 Ling-Lu Zeng†
Ling-Lu Zeng† Wan-Qiu Meng
Wan-Qiu Meng Li Sun
Li Sun- College of Life Sciences, Shihezi University, Shihezi, Xinjiang, China
Eremopyrum triticeum, an annual spring ephemeral plant and a wild relative of wheat (Triticum aestivum), is widely distributed in the Junggar Desert of northern Xinjiang, China. It has several adaptive traits to survive in the desert environments, such as rapid growth in the early spring, flowers quickly, and completes its life cycle within approximately two months. However, the adaptation mechanisms of the fast flowering are still unknown. In this study, high-throughput RNA sequencing (RNA-seq) was performed to identify differentially expressed genes (DEGs) associated with flowering in E. triticeum during three developmental stages. A total of 11,278 DEGs were identified, including 1,632 DEGs specifically expressed during the flowering stage. Pathway analysis showed that these DEGs are mainly enriched in plant-pathogen interaction, plant hormone signal transduction, the MAPK signaling pathway, and so on. A total of 92 DEGs related to the flowering pathway were identified, which are mainly involved in the photoperiod, hormone signaling, autonomous, and vernalization pathway. Multiple transcription factor families related to floral transition were identified, with members of the MADS-box, bHLH, MYB, and AP2 families being the most abundant. In addition, four FLOWERING LOCUS T (FT) genes were identified in E. triticeum, and three of them were highly up-regulated at the flowering stage. The expression of EtFT-1 was induced in darkness, and short-day conditions promote its expression. Overexpression of the EtFT-1 gene accelerates flowering in Arabidopsis.
Introduction
Spring ephemeral plants are a significant component of desert flora. They are able to use winter snowmelt and early spring rainfall to germinate, grow and flower rapidly, and complete their life cycle in two to three months (Wang, 1993; Zhang and Chen, 2002). Spring ephemeral plants are an important part of the flora of the Gurbantunggut desert in Northern Xinjiang, China (Mao and Zhang, 1994). In the month of May, their cover here can be as high as 40%, making them significant for dune stabilization and environmental sustainability during the spring season (Wang et al., 2003). These species have developed specialized adaptive mechanisms that enable them to be effective in the harsh desert environment, including rapid growth and flowering, high photosynthesis ability under high light intensity conditions, high seed production potential, and resistance against abiotic stress factors. Understanding the adaptation mechanisms of ephemeral plants helps to elucidate the survival strategies of plants in extreme desert environments and provides important clues for ecological adaptation and evolutionary studies. Furthermore, these plants play an important role in desert ecosystems by stabilizing sand dunes and maintaining the ecological balance in spring arid environments, which is important for combating desertification and protecting biodiversity.
Eremopyrum triticeum (Gaertn.) Nevski (2n = 2x = 14) is an annual spring ephemeral plant and one of the wild relatives of wheat (Triticum aestivum) (Frederiksen and Bothmer, 1995). In China, it is mainly distributed in the southern part of Junggar Desert and the Ili Valley (Zhang, 2014), where it serves as an important component of desert and steppe grassland ecosystems. This species plays an important ecological role in stabilizing soil, maintaining biodiversity, and providing an essential forage resource in early spring (Wang et al., 2003). To cope with the harsh desert environment, E. triticeum has evolved a number of specialized adaptive traits, including rapid germination, growth, and flowering, which allow it to efficiently utilize winter snowmelt and early spring rainfall. It completes its life cycle in about two months, from late March to early April, before the onset of extreme summer drought (Zhang, 2014; Qiu et al., 2007). In addition, E. triticeum exhibits heterochronic germination, with seeds capable of germinating in both spring and fall, allowing it to take advantage of favorable conditions across seasons and increasing its adaptability to desert environments (Wang et al., 2024). The species exhibits high photosynthetic efficiency under intense sunlight, seed yield, low temperature tolerance, drought and salinity resistance, allowing it to survive and reproduce in extreme desert environments (Zhang, 2014). Despite these ecological and adaptive advantages, the molecular mechanisms underlying its rapid flowering and environmental resistance remain largely unknown.
The flowering transition is a critical process in the life cycle of a plant, as it marks the transition from vegetative growth to reproductive growth. The timing of flowering is regulated by both internal genetic elements and external environmental signals, including photoperiod, temperature change, and stress (Kurokura et al., 2013). In Arabidopsis, six pathways have been identified that regulate flowering: the photoperiod pathway, the gibberellin (GA) pathway, the vernalization pathway, the environmental temperature pathway, the aging pathway, and the autonomous pathway (Fornara et al., 2010; Srikanth and Schmid, 2011; Kinoshita and Richter, 2020). These different pathways are coordinated into a regulatory network controlled by a group of key regulators, including FLOWERING LOCUS T (FT), GIGANTEA (GI), SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), FLOWERING LOCUS C (FLC), TWIN SISTER OF FT (TSF), and LEAFY (LFY) (Parcy, 2005; Moon et al., 2005). Among these regulators, FT is the key regulator controlling the floral transition (Suárez-López et al., 2001). FT is a member of the phosphatidylethanolamine binding protein (PEBP) superfamily that functions as a mobile floral signal. It plays a critical role in the regulation of flowering time. The PBP family in Arabidopsis contains six members that are divided into three subfamilies: TFL1-like, FT-like, and MFT-like. The MFT-like subfamily is the ancestor of the FT-like and TFL1-like subfamilies (Liu et al., 2016). FT expression is controlled by CONSTANS (CO), a member of the CONSTANS-LIKE (COL) family of B-box zinc finger transcription factors. SOC1, a MADS box transcription factor, is a floral activator that regulates flowering time, floral patterning, and meristem identity (Lee and Lee, 2010). Conversely, FLC, another MADS box transcription factor, is a key repressor of flowering through repression of FT and SOC1 expression (Michaels and Amasino, 1999; Helliwell et al., 2006). Activation of floral promoters subsequently induces the expression of floral meristem identity genes such as LFY and APETALA 1 (AP1), which ultimately initiates the flowering process (Blázquez et al., 1997).
In the Arabidopsis photoperiod pathway, FT and CO are essential promoters of flowering, whereas in cereals such as wheat, the vernalization pathway is regulated by genes such as VRN1, VRN2, VRN3, and VRN4 (Yan et al., 2003, 2004, 2006; Kippes et al., 2015). In the autonomous pathway, factors such as FLOWERING LOCUS D (FLD), LUMINIDEPENDENS (LD), FLOWERING LOCUS PA (FPA), FLOWERING LOCUS CA (FCA), FLOWERING LOCUS VE (FVE), and FLOWERING LOCUS Y (FY) facilitate flowering by repressing FLC expression (Michaels and Amasino, 2001).
The age pathway is primarily regulated by microRNA156 (miR156), targets the SQUAMOSA-PROMOTER BINDING LIKE (SPL) transcription factors, and are essential for the regulation of flowering in A. thaliana (Zheng et al., 2019). In addition to GA, other endogenous hormones, such as auxins, abscisic acid (ABA), salicylic acid (SA), jasmonic acid (JA), ethylene, and brassinosteroid (BR) also play positive or negative roles in the regulatory network of flowering (Mutasa-Göttgens and Hedden, 2009; Davis, 2009; Diezel et al., 2011; Campos-Rivero et al., 2017).
Flowering transition is also regulated by environmental factors such as day and night length, drought, salinity, cold, and heat (Kazan and Lyons, 2016; Takeno, 2016; Lee et al., 2023). In response to drought stress, plants tend to accelerate flowering, a process called drought escape (Sherrard and Maherali, 2006; Franks et al., 2007; Bernal et al., 2011). The photoperiodic flowering pathway is closely related to drought-induced flowering regulation. In Arabidopsis, drought stress promotes early flowering under long days by activating FT gene expression, whereas it delays flowering transition under short days by increasing FLC gene expression. In contrast, salinity stress usually delays flowering (Kim et al., 2007), and several key flowering regulators involved in this response have been identified (Ryu et al., 2014; Ma et al., 2015).
To investigate the molecular mechanisms underlying rapid flowering of E. triticeum in desert environments, and to identify key genes and regulatory networks involved in flowering time control, RNA-seq analyses were performed at three developmental stages. Potential candidate differentially expressed genes (DEGs) involved in flowering regulation were identified and their functional properties were investigated to understand the molecular mechanism of early flowering in E. triticeum. In addition, the expression patterns and ectopic expression of the EtFT-1 gene were examined to explore its role in flowering time regulation. This study represents the first attempt to elucidate the genetic regulation of flowering in E. triticeum and provides insights into the molecular regulatory mechanism of flowering that might be shared with the other spring ephemerals.
Materials and methods
Plant material and treatment
In 2021, E. triticeum seeds were sown in the experimental field at Shihezi University, Xinjiang, China (43°26′N, 84°58′E). Tissue samples were collected for RNA sequencing at three different developmental stages: vegetative stage (VS), heading stage (HS), and flowering stage (FS). At the VS stage, 12 days after germination, leaves were collected as controls. Spikelets were collected at the HS stage, which occurred 42 days after germination, when they were just heading, and at the FS stage, which occurred 67 days after germination. Ten E. triticeum plants per sample were used for each developmental stage. In particular: Four leaves from each plant were gathered at the vegetative stage (VS). A single spikelet from each plant was gathered at the heading and flowering stages. Three distinct biological replicates yielded a total of nine samples for transcriptome sequencing. All of the samples were immediately frozen in liquid nitrogen and stored at -80°C.
For diurnal expression analysis, E. triticeum seeds were sown in a pre-mixed soil substrate (nutrient soil:vermiculite:perlite = 3:1:1) and grown in a growth chamber under long-day (LD, 16 h light/8 h dark) or short-day (SD, 8 h light/16 h dark) conditions, with temperatures ranging from 18-22°C and relative humidity between 40% and 60%. After three weeks, young leaves were collected every 4 h for a period of 24 h, starting from time 0 (ZT0) in both LD and SD photoperiods. Collection was completed within 48 h, and leaf samples were collected at the end of each photoperiod. Five leaves from young plants were collected per sample, with three biological replicates for each condition. All samples were immediately snap frozen in liquid nitrogen and stored at -80°C until further analysis.
Illumina sequencing and gene functional annotation
1 µg of RNA was used for RNA sample preparation. Sequencing libraries were prepared using the NEBNext® Ultra™ RNA Library Prep Kit for Illumina® (NEB, USA) according to the manufacturer’s guidelines, and index codes were added to associate sequences with each sample. Briefly, mRNA was purified from total RNA using poly-T oligo-attached magnetic beads. Fragmentation was performed with divalent cations at high temperature in NEBNext First Strand Synthesis Reaction Buffer (5X). First-strand cDNA was synthesized using random hexamer primers and M-MuLV reverse transcriptase. Second-strand cDNA was synthesized using DNA Polymerase I and RNase H. PCR was performed using Phusion High - Fidelity DNA Polymerase, Universal PCR Primers and Index (X) Primer. Finally, the PCR products were purified (AMPure XP system) and the library quality was evaluated on the Agilent Bioanalyzer 2100 system. Transcriptome sequencing was performed on an Illumina HiSeq 2000 platform with three independent biological replicates. The RNA-sequencing was performed by Biomarker Technologies (Beijing, China).
The sequences were further processed with a bioinformatic pipeline tool, BMKCloud (www.biocloud.net) online platform. To ensure high quality clean data, adapter sequences, poly-N sequences and low-quality reads were removed from the raw data. Subsequently, the Q20, Q30, GC content and sequence duplication levels of the clean data were calculated to assess the quality of the sequencing. De novo assembly of the transcriptome was performed using Trinity software (Grabherr et al., 2011). Using DIAMOND software (Buchfink et al., 2015), unigene sequences were compared to several databases, such as NR, Swiss-Prot, COG, KOG, eggNOG4.5, and KEGG. KOBAS (Xie et al., 2011) was used to get KEGG orthology data. The sequences were analyzed using InterProScan (Jones et al., 2014), a program that combines information from several databases, to produce Gene Ontology (GO) annotations. Additionally, HMMER software (Eddy, 1998) was used to compare unigene amino acid sequences to the Pfam database in order to identify functional domains.
Differential expression analysis, gene ontology and KEGG pathway enrichment analysis
Expression levels were quantified using the fragments per kilobase per million reads (FPKM) method (Trapnell et al., 2010). Differential expression between groups was analyzed using the DESeq2 package (Love et al., 2014). To control for false positives, we applied the Benjamini-Hochberg method to adjust p-values, resulting in a false discovery rate (FDR). Differentially expressed genes were identified using the criteria of |log2 fold change (FC)| ≥ 1 and FDR < 0.01. FC represents the expression ratio between the two groups.
Gene Ontology (GO) enrichment analysis of the DEGs was performed using the R package topGO based on the Kolmogorov-Smirnov test. The KEGG database provides high-level information on the functions and utilities of biological systems (Kanehisa et al., 2004). KOBAS software was used to assess the statistical enrichment of DEGs in KEGG pathways (Xie et al., 2011).
Phylogenetic analysis
Protein sequences were obtained from the NCBI database. Homologous sequences were aligned using CLUSTALW, and a phylogenetic tree was constructed using MEGA 7.0 software with the neighbor-joining (NJ) method, with 1000 bootstrap replicates.
