 Freya Maria Rosemarie Ziegler1,2*
Freya Maria Rosemarie Ziegler1,2* Amelia Gaston3
Amelia Gaston3 Karine Guy4Marie Devers3Erika Krüger5Bastienne Brauksiepe6
Karine Guy4Marie Devers3Erika Krüger5Bastienne Brauksiepe6 Klaus Eimert5
Klaus Eimert5 Sonia Osorio7
Sonia Osorio7 Béatrice Denoyes3*
Béatrice Denoyes3* Björn Usadel1,2*
Björn Usadel1,2*- 1Forschungszentrum Jülich, Institute of Bio- and Geosciences, Institute for Bio- and Geosciences (IBG)-4 Bioinformatics, Bioeconomy Science Center (BioSC), Jülich, Germany
- 2Heinrich-Heine University Düsseldorf, Faculty of Mathematics and Natural Sciences, Institute for Biological Data Science, Cluster of Excellence on Plant Sciences (CEPLAS), Düsseldorf, Germany
- 3Université de Bordeaux, National Research Institute for Agriculture, Food and Environment (INRAE), Biologie du Fruit et Pathologie, UMR 1332, Bordeaux, France
- 4INVENIO, MIN de Brienne, Bordeaux, France
- 5Department of Pomology, Hochschule Geisenheim University, Geisenheim, Germany
- 6Department of Molecular Plant Science, Hochschule Geisenheim University, Geisenheim, Germany
- 7Instituto de Hortofruticultura Subtropical y Mediterránea “La Mayora”, Departamento de Biología Molecular y Bioquímica, Universidad de Málaga-Consejo Superior de Investigaciones Científicas, Málaga, Spain
Floral initiation is essential for sexual reproduction in angiosperms and plays a critical role in determining crop yields. In cultivated strawberry, however, the molecular mechanisms underlying floral initiation remain poorly understood, with most studies focusing on a single genotype under controlled conditions. To gain more insight into this process, we conducted a field-based study in two countries using two seasonal flowering cultivars. Our investigation focused on the early stages of floral initiation, analyzing samples that captured key developmental stages of the shoot apical meristem. Differential gene expression analysis of leaf and terminal bud tissues revealed an enrichment of genes involved in carbohydrate metabolism and phytohormone signaling pathways in leaves. Additionally, we observed enrichment of protein classes related to cytoskeleton organization, cell cycle regulation, and chromatin structure during early floral initiation. We also identified genes associated with the photoperiodic pathway, including well-known floral integrators such as TFL1 and SOC1, along with several other genes linked to phytohormone regulation, such as XTH23, PP2 and EIN3.
Introduction
Floral initiation is a pivotal event in the life cycle of angiosperm plants that marks the transition from vegetative to reproductive development. It occurs in the shoot apical meristem (SAM), which generates leaves, shoots and flowers (Benlloch et al., 2007; Figure 1A). Flowering is necessary for reproductive success, significantly affecting pollination, fruit production, and crop yields (Weberling, 1989; Wyatt, 1982).
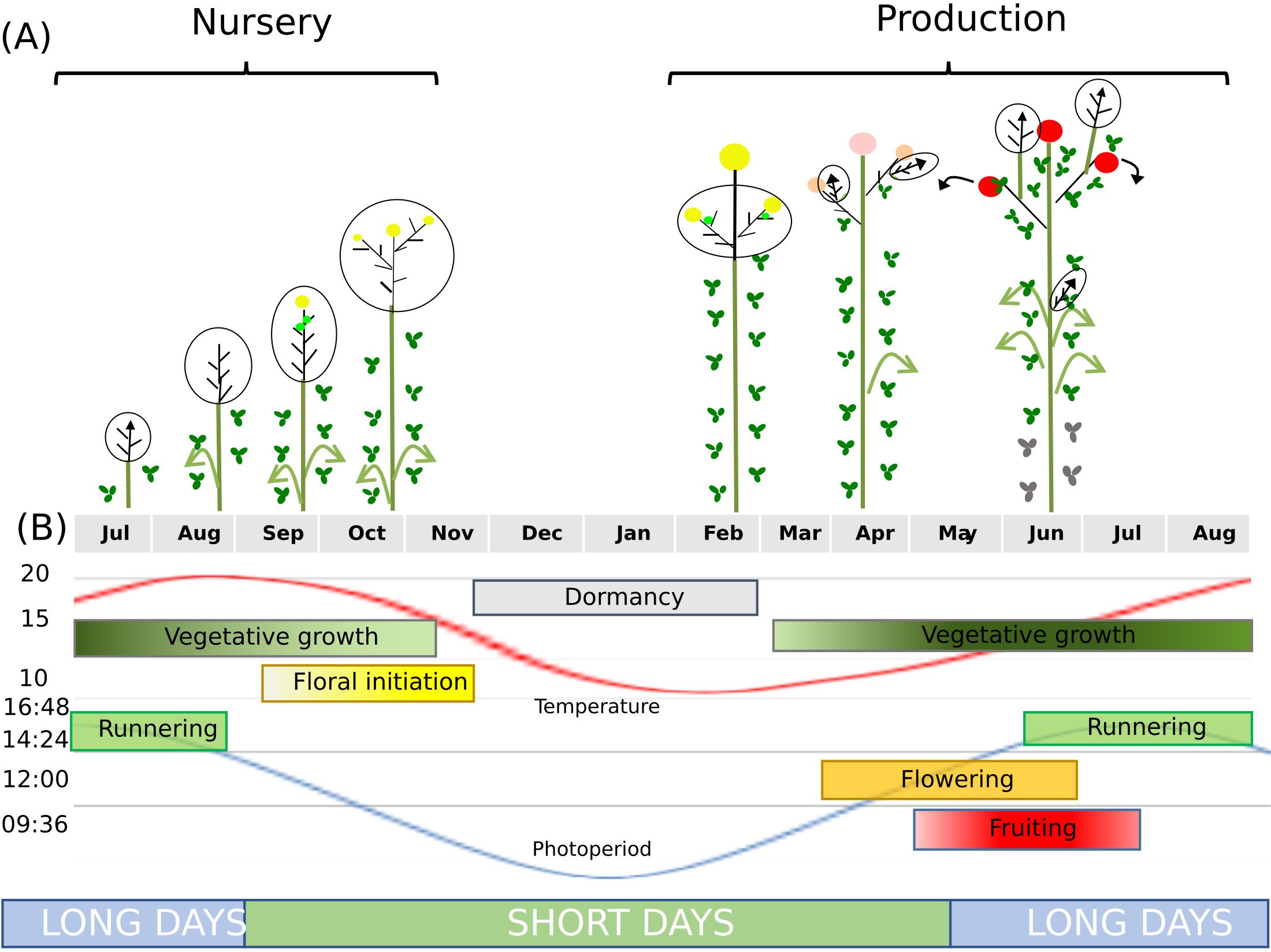
Figure 1. Developmental processes in seasonal flowering varieties of cultivated octoploid strawberry. (A) In the nursery, temperatures and day length decline, allowing floral initiation to occur. The shoot apical meristem transitions from vegetative (arrow) to floral development (yellow sphere). Because strawberry is a rosette, the terminal bud is not visible and is represented here within the dotted line at the base of the plant. (B) Runnering occurs on long days. Dormancy occurs at the end of autumn and during winter, when temperatures do not allow vegetative development. The dashed circle indicates the terminal bud, which includes the foliar primordia.
In strawberry (Fragaria × ananassa), the most widely cultivated berry crop, the fruit yield depends on the timing and duration of floral initiation (Costes et al., 2014). Most strawberry genotypes are seasonal flowering (also called short-day, single cropping or June bearing) varieties, with a single fruiting period in the spring of year N. Floral initiation occurs during the previous autumn (year N – 1) and is triggered by declining temperatures and day length at the end of summer/beginning of autumn (Heide et al., 2013; Figure 1B). Floral initiation is followed by floral development, specifically the organogenesis of flowers in the inflorescence. Following dormancy and the fulfilment of chilling requirements, flowers initiated in autumn begin to emerge in spring.
Strawberry plants also reproduce asexually via stolons, which are elongated stems bearing daughter plants (Tenreira et al., 2017). The choice between sexual and asexual reproduction is made in the axillary meristem, which can produce either an inflorescence-bearing branch or a stolon, and thus determines two antagonistic traits: fruit yield, a major trait for producers, and daughter-plant yield, a major trait for nurseries (Tenreira et al., 2017; Gaston et al., 2021).
The interplay between photoperiod and temperature, and their combined influence on floral initiation have been extensively documented (Stewart and Folta, 2010; Heide et al., 2013). The overarching consensus is that optimal conditions for floral initiation involve temperatures of 12–18°C and photoperiods of 10–12 h maintained over a period of 3–4 weeks. Global radiation also contributes to floral initiation (Krüger et al., 2022). Variations in these parameters are mainly due to cultivar-specific responses (Verheul et al., 2006; Krüger et al., 2022).
Recent work has begun to identify and characterize the strawberry genes controlling floral initiation, plant architecture and yield (Hytonen and Kurokura, 2020). The CENTRORADIALIS/TERMINAL FLOWER 1/SELF-PRUNING (CETS) family, represented by TFL1/FT genes, plays a pivotal role (Wickland and Hanzawa, 2015; Kurokura et al., 2017; Gaston et al., 2021). TFL1 acts as a floral repressor in both diploid Fragaria vesca (Iwata et al., 2011; Koskela et al., 2012) and cultivated strawberry (Koskela et al., 2016), and its expression declines during floral initiation in cultivar Benihoppe (Liang et al., 2022). Three strawberry FT genes have also been identified. FveFT1 encodes a long-day floral activator, as shown in the tfl1 diploid genetic background (Koskela et al., 2012; Rantanen et al., 2014). FveFT2 encodes a non-photoperiodic florigen (Gaston et al., 2021) and operates in tandem with the photoperiodic anti-florigen FveTFL1 (Gaston et al., 2021). Overexpression of FveFT2 confers a very early flowering phenotype. In octoploid cultivated strawberry, FanFT3 encodes a floral repressor (Koembuoy et al., 2020), and FveFT3 overexpression promotes plant branching in the tfl1 diploid genetic background.
The regulation of flowering is intricately linked to the production of stolons. Gibberellin biosynthesis and signaling affect strawberry plant architecture and fruit yield by specifying whether the axial meristem produces a stolon or an inflorescence-bearing branch crown (Tenreira et al., 2017; Caruana et al., 2018). The natural mutation in FveGA20ox4 generates a runnerless phenotype (Tenreira et al., 2017), which can be reversed by mutating FveRGA1 (REPRESSOR OF GIBBERELIC ACID1), encoding a DELLA protein (Caruana et al., 2018). The molecular mechanisms governing the balance between sexual and asexual reproduction in the axial meristem are only beginning to be understood.
The molecular control of floral initiation in the cultivated strawberry F. × ananassa has yet to be studied in detail under natural conditions. The primary objective of this study was to identify and characterize the early molecular events associated with floral initiation under field conditions using transcriptomic analysis. A secondary objective was to evaluate how environmental variables (e.g. photoperiod) and genetic background influence gene expression during the transition from vegetative to reproductive development. Here, we examined the early molecular events of floral initiation by transcriptomic analysis, taking into account both the genotype and environment by studying two cultivars – the Italian cultivar Clery (CL) and the French cultivar Gariguette (GA) – at two locations, one in France and one in Germany. We analyzed RNA-Seq data from leaf and terminal bud tissues during the transition from the vegetative phase to the early stages of floral initiation in the SAM (weeks 29, 32, 33 and 35). As anticipated, we found that both cultivars were enriched during the early period of floral initiation for differentially expressed genes (DEGs) encoding proteins involved in flowering, but also those relating to chromatin structure, cytoskeletal organization, serine/threonine phosphatase signaling, cell division, cell wall organization, and RNA biosynthesis.
