 Joanna N. Soldek1‡Marta Ballesteros-Gutiérrez1†‡
Joanna N. Soldek1‡Marta Ballesteros-Gutiérrez1†‡ Laura Díaz-Sáez2Ignacio Delgado-Santamaría1†
Laura Díaz-Sáez2Ignacio Delgado-Santamaría1† José Manuel Palacios1,3
José Manuel Palacios1,3 Marta Albareda1,3*
Marta Albareda1,3*- 1Centro de Biotecnología y Genómica de Plantas, Universidad Politécnica de Madrid – Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA/CSIC), Madrid, Spain
- 2Instituto de Química Física Blas Cabrera, Consejo Superior de Investigaciones Científicas (CSIC), Madrid, Spain
- 3Departamento de Biotecnología-Biología Vegetal, Escuela Técnica Superior de Ingeniería Agronómica, Alimentaria y de Biosistemas, Universidad Politécnica de Madrid, Madrid, Spain
The establishment of the rhizobium-legume symbiosis requires adjusting the behavior of both partners to nodule conditions in which transition metals are delivered to the bacteria, as many rhizobial metalloenzymes are essential for bacteroid functions and symbiotic performance. A previous proteomic analysis revealed the existence of a relevant number of proteins differentially expressed in bacteroids induced by Rhizobium leguminosarum bv. viciae (Rlv) UPM791 in pea and lentil nodules. Among these proteins, a metal-binding protein (RLV_3444) component of an ABC-transporter system (RLV_3442-3444) was shown to be overexpressed in pea bacteroids, suggesting that metal provision to the bacteroid is more restrictive in the rhizobium-pea symbiosis. In this work, protein sequence analysis and structural modelling have revealed that RLV_3444 is highly similar to the functionally characterized zinc-binding protein ZniA from Klebsiella pneumoniae, so the host-dependent binding protein was renamed as ZniA and the transporter system as ZniCBA. The genome of Rlv UPM791 also encodes the conserved high-affinity ZnuABC transporter system. We demonstrate that at least one of the two systems must be present for Rlv to grow under zinc-limiting conditions and for optimal symbiotic performance with pea and lentil plants. The three conserved histidine residues present in multiple Zn2+-binding proteins have been shown as essential for the function of Rlv ZniA, and in-silico modelling suggests that they might participate in metal coordination. We also demonstrate that both ZniCBA and ZnuA are regulated by zinc in a Zur-dependent manner, consistent with the presence of a Zur box in their regulatory region. The expression patterns revealed that ZniCBA is expressed at lower levels than ZnuA, and its expression increased in a znuA mutant under both free-living and symbiotic conditions. These results, along with the observed increment in the expression of ZniCBA in pea versus lentil bacteroids, suggest that the host-dependent transporter system might play an auxiliary function for zinc uptake under zinc starvation conditions and might play a relevant role in the adaptation of rhizobia to the legume host.
Introduction
Zinc is an essential nutrient for cells, as it is required for catalytic or structural functions in numerous proteins (Blindauer, 2015). Excess concentrations of zinc become toxic by competing with other metal ions for biologically important ligands, and so intracellular zinc levels must be tightly regulated. Bacterial zinc homeostasis requires the balance of import and export systems to control zinc concentration inside the cell, thus preventing metal toxicity (Choi and Bird, 2014). Under zinc-repleted conditions, zinc supply is mediated by low-affinity transporters such as the constitutively expressed ZupT, belonging to the ZIP (zinc-iron permeases) family, which has a broad affinity for divalent metals with a preference for zinc (Grass et al., 2005). On the contrary, high-affinity zinc uptake systems, commonly mediated by members of the ATP-binding cassette (ABC) transporter superfamily, operate as a response to zinc shortage conditions. The expression levels of these transporters are controlled at the transcriptional level by Zur (zinc uptake regulator), a member of the ferric uptake transcriptional regulator (Fur) family. When enough Zn is present in the cell, Zn atoms bind to Zur protein, thus turning it into a repressor of genes involved in Zn uptake (Hantke, 2005; Mikhaylina et al., 2018; Kandari et al., 2021).
ABC transporter systems rely on three components: (i) a membrane-spanning permease, (ii) a cytosolic ATPase, and (iii) a periplasmic substrate-binding protein (SBP) that confers high affinity and selectivity to the system and forms a complex with the membrane component to deliver the metal into the cytosol (Cerasi et al., 2013; Blindauer, 2015; Bui and Inaba, 2024). The SBP structures are characterized by two independent globular α/β domains that interact to create a metal-binding pocket. Different kinds of SBPs have been described, and those from cluster A, distinguished by an α-helix that connects the two domains, comprise the subcluster A-I, including SBPs that directly interact with metal ions (Fe2+, Zn2+, or Mn2+) (Berntsson et al., 2010). Structures of Zn2+-specific SBPs have revealed the existence of three conserved histidine residues that participate in metal coordination together with a conserved glutamate/aspartate residue or a water molecule (Banerjee et al., 2003; Yatsunyk et al., 2008; Ilari et al., 2011; Handali et al., 2015; Maunders et al., 2024). A glutamate residue at the same position of the third histidine participating in the coordination environment has been associated with Mn2+- or Fe2+-specific SBPs (Lawrence et al., 1998; Rukhman et al., 2005; Sun et al., 2009). Furthermore, some Zn2+-binding SBPs contain a histidine-rich domain that has been related to sensing/regulation or zinc acquisition functions (Banerjee et al., 2003; Wei et al., 2007; Neupane et al., 2017).
The symbiotic rhizobium-legume interaction, a key sustainable source of nitrogen in agro-ecosystems, requires adjusting the behavior of both partners by sophisticated plant- and bacterial-dependent mechanisms that lead to the formation of the root nodules (Ledermann et al., 2021; Hawkins and Oresnik, 2022). In the nodules, bacteria are surrounded by a plant-derived membrane, establishing intracellular symbiotic organelles (symbiosomes) where they differentiate into bacteroids, the symbiotic form of rhizobia that express the nitrogenase enzyme catalyzing the conversion of N2 into ammonia (Poole et al., 2018). Bacteroids are fully dependent on plant cells for the acquisition of nutrients, and establishment of the symbiosis triggers metal uptake mechanisms in the bacteroid, as many enzymes required for symbiotic performance are highly expressed metalloenzymes, including the nitrogenase, superoxide dismutases, and others required for bacteroid functions (Abreu et al., 2019; González-Guerrero et al., 2023). Furthermore, metals are transported to the bacteroid in complexes with organic acids, and availability of metals is sometimes compromised due to the speciation of the corresponding cation with strong plant-derived chelators (Moreau et al., 1995; Cacho et al., 2010). Although nodules constitute approximately 5% of the total plant biomass, they may contain 25%–30% of the total transition metals from the plant (González-Guerrero et al., 2023). Consequently, metal-related deficiencies, and in particular zinc soil deficiencies or alterations in the metal delivery pathways to the cytosol of plant-infected cells, have been shown to affect nodulation, symbiotic nitrogen fixation, and legume productivity (O’Hara, 2001; Abreu et al., 2017; León-Mediavilla et al., 2018).
In Gram-negative bacteria, zinc uptake is commonly mediated by the conserved high-affinity ZnuABC transporter system belonging to the ABC transporter superfamily (Patzer and Hantke, 2000), which plays an important role in pathogenic bacteria adaptation to zinc-limiting conditions. This system promotes survival and virulence in the host, in which metal availability is actively reduced in response to bacterial infection (Patzer and Hantke, 1998; Chen and Morse, 2001; Ammendola et al., 2007; Vahling-Armstrong et al., 2012; Akhtar and Turner, 2022). However, metal transport has been shown to involve more than a single system, and redundant functions for zinc uptake under these conditions have been observed (Desrosiers et al., 2010; Corbett et al., 2012; Cerasi et al., 2013; Plumptre et al., 2014; Pederick et al., 2015; Maunders et al., 2024). In contrast to the extensive studies in pathogenic bacteria, functional studies of zinc homeostasis in plant-associated bacteria are scarce. The requirement of Sinorhizobium meliloti ZnuABC for growth under zinc-limiting conditions has been demonstrated (Vahling-Armstrong et al., 2012), but its functional role during symbiosis has not been studied. In Agrobacterium tumefaciens, two zinc ABC transporters have been described (Bhubhanil et al., 2014; Chaoprasid et al., 2016), and the functional role of ZnuABC was revealed in the absence of the second transporter system, TroCBA, that predominantly plays zinc uptake functions under zinc-limiting conditions (Chaoprasid et al., 2016). In this system, the virulence of mutants affected by single mutations in the permease troC or in znuA was not affected (Bhubhanil et al., 2014; Chaoprasid et al., 2016), and the cooperation of both transporter systems in plant virulence has not been demonstrated. A transcriptome-based analysis revealed that the Sinorhizobium fredii znuABC operon was highly induced in soybean bacteroids as compared to free-living cells, and host-dependent upregulation patterns were also observed (Jiao et al., 2016; Jiao et al., 2018). The functional role of the SBP ZnuA has been shown to be dependent on the legume host in the S. fredii-soybean system (Jiao et al., 2018; Zhang et al., 2020), where accessory zinc transporters, such as ZIP transporters, contribute to rhizobia adaptation to the different hosts (Zhang et al., 2020).
Rhizobium leguminosarum bv. viciae (Rlv) UPM791 effectively nodulates legume genera belonging to the Viciae tribe (Pisum, Vicia, Lathyrus, and Lens) (Laguerre et al., 2003), but not all combinations are equally effective, and differences in symbiotic performance associated with the legume host have been described (Den Herder and Parniske, 2009). A proteomic analysis in bacteroids induced by Rlv UPM791 in pea and lentil nodules revealed the existence of a relevant number of proteins differentially expressed (Durán et al., 2021), suggesting that each legume host provides a different environment, inducing specific adaptive responses in the bacteria. Among these proteins, a potential metal-binding protein (RLV_3444, renamed as ZniA), a component of an ABC-transporter system (RLV_3442-3444), here designated as ZniCBA, was shown to be expressed at higher levels in pea bacteroids, suggesting that provision of this element to the bacteroid is more restrictive in the rhizobium-pea symbiosis. In this work, the functional role and expression analysis of this pea-specific SBP have been analyzed. We have demonstrated that ZniCBA is a zinc uptake transporter system that may replace the functional role of the ABC-transporter system ZnuABC under zinc-limiting conditions in free-living cells. The role of both transporter systems in the optimization of the symbiosis of Rlv with pea and lentil plants has also been established. The expression of the zniCBA operon and znuA gene has been shown to be regulated by zinc in a Zur-dependent manner. Both transporters were significantly induced in pea nodules, and their expression pattern suggests that zniCBA might function as an auxiliary zinc uptake system under zinc starvation conditions and might play a relevant role in the adaptation of Rlv to its legume host.
Materials and methods
Bacterial strains, plasmids, and growth conditions
Strains and plasmids used in this study are listed in Supplementary Table S1. R. leguminosarum strains were routinely grown at 28°C in yeast mannitol broth (YMB) (Vincent, 1970), tryptone-yeast extract (TY) (Beringer, 1974), Rhizobium minimal media (Rmin) (O’Gara and Shanmugam, 1976), or universal minimal salt (UMS) medium (Wheatley et al., 2017). Escherichia coli strains were grown at 37°C in LB medium (Bertani, 1951). E. coli DH5α was used for standard cloning procedures (Hanahan, 1983), and E. coli S17.1 (Simon et al., 1983) for conjugative plasmid transfer between E. coli and R. leguminosarum. Antibiotic concentrations used were as follows (µg ml−1): ampicillin, 100; kanamycin, 50; tetracycline, 5 (for R. leguminosarum) or 10 (for E. coli).
For growth curve analyses, bacterial cultures were grown in zinc-free UMS medium up to an OD600 of 0.6 and diluted to an initial OD600 of 0.01 in fresh zinc-free UMS or the same medium containing 50 µM EDTA with or without 50 µM ZnSO4, MnSO4, or FeSO4 supplementation. Growth curves were determined with cell cultures incubated in 100-well Honeycomb plates (Growth Curves Ltd., Piscataway, NJ, USA) with continuous double-orbital shaking in a Bioscreen C Pro device (Growth Curves Ltd., Piscataway, NJ, USA) at 28°C, with OD600 measurement intervals every 30 min for 30 h.