Quantitative real-time PCR analysis
Total RNA was isolated from each sample according to the protocol provided with the RNA-prep Pure Plant Kit (TIANGEN Biotech, China). Then, 1 µg of total RNA was used to synthesize first-strand cDNA using the PrimeScript RT Reagent Kit (Takara, DRR037A). To verify the reliability of the gene expression profiles from the RNA-seq data, nine genes, including EtFT-1 (c164910.graph_c0), EtFKF1 (c162671.graph_c3), EtPP2C (c149628.graph_c0), EtLHY (c162145.graph_c0), EtCO (c160085.graph_c2), EtPIF4 (c162394.graph_c2), EtGID1 (c161985.graph_c0), EtRGL1 (c81081.graph_c0), and EtBRI1 (c154802.graph_c1), involved in flowering regulation and hormone signaling, were selected for qRT-PCR on an ABI 7500 Fast real-time PCR system (Applied Biosystems, USA) using ChamQ Universal SYBR qPCR Master Mix (Vazyme, Nanjing, China). The qRT-PCR validation analyses were performed on the same nine samples as the RNA-seq analyses, with three technical replicates for each sample. Relative expression levels were evaluated using the 2-ΔΔCt method (Livak and Schmittgen, 2001). The 18S rRNA gene of E. triticeum (GenBank accession no. PQ072787) was used as an internal control. The reference treatment was vegetative stage (VS). The results of the statistical analysis were expressed as the mean ± standard deviation of the three replicates. Furthermore, the circadian expression level of EtFT-1 was assessed by qRT-PCR analysis, and the 2−ΔCt technique was used to quantify relative expression. The 18S rRNA gene of E. triticeum was served as an internal control. The specific PCR primers are listed in Supplementary Table 9.
Generation of EtFT-1 transgenic Arabidopsis
The open reading frame (ORF) of EtFT-1 was inserted into the plant expression vector pCAMBIA2300 at the Kpn I and Xba I restriction sites to construct the recombinant plasmid 35S::EtFT-1 under the control of the CaMV 35S promoter. The recombinant vector and empty pCAMBIA2300 vector were then transferred into Agrobacterium tumefaciens strain GV3101 by heat shock treatment. The validated recombinant vector was transformed into Arabidopsis (Col-0) by the floral dipping method (Clough and Bent, 1998). Homozygous transgenic Arabidopsis lines were selected on MS medium supplemented with 50 mg/L kanamycin. The overexpressing T3 transgenic lines were collected for subsequent experiments. Four weeks after planting, samples of wild-type (WT) and transgenic lines were collected for total RNA extraction and reverse transcription-polymerase chain reaction (RT-PCR) to analyze the expression levels of EtFT-1. The specific PCR primers are listed in Supplementary Table 9.
Data analysis
All experimental data were analyzed using SPSS 26.0 software. A one-way ANOVA was first performed, followed by a t-test and Duncan’s multiple comparisons to assess differences between groups. Statistical significance was set at p < 0.05 and p < 0.01, respectively.
Results
Sequencing, assembly and functional annotation
A total of nine samples from three developmental stages (VS, HS, and FS) were analyzed (Figure 1A), yielding 193,417,047 clean reads (equivalent to 57.78 GB) after removing adapter sequences, contaminants, and low-quality reads. The average Q30 and GC content was 94.71% and 53.35%, respectively. A total of 86,950 transcripts and 41,289 unigenes were identified from these contigs, with an average length of 1,149 base pairs and an N50 of 1,875 base pairs (Supplementary Table 1, Supplementary Table 2). To assess gene expression differences between pairs of samples, Pearson correlation coefficients were calculated and the correlation values for all nine datasets (41,289 unigenes in total) were visualized in a heatmap (Figure 1B). The results indicated that the three biological replicates at each time point showed a high degree of correlation.
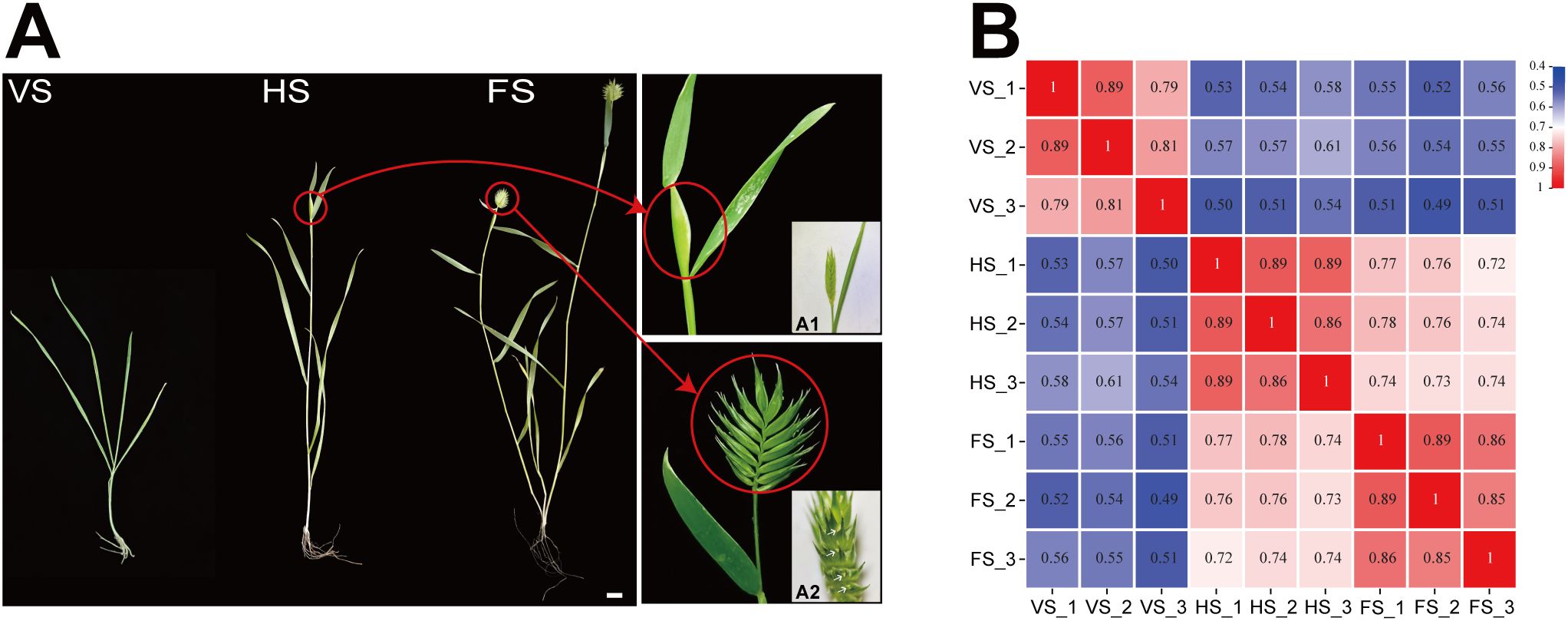
Figure 1. Three developmental stages of the vegetative growth and flowering stages of E. triticeum (A) and Pearson correlation coefficients of nine samples (B). A1. The spikelet at the heading stage under a stereomicroscope; A2. Lateral view of the spikelet at the flowering stage under a stereomicroscope. Arrows point to the florets. Scale bar: 10 mm. VS, vegetative growth stage; HS, heading stage; FS, flowering stage.
Functional annotation was performed against several publicly available nucleotide and protein databases. Homologs of the 41,289 unigenes were identified in the following databases with an E-value cutoff of 1e-5 (Supplementary Table 3): COG (7,573 unigenes, 18.34%), GO (21,179 unigenes, 51.29%), KEGG (16,067 unigenes, 38.91%), KOG (14,943 unigenes, 36.19%), Pfam (19,572 unigenes, 47. 40%), Swiss-Prot (15,576 unigenes, 37.72%), TrEMBL (26,295 unigenes, 63.69%), EggNOG (20,411 unigenes, 49.43%), and NR (26,545 unigenes, 64.29%). After eliminating redundancies from different databases, 28,956 unigenes were annotated at least once, representing about 70.13% of the total. Statistical comparison of the distributed E-values revealed that a remarkable 78.54% of the mapped sequences had high homology (E-value < 1e-30) and 60.22% had very high homology (E-value < 1e-50) when compared with plant sequences annotated in the NR database (Supplementary Figure 1A). Similarity distribution analysis showed that 85.09% of the unigenes had a similarity level of at least 60%, and 53.31% of the unigenes had a similarity level between 90% and 100% (Supplementary Figure 1B). Among the genes annotated in the NR database, the highest percentage of matches was associated with Aegilops tauschii (28.11%), Triticum turgidum (23.39%), and Hordeum vulgare (15.85%) (Supplementary Figure 1C).
Differentially expressed genes identification
To identify the DEGs related to flowering, we analyzed the transcript levels of each unigene at three developmental stages, and the DEGs were defined by a log2 (fold change) ≥ 1 and a false discovery rate (FDR) ≤ 0.01 (Figure 2). A total of 11,278 DEGs were identified in the E. triticeum RNA-seq data. Among them, 3,748 DEGs were found to vary in HS_vs_FS (2,267 up-regulated and 1,481 down-regulated) (Figure 2A). Venn diagram analysis revealed that 596 transcripts were differentially expressed in the flowering stage compared to the heading stage but not differentially expressed over the other comparisons of developmental stages (Figure 2B). In the volcano plot, the number of up-regulated DEGs was more significant than the number of down-regulated DEGs in VS_vs_HS, HS_vs_FS and VS_vs_FS (Figure 2C).
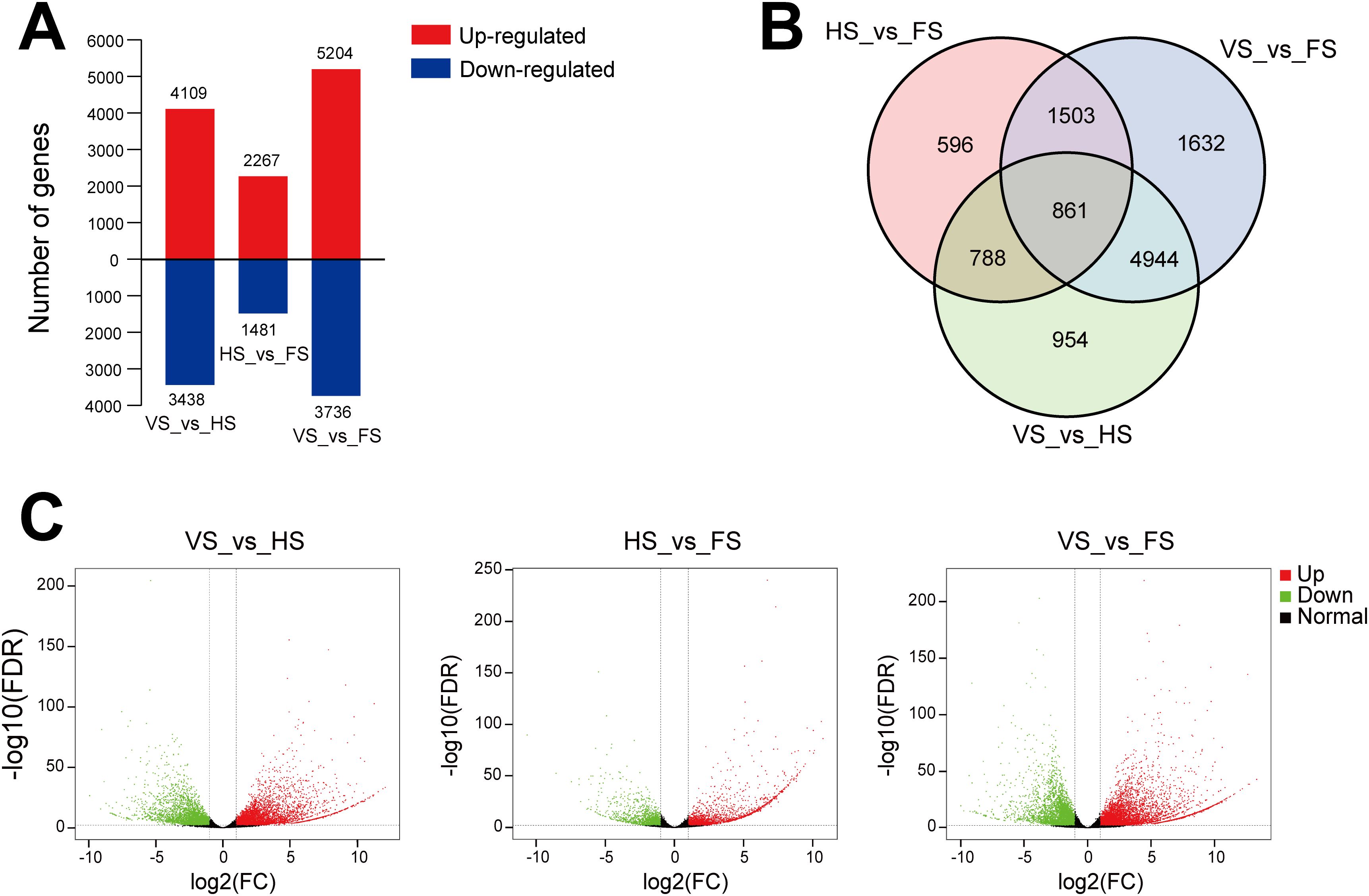
Figure 2. Transcriptome profiling of E. triticeum at three different developmental stages. (A) DEGs identified from different comparisons. (B) Venn diagram of DEGs at the three developmental stages. (C) Volcano plot of DEGs. Red and green dots indicate up- and down-regulated DEGs, respectively, and black dots indicate genes with no significant differences. VS, vegetative stage; HS, heading stage; FS, flowering stage.
GO enrichment analysis was used to determine the function of DEGs, and three main categories were identified, including molecular function, biological process, and cellular components (Figure 3A). A total of 28 functional GO terms were significantly enriched (P ≤ 0.05) in different biological processes (Supplementary Table 4). The most enriched molecular functions were protein kinase activity (GO: 0004672) and ATP binding (GO: 0005524). In the biological process category, macromolecule modification (GO: 0043412) and regulation of transcription (GO: 0006355) were significantly enriched. The most significantly enriched cellular component terms were integral component of membrane (GO: 0016021) and chloroplast thylakoid membrane (GO: 0009535).