Results
Phenotypic variation of floral initiation according to the cultivar and environment
We investigated floral initiation over time by dissecting the terminal bud in cultivars CL and GA and observing the SAM from the middle of July until the end of October over 3 years (2016–2018) in two different environments in France and Germany. The SAM in the terminal bud was always vegetative on the first sampling date in July regardless of the genotype or location (Figure 2). The SAM transitioned to a floral identity when its apical dome rose above the level of the developing stipules (Krüger et al., 2022).
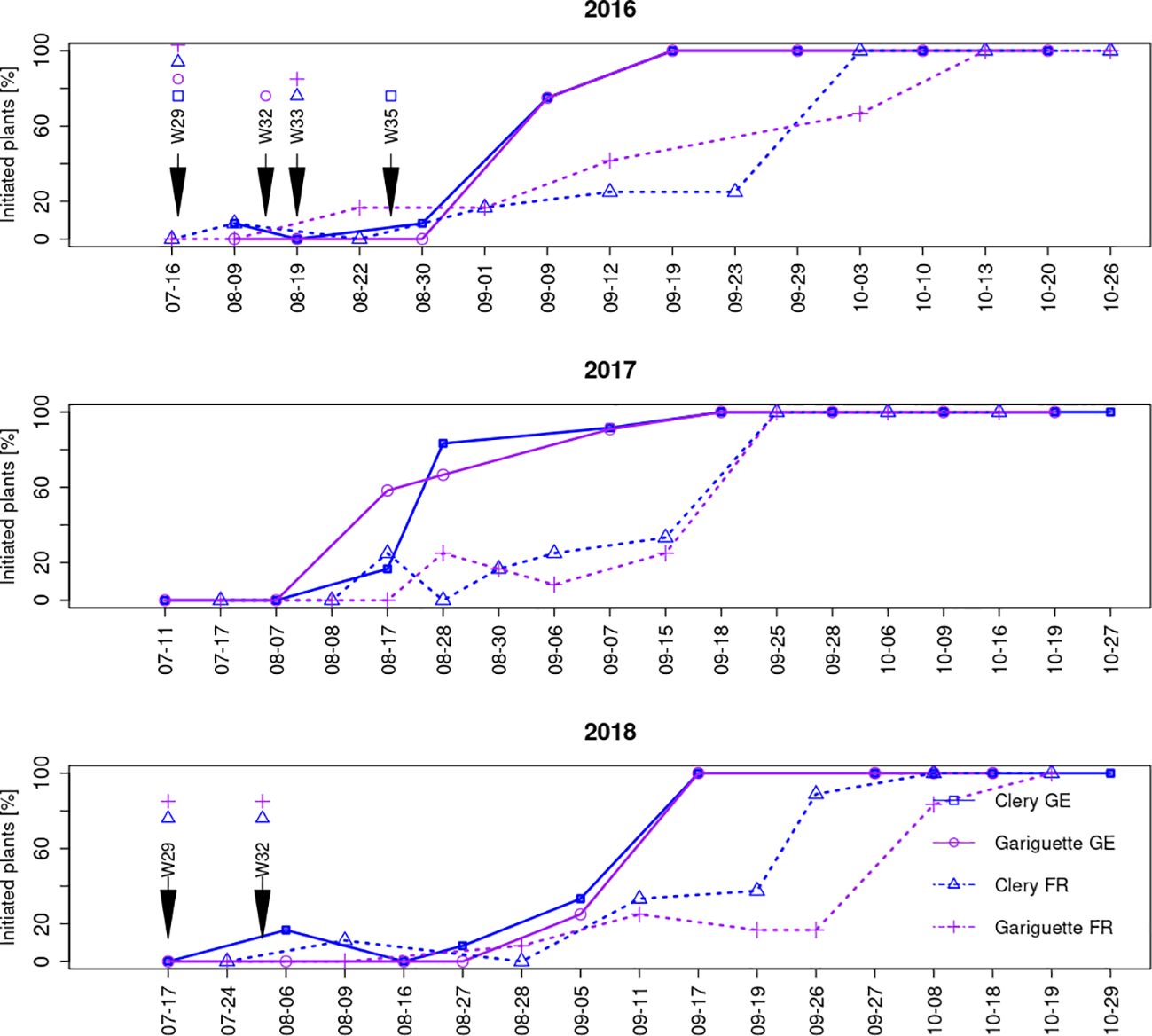
Figure 2. The frequency of plants undergoing floral initiation over time according to cultivar and location. Floral initiation was confirmed when the SAM reached at least stage 2 (terminal bud dome has risen above the developing stipules). Sampling dates for the RNA-Seq analysis of leaf and terminal bud tissues are indicated by arrows labeled with the sampling week: w29 (T0), w32 (T10A), w33 (T10B) and w35 (T50). FR, France; GE, Germany.
Because different plants were tested in each sample, the percentage of initiated plants (those with a floral terminal bud) fluctuated between consecutive sampling dates. The beginning of floral initiation was similar in France and Germany, with a low percentage (<10%) of initiated plants during the first weeks, rising steadily and then jumping from 20–40% to 90–100% in a single week (Figure 2). In Germany, this week was in the middle or at the end of August (depending on the year) whereas in France it was in the middle or at the end of September (Figure 2).
Floral initiation was followed by organogenesis in the SAM, resulting in the formation of an entire inflorescence with differentiated flower organs (still in the terminal bud, Figure 1). SAM development into an inflorescence followed the same tendency every year at both locations, although it occurred earlier for genotype CL than GA, and earlier in Germany than in France, as anticipated given the earlier floral initiation in Germany (Figure 3).
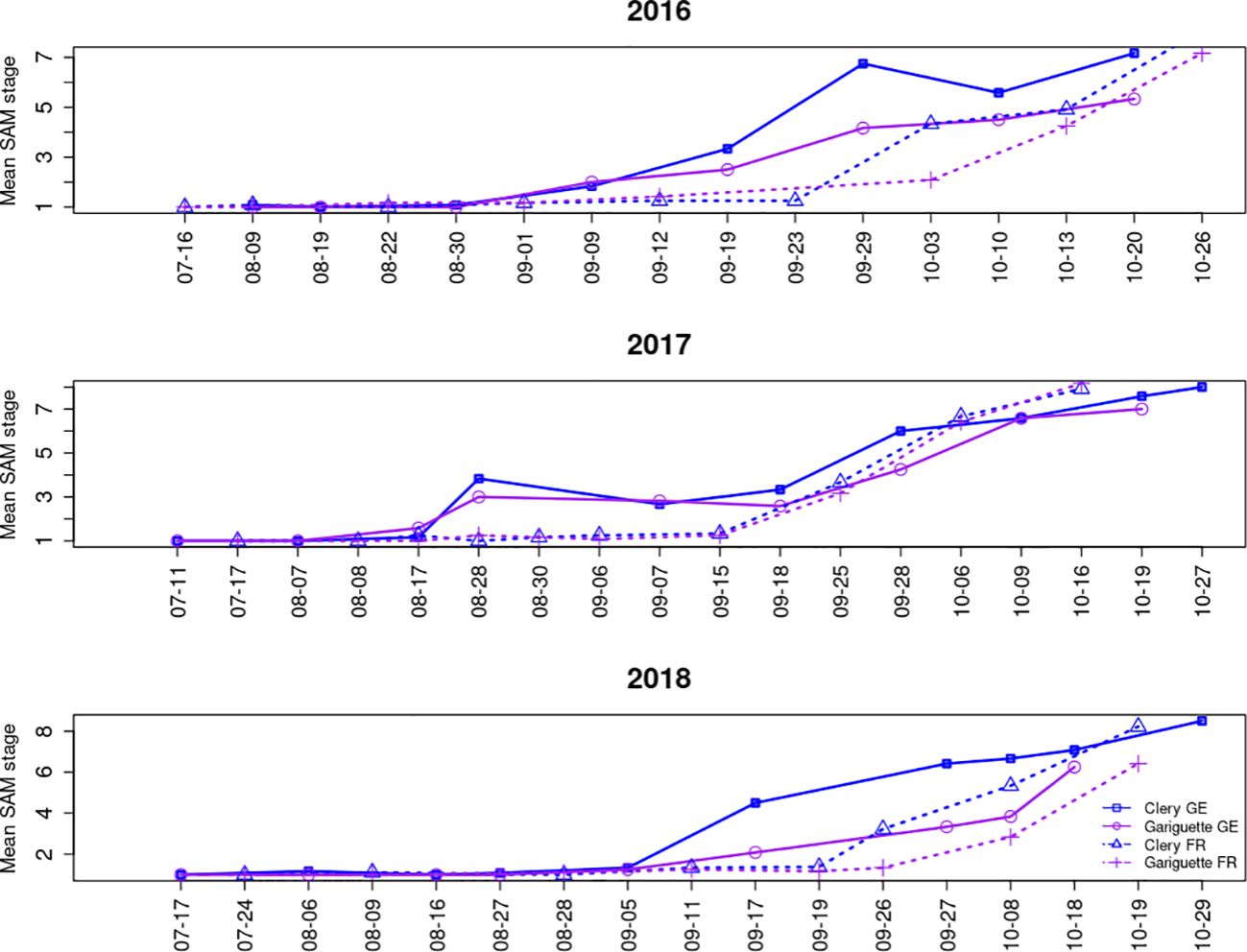
Figure 3. The mean floral developmental stage of the shoot apical meristem over time for the strawberry cultivars Clery and Gariguette in Germany (GE) and France (FR). SAM staging is described in Supplementary Figure S1A.
Choosing relevant sampling dates to study early floral initiation
We collected samples for RNA-Seq at three time points, when all plants were vegetative: T0 with no plants initiated, T10 when ~10% of the plants were initiated, and T50 when ~50% of the plants were initiated. Given the effects of genotype and environment and their interaction noted above and reviewed by Heide et al., 2013, the time points differed by genotype, location and year (Table 1; Figure 2). We also observed that the development of a given genotype could be discontinuous between two dates because the acquisition of the stage of the terminal bud require dissection and is destructive. At each date of observation, nine new plants of the same genotype were analyzed. For T0 samples, leaves and terminal buds were collected from daughter plants maintained on their mother plants during w29 or w30. The sample names were based on genotype, location/year and sampling time point as follows: CL_GE16_T0, GA_GE16_T0, CL_FR16_T0, GA_FR16_T0, CL_FR18_T0, and GA_FR18_T0.