Mutants and plasmid construction
DNA purification, digestion with restriction enzymes, ligation, gel electrophoresis, amplification by polymerase chain reaction (PCR), and E. coli transformation were performed as described in standard protocols (Sambrook and Russell, 2001). Primers used for mutants and plasmid construction are listed in Supplementary Table S2.
Derivatives of Rlv SPF25 incorporating zniA and zniCBA mutations were generated by homologous recombination using the suicide vector pK18mobsac (Schäfer et al., 1994). For this purpose, a first round of PCR reactions amplified two DNA fragments of ca. 1 Kb corresponding to up- and downstream regions of the zniA (rlv_3444) gene using primers P1_rlv_3444_BamHI/P2_rlv_3444 and P3_rlv_3444/P4_rlv_3444_HindIII, respectively, and genomic DNA from the Rlv UPM791 strain as a template. The two PCR products of the flanking DNA regions of the gene were linked via an overlapping GC-rich (C5G5C5) sequence, previously incorporated using P2_rlv_3444 and P3_rlv_3444 primers, by a fusion PCR (Cha-aim et al., 2009) using primers P1_rlv_3444_BamHI/P4_rlv_3444_HindIII. Following the same procedure, regions covering zniC (rlv_3442) upstream and zniA (rlv_3444) downstream were amplified using primers P1_rlv_3442-44_BamHI/P2_rlv_3442–44 and P3_rlv_3444/P4_rlv_3444_HindIII, respectively. A fusion PCR with primers P1_rlv_3442-44_BamHI/P4_rlv_3444_HindIII was performed to amplify the fragment carrying the zniCBA deletion. Fusion PCR products containing zniA and zniCBA deletions were cloned separately in pBlueScript-II KS+ as BamHI-HindIII restriction fragments, sequenced, and moved to the pK18mobsac suicide vector. The resulting plasmids, pK18.ZniA and pK18.ZniCBA, were introduced independently into the SPF25 strain by conjugation, and homologous recombination by a double crossover event was selected by the sacB system in Rhizobium minimal medium (Schäfer et al., 1994). Gene deletions were verified by PCR and sequencing analysis, and the transconjugant strains carrying the mutations in zniA and zniCBA were designated as UPM1629 and UPM1631, respectively.
To generate an in-frame deletion in the znuA gene, regions up- and downstream of the znuA gene were PCR amplified using primers P1_ZnuA_BamHI/P2_ZnuA and P3_ZnuA/P4_ZnuA_HindIII, respectively, and UPM791 genomic DNA as a template. The two PCRs were linked via an overlapping GC-rich sequence by a fusion PCR using primers P1_ZnuA_BamHI/P4_ZnuA_HindIII to amplify the fragment containing the deletion of the znuA gene. The resulting DNA fragment was digested with BamHI-HindIII restriction enzymes and cloned in pBlueScript-II KS+, sequenced, and then moved to the pK18mobsac suicide vector. The new plasmid generated (pK18.ZnuA) was introduced into SPF25 strain and into derivative strains UPM1629 (ΔzniA) and UPM1631 (ΔzniCBA) by conjugation, and homologous recombinations were selected by the sacB system. Gene deletion was verified by PCR and sequencing analysis, and the transconjugant strains carrying mutations in znuA, znuA/zniA, and znuA/zniCBA were designated as UPM1628, UPM1630, and UPM1632, respectively.
To generate mutant derivatives bearing a deletion in the zur gene, two DNA regions up- and downstream of the gene were amplified by PCR using primer pairs P1_Zur/P2_Zur and P3_Zur/P4_Zur, respectively. The resulting DNA fragments were fused by an overlapping GC sequence by PCR using P1_Zur/P4_Zur primers. The PCR product carrying the zur deletion was cloned in plasmid pCR2.1TOPO, sequenced, and moved to the pK18mobsac vector as an XbaI-BamHI restriction fragment, rendering the pK18.Zur plasmid. This construction was introduced into SPF25 and into its derivative strain UPM1630 (ΔznuAΔzniA), and homologous recombination was selected by the sacB system, giving rise to UPM1633 (Δzur) and UPM1634 (ΔzurΔznuAΔzniA). Gene deletion was checked by PCR and sequencing analysis.
For analysis of promoter expression, transcriptional gene fusions were generated with the promoterless gusA gene in plasmid pLMB51 (Tett et al., 2012). DNA fragments containing the upstream regions of zniC and znuA genes were PCR amplified with primers Prom1_rlv_3442-44_BamHI_F/Prom5_rlv_3442-44_XbaI_R and Prom_ZnuA_BamHI_F/Prom_ZnuA_XbaI_R, respectively, and UPM791 genomic DNA as a template. PCR products were cloned separately into pBlueScript-II KS+ plasmid using BamHI-XbaI restriction sites, sequenced, and moved to pLMB51 plasmid, thus generating pLMBZniCBA and pLMBZnuA plasmids, respectively (Tett et al., 2012). For regulatory analysis of the expression of the zniCBA operon, truncated forms of the DNA region upstream of the operon containing 537, 258, and 88 bp from the ATG codon were amplified with primers Prom2_rlv_3442-44_BamHI_F/Prom5_rlv_3442-44_XbaI_R, Prom3_rlv_3442-44_BamHI_F/Prom5_rlv_3442-44_XbaI_R, and Prom4_rlv_3442-44_BamHI_F/Prom5_rlv_3442-44_XbaI_R, respectively. PCR products were cloned separately into pBlueScript-II KS+ plasmid, sequenced, and moved to pLMB51 plasmid as a BamHI-XbaI restriction fragment, rendering pLMB537.ZniCBA, pLMB258.ZniCBA, and pLMB88.ZniCBA, respectively.
For the construction of pBBRZniA, the DNA region upstream of the zniC gene and the zniA gene from its 16 bp upstream sequence were amplified with primers Prom3_rlv_3442-44_BamHI_F/P2_Com_rlv_3444_R and P3_Com_rlv_3444_F/P4_Com_rlv_3444_XbaI_R, respectively, and UPM791 genomic DNA as a template. The two PCRs were linked by an overlapping GC-rich sequence through a fusion PCR using primers Prom3_rlv_3442-44_BamHI_F/P4_Com_rlv_3444_R. The same procedure was performed for the construction of pBBRZniAST, but the P4_Com_rlv_3444Strep_XbaI_R, including the sequence coding for StrepTag II peptide (WSHPQFEK) for in-frame fusion of the tag sequence to the 3´end of the gene, was used. To generate the pBBRZnuA plasmid, DNA covering the znuA gene and its promoter region was PCR-amplified with Prom_ZnuA_BamHI_F/Com_ZnuA_XbaI_R primers. The PCR products obtained were cloned separately in the pBlueScript-II KS+ vector as BamHI-XbaI restriction fragments, sequenced, and moved to the broad host range vector pBBR1MCS-2 (Kovach et al., 1995). Site-directed mutagenesis of the zniAST gene was performed in pBBRZniAST using complementary oligonucleotides with adequate centered nucleotide substitutions (Supplementary Table S2) and DpnI digestion to eliminate the native template. Substitution of target nucleotides as well as the absence of additional mutations was verified by sequencing. The pLMBcZni plasmid was generated by amplifying by PCR the zniCBA operon and its promoter region using primers Prom1_rlv_3442-44_BamHI_F and rlv_3442-44_XbaI_R, and UMP791 genomic DNA as template. The PCR product was cloned in the pBluescript-II KS+ plasmid, sequenced, and then moved as a BamHI-XbaI fragment to the pLMB51 plasmid.
Plant assays
Pea (Pisum sativum L. cv. Frisson) and lentil (Lens culinaris cv. Magda) were surface-sterilized, pregerminated, planted in Leonard jar assemblies under bacteriologically controlled conditions, and inoculated with 1 ml of early stationary phase bacterial cultures grown in YMB as previously described (Sánchez-Cañizares et al., 2011). Plants were grown in a greenhouse with a standard nitrogen-free plant nutrient solution (Brito et al., 1994). When required, the nutrient solution was supplemented with ZnSO4 at the desired concentration. Plants were grown at 18/25°C night/day with 16/8 h of light/dark photoperiod and harvested 21 days (pea) or 28 days (lentil) after inoculation. Shoots were collected and dried (60°C for 48 h), and shoot dry weight was determined. Shoot total nitrogen content was measured using a TruMac C/N analyzer (Leco Corporation).
β-Glucuronidase activity assays
β-Glucuronidase activity assays were performed with free-living cells and bacteroids as described in Lodwig et al. (2004) using X-GlcA (5-bromo-4-chloro-3-indolyl-β-D-glucuronide) as substrate. Values of activity were calculated as OD420 min−1 (mg protein)−1 and expressed as Miller units. Bacteroids were obtained from the nodules as previously described (Cacho et al., 2010). Protein content was obtained following the bicinchoninic acid method (Smith et al., 1985) after alkaline digestion of cells at 90°C in 2 N NaOH for 10 min using bovine serum albumin as the standard.
RNA extraction and qRT-PCR
RNA was extracted from nodule samples as described in Ballesteros-Gutiérrez et al. (2023). RNA concentration was quantified with a NanoDrop spectrophotometer and tested for possible DNA contamination by PCR using the RNA samples as templates and primers specific for rpoD (rpoD_qPCR_F/rpoD_qPCR_R). RNA integrity was confirmed by electrophoresis in a 1% agarose gel. cDNA was synthesized from 500 ng of DNA-free RNA using the PrimeScript RT reagent kit (Takara, Saint-Germain-en-Laye, France), supplemented with RNase out (Invitrogen, Thermo Fisher Scientific, Massachusetts, USA) following the manufacturer’s specifications. Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) was performed with Power SyBR Green master mix (Applied Biosystems) and primers rlv_3444_qPCR_F/rlv_3444_qPCR_R for zniA, znuA_qPCR_F/znuA_qPCR_R for znuA, hupL_qPCR_F/hupL_qPCR_R for hupL, and rpoD_qPCR_F/rpoD_qPCR_R for rpoD genes. Gene expression levels were normalized to the rpoD reference gene (Ballesteros-Gutiérrez et al., 2023). Determinations were performed with RNA extracted from three independent biological samples, with the threshold cycle (CT) determined in triplicate. The relative levels of transcription were calculated using the 2–ΔΔCT method (Livak and Schmittgen, 2001).
Bioinformatic analysis
Protein sequences were obtained from the GenBank database. Multiple sequence alignment was created with SnapGene 6.2 software using the Clustal Omega multiple alignment algorithm v.1.2.4 (Chenna et al., 2003). The phylogenetic tree of the protein alignment was analyzed with MEGA 12 (Tamura et al., 2021) using a neighbor-joining algorithm method (Saitou and Nei, 1987) with bootstrapping (1,000 iterations). The Uniprot database (https://www.uniprot.org/) was used to perform protein BLAST and amino acid sequence identity analysis. Zur box prediction from DNA upstream of zniCBA and znuA genes were generated based on the conserved sequence motifs using the bioinformatics tool Multiple EM for Motif Elicitation (MEME) (Bailey et al., 2006). The analysis included the 42 Zur-binding motifs from Rhizobiales obtained from the Regprecise database of the curated inferences of regulons in prokaryotic genomes (Novichkov et al., 2009). Structural prediction of the mature RLV_3444 protein (25–299 residues) was carried out using the AlphaFold 3 server (Abramson et al., 2024). Structural comparison with the PDB database was performed at the DALI server (http://ekhidna2.biocenter.helsinki.fi/dali/) (Holm et al., 2023). The program was run with the default server parameters. Alignments for further data analysis and figure preparation were done in Pymol (v.3.1.3.1, Schrödinger). Published protein structures were obtained from the PDB database.
Statistical analysis
Statistical analysis was performed by analysis of variance (ANOVA) linear model test following completely random design. Multiple comparisons of means were analyzed as indicated in the corresponding legend figure. Statistical analysis and graph generation were performed with GraphPad Prism 8 (GraphPad Software).