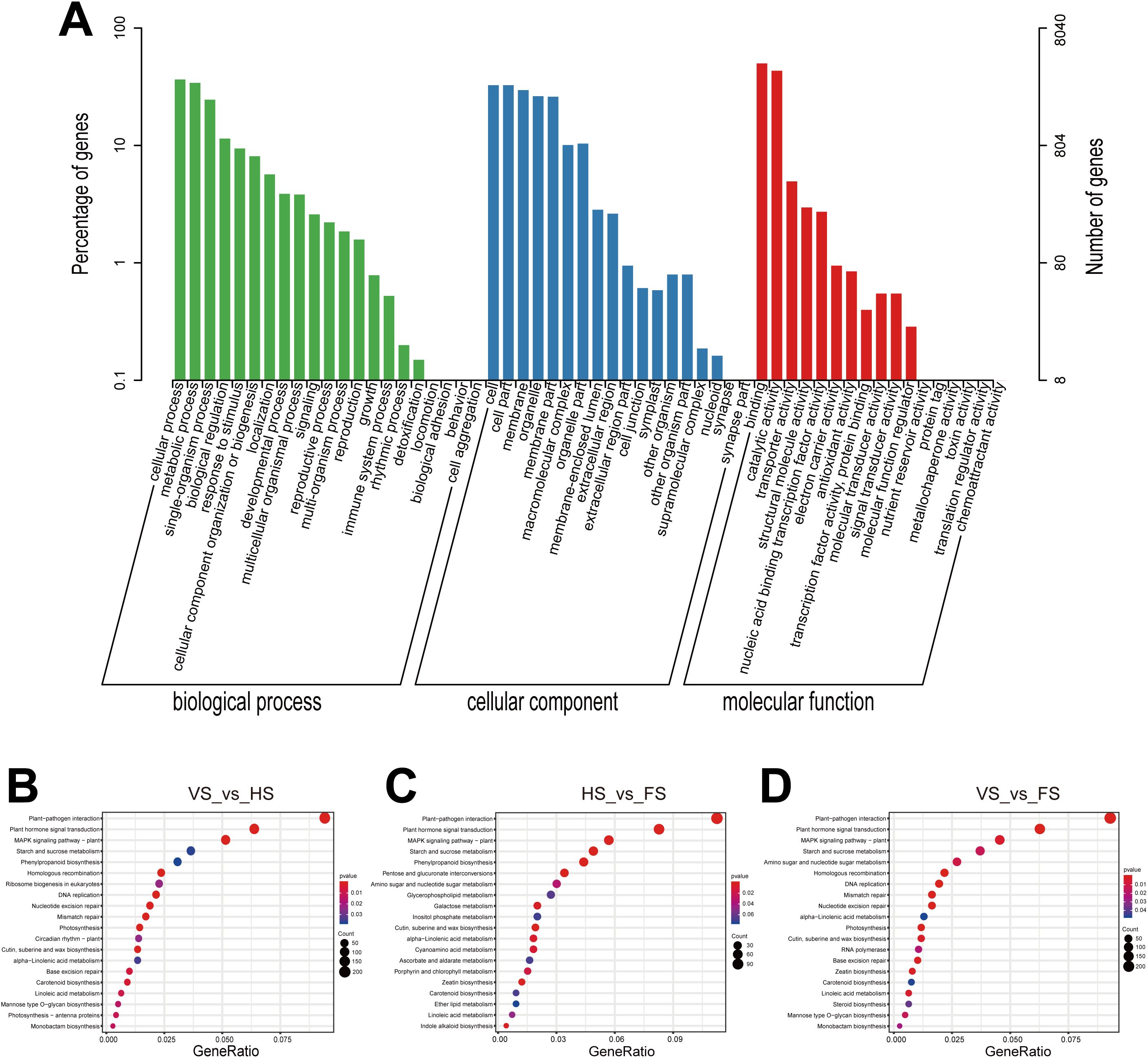
Figure 3. The GO classification and KEGG pathway enrichment of identified DEGs in E. triticeum. (A) GO classification of DEGs. (B) KEGG enrichment bubble plot of DEGs in the VS_vs_HS comparison. (C) KEGG enrichment bubble plot of DEGs in HS_vs_FS comparison. (D) KEGG enrichment bubble plot of DEGs in the VS_vs_FS comparison. The x-axis represents the gene ratio and the y-axis represents the pathways. The size of the bubbles indicates the number of genes enriched in each KEGG pathway. Larger bubbles represent more genes. The color of each bubble indicates the level of significance. VS, vegetative stage; HS, heading stage; FS, flowering stage.
The KEGG pathway enrichment analysis was further used to understand the expression of the DEGs in metabolic pathways, and the p-values ≤ 0.05 were considered as a significant enrichment. The top 20 most enriched KEGG pathways are shown in Figure 3B-D and Supplementary Table 5A-C. The DEGs were mainly enriched in pathways such as plant-pathogen interaction, plant hormone signal transduction, MAPK signaling pathway, starch and sucrose metabolism, phenylpropanoid biosynthesis, pentose and glucuronate interconversion, amino sugar and nucleotide sugar metabolism.
Identification of the DEGs related to flowering
In this study, six major flowering pathways, namely photoperiod (plant circadian rhythm), vernalization, autonomous, aging, temperature, and GA pathways, were analyzed based on transcriptome data. The expression variations of genes associated with these flowering pathways were then compared across three developmental stages. Based on the GO and KEGG annotations of the DEGs, 188 candidate genes related to flowering regulation pathways were initially identified. Among them, 92 DEGs involved in flowering transition were selected (Supplementary Table 6). The results showed that most of the DEGs were associated with the photoperiod pathway (48 DEGs), followed by the GA pathway (22 DEGs), the autonomous pathway (8 DEGs), the vernalization pathway (6 DEGs), the aging pathway (5 DEGs), and the temperature pathway (2 DEGs) (Figure 4A).
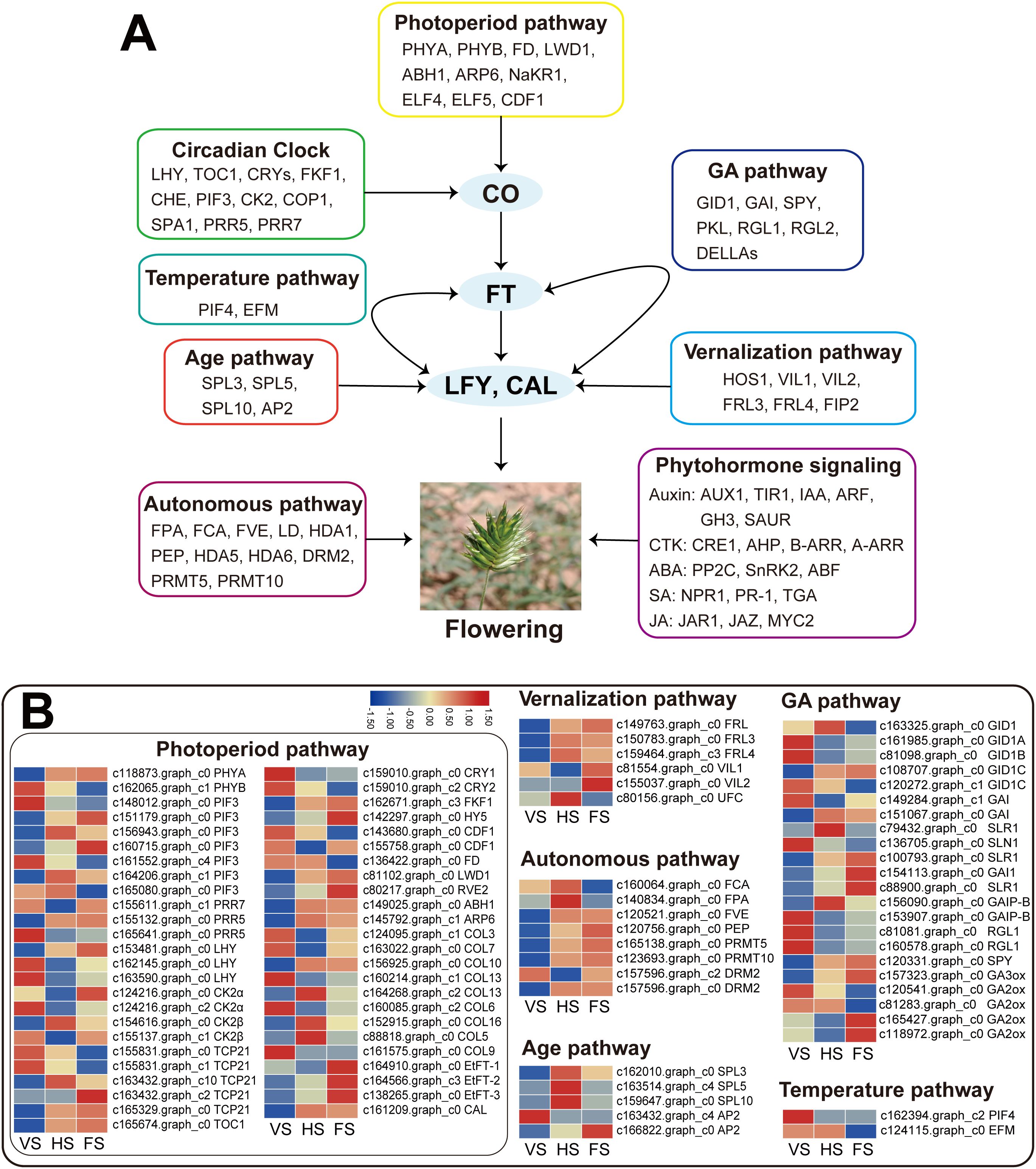
Figure 4. The major flowering pathways in E. triticeum. (A) A putative model of the flowering regulatory network in E. triticeum. (B) The heatmaps of the expression patterns of differentially expressed genes (DEGs) related to flowering in three developmental stages: vegetative stage (VS), heading stage (HS), and flowering stage (FS).
In the E. triticeum RNA-seq data, several important genes in the photoperiod and circadian rhythm pathway were identified. These include RVE2 (REVEILLE 2), PIF3 (phytochrome-interacting factor 3), LATE ELONGATED HYPOCOTYL (LHY), TCP, HY5, COL, and TIMING OF CAB EXPRESSION 1 (TOC1). These genes were found to be up-regulated in the flowering stage compared to the heading stage. Furthermore, four FT-like genes were identified in the three developmental stages, and three of them, including EtFT-1 (c164910.graph_c0), EtFT-2 (c164566.graph_c3), and EtFT-3 (c138265.graph_c0), showed significant up-regulation in the flowering stage (Figure 4B, Supplementary Table 6A). Given that the COL gene is an important core regulator in the flowering pathway, we investigated the evolutionary relationship of the EtCOL gene in E. triticeum with those in other plants. Phylogenetic tree analysis revealed that the EtCOL genes are closely related to the COL gene family in wheat and barley (Supplementary Figure 2A). The floral meristem identity genes AP1 and FRUITFUL (FUL) were not detected in the E. triticeum RNA-seq data. However, an AP1 paralog, the CAULIFLOWER (CAL) gene, was identified and was found up-regulated at both the head and flowering stages (Figure 4, Supplementary Table 6A, B). In addition, the floral organ identity gene LEAFY (LFY) was identified, but no differential expression was observed in the E. triticeum transcriptome data (Supplementary Table 6A).
In the vernalization pathway, two VIN3-like (VIL) genes showed differential expression, with significantly higher expression levels observed at the flowering stage. In addition, FRIGIDA-like (FRL) genes (c149763.graph_c0, c150783.graph_c0, and c159464.graph_c0), which were identified as floral repressors that play a crucial role in regulating flowering time in Arabidopsis (Wang et al., 2006), were found to be up-regulated at the flowering stage compared to the heading stage. Phylogenetic analysis revealed that two EtFRLs were closely related to the wheat TaFRLs, while the third showed high homology to the maize ZmFRL1 (Supplementary Figure 2B). These results suggest that EtFRLs may be involved in the regulation of vernalization-induced flowering in E. triticeum. Notably, the key regulator of vernalization, FLC, which plays a central role in Arabidopsis, was not detected in the E. triticeum transcriptome data, suggesting potential differences in the vernalization pathway between the two species. Similarly, the negative regulator VRN2, known to be involved in wheat vernalization, was not detected in the E. triticeum transcriptome.
In addition, several putative genes associated with the GA, autonomous, aging, and temperature pathways were identified in the RNA-seq data (Figure 4B, Supplementary Table 6). In the GA pathway, genes encoding GAI1 (GA-insensitive 1), SLR1 (slender rice 1), GA2ox (gibberellin 2-beta-dioxygenase), and GA3ox were up-regulated in the flowering stage compared to the heading and vegetative stages.
In the aging pathway, transcriptomic analysis revealed key regulators of floral development in E. triticeum, including orthologs of SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) and APETALA2 (AP2). The SPL gene family, which is critical for floral organ growth and fertility regulation through age-dependent pathways (Chao et al., 2017; Xing et al., 2010), showed stage-specific expression patterns: SPL3, SPL5, and SPL10 showed the highest expression levels at the heading stage. Phylogenetic analysis revealed that EtSPL3 was closely clustered with wheat TaSPL3, and EtSPL5 was closely clustered with wheat TaSPL5 (Supplementary Figure 3C), suggesting a similar role in wheat flowering.
In addition, an AP2 homolog (c166822.graph_c0), which is involved in spikelet determinacy and the transition to floral meristem development (Chuck et al., 2002; Komatsu et al., 2003), was found to be significantly up-regulated at the flowering stage. These results suggest that AP2 may play a critical role in modulating floral meristem identity and shaping inflorescence architecture in E. triticeum.
Identification of hormone-related DEGs involved in flowering
A total of 197 DEGs in the plant hormone signaling pathway (ko04075) were identified by KEGG pathway analysis (Supplementary Table 7). These genes include AUX1, TRANSPORT INHIBITOR RESPONSE 1 (TIR1), gibberellin responsive receptor (GID1), abscisic acid responsive element binding factor (ABF), and ethylene responsive receptor (ETR). These genes are involved in the gibberellin (GA), abscisic acid (ABA), auxin, brassinosteroid (BR), ethylene, gibberellin (GA), jasmonic acid (JA), and salicylic acid (SA) signaling pathways (Table 1).
