 Agustina Rosa Andrés-Hernández
Agustina Rosa Andrés-Hernández Ernesto C. Rodríguez-Ramírez
Ernesto C. Rodríguez-Ramírez- 1Facultad de Ciencias Biológicas, Benemérita Universidad Autónoma de Puebla, Puebla, Pue, Mexico
- 2Laboratorio de Dendrocronología, Universidad Continental, Huancayo, Junín, Peru
Introduction: Tropical montane cloud forests (TMCFs) host specialized plant species reliant on persistent atmospheric humidity, including fog immersion obligates and relict assemblages. Understanding anatomical and morphological adaptations in TMCF woody angiosperms is critical for elucidating their acclimation strategies to hydric stress under shifting fog regimes. This study investigates interspecific variability in wood and leaf traits among 10 TMCF tree species in Mexico’s Medio Monte Natural Protected Area, hypothesizing that distinct anatomical strategies emerge in response to climatic stressors.
Methods: Wood anatomical (e.g., vessel density, hydraulic diameter, fiber length) and leaf morphological traits (e.g., lamina length, vein density, leaf organization) were analyzed across species. Traits were correlated with climatic variables—mean maximum/minimum temperatures, monthly precipitation, and evapotranspiration—to identify adaptive patterns. Statistical analyses quantified interspecific differences and assessed trait-climate relationships.
Results: Significant interspecific divergence occurred in both wood and leaf traits. Wood anatomy was strongly influenced by mean maximum temperature, precipitation, and evapotranspiration, affecting vessel density, vulnerability index, ray dimensions, and fiber length. Leaf traits correlated with temperature extremes and evapotranspiration, driving variation in leaf size, apex/base morphology, venation complexity, and marginal teeth. Notably, hydraulic efficiency (e.g., wider vessels) aligned with higher precipitation, while drought-associated traits (e.g., denser veins) linked to elevated temperatures.
Discussion: TMCF species exhibit trait-based strategies balancing hydraulic safety and efficiency, reflecting niche partitioning under microclimatic gradients. Temperature and water availability differentially shape wood and leaf adaptations, with vessel architecture and venation patterns acting as key regulators of water loss. These findings underscore the functional diversity of TMCF trees and their capacity to acclimate to environmental variability. Conservation efforts must prioritize microclimate preservation to safeguard these adaptive traits amid climate change.
1 Introduction
Tropical Montane Cloud Forests (TMCFs; sensu Bruijnzeel et al., 2010) exhibit substantial biodiversity and contribute significantly to essential ecosystem services in hotspot mountainous areas (Gentry, 1992; Fahey et al., 2016; Richter et al., 2024). The adaptability of TMCF plants to hydric gradients is determined by the frequency of fog, mist and potential evapotranspiration (Bruijnzeel et al., 2011; Crausbay et al., 2014). Along this gradient, tree height, density, and forest species diversity are progressively reduced and morpho-physiological acclimation and/or adaptations to resist or avoid hydric stress are more common (Thybring and Fredriksson, 2021; Rodríguez-Ramírez et al., 2022; López-Calvillo et al., 2023). Nonetheless, the occurrence of longer than typical hydric stress seasons or droughts over an extended number of years in TMCFs can result in reduced plant growth, seedling mortality, and large-scale tree mortality (Choat et al., 2018; Rodríguez‐Ramírez et al., 2024a). Climate change poses a significant potential risk to the future survival of TMCFs (Los et al., 2021; Ramírez-Barahona et al., 2025). Projected climate scenarios predict a temperature increase of 4.1-5°C above current levels and an increase in CO2 emissions to 70.04 gigatons for Shared Socioeconomic Pathways (SSP) 3-7.0 and 116.8 gigatons for SSP 5-8.5 (Taylor et al., 2012).
Typically, hydric stress on plants is studied in controlled greenhouses; however, this approach does not consider the complexity of TMCF systems and the responses of relict endemic trees (Eller et al., 2020). Thus, a mechanistic understanding of how TMCF wetness variation determines community composition and function is required in the field to predict TMCF responses to climate change (Andrés-Hernández et al., 2023). Functional traits have often been defined as morphological, physiological and phenological plant characteristics that influence the success of a genotype (individual), which are in turn influenced by environmental interactions and evolutionary processes, environmental interactions and evolutionary processes (Anderegg and Meinzer, 2015; Tng et al., 2018). New evidence about trait variation in plants, such as leaf vein acclimation (Eller et al., 2020), wood anatomy (Argüelles-Marrón et al., 2023) and seed size/mast (Burns, 2012) convinced ecologists that functional traits are the best approach to achieving predictive understanding (McGill et al., 2006; Sobral, 2021; Andrés-Hernández et al., 2023).
In this context, functional traits are useful parameters for studying plant hydric stress strategies and acclimating organ-level changes in plants when they are hydric-stressed, such as during drought or high precipitation (Rodríguez‐Ramírez et al., 2024a). We hypothesized that TMCF tree species display different wood and leaf anatomical adjustments or strategies in response to the air and soil moisture (Guerin et al., 2012; Thybring and Fredriksson, 2021). Similarly, plant trait information can be incorporated into functional trait theory and used to link plant performance to a range of specific climatic events in TMCF tree species (Báez et al., 2022). This has led to several predictions regarding how climatic variation in habitat drives specific functional and hydraulic architectural traits (Weigelt et al., 2021). To help resolve these questions, the main aims of this field study were to: (i) assess wood and leaf trait variability in ten TMCF tree species in situ; (ii) identify quantitative patterns (trait-trait and trait-climate relationships) among tree species; iii) evaluate how traits are influenced by climatic factors; and (iv) explore how climate influences tree species functional traits. Concatenating climate variation and wood- and leaf-anatomy are essential for understanding the growth, development, and adaptation of TMCF tree species under hydric stress (Pandey, 2021; Andrés-Hernández et al., 2023; Rodríguez‐Ramírez et al., 2024b).
2 Materials and methods
2.1 Study site
The study area (longitudes: 98° 15’W; latitude: 20° 24’ N) is the Medio Monte Natural Protected Area (151.63 km2), altitudes ranging from 1558 to 1913 meters above sea level, located in central-eastern Sierra Madre Oriental, Mexico (Figure 1). The study forest exhibits an annual temperature range of 14.5 to 24.4°C and an average annual rainfall of approximately 2600 mm, with the majority occurring between late July and late October (Argüelles-Marrón et al., 2023). The study forest has acidic soils (pH 4–6), specifically andosol-humic and light sandy-loamy (Andrés-Hernández et al., 2023). The canopy (heights ranging from 4 to 40 m) comprises a mixture of Neotropical angiosperms (Quercus spp. Fagus mexicana Martínez, Magnolia schiedeana Schltdl., Alnus jorullensis Kunth, Sambucus nigra L., Symplocos spp., Styrax glabrescens Benth, Tilia mexicana Schltdl., Turpinia insignis (Kunth) Tul, Beilchsmiedia spp., Clethra spp., Liquidambar styraciflua L., Carya spp., and rarely Ulmus mexicana (Liebm.) Planch) and gymnosperms (Pinus patula Schltdl. & Cham.), among other species (Miranda and Sharp, 1950).
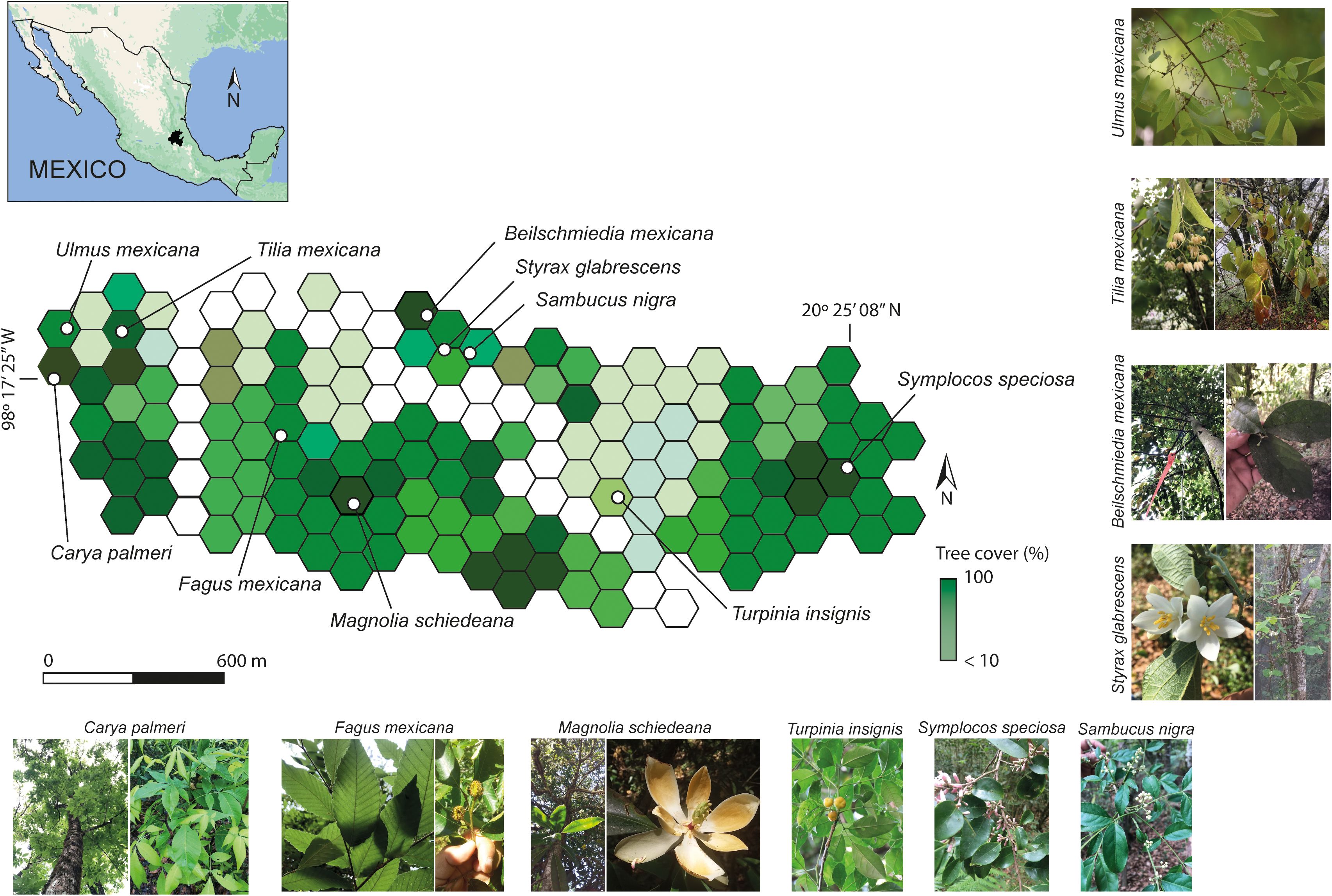
Figure 1. Map of the study area in the Medio Monte Natural Protected Area with hexagonal grids. The color scale represents the proportion of forest canopy cover in each grid (white: low canopy cover; dark green: high canopy cover). The location of the tree species is indicated by dots, and the inset map shows the study area within Mexico (black). Data acquired from https://globalforestwatch.org/map. The Tropical Montane Cloud Forest tree species assessed are represented.
We used mean maximum and minimum temperature (TMAX, TMIN) in °C, vapor pressure deficit (VPD), monthly precipitation (PRE), and evapotranspiration (EVT) in mm from the Climatologies at High resolution for the Earth’s Land Surface Areas database (CHELSA v.2.0; http://chelsa-climate.org/; Karger et al., 2021). The layer resolution was approximately 1 km2 with records from 1980 to 2018. These climatic factors represent key aspects of the TMCF tree growth-climate relationship (Rodriguez-Ramirez et al., 2020a; Argüelles-Marrón et al., 2023).
2.2 Study species
For our study, we sampled ten individuals of each TMCF tree species: Symplocos speciosa Hemsl., Sambucus nigra L, Styrax glabrescens, Turpinia insignis, Magnolia schiedeana, Tilia mexicana, Fagus mexicana, Beilschmiedia mexicana (Mez) Kosterm., Ulmus mexicana, and Carya palmeri W.E. Manning. These species show a high relative abundance within the study area, so we chose them because they are typical of eastern Mexican TMCFs (Luna-Vega et al., 2022).
2.3 Sampling and processing
2.3.1 Wood anatomy
From late December 2022 to early February 2023, exhaustive surveys were conducted to determine the distribution from the most abundant TMCF tree species in the study area (Kindt and Coe, 2005). We selected four individuals of each selected (excluding those exhibiting scars or rot) using a global positioning system (GPS) (Garmin® 101 eTrex 10; Garmin, Olathe, KS, USA). Prior to the beginning of the rainy and growing seasons, we collected four wood samples from trees from each species (Rodríguez-Ramírez et al., 2018). Wood samples were collected by cutting from three to five cm wide piece containing bark and earlywood using a handsaw (Silky Zubat Handsaw, Truper®, CDMX, Mexico), and immediately fixed it in a formalin, acetic acid, and ethanol (FAA; 10:5:85) for 24 h. Histological sections were made in the transverse, longitudinal and radial planes at 25 μm width with the rotary microtome (Leica® 2000R, Wetzlar, Germany). Samples were dehydrated with graded ethyl alcohol from 50% to 96% (v/v). Wood samples were stained with safranin dissolved in 96% alcohol for one hour and fast-green for eight seconds, then washed with 100% ethanol and rinsed with xylol (McCracken and Johansen, 1940). Finally, the samples were mounted on a synthetic resin (Ruzin, 2000). To gather data on vessel elements and fiber lengths, samples were macerated in Jeffrey’s solution as previously described (Berlyn and Miksche, 1976).
