw bending and lodging in landrace barley: Finite element modelling. Sarhad J. Agric. 37, 555–563. doi: 10.17582/journal.sja/2021/37.2.555.563
Jung, J. (1984). “Plant bioregulators in cereal crops: action and uses,” in Bioregulators: chemistry and uses. Eds. Ory, R. L. and Rittig, F. R. (American Chemical Society, Washington), 29–43. doi: 10.1021/bk-1984-0257.ch004
Kamran, M., Ahmad, I., Wu, X., Liu, T., Ding, R., and Han, Q. (2018a). Application of paclobutrazol: a strategy for inducing lodging resistance of wheat through mediation of plant height, stem physical strength, and lignin biosynthesis. Environ. Sci. pollut. Res. 25, 29366–29378. doi: 10.1007/s11356-018-2965-3
Kamran, M., Cui, W., Ahmad, I., Meng, X., Zhang, X., Su, W., et al. (2018b). Effect of paclobutrazol, a potential growth regulator on stalk mechanical strength, lignin accumulation and its relation with lodging resistance of maize. Plant Growth Regul. 84, 317–332. doi: 10.1007/s10725-017-0342-8
Kashiwagi, T., Togawa, E., Hirotsu, N., and Ishimaru, K. (2008). Improvement of lodging resistance with QTLs for stem diameter in rice (Oryza sativa L.). Theor. Appl. Genet. 117, 749–757. doi: 10.1007/s00122-008-0816-1
Kassambara, A. (2019). ggcorrplot: Visualization of a correlation matrix using ‘ggplot2’. R package version 0.1.4.999. Available online at: https://github.com/kassambara/ggcorrplot (Accessed May 07, 2025).
Kline, R. B. (2015). Principles and Practice of Structural Equation Modeling. 4th Ed (New York: Guildford Press).
Kristensen, P. S., Dockter, C., Lundqvist, U., Lu, Q., Gregersen, P. L., Thordal-Christensen, H., et al. (2016). Genetic mapping of the barley lodging resistance locus Erectoids-k. Plant Breed. 135, 420–428. doi: 10.1111/pbr.12377
Kuczyńska, A., Surma, M., Adamski, T., Mikołajczak, K., Krystkowiak, K., and Ogrodowicz, P. (2013). Effects of the semi-dwarfing Sdw1/denso gene in barley. J. Appl. Genet. 54, 381–390. doi: 10.1007/s13353-013-0165-x
Laidig, F., Feike, T., Klocke, B., Macholdt, J., Miedaner, T., Rentel, D., et al. (2022). Yield reduction due to diseases and lodging and impact of input intensity on yield in variety trials in five cereal crops. Euphytica 218, 150. doi: 10.1007/s10681-022-03094-w
Lamb, E., Shirtliffe, S., and May, W. (2011). Structural equation modeling in the plant sciences: An example using yield components in oat. Can. J. Plant Sci. 91, 603–619. doi: 10.4141/cjps2010-035
Li, C., Li, W., Luo, Y., Jin, M., Chang, Y., Cui, H., et al. (2023). Mixed cropping increases grain yield and lodging resistance by improving the canopy light environment of wheat populations. Eur. J. Agron. 147, 126849. doi: 10.1016/j.eja.2023.126849
Li, B., Luo, Y., Guo, C., Yang, Y., Yuan, X., Xing, M., et al. (2022). Effects of wheat straw returning and potassium application rates on the physicochemical properties and lodging resistance of different stem internodes in direct-seeded rice. Int. Agrophys. 36, 309–321. doi: 10.31545/intagr/155271
Liao, P., Bell, S. M., Chen, L., Huang, S., Wang, H., Miao, J., et al. (2023). Improving rice grain yield and reducing lodging risk simultaneously: a meta-analysis. Eur. J. Agron. 143, 126709. doi: 10.1016/j.eja.2022.126709
LMC International (2023). The economic impact of barley on the Canadian economy: 2022. Available online at: https://cerealsCanada.ca/economic-impact/ (Accessed May 07, 2025).