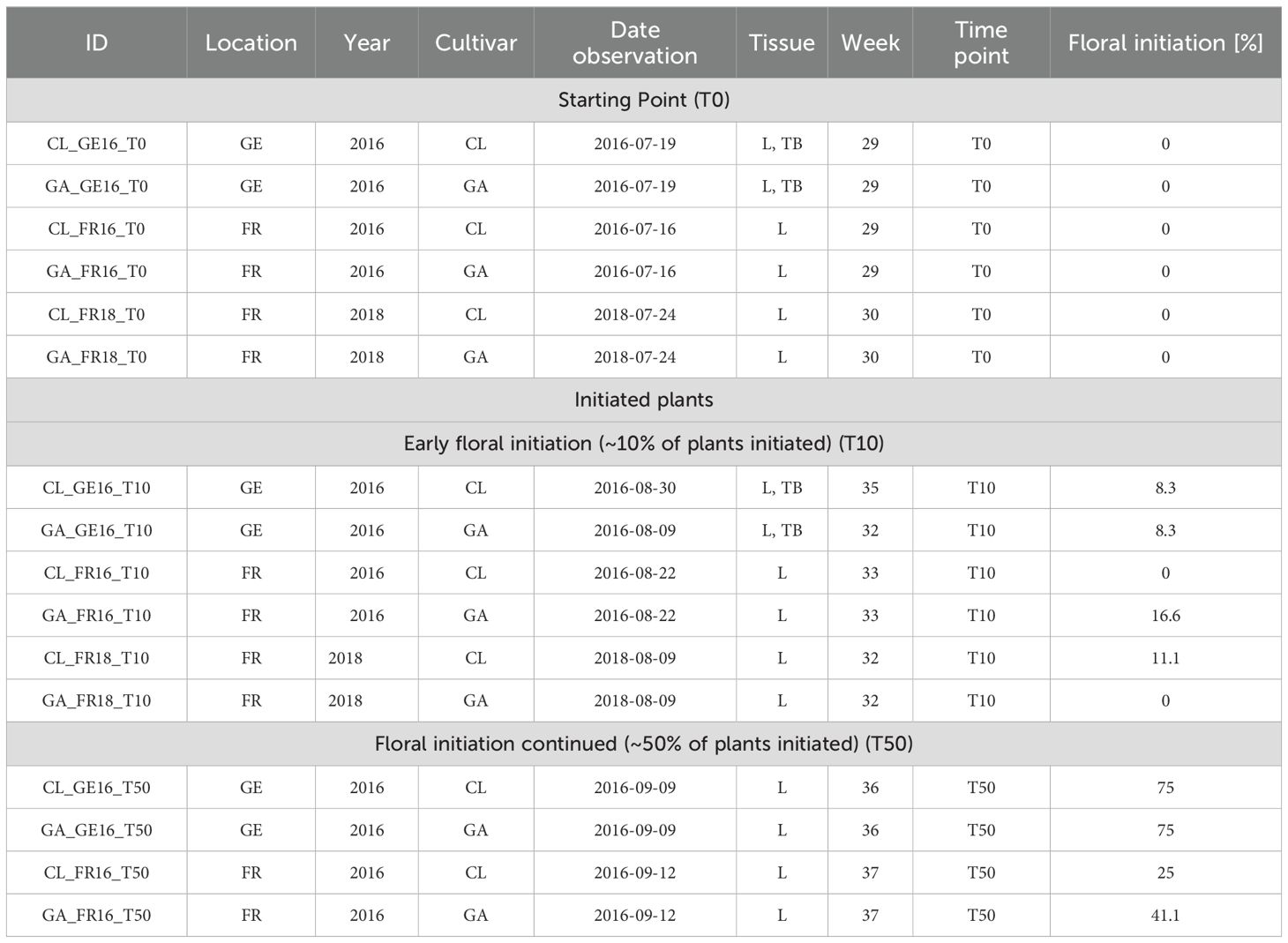
Table 1. Frequency of floral initiation [%] in samples of strawberry leaf (L) and terminal bud (TB) acquired in France (FR) and Germany (GE) over 3 years for the cultivars Clery (CL) and Gariguette (GA).
For T10 samples, leaves and terminal buds were collected when the percentage of initiated plants varied from 0% to 17%. A 0% frequency of initiated plants was observed in sample CL_FR16, but the percentage was 8.3% 10 days before, and in sample GA_FR18, where the percentage increased to 11% the week after (Figure 1; Table 1). The sample names were CL_GE16_T10, GA_GE16_T10, CL_FR16_T10, GA_FR16_T10, CL_FR18_T10, and GA_FR18_T10. We recovered four T50 samples: CL_GE16_T50, GA_GE16_T50, CL_FR16_T50, and GA_FR16_T50.
Transcriptome variation in strawberry tissues during floral initiation
To find differences and similarities between cultivars during early floral initiation, the RNA-Seq samples for leaf and terminal bud were compared using a combination of principal component analysis (PCA) and overrepresentation analysis (ORA). All three replicates of each sample were grouped, confirming their homogeneity. For the leaf samples (Figure 4), the gene expression patterns for cultivars CL and GA grown in France were initially similar (CL_FR16_T0 and GA_FR16_T0) but began to segregate along the PC1 axis by stage T10 (CL_FR16_T10A and GA_FR16_T10A). This was attributed to genes associated with photosynthesis, uptake of transition metal ions, phytohormone activity, lipid metabolism, and RNA biosynthesis. Notably, for both cultivars at both locations at stage T10, a more pronounced separation was observed along the PC2 axis. The separation patterns of both cultivars aligned closely across these time points (Figure 4).
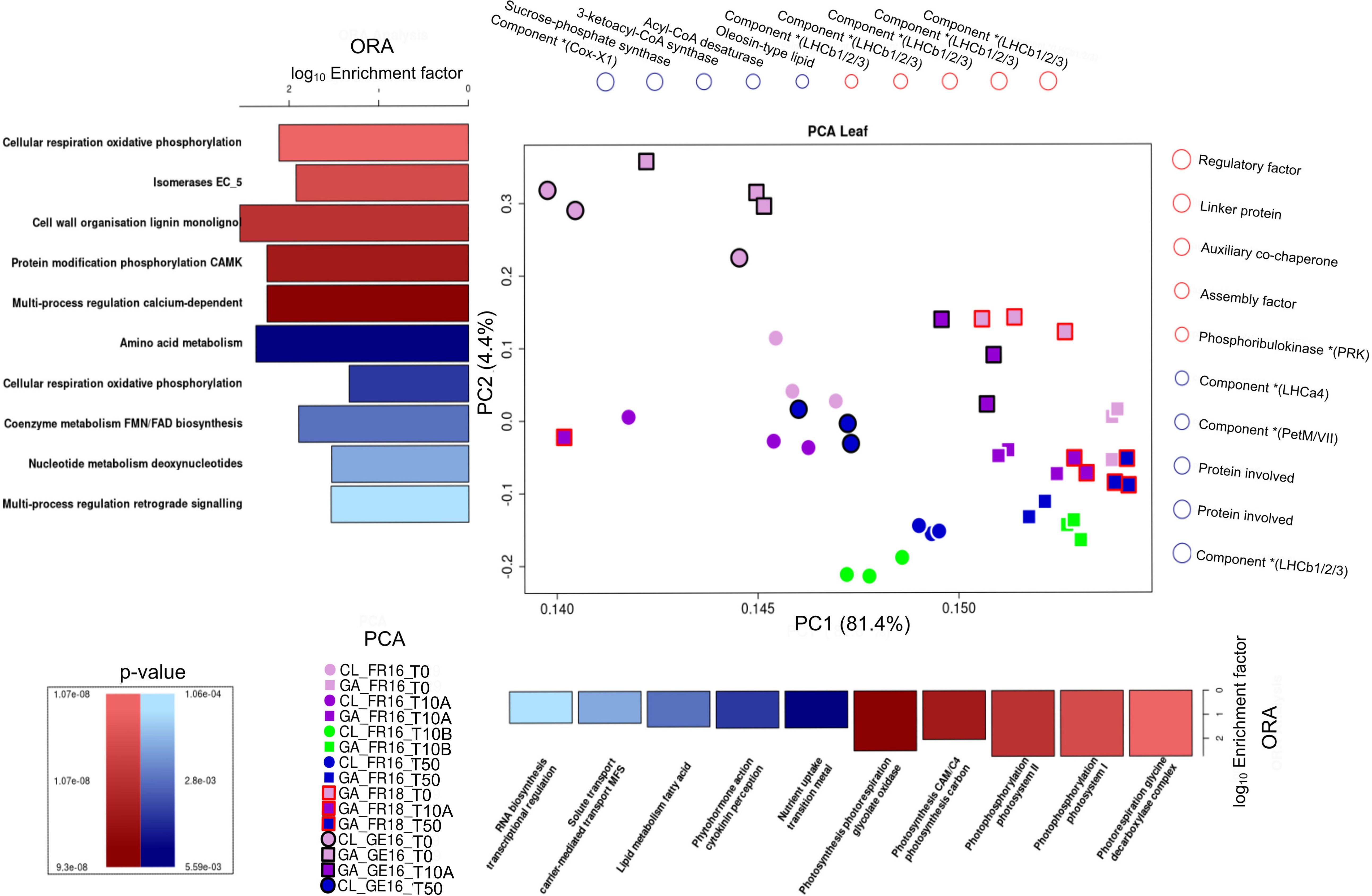
Figure 4. Principal component analysis (PCA) of differentially expressed genes (DEGs) in the leaves of cultivars Clery (CL) and Gariguette (GA) grown in France in 2016 (FR16) and 2018 (FR18), and in Germany in 2016 (GE16), sampled at time points T0 (week 29), T10A week 32), T10B (week 33) and T50 (week 35). Overrepresentation analysis (ORA) was then applied using MapMan protein classes based on the loadings for principal components 1 (PC1) and 2 (PC2). The top 5 MapMan protein classes resulting from ORA (p < 0.01), and their involvement in the separation along PC1 and PC2 in the positive (red) and negative (blue) directions, are shown as bar plots of the log10 enrichment factor. Genes strongly associated with the positive (red) or negative (blue) separation determined by PCA are depicted above (PC1) and to the right (PC2). In the PCA plots, circles and squares represent cultivars CL and GA, respectively. Frame colors represent the location/year: none = France 2016, black = Germany 2016, red = France 2018. Block colors represent the stage: light purple = T0, dark purple = T10A, green = T10B, blue = T50.
In Germany, cultivars CL and GA were separated along the PC2 axis from the first time point T0 (CL_GE16_T0 and GA_GE16_T0), particularly for protein classes involved in cellular respiration, cell wall organization, protein modification, and multi-process regulation (Figure 4). But by stage T10, the cultivars were dispersed clusters, which disguised the separation between them (CL_GE16_T10A and GA_GE16_T10A).
For the terminal bud (Supplementary Figure S3), the floral initiation of GA_FR18 was delineated along the PC1 axis. The separation observed at T0, T10 and T50 was mainly influenced by protein classes related to protein homeostasis, biosynthesis, and multi-process regulation of phosphoinositide. Moreover, the segregation at stage T10A also affected the PC2 axis, and involved protein classes related to cell wall organization (lignin), RNA processing, and secondary metabolism (phenolics). Analogous outcomes were observed for GA_GE16_T0 and GA_GE16_T10A, as well as for CL_GE16_T0 and CL_GE16_T50. Although the cultivars were clearly separated along the PC1 axis at stage T0, the results for both cultivars were similar at stages T10 and T50.
Common DEGs between T0 and T10 during early floral initiation
We focused on early floral initiation under natural conditions by screening for genes that were differentially expressed between the two first time points: T0 (w29) and T10 (w32, w33 or w35 according to the country and the year). We compared pairs of transcript profiles between T0 and T10 samples within each cultivar (CL or GA) and within each organ (leaf or terminal bud). The comparisons are represented by logical names. For example, CL_GE16_T0xT10 refers to the comparison of samples CL_GE16_T0 and CL_GE16_T10. Suffixes L and TB were added to represent leaf and terminal bud tissues, respectively. We identified 71,369 DEGs across all comparisons (FDR-corrected p ≤ 0.05 in all cases).
For leaf samples, the comparison of stages T0 and T10 for cultivar CL grown in Germany (CL_GE16_T0xT10L) and France (CL_FR16_T0xT10L) revealed 7118 DEGs (Supplementary Figure S2A; Supplementary Table S1). The analogous comparisons for cultivar GA (GA_GE16_T0xT10L, GA_FR16_T0xT10L and GA_FR18_T0xT10L) revealed 786 DEGs (Supplementary Figure S2B; Supplementary Table S2). The two cultivars shared 416 common DEGs expressed in leaves (Supplementary Table S3).
For terminal bud samples, the comparisons for cultivar CL (CL_GE16_T0xT10TB) revealed 3385 DEGs (Supplementary Table S4), whereas those for cultivar GA (GA_GE16_T0xT10TB and GA_FR18_T0xT10TB) revealed 1548 DEGs (Supplementary Table S5). The two cultivars shared 1207 common DEGs expressed in the terminal bud (Supplementary Figure S2C; Supplementary Table S6).
Enrichment analysis for MapMan protein classifications was applied to the common DEGs. In CL leaves, the main enriched classes (Figure 5) were plant reproduction, modulation of flowering, enzyme activity, secondary metabolism (terpenoids), and carbohydrate metabolism (starch). Conversely, the main enriched classes in GA leaves were multi-stress responses, cytoskeletal organization (microtubule network), chromatin organization/structure, phytohormone activity, carbohydrate metabolism and RNA biosynthesis (Figure 5). The DEGs shared by both cultivars were enriched for proteins related to solute transport channels, cell wall organization (pectin, rhamnogalacturonan), cell division and photosynthesis (Figure 5).