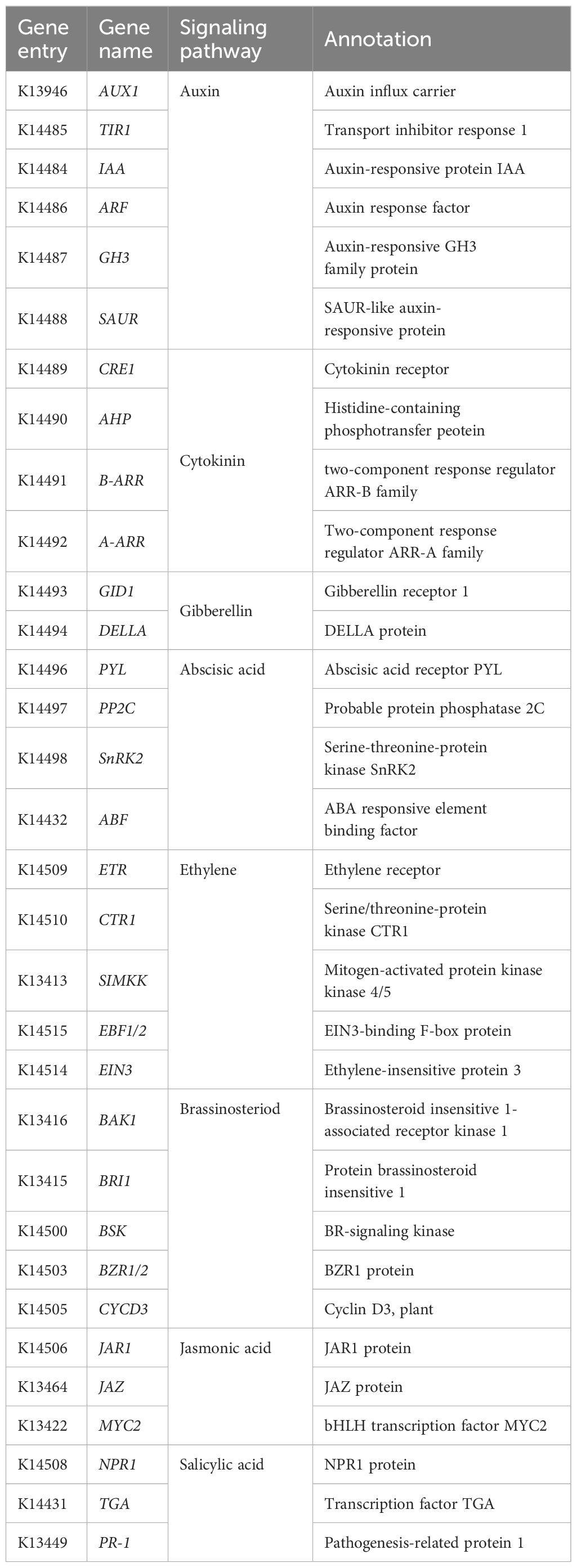
Table 1. The identified DEGs involved in hormone signal transduction pathways in E. triticeum.
In the auxin signaling pathway, six genes encoding indoleacetic acid (IAA), five genes encoding auxin response factor (ARF), three genes encoding Gretchen Hagen 3 (GH3), and six genes encoding small auxin-up RNA (SAUR) were upregulated at the flowering stage. In the ABA signaling pathway, four PP2C, two ABF, and one SnRK2 were up-regulated at the flowering stage. In the BR pathway, BAK1 and BRI1 genes were up-regulated at the flowering stage. Salicylic acid (SA), a hormone signaling molecule, plays an important role in plant immunity and stress response. SA is also involved in the regulation of flowering time. SA regulates flowering time in plants such as Arabidopsis by participating in the regulation of the expression of flowering-related genes such as CO, FLC, FT, FLD, etc. in the vernalization and photoperiodic pathways (Martínez et al., 2004). Disease progression-related (PR) proteins are induced in plants by the application of various biotic stresses (e.g., pathogens) and abiotic stresses (e.g., drought, cold, salinity, heavy metals, etc.). In this study, PR-1 was found to be significantly increased during the flowering stage of E. triticeum, suggesting that it plays an important role in flowering and resistance response in E. triticeum. In addition, the genes TGA in the SA pathway and ARR in the GA pathway were upregulated at the flowering stage (Figure 5). These results suggest that the accumulation of phytohormones and hormone-related transcripts is significantly modulated during the flowering stage of E. triticeum.

Figure 5. Expression patterns of the differentially expressed genes (DEGs) in the plant hormone signaling pathways.
Identification of flowering pathway-related transcription factors
In this study, a total of 410 DEGs belonging to 49 transcription factor (TF) families were identified, including bHLH, MYB, AP2/ERF, WRKY, bZIP, C2C2, and MADS-box. We then detected the expression levels of these genes in the RNA-seq data during the three developmental stages in E. triticeum. Notably, a number of TF families exhibited high expression levels at the head or flowering stage. For example, 11 DEGs belonging to the bHLH family were found to be significantly up-regulated at the FS stage compared to the VS and HS stages (Figure 6). In addition, several WRKY and MYB transcription factors were identified as significantly upregulated at the FS stage, suggesting a potential regulatory role for these TFs in the flowering process of E. triticeum.

Figure 6. Differential expression of transcription factors at three developmental stages in E. triticeum. Red and green represent high and low expression levels, respectively.
MADS-box TFs play critical roles in inflorescence meristem and flower development (Ohmori et al., 2009; Gao et al., 2010). In this study, a total of 12 putative MADS-box genes were identified from the E. triticeum transcriptome data, and six genes were predominantly expressed in the flowering stage compared to the vegetative and heading stages (Figure 6). To better understand the functions of the E. triticeum MADS-box genes, we constructed a phylogenetic tree using 98 MADS-box protein sequences from Arabidopsis and wheat (Supplementary Figure 3). The results indicated that the E. triticeum MADS box could be divided into two groups: M-type (5 genes, 3 Mα and 2 Mγ) and MIKC-type (7 genes). Among the MIKC-type MADS box genes, the Arabidopsis and wheat orthologs of AG, AGL12, SOC, TT16, SVP, and AP3 were identified in E. triticeum. Gene expression patterns revealed that two TT16 (c122821.graph_c0, and c165713.graph_c0), one AG (c162082.graph_c8), and one AP3 (120438.graph_c0) were up-regulated at the flowering stage (Figure 6). These results suggest that these TFs may play an essential role in the regulation of flowering in E. triticeum.
qRT-PCR validation
To validate the accuracy of the transcriptome results, nine genes were selected for qRT-PCR analysis, including seven genes related to growth and flowering regulation and two genes related to hormone signaling (Figure 7). The results showed that the EtFT-1, EtFKF1, and EtPP2C were significantly up-regulated at the flowering stage. These genes have been shown to be important for the circadian clock and the regulation of photoperiodic flowering in Arabidopsis (Nelson et al., 2000). Overall, the expression trends of these genes were highly consistent with the RNA-seq.
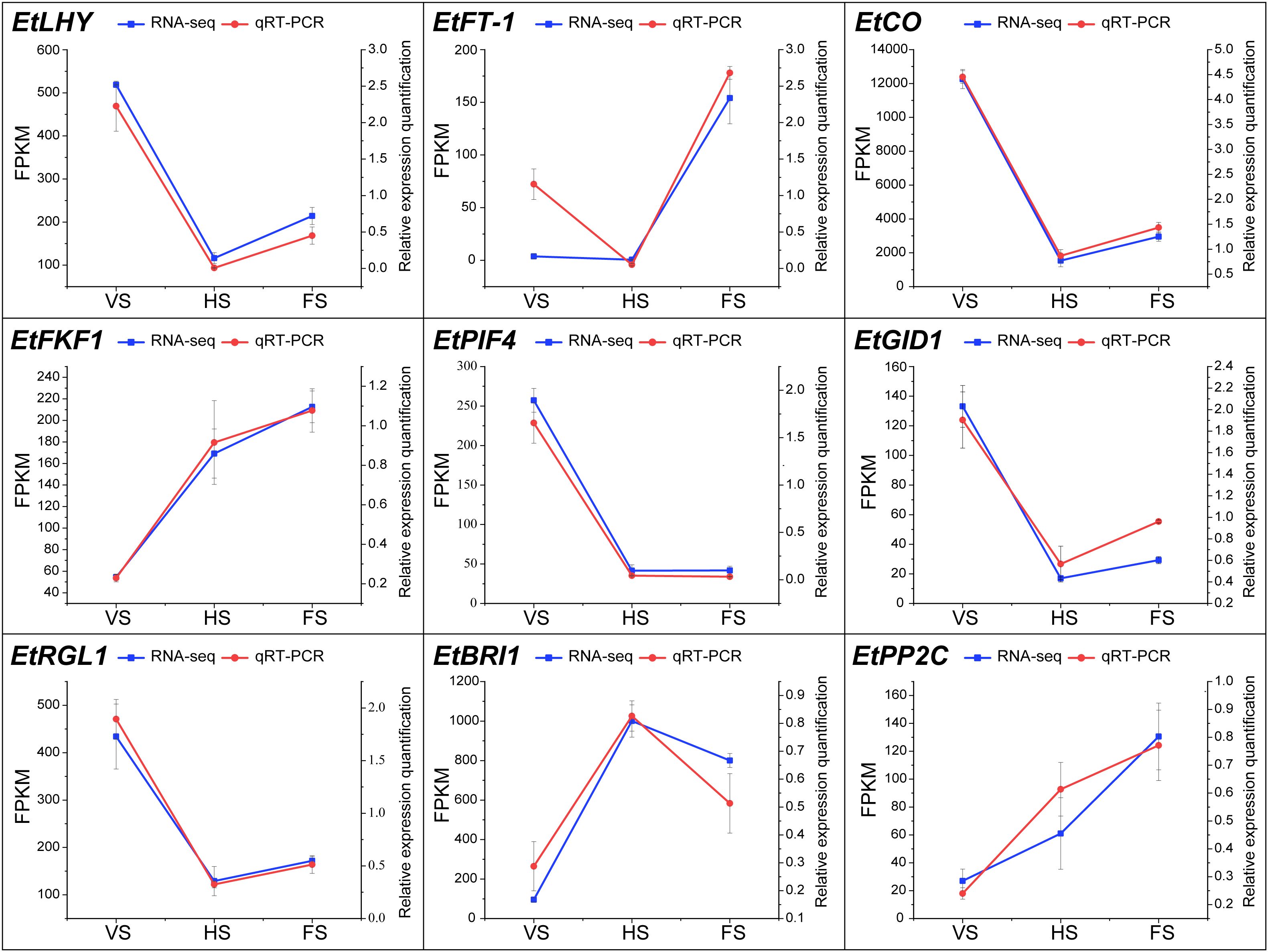
Figure 7. Expression validation of genes between qRT-PCR and RNA-seq at different developmental stages in E. triticeum. Red and blue represent qRT-PCR and RNA-seq data, respectively.
Identification and expression pattern analysis of EtFT genes
The FT and TFL1 subfamilies have undergone functional divergence in flowering regulation, with FT activating and TFL1 repressing flowering, respectively (Wang et al., 2015). In this study, four FT-like genes, namely EtFT-1, EtFT-2, EtFT-3, and EtFT-4, were identified in the transcriptomic data of E. triticeum. Sequence alignment revealed that the EtFT proteins contain a conserved PEBP domain (Figure 8A). The conserved residues Tyr85 (Y) and Gln140 (Q) in Arabidopsis FT were identified in EtFT-1, EtFT-2, and EtFT-3, which are crucial for FT activity and diagnostic for differentiating FT from TFL1 (Hanzawa et al., 2005; Ahn et al., 2006). However, in EtFT-4, the conserved residue Gln140 (Q) was replaced by Ser140 (S), which is similar to Triticum aestivum TaFTL (XP_044320677.1) and Hordeum vulgare HvFTL (XP_044970677.1) (Figure 8A). In addition, the EtFT proteins contained a conserved outer loop structure (LGRQTVYAPGWRQN) in segment B and the LYN/IYN triad in segment C, corresponding to a potential ligand binding site in FT/TFL1 family proteins (Ahn et al., 2006).