Lopez, J. R., Erickson, J. E., Munoz, P., Saballos, A., Felderhoff, T. J., and Vermerris, W. (2017). QTLs associated with crown root angle, stomatal conductance, and maturity in sorghum. Plant Genome 10, 10.3835. doi: 10.3835/plantgenome2016.04.0038
Lynch, J. (1995). Root architecture and plant productivity. Plant Physiol. 109, 7–13. doi: 10.1104/pp.109.1.7
Ma, B. L. and Smith, D. L. (1992a). Growth regulator effects on aboveground dry matter partitioning during grain fill of spring barley. Crop Sci. 32, 741–746. doi: 10.2135/cropsci1992.0011183X003200030033x
Ma, B. L. and Smith, D. L. (1992b). Modification of tiller productivity in spring barley by application of chlormequat or ethephon. Crop Sci. 32, 735–740. doi: 10.2135/cropsci1992.0011183X003200030032x
Ma, J. F. and Yamaji, N. (2006). Silicon uptake and accumulation in higher plants. Trends Plant Sci. 11, 392–397. doi: 10.1016/j.tplants.2006.06.007
Madić, M., Knezevic, D., Paunovic, A., and Durovic, D. (2016). Plant height and internode length as components of lodging resistance in barley. Acta Agric. Serb. 21, 99–106. doi: 10.5937/AASer1642099M
Mangin, A., Heuschele, J., Brûlé-Babel, A., Flaten, D., Wiersma, J., and Lawley, Y. (2022). Rapid in situ nondestructive evaluation of lodging risk in dryland agronomic wheat research. Agron. J. 114, 2822–2829. doi: 10.1002/agj2.21173
Mattupalli, C., Seethepalli, A., York, L. M., and Young, C. A. (2019). Digital imaging to evaluate root system architectural changes associated with soil biotic factors. Phyto. J. 3, 102–111. doi: 10.1094/PBIOMES-12-18-0062-R
Melis, M. and Farina, M. P. W. (1984). Potassium effects on stalk strength, premature death and lodging of maize (Zea mays L.). S. Afr. J. Plant Soil. 1, 122–124. doi: 10.1080/02571862.1984.10634125
Mickelson, H. R. and Rasmusson, D. C. (1994). Genes for short stature in barley. Crop Sci. 34, 1180–1183. doi: 10.2135/cropsci1994.0011183X003400050007x
Nakajima, T., Yoshida, M., and Tomimura, K. (2008). Effect of lodging on the level of mycotoxins in wheat, barley, and rice infected with the Fusarium graminearum species complex. J. Gen. Plant Pathol. 74, 289–295. doi: 10.1007/s10327-008-0103-7
Niu, Y., Chen, T., Zhao, C., and Zhou, M. (2022). Lodging prevention in cereals: Morphological, biochemical, anatomical traits and their molecular mechanisms, management and breeding strategies. Field Crops Res. 289, 108733. doi: 10.1016/j.fcr.2022.108733
Niu, L., Feng, S., Ding, W., and Li, G. (2016). Influence of speed and rainfall on large-scale wheat lodging from 2007 to 2014 in China. PloS One 11, e0157677. doi: 10.1371/journal.pone.0157677
Niu, L., Feng, S., Ru, Z., Li, G., Zhang, Z., and Wang, Z. (2012). Rapid determination of single-stalk and population lodging resistance strengths and an assessment of the stem lodging wind speeds for winter wheat. Field Crops Res. 139, 1–8. doi: 10.1016/j.fcr.2012.10.014
O’Donovan, J. T., Turkington, T. K., Edney, M. J., Clayton, G. W., McKenzie, R. H., Juskiw, P. E., et al. (2011). Seeding rate, nitrogen rate, and cultivar effects on malting barley production. Agron. J. 103, 709–716. doi: 10.2134/agronj2010.0490
O’Donovan, J. T., Turkington, T. K., Edney, M. J., Juskiw, P. E., McKenzie, R. H., Harker, K. N., et al. (2012). Effect of seeding date and seeding rate on malting barley production in western Canada. Can. J. Plant Sci. 92, 321–330. doi: 10.4141/cjps2011-130
Ookawa, T., Hobo, T., Yano, M., Murta, K., Ando, T., Miura, H., et al. (2010). New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 1, 1–11. doi: 10.1038/ncomms1132
Parra-Londono, S., Kavka, M., Samans, B., Snowdon, R., Wieckhorst, S., and Uptmoor, R. (2018). Sorghum root-system classification in contrasting P environments reveals three main rooting types and root-architecture-related marker–trait associations. Ann. Bot. 121, 267–280. doi: 10.1093/aob/mcx157
Piñera-Chavez, F. J., Berry, P. M., Foulkes, M. J., Jesson, M. A., and Reynolds, M. P. (2016b). Avoiding lodging in irrigated spring wheat. I. Stem and root structural requirements. Field Crops Res. 196, 325–336. doi: 10.1016/j.fcr.2016.06.009
Piñera-Chavez, F. J., Berry, P. M., Foulkes, M. J., Molero, G., and Reynolds, M. P. (2016a). Avoiding lodging in irrigated spring wheat. II. Genetic variation of stem and root structural properties. Field Crops Res. 196, 64–74. doi: 10.1016/j.fcr.2016.06.007
Posit Team (2025). RStudio: Integrated development for R (Boston, MA: RStudio, PBC). Available online at: http://www.rstudio.com/ (Accessed May 07, 2025).
