 Yang Liu
Yang Liu Qi ZhuRongkai LiRenyuan WeiXiaoyu GengXiang Zhang
Qi ZhuRongkai LiRenyuan WeiXiaoyu GengXiang Zhang Huanhe Wei
Huanhe Wei Pinglei GaoKe XuQigen Dai
Pinglei GaoKe XuQigen Dai Yinglong Chen*
Yinglong Chen*- Jiangsu Key Laboratory of Crop Cultivation and Physiology/Jiangsu Key Laboratory of Crop Genetics and Physiology/Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops/Key Laboratory of Saline-Alkali Soil Reclamation and Utilization in Coastal Areas, The Ministry of Agriculture and Rural Affairs of China/Research Institute of Rice Industrial Engineering Technology, Yangzhou University, Yangzhou, China
Introduction: How to increase crop yield in coastal saline-alkali land has become a focus and hot topic of concern for researchers.
Methods: Field experiments were conducted to identify whether foliar application of magnesium sulfate (MgSO4·7H2O) can enhance rice salt tolerance and improve rice yield. Treatments with four concentrations of MgSO4·7H2O (10 g L-1, 20 g L-1, 30 g L-1, and 40 g L-1) were applied during the jointing and heading stages of rice in three fields with different salt levels in Yancheng City, Jiangsu Province, China in 2022 and 2023.
Results: Results showed that the application of magnesium sulfate, even the lowest concentration of MgSO4, could significantly increase the rice yield and total biomass under all the three salt treatments, while the increase displayed more obvious under higher salt treatment. Magnesium sulfate treatment enhanced the Rubisco enzyme activity and total chlorophyll content in rice flag leaves, delayed leaf tip wilt, and thus improved the photosynthetic capacity of rice. Additionally, magnesium sulfate treatment significantly reduced the accumulation of toxic sodium ions (Na+) in rice compared to the untreated control, accompanied with notable enhancement of Mg/Na, K/Na, P/Na, and Ca/Na.
Discussion: This study found that magnesium sulfate could enhance the salt tolerance of rice in coastal saline-alkali soils, whereas the effects vary significantly among different concentrations. Under 20 g L-¹ of MgSO4 treatment, rice leaves exhibited the highest net photosynthetic rate and total chlorophyll content, while the incidence of leaf tip wilt and the accumulation of toxic sodium ions (Na+) were minimized, resulting in the highest yield and total biomass. Therefore, 20 g L-¹ of MgSO4 is likely to be recommended as the optimal application concentration in saline-alkali areas.
1 Introduction
Soil salinization is one of the significant abiotic stresses that constrain crop production and yield formation, gradually evolving into a global issue (Zhang et al., 2024; Pu et al., 2014). Approximately 800 million hectares of saline-alkaline land exist worldwide (Fahad et al., 2014). However, this figure continues to rise due to issues such as the use of contaminated water for irrigation and the accumulation of excessive salts resulting from improper irrigation and drainage practices. China is facing increasing scarcity of arable land due to the continuous advancement of urbanization and industrialization, with the per capita arable land area reaching only 40% of the world average (Zhang, 2015). China currently possesses approximately 2.34 million hectares of coastal saline-alkaline land and early 100 million hectares of inland saline-alkaline land, which are considered as potential reserve arable land resources (Li et al., 2022; Geng et al., 2024). Rice (Oryza sativa L.) serves as a pioneering crop for the improvement of saline-alkali lands, with its cultivation area on such lands expanding annually (Yan and He, 2010; Wang et al., 2022). It is also one of the most important staple crops globally, expected to be the primary food source for approximately 9 billion people by 2050. To meet this demand, rice production must increase by at least 60%.
Rice is more susceptible to rhizosphere salt stress than wheat, oilseed rape and other crops. Research indicates that while soil salinity exceeds 3‰, the number of effective panicles and biomass in rice are significantly reduced, ultimately leading to a yield decrease of over 30% (Eynard et al., 2005; Chen et al., 2022b). Excessive accumulation of salt ions affects the growth and development of rice by altering its osmotic homeostasis, ionic homeostasis, and oxidative stress. Under low to moderate salt concentrations, salt stress causes stomata closure and reduces the photosynthetic area in rice leaves. Under moderate to high salt concentrations, leaf tip necrosis is frequently occurred (Tester, 2003), mainly due to the excessive accumulation of toxic sodium (Na+) in plant tissues, which also disrupts ion balance, competitively inhibits the uptake of essential nutrients such as phosphorus (P), potassium (K), and calcium (Ca), and generates reactive oxygen species (ROS). Excessive reactive oxygen species (ROS) damage genetic material, leading to peroxidation of leaf cell membranes, which in turn produces excessive amounts of malondialdehyde (MDA), hydrogen peroxide (H2O2), and other peroxides, thereby disrupting the normal growth and development of rice (Tester, 2003; Wu et al., 2022).
Magnesium, as the second most abundant free divalent cation in plant growth processes (Mao et al., 2014), plays a crucial role in numerous vital physiological and biochemical processes such as chlorophyll synthesis and degradation, and the activation of various antioxidant system enzymes (Chen et al., 2018; Tian et al., 2021). Exogenous magnesium supplementation during the growth of plants such as sunflower (Rivelli et al., 2010) and citrus can effectively regulate chlorophyll content and alleviate the normal growth of plants under abiotic stresses like high light conditions. Research has shown that under salt stress, the uptake of magnesium by plants significantly decreases (Yildirim et al., 2008). The application of exogenous magnesium enhances the antioxidant system and modulates gene expression, thus promotes the accumulation of biomass in leaves and buds in castor oil plants (Brito Neto et al., 2012), chili peppers (Sevgin Zirek and Uzal, 2020), and peanut seedlings (Wang et al., 2024). Besides, exogenous magnesium significantly improves the salt tolerance of rice seedlings through the combined effects of enhancing secondary metabolic synthesis pathways and increasing antioxidant enzyme activity (Xuan et al., 2022). Given that previous studies primarily focused on small-scale ecological experiments such as growth chamber or pot trials, this experiment was designed to investigate the effects of foliar supplementation with magnesium sulfate under field conditions in coastal saline-alkali soils. The study aimed to clarify the effect of exogenous magnesium on alleviating salt stress in rice from the perspective of the whole growth period in coastal filed, meanwhile provide the proper concentration of foliar magnesium and clarify the responded mechanism.
2 Materials and methods
2.1 Materials and treatments
The study was conducted from 2022 to 2023 at the Jinhai Island Base in Sheyang County, Yancheng City, Jiangsu Province (33°57′N, 120°24′E), located less than 5 km from the Yellow Sea. The study area belongs to maritime humid climate with an annual average of approximately 2200 hours of sunshine and an annual rainfall of about 1000 millimeters. The soil type is sandy loam and the pH was 8.0 to 8.5, with 0.46 g kg−1 total nitrogen (N), 20.5 mg kg−1 olsen phosphorus (P), 85.6 mg kg−1 available potassium (K), 55.35 mg g−1 total sodium content, 3.04 mg g−1 total magnesium content, 24.41 mg g−1 total calcium content and 2.7 g kg−1organic carbon in the 0–20 cm soil layer. Rice cultivar Nanjing 9108 was adopted and sown on May 12. Rice seedlings with two fully expanded leaves were transplanted to field on June 5. The electrical conductivity of soil in trial field was measured before the experiment, based on which the field was subsequently categorized into three salt treatments, denoted as S1, S2 and S3, respectively, with the increasing of salt content (Chen et al., 2022a). The magnesium sulfate (MgSO4·7H2O) treatments contained four concentrations of 10 g L-1, 20 g L-1, 30 g L-1 and 40 g L-1, denoted as Mg1, Mg2, Mg3, and Mg4, respectively. The magnesium sulfate was foliar sprayed during the jointing (77 days after sowing in 2022 and 78 days in 2023) and heading stages (106 days after sowing in 2022 and 107 days in 2023) of rice with the volume of 380 L ha-1, while the control (CK) received an equivalent volume of pure water. Using a hand sprayer, the spray was evenly applied to the rice leaves from 5:00 a.m. to 6:00 a.m. on the day of the experiment, with each experimental plot covering an area of 50 m2 and replicated three times. Nitrogen, phosphorus, and potassium were supplied through urea (42% N), superphosphate (12% P2O5), and potassium chloride (60% K2O), respectively. The application rate of pure nitrogen was 262.5 kg ha-¹, with a nitrogen fertilizer ratio of 6:4 for basal and tillering fertilizer to panicle fertilizer. The application rate of pure phosphorus was 126 kg ha-¹, with a phosphorus fertilizer ratio of 5:5 for basal and tillering fertilizer to panicle fertilizer. The application rate of pure potassium was 360 kg ha-¹, with a potassium fertilizer ratio of 4:6 for basal and tillering fertilizer to panicle fertilizer. Other management practices and pest control were conducted according to conventional high-yield cultivation standards.
2.2 Sampling and measurement
2.2.1 Soil salinity dynamic monitoring
From 2022 to 2023, during the transplanting to maturity phase, soil conductivity in each plot was measured every 5 days using the Spectrum ECtestr-2265FS (Spectrum, USA).
2.2.2 Rice grain yield and its components
After the rice maturation, 100 complete plants were selected from each treatment for yield determination, with two replications. The number of panicles per experimental plot, the number of grains per panicle, and the thousand-grain weight were measured, and the filled-grain percentage was calculated.
Thousand-grain weight was calculated by randomly selecting 1000 plump seeds per replicate and averaging their weights.
Filled-grain percentage = (number of filled grains per panicle)/(total number of grains per panicle).
2.2.3 Dry matter weight determination
At the jointing (77 days after sowing in 2022 and 78 days in 2023), heading (106 days after sowing in 2022 and 107 days in 2023), and maturity stages (153 days after sowing in 2022 and 158 days in 2023), 5 holes of rice plants were sampled based on the average number of tillers per plot to determine the dry matter weight. The sampled plants were divided into leaves, stems, and panicles, then inactivation at 105°C for 30 minutes and dried at 80°C for 3 days until constant weight. Crop growth rate (g m-2 d-1) = (W2 - W1)/(t2 - t1), where W1 and W2 represent the dry matter weights of the population measured during two distinct periods, and t1 and t2 denote the time (days) corresponding to the two population dry matter weight measurements.
2.2.4 Photosynthetic parameters and SPAD values
At 20 days post-heading, the photosynthetic parameters including net photosynthetic rate, transpiration rate, stomatal conductance, and intercellular CO2 concentration of the flag leaves were measured using a portable photosynthesis system (LI-6400, LI-COR, Lincoln, NE, USA). The settings were based on Gu (Gu et al., 2017) until stable photosynthetic data were obtained, after which the leaves were removed from the chamber.
SPAD values were measured at the base, middle, and tip of the uppermost fully expanded leaf at the jointing, heading, and grain-filling stages using a SPAD-502 chlorophyll meter (MINOLTA, Japan), with six uniform rice plants selected per treatment.
2.2.5 Leaf physiological measurements
At 20 days post-heading, the flag leaves of each treatment were collected between 9:00-10:00 AM, with 15–20 leaves collected per treatment. After collection, the leaves were trimmed of their tips, tails, and veins, and immediately immersed in liquid nitrogen for freezing. Once fully frozen, the samples were transferred to a -80°C freezer for storage, to be used for subsequent analytical measurements.
2.2.6 Physiological parameter measurements
Leaf tip wilt index was determined of all the rice leaves at the jointing, heading, 15, 30, and 45 days after heading stage using the formula TWI = L1/L2, where L1 is the length of the wilted part of the all leaves and L2 is the total length of all leaves (Chen et al., 2021).
Superoxide dismutase (SOD) activity was measured using the NBT photochemical reduction method (Jin et al., 2008); peroxidase (POD) activity was measured using the guaiacol method (Jiang et al., 2021); catalase (CAT) activity was measured using UV spectrophotometry (Panda et al., 2017); ascorbate peroxidase (APX) activity was measured using UV spectrophotometry (Molina et al., 2002); malondialdehyde (MDA) was measured according to Di (Di et al., 2021); soluble sugar was measured according to Wang (Wang et al., 2014); soluble protein was measured according to Eva M et al (Campion et al., 2011); free proline content was measured according to Vieira (Vieira et al., 2010).
Leaf Rubisco enzyme activity, H+-ATPase enzyme activity, and hydrogen peroxide (H2O2) content were determined using reagent kits from Keming Biotechnology Co., Ltd. (Suzhou).
2.2.7 Element content in organs
At the jointing, heading, and maturity stages, three holes of plants were sampled, divided into stems, leaves, and panicles, inactivation at 105°C for 30 minutes, and dried at 80°C until constant weight. The dried samples were then ground and sieved. The contents of Na, Mg, K, P, and Ca in the samples were measured using an inductively coupled plasma mass spectrometer (AGILENT 7900 ICP-MS, Agilent Technologies, Japan), with three replicates per treatment.
2.3 Data statistics and analysis
Multivariate analyses of variance were carried out to determine the effects of different salinity concentrations and magnesium sulfate treatments, as well as their interactions on rice grain yield and its components, leaf tip wilt index, ion content, and distribution at a significance level of 5%. Additionally, pairwise comparisons were also performed on treatment means by using the LSD test at a significance level of 5%. The clustering is ward’s method and the clustering distance is Euclidean. All analyses were performed using IBM SPSS Statistics 26 (IBM, Armonk, NY, USA). Graphs appearing in articles such as polarized heat maps and Pearson correlation plots were prepared using Origin 2024b (Origin Lab, Hampton, MA, USA).
3 Results
3.1 Dynamic changes of soil electrical conductivity in different saline-alkaline soils
As shown in Figure 1, the analysis reveals that during the 2022–2023 trials, the trends in soil electrical conductivity (EC) after rice transplanting were consistent across the three different saline-alkaline fields. Over the two-year period, the average electrical conductivity in three fields was 2.54 mS cm-1 (S1), 3.48 mS cm-1 (S2) and 4.7 mS cm-1 (S3), respectively.