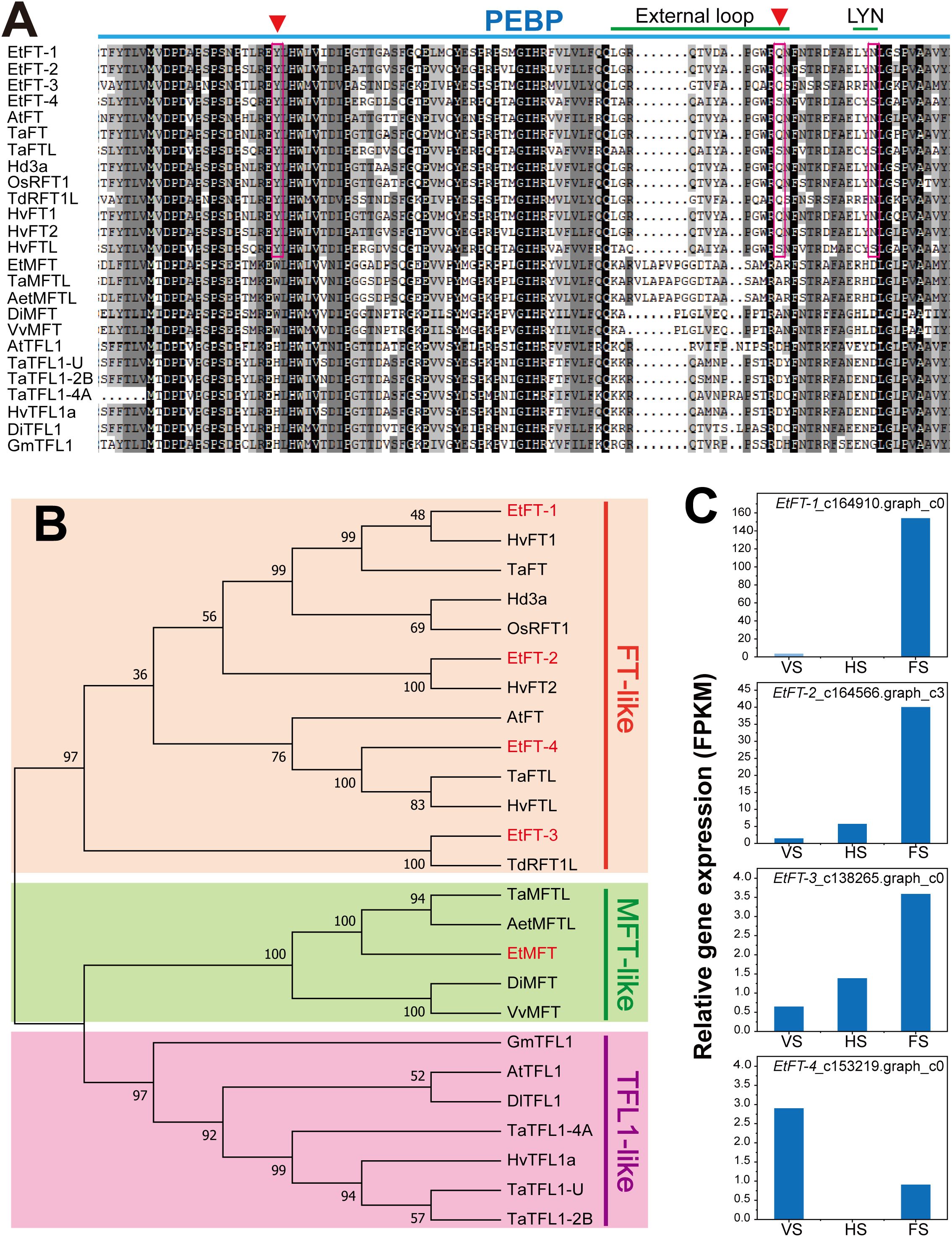
Figure 8. Amino acid alignment, phylogenetic tree and expression analysis of EtFTs. (A) Multiple sequence alignment of EtFTs from E. triticeum, along with FT, TFL and MFT proteins from different plant species. AtFT (Arabidopsis thaliana, At1g65480); TdRFT1L (Triticum dicoccoides, XP_037476013); Hd3a (Oryza sativa, AB052941); OsRFT1 (Oryza sativa, NP_001056859); TaFT (Triticum aestivum, AAW23034); TaFTL (Triticum aestivum, XP_044320677.1); HvFT1 (Hordeum vulgare, AAZ38709); HvFT2 (Hordeum vulgare, ABB99414); HvFTL (Hordeum vulgare, XP_044970677.1); TaMFTL (Triticum aestivum, XP_044432627.1); AetMFTL (Aegilops tauschii, XP_020153395.1); DiMFT (Dimocarpus longan, AUG98253); VvMFT (Vitis vinifera, NP_001267935); TaTFL1-U (Triticum aestivum, TraesCSU02G202000.1); TaTFL1-2B (Triticum aestivum, TraesCS2B02G310700.1); TaTFL1-4A (Triticum aestivum, TraesCS4A02G409200.1); HvTFL1a (Hordeum vulgare, HORVU2Hr1G072750.1); GmTFL1 (Glycine max, FJ573238); AtTFL1 (Arabidopsis thaliana, At5g03840); DlTFL1 (Dimocarpus longan, AHY24028). (B) Phylogenetic analysis of four EtFT encoding proteins: EtFT-1 (c164910.graph_c0), EtFT-2 (c164566.graph_c3), EtFT-3 (c138265.graph_c0), and EtFT-4 (c153219.graph_c0)) with FT, TFL, and MFT from different plant species. The phylogenetic tree was generated by MEGA 7 software using the neighbor-joining (NJ) method, and the percentage of replicate trees from the bootstrap test (1000 replicates) is indicated beside the branches. Red, green and blue branches represent FT-like, MFT-like, and TFL1-like protein, respectively. (C) The relative expression levels of EtFT-1, EtFT-2, EtFT-3 and EtFT-4 genes at three different stages of E. triticeum. Sample labels are as follows: VS, vegetative stage; HS, heading stage; FS, flowering stage.
To further investigate the evolutionary relationships of the PEBP family, a phylogenetic tree of EtFT proteins with other plant species was constructed using the neighbor-joining (NJ) method. The results indicated that EtFT-1 was more closely related to Triticum aestivum TaFT and Hordeum vulgare HvFT1, EtFT-2 was closely related to HvFT2, and EtFT-3 was clustered with Triticum dicoccoides TdRFT1L (Figure 8B).
To predict the biological function of EtFT genes, we analyzed the dynamic expression patterns of EtFT genes in the transcriptional data of E. triticeum. The results showed that EtFT-1, EtFT-2, and EtFT-3 were all significantly up-regulated in the spikelets at the flowering stage. Among them, EtFT-1 showed the highest expression at the flowering stage compared to the other EtFT genes (Figure 8C), suggesting its critical role in the regulation of flowering in E. triticeum.
Expression patterns of EtFT-1 under different day length
Previous studies have shown that FT expression is responsive to day length. For example, in Arabidopsis, FT mRNA accumulates only under long days (LDs) (Suárez-López et al., 2001), whereas rice Hd3a, a homolog of FT, accumulates mRNA under short days (SDs) (Izawa et al., 2002; Kojima et al., 2002). To investigate whether the accumulation of EtFT-1 mRNAs follows a similar pattern related to photoperiod-induced flowering, the levels of EtFT-1 mRNAs were examined under LDs (16 h light/8 h dark) and SDs (8 h light/16 h dark).
The EtFT-1 mRNA exhibited a diurnal rhythm in both LD and SD-grown E. triticeum plants. EtFT expression underwent a more pronounced induction under SD than under LD conditions (Figure 9). When plants were exposed to LD conditions, the EtFT-1 showed a low expression level. However, the transcript level of EtFT-1 was increased after shifting to darkness and peaked after 4 h of dark treatment (Figure 9A). When plants were exposed to SD conditions, EtFT-1 mRNA showed a pronounced peak around the time of the onset of night (Figure 9B). These results demonstrate that unlike FT in Arabidopsis, EtFT-1 expression is likely induced in darkness, and short-day conditions promote its expression.
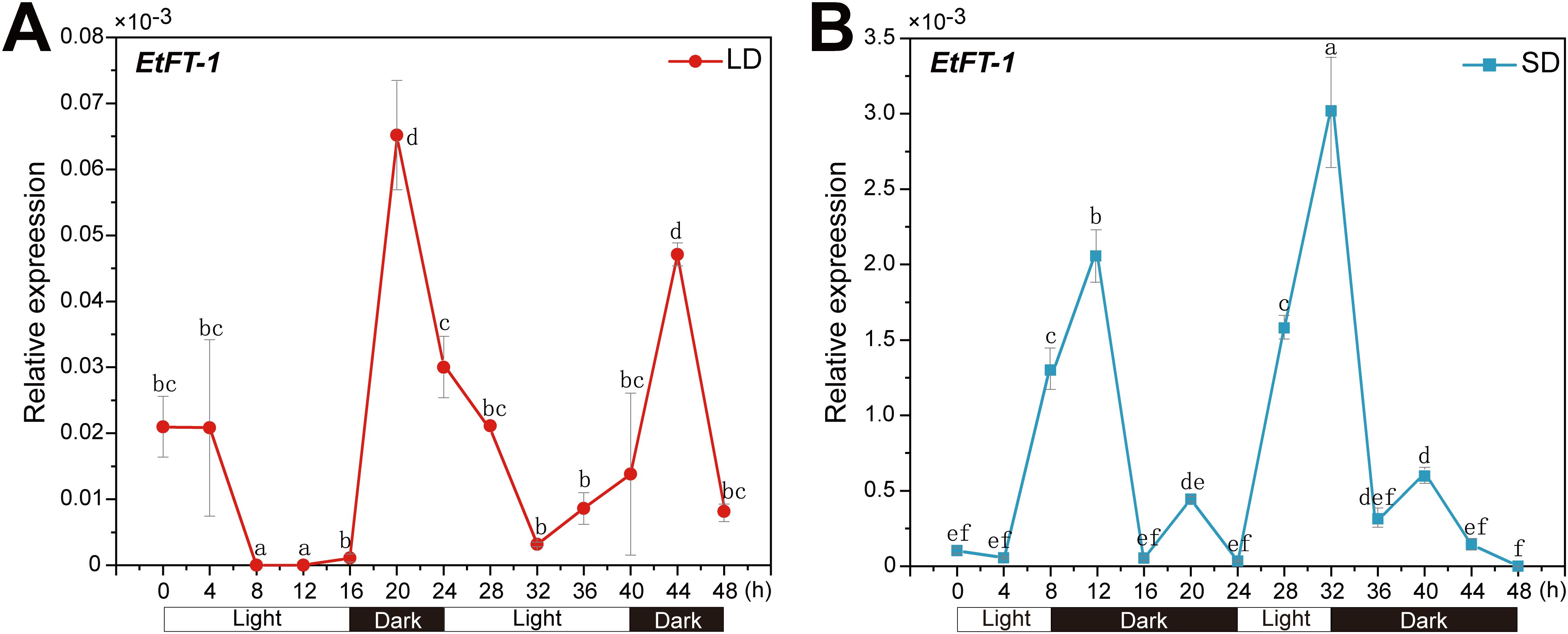
Figure 9. Expression of EtFT-1 under different photoperiods. Transcript levels of EtFT-1 in the leaves of 3-week-old E. triticeum plants grown under long-day (LD, 16 h light/8 h dark) or short-day (SD, 8 h light/16 h dark) conditions by qRT-PCR, with three biological replicates. The 18S rRNA gene of E. triticeum (GenBank accession No. PQ072787) served as the internal control. Relative expression levels were evaluated by the 2−ΔCt method. (A) Expression of EtFT-1 under LD conditions. (B) Expression of EtFT-1 under SD conditions. Different letters (a, b, c, etc.) indicate significant differences between groups (P < 0.05), while the same letter indicates non-significant differences between groups.
Overexpression of EtFT-1 accelerates flowering in Arabidopsis
To study the effect of EtFT-1 on promoting flowering in Arabidopsis transgenic plants, the EtFT-1 overexpression vector driven by the CaMV 35S promoter was constructed and introduced into Arabidopsis (Col-0) by Agrobacterium-mediated transformation. Homozygous transgenic Arabidopsis lines carrying the EtFT-1 gene were confirmed based on their resistance to kanamycin. Phenotypic analysis was performed on these homozygous plants of the T3 generation, using wild-type (WT) Arabidopsis as a control. Genomic DNA was extracted from individual homozygous EtFT-1 transgenic Arabidopsis and WT plants. The target EtFT-1 gene was then amplified by PCR using specific primers for molecular identification of the transgenic lines (Figure 10A). Semi-quantitative RT-PCR was performed to assess the expression of EtFT-1 in the transgenic lines, and three independent lines with higher EtFT-1 expression (OE-2, OE-4, and OE-6) were selected for further functional analysis (Figure 10B).
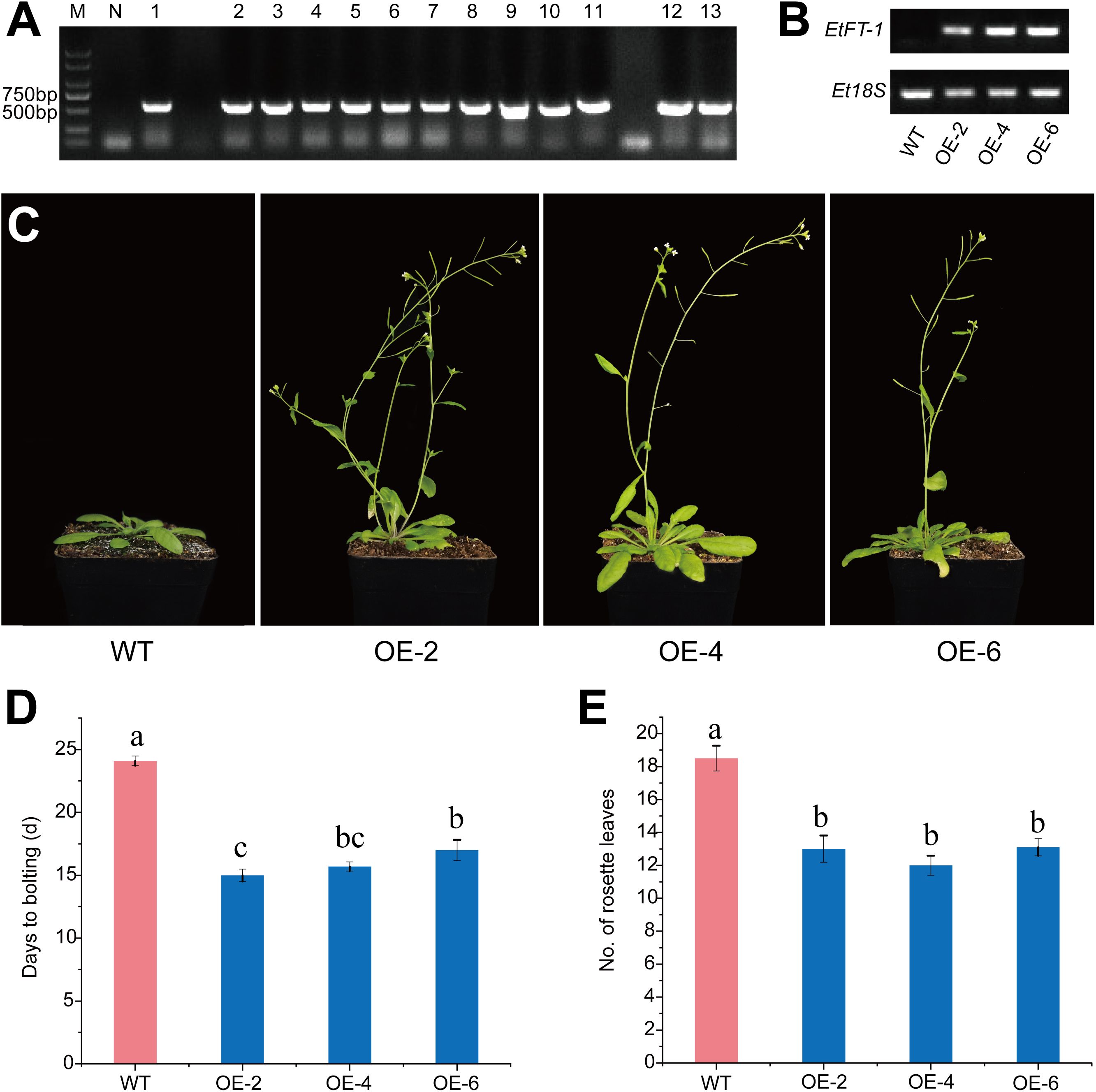
Figure 10. Flowering phenotypes of EtFT-1 overexpressing lines under 16 h light/8 h dark photoperiod. (A) Resistance screening and PCR identification of transgenic A. thaliana. M: Marker 2000; N: Negative, template is ddH2O; 1-13: PCR identification of transgenic A. thaliana. (B) Semiquantitative RT-PCR analysis of EtFT-1 overexpressing Arabidopsis lines (OE-2, OE-4, and OE-6) and wild-type (WT) plants. (C) The flowering phenotypes of EtFT-1 overexpressing lines under a 16 h light/8 h dark photoperiod. (D) The bolting time in WT and transgenic plants grown under a 16 h light/8 h dark photoperiod. (E) The number of rosette leaves of WT and transgenic plants at the bolting stage under a 16 h light/8 h dark photoperiod. Different letters (a, b, c, etc.) indicate significant differences between groups (P < 0.05), while the same letter indicates non-significant differences between groups.