Eight anatomical wood traits were measured for each individual. Fifty measurements were taken per individual for each trait related to water transport (xylem vessel density, hydraulic diameter, vulnerability index, vessel grouping index, solitary vessel index, fiber length, length of uniseriate rays, and width of rays; Table 1; Supplementary Figure S1A), related to the moisture content of wood, and these relationships can vary among different tree species and under microclimatic conditions (Scholz et al., 2014; Schuldt et al., 2013; von Arx et al., 2013). The measurements were performed according to IAWA recommendations (IAWA COMMITTEE, 1989).
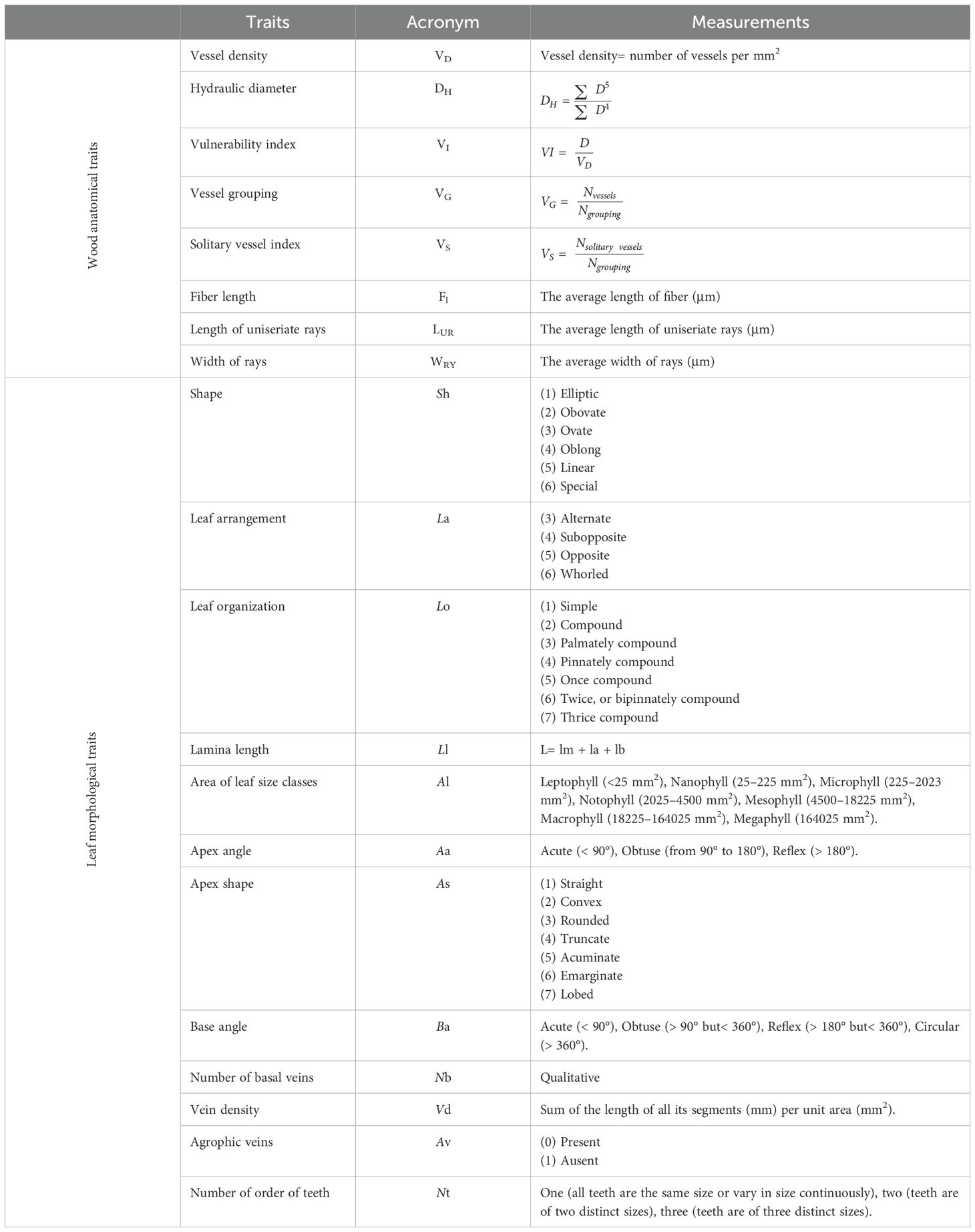
Table 1. Overview of wood and leaf traits, their acronyms, and measurements.
2.3.2 Leaf anatomy
We randomly selected 20 fully mature, healthy and undamaged leaves for each previously selected trees species (see Wood Anatomy section) in the summers (from May to June) of 2023 and 2024 (i.e. when the leaves reached their maximum development; Borchert et al., 2005). The leaves were selected from the basal branches of each tree, to minimize abiotic variation due to their position on the tree, atmospheric humidity levels, and light incidence. After collection, the leaves were placed in plastic bags (40 x 40 cm) with moist paper towels according to the method proposed by Rodríguez-Ramírez et al. (2021a). Leaf samples were transported to the Wood Anatomy Laboratory (Benemérita Universidad Autónoma de Puebla, BUAP, Mexico) within 3 days of collection.
The collected leaves were cleared in a 50% solution (w/v) of Na2CO3 at 85°C for 1 to 2 h. When leaf samples turned bright green, they were removed and rinsed with tap water. The leaves were then washed with a bleach solution 50% (Clorox®) for 45 to 50 min. To remove the epidermis, the leaves were placed in a glass container with bleach solution and tap water (50:50), and then brushed off with a soft marten hairbrush until the samples completely lost their greenness (Rodríguez-Ramírez et al., 2021b).
We digitized the 20 cleared leaves collected from each species. Each leaf was placed directly on the glass of a high-resolution flatbed scanner (HP OfficeJet Pro 7740, Hewlett-Packard Development Company, L.P., Houston, TX, USA). This method allowed us to obtain high-resolution digital images (1.3 µm per pixel resolution) with uniform illumination and minimal sample defects, as the resolution was high enough to zoom in to the finest veins.
We measured twelve data sets of leaf morphological traits: shape, leaf arrangement, leaf organization, lamina length, area of leaf size classes, apex angle, apex shape, base angle, number of basal veins, vein density, agrophic veins, and number of orders of teeth (Table 1; Supplementary Figure S1B) influencing water transport, light capture, and defense mechanisms (Ellis et al., 2009; Eller et al., 2016). Traits were measured at the mid‐leaf, taking care to avoid secondary veins. Leaf measurements (accuracy 0.01 mm) were performed using image analysis software (ImagePro v 4.5, Media Cybernetics, Carlsbad, CA) (Hickey et al., 1999; Guerin et al., 2012).
The assessment of differences between traits among species was carried out through one-way analysis of variance (ANOVA) and post-hoc Tukey’s test. Before analysis, the data were logarithmically transformed to improve normality and homoscedasticity. These analyses were performed using the R v.4.0.2 (R Core Team, 2018) and ggplot2 package (Wickham, 2016).
2.4 Data processing and analysis
2.4.1 Links between functional traits and climatic factors
To assess the relationship between functional traits and climatic factors (TMAX, TMIN, VPD, PRE, and EVT) model-based fourth-corner analysis was employed. This approach addresses the “fourth-corner problem” by examining the connections among three matrices: (1) species by geographic coordinates, (2) species by functional traits, and (3) geographic coordinates by climatic factors, ultimately estimating a matrix that reflects trait-climate concatenations (Borcard et al., 2011). We adhered to the framework outlined by Warton et al. (2015), utilizing the R package mvabund (Wang et al., 2012). This process involved fitting a generalized linear model (GLM) where climatic variables were treated as functional traits, including their interactions. To enhance the accuracy of likelihood estimates, the model was fitted using a quasi-Poisson distribution for model errors, incorporating a LASSO-penalized regression model with the mvabund package (Wang et al., 2012). The model’s performance was then evaluated using diagnostic plots. All analyses were conducted using the fourthcorner() functions available in the R package ade4 (Dray et al., 2007).
2.4.2 Multivariate analysis
We performed principal coordinate analysis (PCoA; Wang et al., 2022a) to identify dimensions of trait variability that maximally correlate with climatic factors (TMAX, TMIN, VPD, PRE, and EVT). In both analyses, presence/absence (1 or 0 respectively) scores were assigned to each trait category including traits which were originally represented by multiple categories (e.g., leaf size and shape). To assess climatic factors (distances) or similarities between functional traits to each TMCF tree species, we used the PCoA’s on the square root of the Bray–Curtis dissimilarities to obtain a fully Euclidean solution. All PCoA axes with eigenvalues >1 were retained, in this case the first four axes. When the distances are based on a complement-to-1 of a non-metric similarity coefficient, we eliminated the presence of negative eigenvalues using square-root transformation which makes it fully metric (Borcard et al., 2011). We extracted the first two axes of the analysis (Axis 1 and Axis 2) and used them in further analyses. The multivariate analyses were performed with CANOCO software v.5.0 (Šmilauer and Lepš, 2014).
2.4.3 Cluster analysis
To explore whether certain trait syndromes (recurrent combinations of multiple traits that have evolved together within or across species; Raffard et al., 2017), we used k-means cluster analysis (Hartigan and Wong, 1979). Consistent occurrence could indicate that there are traits adapted to specific climatic factors, or multiple ecological strategies (e.g., acclimations at the leaf, stem, and root levels) to given environmental conditions. Nonetheless, k-means clustering operates in a Euclidean space by first using the absolute values (Legendre and Legendre, 1998), and is inversely related to simple concordance, which demonstrates an unwanted property that rare traits are treated as similar simply because they have a lot of zeros in common (Šmilauer and Lepš, 2014). Therefore, we created a modified Euclidean space by first, we used the absolute values of the Pearson’s correlation coefficients to perform a cluster analysis between traits, then performing a principal coordinate analysis (Zuur et al., 2007) on these values, before running the clustering in the space generated by the first two principal coordinate axes. We performed the analysis using hmisc (Harrell, 2024) and pheatmap (Kolde, 2022) packages. Based on this assessment, the traits were assigned to a consensus cluster, representing the cluster to which they were most frequently assigned. These clusters were superimposed on the PCoA plots to examine their position in climate space (De Bello et al., 2021).
2.4.4 Climate influence on traits
To estimate smooth functional relationships between climate effects on cluster traits among TMCF tree species, we performed a generalized additive model (GAM; Wood, 2017) using a Poisson distribution. The explanatory factors were climatic factors (TMAX, TMIN, VPD, PRE, and EVT), whereas cluster traits were response variables (wood and leaf functional traits, Table 1; Supplementary Figure S1). To test the normality of the residuals of the response variables, we employed a Wilcoxon-Mann-Whitney test as outlined by Dehaene et al. (2021). A penalized smooth term prevents excess wiggliness and indicates the extent to which the fitted smooth function can vary. A wigglier function can capture intricate patterns in the data, while a less wiggly function results in a smoother more generalized fit (Wood, 2023). All the mixed models from this section can be compared using Akaike information criteria (AIC; Anderson et al., 1994) to determine the best fit model. Likewise, we use the effective degrees of freedom (edf), which at a maximum is the number of coefficients to be estimated in the model, minus any constraints (Pedersen et al., 2019). We performed all GAM analyses with R-software (R Core Team, 2018) using the glm2 (Donoghoe, 2022), lm4 (Bates et al., 2015), marginaleffects (Scholbeck et al., 2024), mumin (Barton, 2022) and ggplot2 (Wickham, 2016) packages.
3 Results
3.1 Trait behavior among TMCF tree species
We found high variations in wood anatomical traits among species (Figure 2A). Noteworthy, high vessel density (VD) values from 10 to 60 vessels per mm2) were identified in S. speciosa and F. mexicana; while T. mexicana, U. mexicana, and C. palmeri showed low vessels per mm2 (from 5 to 20). Likewise, we detected high hydraulic diameter (DH) values from 10 to 250 μm (i.e., M. schiedeana, U. mexicana, and C. palmeri), whereas narrow DH values from 10 to 20 μm were detected in T. insignis, S. nigra, S. glabrescens, B. mexicana, F. mexicana, and T. mexicana). We observed a similar effect of variation among species in M. schiedeana, F. mexicana, U. mexicana, and C. palmeri, where exhibited high vulnerability index (VI) (from 5 to 10 μm/mm-2), and vessel grouping (VG) values (from 2 to 5 Nvessels/Ngrouping); whereas S. speciosa, T. insignis, S. nigra, S. glabrescens, and B. mexicana exhibited constrained VI (from 2 to 5), and VG values (from 2 to 6). Notably, solitary vessel index (VS) values were similar among species (from 2 to 4 Nsolitary vessels/Ngrouping).