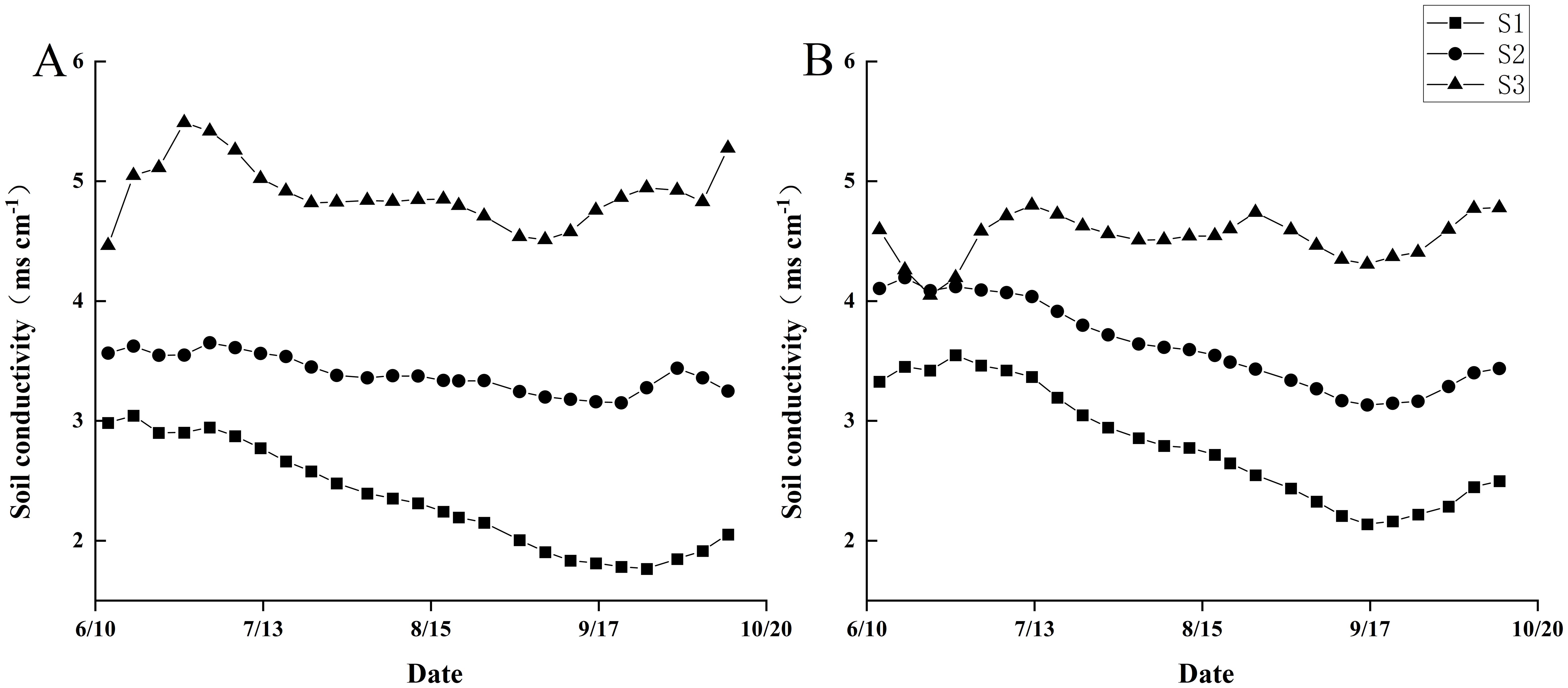
Figure 1. Dynamic changes of soil electrical conductivity in the three trial fields during rice growth season from 2022 to 2023. (A) shows the year 2022 and (B) shows the year 2023.
3.2 Rice grain yield and its components, dry matter weight and harvest index
As shown in Table 1, salinity significantly decreased rice yield and the decrease displayed more obvious with the increasing of salinity level. Compared to mild saline-alkali soil (S1), average yield reduction under severe saline-alkali soil (S3) reached 33%, with an average decline of 18.6% in filled-grain percentage and 7.6% in 1000-grain weight due to the elevated salinity. Overall, the decline in rice yield due to salinity was notably alleviated after foliar applying magnesium sulfate in all the three saline fields. The effects of Mg1 to Mg4 treatments from 2022 to 2023 exhibited a trend of first increasing and then decreasing. However, significant differences were observed in the effects of different magnesium sulfate concentrations. Compared to that of control, rice yield increased the most under spraying concentration of 20 g L-1 MgSO4 under all the three salt levels. The rice yield was increased averagely by 15%, 37.9% and 47.2% under S1, S2 and S3 salt concentration, respectively, across the two years. In terms of the number of panicles, significant difference occurred among different salt concentrations under the same magnesium sulfate concentration, where the number of panicles decreased with increased salt concentration and the most significant difference appeared under the S1 treatment. Under the same salt concentration, the Mg2 treatment was the most effective among different magnesium sulfate concentrations, followed by Mg1 and Mg3, while Mg4 was the least effective and showed no significant difference from the control (CK). At S1 salt concentration, the two-year average increase was 5.5% for Mg1 treatment, 4% for Mg2 treatment, 3.9% for Mg3 treatment, and 2.7% for Mg4 treatment. At S2 salt concentration, the average increase for the treatments was 18.6% (Mg1), 18.3% (Mg2), 17.7% (Mg3), and 12.4% (Mg4), whereas at S3 treatment, the average increase amounted to 14.4% for Mg1 treatment, 8.2% for Mg2 treatment, 13.3% for Mg3 treatment and 4% for Mg4 treatment. The effects of different magnesium sulfate concentrations varied in terms of panicle grain number over the two years, but overall, Mg2 and Mg3 treatments were the most effective, with Mg1 and Mg4 being less so. The Mg2 treatment significantly improved the filled-grain percentage by 10.2% (S1), 17.2% (S2), 24.2% (S3) in 2022, and 4.4% (S1), 11.1% (S2), 23.5% (S3) in 2023, respectively, compared to CK. Across the three salt concentrations, the average filled-grain percentage under Mg1, Mg2, Mg3 and Mg4 were 88%, 91.2%, 83.7% and 83.4%, in 2022, and 78.2%, 85.2%, 73.1% and 73.4%, respectively. The effect of magnesium sulfate treatment on the 1000-grain weight varied at different salt concentrations, with Mg1 being the most effective, increasing by up to 2.9% (2022 S1), followed by Mg2, Mg3, and Mg4. However, there was an overall upward trend. As shown in Table 2, the analysis reveals that across both years of the experiment, the dry matter weight and growth rate of the rice population at various stages significantly decreased with increasing salt concentration. Compared to the S1 and S2, the dry matter weight at maturity decreased more significantly under S3, with values of 10.6 t ha-1 and 9.6 t ha-1 in 2022 and 2023, respectively. Magnesium sulfate treatments at different concentrations significantly enhanced the dry matter weight of the rice population at all stages, with significant differences among treatments. Overall, the Mg2 treatment proved to be the most effective. Compared to the control treatment, the Mg2 treatment in 2022 increased the dry matter weight at maturity by 21.4% (S1), 48.4% (S2), and 41.2% (S3), and in 2023 by 12% (S1), 9.5% (S2), and 48.8% (S3). In the 2022–2023 experiments, the harvest index showed a trend of first decreasing and then increasing with rising salt concentrations. There was a significant interaction effect between magnesium sulfate treatments at different concentrations and different salt concentration treatments on the dry matter weight, crop growth rate, and harvest index of the rice population.
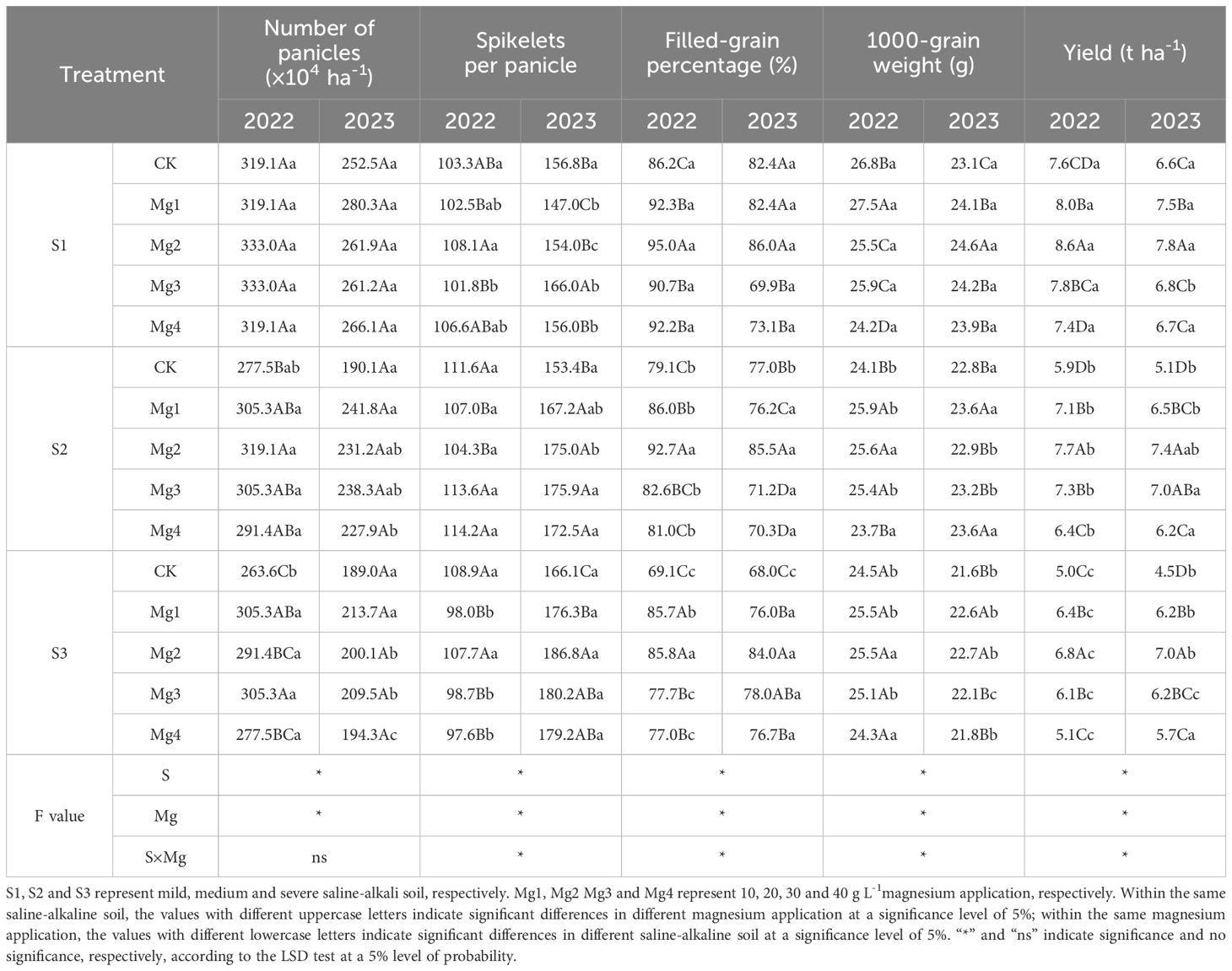
Table 1. Effects of magnesium application on rice yield and its components.

Table 2. Effects of magnesium application dry matter weight and harvest index of rice.
3.3 Effects of magnesium application on rice leaf tip wilt index
The dynamic changes in rice leaf tip necrosis under different concentrations of magnesium sulfate application are shown in Figure 2 for two years of experimentation. The trends of leaf tip necrosis in rice across both years were generally consistent. As the growth process advanced, the condition of leaf tip necrosis gradually worsened. The growth rate was slow before the heading stage, but it accelerated significantly from the heading stage to 45 days post-heading, with the largest increase occurring between 30- and 45- days post-heading. Analysis of the two-year experimental data showed that under different salt concentrations, magnesium sulfate at various concentrations significantly alleviated rice leaf tip necrosis. Compared to the control (CK), the reduction rate was the most significant under Mg2 treatment, which were 14.3% (S1), 17% (S2), and 19.9% (S3) in 2022, and 26.5% (S1), 14.5% (S2), and 10% (S3) in 2023, respectively. At the same salt concentration, there were significant differences among treatments with different concentrations of magnesium sulfate, generally showing a pattern of Mg2 > Mg1 > Mg3 > Mg4. The best treatment effect was observed with Mg2 in both years, with the maximum alleviation of 19.9% in 2022(S3) and 26.5% in 2023(S1) compared to the control.
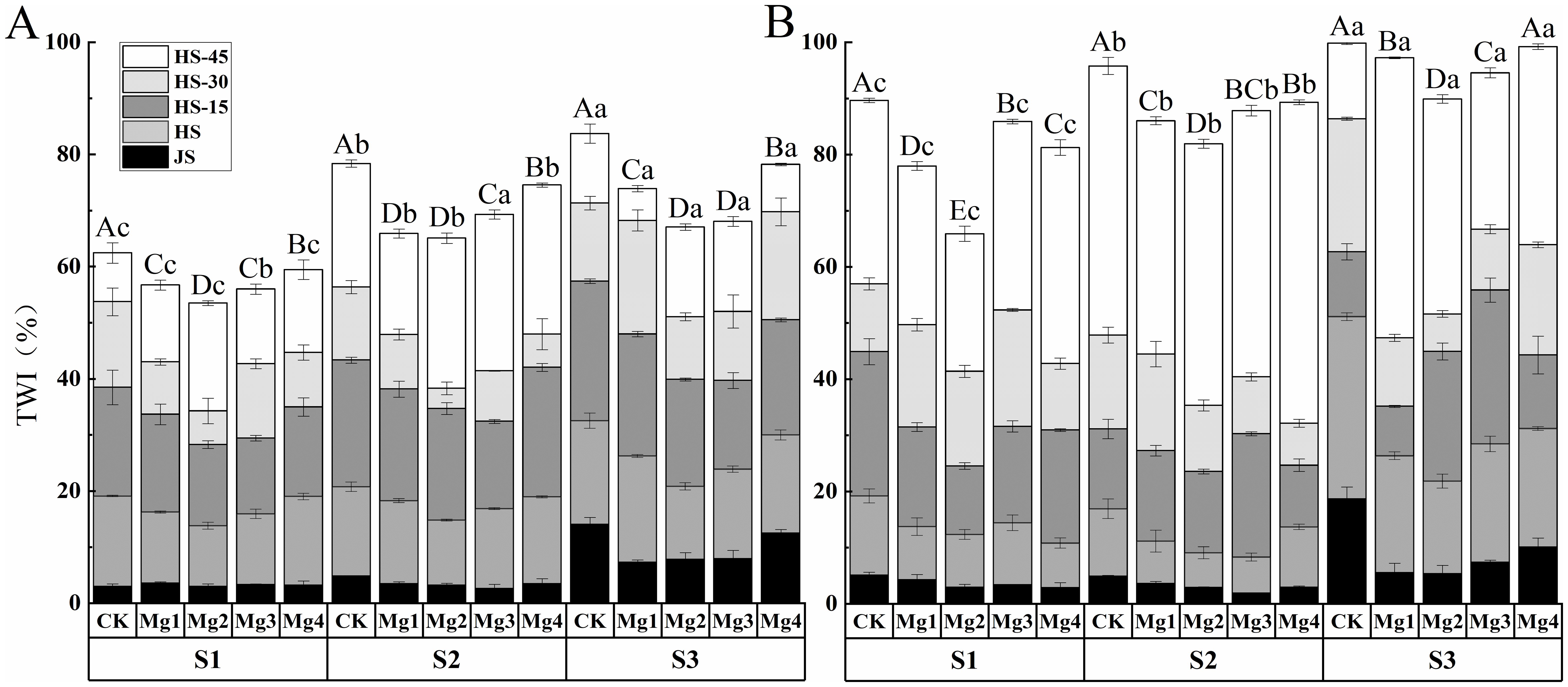
Figure 2. Effects of magnesium application on the rice leaf tip wilt index. (A) shows the year 2022 and (B) shows the year 2023. JS and HS represent jointing stage and heading stage, respectively. HS-15, HS-30 and HS-45 represent 15, 30 and 45 days, respectively, after the heading stage. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1 magnesium, respectively. Within the same saline-alkaline soil, the values with different uppercase letters indicate significant differences in different magnesium application at a significance level of 5%; Within the same magnesium application, the values with different lowercase letters indicate significant differences in different saline-alkaline soil at a significance level of 5%.
3.4 Effects of magnesium application on rice SPAD values
Figure 3 illustrates the trends in SPAD values of rice at various growth stages under different salt concentrations over two years. It is observed that the SPAD values of rice leaves generally followed a consistent pattern across the years, showing a gradual decline as the growth stages progressed. The SPAD values of the fully expanded last leaf tended to decrease over time. Under the same salt concentration, the peak SPAD values of rice leaves varied after treatments with different concentrations of magnesium sulfate. In 2022, except for the Mg2 treatment, which reached its maximum 15 days after heading, all other magnesium sulfate treatments showed peak SPAD values similar to the control at the heading stage. In 2023, both Mg2 and Mg3 treatments reached their maximum 15 days after heading, while the other treatments exhibited patterns consistent with those observed in 2022. Under different salt concentrations, the effects of magnesium sulfate application varied, with the overall performance ranking as Mg2 > Mg3 > Mg1 > Mg4.