Results
Identification of two putative zinc ABC transporter systems in R. leguminosarum bv. viciae UPM791 genome
Inspection of the Rlv UPM791 genome reveals the presence of a chromosomic operon (rlv_5427-5425) encoding the putative high-affinity zinc transporter system ZnuABC. The znuA (rlv_5427) gene is oriented in the opposite direction to the znuC (rlv_5426) and znuB (rlv_5425) genes (Figure 1A). Rlv ZnuA shares a high amino acid sequence identity with its homologues in S. fredii, A. tumefaciens and S. meliloti (identities of 69.1%, 67.31%, and 66.56%, respectively) (Supplementary Figure S1). A Zur-like transcriptional regulator is encoded by gene rlv_ 5424 located immediately downstream of the znuB gene (Figure 1A). RLV_5424 is very similar to Zur from A. tumefaciens C58, S. fredii CCBAU 45436 and S. meliloti 1021 strains (74.24%, 72.52%, and 70.23% sequence identities, respectively). A Zur box containing a 15 bp sequence was identified upstream znuA gene, very similar to the conserved Zur motifs of Rhizobiales, localized 41 bp upstream of the translational start site of the gene (Figure 1B).
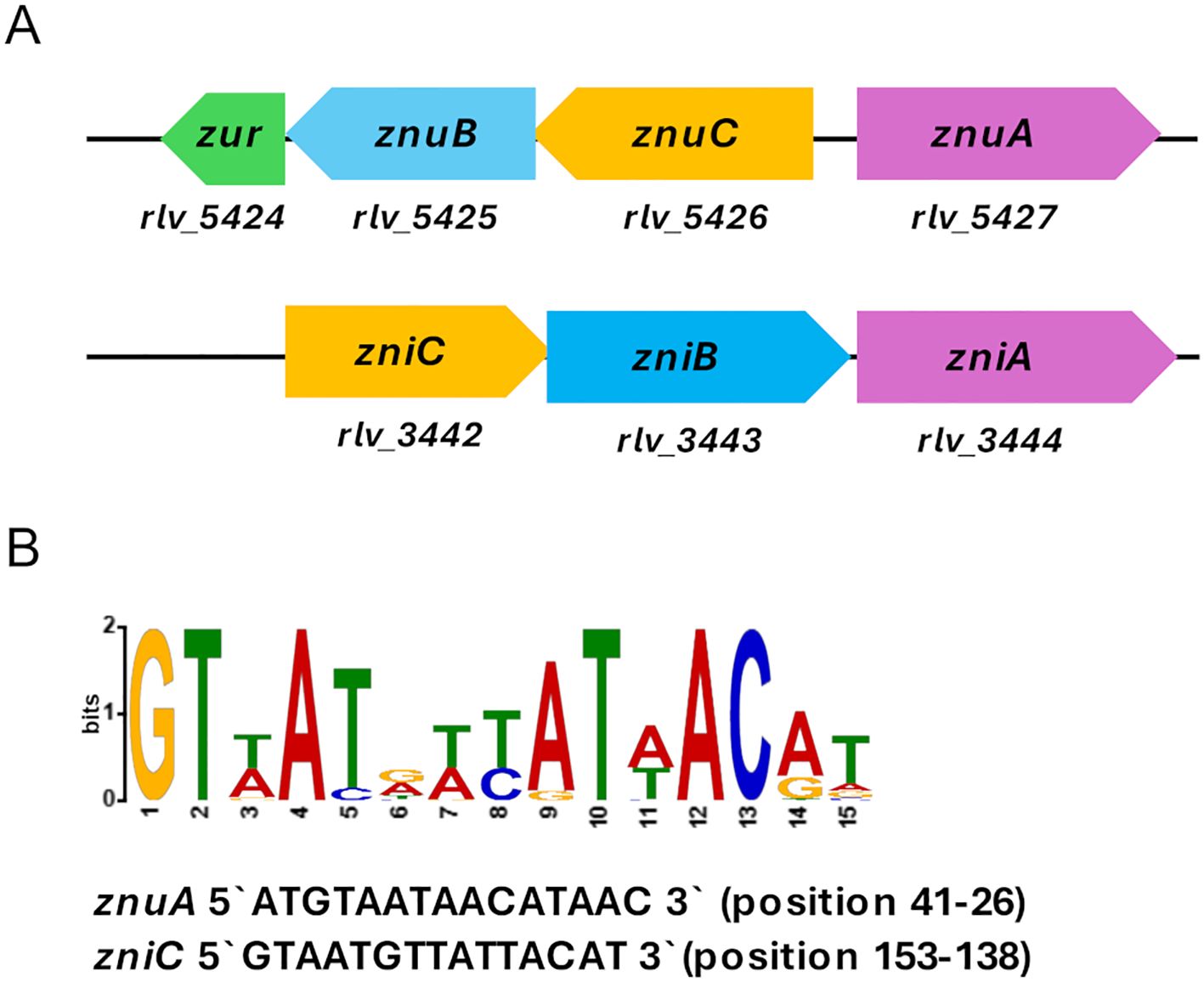
Figure 1. Genomic context of znuA and zniA genes in R. leguminosarum bv. viciae UPM791. (A) Gene designations of putative znuABC and zniCBA operons, and zur gene in Rlv UPM791 genome are shown below the genetic map. (B) The Zur consensus motif determined by comparison of promoter DNA from Rhizobiales and Rlv UPM791 znuA and zniC genes using MEME. More prominent letters denote more frequent usage in the motif. The sequences of Zur boxes and the position indicated as bp upstream of the translational start sites in the promoter regions of znuA and zniC genes are indicated.
The Rlv UPM791 genome encodes a metal ABC transporter system containing the host-dependent metal-binding protein RLV_3444 (Durán et al., 2021). This protein is encoded by the chromosomic rlv_3444 gene, which is part of an operon with a typical organization of bacterial ABC permeases and also encodes an ATP-binding protein (rlv_3442) and a permease (rlv_3443) (Figure 1A). In addition, a conserved Zur motif was also identified 153 bp upstream of the rlv_3442 gene. A comparison of the RLV_3444 amino acid sequence with protein databases revealed a highly significant level of conservation (over 75% identity) with proteins annotated as metal, Zn, or Zn/Mn ABC transporter SBPs in the genomes of several members of Rhizobiaceae. Furthermore, RLV_3444 exhibits a high amino acid sequence identity with the functionally characterized SBPs from K. pneumoniae ZniA (53.42% sequence identity), A. tumefaciens TroA (41.61% identity) and Paracoccus denitrificans AztC (40.88% identity), previously shown to participate in zinc uptake (Chaoprasid et al., 2016; Handali et al., 2015; Maunders et al., 2024) (Supplementary Figure S1). Sequence alignments revealed that RLV_3444 contains the three fully conserved histidine residues (H62, H127, and H193, RLV_3444 numbering) required for the coordination of a zinc atom in the crystal structures of K. pneumoniae ZniA and P. denitrificans AztC (Supplementary Figure S2). In RLV_3444, a glutamate at position 268, conserved in K. pneumoniae ZniA, where it participates in zinc coordination (Maunders et al., 2024), was also identified. In contrast, the short central histidine-rich domain present in P. denitrificans AztC and A. tumefaciens TroA and associated to Zn2+ specificity (Blindauer, 2015) is not present either in RLV_3444 or in K. pneumoniae ZniA.
Structural prediction of mature RLV_3444 protein (25–299 residues) was carried out using the AlphaFold 3 server, and the model obtained was compared to the available structures in the PDB database using the Dali server. Proteins showing the best structural homologies with RLV_3444 were zinc-, manganese-, and iron-binding proteins, with the highest scores for K. pneumoniae ZniA (PDB id: 8SVC; Maunders et al., 2024) and P. denitrificans AztC (PDB id: 5W57; Neupane et al., 2017) (Supplementary Table S3), suggesting similar functions for these proteins. Structural alignment of RLV_3444 with AztC and ZniA has revealed that it shares the two independent domains that interact to create a metal-binding pocket between them connected by a rigid α-helix characteristic of the cluster A SBPs-associated family (Berntsson et al., 2010) (Figure 2A). Structural alignments have also shown a highly conserved metal binding site (Figure 2B).
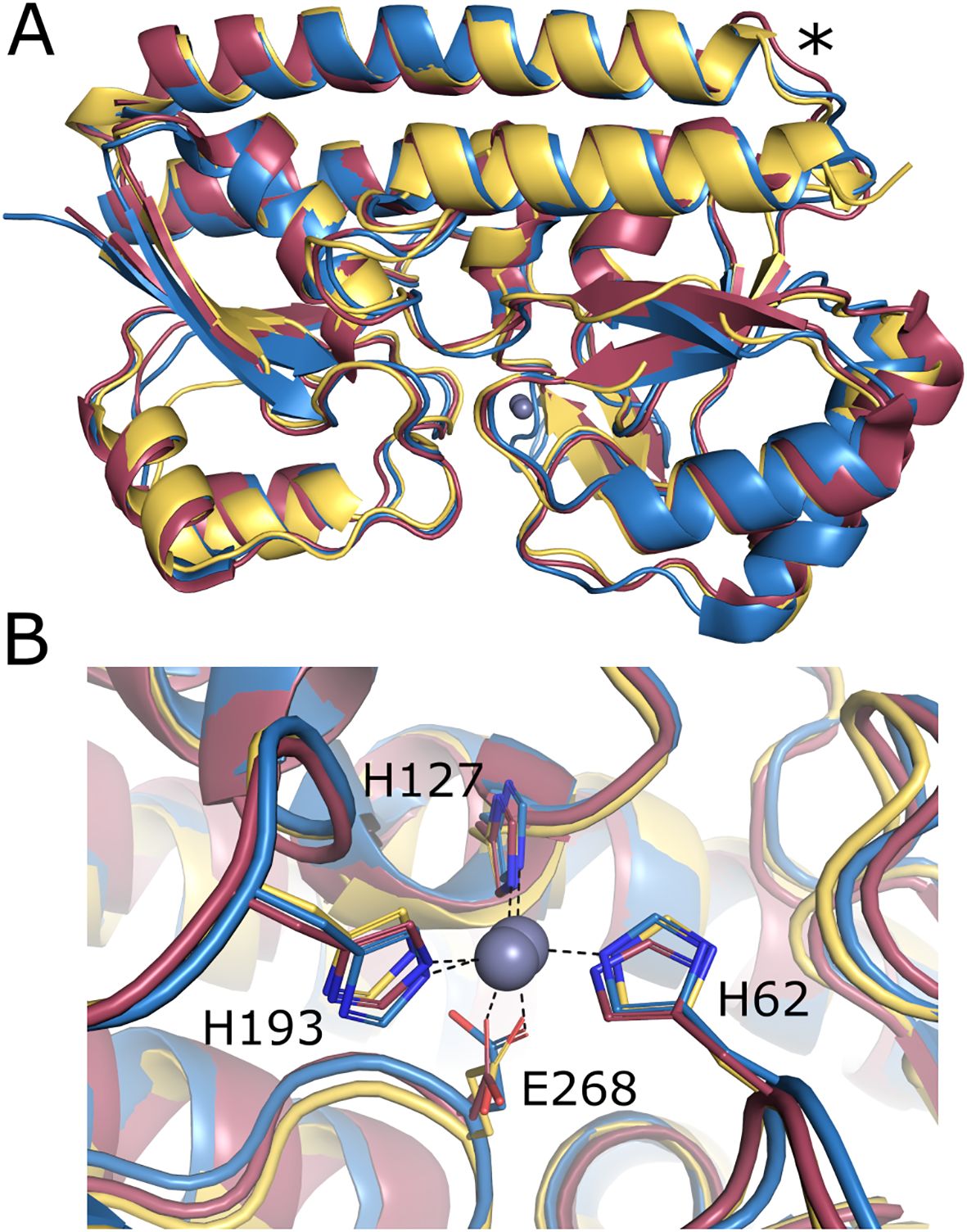
Figure 2. In-silico modelled structure of R. leguminosarum ZniA. (A) Superposition of the Alphafold model structure of R. leguminosarum ZniA (yellow, residues 25-299) and the crystal structure of K. pneumoniae ZniA (red, PDB: 8SVC) and P. denitrificans AztC (blue, PDB: 5W57). The cluster A rigid α-helix is marked with an asterisk. (B) Detail for metal binding site of R. leguminosarum ZniA (yellow) aligned with K. pneumoniae ZniA (red, PDB: 8SVC) and P. denitrificans AztC (blue, PDB: 5W57). The zinc atom is shown in grey.
Taken together, these results suggest that RLV_3444 is a potential zinc-binding protein of an ABC transporter system. Based on the bioinformatic analysis and the functional and regulatory role of this protein (see below), the corresponding gene was designated as zniA (zinc import), and we also designated the rlv_3442–3444 operon as zniCBA (Figure 1).