Qin, L., Walk, T. C., Han, P., Chen, L., Zhang, S., Li, Y., et al. (2019). Adaption of roots to nitrogen deficiency revealed by 3D quantification and proteomic analysis. Plant Physiol. 179, 329–347. doi: 10.1104/pp.18.00716
Rajala, A. and Peltonen-Sainio, P. (2002). Timing applications of growth regulators to alter spring cereal development at high latitudes. Agric. Food Sci. 11, 233–244. doi: 10.23986/afsci.5721
R Core Team (2021). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed May 07, 2025).
Reynolds, M., Foulkes, M. J., Slafer, G. A., Berry, P., Parry, M. A. J., Snape, J. W., et al. (2009). Raising yield potential in wheat. J. Exp. Bot. 60, 1899–1918. doi: 10.1093/jxb/erp016
Richard, C. A., Hickey, L. T., Fletcher, S., Jennings, R., Chenu, K., and Christopher, J. T. (2015). High-throughput phenotyping of seminal root traits in wheat. Plant Methods 11, 13. doi: 10.1186/s13007-015-0055-9
Rosseel, Y., Jorgensen, T. D., De Wilde, L., Oberski, D., Byrnes, J., Vanbrabant, L., et al. (2022). Lavaan: latent variable analysis. R package version 0.6-12. Available online at: https://cran.r-project.org/web/packages/lavaan/index.html (Accessed November 27, 2023).
Rueden, C. T., Schindelin, J., Hiner, M. C., DeZonia, B. E., Walter, A. E., Arena, E. T., et al. (2017). ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinf. 18, 529. doi: 10.1186/s12859-017-1934-z
Sakamoto, T., Shibayama, M., Takada, E., Akihiro, I., Morita, K., Takahashi, W., et al. (2010). Detecting seasonal changes in crop community structure using day and night digital images. Photogramm. Eng. Remote Sens. 76, 713–726. doi: 10.14358/PERS.76.6.713
Seethepalli, A., Dhakal, K., Griffiths, M., Guo, H., Freschet, G. T., and York, L. M. (2021). RhizoVision Explorer: open-source software for root image analysis and measurement standardization. Ann. Bot. 13, plab056. doi: 10.1093/aobpla/plab056
Shao, M. R., Jiang, N., Li, M., Howard, A., Lehner, K., Mullen, J. L., et al. (2021). Complementary phenotyping of maize root system architecture by root pulling force and X-ray imaging. Plant Phenom. 2021, 9859254. doi: 10.34133/2021/9859254
Slafer, G. A., Foulkes, M. J., Reynolds, M. P., Murchie, E. H., Carmo-Silva, E., Flavell, R., et al. (2023). A ‘wiring diagram’ for sink strength traits impacting wheat yield potential. J. Exp. Bot. 74, 40–71. doi: 10.1093/jxb/erac410
Sparks, E. E. (2022). Maize plants and the brace roots that support them. New. Phytol. 237, 48–52. doi: 10.1111/nph.18489
Stanca, A. M., Jenkins, G., and Hanson, P. R. (1979). Varietal responses in spring barley to natural and artificial lodging and to a growth regulator. J. Agric. Sci. 93, 449–456. doi: 10.1017/S0021859600038144
Statistics Canada (2025a). Estimated areas, yield, production, average farm price and total farm value of principal field crops, in metric and imperial units. Available online at: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210035901 (Accessed May 07, 2025).
Statistics Canada (2025b). Farm product prices, crops and livestock. Available online at: https://www150.statcan.gc.ca/t1/tbl1/en/tv.action?pid=3210007701 (Accessed May 07, 2025).
Sterling, M., Barker, C. J., Berry, P. M., and Wade, A. (2003). An experimental investigation of the lodging of wheat. Agric. For. Meteorol. 119, 149–165. doi: 10.1016/S0168-1923(03)00140-0
Tidemann, B. D., O’Donovan, J. T., Izydarczyk, M., Turkington, T. K., Oatway, L., Beres, E., et al. (2020). Effects of plant growth regulator applications on malting barley in western Canada. Can. J. Plant Sci. 100, 653–665. doi: 10.1139/cjps-2019-0200
Topp, C. N., Iyer-Pascuzzi, A. S., Anderson, J. T., Lee, C.-R., Zurek, P. R., Symonova, O., et al. (2013). 3D phenotyping and quantitative trait locus mapping identify core regions of the rice genome controlling root architecture. Proc. Natl. Acad. Sci. U.S.A 110, E1695–E1704. doi: 10.1073/pnas.1304354110
United States Department of Agriculture (2025). Production – barley. Available online at: https://www.fas.usda.gov/data/production/commodity/0430000 (Accessed May 07, 2025).