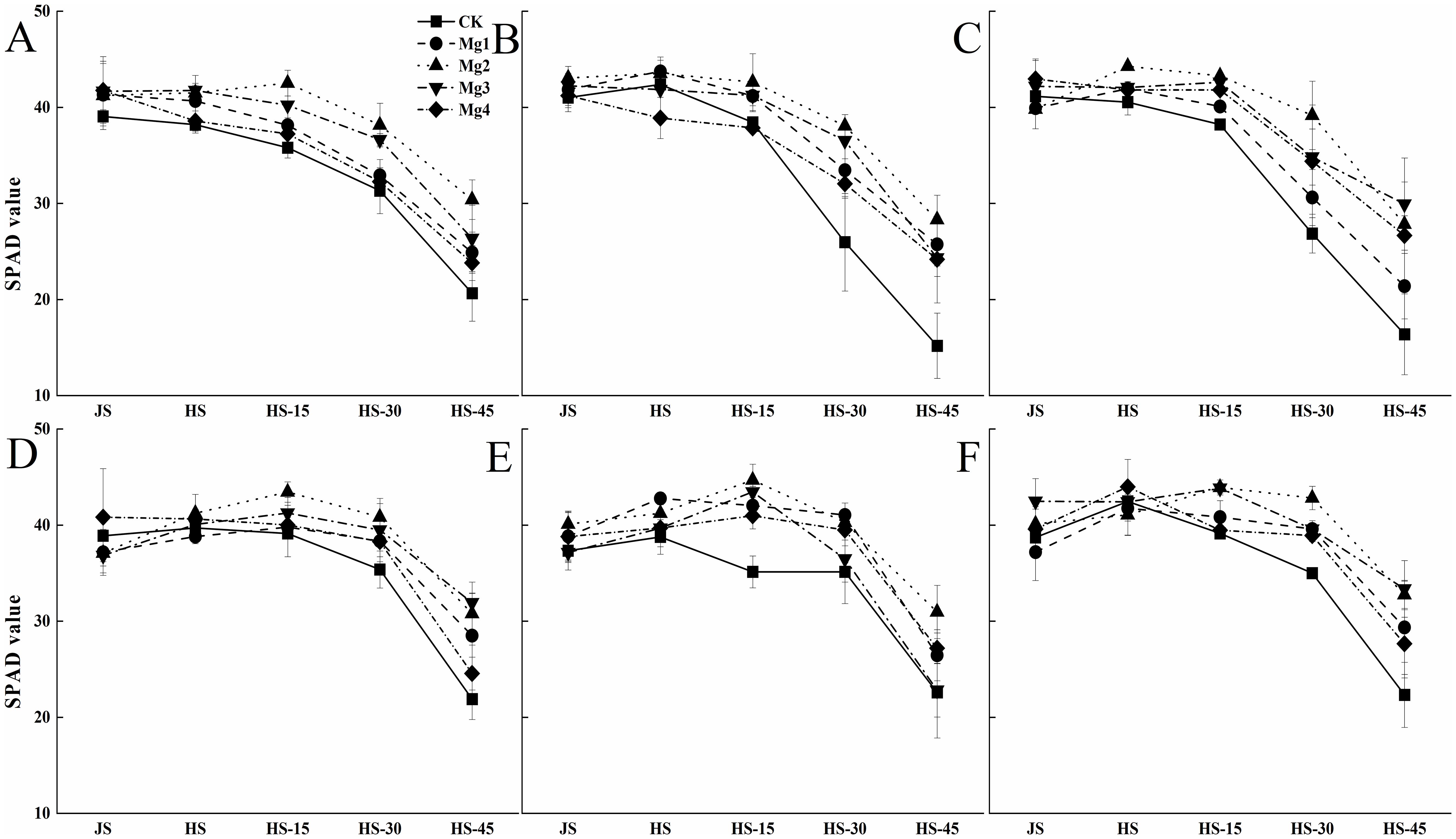
Figure 3. Effects of magnesium application on rice SPAD values. JS and HS represent jointing stage and heading stage, respectively. HS-15, HS-30 and HS-45 represent 15, 30 and 45 days, respectively, after the heading stage. (A–C) represent mild, medium and severe saline-alkali soil of 2022, (D–F) represent mild, medium and severe saline-alkali soil of 2023, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1 magnesium, respectively.
3.5 Effects of magnesium application on the pigment composition of rice flag leaf at 20 days after heading
As shown in Table 3, the application of magnesium sulfate at different concentrations significantly increased the contents of chlorophyll a, chlorophyll b, and carotenoids in the flag leaves of rice under various salt concentrations, with the enhancement effect ranking as Mg2 > Mg1 > Mg3 > Mg4 > CK. In 2022, the interaction between different salt concentrations and magnesium sulfate application significantly affected the contents of chlorophyll a, chlorophyll b, and total chlorophyll in the flag leaves of rice. In 2023, a significant interaction was observed only for the increase in chlorophyll a content. Over the two years of testing, there was no significant interaction between salt treatment and magnesium sulfate application on the increase in carotenoid content. In both 2022 and 2023, the M2 treatment yielded the best results, with the largest increase. Compared to the control, Mg2 treatment significantly enhanced chlorophyll content in rice flag leaves across two consecutive growing seasons. Chlorophyll a content increased by 58.2% (S1), 34% (S2), and 59.8% (S3) in 2022, with further improvements of 64.3% (S1), 42.8% (S2), and 31.2% (S3) in 2023. Chlorophyll b content exhibited increases of 84.9% (S1), 62.5% (S2), and 78.7% (S3) in 2022, followed by 46.6% (S1), 17.6% (S2), and 36.4% (S3) in 2023. Total chlorophyll (a + b) content showed improvements of 63.6% (S1), 39% (S2), and 63.2% (S3) in 2022, and 59.5% (S1), 35.5% (S2), and 32.6% (S3) in 2023, while carotenoid content was elevated by 70.4% (S1), 30.6% (S2), and 40.9% (S3) in 2022, and 34.5% (S1), 29.3% (S2), and 21.7% (S3) in 2023.
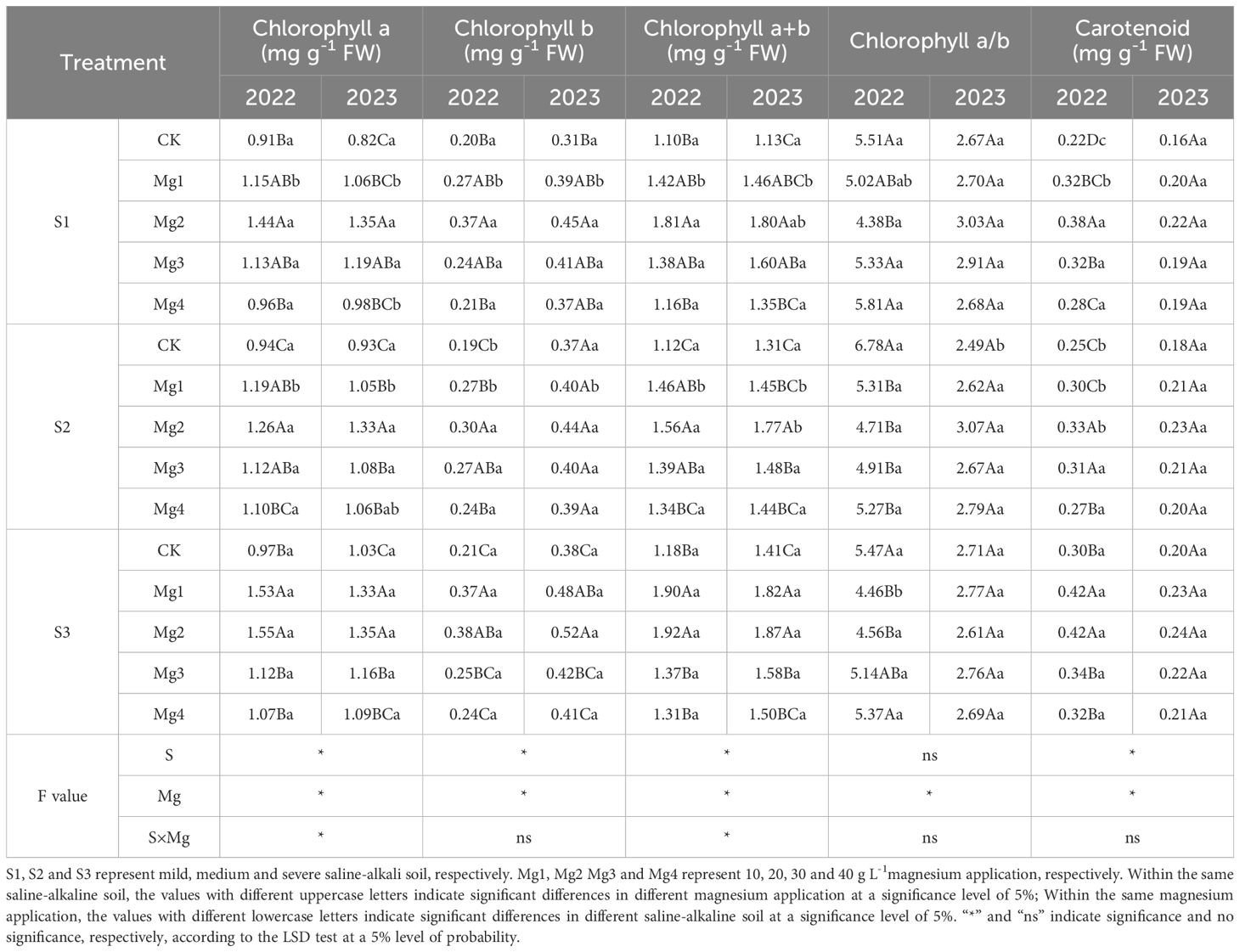
Table 3. Effects of magnesium application on the pigment composition of rice flag leaf at 20 days after heading.
3.6 Effects of magnesium application on photosynthetic characteristics of rice flag leaf in rice at 20 days after heading
The impact of foliar application of different concentrations of magnesium sulfate on photosynthetic characteristics of flag leaf in rice 20 days after heading is shown in Table 4. As indicated in Table 4, the application of magnesium sulfate at different salinity levels significantly affected the photosynthetic characteristics of rice flag leaves, with a highly significant interaction effect on net photosynthetic rate, stomatal conductance, and transpiration rate. Significant differences in photosynthetic characteristics were observed among different concentrations of magnesium sulfate.
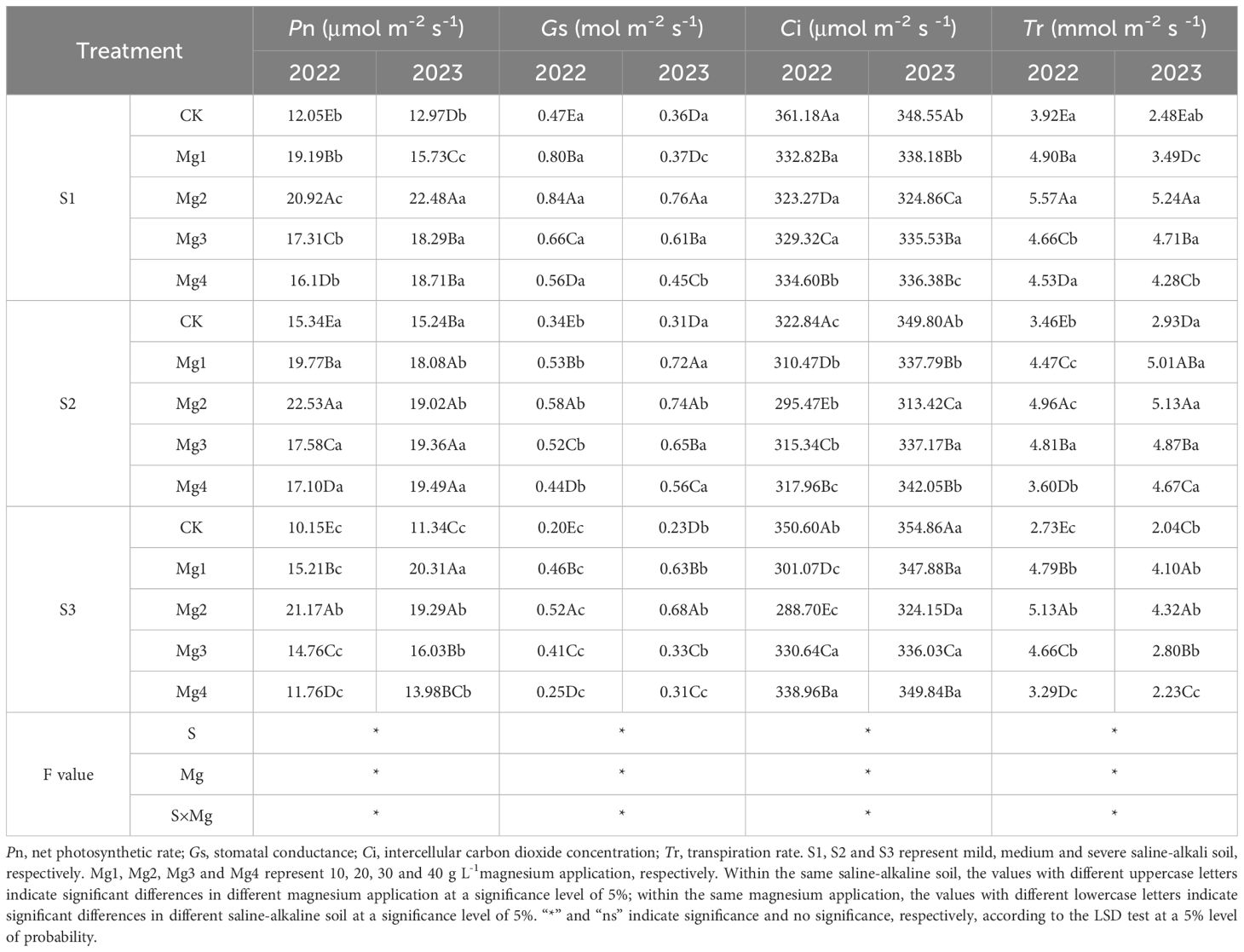
Table 4. Effects of magnesium application on photosynthetic characteristics of rice flag leaf at 20 days after heading.
In the two-year experimental data, compared with control, the application of magnesium sulfate significantly enhanced the net photosynthetic rate, stomatal conductance, and transpiration rate of rice leaves under different salinity levels, while significantly reducing the intercellular CO2 concentration. Comprehensive analysis of Mg treatment effects revealed that foliar magnesium sulfate supplementation significantly improved photosynthetic parameters in rice flag leaves compared to the control. In 2022, the net photosynthetic rate increased by an average of 52.5% (S1), 25.4% (S2), and 54.9% (S3), with further enhancements of 45% (S1), 24.6% (S2), and 53.5% (S3) in 2023. Stomatal conductance exhibited average increases of 53.2% (S1), 51.9% (S2), and 106.3% (S3) in 2022, followed by 53.7% (S1), 116% (S2), and 114.5% (S3) in 2023. Transpiration rate rose by 25.3% (S1), 29% (S2), and 63.5% (S3) in 2022, while in 2023, it showed more pronounced improvements of 78.5% (S1), 67.8% (S2), and 65.1% (S3). Conversely, intercellular CO2 concentration decreased by 8.6% (S1), 4% (S2), and 10.2% (S3) in 2022, with sustained reductions of 4.3% (S1), 4.9% (S2), and 4.3% (S3) in 2023.Significant differences in effects were observed among different concentrations of magnesium sulfate treatments, with the Mg2 treatment being the most effective. The overall performance over the two years was Mg2 > Mg1 > Mg3 > Mg4. The Mg2 treatment demonstrated the highest improvements in photosynthetic performance across both experimental years under all the four magnesium sulfate concentration treatments. Compared to those of control, the net photosynthetic rate of rice leaves increased by 73.6% and 73.3% (S1), 46.8% and 24.8% (S2), and 108.5% and 70.1% (S3) in 2022 and 2023, respectively. Stomatal conductance showed marked enhancements by 79.8% and 113.6% (S1), 70% and 141.6% (S2), and 164.1% and 201.4% (S3) in 2022 and 2023, respectively., Similarly, transpiration rate exhibited rises of 42.1% and 111.1% (S1), 43.4% and 75.1% (S2), and 87.8% and 112.3% (S3) in 2022 and 2023, respectively. In contrast, intercellular CO2 concentration displayed consistent reductions of 10.5% and 6.8% (S1), 8.5% and 10.4% (S2), and 17.7% and 8.7% (S3) in 2022 and 2023, respectively.