Figure 5. Overrepresentation analysis (ORA) of common genes expressed in the leaf and terminal bud (meristem) tissues (separated by dotted line) of cultivars Clery (CL), Gariguette (GA), or both (CL+GA). Bubble chart shows the log2 enrichment factor (log2 ERF) for MapMan protein annotation (y-axis) for common genes and the green color gradient represents the adjusted p-value for enriched protein classes.
In the CL terminal bud, the enriched protein classes were similar to those found in leaves, including carbohydrate metabolism, aspartate metabolism, and cell wall organization (hydroxyproline-rich glycoprotein). The DEGs were also enriched for RNA biosynthesis (bZIP superfamily), the circadian clock system, and photosynthesis (Figure 5). In the GA terminal bud, the most enriched protein classes were related to secondary metabolism (terpenoids), responses to external stimuli, cell wall organization (pectin, rhamnogalacturonan), redox homeostasis and photosynthesis (Figure 5). The DEGs shared by both cultivars were enriched for protein classes involved in chromatin organization/structure, cytoskeletal organization (microtubule network), serine/threonine phosphatase superfamily, cell division, cell wall organization (cellulose), RNA biosynthesis (bZIP superfamily) and plant reproduction/flowering (Figure 5).
The geographical location significantly affected the differences observed during floral initiation in both tissues. Protein classes involved in multi-process signaling (phosphoinositides), cytoskeletal organization, and cellular respiration differed most between Germany and France.
DEGs in early floral initiation highlight known genes involved in the flowering pathway
The common DEGs shared by different genotypes and tissues included those encoding transcription factors and signaling proteins related to plant reproduction and flowering. Some displayed the same tendency regardless of location whereas others showed different behaviors in Germany and France, or in leaves and the terminal bud, or combinations of the location and tissue (Figure 6).
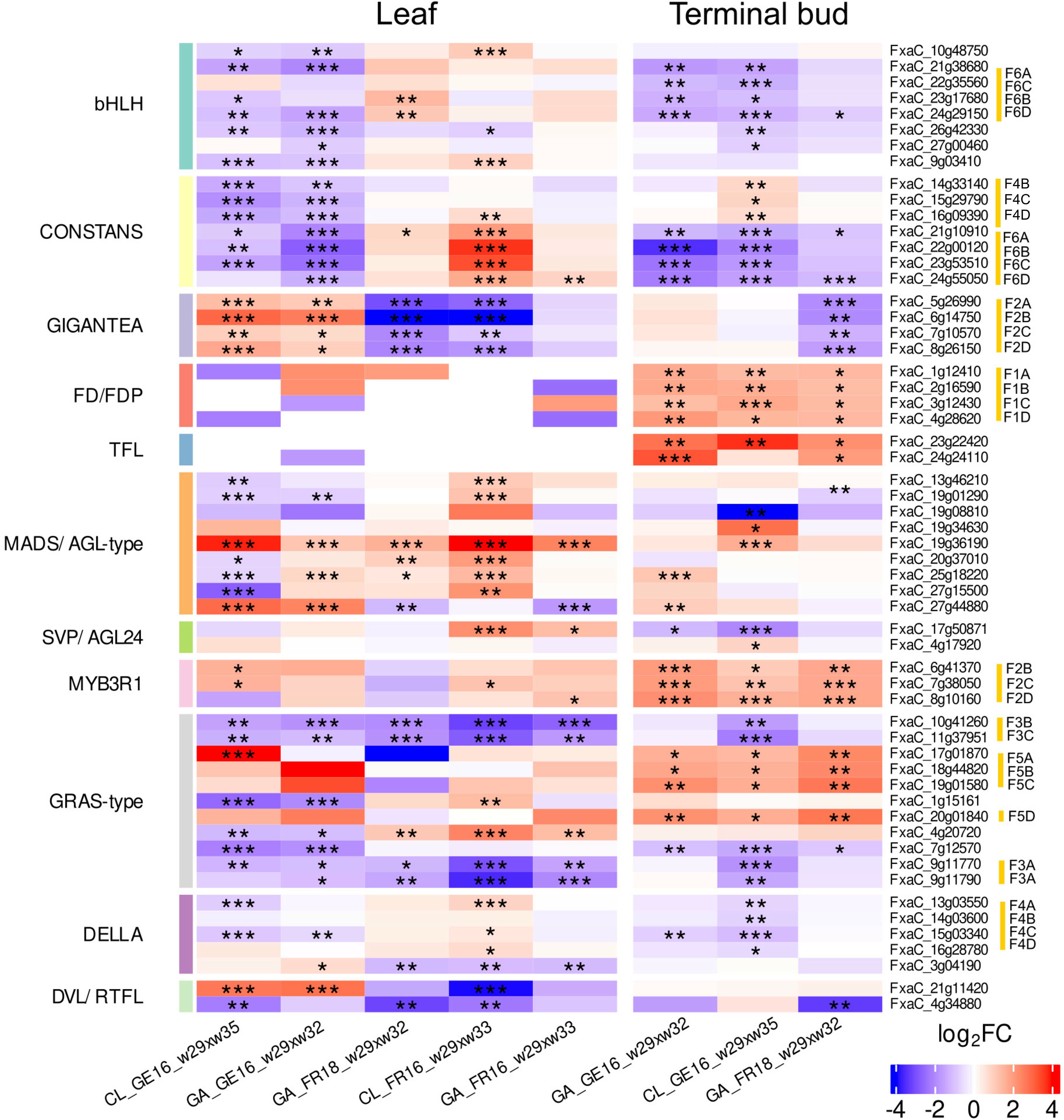
Figure 6. Heat map showing the expression of genes in leaf and terminal bud tissues of strawberry cultivars Clery (CL) and Gariguette (GA) sampled from France (FR) in 2016 and 2018, and from Germany (GE) in 2016, during floral initiation. Differentially expressed genes (named in rows) were chosen based on their protein function (MapMan protein classes) and significance level (***FDR < 0.001, **FDR < 0.01, *FDR < 0.05) for specific date contrasts (shown in columns) during w29 (T0), w32 (T10A), w33 (T10B) and w35 (T50). Gene expression is scaled to range between 5 and –5. Red corresponds to stronger upregulation and blue to downregulation. MapMan protein classification for genes was added on the left and color bars show different protein classes. Homologous genes and their corresponding haplotype in the F. × ananassa genome are highlighted in yellow on the right.
Seven genes encoding CONSTANS-like (COL) proteins were differentially expressed between T0 and T10 (Figure 7). The downregulation of COL genes was observed in all terminal bud samples at T10 compared to T0, with five showing strong, shared downregulation across both cultivars and the other two only significantly downregulated in cultivar GA (Figure 6). In contrast, the differential expression of these COL genes in leaves between T0 and T10 was highly dependent on the location. We observed significant downregulation in both cultivars in Germany (CL_GE16_T0xT10L and GA_GE16_T0xT10L) but an increase in France (GA_FR16_T0xT10L and CL&GA_FR18_T0xT10L). The CL_FR16_T0xT10L comparison revealed the significant upregulation of five COL genes in leaf tissue.
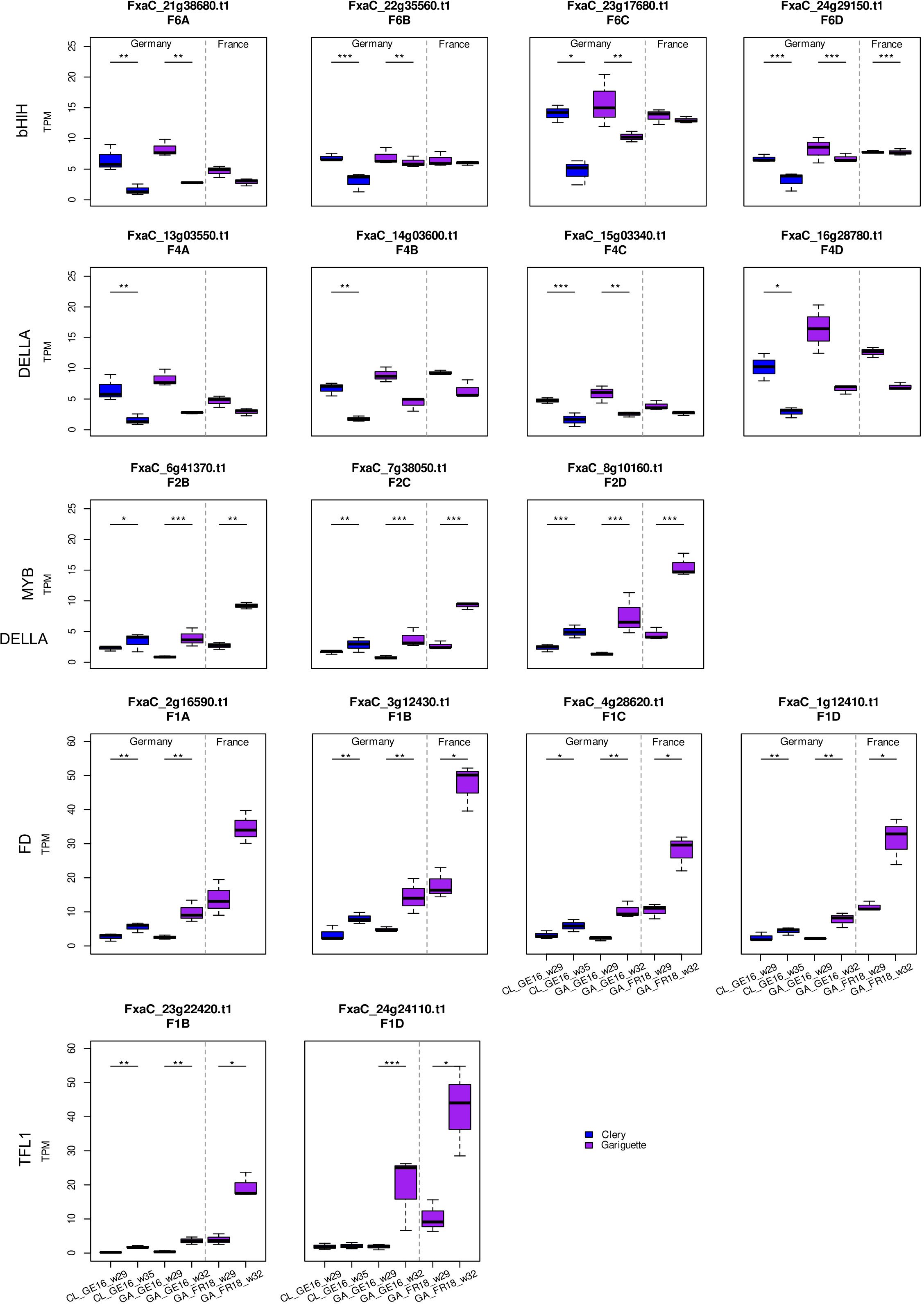
Figure 7. Transcript per million (TPM) values for bHLH, DELLA, MYB, FD and TFL1 candidate genes and their homologous sequences. Comparison of sampling dates during floral initiation in Clery (CL, blue) and Gariguette (GA, purple) terminal buds from plants growing in Germany (GE) and France (FR). Significance levels: ***FDR < 0.001, **FDR < 0.01 and *FDR < 0.05.
Eight genes encoding the bHLH-type transcriptional co-activator FBH were predominantly downregulated between T0 and T10 in the leaf tissue of both cultivars in Germany but tended to be upregulated in France. DEGs encoding the regulatory protein GIGANTEA (GI) were strongly downregulated between T0 and T10 in both tissues of plants grown in France, but were upregulated in leaf samples over the whole floral initiation period in Germany. FD-like and TFL1 genes were minimally expressed in leaves during early floral initiation but were strongly upregulated between T0 and T10 in the terminal buds.