The results showed that the EtFT-1 overexpressing lines exhibited earlier bolting (by 6–10 days) and flowering (by 9–10 days) compared to WT plants (Figures 10C, D, Supplementary Table 8). In addition, the number of rosette leaves was significantly reduced in the EtFT-1 overexpressing lines compared to WT plants (Figure 10E). These results indicate that ectopic overexpression of EtFT-1 promotes early flowering in Arabidopsis.
Discussion
DEGs associated with flowering in E. triticeum are mainly related to the photoperiod and vernalization pathways
In the plant life cycle, floral transition has been extensively studied in the model plants Arabidopsis and rice in recent years. However, the molecular mechanisms of floral transition in desert ephemerals remain poorly understood. E. triticeum is a widely distributed annual ephemeral species in northern Xinjiang, especially in the Junggar Desert. In extreme desert environments, E. triticeum is capable of rapid growth, flowering, and reproduction, completing its life cycle in about two months to avoid severe summer drought conditions. However, little is known about the molecular mechanisms of the floral transition in E. triticeum, largely due to the limited genomic data available in public databases. RNA-seq is a powerful tool that has been used to discover novel transcripts and gene regulatory networks. It has become a valuable research tool in biological studies. In this study, 92 flowering-related DEGs were identified (Figure 4; Supplementary Table 6). These genes were mainly associated with photoperiod, vernalization, gibberellin, autonomous, and age pathways. Notably, several homologs were identified that function as key activators and transcription factors related to flowering in E. triticeum, such as the EtFT gene family in the photoperiod pathway, the EtFRL family in the vernalization pathway, and the EtAP2 family in the aging pathway.
The influence of the photoperiodic pathway on flowering time is closely linked to circadian rhythm. In many annual plants, flowering time is influenced by the duration of daily light exposure. In Arabidopsis, long days promote flowering, while short days inhibit flowering. The COL gene, a downstream gene in the photoperiod pathway, controls the transition to flowering by transducing the light signal to the flowering integrator FT (Imaizumi, 2010). Both COL and FT are important mediators in the photoperiodic flowering pathway in Arabidopsis. Under long-day conditions, AtCO stimulates FT expression in leaves (Wong et al., 2013). In rice, the CO homolog Hd1 promotes FT expression under short days, but suppresses Hd3a transcription under long days, thereby delaying flowering (Kojima et al., 2002). COL gene expression is regulated by the circadian clock, and the clock component GI contributes to the formation of the GI-FKF1 complex, which is triggered by light to upregulate CO expression under long days (Sawa et al., 2007; Yang et al., 2024). In this study, we found that most of the flowering-related genes in E. triticeum are associated with the photoperiod pathway, suggesting its central role in regulating flowering in E. triticeum. However, the detailed regulatory mechanism requires further investigation.
Previous studies have shown that the molecular mechanisms of flowering transition are conserved to some extent between the dicot Arabidopsis and the monocot wheat, although clear differences remain. For example, the key Arabidopsis vernalization regulator gene FLC has not been identified in wheat (Milec et al., 2014; Xu and Chong, 2018). In wheat, vernalization is regulated by the VRN1, VRN2, and VRN3 genes. The wheat VRN1 gene is not related to Arabidopsis VRN1, but is homologous to the Arabidopsis AP1/CAL/FUL genes (Blümel et al., 2015). VRN2, a CCT protein, has no Arabidopsis homologue and acts as a negative regulator of wheat flowering (Yan et al., 2004). The wheat VRN3 gene is an ortholog of FT and promotes flowering in a manner similar to FT in Arabidopsis (Yan et al., 2006). In this study, we identified a VRN1 gene homolog, the CAL gene, in the E. triticeum RNA-seq data, and it was significantly up-regulated at the flowering stage (Figure 4). This suggests that it may play an important role in the vernalization induction of E. triticeum. However, the negative regulator of wheat vernalization, VRN2, was not detected in the E. triticeum transcriptome, possibly due to its non-expression or low expression in the tissues tested. This suggests that the vernalization-mediated flowering pathway in E. triticeum may differ from that in wheat. Further research is needed to fully elucidate this mechanism.
Complex transcriptional regulatory network of flowering in E. triticeum
An increasing number of studies have shown that several transcription factor families, including bHLH, MYB, Dof, bZIP, WRKY, and MADS-box, play essential roles in regulating the flowering transition (Waseem et al., 2019; Song et al., 2024; Zhang et al., 2020; Zhou et al., 2019a). In this study, 49 transcription factor families were identified from the E. triticeum transcriptome data, with many members of the bHLH, MYB, and MADS-box families showing up-regulated expression at the flowering stage (Figure 6). MYB FTs play an important role in plant development, participating in various biological processes such as stress and protective responses, seed and flower development (Zhou et al., 2019b). For example, overexpression of the wheat MYB transcription factor TaMYB72 in rice reduced flowering time by 12 days under long-day conditions (Zhang et al., 2016). In addition, members of the bHLH and MADS-box gene families are essential for plant reproductive development. For example, the Arabidopsis cib2 mutant exhibits an early flowering phenotype, and ectopic expression of the pineapple bHLH transcription factor AcCIB2 in the Arabidopsis Atcib2 mutant can complement the phenotype, indicating that AcCIB2 shares a similar biological function with AtCIB2 (Aslam et al., 2020). Similarly, ectopic overexpression of the PeMADS5 gene from Phyllostachys edulis resulted in early flowering and abnormal flower phenotypes in Arabidopsis (Zhang et al., 2018). We found that most of the bHLH, MYB, and MADS-box transcription factor genes in E. triticeum were highly up-regulated at the flowering stage (Figure 6), suggesting that these transcription factors play important roles in regulating flowering in E. triticeum.
These transcription factors are known to play important roles in plant responses to abiotic stresses. For example, bHLH transcription factors have been implicated in the regulation of iron homeostasis and stress responses in plants (Qian et al., 2021). Similarly, MYB transcription factors are involved in various stress responses (Li et al., 2019), in addition to the seed and flower development (Chopy et al., 2023). MADS-box transcription factors are critical for reproductive development and can interact with bHLH proteins to regulate floral organ formation. In addition, certain MADS-box genes are involved in responses to abiotic stresses such as salinity, drought, and cold (Di Marzo et al., 2020; Castelán-Muñoz et al., 2019). The up-regulated expression of these transcription factors during flowering may enhance the ability of E. triticeum to adapt to the arid desert environment by optimizing reproductive success before the onset of drought.
Phytohormones regulating flower development in E. triticeum
Flowering is controlled by a complex network of genes that work together to make sure it happens at the right time. Hormones, signaling, and internal balance are all important parts of this process (Campos-Rivero et al., 2017). KEGG enrichment analysis of the DEGs showed significant enrichment of plant hormone biosynthesis and signaling pathways during the flowering process in E. triticeum (Figure 5). These results were consistent with those observed in Camellia sasanqua and Mikania micrantha (Fan et al., 2022; Liang et al., 2023), suggesting that phytohormone biosynthesis and signal transduction play an important role in regulating the floral transition in E. triticeum.
Gibberellins (GAs) are essential for regulating several processes of plant growth and development, including seed germination, stem elongation, and the timing of flowering. In the established GA signaling pathway, GA binds to the soluble GA receptor GID1, which then interacts with the DELLA repressor protein. In petunia, three GID1 genes, namely PhGID1A, PhGID1B, and PhGID1C, have been identified, and their suppression by virus-induced gene silencing (VIGS) results in reduced growth and delayed flowering (Liang et al., 2014). In the present study, five putative EtGID1 genes were identified in E. triticeum, with the EtGID1C gene (c108707.graph_c0) showing high expression levels during the flowering stage (Figure 4). This finding suggests that the EtGID1C gene plays a role in the flowering process of E. triticeum.
Previous studies have shown that SA can affect flowering in several plants, including Sinningia speciosa, Lemna gibba, and Gazania rigens (Martín-Mex et al., 2015; Fu et al., 2020; Abdul Kareem and Saeed, 2020). In some species, SA is essential for the induction of flowering (Wada and Takeno, 2010). For example, stress-induced SA accumulation promotes earlier flowering in Arabidopsis and Lemna paucicostata (Martínez et al., 2004; Shimakawa et al., 2012). The SA signaling pathway genes PR-1 and TGA have been implicated in the regulation of floral organ development and flowering (Luo et al., 2022). For example, the tga7 mutant in Arabidopsis showed a delayed flowering phenotype under both long-day (LD) and short-day (SD) conditions (Xu et al., 2021). In this study, we found that most of the EtTGA and EtPR-1 genes were up-regulated at the flowering stage (Figure 5), suggesting that the SA pathway may play a regulatory role in flowering in E. triticeum.
EtFT-1 is involved in flowering time regulation in E. triticeum
The FLOWERING LOCUS T (FT) protein, a conserved florigen in angiosperms, integrates signals from multiple flowering pathways (e.g. photoperiod, vernalization) and is translocated from leaves to shoot apical meristems to initiate floral transition (Liu et al., 2021). Loss of FT gene function results in delayed flowering under long-day (LD) conditions, whereas its overexpression results in exceptionally early flowering regardless of day length (Kardailsky et al., 1999; Kobayashi et al., 1999). For example, overexpression of the Arabidopsis AtFT gene has been shown to promote early flowering in species such as cotton, apple, and poplar (McGarry et al., 2013; Yamagishi et al., 2011; Zhang et al., 2010).
In E. triticeum, we identified four FT homologs (EtFT-1, 2, 3, and 4), with EtFT-1 exhibiting the highest expression level during the flowering stage, suggesting its central role in flower induction. To investigate the function of EtFT-1, EtFT-1 was overexpressed in Arabidopsis, resulting in transgenic lines with significantly earlier bolting and flowering times compared to wild-type plants. However, the number of the rosette leaves in transgenic plants was reduced, which is a key feature in determining flowering time in Arabidopsis (Andrés and Coupland, 2012). These results suggest that EtFT-1 may plays a critical role in promoting flowering in E. triticeum.
Comparative research on a range of plant species has shown that FT homologs are always essential for controlling flowering timing. In rice, FT-like genes such as Hd3a and RFT1 are essential for flowering induction under different photoperiods (day and night lengths) (Komiya et al., 2009). Similarly, six FT homologs have been identified in soybean (Glycine max), with GmFT2a significantly promoting flowering under short-day conditions (Sun et al., 2011; Zhao et al., 2016). Four FT homologous genes are found in tobacco. While overexpression of NtFT1, NtFT2, and NtFT3 delays flowering, overexpression of NtFT4 promotes flowering (Harig et al., 2012). These investigations demonstrate the evolutionary conservation and functional diversity of FT homologs in controlling flowering in a range of plant species.
In this study, we characterized EtFT-1 through transgenic expression in Arabidopsis and confirmed its function in promoting flowering. The roles of the other EtFT homologs in the regulation of flowering, however, have yet to be clarified. Further studies should more closely examine the functional divergence of EtFT genes using transgenic methods in Arabidopsis and monocot species that are more closely related to E. triticeum, such as wheat. Furthermore, clarification of their regulatory networks, interacting proteins, and reactions to environmental signals like drought and photoperiod will offer a more profound understanding of the molecular mechanisms that control flowering in desert-adapted ephemerals.
Conclusion
In this research, RNA-seq was used to investigate gene expression patterns associated with heading and flowering in E. triticeum, revealing specific gene networks and key regulators. Transcripts associated with major flowering pathways, including photoperiod, vernalization, GA, and autonomous pathways, were identified, highlighting the importance of genes such as FT, FD, COL, SPL, GID, and transcription factors from the AP2, bHLH, and MADS-box families. In particular, we identified a novel FT-like gene, EtFT-1, which promotes flowering when overexpressed in Arabidopsis. These findings provide valuable insights into the molecular mechanisms of flowering in E. triticeum, and the identified homologs provide a useful resource for plant breeding to regulate flowering time.
Data availability statement
All the data in this study are included in this manuscript. The raw RNA-seq reads have been deposited in the NCBI database (accession PRJNA1198393).