3.7 Effects of magnesium application on photosynthetic carbon assimilation and stomatal regulation in flag leaves of rice 20 days after heading
As shown in Figure 4, the application of magnesium sulfate at different concentrations significantly increased the activity of Rubisco in rice flag leaves 20 days after heading under different saline-alkaline soils in both years of the study. The effects varied significantly among different magnesium sulfate treatments, with the enhancement being Mg2 > Mg1 > Mg3 > Mg4 > CK. The Mg2 treatment yielded the best results, showing increases of 118.4% and 35.5% (S1), 160.8% and 68% (S2), 104.9% and 79.6% (S3) compared to the control in the two years, respectively. As shown in Figure 4, foliar magnesium sulfate application differentially modulated H+-ATPase activity and Rubisco carboxylation in rice flag leaves, with Mg2 treatment exhibiting the most pronounced effects. Under high salinity (S3), Mg2 elevated H+-ATPase activity by 119.1% (2022) and 87.3% (2023), which mechanistically linked to enhanced stomatal conductance (Gs improved on average by 177.8%) and Na+/K+ homeostasis. This proton-pumping activity energized guard cell turgor, facilitating CO2 influx and synergistically amplifying Rubisco activity and net photosynthetic rate. Similar trends were observed in S1 as elevated H+-ATPase activity by 143.6% (2022) and 29.4% (2023) also and S2 as 156% (2022) and 74.4% (2023). Though low-salinity conditions (S1) showed attenuated physiological gains due to redundant Mg²+ uptake and lower oxidative stress.
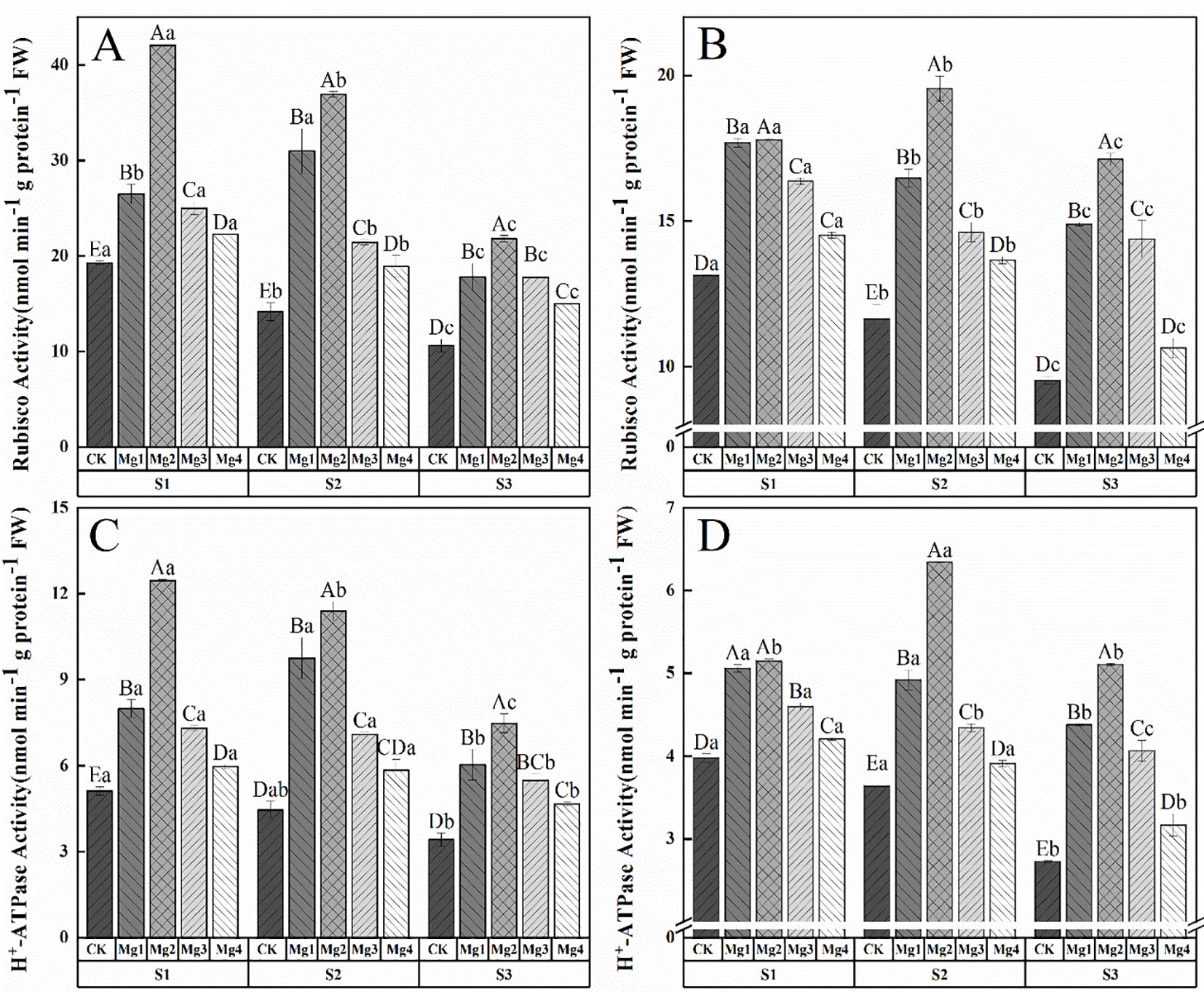
Figure 4. Effects of magnesium application on photosynthetic carbon assimilation and stomatal regulation in flag leaves of rice 20 days after heading. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1magnesium application, respectively. Rubisco enzyme activity of 2022 and 2023 (A, B), H+-ATP enzyme activity of 2022 and 2023 (C, D). Within the same saline-alkaline soil, the values with different uppercase letters indicate significant differences in different magnesium application at a significance level of 5%; within the same magnesium application, the values with different lowercase letters indicate significant differences in different saline-alkaline soil at a significance level of 5%.
3.8 Effects of magnesium application on indicators of oxidative damage and osmotic homeostasis in rice leaves at 20 days after heading
As analyzed in Table 5, there was a highly significant interaction effect between different concentrations of magnesium sulfate and salt treatments on the content of osmoregulatory substances in rice flag leaves 20 days post-heading. In the 2022–2023 trials, the contents of soluble sugars and proline in the flag leaves 20 days post-heading showed a trend of first increasing and then decreasing with the increase of magnesium sulfate concentrations, with the maximum values generally observed under the S3 treatment. The contents of soluble proteins, malondialdehyde, and hydrogen peroxide, as peroxidation products, generally increased with the rise in salt concentrations. The treatment effects among different concentrations of magnesium sulfate were significantly different, with Mg2 showing the most optimal effect overall. Over the two years of trials, compared to the control treatment, the Mg2 treatment increased the content of soluble sugars by 96.8% and 46.6% (S1), 30% and 15.8% (S2), 55.6% and 86.2% (S3); proline content by 15.5% and 12.5% (S1), 16.5% and 25.3% (S2), 20.8% and 35% (S3); soluble protein content decreased by 50.9% and 13.7% (S1), 54.8% and 26.8% (S2), 32.9% and 31.4% (S3); malondialdehyde content decreased by 70.6% and 48.2% (S1), 48.3% and 43.6% (S2), 80% and 44.1% (S3); hydrogen peroxide content decreased by 49.5% and 32.3% (S1), 35.5% and 36.6% (S2), 39.5% and 59.6% (S3).
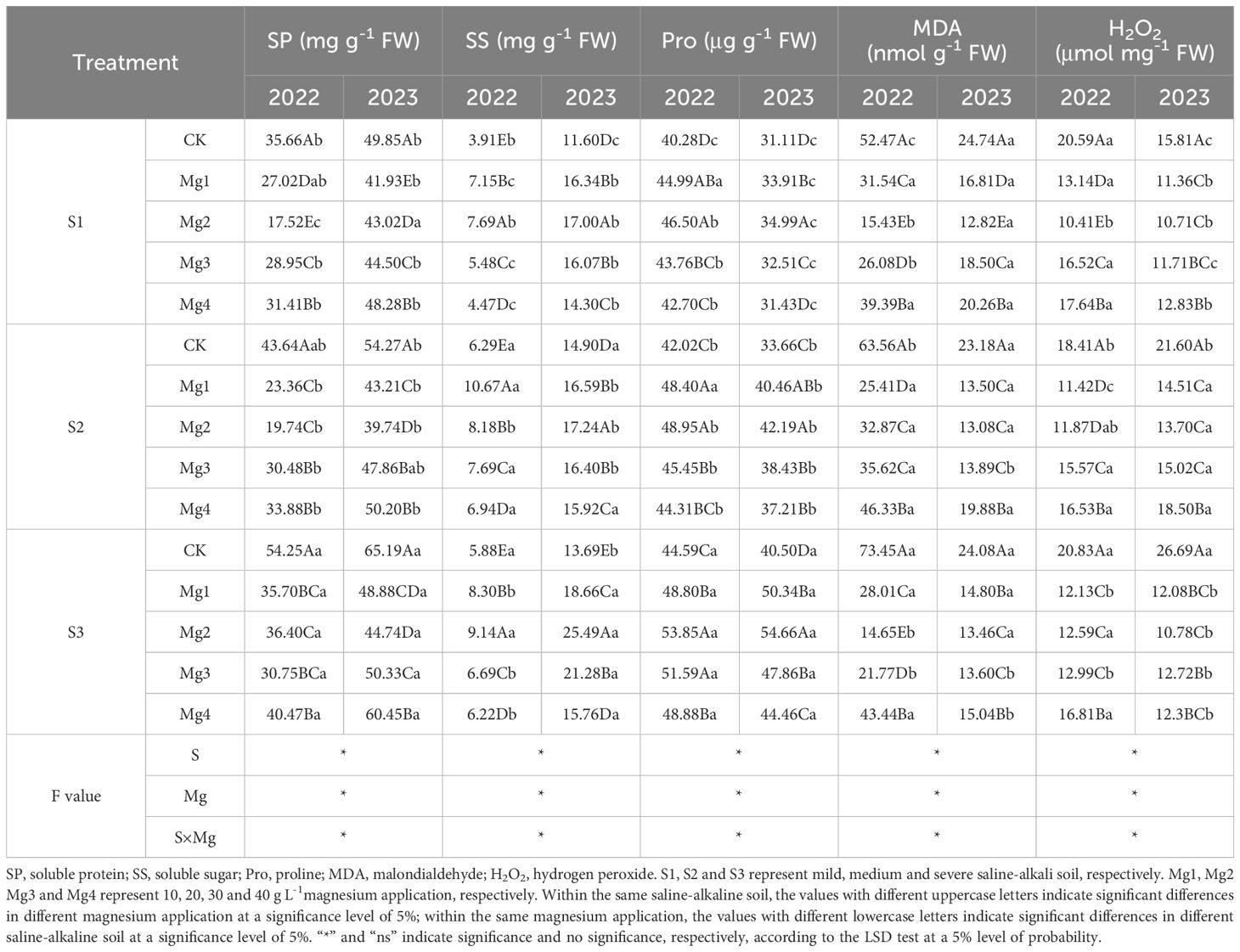
Table 5. Effects of magnesium application on indicators of oxidative damage and osmotic homeostasis in rice leaves at 20 days after heading.
3.9 Effect of magnesium application on the antioxidant system of flag leaves in rice 20 days after heading
As shown in Figure 5, the application of magnesium sulfate at different concentrations significantly increased the CAT enzyme activity in rice leaves 20 days after heading in both years of the study. The enhancement of rice leaf resistance varied with the concentration of magnesium sulfate, showing significant differences among treatments. Overall, the Mg2 treatment was the most effective across different salt concentrations, significantly increasing by 299.5%and 109.1%(S1), 269.6%, and 161%(S2), 232.2%and 160.4%(S3) compared to the control (CK) in 2022-2023, respectively. Figure 5 indicate that the SOD enzyme activity in rice leaves initially increased and then decreased with the rise in magnesium sulfate concentration over the two years, with the treatment effect ranking as Mg2 > Mg1 > Mg3 > Mg4 > CK. In 2022, the maximum SOD activity of 38.35 U g protein-1 FW was achieved under S1 treatment, a157.9% increase compared to the CK. In 2023, the maximum activity of 13.66 U g protein-1 FW was reached under S2 treatment, a 125.6% increase over the CK. Figure 5 show that the APX enzyme activity in rice leaves under different salt concentrations was significantly enhanced by magnesium sulfate treatments, with significant differences among treatments. In 2022, the Mg1 treatment increased APX activity by 62.3% (S1), 186.9% (S2), and 136.6% (S3) compared to the control, while the Mg2 treatment increased it by 154.4% (S1), 258.4% (S2), and 191.9% (S3). The Mg3 and Mg4 treatments showed increases of 43.7% (S1), 97.6% (S2), 119.2% (S3) and 27.3% (S1), 55.4% (S2), 75% (S3), respectively. In 2023, the Mg1, Mg2, Mg3, and Mg4 treatments increased APX activity by 53.8% (S1), 61.7% (S2), 95.9% (S3), 67% (S1), 103.6% (S2), 149.9% (S3), 32.4% (S1), 63.5% (S2), 82.3% (S3), and 11.9% (S1), 23.1% (S2), 36.8% (S3), respectively. Figure 5 illustrate that the POD enzyme activity in untreated rice leaves decreased with increasing salt concentration, while magnesium sulfate treatments significantly enhanced POD activity. The overall trend was similar to that of SOD activity, initially increasing and then decreasing with the rise in magnesium sulfate concentration, with the Mg2 treatment being the most effective. The maximum POD activity of 556.7 U g protein-1 FW and 276.1 U g protein-1 FW was achieved under S3 treatment in both years, representing increases of 791.2% and 446.2% over the control, respectively.