Several MADS/AGL-type genes also showed differential expression. Specifically, the FxaC_19g36190 gene was significantly upregulated across almost all comparisons in both tissues, except the terminal buds of cultivar GA. Among the DEGs associated with SVP/AGL24, the gene FxaC_17g50871 was upregulated in the leaves of both cultivars in France but was downregulated in the terminal buds of plants grown in Germany.
DEGs encoding MYB3R1 transcription factors were upregulated in all comparisons of T0 and T10, especially in terminal buds. Transcription factor genes involved in the gibberellin pathway (e.g., encoding DELLA and GRAS-type proteins) were also modulated. Some were significantly downregulated in the leaves of CL plants grown in Germany, and also in the terminal buds of both cultivars, but most such genes in CL_FR16 leaves were upregulated between T0 and T10. Four DEGs encoding GRAS-type proteins were downregulated in all leaf samples, as was one gene in the terminal buds in the case of CL_GE16_ T0xT10, but other DEGs were upregulated in all terminal bud samples. The GA samples in general showed less significant differential expression than CL across locations and years. We identified two DEGs encoding DEVIL/ROT-FOUR-LIKE (DVL/RTFL) proteins with similar expression profiles to GI genes. One of these (FxaC_4g34880) was significantly downregulated only in the leaves and terminal buds of GA plants grown in France in 2018 (Figure 6).
Given that cultivated strawberry is octoploid (2n = 8x = 56), up to eight homoeoalleles for each gene located at orthologous positions in the four subgenomes (A, B, C and D) of F. × ananassa could potentially be expressed, although only some of the loci may be active (Hardigan et al., 2021). We discuss different situations for different genes below. The expression of homoeoalleles belonging to all four subgenomes was observed in terminal buds for the bHLH transcription factor gene on homoeologous group 6, all of which were significantly downregulated in both cultivars grown in Germany (Figure 7). Similarly, the DELLA transcription factor gene FanRGA was expressed in the four subgenomes of homoeologous group 4. They were all significantly downregulated in cultivar CL in Germany, whereas only the alleles of one subgenome (FxaC_15g03340) were significantly downregulated in cultivar GA (Figure 7). Finally, the flowering gene FanFD was expressed in all four subgenomes whereas FanTFL1 was only expressed in two subgenomes. These genes were significantly upregulated in most terminal bud samples but no significant differential expression was observed in leaves (Figures 6 and 7; FDR < 0.05).
Comparative analysis of selected floral initiation genes based on reference data in cultivated strawberry
We compared our DEGs to those reported in a previous study of floral initiation in the cultivated strawberry cv. Benihoppe (Liang et al., 2022). This revealed 19 common DEGs in leaf or terminal bud tissues, from which we selected six expressed in the terminal bud and associated with hormone pathways or carbohydrate metabolism for analysis at the subgenomic level (Figure 8).
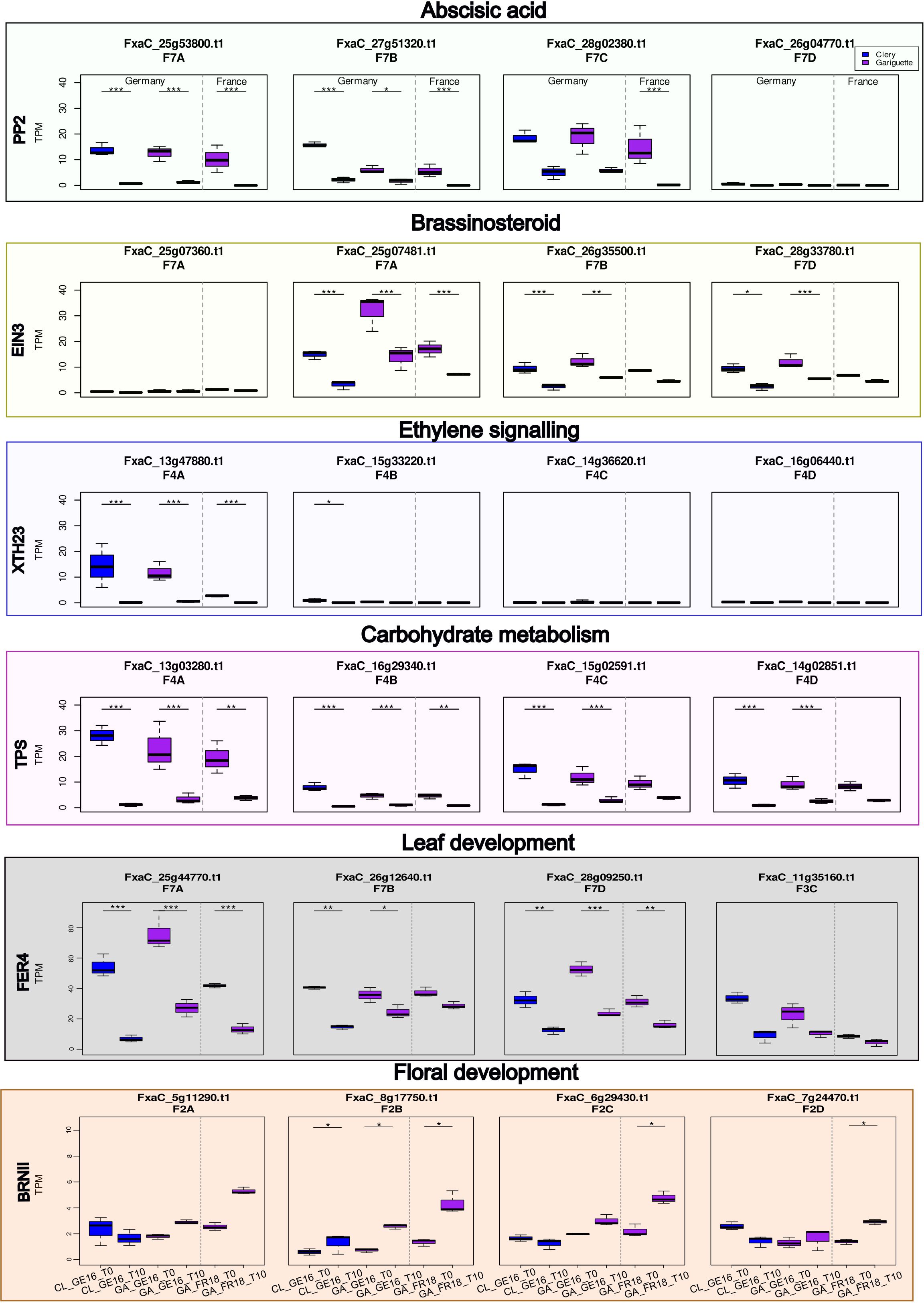
Figure 8. Whisker boxplots illustrating transcript per million (TPM) values for the candidate genes PP2, EIN3, XTH23, TPS, FER4 and BRNII involved in the abscisic acid, brassinosteroid and ethylene signaling pathways, and in carbohydrate metabolism. We analyzed the expression of homologous sequences between dates during floral initiation in terminal buds, comparing the Clery (CL, blue) and Gariguette (GA, purple) genotypes growing in Germany (GE) and France (FR). Significance levels: ***FDR < 0.001, **FDR < 0.01 and *FDR < 0.05.
One gene encoding a type 2C protein phosphatase (PP2) involved in the abscisic acid (ABA) pathway was significantly downregulated in three subgenomes across both locations and cultivars, except subgenome 7C (FxaC_28g02380) in Germany and subgenome 7D (FxaC_26g04770) in the terminal bud (Figure 8). Furthermore, the EIN3 gene encoding a brassinosteroid ethylene-like transcription factor was downregulated in three subgenomes but strongly expressed in subgenome 7A (FxaC_25g07481) in the terminal bud in both Germany and France (Figure 8; FDR < 0.001). The XTH23 gene involved in ethylene signaling was only significantly downregulated in subgenome 4A for both locations and both cultivars (FDR < 0.001). The expression of FxaC_15g33220 on subgenome 4B was significantly downregulated for cultivar CL in Germany (Figure 8; FDR < 0.05). In the context of carbohydrate metabolism, a TPS gene in homoeology group 4 was expressed at different levels depending on the subgenome, with significant downregulation in subgenomes 4C and 4D only in Germany, but significant downregulation in subgenomes 4A and 4B for both cultivars at both locations (Figure 8).
We analyzed two DEGs associated with in leaf and/or floral development. The FanFER4 gene, involved in leaf development, was significantly downregulated in both cultivars at both locations (FDR < 0.01) in subgenomes 7A and 7D, but the expression in subgenome 7B was only significant in Germany (FDR < 0.001). Three homeologous alleles of FER4 were identified on chromosome 7 (FDR < 0.05), and the fourth was located on chromosome 3 (Figure 8). The BRN1 gene encoding an RNA-binding protein involved in the regulation of flowering time was significantly upregulated in subgenomes 2C and 2D for cultivar GA in France and in subgenome 2B at both locations (Figure 8; FDR < 0.05).
Discussion
Floral initiation is a highly orchestrated developmental phase influenced by endogenous and exogenous signals (Korneef et al., 1998; Wellmer and Riechmann, 2010). In cultivated strawberry, a major crop species, the flowering time influences fruit yields, and recent studies have shown that both the genotype and the environment influence floral initiation (Krüger et al., 2022) and flowering time (Prohaska et al., 2024). Floral initiation occurs when temperatures and day length decrease, which reflects interactions between these two environmental factors (Heide et al., 2013). The process therefore varies from year to year at the same location. To accommodate this variation, we based our sampling dates on the percentage of initiated plants rather than a fixed calendar date. By tracking this percentage of initiated plants over time and sampling at 0%, 10% and 50% of initiated plants, we also highlighted the much more rapid floral initiation process in Germany compared to France, probably resulting from the steeper decrease in temperature and day length in Germany than France (Krüger et al., 2022).
The analysis of floral initiation in the model plant Arabidopsis (Arabidopsis thaliana) and various crop species has identified several regulatory pathways, including an autonomous pathway as well as those responding to photoperiod, temperature, vernalization, gibberellins, aging and carbohydrate levels (Mouradov et al., 2002; Boss et al., 2004). By studying two cultivars (Gariguette and Clery) cultivated in two different countries (France and Germany) across 3 years, we identified components of these pathways in the regulation of floral initiation in strawberry, as discussed in more detail below.
Environmental differences in floral initiation of strawberry
Our results demonstrate clear environmental modulation of floral initiation timing and gene expression in Fragaria × ananassa, shaped by both genotype and location. Floral initiation consistently occurred earlier in Germany (mid/late August) than in France (mid/late September), regardless of cultivar, suggesting that local climatic cues—likely temperature and photoperiod—accelerate meristem transition in northern latitudes (Krüger et al., 2022; Heide & Sønsteby, 2007).
Transcriptomic analyses revealed early divergence in gene expression patterns between sites. In Germany, genotype-specific differences in leaves and terminal buds were already evident at the vegetative stage (T0), marked by enriched expression of genes related to cellular respiration, protein modification, and cytoskeletal organization. In contrast, divergence in France emerged at stage T10 and involved genes regulating photosynthesis, phytohormone signaling, and RNA metabolism—indicating delayed environmental responsiveness (Liang et al., 2022).
Notably, CONSTANS-like (COL) genes and other flowering-time regulators such as FBH and GIGANTEA (GI) displayed environment-dependent expression. COL genes were downregulated in terminal buds across all samples at T10 but showed opposite trends in leaf tissues—downregulated in Germany and upregulated in France. Similar patterns were observed for GI and FBH, suggesting location-specific integration of photoperiod and circadian cues (Suárez-López et al., 2001; Imaizumi and Kay, 2006).
Enrichment of DEGs further supported environment-driven divergence. In Germany, stress-related and signaling-related pathways dominated, while in France, chromatin remodeling, cytoskeletal organization, and secondary metabolism were more prominent. These trends suggest different adaptive strategies between locations in response to seasonal transitions (Wellmer and Riechmann, 2010).
The octoploid genome structure of cultivated strawberry allowed for a nuanced view of subgenome-specific expression. Several floral regulators, including DELLA, TPS, and FER4 genes, exhibited differential expression across homoeoalleles, with stronger modulation in Germany. This supports the idea that specific subgenomes may contribute disproportionately to environmental sensitivity (Hardigan et al., 2021).