Author contributions
LZ: Writing – review & editing, Methodology, Writing – original draft. WM: Data curation, Writing – review & editing. XZ: Writing – review & editing, Formal analysis. JP: Data curation, Writing – review & editing. XY: Data curation, Writing – review & editing. LS: Conceptualization, Supervision, Writing – review & editing, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Third Xinjiang Scientific Expedition Program (grant no. 2022xjkk150301).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1576519/full#supplementary-material
Supplementary Figure 1 | The NR classification description. (A, B) and (C) represent the e-value distribution, similarity distribution and species distribution of NR annotations, respectively.
Supplementary Figure 2 | Phylogenetic analysis of COL (A), FRL (B) and SPL (C) proteins in E. triticeum and other plant species.
Supplementary Figure 3 | Phylogenetic relationship of MADS-box genes in E. triticeum (Et), Triticum aestivum (Traes), and Arabidopsis thaliana (At).
Supplementary Table 1 | Summary for the transcriptome assembly.
Supplementary Table 2 | Statistics of the transcriptome database of E. triticeum.
Supplementary Table 3 | Summary statistics of functional annotation of the unigenes of E. triticeum in public databases.
Supplementary Table 4 | GO enrichment analysis for differentially expressed transcripts with corrected P-valuess ≤ 0.05.
Supplementary Table 5 | KEGG pathway enrichment results with upregulated DEGs from the VS_vs_HS (A), HS_vs_FS (B), and VS_vs_FS (C) comparison.
Supplementary Table 6 | Differentially expressed genes and the related flowering pathway in E. triticeum. (A) All candidate differentially expressed genes (DEGs) related to flowering in E. triticeum. (B) Differentially expressed genes associated with the flowering pathway in E. triticeum.
Supplementary Table 7 | The differentially expressed genes (DEGs) involved in hormone signal transduction pathway in E. triticeum.
Supplementary Table 8 | Flowering phenotypic analysis of wild-type (WT) and EtFT-1 overexpressing transgenic Arabidopsis thaliana plants.
Supplementary Table 9 | Primers used in this study.
References
Abdul Kareem, A. J., Saeed, M. (2020). Effect of ascorbic and salicylic acids on growth and flowering of Gazania cv. Frosty Kiss Mixed. Ornam. Hortic. 26, 537–544. doi: 10.1590/2447-536x.v26i4.2118
Ahn, J. H., Miller, D., Winter, V. J., Banfield, M. J., Lee, J. H., Yoo, S. Y., et al. (2006). A divergent external loop confers antagonistic activity on floral regulators FT and TFL1. EMBO. J. 25, 605–614. doi: 10.1038/sj.emboj.7600950
Andrés, F., Coupland, G. (2012). The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 13, 627–639. doi: 10.1038/nrg3291
Aslam, M., Jakada, B. H., Fakher, B., Greaves, J. G., Niu, X., Su, Z., et al. (2020). Genome-wide study of pineapple (Ananas comosus L.) bHLH transcription factors indicates that cryptochrome-interacting bHLH2 (AcCIB2) participates in flowering time regulation and abiotic stress response. BMC Genom. 21, 735. doi: 10.1186/s12864-020-07152-2
Bernal, M., Estiarte, M., Peñuelas, J. (2011). Drought advances spring growth phenology of the Mediterranean shrub Erica multiflora. Plant Biol. (Stuttg). 13, 252–257. doi: 10.1111/j.1438-8677.2010.00358.x
Blázquez, M. A., Soowal., L. N., Lee, I., Weigel, D. (1997). LEAFY expression and flower initiation in Arabidopsis. Development. 124, 3835–3844. doi: 10.1242/dev.124.19.3835
Blümel, M., Dally, N., Jung, C. (2015). Flowering time regulation in crops-what did we learn from Arabidopsis? Curr. Opin. Biotechnol. 32, 121–129. doi: 10.1016/j.copbio.2014.11.023
Buchfink, B., Xie, C., Huson, D. H. (2015). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60. doi: 10.1038/nmeth.3176
Campos-Rivero, G., Osorio-Montalvo, P., Sánchez-Borges, R., Us-Camas, R., Duarte-Aké, F., De-la-Peña, C. (2017). Plant hormone signaling in flowering: An epigenetic point of view. J. Plant Physiol. 214, 16–27. doi: 10.1016/j.jplph.2017.03.018
Castelán-Muñoz, N., Herrera, J., Cajero-Sánchez, W., Arrizubieta, M., Trejo, C., García-Ponce, B., et al. (2019). MADS-box genes are key components of genetic regulatory networks involved in abiotic stress and plastic developmental responses in plants. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00853
Chao, L. M., Liu, Y. Q., Chen, D. Y., Xue, X. Y., Mao, Y. B., Chen, X. Y. (2017). Arabidopsis transcription factors SPL1 and SPL12 confer plant thermotolerance at reproductive stage. Mol. Plant 10, 735–748. doi: 10.1016/j.molp.2017.03.010
Chopy, M., Binaghi, M., Cannarozzi, G., Halitschke, R., Boachon, B., Heutink, R., et al. (2023). A single MYB transcription factor with multiple functions during flower development. New Phytol. 239, 2007–2025. doi: 10.1111/nph.19096
Chuck, G., Muszynski, M., Kellogg, E., Hake, S., Schmidt, R. J. (2002). The control of spikelet meristem identity by the branched silkless1 gene in maize. Science. 298, 1238–1241. doi: 10.1126/science.1076920
Clough, S. J., Bent, A. F. (1998). Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743. doi: 10.1046/j.1365-313x.1998.00343.x
Davis, S. J. (2009). Integrating hormones into the floral-transition pathway of Arabidopsis thaliana. Plant Cell Environ. 32, 1201–1210. doi: 10.1111/j.1365-3040.2009.01968.x
Diezel, C., Allmann, S., Baldwin, I. T. (2011). Mechanisms of optimal defense patterns in Nicotiana attenuata: flowering attenuates herbivory-elicited ethylene and jasmonate signaling. J. Integr. Plant Biol. 53, 971–983. doi: 10.1111/j.1744-7909.2011.01086.x
Di Marzo, M., Roig-Villanova, I., Zanchetti, E., Caselli, F., Gregis, V., Bardetti, P., et al. (2020). MADS-box and bHLH transcription factors coordinate transmitting tract development in Arabidopsis thaliana. Front. Plant Sci. 11. doi: 10.3389/fpls.2020.00526
Eddy, S. R. (1998). Profile hidden Markov models. Bioinformatics. 14, 755–763. doi: 10.1093/bioinformatics/14.9.755
Fan, M., Li, X., Zhang, Y., Wu, S., Song, Z., Yin, H., et al. (2022). Floral organ transcriptome in Camellia sasanqua provided insight into stamen petaloid. BMC Plant Biol. 22, 474. doi: 10.1186/s12870-022-03860-x
Fornara, F., de Montaigu, A., Coupland, G. (2010). SnapShot: control of flowering in arabidopsis. Cell. 141, 550–550. doi: 10.1016/j.cell.2010.04.024
Franks, S. J., Sim, S., Weis, A. E. (2007). Rapid evolution of flowering time by an annual plant in response to a climate fluctuation. Proc Natl Acad Sci USA. 104, 1278–1282. doi: 10.1073/pnas.0608379104
Frederiksen, S., Bothmer, R. (1995). Intergeneric hybridizations with eremopyrum (Poaceae). Nord. J. Bot. 15, 39–47. doi: 10.1111/j.1756-1051.1995.tb00119.x
Fu, L., Tan, D., Sun, X., Ding, Z., Zhang, J. (2020). Transcriptional analysis reveals potential genes and regulatory networks involved in salicylic acid-induced flowering in duckweed (Lemna gibba). Plant Physiol. Biochem. 155, 512–522. doi: 10.1016/j.plaphy.2020.08.001
Gao, X., Liang, W., Yin, C., Ji, S., Wang, H., Su, X., et al. (2010). The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol. 153, 728–740. doi: 10.1104/pp.110.156711
Grabherr, M. G., Haas, B. J., Yassour, M., Levin, J. Z., Thompson, D. A., Amit, I., et al. (2011). Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644–652. doi: 10.1038/nbt.1883
Hanzawa, Y., Money, T., Bradley, D. (2005). A single amino acid converts a repressor to an activator of flowering. Proc. Natl. Acad. Sci. USA. 102, 7748–7753. doi: 10.1073/pnas.0500932102
Harig, L., Beinecke, F. A., Oltmanns, J., Muth, J., Müller, O., Rüping, B., et al. (2012). Proteins from the FLOWERING LOCUS T-like subclade of the PEBP family act antagonistically to regulate floral initiation in tobacco. Plant J. 72, 908–921. doi: 10.1111/j.1365-313X.2012.05125.x
Helliwell, C. A., Wood, C. C., Robertson, M., James Peacock, W., Dennis, E. S. (2006). The Arabidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is part of a high-molecular-weight protein complex. Plant J. 46, 183–192. doi: 10.1111/j.1365-313X.2006.02686.x
Imaizumi, T. (2010). Arabidopsis circadian clock and photoperiodism: time to think about location. Curr. Opin. Plant Biol. 13, 83–89. doi: 10.1016/j.pbi.2009.09.007
Izawa, T., Oikawa, T., Sugiyama, N., Tanisaka, T., Yano, M., Shimamoto, K. (2002). Phytochrome mediates the external light signal to repress FT orthologs in photoperiodic flowering of rice. Genes Dev. 16, 2006–2020. doi: 10.1101/gad.999202
Jones, P., Binns, D., Chang, H. Y., Fraser, M., Li, W., McAnulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics. 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Kanehisa, M., Goto, S., Kawashima, S., Okuno, Y., Hattori, M. (2004). The KEGG resource for deciphering the genome. Nucleic Acids Res. 32, 277–280. doi: 10.1093/nar/gkh063
Kardailsky, I., Shukla, V. K., Ahn, J. H., Dagenais, N., Christensen, S. K., Nguyen, J. T., et al. (1999). Activation tagging of the floral inducer FT. Science. 286, 1962–1965. doi: 10.1126/science.286.5446.1962
Kazan, K., Lyons, R. (2016). The link between flowering time and stress tolerance. J. Exp. Bot. 67, 47–60. doi: 10.1093/jxb/erv441
Kim, S. G., Kim, S. Y., Park, C. M. (2007). A membrane-associated NAC transcription factor regulates salt-responsive flowering via FLOWERING LOCUS T in Arabidopsis. Planta. 226, 647–654. doi: 10.1007/s00425-007-0513-3
Kinoshita, A., Richter, R. (2020). Genetic and molecular basis of floral induction in Arabidopsis thaliana. J. Exp. Bot. 71, 2490–2504. doi: 10.1093/jxb/eraa057
Kippes, N., Debernardi, J. M., Vasquez-Gross, H. A., Akpinar, B. A., Budak, H., Kato, K., et al. (2015). Identification of the VERNALIZATION 4 gene reveals the origin of spring growth habit in ancient wheats from South Asia. Proc. Natl. Acad. Sci. USA. 112, 5401–5410. doi: 10.1073/pnas.1514883112
Kobayashi, Y., Kaya, H., Goto, K., Iwabuchi, M., Araki, T. (1999). A pair of related genes with antagonistic roles in mediating flowering signals. Science. 286, 1960–1962. doi: 10.1126/science.286.5446.1960
Kojima, S., Takahashi, Y., Kobayashi, Y., Monna, L., Sasaki, T., Araki, T., et al. (2002). Hd3a, a rice ortholog of the Arabidopsis FT gene, promotes transition to flowering downstream of Hd1 under short-day conditions. Plant Cell Physiol. 43, 1096–1105. doi: 10.1093/pcp/pcf156
Komatsu, M., Chujo, A., Nagato, Y., Shimamoto, K., Kyozuka, J. (2003). FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development. 130, 3841–3850. doi: 10.1242/dev.00564
Komiya, R., Yokoi, S., Shimamoto, K. (2009). A gene network for long-day flowering activates RFT1 encoding a mobile flowering signal in rice. Development. 136, 3443–3450. doi: 10.1242/dev.040170
Kurokura, T., Mimida, N., Battey, N. H., Hytönen, T. (2013). The regulation of seasonal flowering in the Rosaceae. J. Exp. Bot. 64, 4131–4141. doi: 10.1093/jxb/ert233
Lee, Z., Kim, S., Choi, S. J., Joung, E., Kwon, M., Park, H. J., et al. (2023). Regulation of flowering time by environmental factors in plants. Plants. 12, 3680. doi: 10.3390/plants12213680
Lee, J., Lee, I. (2010). Regulation and function of SOC1, a flowering pathway integrator. J. Exp. Bot. 61, 2247–2254. doi: 10.1093/jxb/erq098
Li, J., Han, G., Sun, C., Sui, N. (2019). Research advances of MYB transcription factors in plant stress resistance and breeding. Plant Signal Behav. 14, 1613131. doi: 10.1080/15592324.2019.1613131
Liang, C., Liu, L., Zhang, Z., Ze, S., Pei, L., Feng, L., et al. (2023). Transcriptome analysis of critical genes related to flowering in Mikania micrantha at different altitudes provides insights for a potential control. BMC Genom. 24, 14. doi: 10.1186/s12864-023-09108-8
Liang, Y. C., Reid, M. S., Jiang, C. Z. (2014). Controlling plant architecture by manipulation of gibberellic acid signalling in petunia. Hortic. Res. 1, 14061. doi: 10.1038/hortres.2014.61
Liu, K., Feng, S., Pan, Y., Zhong, J., Chen, Y., Yuan, C., et al. (2016). Transcriptome analysis and identification of genes associated with floral transition and flower development in sugar apple (Annona squamosa L.). Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01695