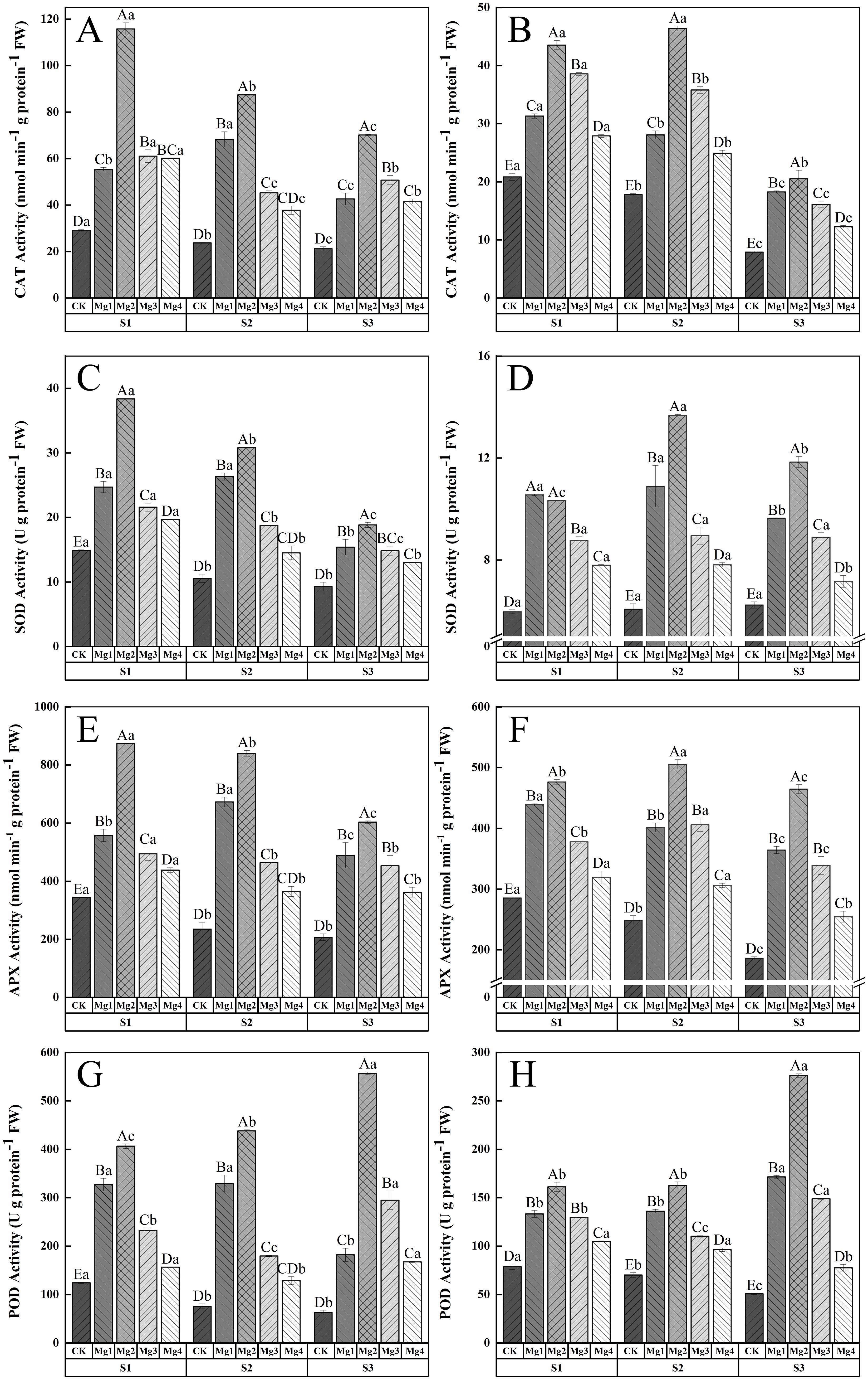
Figure 5. Effect of magnesium application on the antioxidant system of flag leaves in rice 20 days after heading. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1magnesium application, respectively. CAT enzyme activity of 2022 and 2023 (A, B), SOD enzyme activity of 2022 and 2023 (C, D), APX enzyme activity of 2022 and 2023 (E, F), POD enzyme activity of 2022 and 2023 (G, H), Within the same saline-alkaline soil, the values with different uppercase letters indicate significant differences in different magnesium application at a significance level of 5%. Within the same magnesium application, the values with different lowercase letters indicate significant differences in different saline-alkaline soil at a significance level of 5%.
3.10 Effects of magnesium application on element contents in various organs of rice at maturity stage
In the experiments conducted in 2022 (Figures 6-1) and 2023 (Figures 6-2), the elemental content in various organs of rice at maturity varied significantly. The distribution of Mg content was highest in leaves, followed by stems and sheaths, and then panicle; the distribution of Na and K content was highest in stems and sheaths, followed by leaves, and then panicle; the distribution of P content was highest in panicle, followed by stems and sheaths, and then leaves; and Ca was predominantly found in leaves. As the salinity concentration increased, the contents of Mg, K, P, and Ca in the stems, sheaths, and leaves of rice at maturity showed a gradual decline, while the contents of Mg, K, P, and Ca in the panicle first increased and then decreased, and the content of Na increased with the rise in salinity concentration. In both experimental years, the Mg content in different parts of the rice at maturity reached its maximum under the Mg2 treatment, with stems and sheaths content increasing by an average of 44.3% and 31.3% compared to the control treatment, leaf content increasing by an average of 25.6% and 19.8%, and panicle content increasing by an average of 45.6% and 9.3%. Similarly, Na content reached its minimum under this treatment, with stems and sheaths content decreasing by an average of 24.8% and 42.9% compared to the control treatment, leaf content decreasing by an average of 40.2% and 43.2%, and panicle content decreasing by an average of 35.8% and 36.1%. The application of magnesium sulfate at varying concentrations significantly enhanced the elemental content in various organs of rice at maturity, with significant differences among treatments. Overall, except for the Na element, which initially decreased and then increased, the content of other elements generally increased first and then decreased with the increase in magnesium sulfate concentration. The effectiveness of different concentrations of magnesium sulfate treatments was ranked as Mg2 > Mg1 > Mg3 > Mg4.
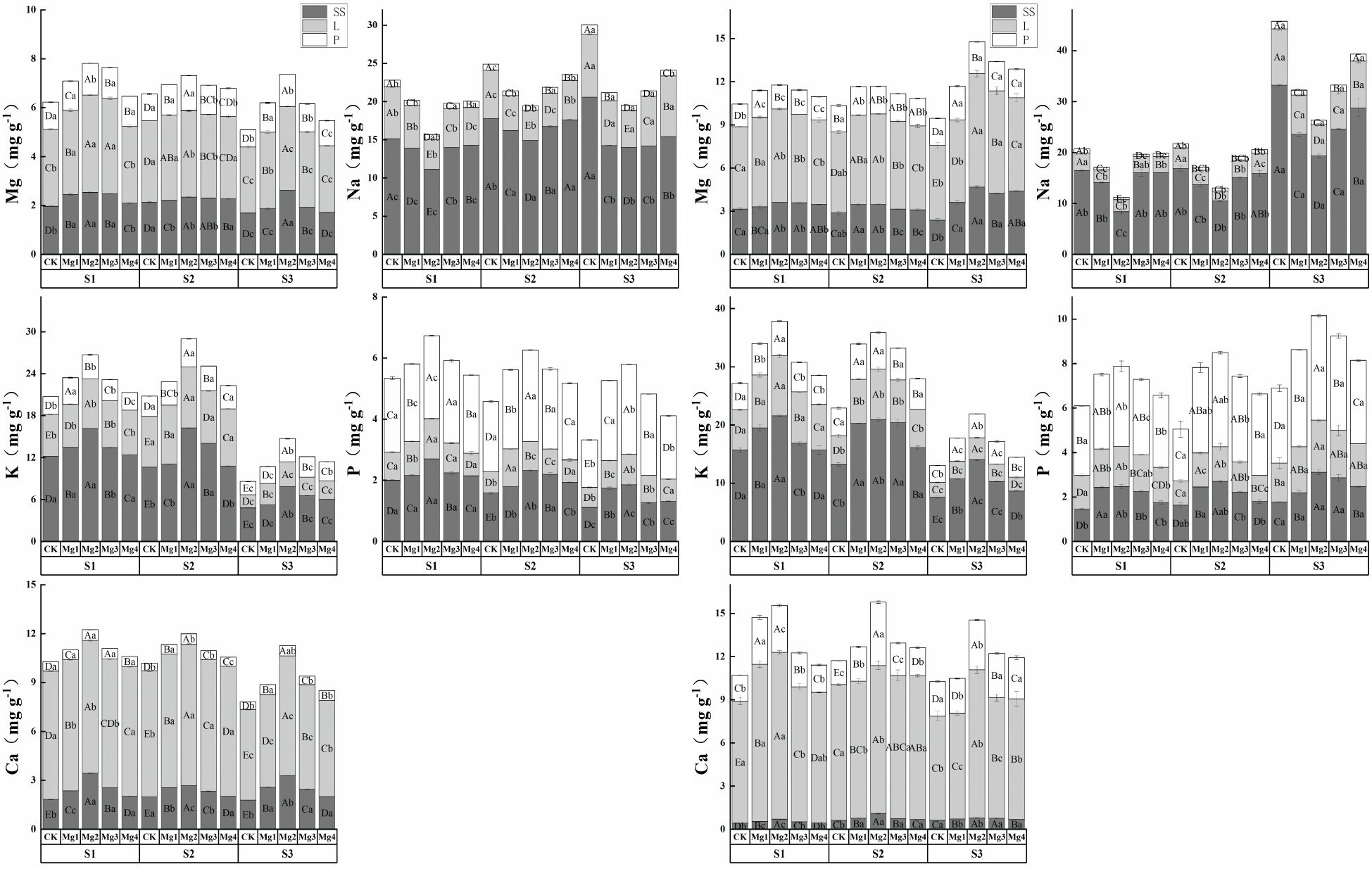
Figure 6. (1) Effects of magnesium application on elemental content in various organs of rice at maturity stage in 2022. SS represents stem sheath; L represents leave; P represents panicle. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1magnesium application, respectively. Within the same saline-alkaline soil, the values with different uppercase letters indicate significant differences in different magnesium application at a significance level of 5%; within the same magnesium application, the values with different lowercase letters indicate significant differences in different saline-alkaline soil at a significance level of 5%. (2) Effects of Magnesium Application Elemental Content in Various Organs of Rice at Maturity Stage in 2023. In the figure, SS represents stem sheath; L represents leave; P represents panicle. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1magnesium application, respectively. Within the same saline-alkaline soil, the values with different uppercase letters indicate significant differences in different magnesium application at a significance level of 5%; within the same magnesium application, the values with different lowercase letters indicate significant differences in different saline-alkaline soil at a significance level of 5%.
3.11 Effects of magnesium application on element ratios in various organs of rice at maturity stage
Analysis of Figure 7 indicates that, overall, during the two-year experiment, the ratios of Mg/Na, K/Na, P/Na, and Ca/Na in various organs of rice at maturity decreased with increasing salt concentration. Notably, the decline in ion ratios was more pronounced in the leaves compared to other parts. Following the application of different concentrations of exogenous magnesium sulfate, the ion ratios in the rice organs initially increased and then decreased as the concentration of exogenous magnesium sulfate rose, with the highest values observed in the spike under the Mg2 treatment. The effectiveness of exogenous magnesium sulfate treatments was ranked as follows: Mg2 > Mg1 > Mg3 > Mg4 > CK, with the ion ratios in rice organs exhibiting the order: panicle > leaves > sheath.
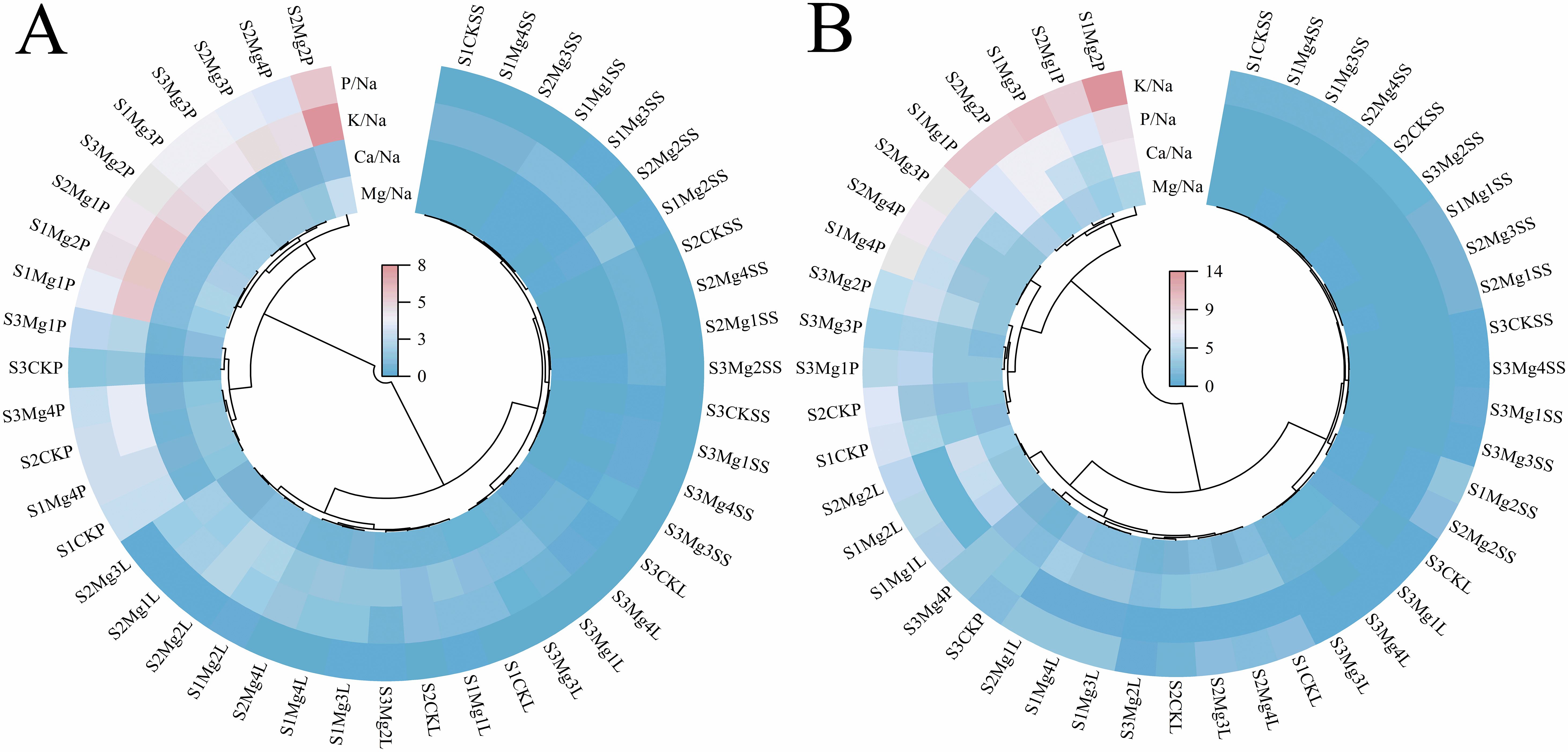
Figure 7. Analysis of elemental ratio heat maps in various organs of rice at maturity stage of magnesium application. (A) shows the year 2022 and (B) shows the year 2023. S1, S2 and S3 represent mild, medium and severe saline-alkali soil, respectively. Mg1, Mg2 Mg3 and Mg4 represent 10, 20, 30 and 40 g L-1magnesium application, respectively. MS is expressed as the Maturity stage; SS represents stem sheath; L represents leave; P represents panicle.
3.12 Effects of magnesium application on economic benefits of rice production
As shown in Table 6, across the two-year field trials, rice earnings peaked at S1 and gradually decreased as salt concentration increased. Foliar application of magnesium sulfate significantly increased rice earnings, with the Mg2 treatment consistently outperforming other treatments under different saline-alkali soil conditions. Compared to the control treatment, the Mg2 treatment achieved a maximum rice revenue of 11,825 CNY ha-¹, representing a 33.9% increase.