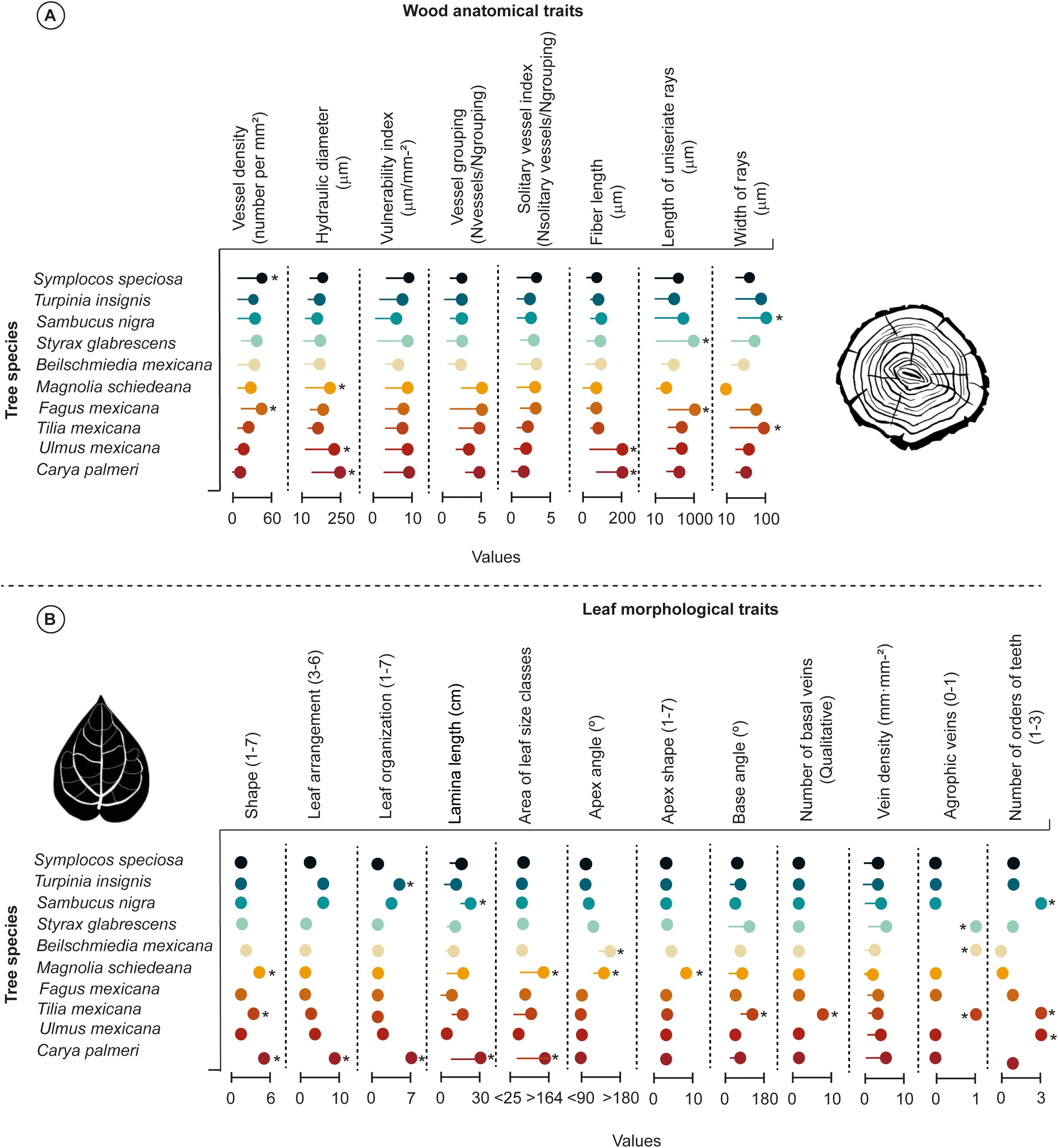
Figure 2. Lollipop plot illustrates all wood anatomical (A) and leaf morphological (B) traits variations among Tropical Montane Cloud Forest tree species. Lollipop plots with asterisks are significantly different as tested using a post-hoc Tukey test (p< 0.05). The asterisk denotes statistically significant differences between group means (p< 0.05).
Regarding the vessel elements, specific deciduous tree species (i.e., U. mexicana and C. palmeri) showed high fiber length (FL) values from 5 to 200 μm compared to the other TMCF tree species, whereas in S. speciosa, T. insignis, S. nigra, B. mexicana, M. schiedeana, F. mexicana, and T. mexicana displayed narrow FL values (from 2 to 30 μm). The length of uniseriate rays (LUR) was similar between S. glabrescens and F. mexicana (from 10 to 1000 μm), while M. schiedeana and B. mexicana showed the narrower LUR values (LUR; from 10 to 20 μm). Finally, higher ray width (WRY) values were detected in S. nigra and T. mexicana (WRY; from 10 to 100 μm), whereas M. schiedeana possessed narrower WRY values (10 μm) (Figure 2A).
Our analysis demonstrated differences in leaf morphological traits among tree species using one-way ANOVA and post-hoc Tukey tests, that can strongly influence the ability of the species to adapt to fluctuations in the humidity of the environment (Figure 2B). Remarkably, M. schiedeana (obovate), T. mexicana (ovate), and C. palmeri (odd-pinnate) showed high plasticity in leaf shape (Sh; from 5 to 6) compared to the other TMCF tree species surveyed. Leaf arrangement (La) values were considerably narrower among tree species (from 3 to 5); notwithstanding, C. palmeri exhibited high La values (from 5 to 6). Furthermore, T. insignis and C. palmeri showed high leaf organization (Lo; from 6 to 7), where the values ranged from 1 to 4. Noteworthy, M. schiedeana and C. palmeri demonstrated high variation in lamina length (Ll; from 15 to 30 cm) and area of leaf size classes (Al; ranged from > 30 to > 160; nanophyll) in relation to other species. Similar significant differences in apex angle (Aa; ranging from 7 to 10°) and apex shape (As; ranging from 8 to 10) were observed in B. mexicana and M. schiedeana. Likewise, T. mexicana displayed differences in base angle (Ba; ≈170°), and number of basal veins (Nb; ≈9) regarding to other species. Furthermore, S. glabrescens, B. mexicana, and T. mexicana showed high agrophic vein values (Av; 1); notwithstanding, S. nigra, T. mexicana and U. mexicana demonstrated high number of orders of teeth (Nt; 3) (Figure 2B).
3.2 Relationship between traits and climatic factors
Overall, the fourth-corner analyses demonstrated significant association between trait-climatic factor interactions (Figure 3), 753 correlations significantly differed from zero, with an inertia of the overall model of 1345 (p< 0.001). Most TMCF tree species’ wood anatomical traits and leaf morphological traits and all climatic factors were significantly correlated with at least one other variable; VD and DH positively with TMAX (from ≈0.05 to 0.15), whereas VI, VG. VS, Fl, LUR and WRY exhibited negative correlation with TMAX (from ≈-0.05 to -0.20; Figure 3A). The DH and Fl showed a negative correlation with PRE (-0.20; Figure 3A).
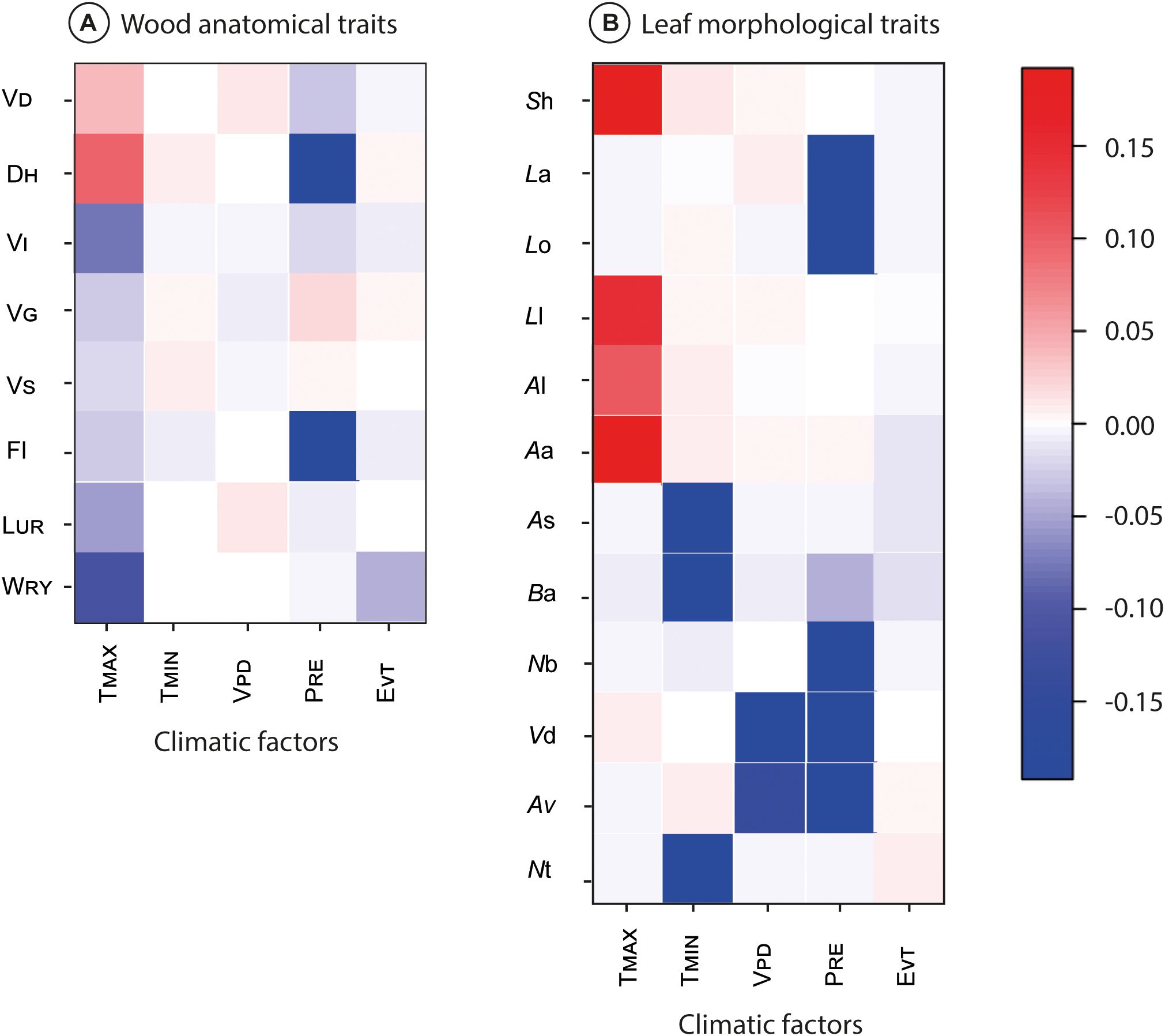
Figure 3. Results of the fourth‐corner analysis. Significant correlations between traits and climatic variables are represented by colored squares. Darker colors indicate stronger correlations. Wood anatomical traits: xylem vessel density (VD), hydraulic diameter (DH), vulnerability index (Vl), vessel grouping index (VG), solitary vessel index (VS), fiber length (Fl), length of uniseriate rays (LUR), and width of rays (WRY) (A). Leaf morphological traits: leaf arrangement (La), leaf organization (Lo), lamina length (Ll), area of leaf size classes (Al), apex angle (Aa), apex shape (As), base angle (Ba), number of basal veins (Nb), vein density (Vd), agrophic veins (Av), and number of orders of teeth (Nt) (B).
Of the leaf morphological traits, the Sh, Ll, Al, and Aa demonstrated a positive correlation with TMAX (from ≈0.10 to 0.20; Figure 3B); whereas As, Ba and Nt showed negative correlation with TMIN (-0.20), the Vd and Av exhibited negative correlation with VPD (from -0.10 to -0.15). Finally, the La, Lo, Nb, vd, and Av showed negative correlation with PRE (-0.20; Figure 3B).
3.3 Trait relationship
The heatmap of the eight-wood anatomical and twelve-leaf morphological traits demonstrated different patterns and relationships between traits. Therefore, wood anatomical traits showed positive correlations between LUR vs VD and Fl vs DH, (r= 1.0), whereas a negative correlation between VS vs VD (Figure 4A). Moreover, regarding the leaf traits, the relation between Ba vs As and Nt vs Ba exhibited positive correlation (r= 1.0), and a noteworthy negative correlation between Vd vs Sh, Nt vs Al, Vd vs As, and Nt vs Av (r= -1.0) (Figure 4B).