ZniCBA replaces the functional role of ZnuA in R. leguminosarum free-living cells under zinc-limiting conditions
In order to investigate the functional role of ZniA under free-living conditions, growth curves of SPF25 and its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) strains were determined in UMS medium with no zinc added. The results indicated that all bacterial strains exhibited similar growth under these zinc-depleted conditions (Figure 3A). Atomic absorption spectromety analysis of zinc in the “Zn-free” medium revealed the presence of ca. 1 µM Zn2+ traces, likely too high to induce real zinc deprivation conditions (Outten and O’Halloran, 2001), so we added EDTA (50 μM) to chelate zinc cations. In the presence of EDTA, the growth of znuA- and zniA-deleted single mutants was similar to that observed in the wild-type strain. Conversely, the bacterial strain carrying deletions in both znuA and zniA genes (UPM1630) showed a clearly defective growth phenotype (Figure 3A). This phenotype was fully reverted by the expression of znuA or zniA genes in pBBRZnuA or pBBRZniA plasmids, respectively. Furthermore, the phenotype of the double mutant was also reverted by supplementing the medium with Zn2+ (50 µM ZnSO4) indicating that the defective phenotype of UPM1630 was due to its inability to take up zinc (Figure 3B). Supplementation of EDTA-containing medium with other metals (Mn2+ and Fe2+) did not restore the growth of the znuA/zniA-deficient derivative UPM1630 (Figure 3B), indicating that the growth defect was specifically related to a zinc deficiency. Altogether, these data indicate that ZniA plays a functional role similar to that of ZnuA, both proteins being able to support wild-type levels of growth under zinc-limiting conditions in R. leguminosarum free-living cells.
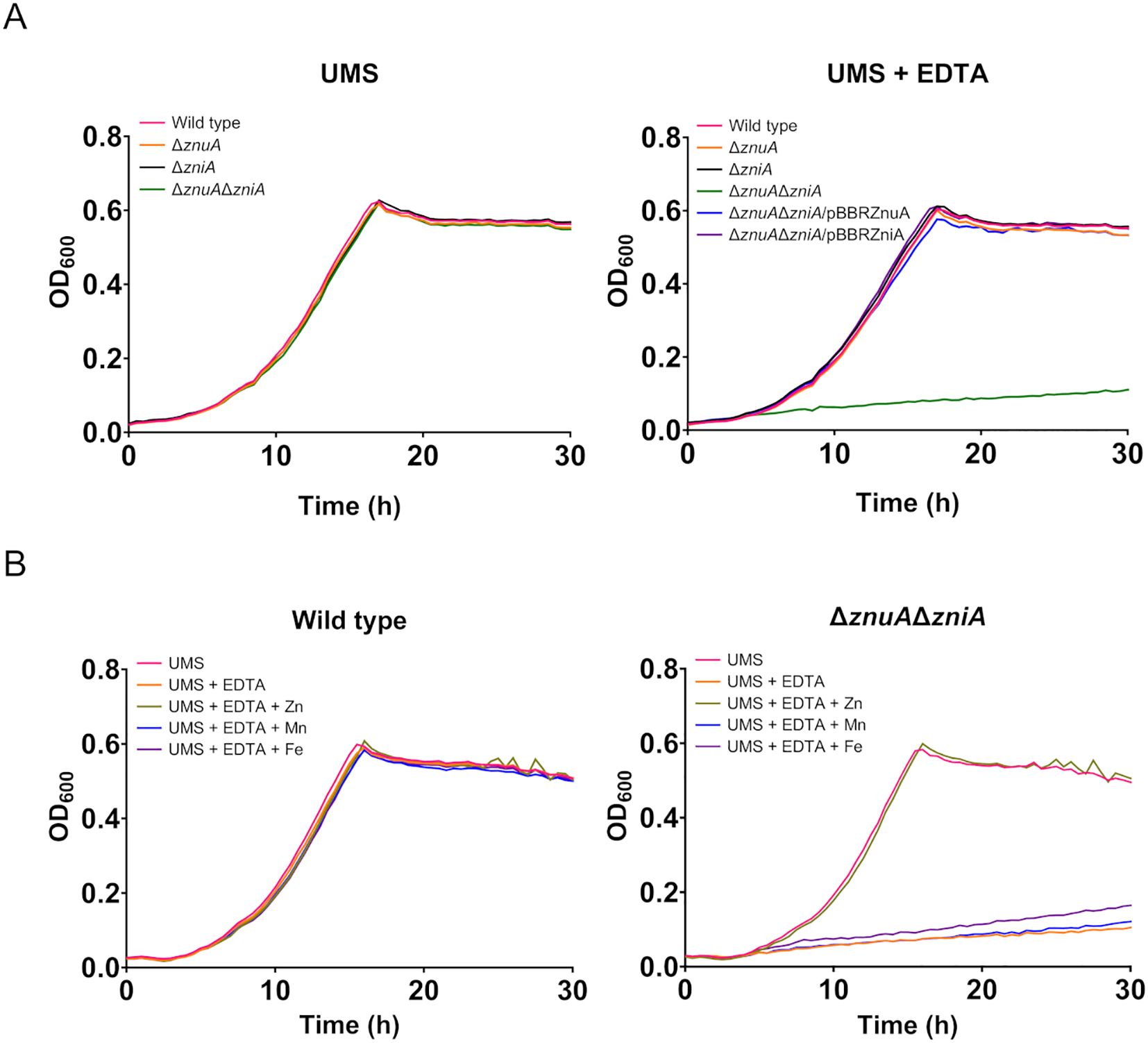
Figure 3. Growth curves of Rlv strains under metal-limiting conditions. (A) Effect of ZnuA and ZniA on Rlv SPF25 growth. Rlv strains were grown until the stationary phase in zinc-depleted (no zinc added) UMS medium (UMS) or in the same medium supplemented with EDTA (50 µM EDTA; UMS+EDTA). (B) Effect of metal supplementation on the growth of Rlv strains. Rlv strains were grown until the stationary phase in zinc-depleted (no zinc added) UMS medium (UMS), in EDTA-chelated medium (no zinc added, 50 µM EDTA; UMS+EDTA) or in the same medium supplemented with 50 µM of either ZnSO4, MnSO4 or FeSO4, as indicated. Each OD determination represents the mean of three replicates. All standard errors were below 10%. Strains: wild type (SPF25, pBBR), ΔznuA (UPM1628, pBBR), ΔzniA (UPM1629, pBBR), ΔznuAΔzniA (UPM1630, pBBR). znuA and zniA were provided cloned in pBBR1MCS-2 as indicated (pBBRZnuA and pBBRZniA plasmids, respectively). pBBR: pBBR1MCS-2.
ZniA and ZnuA are not interchangeable for providing zinc to Znu permease
With the aim to consider the possibility of cross-talk between Zni and Znu systems via their respective SBPs, we constructed a mutant (UPM1631) carrying a deletion of the whole zniCBA operon. This mutant showed normal growth under all conditions tested, as expected from the presence of an active Znu system (Figure 4). These results indicate that ZniCBA and ZnuABC were functionally redundant under metal-limitation conditions in the presence of EDTA (0 µM ZnSO4, 50 µM EDTA). In order to check whether ZniA could be providing Zn2+ to the bacterial cell via the Znu permease in the absence of ZnuA, a znuA deletion was generated in this zniCBA mutant, thus resulting in the UPM1632 strain. The growth of the UPM1632 double mutant was similar to that observed in the wild-type strain in UMS under zinc-depleted conditions (0 µM ZnSO4 added), but it was impaired in the presence of EDTA. The defective phenotype was reverted back to wild-type levels by supplementing the medium with ZnSO4 (50 µM) and also by the expression of the znuA gene from the pBBRZnuA plasmid. In contrast, such complementation was not induced by the expression of zniA from pBBRZniA. As a control, we checked that the UPM1632 defective growth phenotype under metal-chelated conditions was rescued to wild-type levels with the expression of ZniCBA under the control of its own promoter in the pLMBcZni plasmid (Supplementary Figure S3). These data demonstrate that ZnuA function cannot be substituted by ZniA when Zn provision depends on Znu permease.
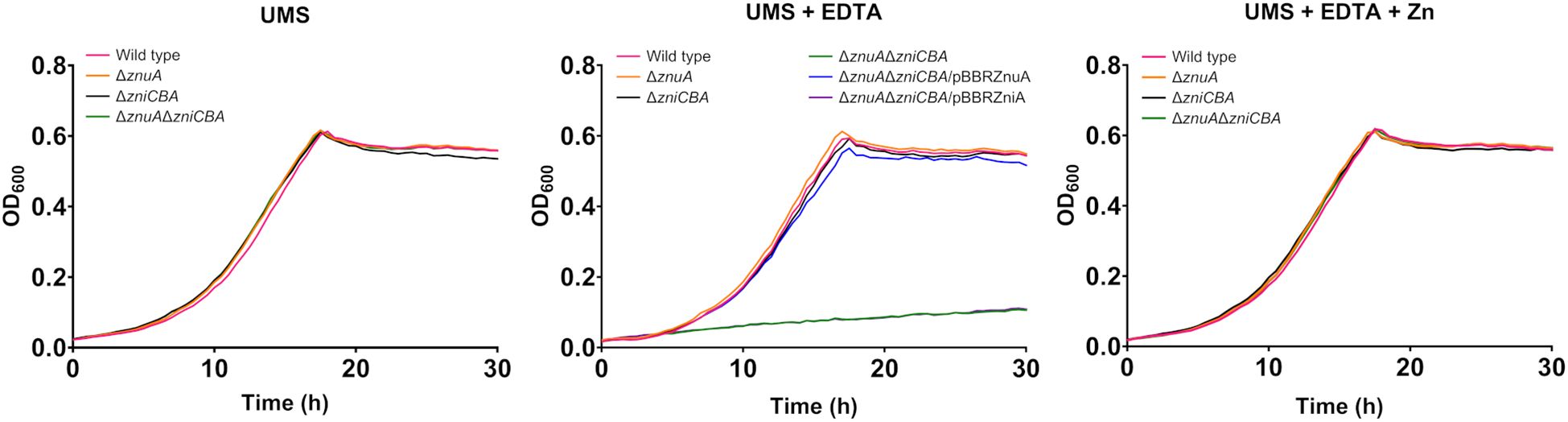
Figure 4. Effect of ZniA expression on the complementation of ZnuA under metal limiting conditions. Rlv strains were grown until the stationary phase in zinc-depleted (no zinc added) UMS medium (UMS), in EDTA-chelated medium (no zinc added, 50 µM EDTA; UMS+EDTA) or in the same medium supplemented with 50 µM ZnSO4 (UMS+EDTA+Zn). Each OD determination represents the mean of three replicates. All standard errors were below 10%. Strains: wild type (SPF25, pBBR), ΔznuA (UPM1628, pBBR), ΔzniCBA (UPM1631, pBBR), ΔznuAΔzniCBA (UPM1632, pBBR). znuA and zniA were provided cloned in pBBR1MCS-2 as indicated (pBBRZnuA and pBBRZniA plasmids, respectively). pBBR: pBBR1MCS-2.
Functional analysis of ZniA residues potentially involved in metal coordination
The potential involvement of ZniA histidine residues (H62, H127, and H193) in ZniA function has been investigated. To carry out these studies, plasmids expressing ZniAST, incorporating a C-terminal Strep-Tag affinity tail for future monitoring and purification purposes, and variants harboring H62A, H127A, and H193A changes were generated and transferred separately by conjugation into the znuA/zniA-deleted strain UPM1630. Bacterial cultures of the corresponding transconjugant strains were analyzed for growth in UMS medium under zinc-depleted (0 µM ZnSO4), metal-chelated (0 µM ZnSO4, 50 µM EDTA), and zinc-repleted (50 µM ZnSO4, 50 µM EDTA) conditions. The growth of all strains was similar to that observed in the SPF25 strain under zinc-depleted conditions (Figure 5). The expression of ZniAST in pBBRZniAST complemented the defective growth phenotype of the UPM1630 strain up to wild-type levels under zinc-chelated conditions, indicating that the incorporation of the affinity tail did not affect the function of the protein. In these experiments, we observed that zniAST carrying the mutations H62A, H127A, or H193A did not complement the ΔzniAΔznuA mutant for growth in the medium containing EDTA (0 µM ZnSO4, 50 µM EDTA). The defective growth associated with the mutation in the histidine residues was restored with the addition of zinc to the EDTA-chelated medium. These results suggest that these histidine residues are critical for ZniA function, likely by contributing to metal coordination along the transport process.