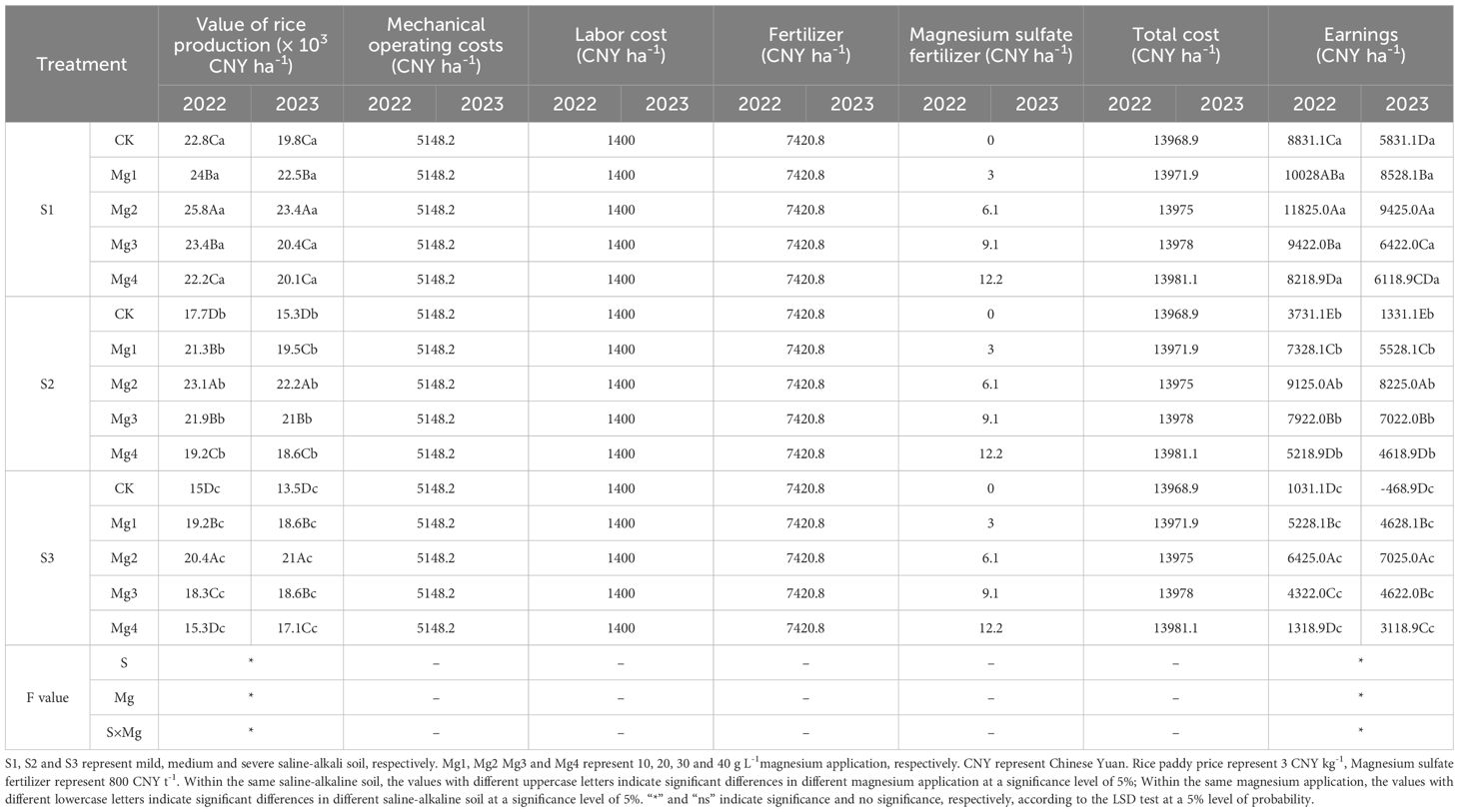
Table 6. Effects of magnesium application on economic benefits of rice production.
4 Correlation analysis
As shown in Figures 8, upon conducting a correlation analysis of rice yield and related physiological indicators across two years of trials, it was observed that rice yield exhibited a significant positive correlation with the activities of antioxidant enzymes such as CAT and POD, the activity of photosynthetic assimilatory enzymes like Rubisco, and the content of mature-stage nutrients including Mg, K, P, and Ca, with correlation coefficients greater than 0.5 (Figure 8). Conversely, there was a significant negative correlation with correlation coefficients less than -0.5 between rice yield and leaf membrane lipid peroxidation products MDA and H2O2, soluble protein, and the content of toxic sodium ions at maturity. Additionally, it was noted that net photosynthetic rate and chlorophyll content had varying impacts on the physiological indicators of rice leaves.
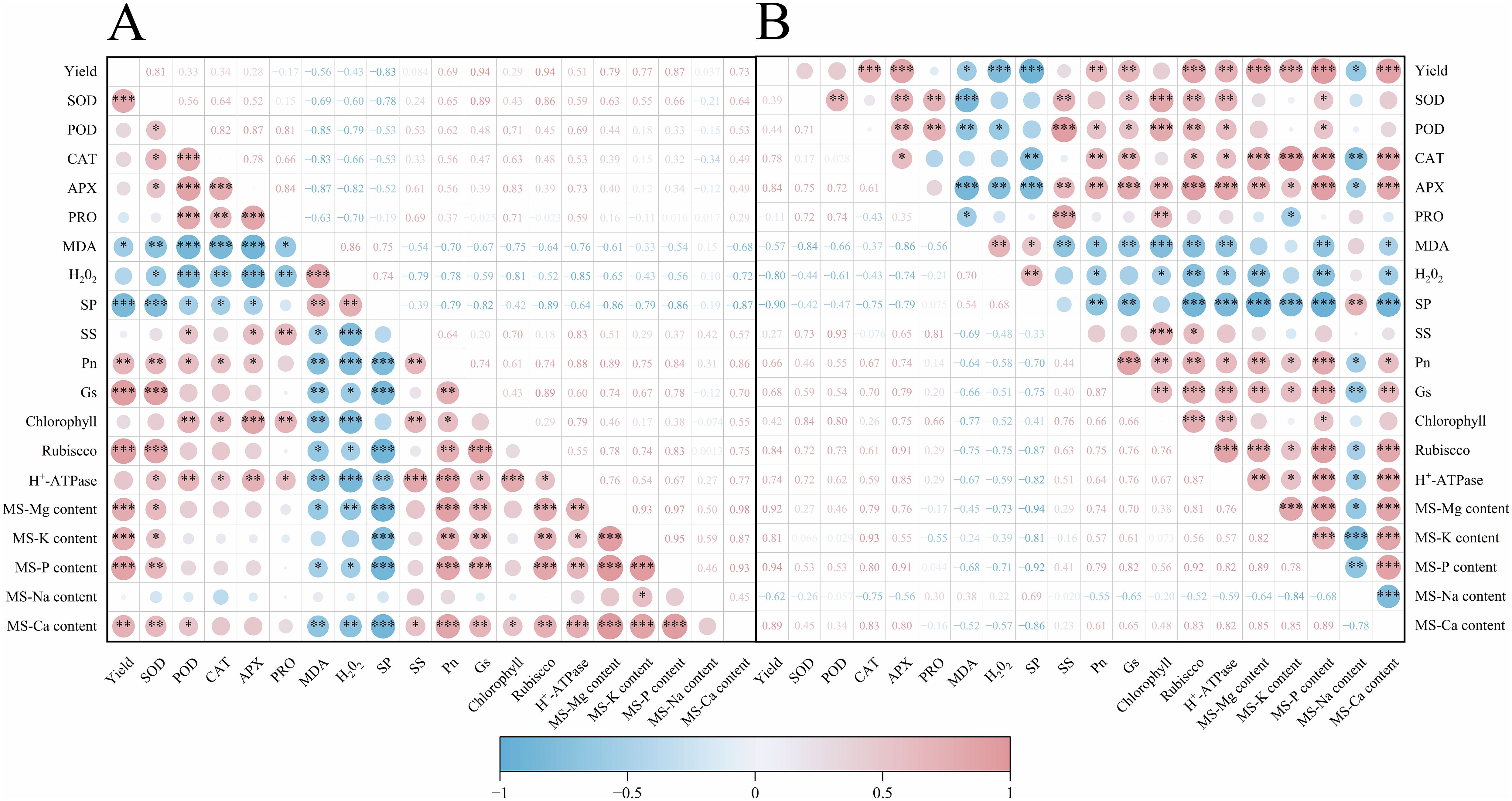
Figure 8. Effects of magnesium application on rice yield and correlation analysis of related indicators in 2022-2023. (A) shows the year 2022 and (B) shows the year 2023. SP refers to soluble protein; SS denotes soluble sugar; Pn represents the net photosynthetic rate; Gs denotes the stomatal conductance; MS-Mg content represents the magnesium content at maturity stage; MS-K content denotes the potassium content at maturity stage; MS-P content signifies the phosphorus content at maturity stage; MS-Na content indicates the sodium content at maturity stage; MS-Ca content refers to the calcium content at maturity stage. "*" denotes a significant interaction at the 0.05 level; "**" denotes a significant interaction at the 0.01 level; "***" denotes a significant interaction at the 0.001 level.
5 Discussion
Over the past decade, rice has been recognized as a pioneer crop for saline-alkali soil reclamation, and the impacts of salt stress on its yield formation process, along with the underlying mechanisms, have been extensively studied and well-documented (Leung et al., 2002; Ganie et al., 2019; Jiang et al., 2024). Studies by Wei (Wei et al., 2024) and Geng (Geng et al., 2024) have demonstrated that under saline-alkaline conditions, key indicators such as the number of grains per panicle, 1000-grain weight, and seed setting rate in rice significantly decrease, collectively leading to a substantial reduction in rice yield. In the current study, rice yield in severe saline-alkaline soil was approximately 33.3% and 31.8% lower than that in median saline-alkaline soil over two years. Compared to that in 2022, rice yield under varying salt concentrations in 2023 declined by 12% (S1), 13.6% (S2), and 10% (S3), respectively. It showed that all components of rice yield are affected by salt stress, leading to a significant decline in yield. In the current study, magnesium sulfate sparaying displayed significant improvements in rice grain filling rate and thousand-grain weight and increased rice yield by an average of 6.7% (S1), 27.3% (S2), and 30.2% (S3) across the two years, whereas Mg2 treatment showed the most significant yield increasing effect among all the four application rates. The difference in the yield improvement effect of magnesium sulfate application on rice under different salt concentrations originates from the threshold-dependent interaction between magnesium-mediated stress tolerance and the plant’s physiological requirements. Under the high-salinity condition (S3: 4.7 mS cm-1), Mg2+ treatment significantly alleviated Na+ toxicity by reducing Na+ accumulation in the aboveground parts, promoting Mg2+ absorption, and enhancing chloroplast integrity. Meanwhile, Mg2+ synergistically activated the antioxidant network, systematically reducing oxidative damage and improving photosynthetic carbon assimilation efficiency. These reparative effects facilitated the allocation of dry matter to the grains, increasing the filled-grain percentage and achieving an average yield increase of 30.2%. Under the low-salinity condition (S1: 2.54 mS cm-1), the effect of Mg2+ sparying was weakened in some degree due to the relatively lower Na+ stress accompanied with higher aboveground Mg2+ level compared to those under higher salinity stress. Despite minor improvements in the antioxidant system and leaf pigments, photosynthesis approaching saturation and nutrient redundancy resulted in a small yield increase in rice. This result was consistent with the findings of Hao’s research (Hao, 2023), whereas both studies did not provide a detailed analysis of the specific mechanisms by which Mg2+ alleviate Na+ accumulation, improve antioxidant systems, and enhance photosynthesis.
Magnesium, as an essential macronutrient in photosynthesis, plays a crucial role in maintaining carbohydrate synthesis and promoting the transport between “source and sink,” thereby enhancing crop yield and quality, such as in cowpea (Ciscomani-Larios et al., 2021), sugar beet (Pogłodziński et al., 2021), tobacco (Ishfaq et al., 2022), and tea (Ruan et al., 2011). Previous studies have shown that exogenous magnesium fertilizer effectively enhances biomass accumulation and chlorophyll content in timothy grass (Radkowski et al., 2017) and faba beans (Neuhaus et al., 2014).It has also been demonstrated to improve carbohydrate and nutrient distribution (nitrogen, phosphorus, potassium) in bananas under nutrient-deficient conditions (He et al., 2020).In wheat, magnesium application significantly increases photosynthetic system efficiency and promotes carbohydrate transport (Ba et al., 2020).Additionally, magnesium elevates antioxidant enzyme activity in pepper (Sevgin Zirek and Uzal, 2020) and rice seedlings (Xuan et al., 2022) exposed to salt stress. These combined effects ultimately promote normal plant growth under various abiotic stresses. In this study, we revealed that the physiological mechanisms by which foliar application of magnesium sulfate alleviates salt stress in rice are salt-dependent. Under conditions of severe salinity and alkalinity(S3), the application of an appropriate amount of magnesium sulfate via leaf spraying resulted in an average increase of 45.4% in chlorophyll a content, optimizing the chlorophyll a/b ratio. This is attributed to magnesium ions serving as the central metal ion in the chlorophyll porphyrin ring, where their concentration directly influences the pigment content and photosynthetic capacity of the leaves. The effective supplementation of exogenous magnesium ions may have inhibited the expression of the SGR gene, reduced the activity of de-magnesium chlorophyllase, delayed chlorophyll degradation, and maintained the thylakoid pH gradient and membrane potential. This ensured the smooth transfer of electrons from PSII to PSI, preventing the formation of triplet state chlorophyll, thereby reducing the generation of singlet oxygen (¹O2) at its source (Meng et al., 2023). Research indicates that the OsMGT3 magnesium transporter on the chloroplast membrane facilitates the import of exogenous Mg²+ from the cytoplasm into the chloroplast, allowing the lysine residues at the active site of the Rubisco large subunit to effectively bind with Mg²+ and CO2, forming the Rubisco-Mg²+-CO2 complex, which maintains the magnesium ion concentration required for Rubisco activation (Chen et al., 2022). Furthermore, the exogenous magnesium sulfate indirectly prevents the oxidative modification of the Rubisco large subunit by reducing ROS production at its source, thereby preserving its structural stability and enzymatic activity. This aligns with our experimental observations, which showed an approximate 92.2% increase in Rubisco enzyme activity and an approximately 89.3% increase in net photosynthetic rate compared to the control treatment. However, since the gene expression or protein abundance related to pigment synthesis, degradation, and Rubisco activation were not determined in the current study, further molecular biology experiments are necessary to validate the inferred mechanisms regarding the delay in chlorophyll synthesis and the maintenance of Rubisco function. In studies involving leguminous plants, the application of exogenous magnesium sulfate has been found to significantly enhance the expression of the H+-ATPase gene (primarily vha2). This activation aids in enhancing proton pump activity, promoting the transmembrane transport of other metabolic products such as sucrose, and providing dynamic support for the distribution of photosynthetic products (Neuhaus et al., 2013). In our experiment, H+-ATPase activity increased by approximately 119.1%, which may correlate with these findings. Numerous studies have demonstrated that magnesium ions can enhance cell membrane integrity, mitigate ROS accumulation induced by salt stress, reduce the stress load on the antioxidant system, and improve enzyme activity stability. As a cofactor or structural stabilizer for enzymes, magnesium may directly participate in maintaining the conformation and catalytic activity of antioxidant enzymes under optimal pH and redox conditions. Although there is currently a lack of conclusive molecular evidence regarding the direct regulation of antioxidant enzyme gene expression by Mg²+, it is possible that it indirectly promotes their transcription or translation by influencing ROS-mediated signaling pathways (such as Ca²+/MAPK). This is also reflected in our experiment, where antioxidant enzyme activity was significantly enhanced compared to the control treatment. However, the lack of analysis regarding the changes in the expression of ROS signaling pathways and antioxidant enzyme-encoding genes means that the signal regulation inferred from the literature still requires molecular evidence support.