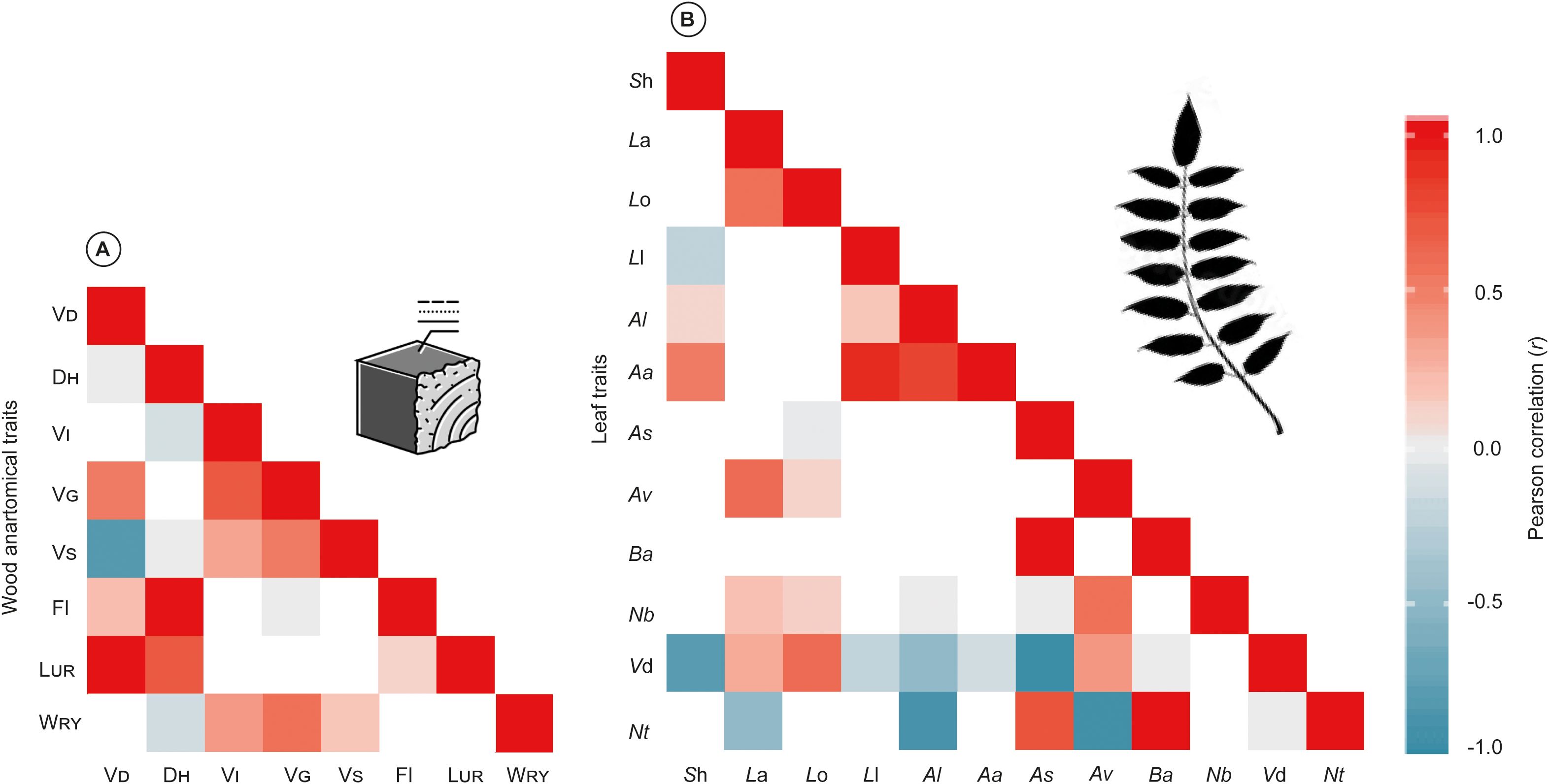
Figure 4. Pearson correlation (r) between wood anatomical (A), and leaf morphological traits (B). Red indicates the positive correlation, and blue demonstrates the negative correlation (p< 0.05 and p< 0.01, respectively). Trait abbreviations are given in Table 1.
3.4 Trait syndromes
Principal coordinate analysis (PCoA) revealed that traits were consistently grouped (i.e., Clusters I-V), and the location of these trait ‘syndromes’ in climate space was dissimilar between wood and leaf traits (Figure 5). They are a foundational concept for studying convergent evolution and the functional integration of traits in ecology and evolutionary biology. The PCoA did not suffer from collinearity among variables (for Pearson’s correlation coefficients) between the eight wood anatomical and twelve leaf traits surveyed (Figure 4). The trait syndromes were numbered according to their location on the dominant climate (Cluster I= VS, VG, WRY; Cluster II= LUR, VD Fl and DH; Cluster III= As, Ba and Nt; Cluster IV= Lo, Vd, Av, La, and Nb; and Cluster V= Al, Aa, La, and Sh).
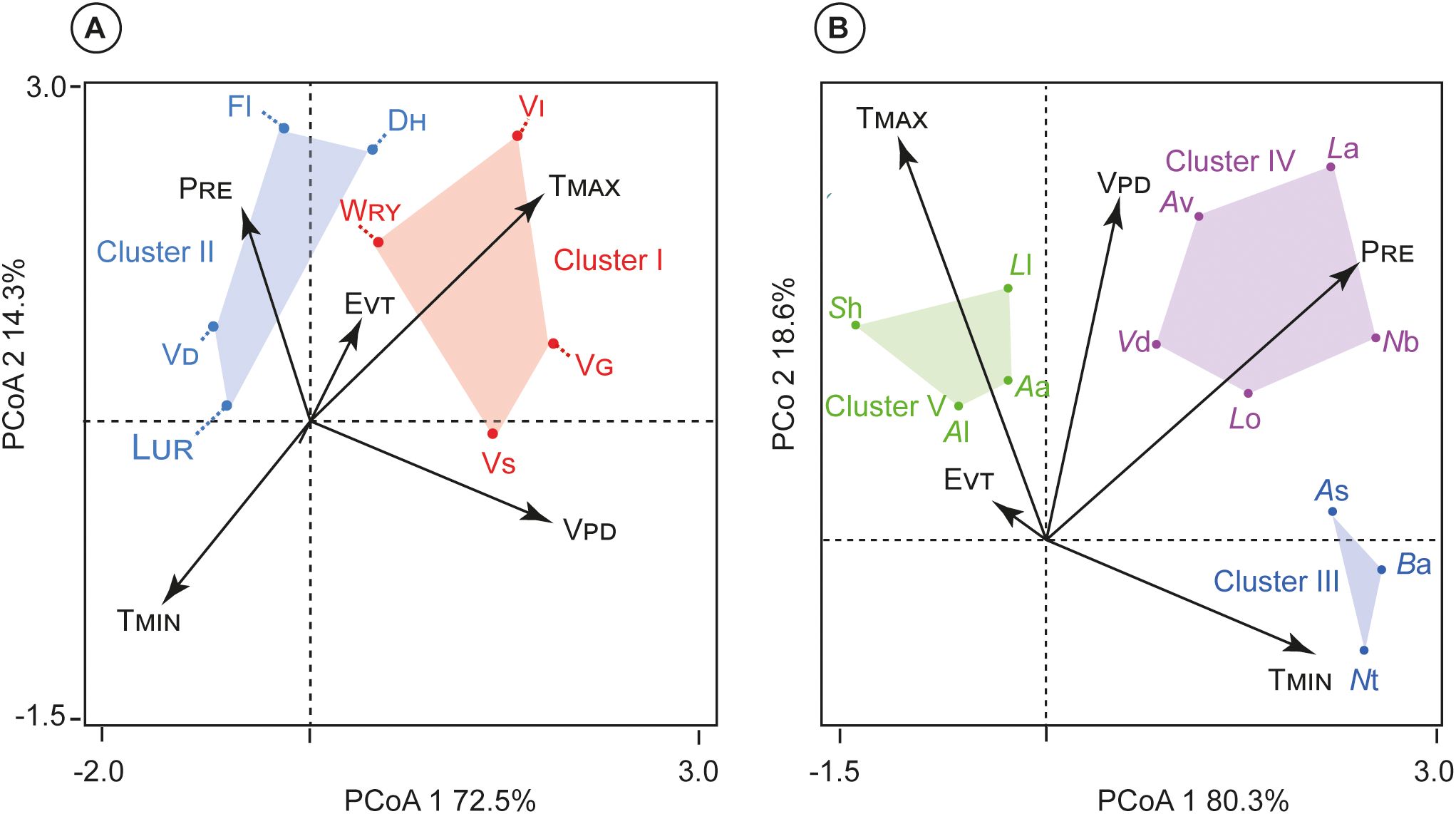
Figure 5. Clusters of traits from k-means cluster analysis. Traits in the same cluster are shown in space defined by climate-related trait dimensions from Principal Coordinate Analysis (PCoA). (A) Wood anatomical traits vs climatic factors; and (B) Leaf anatomical traits vs climatic factors. Trait abbreviations are given in Table 1.
Along the wood anatomical (Clusters I and II), axis 1 of PCoA was positively correlated with VPD (r= 0.882) but negatively with TMIN (r= −0.762). Axis 2 of PCoA was highly positively correlated with TMAX (r= 0.843) and EVT (r= 0.743) but negatively with PRE (r= -0.876) (Figure 5A). The wood anatomical traits PCoA explained a total of 86.8% of the variability, with the first axis (72.5%) associated with TMIN and VPD, whereas the second axis (14.3%) linked with PRE, and TMAX.
Along the leaf anatomical traits (Clusters III-V), axis 1 of PCoA was positively correlated with TMIN (r= 0.896). Axis 2 of PCoA was highly correlated with PRE (r= 0.978), VPD (r= 0.651), but negatively correlated with TMAX (r= -0.891), and EVT (r= -0.631) (Figure 5B). Furthermore, the leaf traits PCoA described a total of 98.9% of the variability, the first axis (80.3%) related with TMIN; while the second axis (18.6%) associated with PRE, EVT, VPD, and TMAX.
3.5 Functional relationship between cluster-traits and climatic factors
The results of the GAM models (Supplementary Table S1) influenced the smooth function of the response on Clusters [wood anatomical (Cluster I-II), and leaf traits (Cluster III-V)] and the climatic relationship (Figure 6). The TMAX effect on Cluster I (VI, VG, VS, WRY) demonstrated that the slope of B. mexicana, M. schiedeana, and F. mexicana exhibited similar shape, wiggliness and were positive up until TMAX values approached 18°C. Similarly, T. insignis, S. nigra, S. speciosa, T. mexicana, and C. palmeri curves exhibited values ranged from 17 to 20°C, whereas U. mexicana and S. glabrescens showed wider values ranging from 21 to 23°C (Figure 6A). Likewise, the PRE directly influenced on the Cluster II (DH, LUR, VD, and Fl) where T. insignis, S. nigra and S. glabrescens curves showed dissimilar shape and wiggliness with restricted values of 800 to 1000 mm, whereas U. mexicana, S. speciosa ranged from ≈1000 to 1300 mm. Noteworthy that, B. mexicana, M. schiedeana, C. palmeri, T. mexicana, and F. mexicana exhibit hydric efficiency with high precipitation values (> 2000 mm) (Figure 6B).
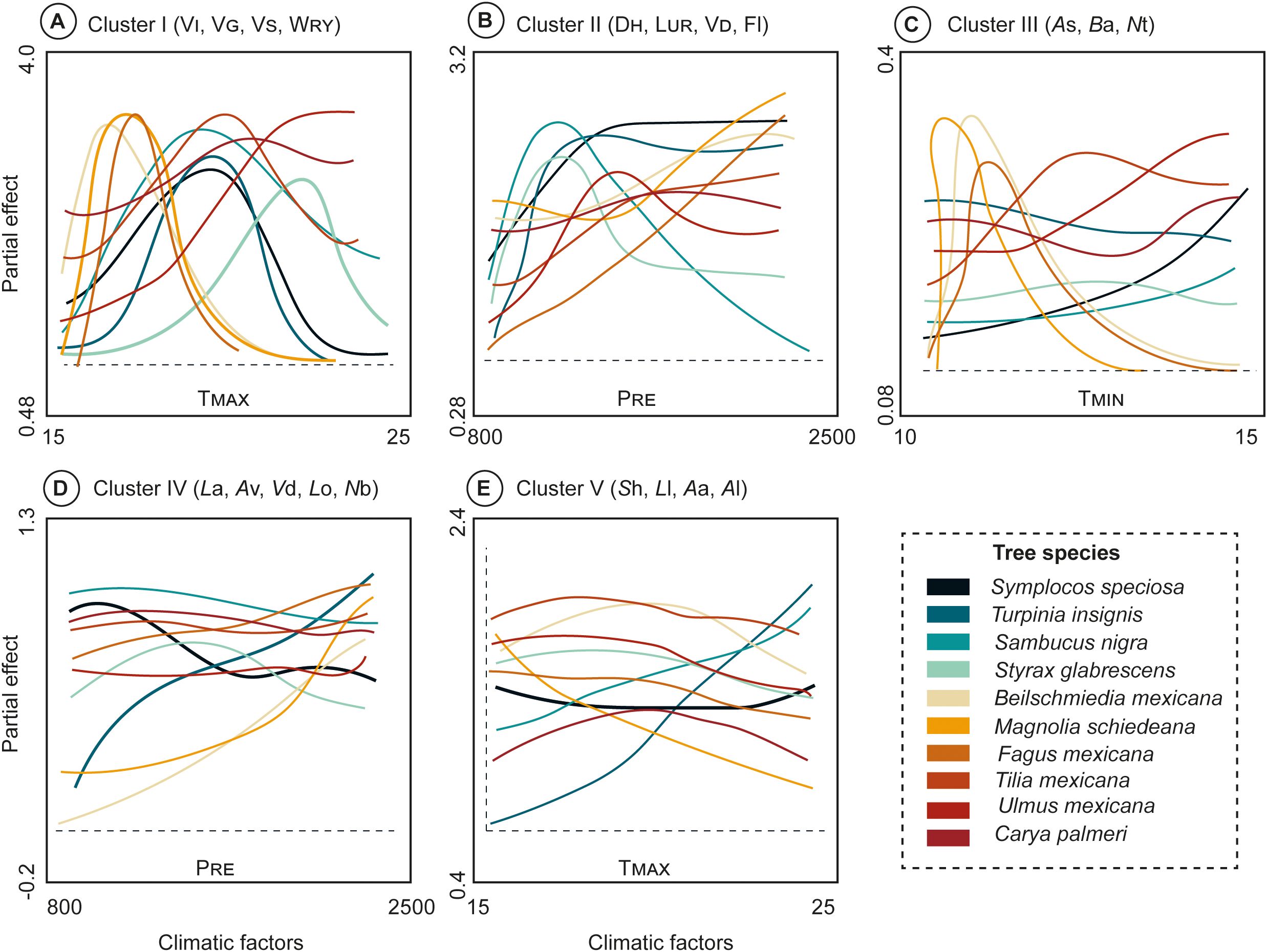
Figure 6. Response curves from the generalized additive models (GAM) according PCoA results of 10 TMCF tree species against specific climatic factors (TMAX, TMIN, PRE and clusters. (A) Cluster I (VI, VG, VS, WRY) with TMAX; (B) Cluster II (DH, LUR, VD, Fl) with PRE; (C) Cluster III (As, Ba, Nt) with TMIN; (D) Cluster IV (La, Av, Vd, Lo, Nb) with PRE; and (E) Cluster V (Sh, Ll, Aa, Al) with TMAX. Trait abbreviations are given in Table 1.