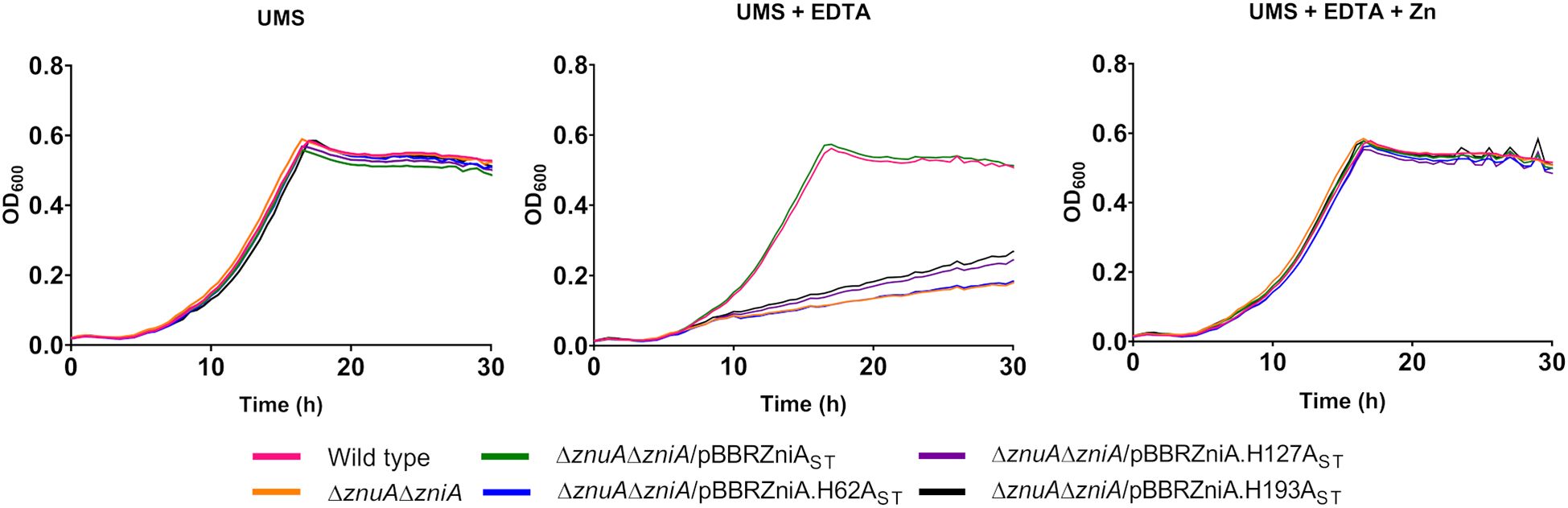
Figure 5. Functional analysis of conserved histidine residues from Rlv ZniA. The indicated Rlv strain complemented with plasmid-based zniA or with zniA versions with the indicated alterations in histidine residues were grown until stationary phase in zinc-depleted (no zinc added) UMS medium (UMS), in EDTA-chelated medium (no zinc added, 50 µM EDTA; UMS+EDTA) or in the same medium supplemented with 50 µM ZnSO4 (UMS+EDTA+Zn). Each OD determination represents the mean of three replicates. All standard errors were below 10%. Strains: wild type (SPF25, pBBR), ΔznuAΔzniA (UPM1630, pBBR). pBBR: pBBR1MCS-2.
Rlv ZniA and ZnuA contribute to symbiotic performance in pea and lentil plants
The metal-binding protein ZniA was shown to be overexpressed in pea bacteroids, with protein expression levels ca. twofold higher in pea than in lentil bacteroids (Durán et al., 2021). Our proteomic analysis also revealed that ZnuA was detected in bacteroids from pea but not from lentil (Durán et al., 2021). Furthermore, previous studies revealed a host-dependent contribution of ZnuA in S. fredii-soybean system (Jiao et al., 2018; Zhang et al., 2020). This prompted us to study the potential requirement for ZniA and ZnuA for efficient symbiosis of Rlv with pea and lentil plants. To carry out these studies, plants inoculated with Rlv SPF25 and its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) strains were grown in a standard N-free nutrient solution (0.8 µM ZnSO4) or in the same solution containing 80 µM ZnSO4. Symbiotic performance of znuA- and zniA-deleted strains was similar to that of the wild-type strain in both legumes under either standard- or high-zinc concentrations (Figure 6). In contrast, pea and lentil plants inoculated with znuA/zniA double mutant UPM1630 accumulated significantly less shoot dry weight and less nitrogen in comparison to the wild-type strain at both zinc concentrations tested. This result indicates that high-affinity zinc transport is essential for optimal development of symbiosis of R. leguminosarum with legume plants. The impaired phenotype of the UPM1630 strain was complemented by the overexpression of wild-type copies of ZniA or ZnuA in pBBRZniA or pBBRZnuA plasmids, respectively (Figure 6). In contrast, the observed impairment of plant growth in the double mutant was not corrected by the addition of additional Zn2+ to the nutrient solution (80 μM ZnSO4, 100-fold higher than the original concentration). This observation indicates that, in the absence of functional Znu and Zni systems, potential alternative metal transporters do not provide the bacteroids with enough zinc for the symbiotic performance of this strain. These results indicate that both ZnuABC and ZniCBA transporter systems play interchangeable, essential functional roles in the provision of Zn2+ to the bacteroids under symbiotic conditions.
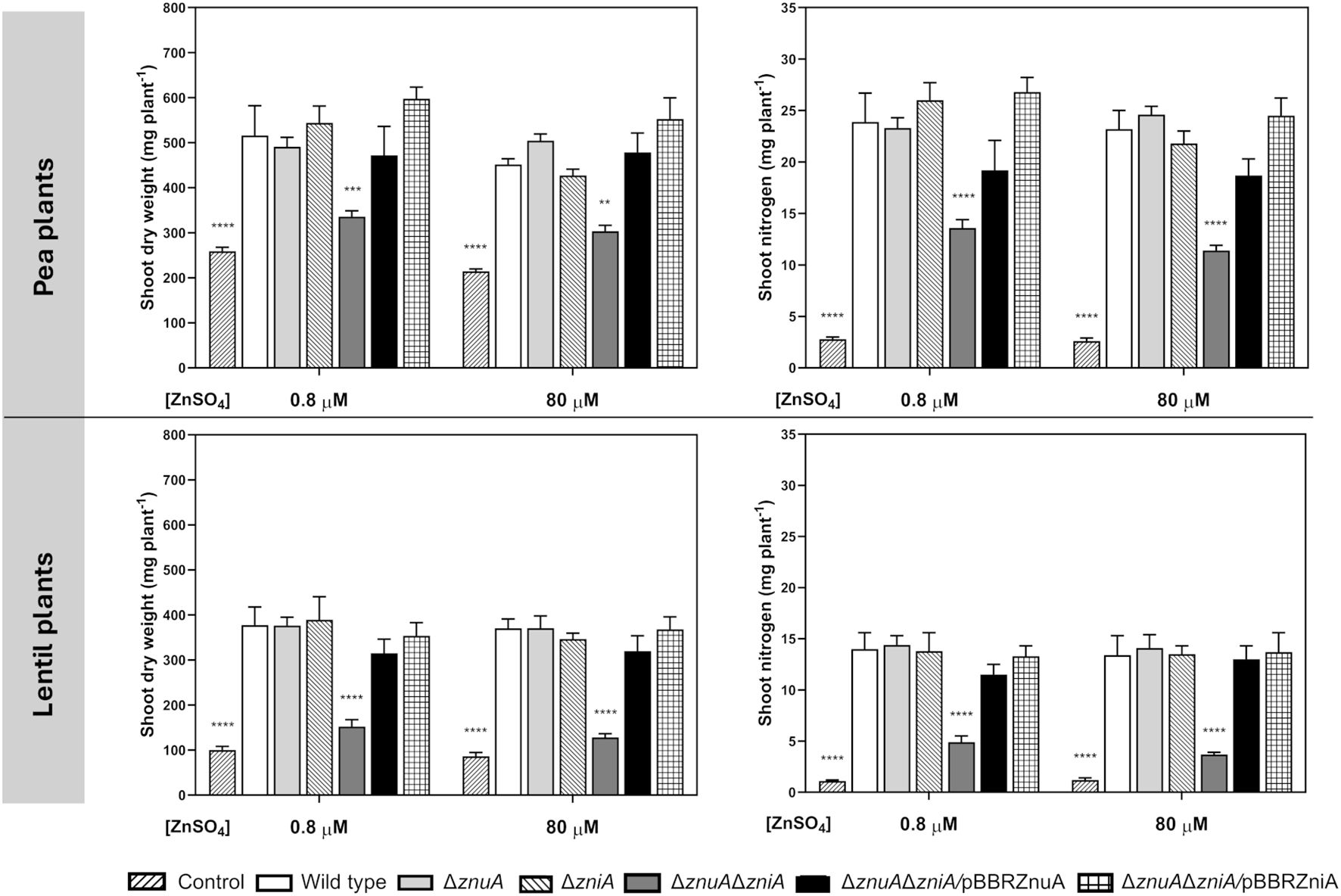
Figure 6. Effect of ZnuA and ZniA on the symbiotic performance of Rlv SPF25 with pea and lentil as host plants. Histograms represent the shoot dry weight and shoot nitrogen content of pea and lentil plants inoculated with the indicated Rlv strains. Plants were grown in a N-free standard nutrient solution (0.8 µM ZnSO4) or in the same solution containing 80 µM ZnSO4 as specified. Bars are the mean of four replicates ± standard error. Data were analyzed by one-way ANOVA and Dunnett´s test for multiple comparisons of means with wild type strain. **P < 0.01, ***P < 0.001, ****P < 0.0001. Control: uninoculated plants. Strains: wild type (SPF25), ΔznuA (UPM1628), ΔzniA (UPM1629), ΔznuAΔzniA (UPM1630), ΔznuAΔzniA/pBBRZnuA (UPM1630, pBBRZnuA), ΔznuAΔzniA/pBBRZniA (UPM1630, pBBRZniA).
Analysis of zniCBA and znuA genes expression under free-living conditions
In order to investigate the regulatory mechanisms governing the expression of zni and znu genes, independent transcriptional fusions to DNA upstream from the zniCBA operon and from the znuA gene to a promoterless gusA reporter gene in the pLMB51 plasmid were constructed, thus generating plasmids pLMBZniCBA (zniCBA´-gusA) and pLMBZnuA (znuA´-gusA). These plasmids were transferred separately by conjugation to SPF25 and to its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) strains.
We first analyzed zni gene expression under free-living conditions. To this aim, reporter activity was determined in bacterial cultures grown in UMS medium or in the same medium supplemented with EDTA (50 µM). In SPF25 and UPM1629 (ΔzniA) strains harboring the zniCBA´-gusA reporter fusion, only basal levels of enzymatic activity (ca. 20 units) were observed when cells were grown in UMS medium (Figure 7). In contrast, significant levels of activity were reported in both strains under metal-chelated conditions. Interestingly, under the same conditions, reporter activity increased in the znuA-deficient UPM1628 strain (ca. 8-fold over basal levels); finally, the expression of the reporter enzyme was highly induced (ca. 60-fold over basal levels) in the znuA/zniA-deficient UPM1630 strain. A similar pattern of results was observed when analyzing expression of the znuA gene through the use of znuA´-gusA fusion (Figure 7). In this case, the level of reporter activity was always higher than in the case of zniCBA´-gusA fusion, suggesting that ZnuA is the major transporter supporting Zn provision to the bacterium. In all cases, enzymatic activities associated with either reporter fusion were reduced to basal levels when the metal-chelated medium was supplemented with zinc (50 µM ZnSO4) in all the genetic backgrounds. These results confirm that the expression of both transporter systems is negatively regulated by zinc and that ZniCBA functions as an auxiliary transporter system in the absence of ZnuABC.