In contrast, under mild saline-alkali soil (S1), although magnesium sulfate application also increased chlorophyll a content (by approximately 58.2%) and antioxidant enzyme activity, the mild oxidative stress (with H2O2 content decreasing by approximately 32.3%) and relatively stable ion balance resulted in diminished physiological benefits from exogenous magnesium sulfate. Compared to high salinity conditions, the redundant absorption of nutritional elements in rice leaves and stems at low salinity concentrations failed to surpass the photosynthetic saturation threshold, indicating a diminishing marginal benefit of the antioxidant system and ion homeostasis investment. Overall, the treatment effects at concentrations of 10, 20, 30, and 40 g L-1 showed a decreasing trend with increasing magnesium sulfate concentration. This indicates that excessive magnesium sulfate fertilizer negatively impacts rice growth, yield, and physiological responses, consistent with findings by Suman (Lamichhane et al., 2023) although it still improved rice growth to some extent in saline-alkali soil.
Research has indicated that under salt stress, rice can synthesize soluble organic osmotic adjustment substances to alter cell osmotic potential, thereby alleviating physiological drought caused by the accumulation of high concentrations of sodium ions (Rao et al., 2013). Proline, as an osmotic pressure-regulating solute, can stabilize cell structures and maintain organelle functionality under stress conditions such as salt stress to support normal plant growth; sugars, as the final products of photosynthetic assimilation, together with proline, participate in the defense mechanism against abiotic stress in rice (Alavilli et al., 2023; Zhang et al., 2023). In this experiment, the contents of proline and soluble sugar in rice leaves both gradually increased with the increase in salt concentration. Moreover, the levels of both substances showed a trend of first increasing and then decreasing as the concentration of magnesium sulfate increased, indicating that foliar application of magnesium sulfate, at appropriate concentrations, enhances the content of osmoregulatory substances in rice leaves, thereby alleviating the osmotic stress caused by sodium ions. With increasing salt concentrations, the accumulation of toxic sodium ions in various rice organs also increased, leading to a gradual disruption of ionic homeostasis, exacerbated physiological drought, and increased leaf tip necrosis; leaf cell membranes suffered more severe damage, with increased levels of peroxidative substances such as malondialdehyde and hydrogen peroxide. Foliar magnesium sulfate supplementation significantly enhanced chlorophyll content (a, b, and total) in rice flag leaves 20 days after heading, particularly under high salinity conditions (S3), where chlorophyll an increased by 59.8% compared to the control enhancing the net photosynthetic rate, increasing the synthesis and transport of photosynthates; it also elevated the activity of antioxidant enzymes and accumulated substantial osmoregulatory substances to maintain normal physiological and biochemical activities in the leaves. Foliar magnesium sulfate application alleviated salt stress-induced damage in rice, maintained high crop yields, and minimized losses in rice production on saline-alkali lands.
During the growth of rice, the absorption of various mineral elements is essential to maintain normal physiological and biochemical activities, as these elements play crucial roles in enzyme activity and metabolism (Zhao et al., 2004). However, numerous studies have shown that under saline-alkali stress, crops experience excessive accumulation of Na+, leading to a decline in the levels of essential nutrients such as K+, Ca2+, and Mg2+ within the plant. Additionally, the high concentration of Na+ competes for translocation sites on the cell membrane, significantly reducing the uptake of ions such as Mg2+ (Wang et al., 2011; Zhang, 2024). At the same time, the content and balance of mineral elements in plants vary significantly among different species and organs (Liu et al., 2017). In rice, Na+ primarily accumulates in the roots, with higher accumulation in the stem and sheath than in the leaves, which is a significant physiological mechanism for adapting to growth under saline-alkaline stress (Zhao et al., 2004). When the Na+ accumulation in leaves exceeds the cell compartmentalization capacity, it disrupts the plant’s internal ion homeostasis, leading to salt damage (Zhao et al., 2020). In this study, the Na content in various organs of rice at maturity was ranked as follows: stem and sheath > leaf > panicle. (Figures 6-1, 2), which is consistent with the findings of Jing and Zhang (Jing et al., 2017; Zhang, 2024). Under salt stress, Na+ primarily enters root cells passively through non-selective cation channels (NSCCs). Studies have shown that the application of exogenous Mg²+ can maintain membrane potential stability, reduce the passive leakage of Na+ through NSCCs, and may indirectly affect the activity of OsHKT1;5 to decrease Na+ transport activity, thereby reducing Na+ accumulation and alleviating Na+ toxicity under salt stress, which enhances plant salt tolerance (Sevgin Zirek and Uzal, 2020). In experiments, the application of exogenous magnesium sulfate resulted in an average reduction of Na+ content in rice sheaths by 33.9%, in leaves by 41.7%, and in spikelets by 36% compared to the control. Concurrently, the plant’s ability to absorb Mg²+ was restored, likely relying on the activation of transport proteins OsMGT1/OsMGT6. High concentrations of Na+ competitively inhibit the activity of Mg²+ transport proteins, reducing magnesium absorption. After the application of exogenous magnesium sulfate, this process was partially reversed according to the law of mass action (Roy and Chowdhury, 2021). In the experiments, the Mg²+ content in rice leaves increased by approximately 22.7% (S1). Some studies suggest that Mg²+ may activate the CBL-CIPK23 complex, which plays a crucial role in the regulation of Na+ and K+ transport, potentially also indirectly affecting the activity of OsNHX1, promoting Na+ compartmentalization and reducing Na+ toxicity in the cytoplasm (Pan et al., 2018). The AtMHX protein in vacuoles stores excess Mg²+, preventing Mg²+/Na+ imbalance in the cytoplasm, which aligns with the observation that the Mg²+/Na+ ratio under Mg²+ treatment was greater than that of the control (Figure 7). AtMHX also aids in storing cytoplasmic Mg²+ in vacuoles, maintaining the transmembrane driving force for potassium absorption (Chen et al., 2021), resulting in a 39.2% increase in potassium content in rice under Mg²+ treatment compared to the control (S1). Following the application of exogenous magnesium sulfate, the increased magnesium content in rice activated the phosphorylation of the Ca²+/H+ antiporter OsCAX1, which may synergistically promote the compartmental accumulation of Ca²+, aiding in cytosolic calcium homeostasis. Under high salt stress, high concentrations of Na+ occupy the Ca²+ binding sites of CNGC-type channels, leading to a decrease in the rate of calcium absorption. Experimental results indicated that the Ca²+ content in the S3 treatment decreased by approximately 14.8% compared to S1. Additionally, Na+ occupies the carboxyl groups of pectin, reducing the binding capacity of Ca²+, weakening the mechanical strength of the cell wall, and exacerbating ion leakage (Li et al., 2022). The application of exogenous magnesium sulfate may competitively bind to the EF-hand domain of SOS3, affecting its interaction with SOS2, thereby regulating the excessive activation of SOS1 and preventing the collapse of the proton gradient. Furthermore, by synergistically enhancing the activity of antioxidant enzymes, it mitigates oxidative damage to Ca²+ channels (Yi et al., 2022), which is consistent with the observation that Ca²+ content under Mg²+ treatment increased by approximately 45.3% compared to the control (S1). However, due to the absence of key ion transport protein genes’ expression analysis (such as OsHKT1;5, OsNHX1, OsMGT1, and OsCAX1), the regulatory mechanisms of magnesium sulfate on rice ion balance remains at a hypothetical level.
It is noteworthy that in this experiment, the experimental site was located in a coastal saline-alkali land within 5 km of the sea, where the sandy loam soil texture and inherently high background levels of sodium (Na+) and potassium (K+) collectively resulted in significantly reduced magnesium (Mg²+) availability. This substantially diminished rice uptake and utilization of soil Mg²+, despite its total content falling within the normal range (Yan and Hou, 2018). Ion analysis further revealed that the application of exogenous magnesium sulfate (MgSO4) did not reduce rice uptake of other essential nutrients (e.g., P, K+, Ca2+), thereby excluding potential ionic interference effects. Despite a series of significant physiological responses were obtained in this study, the inability to establish a complete control treatment (no salt + no magnesium sulfate application) due to field trial conditions somewhat limits our analysis of the independent effects of each treatment. Besides, assessments of related gene expression or protein abundance were lacked regardless of enzyme activity, ion content, and photosynthetic parameters. This has led to a mechanism analysis that is largely based on literature speculation, lacking direct molecular evidence, which presents several limitations in our experimental research. And the same time the application of magnesium sulfate via foliar spraying not only supplemented the exogenous magnesium sulfate for rice grown in saline-alkali soils but also introduced SO4²-, which is an important component of sulfur assimilation and tolerance to abiotic stress. Previous studies have indicated that appropriate concentrations of SO4²- can enhance the antioxidant capacity of crops through the synthesis of glutathione (Barlas et al., 2023). The concentrations of magnesium sulfate applied in this study were 10 g L-1, 20 g L-1, 30 g L-1, and 40 g L-1, corresponding to sulfur contents ranging from 1.3 g L-1 to 5.2g L-1. This is significantly higher than the recommended suitable concentrations reported in earlier studies (4.66–9.32 mg L-1) (Sumardiyono et al., 2015). This suggests that SO4²- may partially contribute to alleviating oxidative stress, while our data strongly support that Mg²+ is the primary driving force behind the observed phenomena.
6 Conclusions
This study pioneers the investigation of the effects of foliar application of magnesium sulfate (MgSO4·7H2O) on the salt tolerance of rice under actual coastal saline-alkali soil conditions, bridging the gap between greenhouse/potted experiments and practical agricultural applications. The findings indicate that the response of rice to magnesium sulfate supplementation exhibits a threshold dependency under coastal saline-alkali conditions, with Mg²+ treatment achieving optimal results in both physiological and yield parameters. Under varying salinity levels of coastal saline-alkali soils, exogenous magnesium sulfate supplementation significantly enhanced rice yield by increasing chlorophyll content and Rubisco enzyme activity, thereby improving photosynthesis. Additionally, it minimized sodium (Na+) toxicity by regulating ion balance (increasing the ratios of Mg²+/Na+, K+/Na+, Ca²+/Na+, and P/Na+) and enhanced antioxidant enzyme activity to mitigate oxidative damage. The optimal exogenous magnesium sulfate treatment not only increased rice yield but also improved economic returns in saline-alkali environments, providing a sustainable and cost-effective strategy for rice cultivation in coastal saline-alkali soils. Even the study holds significant implications for guiding high-yield cultivation of rice in coastal saline-alkali areas and for the rational application of magnesium sulfate in the field, the lack of the control without salinity stress due to field conditions and specific molecular analysis on ion balance and antioxidant systems limited the systematicness and depth of the study. Thus, the molecular and omics approaches need to be integrated in the future work to elaborate on the mechanism by which magnesium regulates salt tolerance in rice.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
YL: Writing – original draft, Data curation, Formal Analysis, Investigation, Project administration, Software, Validation. QZ: Data curation, Investigation, Software, Writing – review & editing. RL: Data curation, Investigation, Writing – review & editing. RW: Data curation, Writing – review & editing. XG: Data curation, Investigation, Writing – review & editing. XZ: Data curation, Software, Writing – review & editing. HW: Writing – review & editing. PG: Data curation, Validation, Writing – review & editing. KX: Writing – review & editing. Q-gD: Conceptualization, Funding acquisition, Resources, Supervision, Writing – review & editing. YC: Conceptualization, Funding acquisition, Methodology, Project administration, Resources, Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was financially supported by the National Key Research and Development Program (2022YFD1900704-05), the Jiangsu Agriculture Science and Technology Innovation Fund CX (23) 1020, Natural Science Foundation of the Jiangsu Higher Education Institutions of China (24KJA210002), the Postgraduate Research & Practice Innovation Program of Jiangsu Province (SJCX22-1781), Scientific and Technological Innovation Fund of Carbon Emissions Peak and Neutrality of Jiangsu Provincial Department of Science and Technology (BE2022304), and the Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Acknowledgments
We would like to thank Yangzhou University and Jiangsu Coast Development Group Co., Ltd for providing the experimental site, Prof. Yinglong Chen for his insightful revisions of the manuscript, Dr. Pinglei Gao for his guidance in data analysis, Yadong Yan and Hu Jun of Jiangsu Coast Development Group Co., Ltd for their help and support in the field experiment and Qi Zhu for her help and support during the experimental process.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alavilli, H., Yolcu, S., Skorupa, M., Aciksoz, S. B., and Asif, M. (2023). Salt and drought stress-mitigating approaches in sugar beet (Beta vulgaris L.) to improve its performance and yield. Planta 258, 30. doi: 10.1007/s00425-023-04189-x
Ba, Q., Zhang, L., Chen, S., Li, G., and Wang, W. (2020). Effects of foliar application of magnesium sulfate on photosynthetic characteristics, dry matter accumulation and its translocation, and carbohydrate metabolism in grain during wheat grain filling. Cereal Res. Commun. 48, 157–163. doi: 10.1007/s42976-020-00026-z
Barlas, N. T., Bahamonde, H. A., Pimentel, C., Domínguez-Huidobro, P., Pina, C. M., and Fernández, V. (2023). Evaluating leaf wettability and salt hygroscopicity as drivers for foliar absorption. Plants 12, 2357. doi: 10.3390/plants12122357
Brito Neto, J. F., Beltrão, N. D. E. M., Rigon, J. P. G., and Capuani, S. (2012). Effect of calcium and magnesium silicate on the growth of the castor oil plant subjected to salinity levels. Sem. Ci. Agr. 33, 2949–2958. doi: 10.5433/1679-0359.2012v33supl1p2949
Campion, E. M., Loughran, S. T., and Walls, D. (2011). “Protein Quantitation and Analysis of Purity,” in Protein Chromatography: Methods and Protocols. Eds. Walls, D. and Loughran, S. T. (Humana Press, Totowa, NJ), 229–258.
Chen, R., Deng, Y., Ding, Y., Guo, J., Qiu, J., Wang, B., et al. (2021). Rice functional genomics: decades’ efforts and roads ahead. Sci. China Life Sci. 65, 33–92. doi: 10.1007/s11427-021-2024-0
Chen, Y. L., Li, R. K., Ge, J. F., Liu, J. G., Wang, W. B., Xu, M. F., et al. (2021). Exogenous melatonin confers enhanced salinity tolerance in rice by blocking the ROS burst and improving Na+/K+ homeostasis. Environ. Exp. Bot. 189, 104530. doi: 10.1016/j.envexpbot.2021.104530
Chen, Y., Liu, Y., Dong, S., Liu, J., Wang, Y., Hussain, S., et al. (2022a). Response of rice yield and grain quality to combined nitrogen application rate and planting density in saline area. Agriculture 12, 1788. doi: 10.3390/agriculture12111788
Chen, Y., Liu, Y., Ge, J., Li, R., Zhang, R., Zhang, Y., et al. (2022b). Improved physiological and morphological traits of root synergistically enhanced salinity tolerance in rice under appropriate nitrogen application rate. Front. Plant Sci. 13, 104530. doi: 10.3389/fpls.2022.982637
Chen, Z. C., Peng, W. T., Li, J., and Liao, H. (2018). Functional dissection and transport mechanism of magnesium in plants. Semin. Cell Dev. Biol. 74, 142–152. doi: 10.1016/j.semcdb.2017.08.005
Chen, C.-Q., Tian, X.-Y., Li, J., Bai, S., Zhang, Z.-Y., Li, Y., et al. (2022). Two central circadian oscillators OsPRR59 and OsPRR95 modulate magnesium homeostasis and carbon fixation in rice. Mol. Plant 15, 1602–1614. doi: 10.1016/j.molp.2022.09.008
Ciscomani-Larios, J. P., Sánchez-Chávez, E., Jacobo-Cuellar, J. L., Sáenz-Hidalgo, H. K., Orduño-Cruz, N., Cruz-Alvarez, O., et al. (2021). Biofortification efficiency with magnesium salts on the increase of bioactive compounds and antioxidant capacity in snap beans. Cienc. Rural 51, e20200442. doi: 10.1590/0103-8478cr20200442
Di, X. R., Zheng, F., Norton, G. J., Beesley, L., Zhang, Z. L., Lin, H., et al. (2021). Physiological responses and transcriptome analyses of upland rice following exposure to arsenite and arsenate. Environ. Exp. Bot. 183, 104366. doi: 10.1016/j.envexpbot.2020.104366
Eynard, A., Lal, R., and Wiebe, K. (2005). Crop response in salt-affected soils. J. Sustain. Agric. 27, 5–50. doi: 10.1300/j064v27n01_03
Fahad, S., Hussain, S., Matloob, A., Khan, F. A., Khaliq, A., Saud, S., et al. (2014). Phytohormones and plant responses to salinity stress: a review. Plant Growth Regul. 75, 391–404. doi: 10.1007/s10725-014-0013-y
Ganie, S. A., Molla, K. A., Henry, R. J., Bhat, K. V., and Mondal, T. K. (2019). Advances in understanding salt tolerance in rice. Theor. Appl. Genet. 132, 851–870. doi: 10.1007/s00122-019-03301-8
Geng, X., Zhang, X., Liu, Y., Zuo, B., Zhu, W., Ma, W., et al. (2024). Grain yield and its characteristics of japonica/indica hybrids rice in coastal saline-alkali lands. Acta Agronom Sin., 1–20. Available online at: https://link.cnki.net/ur/uid/11.1809.S.20240129.1133.002.