In addition, the Tmin demonstrated a strong influence on the Cluster III (As, Ba, Nt), where M. schiedeana, B. mexicana, and F. mexicana curves were constrained to values ranging from 11 to 13°C; even though S. speciosa, T. insignis, S. nigra, S. glabrescens, T. mexicana, U. mexicana, and C. palmeri curves showed wider values from 13 to 15°C (Figure 6C). In brief to assess the PRE effect, the Cluster IV (La, Av, Vd, Lo, Nb), the S. speciosa, T. insignis, S. nigra, S. glabrescens, F. mexicana, T. mexicana, U. mexicana, and C. palmeri curves demonstrated a very similar overall shape but differ in their wiggliness, with values ranging from 800 to 2500 mm; whereas B. mexicana and M. schiedeana curves showed similar shape and wiggliness with values of ≈2500 mm (Figure 6D).
Finally, the Tmax on Cluster V (Sh, Ll, Aa, Al), we observed a similar shape among F. mexicana, S. glabrescens, S. speciosa, B. mexicana, C. palmeri, T. mexicana, and U. mexicana curves; notwithstanding different wiggliness with values ranging from 15 to 25 °C. Moreover, T. insignis and S. nigra demonstrated similar shape but differ in wiggliness with values fluctuating from 20 to 25 °C, whereas M. schiedeana curve exhibited negative effect (Figure 6E).
4 Discussion
4.1 Towards an understanding of wood and leaf anatomical traits
Although we noted similar environmental conditions across the TMCF study area, our research suggests that each tree species has developed unique adaptations that allow it to adjust to the region’s macroclimatic changes (Eller et al., 2020; Zahedi et al., 2024). Nonetheless, the wood and leaf traits identified in this research indicate a particular ability to acclimate. The presence of TMCF tree species, such as F. mexicana, U. mexicana, C. palmeri, and M. schiedeana, are sensitive to changes in climate, as noted by several authors (Vásquez-Morales et al., 2014; Rodríguez-Ramírez et al., 2018; Ames-Martínez et al., 2022), indicate that they possess specific morphological characteristics that may increase their adaptive or resilience responses to fog and/or mist immersion changes (Sánchez-Velásquez et al., 2023).
It is noteworthy that M. schiedeana, U. mexicana, and C. palmeri exhibited significantly higher hydraulic diameter values than the other species, suggesting that they may influence the resilience of TMCF tree species to drought by enhancing water transport capacity, reducing vulnerability to cavitation, facilitating adaptive plasticity, improving growth recovery rates, and supporting ecological interactions within these unique forest ecosystems (Rodríguez-Ramírez et al., 2020a, 2022; Rodríguez‐Ramírez et al., 2024a). High-vessel density enhances drought resilience through thicker cell walls but reduces growth rates by 30-50% compared to low-vessel density (Eller et al., 2020; Mo et al., 2024). The high-vessel density presence in particular TMCF tree species (F. mexicana and S. speciosa) contribute to habitat diversity by supporting different microhabitats (López-Calvillo et al., 2023) and shaping the functioning of TMCFs (Hu and Riveros-Iregui, 2016; Eller et al., 2020).
The presence of high fiber length in U. mexicana, and C. palmeri contribute to the formation of strong, flexible and cohesive wood tissue. This structural advantage allows trees to grow larger and taller, increasing the potential for above-ground biomass accumulation and consequently higher carbon storage in living trees (Bukoski et al., 2022). It also increases mechanical strength to prevent canopy collapse under hydric stress (Brum et al., 2023). This is particularly important during periods of hydric stress (Rodríguez‐Ramírez et al., 2024b), as trees may experience increased vulnerability to physical damage from wind or heavy rainfall. A robust fiber structure helps maintain stability, allowing trees to withstand climatic stresses (i.e., soil moisture availability) that often accompany drought conditions (DeBell et al., 2002; Yang et al., 2024). This stress affects traits in TMCF tree species, influencing growth, and anatomy (Eller et al., 2020; Rodríguez‐Ramírez et al., 2024b). The high varying uniseriate ray length (i.e., S. glabrescens and F. mexicana) and wide width of rays (S. nigra and T. mexicana) facilitate hydraulic pathways, and store non-structural carbohydrates and other nutrients (von Arx et al., 2015). This storage is vital for the TMCF tree’s energy management, particularly during specific phenological processes (i.e., masting events; Rodríguez-Ramírez et al., 2019) or environmental stress (i.e., drought events, freezing; Rodríguez-Ramírez et al., 2023a). Likewise, the wood mechanical properties of these rays help trees to survive in windy, rainy conditions (Ayala-Usma et al., 2019). This connectivity is crucial for maintaining physiological balance, especially in the humid and variable conditions of TMCFs, where moisture levels can daily fluctuate significantly (Borchert et al., 2005; Fahey et al., 2016). This acclimation is crucial in dense TMCFs (i.e., Mexican beech forests; Rodríguez-Ramírez et al., 2016), where light is in short supply.
Nevertheless, the distinctive leaf morphology suggests that the TMCF tree species evolved to thrive in the diverse moisture conditions. Even though these phenotypic traits could be coded by independent genes, and modified by different selective pressures (Sobral, 2021), the leaf anatomical traits are mechanistically related and will cohere (Wright et al., 2017; Wang et al., 2022a), and may have advantages in one moisture environment and costs in another (Givnish, 1984; Guerin et al., 2012; Sánchez-Velásquez et al., 2023). Many tree species in TMCFs exhibit structural characteristics of leaves that enhance their ability to utilize the condensation of water droplets from fog and/or mist, which can then be absorbed directly into the plant system (Royer et al., 2012; Oliveira et al., 2014; Hughes et al., 2024). This morphological variation within species along TMCF communities suggests adaptation, which may allow local persistence and migration of adaptive potential, or at least response to moisture variation (Ackerly et al., 2002; Hughes et al., 2024). The presence the M. schiedeana, T. mexicana, and C. palmeri displayed a larger size of leaf, area of leaf size classes compared with smaller leaf species (i.e., F. mexicana, S. speciosa, U. mexicana, T. insignis, and B. mexicana), maximizing surface area (i.e., nanophyll with intermediate precipitation seasonality; Givnish, 1984; Wang et al., 2022b) for absorbing moisture from the humid air (i.e., vapor plumes, fog, mist, and drizzle), maintaining hydration during drier periods when rainfall may be scarce (Liancourt et al., 2015; Wright et al., 2017; Wang et al., 2022b). Nonetheless, M. schiedeana and B. mexicana with apex angle (≥ 180°), and apex shape (i.e., convex, rounded, and acuminate) facilitate the runoff of water from fog or mist, ensuring that excess moisture does not accumulate on the leaf surface (Graham and Christopher, 2023). This adaptation prevents fungal infections and other moisture-related problems in the subcanopy of TMCFs (Cheek et al., 2023).
In particular, T. mexicana exhibited a notably wider leaf angle base (> 170°, asymmetrical leaf bases) than the other TMCF tree species, which allows for better light capture and photosynthesis (McCarthy and Mason-Gamer, 2020; Ramírez-Díaz et al., 2024), especially in Lower TMCFs where light conditions can be variable (Rodríguez-Ramírez et al., 2024a). Besides, the presence of agrophic veins in specific tree species (i.e., S. glabrescens, B. mexicana, and T. mexicana) enabling the passive diffusion of water, nutrients (e.g., sugar, hormone auxin), regulating cell-to-cell communication (Band, 2022). This characteristic has been reported in other species from moist environments, such as Bernardia species (Cervantes et al., 2009), and extinct species as Tilia populifolia H.T. Chang, Corylopsis reedae Radtke, Pigg et Wehr, Fothergilla malloryi Radtke, Pigg et Wehr, and Apeibopsis atwoodii Hollick (Radtke et al., 2005; Carvalho et al., 2011).
Finally, toothed leaves are more common in areas with high rainfall and lower temperatures, such as the TMCF, demonstrating a complex concatenation among morphology, physiology, and environmental adaptation. Similarly, toothed leaves in TMCF tree species are primarily associated with strategies that maximize early-season photosynthesis and rapid growth in cool, wet, and water-abundant environments, leveraging the unique climatic and hydrological conditions of TMCFs (Givnish, 1984; Royer and Wilf, 2006). U. mexicana, T. mexicana and S. nigra exhibited high number of the order teeth, leaf hydration and function during dry periods (Iszkulo et al., 2024); notwithstanding, this limits leaf longevity, offering probably an early season opportunity for rapid photosynthesis (Baker-Brosh and Peet, 1997). Notably, M. schiedeana displays leaves that are broadly oblong-elliptical in shape with untoothed margins (Rodríguez-Ramírez et al., 2020a), which may help to low leaf nitrogen concentration and reduce the likelihood of hydric stress during drought events (Royer et al., 2012). This is because loss of cellular turgor would not result in leaf wilting or collapse (Postek, 1981), involving the movement of xylem sap and the resultant carbon economy of the TMCF tree species (Royer and Wilf, 2006; Royer et al., 2012).
4.2 Climatic effect on functional trait syndromes
Several of the individual functional trait–climatic factor correlations that we observed support the functional trait theory (Kearney et al., 2021; Viliani et al., 2024), such as specific wood anatomical traits (DH and VD) and leaf morphological traits (Sh, Ll, Al, and Aa). According to Kühn et al. (2021), functional trait stability may influence performance or fitness in fluctuating climates. Although the observed correlations are descriptive, they are predictive of traits that are likely to change in response to environmental variation (Sánchez‐Velásquez et al., 2023). Correlations between functional traits and climatic factors provide a reference point for identifying environmental variables associated with traits exhibited by relict-endemic TMCF tree species (Rodríguez-Ramírez et al., 2023b; Rodrı́guez-Ramı́rez et al., 2024a).
The wood and leaf traits adjust among TMCF tree species because of the changing environmental conditions (Toledo-Aceves et al., 2019; Yang et al., 2021; Rodríguez-Ramírez et al., 2024b). TMAX triggers a synergy among wood anatomical traits (Cluster I: VS, VG, WRY and VI) is an adaptive trait that allows plants to optimize water transport according to their ecological niches (von Arx et al., 2013; Scholz et al., 2014). The combination of historical maximum temperature ranges with seasonal variations and anticipated future climate data for tropical tree species (Ponce-Reyes et al., 2012), could result in a reduction in trait-fitness in climate change (Vasseur et al., 2014; Wright and Francia, 2024). This is because of an increased likelihood of these species experiencing temperatures above their critical thermal tolerances, which may lead to slower growth (Guillemot et al., 2022; Feeley et al., 2023).
Considering the crucial role that cloud uplift events play in the TMCF water budget, a reduction in the frequency of fog, but rainier climate is likely to result in increased evapotranspiration, vegetation hydric stress and, subsequently, plant mortality (Oliveira et al., 2014; Zhang et al., 2024). For instance, TMCF species in less moist environments might have evolved narrower vessels to enhance safety (Anfodillo and Olson, 2021; Argüelles-Marrón et al., 2023), whereas those in wetter environments might allowing for adequate hydraulic connection in specific xylem vessel traits (i.e., LUR, DH, VD, and Fl), preventing mortality through reduced resilience to hydric stress (Rodríguez‐Ramírez et al., 2024b).
Tropical trees subjected to hydric stress exhibit morpho-anatomical changes that are closely linked to cellular, physiological and biochemical acclimations aimed at minimizing water loss through transpiration and optimizing water use efficiency (Amesalu and Kebede, 2020; Eller et al., 2020; Zahedi et al., 2024). Significant concatenation among leaf morphological traits can indicate adaptation in response to shifts in climate (Givnish, 1984; Doughty et al., 2018; Manishimwe et al., 2022). It is evident that the identified clusters are subject to influence from climatic factors, including VPD, TMAX, PRE, and TMIN. These syndromes allow for maximum fitness under hydric stress, resulting in specific morphological adaptations to TMCF conditions (Andrés-Hernández et al., 2023; Rodríguez-Ramírez et al., 2023a; Rodrı́guez-Ramı́rez et al., 2023b). Due to the considerable diversity in hydraulic trait assemblages, it is currently impossible to determine the leaf hydraulic conductivity and vulnerability to hydric stress of TMCFs (Liancourt et al., 2015). Furthermore, the potential risk of hydraulic failure is also unknown (Mackay, 2024; Robbins et al., 2024), given the lack of available data on climatic factors such as the vapor pressure deficit (VPD), precipitation (PRE), maximum temperature (TMAX) and minimum temperature (TMIN). It is therefore possible that ecological niche conservatism has influenced the persistence of TMCF tree species, limiting the distribution of ecologically dissimilar functional traits across moist or temperature variation in specific regions (Saldaña-Acosta et al., 2008).