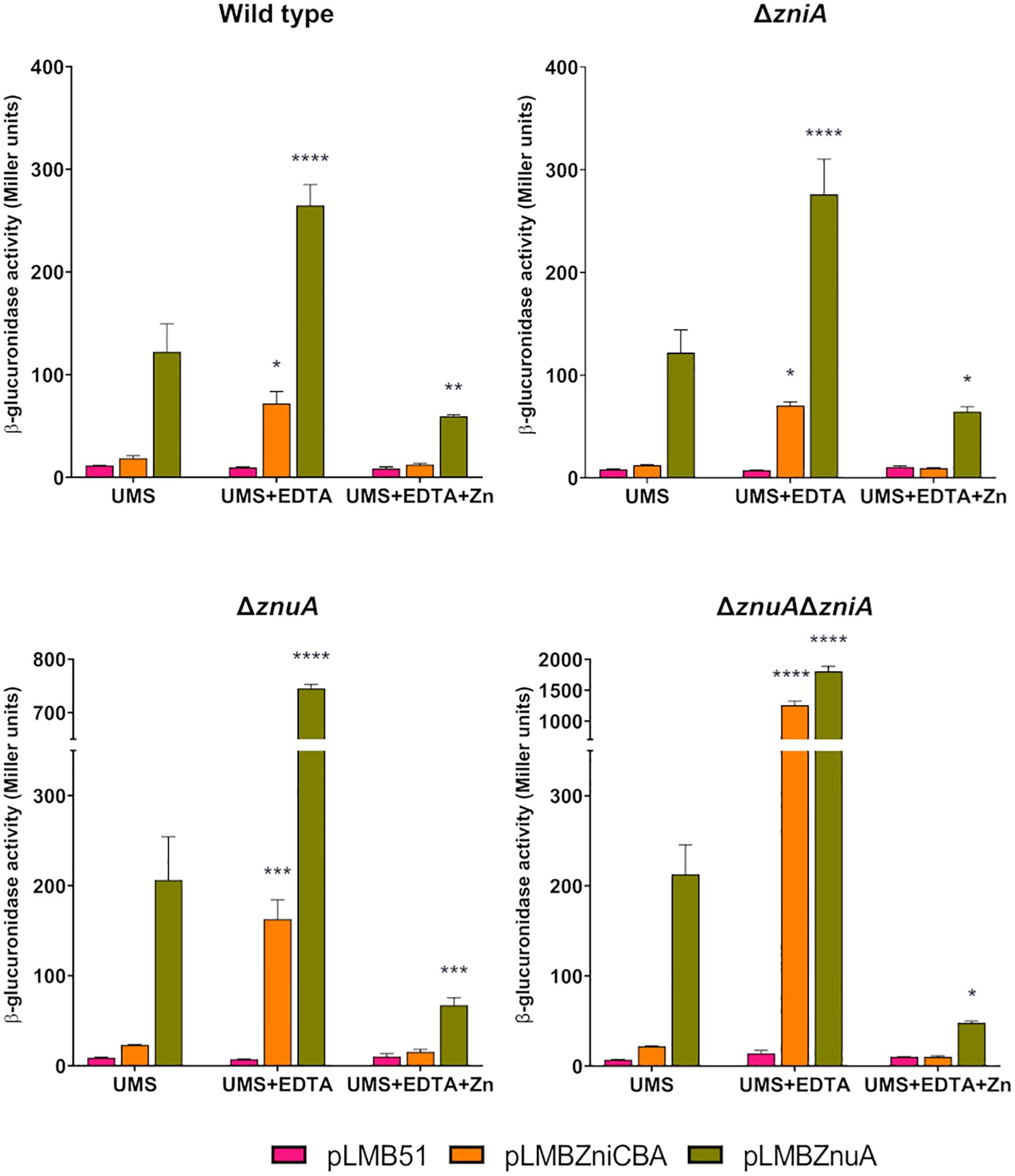
Figure 7. Expression analysis of zniCBA and znuA genes as a function of the zinc concentration in the culture medium. Rlv SPF25 (wild type) and its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) strains harboring empty pLMB51 plasmid, pLMBZniCBA or pLMBZnuA reporter fusion plasmids were grown in UMS medium (UMS), in EDTA-chelated medium (50 µM EDTA; UMS+EDTA) or in the same medium supplemented with zinc (50 µM EDTA, 50 µM ZnSO4; UMS+EDTA+Zn). β-glucuronidase activities are the mean of two independent experiments with two replicates each ± standard error. Data were analyzed by one-way ANOVA and Dunnett´s test for multiple comparisons of means comparing reporter activities associated to each plasmid to those in UMS medium. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
The identification of potential Zur-binding sites in the promoter region of the zniCBA operon and znuA gene (Figure 1) prompted us to investigate whether the expression of both systems is regulated by the transcriptional regulator Zur. To carry out these studies, pLMBZniCBA and pLMBZnuA plasmids were transferred into UPM1633 and UPM1634 strains (Δzur and ΔznuAΔzniAΔzur derivatives of SPF25 and UPM1630, respectively), and β-glucuronidase activity was determined following incubation in the different media. In these experiments, the levels of reporter activity were highly induced in all zur-deleted strains carrying pLMBZniCBA or pLMBZnuA plasmids, regardless the levels of ZnSO4 in the medium (Figure 8). These findings indicate that the zniCBA operon and znuA are negatively regulated by Zur.
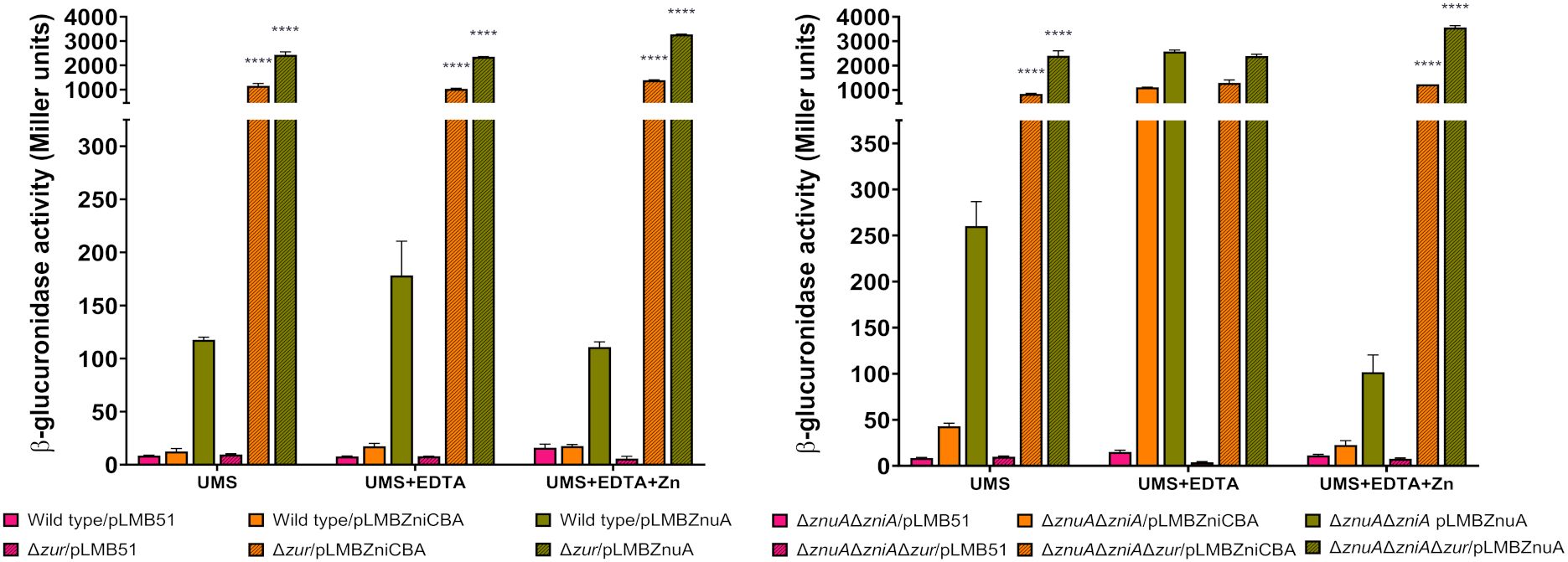
Figure 8. Analysis of Zur-dependent expression of zniCBA and znuA genes. Rlv SPF25 (wild type), UPM1630 (ΔznuAΔzniA) and its zur-mutant derivatives UPM1633 (Δzur) and UPM1634 (ΔznuAΔzniAΔzur) strains harboring empty pLMB51 plasmid, pLMBZniCBA or pLMBZnuA reporter fusion plasmids were grown in UMS medium (UMS), in EDTA-chelated medium (50 µM EDTA; UMS+EDTA) or in the same medium supplemented with zinc (50 µM EDTA, 50 µM ZnSO4; UMS+EDTA+Zn). β-glucuronidase activities are the mean of two independent experiments with two replicates each ± standard error. Data were analyzed by one-way ANOVA and Dunnett´s test for multiple comparisons of means comparing reporter activities associated to each plasmid between SPF25 or UPM1630 and its zur-deficient derivative strains at each condition assayed. ****P < 0.0001.
The metal specificity of responses of zniCBA and znuA was investigated in SPF25 and UPM1630 (ΔznuAΔzniA) strains harboring pLMBZniCBA or pLMBZnuA plasmids through experiments similar to those described above, but adding other metal cations (Mn2+, Fe2+) instead of Zn2+ to EDTA-supplemented media (Figure 9). There was no effect of the addition of these metals in the case of double mutant UPM1630, indicating that the observed effect of Zn blocking was specific for this element. Interestingly, a significant repression of β-glucuronidase activity was shown when the same experiment was carried out in the wild-type background. This observation might be associated with a displacement of zinc-chelated to EDTA by these metals, thus decreasing the effective metal chelator concentration for the complexation of zinc in the medium, as previously suggested in E. coli znu regulation studies (Patzer and Hantke, 2000). This effect was not observed in UPM1630, likely because not enough zinc to repress the expression of the system enters into the cell in the absence of both transporter systems. Furthermore, a low but significant increment of the β-glucuronidase activity associated with the pLMBZnuA reporter plasmid was observed in UPM1630 strain grown in EDTA-chelated medium supplemented with manganese and iron. These results suggest that Rlv ZnuABC and ZniCBA are inducible, specifically by zinc starvation conditions, in a Zur-dependent manner.