Gu, J. F., Chen, Y., Zhang, H., Li, Z. K., Zhou, Q., Yu, C., et al. (2017). Canopy light and nitrogen distributions are related to grain yield and nitrogen use efficiency in rice. Field Crops Res. 206, 74–85. doi: 10.1016/j.fcr.2017.02.021
Hao, J. (2023). Effects of Spraying Magnesium Fertilizer on Nutrient, Yield and Quality of Rice (Shen Yang: Shenyang Agricultural University). doi: 10.27327/d.cnki.gshnu.2022.000604
He, H., Jin, X., Ma, H., Deng, Y., Huang, J., and Yin, L. (2020). Changes of plant biomass partitioning, tissue nutrients and carbohydrates status in magnesium-deficient banana seedlings and remedy potential by foliar application of magnesium. Scientia Hortic. 268, 109377. doi: 10.1016/j.scienta.2020.109377
Ishfaq, M., Wang, Y., Yan, M., Wang, Z., Wu, L., Li, C., et al. (2022). Physiological essence of magnesium in plants and its widespread deficiency in the farming system of China. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.802274
Jiang, D. X., Hou, J. J., Gao, W. W., Tong, X., Li, M., Chu, X., et al. (2021). Exogenous spermidine alleviates the adverse effects of aluminum toxicity on photosystem II through improved antioxidant system and endogenous polyamine contents. Ecotoxicol Environ. Saf. 207, 111265. doi: 10.1016/j.ecoenv.2020.111265
Jiang, W., Wang, X., Wang, Y., Du, Y., Zhang, S., Zhou, H., et al. (2024). S-ABA enhances rice salt tolerance by regulating na+/K+ Balance and hormone homeostasis. Metabolites 14, 181. doi: 10.3390/metabo14040181
Jin, X. F., Yang, X. E., Islam, E., Liu, D., and Mahmood, Q. (2008). Effects of cadmium on ultrastructure and antioxidative defense system in hyperaccumulator and non - hyperaccumulator ecotypes of Sedum alfredii Hance. J. Hazard Mater. 156, 387–397. doi: 10.1016/j.jhazmat.2007.12.064
Jing, P., Cui, M., Qing, T., Zhou, Z., and Dai, Q. (2017). Effects of different saline stress on yield and physiological properties of rice in soil culture. China Rice 23, 26–33. doi: 10.3969/j.issn.1006-8082.2017.04.005
Lamichhane, S., Tarpley, L., and Dou, F. (2023). Impact of excess magnesium salt supply on rice yield, physiological response, and grain mineral content. Sustainability 15, 15741. doi: 10.3390/su152215741
Leung, H., Hettel, G. P., and Cantrell, R. P. (2002). International Rice Research Institute: roles and challenges as we enter the genomics era. Trends Plant Sci. 7, 139–142. doi: 10.1016/S1360-1385(01)02226-9
Li, Y., Fu, L., Lei, Y., Li, Y., and Cheng, H. (2022). New distribution and modification of morphological characteristics of firmiana danxiaensis, an endemic species of China. J. Trop. Subtropical Bot. 30, 735–741.
Li, X., Gao, S., Wu, H., Yang, X., and Wang, L. (2022). Effects of naCl stress on ion absorption and transport in three osmanthus fragrans varieties. Jiangsu Agric. Sci. 50, 116–122. doi: 10.15889/j.issn.1002-1302.2022.23.017
Liu, Z., Wei, Q., and Zhang, H. (2017). Effects of salt stress on mineral element contents in different parts of Elaeagnus angustifo lia seedlings. Chin. J. Ecol. 36, 3501–3509. doi: 10.13292/j.1000-4890.201712.006
Mao, D., Chen, J., Tian, L., Liu, Z., Yang, L., Tang, R., et al. (2014). Arabidopsis transporter MGT6 mediates magnesium uptake and is required for growth under magnesium limitation. Plant Cell 26, 2234–2248. doi: 10.1105/tpc.114.124628
Meng, X., Bai, S., Wang, S., Pan, Y., Chen, K., Xie, K., et al. (2023). The sensitivity of photosynthesis to magnesium deficiency differs between rice (Oryza sativa L.) and cucumber (Cucumis sativus L.). Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1164866
Molina, A., Bueno, P., Marín, M. C., Rodríguez - Rosales, M. P., Belver, A., Venema, K., et al. (2002). Involvement of endogenous salicylic acid content, lipoxygenase and antioxidant enzyme activities in the response of tomato cell suspension cultures to NaCl. New Phytol. 156, 409–415. doi: 10.1046/j.1469-8137.2002.00527.x
Neuhaus, C., Geilfus, C., and Mühling, K. (2014). Increasing root and leaf growth and yield in Mg-deficient faba beans ( Vicia faba ) by MgSO 4 foliar fertilization. J. Plant Nutr. Soil Sci. 177, 741–747. doi: 10.1002/jpln.201300127
Neuhaus, C., Geilfus, C.-M., Zörb, C., and Mühling, K. H. (2013). Transcript expression of Mg-chelatase and H+-ATPase isogenes in Vicia faba leaves as influenced by root and foliar magnesium supply. Plant Soil 368, 41–50. doi: 10.1007/s11104-013-1711-3
Pan, J., Huang, C., Luo, J., Peng, F., and Xue, X. (2018). Effects of salt stress on plant and the mechanism of arbuscular mycorrhizal fungi enhancing salt tolerance of plants. Adv. Earth Sci. 33, 361–372. doi: 10.11867/j.issn.1001-8166.2018.04.0361
Panda, A., Rangani, J., Kumari, A., and Parida, A. K. (2017). Efficient regulation of arsenic translocation to shoot tissue and modulation of phytochelatin levels and antioxidative defense system confers salinity and arsenic tolerance in the halophyte Suaeda maritima. Environ. Exp. Bot. 143, 149–171. doi: 10.1016/j.envexpbot.2017.09.007
Pogłodziński, R., Barłóg, P., and Grzbisz, W. (2021). Effect of nitrogen and magnesium sulfate application on sugar beet yield and quality. Plant Soil Environ. 67, 507–513. doi: 10.17221/336/2021-pse
Pu, L., Han, M., Zhu, M., Zhang, R., and Xiang, Y. (2014). Soil salinization research in China: Advances and prospects. J. Geogr. Sci. 24, 943–960. doi: 10.1007/s11442-014-1130-2
Radkowski, A., Wolski, K., Radkowska, I., and Rapacz, M. (2017). Effect of foliar fertilization with magnesium sulfate and supplemental L-ascorbic acid on dry matter yield and chemical composition of cv. Egida timothy grass. J. Elem. 22 (2), 545–558. doi: 10.5601/jelem.2016.21.3.1167
Rao, P. S., Mishra, B., and Gupta, S. R. (2013). Effects of soil salinity and alkalinity on grain quality of tolerant, semi-tolerant and sensitive rice genotypes. Rice Sci. 20, 284–291. doi: 10.1016/s1672-6308(13)60136-5
Rivelli, A. R., De Maria, S., Pizza, S., and Gherbin, P. (2010). Growth and physiological response of hydroponically-grown sunflower as affected by salinity and magnesium levels. J. Plant Nutr. 33, 1307–1323. doi: 10.1080/01904167.2010.484092
Roy, S. and Chowdhury, N. (2021). “Salt Stress in Plants and Amelioration Strategies: A Critical Review,” in Abiotic Stress in Plants (London: IntechOpen). doi: 10.5772/intechopen.93552
Ruan, J., Ma, L., and Yang, Y. (2011). Magnesium nutrition on accumulation and transport of amino acids in tea plants. J. Sci. Food Agric. 92, 1375–1383. doi: 10.1002/jsfa.4709
Sevgin Zirek, N. and Uzal, O. (2020). The developmental and metabolic effects of different magnesium dozes in pepper plants under salt stress. Not Bot. Horti Agrobo 48, 967–977. doi: 10.15835/nbha48211943
Sumardiyono, C., Suryanti, Rositasari, P., and Dwi Chinta, Y. (2015). Deteksi Pengimbasan Ketahanan Pisang Terhadap Penyakit Layu Fusarium Dengan Asam Fusarat Induced Resistance Detection of Banana Against Fusarium Wilt Using Fusaric Aacid. J Perlindung Tanam Indones. 19 (1), 40–44.
Tester, M. (2003). Na+ Tolerance and na+ Transport in higher plants. Ann. Bot. 91, 503–527. doi: 10.1093/aob/mcg058
Tian, X.-Y., He, D.-D., Bai, S., Zeng, W.-Z., Wang, Z., Wang, M., et al. (2021). Physiological and molecular advances in magnesium nutrition of plants. Plant Soil 468, 1–17. doi: 10.1007/s11104-021-05139-w
Vieira, S. M., Silva, T. M., and Glória, M. B. A. (2010). Influence of processing on the levels of amines and proline and on the physico - chemical characteristics of concentrated orange juice. Food Chem. 119, 7–11. doi: 10.1016/j.foodchem.2008.12.069
Wang, R., Dong, X., Gao, Y., Hao, F., Zhang, H., and Lin, G. (2024). Molecular mechanism of exogenous magnesium in regulating cation homeostasis in roots of peanut seedlings under salt stress. Agronomy. (Basel). 14, 724. doi: 10.3390/agronomy14040724
Wang, L., Wang, Y. F., Wang, X. Y., Li, Y. Y., Peng, F. L., and Wang, L. S. (2014). Regulation of POD activity by pelargonidin during vegetative growth in radish (Raphanus sativus L.). Sci. Hortic. (Amst). 174, 105–111. doi: 10.1016/j.scienta.2014.05.014
Wang, Y., Xue, Y., Yang, K., and Wang, Q. (2011). Effects of NaCl stress on ion contents in different organs of maize (Zea mays L.) seed ling. Chin. J. Ecol. 30, 1654–1661. doi: 10.13292/j.1000-4890.2011.0247
Wang, Y., Zhang, R., Liu, Y., Li, R., Ge, J., Deng, S., et al. (2022). Rice response to salt stress and research progress in salt tolerance mechanism. Chin. J. Rice Sci. 36, 105–117. doi: 10.16819/j.1001-7216.2022.210609
Wei, H., Zhang, X., Zhu, W., Geng, X., Ma, W., Zuo, B., et al. (2024). Effects of salinity stress on grain-filling characteristics and yield of rice. Acta Agronom Sin. 50, 734–746. doi: 10.3724/sp.j.1006.2024.32021
Wu, F., Chen, Z., Zhang, F., Zheng, H., Li, S., Gao, Y., et al. (2022). Identification and transcriptome analysis of genes related to membrane lipid regulation in sweet sorghum under salt stress. IJMS 23, 5465. doi: 10.3390/ijms23105465
Xuan, T. D., Huong, C. T., Quan, N. V., Anh, L. H., Khanh, T. D., and Rayee, R. (2022). Improvement of salinity tolerance in rice seedlings by exogenous magnesium sulfate application. Soil. Syst. 6, 69. doi: 10.3390/soilsystems6030069
Yan, H. and He, W. (2010). Improvement of saline-alkali soil and rice cultivation technology in Northern Yinchuan of Ningxia. Hubei Agric. Sci. 49, 2693–2695+2708. doi: 10.14088/j.cnki.issn0439-8114.2010.11.037
Yan, B. and Hou, Y. (2018). Effect of soil magnesium on plants: a review. IOP Conf. Ser.: Earth Environ. Sci. 170, 22168. doi: 10.1088/1755-1315/170/2/022168
Yi, S., Wang, C., Li, H., Zhou, B., and Sun, X. (2022). Study on mechanisms of arabidopsis thaliana MGT6 in regulating ca2+/mg2+ Homeostasis under high-mg2+ Conditions. Life Sci. Res. 26, 219–223. doi: 10.16605/j.cnki.1007-7847.2021.04.0149
Yildirim, E., Turan, M., and Guvenc, I. (2008). Effect of foliar salicylic acid applications on growth, chlorophyll, and mineral content of cucumber grown under salt stress. J. Plant Nutr. 31, 593–612. doi: 10.1080/01904160801895118
Zhang, Y. (2015). Study on the grain security issues in China in the perspective of globalization (Shanxi: Shanxi Normal University).
Zhang, R. (2024). Characteristics of Different in Salt Tolerance and Betaine Regulation Effect of Rice Cultivars (Yangzhou: YangZhou University). doi: 10.27441/d.cnki.gyzdu.2023.000005
Zhang, X., He, P., Guo, R., Huang, K., and Huang, X. (2023). Effects of salt stress on root morphology, carbon and nitrogen metabolism, and yield of Tartary buckwheat. Sci. Rep. 13, 12483. doi: 10.1038/s41598-023-39634-0
Zhang, M. X., Yu, J., Cao, Y., Ma, J., Bao, C., Wang, X., et al. (2024). Effects of salt stress on physiological and biochemical indicators and total saponin accumulation of Platycodon grandiflorus Jacq. A. DC. J. Biol. 41 (4), 71–77. doi: 10.3969/j.issn.2095-1736.2024.04.071
Zhao, J., Cheng, Y., Liu, Y., Xiang, Y., Huang, Y., and Wang, X. (2020). Effects of salt stress on growth of Oceanrice 86 and absorption transportation and distribution of mineral elements. J. Huazhong Agric. Univ. 39, 7–14. doi: 10.13300/j.cnki.hnlkxb.2020.04.002
Keywords: rice (Oryza sativa L.), salinity stress, magnesium, coastal saline-alkali land, ion homeostasis
Citation: Liu Y, Zhu Q, Li R, Wei R, Geng X, Zhang X, Wei H, Gao P, Xu K, Dai Q and Chen Y (2025) Foliar magnesium enhances rice salt tolerance by improving photosynthesis and regulating ion homeostasis. Front. Plant Sci. 16:1578023. doi: 10.3389/fpls.2025.1578023
Received: 17 February 2025; Accepted: 22 May 2025;
Published: 16 June 2025.
Edited by:
Muhammad Habib ur Rahman, University of Bonn, GermanyReviewed by:
Gian Attilio Sacchi, University of Milan, ItalyThounaojam Thorny Chanu, Assam Don Bosco University, India
Mahmood Ul Hassan, China Agricultural University, China
Copyright © 2025 Liu, Zhu, Li, Wei, Geng, Zhang, Wei, Gao, Xu, Dai and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yinglong Chen, Y2hlbnlpbmdsb25nQHl6dS5lZHUuY24=