The adaptative interspecific capability differences in leaf anatomical traits suggest a wide diversification of the hydraulic strategy among TMCF tree species (Apgaua et al., 2022; Manishimwe et al., 2022). Nevertheless, our research demonstrates that the probability of leaf morphological variations during low moisture periods depend on an understanding of the interaction between leaf anatomical trait assemblages (Robbins et al., 2024). Conversely, the clustering of traits (i.e., As, Ba, Nt, La, Av, Vd, Lo, Nb, Sh, Ll, Aa, and Al) may demonstrate hydraulic adjustments that triggers a mechanical safety versus water transport efficiency tradeoff (Tng et al., 2018), indicating that leaf morphology and phenology reflect a set of ecological strategies that can co-vary with anatomical hydraulic traits (Apgaua et al., 2022). Additionally, the leaf morphological in some evergreen species (e.g., M. schiedeana, S. speciosa, B. mexicana) and semideciduous tree species (e.g., F. mexicana) (Figure 2) supports the argument that these species may have a hydraulic adjustment that makes them highly sensitive to hotter climates (Nishida and Christophel, 1999; Rodríguez-Ramírez et al., 2020b; Rodrı́guez-Ramı́rez et al., 2021a; Reyes-Ortiz et al., 2024).
Functional trait syndromes varied significantly among TMCF tree species, with unique trait combinations (Clusters) emerging for specific climatic responses. These results are in accordance with those previously reported by (Nishida and Christophel, 1999; Vogt, 2001; Rodríguez-Ramírez et al., 2018; Rodrı́guez-Ramı́rez et al., 2020b; Rodrı́guez-Ramı́rez et al., 2021b; Rodrı́guez-Ramı́rez et al., 2024a; Bartholomew et al., 2020; López-Calvillo et al., 2023; Aranda et al., 2024; Ramírez-Díaz et al., 2024; Reyes-Ortiz et al., 2024), where the TMCF species as F. mexicana, T. mexicana, M. schiedeana, B. mexicana, S. nigra, S. speciosa, and C. palmeri exhibited specific wood and/or leaf trait acclimation and/or adaptation strategies to moisture variation, suggesting divergent relationships between trait values and fitness in a particular context are defined by multiple selective coefficients (Sobral, 2021), that shape the evolutionary course of TMCF communities (Price et al., 2011). Further research is required to gain a deeper understanding of the direct influence of climate on the relict-endemic TMCF tree species in phenological processes (e.g., mass flowering, cribo-xylogenesis).
The resilience of TMCFs depends on balancing trait-mediated trade-offs between drought tolerance and productivity (Locatelli et al., 2015). While conservative traits buffer against moderate drought, their slow growth rates may hinder recovery from extreme disturbances (Saatchi et al., 2021). Conversely, acquisitive strategies risk hydraulic failure under prolonged drought but maintain faster carbon cycling (Peters et al., 2019). The interplay between fog persistence, temperature rise, and moisture variability will determine whether these ecosystems can functionally adapt or face compositional collapse. This will enable us to provide a better understanding of resilience, resistance and recovery to changes in moisture conditions, and identify a reliable and effective choice for future restoration and conservation strategies. Targeted conservation should prioritize areas of high functional diversity to maintain adaptive capacity (Goldsmith et al., 2017; Forzieri et al., 2022).
5 Conclusions
In this paper we demonstrated that some TMCF tree species exhibit different wood and leaf anatomical adjustments or strategies in response to the wet environment. This highlights the importance of wood and leaf anatomy trait study for TMCF tree species and indicates that climate variation may alter acclimation and adaptation capacity among species. Finally, our study provides a framework that could be used to begin using functional trait theory to understand the relative importance of different growth patterns, total photosynthetic rate, leaf chlorophyll content and transpiration on the ability of TMCF tree species to survive periods of hydric stress. Future work should test the observed patterns using bark, cribo-xylogenesis to three distinct types of TMCFs: Lower (LTMCF; >700–1700 m asl), Upper (UTMCF; 1701–1799 m asl), and Subalpine (STMCF; 1800–3500 m asl). These studies will improve the representation of plant hydraulics within the montane ecosystem and help to understand phenological processes such as growth, development, and acclimation of TMCF tree species during periods of hydric deficit and refine predictions of how future climate change will affect TMCF functional traits.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author contributions
AA: Conceptualization, Data curation, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. ER: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Software, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We would like to show our thanks to Brenda Muñoz-Vazquez, Santiago Hernández-Rosales, and Fressia N. Ames Martínez for his assistance with fieldwork.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1555607/full#supplementary-material
References
Ackerly, D. D., Knight, C. A., Weiss, S. B., Barton, K., and Starmer, K. P. (2002). Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: Contrasting patterns in species level and community level analyses. Oecologia 130, 449–457. doi: 10.1007/s004420100805
Amesalu, F. and Kebede, G. (2020). Anatomical, morphological and physiological responses of plants to drought stress. Afr J. Agric. Res. 38, 486–496. doi: 10.5829/idosi.wasj.2020.486.496
Ames-Martínez, F. N., Luna-Vega, I., Dieringer, G., and Rodríguez-Ramírez, E. C. (2022). The effect of climate change on Arcto-Tertiary Mexican beech forests: Exploring their past, present, and future distribution. Ecol. Evol. 12, 1–12. doi: 10.1002/ece3.9228
Anderegg, W. R. L. and Meinzer, F. C. (2015). “Wood anatomy and plant hydraulics in a changing climate,” in Functional and Ecological Xylem Anatomy. Ed. Hacke, U. (Springer International Publishing, Switzerland), 235–253. doi: 10.1007/978-3-319-15783-2_9
Anderson, D. R., Burnham, K. P., and White, G. C. (1994). AIC model selection in overdispersed capture-recapture data. Ecology 75, 1780–1793. doi: 10.2307/1939637
Andrés-Hernández, A. R., Luna-Vega, I., and Rodríguez-Ramírez, E. C. (2023). Functional traits and adaptive capacity of cloud forest Ternstroemia species in response to climatic variation. Flora: Morphol Distrib Funct. Ecol. Plants 307. doi: 10.1016/j.flora.2023.152383
Anfodillo, T. and Olson, M. E. (2021). Tree mortality: Testing the link between drought, embolism vulnerability, and xylem conduit diameter remains a priority. Front. For Glob Change 4. doi: 10.3389/ffgc.2021.704670
Apgaua, D. M. G., Tng, D. Y. P., and Laurance, S. G. W. (2022). Tropical wet and dry forest tree species exhibit contrasting hydraulic architecture. Flora: Morphol Distrib Funct. Ecol. Plants 291, 152072. doi: 10.1016/j.flora.2022.152072
Aranda, I., Martin-Benito, D., Sánchez-Gómez, D., de Simón, B. F., and Gea-Izquierdo, G. (2024). Different drought-tolerance strategies of tree species to cope with increased water stress under climate change in a mixed forest. Physiol. Plant 176, e14562. doi: 10.1111/ppl.14562
Argüelles-Marrón, B., Meave, J. A., Luna-Vega, I., Crispín-DelaCruz, D. B., Szejner, P., Ames-Martínez, F. N., et al. (2023). Adaptation potential of Neotropical montane oaks to drought events: Wood anatomy sensitivity in Quercus delgadoana and Quercus meavei. Funct. Ecol. 37, 2040–2055. doi: 10.1111/1365-2435.14362
Ayala-Usma, D. A., Lozano-Gutiérrez, R. E., and González Arango, C. (2019). Wood anatomy of two species of the genus Chrysochlamys (Clusiaceae: Clusioideae: Clusieae) from the northern Andes of Colombia. Heliyon 5, 1–7. doi: 10.1016/j.heliyon.2019.e02078
Báez, S., Fadrique, B., Feeley, K., and Homeier, J. (2022). Changes in tree functional composition across topographic gradients and through time in a tropical montane forest. PloS One 17, e0263508. doi: 10.1371/journal.pone.0263508
Baker-Brosh, K. F. and Peet, R. K. (1997). The ecological significance of lobed and toothed leaves in temperate forest trees. Ecology 78, 1250–1255. doi: 10.1016/j.jaci.2012.05.050
Band, L. R. (2022). Plasmodesmata play a key role in leaf vein patterning. PloS Biol. 20, 8–11. doi: 10.1371/journal.pbio.3001806
Bartholomew, D. C., Bittencourt, P. R. L., da Costa, A. C. L., Banin, L. F., de Britto Costa, P., Coughlin, S. I., et al. (2020). Small tropical forest trees have a greater capacity to adjust carbon metabolism to long-term drought than large canopy trees. Plant Cell Environ. 43, 2380–2393. doi: 10.1111/pce.13838
Barton, K. (2022). Package ‘MuMIn’ Version 1.46.0 (Vienna, Austria: R Foundation for Statistical Computing) Available online at: https://cran.r-project.org/package=MuMIn.
Bates, D., Mächler, M., Bolker, B. M., and Walker, S. C. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw 67, 1–48. doi: 10.18637/jss.v067.i01
Berlyn, G. P. and Miksche, J. P. (1976). Botanical Microtechnique and Cytochemistry (Ames, IA, USA: Iowa State University Press).
Borcard, D., Gillet, F., and Legendre, P. (2011). Numerical Ecology with R (New York, NY: Springer New York). doi: 10.1007/978-1-4419-7976-6
Borchert, R., Robertson, K., Schwartz, M. D., and Williams-Linera, G. (2005). Phenology of temperate trees in tropical climates. Int. J. Biometeorol 50, 57–65. doi: 10.1007/s00484-005-0261-7
Bruijnzeel, L. A., Mulligan, M., and Scatena, F. N. (2011). Hydrometeorology of tropical montane cloud forests: emerging patterns. Hydrol Process 25, 465–498. doi: 10.1002/hyp.7974
Bruijnzeel, L. A., Scatena, F. N., and Hamilton, L. S. (2010). Tropical montane cloud forests: Science for Conservation and Management (Cambridge, UK: Cambridge University Press).
Brum, M., Vadeboncoeur, M., Asbjornsen, H., Puma Vilca, B. L., Galiano, D., Horwath, A. B., et al. (2023). Ecophysiological controls on water use of tropical cloud forest trees in response to experimental drought. Tree Physiol. 43, 1514–1532. doi: 10.1093/treephys/tpad070
Bukoski, J. J., Cook-Patton, S. C., Melikov, C., Ban, H., Chen, J. L., Goldman, E. D., et al. (2022). Rates and drivers of aboveground carbon accumulation in global monoculture plantation forests. Nat. Commun. 13, 4206. doi: 10.1038/s41467-022-31380-7
Burns, K. C. (2012). Masting in a temperate tree: Evidence for environmental prediction? Austral Ecol. 37, 175–182. doi: 10.1111/j.1442-9993.2011.02260.x
Carvalho, M. R., Herrera, F. A., Jaramillo, C. A., Wing, S. L., and Callejas, R. (2011). Paleocene Malvaceae from northern South America and their biogeographical implications. Am. J. Bot. 98, 1337–1355. doi: 10.3732/ajb.1000539
Cervantes, A., Terrazas, T., and Hernández, H. M. (2009). Foliar architecture and anatomy of Bernardia and other genera of Acalyphoideae (Euphorbiaceae). Brittonia 61, 375–391. doi: 10.1007/s12228-009-9084-x
Cheek, M., Edwards, S., and Onana, J. M. (2023). A massive Critically Endangered cloud forest tree, Microcos rumpi (Grewiaceae) new to science from the Rumpi Hills, SW Region, Cameroon. Kew Bull. 78, 247–258. doi: 10.1007/s12225-023-10119-7
Choat, B., Brodribb, T. J., Brodersen, C. R., Duursma, R. A., López, R., and Medlyn, B. E. (2018). Triggers of tree mortality under drought. Nature 558, 531–539. doi: 10.1038/s41586-018-0240-x
Crausbay, S. D., Frazier, A. G., Giambelluca, T. W., Longman, R. J., and Hotchkiss, S. C. (2014). Moisture status during a strong El Niño explains a tropical montane cloud forest’s upper limit. Oecologia 175, 273–284. doi: 10.1007/s00442-014-2888-8
DeBell, D. S., Singleton, R., Harrington, C. A., and Gartner, B. L. (2002). Wood density and fiber length in young Populus stems: Relation to clone, age, growth rate, and pruning. Wood Fiber Sci. 34, 529–539.
De Bello, F., Carmona, C. P., Dias, A. T., Götzenberg, L., Moretti, M., and Berg, M. P. (2021). Handbook of Trait-Based Ecology From Theory to R Tools. 1st (United Kingdom: Cambridge University Press). doi: 10.1017/9781108628426
Dehaene, H., De Neve, J., and Rosseel, Y. (2021). A Wilcoxon–Mann–Whitney test for latent variables. Front. Psychol. 12. doi: 10.3389/fpsyg.2021.754898
Donoghoe, M. M. W. (2022). Package ‘glm2.’ (Vienna, Austria: R Foundation for Statistical Computing).