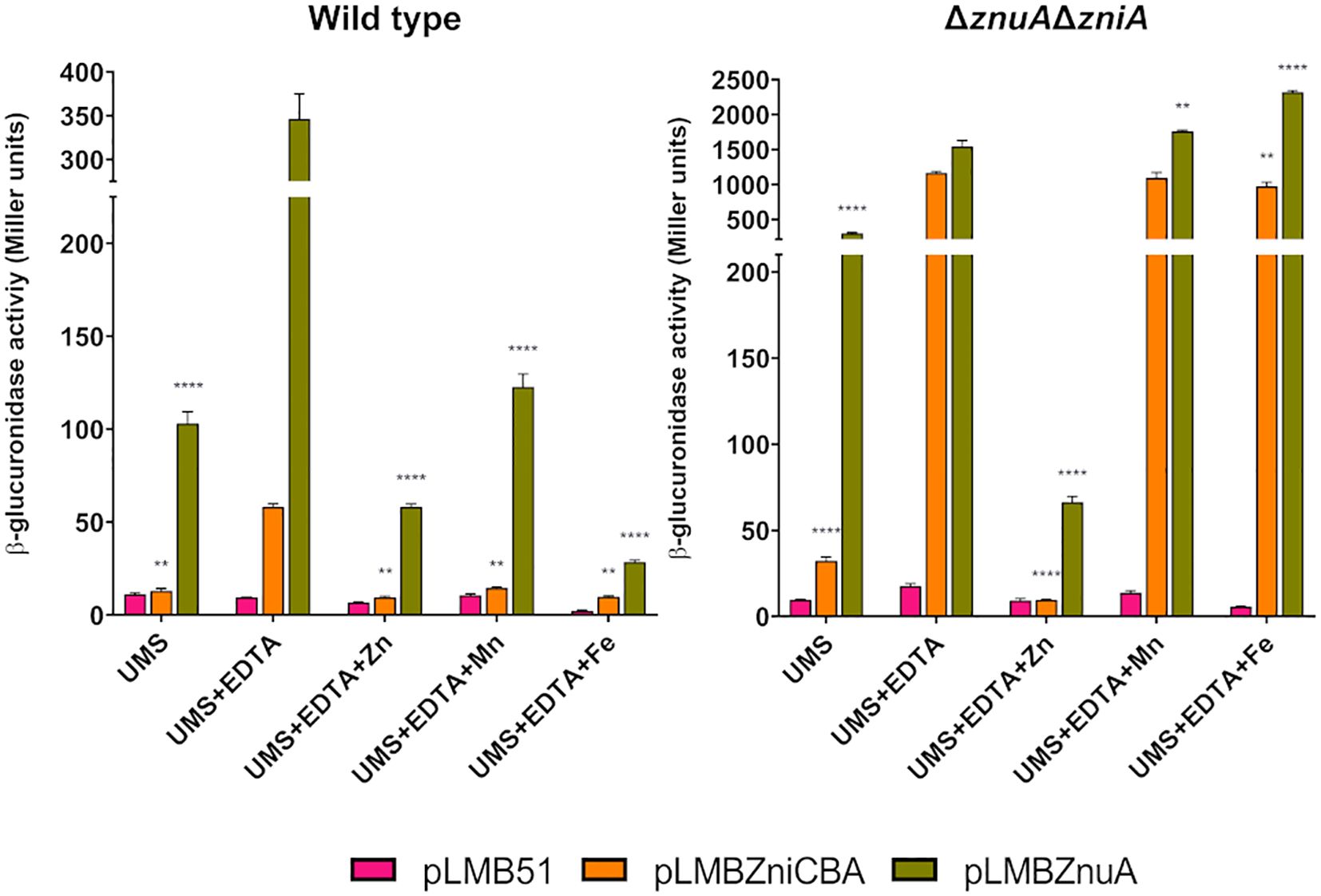
Figure 9. Effect of metal addition on zniCBA and znuA gene expression under metal-limiting conditions. Rlv SPF25 (wild type) and its derivative UPM1630 (ΔznuAΔzniA) strain harboring empty pLMB51 plasmid, pLMBZniCBA or pLMBZnuA reporter fusion plasmids were grown in UMS medium (UMS), in EDTA-chelated medium (50 µM EDTA; UMS + EDTA) or in the same medium supplemented with 50 µM ZnSO4 (UMS + EDTA + Zn), MnSO4 (UMS + EDTA + Mn), or FeSO4 (UMS + EDTA + Fe). β-glucuronidase activities are the mean of two independent experiments with two replicates each ± standard error. Data were analyzed by one-way ANOVA and Dunnett´s test for multiple comparisons of means comparing reporter activities associated to each plasmid under each condition assayed to that in UMS + EDTA condition. **P < 0.01, ****P < 0.0001.
We also wanted to better define the regulatory region controlling the expression of the zniCBA operon. To this aim, a deletion analysis of its promoter region was carried out using transcriptional fusions to DNA regions of the promoter truncated 537, 258, and 88 bp upstream of the operon (plasmids pLMB537.ZniCBA, pLMB258.ZniCBA, and pLMB88.ZniCBA, respectively). The new plasmids, together with pLMBZniCBA, were independently transferred to wild-type (SPF25) and zniA/znuA double mutant (UPM1630) strains. Reporter activities were determined in bacterial cultures grown in UMS and in the same medium supplemented with the metal chelator EDTA with or without zinc (50 µM ZnSO4). In these experiments, fusions including the 537- and 258-bp fragments of the promoter region induced reporter activities that were similar to those associated with the original fusion spanning 775 bp (Figure 10). In contrast, strains carrying the shortest fragment (88 bp) induced only basal levels of reporter activity under all the conditions assayed. These observations indicate that a relevant regulatory region of the ZniCBA transporter system is located between positions 258 and 88 pb upstream of the first ATG codon of the operon, which is consistent with the presence of the Zur box in this region (Figure 1B).
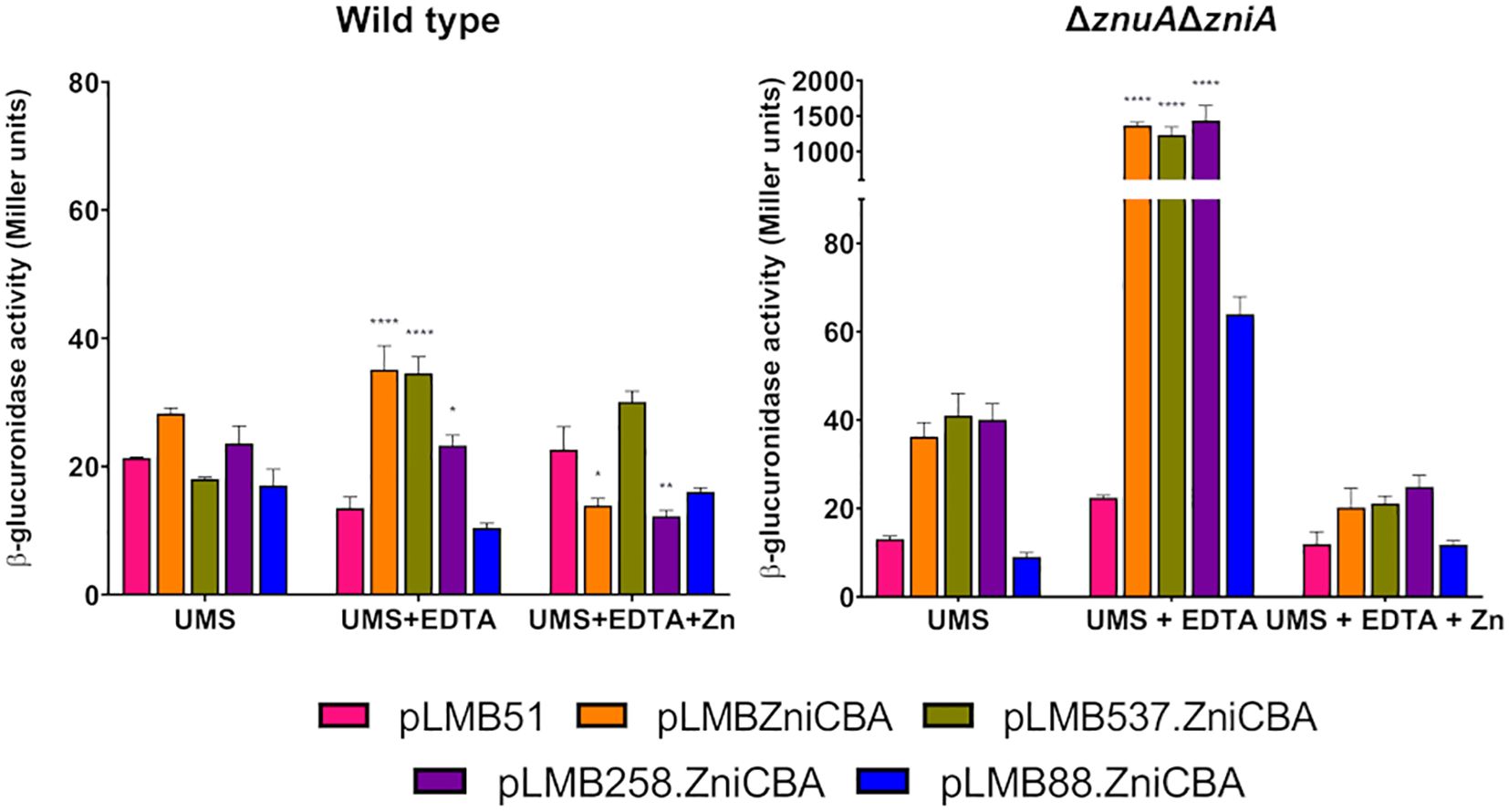
Figure 10. Deletion analysis of the promoter region of zniCBA operon under zinc-limiting conditions. Rlv SPF25 (wild type) and its derivative UPM1630 (ΔznuAΔzniA) strains harboring empty pLMB51 plasmid, pLMBZniCBA, pLMB537.ZniCBA pLMB258.ZniCBA or pLMB88.ZniCBA reporter fusion plasmids were grown in UMS medium (UMS), in EDTA-chelated medium (50 µM EDTA; UMS+EDTA) or in the same medium supplemented with 50 µM ZnSO4 (UMS+EDTA+Zn). β-glucuronidase activities are the mean of two independent experiments with two replicates each ± standard error. Data were analyzed by one-way ANOVA and Dunnett´s test for multiple comparisons of means comparing reporter activities between each reporter plasmid and empty vector pLMB51 at each condition assayed. *P < 0.05, ** P< 0.01, ****P < 0.0001.
Analysis of zniCBA and znuA gene expression under symbiotic conditions
Expression analysis of the zniCBA operon and znuA gene was also carried out under symbiotic conditions. We first analyzed by qRT-PCR whether the control of the host-dependent expression of zniA was regulated at the transcriptional level. This analysis was also extended to the znuA gene, as our proteomic analysis revealed that its encoded protein was detected in pea bacteroids but not in those developed in lentil plants (Durán et al., 2021). Quantification of the levels of transcripts revealed that both zniA and znuA genes were significantly upregulated in bacteroids from both pea and lentil (ca. 10-fold and 3-fold, respectively, for zniA and znuA) (Figure 11A). In this analysis, transcripts of the gene encoding the [NiFe] hydrogenase large structural subunit HupL, governed by a strong host-dependent control (Brito et al., 2008), exhibited expression increments of 100-fold in pea versus lentil bacteroids (20.2 ± 3.0 and 0.18 ± 0.02 relative expression levels in pea and lentil bacteroids, respectively). These data indicate that the host-dependent expression of zniA and znuA is controlled at the transcriptional level. In addition, these observations suggest that the metal provision to bacteroids might be more restrictive in pea than in lentil.
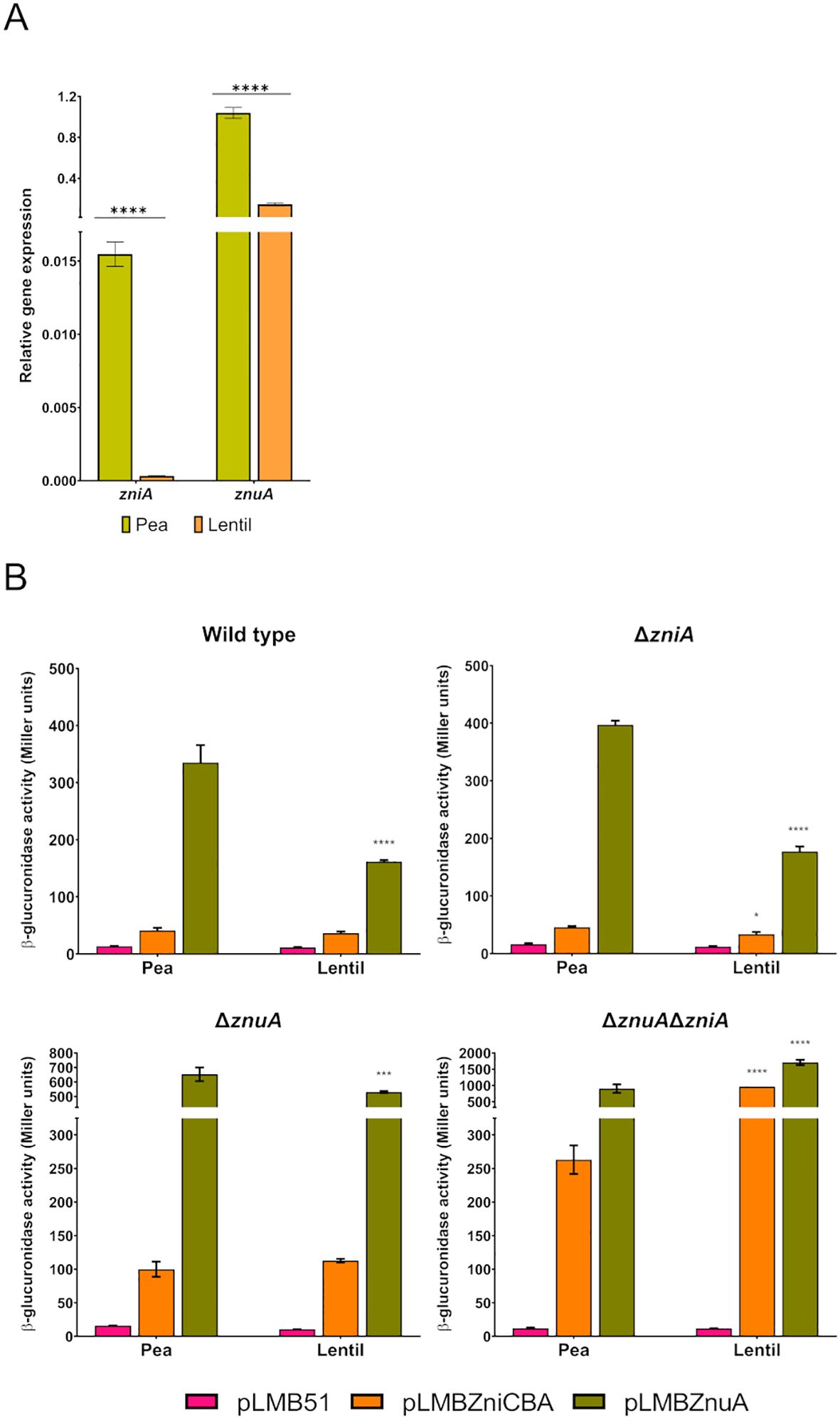
Figure 11. Symbiotic expression of zniCBA and znuA genes in pea and lentil nodules. (A) qRT-PCR analysis. Histogram shows the relative expression levels of zniA and znuA genes in bacteroids induced by Rlv SPF25 strain in pea and lentil nodules using the rpoD gene as standard for normalization. Plants were grown in a standard N-free nutrient solution (0.8 µM ZnSO4). Values are the means of three experimental replicates ± standard error. Data were analyzed by one-way analysis of variance (ANOVA) and Tukey test for multiple comparisons of means comparing expression levels between the two legume hosts. Values significantly different at P < 0.0001 (****) are indicated. (B) Reporter gene analysis of the expression of zniCBA and znuA genes. Histograms show β-glucuronidase activities of pea and lentil bacteroids induced by Rlv SPF25 (wild type) and its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) strains harboring empty pLMB51 plasmid, pLMBZniCBA or pLMBZnuA reporter fusion plasmids as indicated. Plants were grown in a standard N-free nutrient solution (0.8 µM ZnSO4). Data are the mean of two independent experiments with two replicates each ± standard error. Data were analyzed by one-way ANOVA and Tukey test for multiple comparisons of means between the two legume host associated to each plasmid in each strain. *P < 0.05, ***P < 0.001, ****P < 0.0001.