Wang, H., Tang, X., Yang, X., Fan, Y., Xu, Y., Li, P., et al. (2021). Exploiting natural variation in crown root traits via genome-wide association studies in maize. BMC Plant Biol. 21, 346. doi: 10.1186/s12870-021-03127-x
Wickham, H. (2016). ggplot2: Elegant Graphics for Data Analysis. 2nd Ed (New York: Springer-Verlag). doi: 10.1007/978-0-387-98141-3
Wiersma, D. W., Oplinger, E. S., and Guy, S. O. (1986). Environment and cultivar effects on winter wheat response to ethephon plant growth regulator. Agron. J. 78, 761–764. doi: 10.2134/agronj1986.00021962007800050002x
Wilke, N., Siegmann, B., Frimpong, F., Muller, O., Klingbell, L., and Rascher, U. (2019). Quantifying lodging percentage, lodging development and lodging severity using a UAV-based canopy height model. Int. Arch. Photogramm. Remote Sens. Spatial Inf. Sci. 42, 649–655. doi: 10.1371/journal.pone.0090144
World Bank (2025). Cereals: barley exports by country in 2023. Available online at: https://wits.worldbank.org/trade/comtrade/en/country/ALL/year/2023/tradeflow/Exports/partner/WLD/product/100300 (Accessed May 07, 2025).
Wu, W. and Ma, B.-L. (2018). Assessment of canola crop lodging under elevated temperatures for adaptation to climate change. Agric. Meteorol. 248, 455–465. doi: 10.1016/j.agrformet.2017.09.017
Wu, W., Ma, B.-L., Fan, J.-J., Sun, M., Yi, Y., Guo, W.-S., et al. (2019). Management of nitrogen fertilization to balance reducing lodging risk and increasing yield and protein content in spring wheat. Field Crops Res. 241, 107584. doi: 10.1016/j.fcr.2019.107584
Xia, Y. and Yang, Y. (2018). RMSEA, CFI and TLI in structural equation modelling with ordered categorical data: The story they tell depends on the estimation methods. Behav. Res. Methods 51, 409–428. doi: 10.3758/s13428-018-1055-2
Xiao, Y., Liu, J., Li, H., Cao, X., Xia, X., and He, Z. (2015). Lodging resistance and yield potential of winter wheat: effect of planting density and genotype. Front. Agr. Sci. Eng. 2, 168–178. doi: 10.15302/J-FASE-2015061
Yao, J., Ma, H., Pingping., Z., Ren, L., Yang, X., Yao, G., et al. (2011). Inheritance of stem strength and its correlations with culm morphological traits in wheat (Triticum aestivum L.). Can. J. Plant Sci. 91, 1065–1070. doi: 10.4141/cjps2011-033
Zhang, H.-J., Li, T., Liu, H.-W., Mai, C.-Y., Yu, G.-J., Li, H.-L., et al. (2020). Genetic progress in stem lodging resistance of the dominant wheat cultivars adapted to Yellow-Huai River Valleys Winter Wheat Zone in China since 1964. J. Integr. Agric. 19, 438–448. doi: 10.1016/S2095-3119(19)62627-4
Zhang, Y., Xu, W., Wang, H., Fang, Y., Dong, H., and Qi, X. (2016). Progress in improving stem lodging resistance of Chinese wheat cultivars. Euphytica. 212, 275–286. doi: 10.1007/s10681-016-1768-1
Zhang, P., Yan, Y., Gu, S., Wang, Y., Xu, C., Sheng, D., et al. (2022). Lodging resistance in maize: A function of root-shoot interactions. Eur. J. Agron. 132, 126393. doi: 10.1016/j.eja.2021.126393
Zheng, M., Chen, J., Shi, Y., Li, Y., Yin, Y., Yang, D., et al. (2017). Manipulation of lignin metabolism by plant densities and its relationship with lodging resistance in wheat. Sci. Rep. 7, 41805. doi: 10.1038/srep41805
Keywords: barley, lodging, root system architecture, crown root angle, solidity, plant height
Citation: Taylor MW, Ferré CAM, Ye S, Yang X, Feurtado JA and Beattie AD (2025) Validating the contributions of stem and root traits to barley lodging in western Canada. Front. Plant Sci. 16:1547207. doi: 10.3389/fpls.2025.1547207
Received: 17 December 2024; Accepted: 06 June 2025;
Published: 28 August 2025.
Edited by:
Francis Chuks Ogbonnaya, Grains Research and Development Corporation, AustraliaCopyright This work is authored by Taylor, Ferré, Ye, Yang, Feurtado and Beattie. © 2025 His Majesty the King in Right of Canada and Taylor, Ferré, Beattie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: J. Allan Feurtado, YWxsYW4uZmV1cnRhZG9AbnJjLWNucmMuZ2MuY2E=; Aaron D. Beattie, YWFyb24uYmVhdHRpZUB1c2Fzay5jYQ==