Liu, L., Xuan, L., Jiang, Y., Yu, H. (2021). Regulation by FLOWERING LOCUS T and TERMINAL FLOWER 1 in flowering time and plant architecture. Small Struct. 2, 2000125. doi: 10.1002/sstr.202000125
Livak, K. J., Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 25, 402–408. doi: 10.1006/meth.2001.1262
Love, M. I., Huber, W., Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15, 550. doi: 10.1186/s13059-014-0550-8
Luo, Y., Liu, M., Cao, J., Cao, F., Zhang, L. (2022). The role of salicylic acid in plant flower development. For Res. (Fayettev). 2, 14. doi: 10.48130/FR-2022-0014
Ma, X., Qiao, Z., Chen, D., Yang, W., Zhou, R., Zhang, W., et al. (2015). CYCLIN-DEPENDENT KINASE G2 regulates salinity stress response and salt mediated flowering in Arabidopsis thaliana. Plant Mol. Biol. 8, 287–299. doi: 10.1007/s11103-015-0324-z
Mao, Z. M., Zhang, D. M. (1994). The conspectus of ephemeral flora in northern Xinjiang. Arid Zone Res. 11, 1–26. doi: 10.13866/j.azr.1994.03.001
Martínez, C., Pons, E., Prats, G., León, J. (2004). Salicylic acid regulates flowering time and links defence responses and reproductive development. Plant J. 37, 209–217. doi: 10.1046/j.1365-313x.2003.01954.x
Martín-Mex, R., Nexticapan-Garcéz, Á., Villanueva-Couoh, E., Uicab-Quijano, V., Vergara-Yoisura, S., Larqué-Saavedra, A. F. (2015). Salicylic acid stimulates flowering in micropopagated gloxinia plants. Rev. Fitotec. Mex. 38, 115–118. doi: 10.35196/rfm.2015.2.115
McGarry, R. C., Prewitt, S., Ayre, B. G. (2013). Overexpression of FT in cotton affects architecture but not floral organogenesis. Plant Signal Behav. 8, e23602. doi: 10.4161/psb.23602
Michaels, S. D., Amasino, R. M. (1999). FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell. 11, 949–956. doi: 10.1105/tpc.11.5.949
Michaels, S. D., Amasino, R. M. (2001). Loss of FLOWERING LOCUS C activity eliminates the late-flowering phenotype of FRIGIDA and autonomous pathway mutations but not responsiveness to vernalization. Plant Cell. 13, 935–941. doi: 10.1105/tpc.13.4.935
Milec, Z., Valarik, M., Bartos, J., Safar, J. (2014). Can a late bloomer become an early bird? Tools for flowering time adjustment. Biotechnol. Adv. 2, 200–214. doi: 10.1016/j.bioteChadv.2013.09.008
Moon, J., Lee, H., Kim, M., Lee, I. (2005). Analysis of flowering pathway integrators in Arabidopsis. Plant Cell Physiol. 46, 292–299. doi: 10.1093/pcp/pci024
Mutasa-Göttgens, E., Hedden, P. (2009). Gibberellin as a factor in floral regulatory networks. J. Exp. Bot. 60, 1979–1989. doi: 10.1093/jxb/erp040
Nelson, D. C., Lasswell, J., Rogg, L. E., Cohen, M. A., Bartel, B. (2000). FKF1, a clock-controlled gene that regulates the transition to flowering in Arabidopsis. Cell. 101, 331–340. doi: 10.1016/s0092-8674(00)80842-9
Ohmori, S., Kimizu, M., Sugita, M., Miyao, A., Hirochika, H., Uchida, E., et al. (2009). MOSAIC FLORAL ORGANS1, an AGL6-like MADS box gene, regulates floral organ identity and meristem fate in rice. Plant Cell. 21, 3008–3025. doi: 10.1105/tpc.109.068742
Parcy, F. (2005). Flowering: a time for integration. Int. J. Dev. Biol. 49, 585–593. doi: 10.1387/ijdb.041930fp
Qian, Y., Zhang, T., Yu, Y., Gou, L., Yang, J., Xu, J., et al. (2021). Regulatory mechanisms of bHLH transcription factors in plant adaptive responses to various abiotic stresses. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.677611
Qiu, J., Tan, D. Y., Fan, D. Y. (2007). Characteristics of photosynthesis and biomass allocation of spring ephemerals in the Junggar Desert. Chin. J. Plant Ecol. 31, 883–891. doi: 10.17521/cjpe.2007.0111
Ryu, J. Y., Lee, H. J., Seo, P. J., Jung, J. H., Ahn, J. H., Park, C. M. (2014). The Arabidopsis floral repressor BFT delays flowering by competing with FT for FD binding under high salinity. Mol. Plant 7, 377–387. doi: 10.1093/mp/sst114
Sawa, M., Nusinow, D. A., Kay, S. A., Imaizumi, T. (2007). FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science. 318, 261–265. doi: 10.1126/science.1146994
Sherrard, M. E., Maherali, H. (2006). The adaptive significance of drought escape in Avena barbata, an annual grass. Evolution. 60, 2478–2489. doi: 10.1111/j.0014-3820.2006.tb01883.x
Shimakawa, A., Shiraya, T., Ishizuka, Y., Wada, K. C., Mitsui, T., Takeno, K. (2012). Salicylic acid is involved in the regulation of starvation stress-induced flowering in Lemna paucicostata. J. Plant Physiol. 169, 987–991. doi: 10.1016/j.jplph.2012.02.009
Song, H., Duan, Z., Zhang, J. (2024). WRKY transcription factors modulate flowering time and response to environmental changes. Plant Physiol. Biochem. 210, 108630. doi: 10.1016/j.plaphy.2024.108630
Srikanth, A., Schmid, M. (2011). Regulation of flowering time: all roads lead to Rome. Cell Mol. Life Sci. 68, 2013–2037. doi: 10.1007/s00018-011-0673-y
Suárez-López, P., Wheatley, K., Robson, F., Onouchi, H., Valverde, F., Coupland, G. (2001). CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature. 410, 1116–1120. doi: 10.1038/35074138
Sun, H., Jia, Z., Cao, D., Jiang, B., Wu, C., Hou, W., et al. (2011). GmFT2a, a soybean homolog of FLOWERING LOCUS T, is involved in flowering transition and maintenance. PloS One 6, e29238. doi: 10.1371/journal.pone.0029238
Takeno, K. (2016). Stress-induced flowering: the third category of flowering response. J. Exp. Bot. 67, 4925–4934. doi: 10.1093/jxb/erw272
Trapnell, C., Williams, B. A., Pertea, G., Mortazavi, A., Kwan, G., van Baren, M. J., et al. (2010). Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28, 511–515. doi: 10.1038/nbt.1621
Wada, K. C., Takeno, K. (2010). Stress-induced flowering. Plant Signal Behav. 5, 944–947. doi: 10.4161/psb.5.8.11826
Wang, Y. (1993). Phenological observation of the early spring ephemeral and ephemeroid plant in Xinjiang. Arid Zone Res. 10, 34–39. doi: 10.13866/j.azr.1993.03.007
Wang, X. Q., Jiang, J., Lei, J. Q., Zhang, W. M., Qian, Y. B. (2003). Distribution of ephemeral plants and their significance in dune stabilization in Gurbantunggut Desert. J. Geogr. Sci. 13, 323–330. doi: 10.1007/BF02837507
Wang, L. X., Shao, S., Lou, Y. H. (2024). The effect of heterochronic germination on the growth and quality of four forage species in the Eremopyrum. Acta Agrestia Sin. 32, 495–502. doi: 10.11733/j.issn.1007-0435.2024.02.017
Wang, J., Tian, L., Lee, H. S., Chen, Z. J. (2006). Nonadditive regulation of FRI and FLC loci mediates flowering-time variation in Arabidopsis allopolyploids. Genetics. 173, 965–974. doi: 10.1534/genetics.106.056580
Wang, Z., Zhou, Z., Liu, Y., Liu, T., Li, Q., Ji, Y., et al. (2015). Functional evolution of phosphatidylethanolamine binding proteins in soybean and Arabidopsis. Plant Cell. 27, 323–336. doi: 10.1105/tpc.114.135103
Waseem, M., Li, N., Su, D., Chen, J., Li, Z. (2019). Overexpression of a basic helix-loop-helix transcription factor gene, SlbHLH22, promotes early flowering and accelerates fruit ripening in tomato (Solanum lycopersicum L.). Planta. 250, 173–185. doi: 10.1007/s00425-019-03157-8
Wong, A. C., Hecht, V. F., Picard, K., Diwadkar, P., Laurie, R. E., Wen, J., et al. (2013). Isolation and functional analysis of CONSTANS-LIKE genes suggests that a central role for CONSTANS in flowering time control is not evolutionarily conserved in Medicago truncatula. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00486
Xie, C., Mao, X., Huang, J., Ding, Y., Wu, J., Dong, S., et al. (2011). KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 39, W316–W322. doi: 10.1093/nar/gkr483
Xing, S., Salinas, M., Höhmann, S., Berndtgen, R., Huijser, P. (2010). miR156-targeted and nontargeted SBP-box transcription factors act in concert to secure male fertility in Arabidopsis. Plant Cell. 22, 3935–3950. doi: 10.1105/tpc.110.079343
Xu, S., Chong, K. (2018). Remembering winter through vernalisation. Nat. Plants. 4, 997–1009. doi: 10.1038/s41477-018-0301-z
Xu, X., Xu, J., Yuan, C., Hu, Y., Liu, Q., Chen, Q., et al. (2021). Characterization of genes associated with TGA7 during the floral transition. BMC Plant Biol. 21, 367. doi: 10.1186/s12870-021-03144-w
Yamagishi, N., Sasaki, S., Yamagata, K., Komori, S., Nagase, M., Wada, M., et al. (2011). Promotion of flowering and reduction of a generation time in apple seedlings by ectopical expression of the Arabidopsis thaliana FT gene using the apple latent spherical virus vector. Plant Mol. Biol. 75, 193–204. doi: 10.1007/s11103-010-9718-0
Yan, L., Fu, D., Li, C., Blechl, A., Tranquilli, G., Bonafede, M., et al. (2006). The wheat and barley vernalization gene VRN3 is an orthologue of FT. Proc. Natl. Acad. Sci. USA. 103, 19581–19586. doi: 10.1073/pnas.0607142103
Yan, L., Loukoianov, A., Blechl, A., Tranquilli, G., Ramakrishna, W., SanMiguel, P., et al. (2004). The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science. 303, 1640–1644. doi: 10.1126/science.1094305
Yan, L., Loukoianov, A., Tranquilli, G., Helguera, M., Fahima, T., Dubcovsky, J. (2003). Positional cloning of the wheat vernalization gene VRN1. Proc. Natl. Acad. Sci. USA. 100, 6263–6268. doi: 10.1073/pnas.0937399100
Yang, M., Lin, W., Xu, Y., Xie, B., Yu, B., Chen, L., et al. (2024). Flowering-time regulation by the circadian clock: from Arabidopsis to crops. Crop J. 12, 17–27. doi: 10.1016/j.cj.2023.09.002
Zhang, G. F. (2014). Research on Eremopyrum wild plant resources and their important characteristics. Crops. 28, 10–13. doi: 10.16035/j.issn.1001-7283.2014.03.009
Zhang, L. Y., Chen, C. D. (2002). On the general characteristics of plant diversity of Gurbantunggut sandy desert. Acta Ecol. Sin. 22, 1923–1932. doi: 10.3321/j.issn:1000-0933.2002.11.018
Zhang, H., Harry, D. E., Ma, C., Yuceer, C., Hsu, C. Y., Vikram, V., et al. (2010). Precocious flowering in trees: the FLOWERING LOCUS T gene as a research and breeding tool in Populus. J. Exp. Bot. 61, 2549–2560. doi: 10.1093/jxb/erq092
Zhang, L., Liu, G., Jia, J., Zhao, G., Xia, C., Zhang, L., et al. (2016). The wheat MYB-related transcription factor TaMYB72 promotes flowering in rice. J. Integr. Plant Biol. 58, 701–704. doi: 10.1111/jipb.12461
Zhang, Y., Tang, D., Lin, X., Ding, M., Tong, Z. (2018). Genome-wide identification of MADS-box family genes in moso bamboo (Phyllostachys edulis) and a functional analysis of PeMADS5 in flowering. BMC Plant Biol. 18, 176. doi: 10.1186/s12870-018-1394-2
Zhang, Y., Zhang, B., Yang, T., Zhang, J., Liu, B., Zhan, X., et al. (2020). The GAMYB-like gene SlMYB33 mediates flowering and pollen development in tomato. Hortic. Res. 7, 133. doi: 10.1038/s41438-020-00366-1
Zhao, C., Takeshima, R., Zhu, J., Xu, M., Sato, M., Watanabe, S., et al. (2016). A recessive allele for delayed flowering at the soybean maturity locus E9 is a leaky allele of FT2a, a FLOWERING LOCUS T ortholog. BMC Plant Biol. 16, 20. doi: 10.1186/s12870-016-0704-9
Zheng, C., Ye, M., Sang, M., Wu, R. (2019). A Regulatory Network for miR156-SPL Module in Arabidopsis thaliana. Int. J. Mol. Sci. 20, 6166. doi: 10.3390/ijms20246166
Zhou, Y., Hu, L., Song, J., Jiang, L., Liu, S. (2019a). Isolation and characterization of a MADS-box gene in cucumber (Cucumis sativus L.) that affects flowering time and leaf morphology in transgenic Arabidopsis. Biotechnol. Biotechnol. Equip. 33, 54–63. doi: 10.1080/13102818.2018.1534556
Keywords: Eremopyrum triticeum, transcriptome, flowering pathway, differentially expressed genes, flowering locus T
Citation: Zeng L-L, Meng W-Q, Zhong X-N, Peng J-J, Yang X-L and Sun L (2025) Transcriptome analyses reveal the flowering regulatory networks in the desert ephemeral plant Eremopyrum triticeum. Front. Plant Sci. 16:1576519. doi: 10.3389/fpls.2025.1576519
Received: 14 February 2025; Accepted: 08 April 2025;
Published: 08 May 2025.
Edited by:
Martin Raspor, University of Belgrade, SerbiaReviewed by:
Nishanth M. J., St Joseph’s University, IndiaTianfu Han, Zhengzhou University, China
Maja Belic, University of Belgrade, Serbia
Copyright © 2025 Zeng, Meng, Zhong, Peng, Yang and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Li Sun, bGlzQHNoenUuZWR1LmNu
†These authors have contributed equally to this work