Overall, our findings emphasize that floral initiation in strawberry is a complex trait governed by both genetic background and environmental signals, with location-dependent expression of key flowering genes and subgenomic specialization. This has direct implications for breeding programs targeting climate resilience and photoperiod adaptability.
Photoperiodic pathway and floral integrators
The photoperiodic pathway of floral initiation involves a network of regulators that respond to day length. In Arabidopsis, the key genes involved in this process are well known. GI regulates circadian rhythms and promotes flowering under long-day conditions by stabilizing the CO protein (Michaels et al., 2003; Brandoli et al., 2020). CO plays a central role in sensing day length, accumulating during long days to directly activate the expression of the florigen gene FT (Abe et al., 2005; Wigge et al., 2005; Corbesier et al., 2007). In the SAM, FT competes with the floral repressor TFL1 to bind FD (Zhu et al., 2020). SOC1 is an integrator of the photoperiod, gibberellin and temperature pathways to promote flowering at the shoot apex (Li et al., 2008; Jung et al., 2012).
We found that the strawberry orthologs of these genes (FanCO, FanSOC1, FanFD, FanTFL1 and FanGI) were differentially expressed during early floral initiation. The involvement of FanCO, FanSOC1 and FanTFL1 in the seasonal control of floral initiation has already been demonstrated (Iwata et al., 2011; Koskela et al., 2012; Mouhu et al., 2013; Koskela et al., 2016; Kurokura et al., 2017; Munoz-Avila et al., 2022). In addition, we and others (Liang et al., 2022) showed that FanBRN1 is also differentially expressed during floral initiation in strawberry. The Arabidopsis ortholog AtBRN1 acts as a repressor of SOC1 activity (Kim et al., 2013).
Phytohormone signaling
We identified multiple DEGs associated with the gibberellic acid, brassinosteroid, ABA, and jasmonic acid pathways. The gibberellic acid pathway promotes the transition from vegetative growth to flowering (Yu et al., 2004; Mutasa-Göttgens and Hedden, 2009; Achard and Genschik, 2009). We found that FanRGA was differentially expressed in the terminal buds of both cultivars. In diploid strawberry, the ortholog FveRGA encodes a DELLA protein that suppresses stolon development (Li et al., 2019; Caruana et al., 2018). DELLA proteins are negative regulators of gibberellin signaling that act immediately downstream of the gibberellic acid receptor (Eckhardt, 2007).
Brassinosteroids promote flowering in Arabidopsis, (Liz and He, 2020). We found that FanXTH2, a component of the brassinosteroid pathway encoding the enzyme xyloglucan endotransglucosylase/hydrolase, was differentially expressed between cultivars and locations, as previously shown in strawberry terminal bud tissue (Liang et al., 2022) and loquat bud tissue (Xia et al., 2020).
We observed the downregulation of FanPP2 during floral initiation in both cultivars and locations, as also reported in a previous study (Liang et al., 2022). The Arabidopsis ortholog encodes a negative regulator of ABA signaling, suggesting that it influences the timing of floral initiation by repressing ABA-responsive genes. We also observed the downregulation of FanEIN3, encoding an ethylene-responsive transcription factor, consistently with previous findings (Liang et al., 2022). The Arabidopsis ortholog delays flowering by activating ERF1 and the APETALA2 (AP2)/ERF1 protein family (Guo and Ecker, 2004).
Transcription factors
Multiple transcription factor families, including bHLH, MADS-box and MYB, were differentially expressed during floral initiation, and are known to be involved in the transition from vegetative growth to reproductive development (Bemer and Angenent, 2009; Woodger et al., 2003; Liang et al., 2022). For example, we found two differentially expressed members of the DVL/RTFL family, which are known to be involved in organogenesis (Guo et al., 2015). One was upregulated in both cultivars and locations, but the other was downregulated in German samples and upregulated in French samples of the cultivar CL. Similarly, Arabidopsis DVL/RTFL proteins influence traits such as leaf shape and plant architecture. The overexpression of DEVIL1 (DVL1) and ROTUNDIFOLIA4 (ROT4/DVL16) in Arabidopsis produced a pleiotropic phenotype with short stature, rounder rosette leaves, and clustered inflorescences (Wen et al., 2004; Valdivia et al., 2012).
Plant peptides also play important roles in development, regulating terminal bud organization, root growth, and leaf shape (Matsubayashi, 2014; Matsubayashi and Sakagami, 2006; Guo et al., 2015). We observed the tissue-specific expression of GRAS-type proteins, which are putative transcriptional regulators, consistent with their documented roles. Most GRAS-type genes were dominantly expressed in roots with a subset also expressed in shoots and leaves (Heckmann et al., 2006; Liang et al., 2022). The findings emphasize the interplay between transcriptional regulation and tissue-specific signaling during floral initiation.
Carbohydrate and energy metabolism
DEGs related to carbohydrate (particularly starch) metabolism were identified in both cultivars, as well as certain secondary metabolic pathways, such as the terpenoid pathway (Figure 5). Trehalose-6-phosphate (T6P) is a sugar derivative that is proposed to influence flowering both in the terminal bud and the SAM. Accordingly, the TPS1 gene encoding trehalose-6-phosphate synthase contributes to the genetic framework that controls flowering time (Wahl et al., 2013; Rojas et al., 2023). We showed that FanTPS1 was downregulated in terminal buds in both cultivars and locations, as previously reported (Liang et al., 2022).
The carbohydrate pathway not only controls floral initiation but also contributes more widely to plant development (Zhang et al., 2013; Quiao et al., 2021). Carbohydrate metabolism provides the energy and building blocks needed for flower development so the interplay between carbohydrate metabolism, phytohormonal signaling and environmental cues ensures that the transition from vegetative growth to flowering is timed to maximize reproductive success (Corbesier et al., 1998; Sawicki et al., 2015). In the terminal bud of cultivar CL, we observed the overrepresentation of DEGs linked to starch metabolism, which serves as an energy reserve. Mobilized starch, converted into sucrose in leaves and stems, provides an early signal for floral induction (Bernier et al., 1993). Key differences between the cultivars included higher asparagine and rhamnogalacturonan I (RG-I) production in GA. Asparagine provides nitrogen for signaling proteins that coordinate floral induction with internal and external pathways (Khurana et al., 1988).
The FERONIA 4 (FER4) gene encodes a product that contributes to carbohydrate metabolism (particularly glycolysis) by interacting with the cytosolic enzyme glyceraldehyde-3-phosphate dehydrogenase (GAPDH). FER deficiency has been shown to reduce GAPDH activity, causing the of accumulation starch (Yang et al., 2015). We found that FER4 was downregulated in both cultivars, and that protein classes related to starch metabolism were enriched. FER4 was also downregulated in the cultivated strawberry cultivar Benihoppe (Liang et al., 2022), where it was linked to leaf development.
Cytoskeleton, cell division and cell wall dynamics
In the terminal buds of cultivar GA, we identified DEGs associated with cytoskeletal organization, specifically the microtubule network required for cell division and expansion, thereby influencing tissue patterning during floral initiation (Chandler, 2012; Denay et al., 2017). Interestingly, DEGs related to cell division were overrepresented in the terminal buds of cultivar CL, whereas DEGs related tRNA biosynthesis, bZIP transcription factors and RG-I biosynthesis, the latter needed for cell wall integrity (Yapo, 2011), were overrepresented in the terminal buds of cultivar GA. The role of RG-I in floral induction remains unclear. Disparities in the representation of protein classes between cultivars may reflect the field experiment setup and environmental fluctuations, masking inherent differences between the cultivars.
Regulation of subgenomic expression
In the octoploid cultivated strawberry, the plasticity of traits such as flowering (Prohaska et al., 2024) can confer polyploid advantage in heterogeneous environments (Wei et al., 2019). In this species, each gene may be represented by up to eight homoeoalleles located on the four homoeologous subgenomes (Rousseau-Gueutin et al., 2009). However, polyploidization is followed by a process of diploidization, whereby gene redundancy is reduced by processes such as gene silencing, sequence elimination and rearrangement (Chen, 2007; Doyle et al., 2008). In strawberry, the analysis of gene redundancy revealed that only 46% of genes retain alleles in all four subgenomes, whereas 7%, 23% and 14% retain alleles on one, two and three homoeologous chromosomes, respectively (Jin et al., 2023). For most of the F. × ananassa genes we identified, such as those encoding FBH, CO, GI, GRAS-type, DELLA, FD/FDP, PP2, XTH23, TPS and BRN1 proteins, we found sequences in all four subgenomes, whereas MYB3R1, EIN3 and FER4 sequences were present in three subgenomes and were similarly expressed. The expression of these genes is therefore likely to be finely regulated, with mutations in the promoter and/or 5′ untranslated region.
Conclusion
Our study provides new insights into the molecular basis of early floral initiation in F. × ananassa under natural environmental conditions. By comparing gene expression profiles between two cultivars, CL and GA, across two environments (Germany and France), we identified key genetic differences that underpin this developmental transition. Notably, we highlighted the involvement of genes such as XTH23, TPS and FER4, as well as transcription factors including FBH, CO and GI, in regulating floral initiation. This process appears to involve a coordinated reprogramming of carbohydrate metabolism, phytohormone signaling, and photoperiod-responsive pathways. These findings contribute to a better understanding of strawberry flowering and offer promising directions for optimazing cultivation practices and breeding strategies. However, further research is needed to elucidate the precise functions of these candidate genes and the significance of their differential expression in diverse environmental contexts.
Materials and methods
Floral initiation
The timing of floral initiation was determined by invasive architectural analysis of the Fragaria × ananassa cultivars Gariguette (GA) and Clery (CL), discriminating between the vegetative and floral stages of the SAM in the terminal bud. By dissecting the buds, the meristems were exposed. Plant architecture assessments were conducted over time throughout the summer and autumn at two locations: Bordeaux in France and Geisenheim in Germany. The first samples were collected in mid-July (T0, w29), coinciding with the presence of daughter plants featuring only small root primordia on stolons. Subsequent sampling took place in August (w32), 3 weeks after the transplantation of rooted plants to the field or nursery, and at 10-day intervals until early October (w40), making eight sampling dates in total.
Plant architecture was assessed as previously described (Krüger et al., 2022). Briefly, we described the daughter plants (including the number of developed leaves and the stage of the terminal bud) using a stereomicroscope with 40–60× magnification. A single terminal bud from the main crown of each plant was dissected for analysis (Figure 1A). We dissected 9–12 plants representing each cultivar and environment at each sampling date. The vegetative or floral status of the terminal buds was assessed as previously described (Krüger et al., 2022) as adapted from earlier methods (Jahn & Dana 1970; Taylor et al., 1997). As we didn’t know in advance on which dates we would observe about 10% of initiated plants, we stored leaf and terminal meristem samples in RNA later at each sampling date for their future use.
RNA-Seq sampling
RNA-Seq sampling time points were determined by the percentage of initiated SAM. Based on the assumption that samples at 0% (time point T0) and 5–20% (time point T10) of initiated plants would capture early floral initiation steps, samples of leaves and terminal buds were collected accordingly at three or four dates depending on the location and year. For each date, cultivar and organ, we collected three replicate samples of nine leaf discs or nine terminal buds from nine separate plants in one 1.5-ml Eppendorf tube (Figure 1B). RNA was extracted as previously described (Gaston et al., 2021) and 3 µg per sample was sent to Sistemas Genomicos (Spain) for sequencing on the Illumina HiSeq 2500 platform.