Doughty, C. E., Santos-Andrade, P. E., Shenkin, A., Goldsmith, G. R., Bentley, L. P., Blonder, B., et al. (2018). Tropical forest leaves may darken in response to climate change. Nat. Ecol. Evol. 2, 1918–1924. doi: 10.1038/s41559-018-0716-y
Dray, S., Dufour, A. B., and Chessel, D. (2007). The ade4 package-II: Two-table and K-table methods. R News 7, 47–52. doi: 10.18637/jss.v022.i04
Eller, C. B., Lima, A. L., and Oliveira, R. S. (2016). Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change. New Phytol. 211, 489–501. doi: 10.1111/nph.13952
Eller, C. B., Meireles, L. D., Sitch, S., Burgess, S. S. O., and Oliveira, R. S. (2020). How climate shapes the functioning of tropical montane cloud forests. Curr. For Rep. 6, 97–114. doi: 10.1007/s40725-020-00115-6
Ellis, B., Daly, D. C., Hickey, L. J., Johnson, K. R., Mitchell, J. D., Wilf, P., et al. (2009). Manual of Leaf Architecture. The New York Botanical Garden Press (Ithaca, New York, USA: Cornell University Press).
Fahey, T. J., Sherman, R. E., and Tanner, E. V. J. (2016). Tropical montane cloud forest: environmental drivers of vegetation structure and ecosystem function. J. Trop. Ecol. 32, 355–367. doi: 10.1017/S0266467415000176
Feeley, K. J., Feeley, K. J., Bernal-Escobar, M., Fortier, R., and Kullberg, A. T. (2023). Tropical trees will need to acclimate to rising temperatures. But can they? Plants 12, 3142. doi: 10.20944/preprints202306.1937
Forzieri, G., Dakos, V., McDowell, N. G., Ramdane, A., and Cescatti, A. (2022). Emerging signals of declining forest resilience under climate change. Nature 608, 534–539. doi: 10.1038/s41586-022-04959-9
Gentry, A. H. (1992). Tropical forest biodiversity: distributional patterns and their conservational significance. Oikos 63, 19–28. doi: 10.2307/3545512
Givnish, T. J. (1984). “Leaf and canopy adaptations in tropical forests,” in Physiological ecology of plants of the wet tropics. Eds. Medina, E., Mooney, H. A., and C., V.-Y. (Springer Dordrecht, Mexico), 51–84. doi: 10.1007/978-94-009-7299-5_6
Goldsmith, G. R., Bentley, L. P., Shenkin, A., Salinas, N., Blonder, B., Martin, R. E., et al. (2017). Variation in leaf wettability traits along a tropical montane elevation gradient. New Phytol. 214, 989–1001. doi: 10.1111/nph.14121
Graham, J. H. and Christopher, R. (2023). Leaf shape and self-mulching by trees: A hypothesis. Symmetry 15, 1–35. doi: 10.3390/sym15061198
Guerin, G. R., Wen, H., and Lowe, A. J. (2012). Leaf morphology shift linked to climate change. Biol. Lett. 8, 882–886. doi: 10.1098/rsbl.2012.0458
Guillemot, J., Martin-StPaul, N. K., Bulascoschi, L., Poorter, L., Morin, X., Pinho, B. X., et al. (2022). Small and slow is safe: On the drought tolerance of tropical tree species. Glob Chang Biol. 28, 2622–2638. doi: 10.1111/gcb.16082
Hartigan, J. A. and Wong, M. A. (1979). Algorithm AS 136: A K-means clustering algorithm. J. R Stat. Soc. Ser. B Methodol 28, 100–108. doi: 10.2307/2346830
Hickey, L., Ash, A., Ellis, B., Johnson, K., Wilf, P., and Wing, S. (1999). Manual of leaf architecture- morphological description and categorization of dicotyledonous and net-veined monocotyledonous angiosperms. 1st. Eds. Ash, A., Ellis, B., Hickey, L. J., Johnson, K., Wilf, P., and Wing, S. (Washington, D. C: Smithsonian Institution).
Hu, J. and Riveros-Iregui, D. A. (2016). Life in the clouds: are tropical montane cloud forests responding to changes in climate? Oecologia 180, 1061–1073. doi: 10.1007/s00442-015-3533-x
Hughes, N. M., Sanchez, A., Berry, Z. C., and Smith, W. K. (2024). Clouds and plant ecophysiology: missing links for understanding climate change impacts. Front. For. Glob. Change 7. doi: 10.3389/ffgc.2024.1330561
IAWA COMMITTEE (1989). IAWA list of microscopic features for hardwood identification by an IAWA Committee. IAWA J. 10, 219–332. doi: 10.2307/4110625
Iszkulo, G., Tyrala-Wierucka, Z., Terlecka, M., Walas, L., and Tomaszewski, D. (2024). “What’s the point of toothed leaves?,” in Research and Practice in Forest Ecology: International Conference, Vol. 101–105 (Kórnik, Poland: Institute of Dendrology, Polish Academy of Sciences).
Karger, D. N., Nobis, M. P., Normand, S., Graham, C. H., and Zimmermann, N. E. (2021). CHELSA-TraCE21k v1.0. Downscaled transient temperature and precipitation data since the last glacial maximum. Clim Past Discuss. doi: 10.5194/cp-2021-30
Kearney, M. R., Jusup, M., McGeoch, M. A., Kooijman, S. A., and Chown, S. L. (2021). Where do functional traits come from? The role of theory and models. Funct. Ecol. 35, 1385–1396. doi: 10.1111/1365-2435.13829
Kindt, R. and Coe, R. (2005). Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies (Nairobi: World Agroforestry Centre (ICRAF).
Kolde, R. (2022). Package “pheatmap”: Pretty heatmaps (Vienna, Austria: R Foundation for Statistical Computing).
Kühn, N., Tovar, C., Carretero, J., Vandvik, V., Enquist, B. J., and Willis., K. J. (2021). Globally important plant functional traits for coping with climate change. Front. Biogeogr 13, 1–18. doi: 10.21425/F5FBG53774
Liancourt, P., Boldgiv, B., Song, D. S., Spence, L. A., Helliker, B. R., Petraitis, P. S., et al. (2015). Leaf-trait plasticity and species vulnerability to climate change in a Mongolian steppe. Glob Chang Biol. 21, 3489–3498. doi: 10.1111/gcb.12934
Locatelli, B., Catterall, C. P., Imbach, P., Kumar, C., Lasco, R., Marín-Spiotta, E., et al. (2015). Tropical reforestation and climate change: beyond carbon. Restor. Ecol. 23, 337–343. doi: 10.1111/rec.12209
López-Calvillo, L. F., Carbó-Ramírez, P., and Rodríguez-Ramírez, E. C. (2023). Small-fragment, high turnover: soil microenvironment fluctuation effect on tree diversity in a Neotropical montane oak forest. PeerJ 11, e15415. doi: 10.7717/PEERJ.15415
Los, S. O., Street- Perrott, F. A., Loader, N. J., and Froyd, C. A. (2021). Detection of signals linked to climate change, land-cover change and climate oscillators in tropical montane cloud forests. Remote Sens Environt 260, 112431. doi: 10.1016/j.rse.2021.112431
Luna-Vega, I., Alcántara-Ayala, O., García-Morales, L. J., Espinosa, D., Ramírez-Martínez, J. C., and Contreras-Medina, R. (2022). Threatened trees characteristic of Mexican Tropical Montane Cloud Forests. Diversity 15, 1–16. doi: 10.3390/d15010042
Mackay, D. S. (2024). Future tree mortality is impossible to observe, but a new model reveals why tropical tree traits matter more than climate change variability for predicting hydraulic failure. New Phytol. 244, 2115–2117. doi: 10.1111/nph.20049
Manishimwe, A., Ntirugulirwa, B., Zibera, E., Nyirambangutse, B., Mujawamariya, M., Dusenge, M. E., et al. (2022). Warming responses of leaf morphology are highly variable among tropical tree species. Forests 13, 1–24. doi: 10.3390/f13020219
McCarthy, D. M. and Mason-Gamer, R. J. (2020). Morphological variation in North American Tilia and its value in species delineation. Int. J. Plant Sci. 181, 175–195. doi: 10.1086/706781
McCracken, E. and Johansen, D. A. (1940). Plant Microtechnique. 1st (New York, NY: McGraw-Hill Book Co. Ltd). doi: 10.2307/3222557
McGill, B. J., Enquist, B. J., Weiher, E., and Westoby, M. (2006). Rebuilding community ecology from functional traits. Trends Ecol. Evol. 21, 178–185. doi: 10.1016/j.tree.2006.02.002
Miranda, F. and Sharp, A. J. (1950). Characteristics of the vegetation in certain temperate regions of eastern Mexico. Ecology 31, 313–333. doi: 10.1017/CBO9781107415324.004
Mo, L., Crowther, T. W., Maynard, D. S., Van den Hoogen, J., Ma, H., Bialic-Murphy, L., et al. (2024). The global distribution and drivers of wood density and their impact on forest carbon stocks. Nat. Ecol. Evol. 8, 2195–2212. doi: 10.1038/s41559-024-02564-9
Nishida, S. and Christophel, D. C. (1999). Leaf anatomy of Beilschmiedia (Lauraceae) in the neotropics. Nat. Hum. Activities 4, 9–43. doi: 10.1007/978-3-662-02899-5_46
Oliveira, R. S., Eller, C. B., Bittencourt, P. R. L., and Mulligan, M. (2014). The hydroclimatic and ecophysiological basis of cloud forest distributions under current and projected climates. Ann. Bot. 113, 909–920. doi: 10.1093/aob/mcu060
Pandey, S. (2021). Climatic influence on tree wood anatomy: a review. J. Wood Sci. 67, 24. doi: 10.1186/s10086-021-01956-w
Pedersen, E. J., Miller, D. L., Simpson, G. L., and Ross, N. (2019). Hierarchical generalized additive models in ecology: An introduction with mgcv. PeerJ 7, e6876. doi: 10.7717/peerj.6876
Peters, M. K., Hemp, A., Appelhans, T., Behler, J., Classen, A., Detsch, F., et al. (2019). Climate–land-use interactions shape tropical mountain biodiversity and ecosystem functions. Nature 568, 88–92. doi: 10.1038/s41586-019-1048-z
Ponce-Reyes, R., Reynoso-Rosales, V. H., Watson, J. E. M., Vanderwal, J., Fuller, R. A., Pressey, R. L., et al. (2012). Vulnerability of cloud forest reserves in Mexico to climate change. Nat. Clim Chang 2, 448–452. doi: 10.1038/nclimate1453
Postek, M. T. (1981). The occurrence of silica in the leaves of Magnolia grandiflora L. Bot. Gaz 142, 124–134. doi: 10.1086/337202
Price, M. F., Gratzer, G., Duguma, L. A., Kohler, T., Maselli, D., and Romeo, R. (2011). Mountain Forests in a Changing World Mountain Forests in a Changing World Realizing values, addressing challenges (Rome: FAO/MPS and SDC).