RNA-Seq data processing
Adaptors were removed from the Illumina RNA-Seq dataset using Trimmomatic v0.38 (Bolger et al., 2014) followed by alignment to the Fragaria_ananassa_v1.0.a2 cv. Camarosa reference genome (GDR database https://www.rosaceae.org; Liu et al., 2021) using the pseudo-aligner Salmon v1.10.1 for read count quantification (Patro et al., 2017). DEGs were identified using edgeR v3.40.1 (Robinson et al., 2010). Contrasts were calculated between the different sampling points (T0, T10A, T10B and T50) and cultivars separately for both tissues. The transcripts per million (TPM) were evaluated by PCA, and ORA was applied to the PC1 and PC2 axes. This provided insights into the leading MapMan (Schwacke et al., 2019) protein classes contributing to the separation along PC1 and PC2 in the positive and negative directions. The outcomes were visually represented as bar charts illustrating the enrichment factor along these axes. The top five genes in both directions along PC1 and PC2 were also depicted on the PCA plot. Venn diagrams were generated to identify shared DEGs in different comparisons using VennDiagram v1.7.3 (Chen and Boutros, 2011) in R. A threshold of FDR < 0.05 was used to filter significant DEGs. Heat map clustering was applied to discern gene relationships within each tissue.
Gene correspondence and functional analysis
To link our work to the published literature (Li et al., 2019; Liang et al., 2022), we matched gene names between the current Camarosa F. × ananassa annotation (v2) and the diploid genome of F. vesca or the Camarosa v1 annotation used by Liang et al. (2022). The genes were checked bidirectionally against the diploid transcriptome of F. vesca and F. × ananassa cv. Camarosa using Blast+ v2.15.0 (Camacho et al., 2009). We focused on the DEGs in our study and applied ORA to the overlapping genes to identify MapMan protein classes that potentially influence floral initiation. Statistical analysis was carried out using R Studio base version 2023.06.0 (R Core Team, 2023).
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ebi.ac.uk, PRJEB83679.
Author contributions
FZ: Writing – original draft, Writing – review & editing, Conceptualization, Data curation, Methodology, Visualization, Formal Analysis, Investigation. AG: Formal Analysis, Investigation, Methodology, Writing – review & editing, Writing – original draft. KG: Methodology, Project administration, Writing – review & editing. MD: Methodology, Writing – review & editing. EK: Conceptualization, Funding acquisition, Methodology, Writing – review & editing. BB: Methodology, Writing – review & editing. KE: Methodology, Writing – review & editing. SO: Funding acquisition, Methodology, Project administration, Writing – review & editing. BD: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing. BU: Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by the European Union’s Horizon 2020 Research and Innovation Program (Goodberry project, grant agreement number 679303).
Acknowledgments
We thank Dr. Richard M Twyman for manuscript proofreading.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1563658/full#supplementary-material
References
Abe, M., Kobayashi, Y., Yamamoto, S., Daimon, Y., Yamaguchi, A., Ikeda, Y., et al. (2005). FD, a bZIP protein mediating signals from the floral pathway integrator FT at the shoot apex. Science (New York, N.Y.), 309(5737), 1052–1056. doi: 10.1126/science.1115983
Achard, P. and Genschik, P. (2009). Releasing the brakes of plant growth: how GAs shutdown DELLA proteins. J. Exp. Bot. 60, 1085–1092. doi: 10.1093/jxb/ern301
Bemer, M. and Angenent, G. C. (2009). Floral organ initiation and development. Plant Dev. Biology-Biotechnol. Perspect., 173–194. doi: 10.1007/978-3-642-02301-9_9
Benlloch, R., Berbel, A., Serrano-Mislata, A., and Madueño, F. (2007). Floral initiation and inflorescence architecture: a comparative view. Ann. Bot. 100, 659–676. doi: 10.1093/aob/mcm146
Bernier, G., Havelange, A., Houssa, C., Petitjean, A., and Lejeune, P. (1993). Physiological signals that induce flowering. Plant Cell 5, 1147–1155. doi: 10.2307/3869768
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Boss, P. K., Bastow, R. M., Mylne, J. S., and Dean, C. (2004). Multiple pathways in the decision to flower: enabling, promoting, and resetting. Plant Cell 16, 16. doi: 10.1105/tpc.015958
Brandoli, C., Petri, C., Egea-Cortines, M., and Weiss, J. (2020). Gigantea: Uncovering new functions in flower development. Genes 11, 1142. doi: 10.3390/genes11101142
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinf. 10, 421. doi: 10.1186/1471-2105-10-421
Caruana, J. C., Sittman, J. W., Wang, W., and Liu, Z. (2018). Suppressor of Runnerless Encodes a DELLA protein that controls runner formation for asexual reproduction in strawberry. Mol. Plant 11, 230–233. doi: 10.1016/j.molp.2017.11.001
Chandler, J. W. (2012). Floral meristem initiation and emergence in plants. Cell. Mol. Life Sci. 69, 3807–3818. doi: 10.1007/s00018-012-0999-0
Chen, Z. J. (2007). Genetic and epigenetic mechanisms for gene expression and phenotypic variation in plant polyploids. Annu. Rev. Plant Biol. 58, 377–406. doi: 10.1146/annurev.arplant.58.032806.103835
Chen, H. and Boutros, P. C. (2011). VennDiagram: a package for generation of highly customizable Venn and Euler diagrams in R. BMC Bioinf. 12. doi: 10.1186/1471-2105-12-35
Corbesier, L., Vincent, C., Jang, S., Fornara, F., Fan, Q., Searle, I., et al. (2007). FT protein movement contributes to long-distance signaling in floral induction of Arabidopsis. Science (New York, N.Y.), 316(5827), 1030–1033. doi: 10.1126/science.1141752
Corbesier, L., Lejeune, P., and Bernier, G. (1998). The role of carbohydrates in the induction of flowering in Arabidopsis thaliana: comparison between the wild type and a starchless mutant. Planta 206, 131–137. doi: 10.1007/s004250050383
Costes, E., Crespel, L., Denoyes, B., Morel, P., Demene, M. N., Lauri, P. E., et al. (2014). Bud structure, position and fate generate various branching patterns along shoots of closely related Rosaceae species: a review. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00666
Denay, G., Chahtane, H., Tichtinsky, G., and Parcy, F. (2017). A flower is born: an update on Arabidopsis floral meristem formation. Curr. Opin. Plant Biol. 35, 15–22. doi: 10.1016/j.pbi.2016.09.003
Doyle, J. J., Flagel, L. E., Paterson, A. H., Rapp, R. A., Soltis, D. E., Soltis, P. S., et al. (2008). Evolutionary genetics of genome merger and doubling in plants. Annu. Rev. Genet. 42, 443–461. doi: 10.1146/annurev.genet.42.110807.091524
Eckhardt, N. A. (2007). GA signaling: direct targets of DELLA proteins. Plant Cell 19, 2970. doi: 10.1105/tpc.107.191010
Gaston, A., Potier, A., Alonso, Sabbadini, S., Delmas, F., Tenreira, T., et al. (2021). The FveFT2 florigen/FveTFL1 antiflorigen balance is critical for the control of seasonal flowering in strawberry while FveFT3 modulates axillary meristem fate and yield. New Phytol. 232, 372–387. doi: 10.1111/nph.v232.1
Guo, H. W. and Ecker, J. R. (2004). The ethylene signaling pathway: New insights. Curr. Opin. Plant Biol. 7, 40–49. doi: 10.1016/j.pbi.2003.11.011
Guo, P., Yoshimura, A., Ishikawa, N., Yamaguchi, T., Guo, Y., and Tsukaya, H.. Comparative analysis of the RTFL peptide family on the control of plant organogenesis (2015). J. Plant Res. 128, 497–510. doi: 10.1007/s10265-015-0703-1
Hardigan, M. A., Lorant, A., Pincot, D. D.A., Feldmann, M. J., Famula, R. A., Acharya, C. B., et al (2021). Unraveling the Complex Hybrid Ancestry and Domestication History of Cultivated Strawberry. Molecular biology and evolution, 38(6), 2285–2305. doi: 10.1093/molbev/msab024
Heckmann, A. B., Lombardo, F., Miwa, H., Perry, J. A., Bunnewell, S., Parniske, M., et al. (2006). Lotus Japanicus nodulation requires two GRAS domain regulators, one which is functionally conserved in a non-legume. Plant Physiol. 142, 1739–1750. doi: 10.1104/pp.106.089508
Heide, O.M. and Sønsteby, A. (2007) Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca). Physiol. Plant. 130, 280–289.
Heide, O. M., Stavang, J. A., and Sønsteby, A. (2013). Physiology and genetics of flowering in cultivated and wild strawberries-a review. J. Hortic. Sci. Biotechnol. 88, 1–18. doi: 10.1080/14620316.2013.11512930
Hytonen, T. and Kurokura (2020). Control of flowering and runnering in Strawberry. Horticult. J. 89, 96–107. doi: 10.2503/hortj.UTD-R011
Imaizumi, T. and Kay, S. A. (2006). Photoperiodic control of flowering: not only by coincidence. Trends Plant Sci. 11, 550–558. doi: 10.1016/j.tplants.2006.09.004
Iwata, H., Gaston, A., Remay, A., Thouroude, T., Jeauffre, J., Kawamura, K., et al. (2011). The TFL1 homologue KSN is a regulator of continuous flowering in rose and strawberry. Plant J. 69, 116–125. doi: 10.1111/j.1365-313X.2011.04776.x
Jahn, OL and Dana, MN. 1970. Crown and inflorescence development in the strawberry Fragaria ananassa. American Journal of Botany 57, 605–612.
Jin, X., Du, H., Zhu, C., Wan, H., Liu, F., Ruan, J., et al. (2023). Haplotype-resolved genomes of wild octoploid progenitors illuminate genomic diversifications from wild relatives to cultivated strawberry. Nat. Plants 9, 1252–1266. doi: 10.1038/s41477-023-01473-2
Jung, J. H., Ju, Y., Seo, P. J., Lee, J. H., and Park, C. M. (2012). The SOC1-SPL module integrates photoperiod and gibberellic acid signals to control flowering time in Arabidopsis. The Plant journal : for cell and molecular biology, 69(4), 577–588. doi: 10.1111/j.1365-313X.2011.04813.x
Khurana, J. P., Tamot, B. K., and Maheshwari, S. C. (1988). Floral induction in a photoperiodically insensitive duckweed, Lemna paucicostata LP6: Role of glutamate, aspartate, and other amino acids and amides. Plant Physiol. 86, 904–907. doi: 10.1104/pp.86.3.904
Kim, H. S., Abbasi, N., and Choi, S. B. (2013). Bruno-like proteins modulate flowering time via 3′ UTR-dependent decay of SOC1 mRNA. New Phytol. 198, 747–756. doi: 10.1111/nph.2013.198.issue-3
Koembuoy, K., Hasegawa, S., Otagaki, S., Takahashi, H., Nagano, S., Isobe, S., et al. (2020). RNA-Seq analysis pf meristem cells indentifies the FaFT3 gene as a common floral inducer in Japaneses cultivated strawberry. Horticult. J. 89, 138–146. doi: 10.2503/hortj.UTD-126
Korneef, M., Alonso-Blanco Cm Peeters, J. M., and Soppe, W. (1998). Genetic control of flowering time in Arabidopsis. Annu. Rev. Plant Biol. 49, 345–370. doi: 10.1146/annurev.arplant.49.1.345
Koskela, E. A., Mouha, K., Albani, M. C., Kuokura, T., Rantanen, M., Sargent, D. J., et al. (2012). Mutation in TERMINAL FLOWER 1 reverses the photoperiodic requirement for flowering in the wild strawberry Fragaria vesca. Plant Physiol. 159, 1043–1054. doi: 10.1104/pp.112.196659
Koskela, E. A., Sonsteby, A., Flachowsky, H., Heide, Om., Hanke, M. V., Elomaa, P., et al. (2016). TERMINAL FLOWER 1 is a breeding target for a novel everbearing trait and tailored flowering responses in cultivated strawberry (Fragaria x ananassa Duch.). Plant Biotechnol. J. 14, 1852–1861. doi: 10.1111/pbi.2016.14.issue-9
Krüger, E., Woznicki, T. L., Heide, O. M., Kusnierek, K., Rivero, R., Masny, A., et al. (2022). Flowering phenology of six seasonal-flowering strawberry cultivars in a coordinated European study. Horticulturae 8, 933. doi: 10.3390/horticulturae8100933
Kurokura, T., Samad, S., Koskela, E., Mouhu, K., and Hytönen, T. (2017). Fragaria vesca CONSTANS controls photoperiodic flowering and vegetative development. J. Exp. Bot. 68, 4839–4850. doi: 10.1093/jxb/erx301
Liang, J., Zheng, J., Wu, Z., and Wang, H. (2022). Time-course transcriptomic profiling of floral induction in cultivated strawberry. Int. J. Mol. Sci. 23, 6126. doi: 10.3390/ijms23116126
Li, D., Liu, C., Shen, L., Wu, Y., Chen, H., Robertson, , et al. (2008). A repressor complex governs the integration of flowering signals in Arabidopsis. Developmental cell, 15(1), 110–120. doi: 10.1016/j.devcel.2008.05.002
Liu, T., Li, M., Liu, Z., Ai, X., and Li, Y. (2021). Reannotation of the cultivated strawberry genome and establishment of a strawberry genome database. Horticult. Res. 8, 41. doi: 10.1038/s41438-021-00476-4
Li, Y., Feng, J., Cheng, L., Dai, C., Gao, Q., Liu, Z, et al. (2019). Gene Expression Profiling of the Shoot Meristematic Tissues in Woodland Strawberry Fragaria vesca. Frontiers in plant science, 10, 1624. doi: 10.3389/fpls.2019.01624
Li, Z. and He, Y. (2020). Roles of Brassinosteroids in Plant Reproduction. International journal of molecular sciences, 21(3), 872. doi: 10.3390/ijms21030872
Matsubayashi, Y. (2014). Posttranslationally modified small-peptides signals in plants. Annu. Rev. Plant Biol. 65, 385–413. doi: 10.1146/annurev-arplant-050312-120122
Matsubayashi, Y. and Sakagami, Y. (2006). Peptide hormones in plants. Annu. Rev. Plant Biol. 57, 649–674. doi: 10.1146/annurev.arplant.56.032604.144204
Michaels, S. D., Ditta, G., Gustafson, Brown, C., Pelaz, S., Yanofsky, M., et al. (2003). AGL24 acts as promoter of flowering in Arabidopsis and is positively regulated by vernalization. Plant J. 33, 867–874. doi: 10.1046/j.1365-313X.2003.01671.x
Mouhu, K., Kurokura, T., Koskela, E. A., Albert, V. A., Elomaa, P., Hytönen, T., et al (2013). The Fragaria vesca homolog of suppressor of overexpression of constans1 represses flowering and promotes vegetative growth. The Plant cell, 25(9), 3296–3310.doi: 10.1105/tpc.113.115055
Mouradov, A., Cremer, F., and Coupland, G. (2002). Control of flowering time: Interacting pathways as a basis for diversity. Plant Cell 14, 111–130. doi: 10.1105/tpc.001362
Muñoz-Avila, J. C., Prieto, C., Sánchez-Sevilla, J. F., and Amaya Castillejo, I. C. (2022).Role of FaSOC1 and FaCO in the seasonal control of reproductive and vegetative development in the perennial crop Fragaria × ananassa. Frontiers in plant science, 13, 971846. doi: 10.3389/fpls.2022.971846
Mutasa-Göttgens, E. and Hedden, P. (2009). Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 60, 1979–1989. doi: 10.1093/jxb/erp040
Patro, R., Guggal, G., Love, M. I., Irizarry, R. A., and Kingsford, C. (2017). Salmon: fast and bias-aware quantification of transcript expression using dual-phase inference. Nat. Methods 14, 417–419. doi: 10.1038/nmeth.4197
Prohaska, A., Petit, A., Lesemann, S., Rey-Serra, P., Mazzoni, L., Masny, A., et al. (2024). Strawberry phenotypic plasticity in flowering time is driven by the interaction between genetic loci and temperature. J. Exp. Bot. 75, 5923–5939. doi: 10.1093/jxb/erae279
Quiao, Z., Hu, H., Shi, S., Yuan, X., Yan, B., and Chen, L. (2021). An update on the function, biosynthesis and regulation of floral volatile terpenoids. Horticulturae 7, 451. doi: 10.3390/horticulturae7110451
Rantanen, M., Kurokura, T., Mouhu, K., Pinho, P., Tetri, E., Halonen, L., et al. (2014). Light quality regulates flowering in FvFT1/FvTFL1 dependent manner in the woodland strawberry Fragaria vesca. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00271
R Core Team. (2023). R: A language and environment for statistical computing. In: R Foundation for Statistical Computing. Vienna, Austria. Available online at: https://www.R-project.org/.
Robinson, M. D., McCarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rojas, B. E., Tonetti, T., and Figueroa, C. M. (2023). Trehalose 6-phosphate metabolism in C4 species. Curr. Opin. Plant Biol. 72:102347. doi: 10.1016/j.pbi.2023.102347
Rousseau-Gueutin, M., Gaston, A., Aïnouche, A., Aïnouche, M. L., Olbricht, K., Staudt, G., et al. (2009). Tracking the evolutionary history of polyploidy in Fragaria L. (strawberry): new insights from phylogenetic analyses of low-copy nuclear genes. Mol. Phylogenet. Evol. 51, 515–530. doi: 10.1016/j.ympev.2008.12.024
Sawicki, M., Jacquens, L., Baillieul, F., Clément, C., Vaillant-Gaveau, N., and Jacquard, C. (2015). Distinct regulation in inflorescence carbohydrate metabolism according to grapevine cultivars during floral development. Physiol. Plant. 154, 447–467. doi: 10.1111/ppl.2015.154.issue-3
Schwacke, R., Ponce-Soto, G. Y., Krause, K., Bolger, A. M., Arsova, B., Hallab, A., et al. (2019). MapMan4: A refined protein classification and annotation framework applicable to multi-omics data analysis. Mol. Plant 12, 879–892. doi: 10.1016/j.molp.2019.01.003
Stewart, P. J. and Folta, K. M. (2010). A review of photoperiodic flowering research in strawberry (Fragaria spp.). Crit. Rev. Plant Sci. 29, 1–13. doi: 10.1080/07352680903436259
Suárez-López, P., Wheatly, K., Robson, F., Onouchi, H., Valverde, F., and Goupland (2001). CONSTANS mediates between the circadian clock and the control of flowering in Arababidopsi. Mature 410, 1116–1120. doi: 10.1038/35074138
Tenreira, T., Lange, M. J. P., Lange, T., Bres, C., Labadie, M., Monfort, A., et al. (2017). A specific gibberellin 20-oxidase dictates the flowering-runnering decision in diploid strawberry. Plant Cell 29, 2168–2182. doi: 10.1105/tpc.16.00949
Valdivia, E. R., Chevalier, D., Sampedro, J., Taylor, I., Niederhuth, C. E., and Walker, J. C. (2012). DVL genes play a role in the coordination of socket cell recruitment and differentiation. J. Exp. Bot. 63, 1405–1412. doi: 10.1093/jxb/err378
Verheul, M. J., Sønsteby, A., and Grimstad, S. O. (2006). Interactions of photoperiod, temperature, duration of short-day treatment and plant age on day treatment and plant age on flowering of Fragaria x ananassa Duch. cv. Korona. Sci. Hortic. 107, 164–170. doi: 10.1016/j.scienta.2005.07.004
Wahl, V., Punnu, J., Schlereth, A., Arrivault, S., Langenecker, T., Franke, A., et al. (2013). Regulation of flowering by trehalose-6-phosphate signaling in arabidopsis thaliana. Science 339, 704–707. doi: 10.1126/science.1230406
Weberling, F. (1989). Morphology of flowers and inflorescences. 1st Edition (Cambridge, UK: Cambridge University Press).
Wei, N., Cronn, R., Liston, A., and Ashman, T. L. (2019). Functional trait divergence and trait plasticity confer polyploid advantage in heterogeneous environments. New Phytol. 221, 2286–2297. doi: 10.1111/nph.2019.221.issue-4
Wellmer, F. and Riechmann, J. L. (2010). Gene networks controlling the initiation of flower development. Rev. Trends Genet. 26, 519–527. doi: 10.1016/j.tig.2010.09.001
Wen, J., Lease, K. A., and Walker, J. C. (2004). DVL, a novel class of small polypeptides: overexpression alters Arabidopsis development. Plant J. 37, 668–677. doi: 10.1111/j.1365-313X.2003.01994.x
Wickland, D. P. and Hanzawa, Y. (2015). The FLOWERING LOCUS TERMINAL FLOWER 1 Gene Family: Functional evolution and molecular mechanisms. Mol. Plant 8, 983–997. doi: 10.1016/j.molp.2015.01.007
Wigge, P. A., Kim, M. C., Jaeger, K. E., Busch, W., Schmid, M., Lohmann , J. U., et al. (2005). Integration of spatial and temporal information during floral induction in Arabidopsis. Science (New York, N.Y.), 309(5737), 1056–1059. doi: 10.1126/science.1114358
Woodger, F. J., Millar, A., Murray, F., Jacobson, J. V., and Gubler, F. (2003). The role of GAMYB transcription factors in GA-regulated gene expression. J. Plant Growth Regul. 22, 176–184. doi: 10.1007/s00344-003-0025-8
Wyatt, R. (1982). Inflorescence architecture: how flower number, arrangement, and phenology affect pollination and fruit-set. Am. J. Bot. 69, 585–594. doi: 10.1002/j.1537-2197.1982.tb13295.x
Xia, Y., Xue, B., Shi, M., Zhan, F., Wu, D., Jing, D., et al. (2020). Comparative transcriptome analysis of flower bud transition and functional characterization of EjAGL17 involved in regulating floral initiation in loquat. PloS One 15, e0239382. doi: 10.1371/journal.pone.0239382
Yang, T., Wang, L., Li, C., Liu, Y., Zhu, S., Qi, Y., et al. (2015). Receptor protein kinase FERONIA controls leaf starch accumulation by interacting with glyceraldehyde-3-phosphate dehydrogenase. Biochem. Biophys. Res. Commun. 465, 77–82. doi: 10.1016/j.bbrc.2015.07.132
Yapo, B. M. (2011). Rhamnogalacturonan-I: A structurally puzzling and functionally versatile polysaccharide from plant cell walls and mucilages. Polymer. Rev. 51, 391–413. doi: 10.1080/15583724.2011.615962
Yu, H., Ito, Y., Zhao, Y., Peng, J., Kumar, P., and Meyerowitz, E. (2004). Floral homeotic genes are targets of gibberellin signaling in flower development. Biol. Sci. 101, 7827–7832. doi: 10.1073/pnas.0402377101
Keywords: strawberry, organogenesis, floral initiation, differentially expressed genes, Fragaria × ananasa
Citation: Ziegler FMR, Gaston A, Guy K, Devers M, Krüger E, Brauksiepe B, Eimert K, Osorio S, Denoyes B and Usadel B (2025) Molecular mechanisms underlying the early steps of floral initiation in seasonal flowering genotypes of cultivated strawberry. Front. Plant Sci. 16:1563658. doi: 10.3389/fpls.2025.1563658
Received: 21 January 2025; Accepted: 28 May 2025;
Published: 19 June 2025.
Edited by:
Sagheer Ahmad, Guangdong Academy of Agricultural Sciences, ChinaReviewed by:
Tabibul Islam, The University of Tennessee, Knoxville, United StatesEmil Khavkin, All-Russia Research Institute of Agricultural Biotechnology, Russia
Anju Bajpai, CISH, India
Copyright © 2025 Ziegler, Gaston, Guy, Devers, Krüger, Brauksiepe, Eimert, Osorio, Denoyes and Usadel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Freya Maria Rosemarie Ziegler, Zi56aWVnbGVyQGZ6LWp1ZWxpY2guZGU=; Béatrice Denoyes, YmVhdHJpY2UuZGVub3llc0BpbnJhZS5mcg==; Björn Usadel, Yi51c2FkZWxAZnotanVlbGljaC5kZQ==