Radtke, M. C., Pigg, K. B., and Wehr, W. C. (2005). Fossil Corylopsis and Fothergilla leaves (Hamamelidaceae) from the lower Eocene flora of Republic, Washington, U.S.A., and their evolutionary and biogeographic significance. Int. J. Plant Sci. 166, 347–356. doi: 10.1086/427483
Raffard, A., Lecerf, A., Cote, J., Buoro, M., Lassus, R., and Cucherousset, J. (2017). The functional syndrome: linking individual trait variability to ecosystem functioning. Proc. R Soc. B 284, 20171893. doi: 10.1098/rspb.2017.1893
Ramírez-Barahona, S., Cuervo-Robayo, Á. P., Feeley, K. J., Ortiz-Rodríguez, A. E., Vásquez-Aguilar, A. A., Ornelas, J. F., et al. (2025). Upslope plant species shifts in Mesoamerican cloud forests driven by climate and land use change. Science 387, 1058–1063. doi: 10.1126/science.adn2559
Ramírez-Díaz, M., Gutiérrez, J., and Terrazas, T. (2024). Leaf architecture and anatomy of eight species of Tilia (Malvaceae). Acta Bot. Mex. 131, e2332. doi: 10.21829/abm131.2024.2332
R Core Team (2018). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.r-project.org. (Accessed October 2, 2024)
Reyes-Ortiz, M., Lira-Noriega, A., Osorio-Olvera, L., Luna-Vega, I., and Williams-Linera, G. (2024). Leaf functional traits and ecological niche of Fagus grandifolia and Oreomunnea mexicana in natural forests and plantings as a proxy of climate change. Am. J. Bot. 111, 1–14. doi: 10.1002/ajb2.16322
Richter, T., Geres, L., König, S., Braziunas, K. H., Senf, C., Thom, D., et al. (2024). Effects of climate and forest development on habitat specialization and biodiversity in Central European mountain forests. Commun. Biol. 15, 1518. doi: 10.1038/s42003-024-07239-6
Robbins, Z., Chambers, J., Chitra-Tarak, R., Christoffersen, B., Dickman, L. T., Fisher, R., et al. (2024). Future climate doubles the risk of hydraulic failure in a wet tropical forest. New Phytol. 244, 2239–2250. doi: 10.1111/nph.19956
Rodríguez-Ramírez, E. C., Arroyo, F., Ames-Martínez, F. N., and Andrés-Hernández, A. R. (2024a). Tracking climate vulnerability across spatial distribution and functional traits in Magnolia gentryi from the Peruvian tropical montane cloud forest. Am. J. Bot. 11, 1–14. doi: 10.1002/ajb2.16400
Rodríguez-Ramírez, E. C., Crispín-DelaCruz, D. B., Morales, M. S., Ticse-Otarola, G. A., Ames-Martínez, F. N., Guerra, A., et al. (2023a). Exploring monsoon precipitation signal in the Peruvian tropical montane cloud forest: Andean Walnut wood anatomy plasticity. IAWA J. 45, 65–81. doi: 10.1163/22941932-bja10132
Rodríguez-Ramírez, E. C., Ferrero, M. E., Acevedo-Vega, I., Crispín-DelaCruz, D. B., Ticse-Otarola, G., and Requena-Rojas, E. J. (2022). Plastic adjustments in xylem vessel traits to drought events in three Cedrela species from Peruvian Tropical Andean forests. Sci. Rep. 12, 1–14. doi: 10.1038/s41598-022-25645-w
Rodríguez-Ramírez, E. C., Frei, J., Ames-Martínez, F. N., Guerra, A., and Andrés-Hernández, A. R. (2024b). Ecological stress memory in wood architecture of two Neotropical hickory species from central-eastern Mexico. BMC Plant Biol. 24, 1–15. doi: 10.1186/s12870-024-05348-2
Rodríguez-Ramírez, E. C., García-Morales, L. J., Alcántara-Ayala, O., Antonio Vázquez-García, J., and Luna-Vega, I. (2021a). Leaf vein morphological variation in four endangered Neotropical Magnolia species along an elevation gradient in the Mexican tropical montane cloud forests. Plants 10, 2595. doi: 10.3390/plants10122595
Rodríguez-Ramírez, E. C., Luna-Vega, I., and Rozas, V. (2018). Tree-ring research of Mexican beech (Fagus grandifolia subsp. mexicana) a relict tree endemic to eastern Mexico. Tree Ring Res. 74, 94–107. doi: 10.3959/1536-1098-74.1.94
Rodríguez-Ramírez, E. C., Martínez-Mejía, M., del, R., Ghimire, B., and Luna-Vega, I. (2021b). Response of leaf morphological traits of relict-endemic Symplocos species (S. coccinea and S. speciosa) to elevation and abiotic fluctuations. Plant Ecol. 222, 693–704. doi: 10.1007/s11258-021-01138-0
Rodríguez-Ramírez, E. C., Ruiz-Santiago, D., Requena-Rojas, E. J., Longhi-Santos, T., Suárez-Islas, A., and Luna-Vega, I. (2023b). Assessing the drought effect on tree rings’ width and xylem traits plasticity in Symplocos coccinea and S. speciosa: two rare and endemic Neotropical montane oak forest trees from eastern Mexico. Trees-Struct Funct. 37, 555–566. doi: 10.1007/s00468-022-02371-4
Rodríguez-Ramírez, E. C., Sánchez-González, A., and Ángeles-Pérez, G. (2016). Relationship between vegetation structure and microenvironment in Fagus grandifolia subsp. mexicana forest relicts in Mexico. J. Plant Ecol. 11, 237–247. doi: 10.1093/jpe/rtw138
Rodríguez-Ramírez, E. C., Terrazas, T., and Luna-Vega, I. (2019). The influence of climate on the masting behavior of Mexican beech: growth rings and xylem anatomy. Trees-Struct Funct. 33, 23–35. doi: 10.1007/s00468-018-1755-3
Rodríguez-Ramírez, E. C., Valdez-Nieto, J. A., Vázquez-García, J. A., Dieringer, G., and Luna-Vega, I. (2020a). Plastic responses of Magnolia schiedeana Schltdl., a relict-endangered Mexican cloud forest tree, to climatic events: Evidences from leaf venation and wood vessel anatomy. Forests 11, 1–18. doi: 10.3390/F11070737
Rodríguez-Ramírez, E. C., Vázquez-García, J. A., García-González, I., Alcántara-Ayala, O., and Luna-Vega, I. (2020b). Drought effects on the plasticity in vessel traits of two endemic Magnolia species in the tropical montane cloud forests of eastern Mexico. J. Plant Ecol. 13, 331–340. doi: 10.1093/jpe/rtaa019
Royer, D. L., Peppe, D. J., Wheeler, E. A., and Ülo, N. (2012). Roles of climate and functional traits in controlling toothed vs. untoothed leaf margins. Am. J. Bot. 99, 915–922. doi: 10.3732/ajb.1100428
Royer, D. L. and Wilf, P. (2006). Why do toothed leaves correlate with cold climates? Gas exchange at leaf margins provides new insights into a classic paleotemperature proxy. Int. J. Plant Sci. 167, 11–18. doi: 10.1086/497995
Saatchi, S., Longo, M., Xu, L., Yang, Y., Abe, H., André, M., et al. (2021). Detecting vulnerability of humid tropical forests to multiple stressors. One Earth 4, 988–1003. doi: 10.1016/j.oneear.2021.06.002
Saldaña-Acosta, A., Meave, J. A., Paz, H., Sánchez-Velásquez, L. R., Villaseñor, J. L., and Martínez-Ramos, M.. (2008). Variation of functional traits in trees from a biogeographically complex Mexican cloud forest. Acta Oecol. 34, 111–121. doi: 10.1016/j.actao.2008.04.004
Sánchez-Velásquez, L. R., Hernández-Vargas, G., Ibarra-Zavaleta, S. P., Pineda-López, M., del, R., Saldaña-Vázquez, R. A., et al. (2023). Climatic variability and plant functional traits in tropical montane cloud forest refugia along the Trans Mexican Volcanic Belt. Flora: Morphol Distrib Funct. Ecol. Plants 308, 152397. doi: 10.1016/j.flora.2023.152397
Scholbeck, C. A., Casalicchio, G., Molnar, C., Bischl, B., and Heumann, C. (2024). Marginal effects for non-linear prediction functions. Data Min Knowl. Discov. 38, 2997–3042. doi: 10.1007/s10618-023-00993-x
Scholz, A., Stein, A., Choat, B., and Jansen, S. (2014). How drought and deciduousness shape xylem plasticity in three Costa Rican woody plant species. IAWA J. 35, 337–355. doi: 10.1163/22941932-00000070
Schuldt, B., Leuschner, C., Brock, N., and Horna, V. (2013). Changes in wood density, wood anatomy and hydraulic properties of the xylem along the root-to-shoot flow path in tropical rainforest trees. Tree Physiol. 33, 161–174. doi: 10.1093/treephys/tps122
Šmilauer, P. and Lepš, J. (2014). Multivariate analysis of ecological data using CANOCO 5. 2nd. Eds. Šmilauer, P. and Lepš, J. (Czech Republic: Cambridge University Press). doi: 10.1017/CBO9781139627061
Sobral, M. (2021). All traits are functional: An evolutionary viewpoint. Trends Plant Sci. 26, 674–676. doi: 10.1016/j.tplants.2021.04.004
Taylor, K. E., Stouffer, R. J., and Meehl, G. A. (2012). An overview of CMIP5 and the experiment design. Bull. Am. Meteorol Soc. 93, 485–498. doi: 10.1175/BAMS-D-11-00094.1
Thybring, E. E. and Fredriksson, M. (2021). Wood modification as a tool to understand moisture in wood. Forests 12, 372. doi: 10.3390/f12030372
Tng, D. Y. P., Apgaua, D. M. G., Ishida, Y. F., Mencuccini, M., Lloyd, J., Laurance, W. F., et al. (2018). Rainforest trees respond to drought by modifying their hydraulic architecture. Ecol. Evol. 8, 12479–12491. doi: 10.1002/ece3.4601
Toledo-Aceves, T., García-Hernández, M., de los, Á., and Paz, H. (2019). Leaf functional traits predict cloud forest tree seedling survival along an elevation gradient. Ann. For Sci. 76, 111. doi: 10.1007/s13595-019-0900-5
Vásquez-Morales, S. G., Téllez-Valdés, O., Del Rosario Pineda-López, M., Sánchez-Velásquez, L. R., Flores-Estevez, N., and Viveros-Viveros, H. (2014). Effect of climate change on the distribution of Magnolia schiedeana: A threatened species. Bot. Sci. 92, 575–585. doi: 10.17129/botsci.116
Vasseur, D. A., DeLong, J. P., Gilbert, B., Greig, H. S., Harley, C. D. G., McCann, K. S., et al. (2014). Increased temperature variation poses a greater risk to species than climate warming. Proc. R Soc. B-Biol Sci. 281, 20132612. doi: 10.1098/rspb.2013.2612
Viliani, L., Bonelli, S., Gentile, G., Parile, E., and Riva, F. (2024). Diverging sub-fields in functional ecology. Front. Ecol. Evol. 12. doi: 10.3389/fevo.2024.1267605
Vogt, U. K. (2001). Hydraulic vulnerability, vessel refilling, and seasonal courses of stem water potential of Sorbus aucuparia L. and Sambucus nigra L. J. Exp. Bot. 52, 1527–1536. doi: 10.1093/jexbot/52.360.1527
von Arx, G., Arzac, A., Olano, J. M., and Fonti, P. (2015). Assessing conifer ray parenchyma for ecological studies: Pitfalls and guidelines. Front. Plant Sci. 6. doi: 10.3389/fpls.2015.01016
von Arx, G., Kueffer, C., and Fonti, P. (2013). Quantifying plasticity in vessel grouping - Added value from the image analysis tool ROXAS. IAWA J. 34, 433–445. doi: 10.1163/22941932-00000035
Wang, Y., Naumann, U., Wright, S. T., and Warton, D. I. (2012). Mvabund- an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 3, 471–474. doi: 10.1111/j.2041-210X.2012.00190.x
Wang, Y., Sun, F., Lin, W., and Zhang, S. (2022b). AC-PCoA: Adjustment for confounding factors using principal coordinate analysis. PloS Comput. Biol. 18, 1–21. doi: 10.1371/journal.pcbi.1010184
Wang, H., Wang, R., Harrison, S. P., and Prentice, I. C. (2022a). Leaf morphological traits as adaptations to multiple climate gradients. J. Ecol. 110, 1344–1355. doi: 10.1111/1365-2745.13873
Warton, D. I., Shipley, B., and Hastie, T. (2015). CATS regression - a model- based approach to studying trait-based community assembly. Methods Ecol. Evol. 6, 389–398. doi: 10.1111/2041-210X.12280
Weigelt, A., Mommer, L., Andraczek, K., Iversen, C. M., Bergmann, J., Bruelheide, H., et al. (2021). An integrated framework of plant form and function: the belowground perspective. New Phytol. 232, 42–59. doi: 10.1111/nph.17590
Wickham, H. (2016). ggplot2: elegant graphics for data analysis. 2nd. Eds. Gentleman, R., Hornik, K., and Parmigiani, G. (Houston, Texas, USA: Springer). doi: 10.1007/978-0-387-98141-3
Wood, S. N. (2017). Generalized additive models: An introduction with R, 2nd. doi: 10.1201/9781315370279
Wood, S. N. (2023). mgcv: Mixed GAM computation vehicle with automatic smoothness estimation (Vienna, Austria: R Foundation for Statistical Computing). Available online at: https://cran.r-project.org/package=mgcv (Accessed October 10, 2024).
Wright, I. J., Dong, N., Maire, V., Prentice, I. C., Westoby, M., Díaz, S., et al. (2017). Global climatic drivers of leaf size Downloaded from. Available online at: http://science.sciencemag.org/ (Accessed November 23, 2024).
Wright, A. J. and Francia, R. M. (2024). Plant traits, microclimate temperature and humidity: A research agenda for advancing nature-based solutions to a warming and drying climate. J. Ecol. 112, 1–9. doi: 10.1111/1365-2745.14313
Yang, Y., Xiao, C., Wu, X., Long, W., Feng, G., and Liu, G. (2021). Differing trade-off patterns of tree vegetative organs in a tropical cloud forest. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.680379
Yang, X., Yan, H., Hao, C., Hu, J., Yang, G., An, S., et al. (2024). Climate of origin shapes variations in wood anatomical properties of 17 Picea species. BMC Plant Biol. 24, 1–17. doi: 10.1186/s12870-024-05103-7
Zahedi, S. M., Karimi, M., Venditti, A., Zahra, N., Siddique, K. H. M., and Farooq, M. (2024). Plant adaptation to drought stress: The role of anatomical and morphological characteristics in maintaining the water status. J. Soil Sci. Plant Nutr 24, 1113–1132. doi: 10.1007/s42729-024-02141-w
Zhang, C., Gu, R., and Russo, S. E. (2024). Functional traits and ecological niches as correlates of the interspecific growth–mortality trade-off among seedlings of 14 tropical tree species. Funct. Ecol. 38, 1888–1901. doi: 10.1111/1365-2435.14624
Keywords: climate variability, drought stress, endangered species acclimation, environmental adaptation, wood anatomy, leaf morphological traits
Citation: Andrés-Hernández AR and Rodríguez-Ramírez EC (2025) Does climatic variation drive the adjustment of functional traits? An assessment of Tropical Montane Cloud Forest tree species. Front. Plant Sci. 16:1555607. doi: 10.3389/fpls.2025.1555607
Received: 05 January 2025; Accepted: 20 May 2025;
Published: 04 June 2025.
Edited by:
Ignacio García-González, University of Santiago de Compostela, SpainReviewed by:
Kyriaki Giagli, Mendel University in Brno, CzechiaDavid Law, Lakehead University, Canada
Copyright © 2025 Andrés-Hernández and Rodríguez-Ramírez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ernesto C. Rodríguez-Ramírez, ZXJvZHJpZ3VlenJAY29udGluZW50YWwuZWR1LnBl