The symbiotic induction of zniA and znuA genes was also analyzed by using transcriptional fusions of zniCBA and znuA promoter regions to the gusA reporter gene in the pLMB51 plasmid. To this aim, we analyzed reporter activities in bacteroids prepared from plants grown in standard nutrient solution (0.8 µM ZnSO4) inoculated with SPF25 (wild type) and its derivatives UPM1628 (ΔznuA), UPM1629 (ΔzniA), and UPM1630 (ΔznuAΔzniA) bearing transcriptional fusions for zniCBA, znuA, or just empty plasmid. Data revealed that expression levels of the reporter gene associated with znuA were higher than those related to zniCBA in all genetic backgrounds and in both legume hosts, which is consistent with qRT-PCR data (Figure 11B). Also, the symbiotic expression of the β-glucuronidase enzyme associated with both reporter fusion plasmids was higher in strains carrying the deletion in znuA (UPM1628) than in the wild type, and even higher levels of reporter activity were scored in the double mutant UPM1630, similar to what was previously observed in free-living cells. In addition, the activity profile recorded in a zniA-deleted mutant harboring pLMBZnuA plasmid increased in comparison to that in SPF25 in pea-bacteroids, suggesting compensatory levels of expression of this system in the absence of ZniA for the potential restriction to zinc in this legume. Enzymatic activities derived from both reporter fusion plasmids increased in lentil bacteroids induced by the UPM1630 strain, suggesting a higher level of zinc deprivation in this host in the absence of both transport systems.
Discussion
In this work, we have characterized two zinc transporter systems, the canonical conserved ZnuABC and ZniCBA, that display similar, interchangeable functional roles. The presence of at least one of the systems is essential for Rlv SPF25 to grow under zinc-limiting conditions and for full symbiotic performance with pea and lentil plants. Bioinformatic analysis revealed high structural similarity of Rlv ZniA with other periplasmic zinc-binding proteins. The protein contains three conserved histidine residues also present in homologous ZniA from K. pneumoniae, AztC from P. denitrificans, and TroA from A. tumefaciens. The third conserved histidine residue (H193, Rlv ZniA numbering) correlates with Zn2+ as a ligand of the binding protein, whereas the presence of a glutamate residue at the same position has been associated with Fe2+ or Mn2+ specificity (Claverys, 2001; Hantke, 2001; Blindauer, 2015). Homology modelling of the Rlv ZniA structure confirms that its metal substrate might be coordinated by these histidines and a glutamate residue (E268, Rlv ZniA numbering) since they occupy the same position in the metal binding site of K. pneumoniae ZniA (Maunders et al., 2024). This is also in line with our results on ZniA site-directed mutant analysis revealing that the conserved histidines are essential for the function of the ZniCBA transporter system under zinc-limiting conditions. In addition, Rlv ZniA does not contain the central histidine domain present in AztC and TroA that has also been associated with zinc specificity (Blindauer, 2015).
Functional evidence for a direct involvement of Rlv ZniA in zinc transport has been established in this work by the rescue of UPM1630 by zinc addition. The reversion of the double mutant phenotype by expressing either ZniA or ZnuA suggests that ZnuABC and ZniCBA play redundant roles under these conditions. It has been reported that, in addition to ZnuABC, some bacterial species express a second zinc transporter playing similar and redundant functions supporting growth under zinc-limiting conditions (Desrosiers et al., 2010; Corbett et al., 2012; Plumptre et al., 2014; Pederick et al., 2015; Handali et al., 2015; Neupane et al., 2019; Maunders et al., 2024). These transporter systems might play an essential role in bacteria adaptation to environments with low zinc availability, as previously suggested (Neupane et al., 2019). Interestingly, Rlv ZniA did not revert the defective phenotype of a znuA zniCBA mutant, indicating that ZniA does not exert its functional role through a crosstalk with ZnuB permease. This is consistent with the low sequence identity between both SBPs (26.04%) and strongly supports the high affinity zinc uptake contribution of Rlv ZniCBA under extreme zinc-limiting conditions. Similar results were observed in P. denitrificans, in which a specificity in the interaction between the SBP and its cognate permease from ZnuABC and AztABCD transporter systems was also demonstrated (Meléndez et al., 2020). In contrast, the Streptococcus pneumoniae AdcABC transporter system, the homologue to ZnuABC in Gram-positive bacteria, shares the permease AdcB and ATPase AdcC with the orphan SBP AcdII, with both SBPs contributing to the growth of the bacteria under zinc-limiting conditions (Bayle et al., 2011). The expression levels of ZniCBA were lower than ZnuA but high enough to support the growth at wild-type levels in the absence of ZnuABC, as previously observed with the AztABCD system in P. denitrificans (Handali et al., 2015). Since ZniCBA upregulation in response to the metal chelator EDTA increases in the absence of ZnuA, ZniCBA might function as a backup system for high-affinity zinc uptake to ensure adequate metal levels in the cell.
Reporter gene analyses have shown that the expression of zniCBA and znuA genes is dependent on zinc levels in a Zur-dependent manner in free-living cells. The transcriptional regulator Zur controls the expression of many zinc uptake transporters, repressing their synthesis in the presence of zinc (Mikhaylina et al., 2018). In our system, deletion of zur greatly increases zni and znu expression, consistent with the presence of a Zur box in their promoter regions. Furthermore, promoter deletion analysis showed that removal of the Zur box-like sequence also affects the regulation of zniCBA expression. The reduced levels of zniCBA expression when using the shortest form of the promoter suggest that, in addition to the Zur box, the 257–88 upstream region might contain other regulatory signals related to the transcription of zniCBA genes.
Analysis of metal specificity of the transcriptional repression in EDTA-chelated medium revealed that in the absence of both SBPs, Zur responds specifically to zinc, supporting the idea that both transporter systems are involved in zinc acquisition. The higher derepression of the ZnuABC system in the presence of Mn2+ or Fe2+ might be accounted for a competitive inhibition of Zn+2 uptake by an unspecific transporter, as previously stated (Patze and Hantke, 2000). Our results also showed that ZniCBA and ZnuA responded to Mn2+ and Fe2+ in a Zur-dependent manner in the wild-type strain. Previous studies reported that A. tumefaciens ZnuA and TroC repressed its EDTA-induced expression in the presence of these metals, accounting for zinc for the highest capacity (Bhubhanil et al., 2014; Chaoprasid et al., 2016). Furthermore, S. fredii ZnuA was also observed to respond to cobalt in a Zur-dependent manner that was explained by a mismetalation of cobalt to Zur (Zhang et al., 2020). In vitro analysis of metal binding to Zur has revealed the formation of a Zur-DNA complex with manganese in addition to zinc (Gaballa and Helmann, 1998; Patzer and Hantke, 2000; Schröder et al., 2010) and a correlation between metal sensor KDS and cytosolic-free concentration has been reported (Osman et al., 2017). In our system, a mismetalation of manganese and iron to Zur would not explain the repression pattern observed in the absence of ZniA and ZnuA in the UPM1630 strain. Our hypothesis is that an excess of these metals in the cell leads to a displacement of zinc chelated by EDTA to free zinc, which might be transported inside, bind to Zur, and thus repress the expression of the systems. This might happen in the wild-type strain, where these low amounts of free zinc would be internalized by the transporters, but not in the UPM1630 strain, where the Zni and Znu systems are not active and extracellular zinc content might not be enough to repress their expression in a Zur-dependent manner.
We have demonstrated that ZnuABC and ZniCBA are required for the Rlv-pea and -lentil symbiosis. Mutation of both SBPs led to a drastic reduction of symbiotic performance in pea and lentil plants, indicating that both transporter systems are involved in the adaptation and survival of Rlv in pea and lentil nodules. The defective symbiotic phenotype of the double mutant strain Rlv UPM1630 was restored by the expression of ZniA or ZnuA, indicating that pea and lentil bacteroids induced by this strain are affected in zinc uptake. Previous studies have shown that the contribution of ZnuA to the symbiotic performance of S. fredii depends on the legume host. A significant decrease in shoot dry weight was observed in Glycine max and Cajanus cajan plants inoculated with the CCBA45436 strain, but not in Glycine soja (Jiao et al., 2018; Zhang et al., 2020). These results suggest that alternative zinc uptake systems are operating, governed by the legume host. As this strain lacks an Rlv ZniCBA-like transporter system, alternative zinc transporters might not supply enough zinc for CCBA45436 strain symbiotic performance in G. max or C. cajan plants where the metal availability might be lower in the nodule. In addition, it was observed a higher accumulative contribution of ZnuA and accessory zinc uptake proteins (Zip1, Zip2, and c06450) to nodulation of this strain in G. soja and C. cajan but not in G. max (Zhang et al., 2020). Our results indicate that zinc is not entering into the bacteroids induced by ΔznuAΔzniA or, alternatively, that the zinc supplied by potential alternative metal transporters is not enough to inhibit the Zur-dependent expression of both systems. These results align with the observation that the defective symbiotic phenotype of UPM1630 was not rescued with a nutrient solution containing zinc concentrations 100 times higher (80 µM ZnSO4) than those of the standard nutrient solution (0.8 µM ZnSO4) in both legumes (Figure 6). Furthermore, even in the presence of zinc levels 1000-fold higher than in the normal nutrient solution, the double mutant UPM1630 was unable to obtain zinc, neither to correct the deficient plant growth (data not shown) nor to repress transcription of zni/znu genes (Supplementary Figure S4). This “impermeability” to Zn in zinc transporter mutants has not been observed in other symbiotic systems, such as the Sinorhizobium/Glycine max system, where mutations in zinc transporters were complemented for nodulation by the addition of 700 µM ZnSO4 (Zhang et al., 2020).
Our analysis has shown that expression levels of ZniCBA and ZnuA increase in the absence of ZnuA, and both transporter systems are highly induced in bacteroids from pea and lentil nodules induced by a ΔzniAΔznuA mutant strain. These data support the hypothesis that ZniCBA functions as an auxiliary zinc transporter system under these conditions. Although the levels of ZniCBA expression were also lower than ZnuA under symbiotic conditions, they were high enough to optimize the symbiotic performance of Rlv with the legume host. Consistent with our results, the presence of more than one zinc transporter system has been extensively reported to allow bacterial survival under conditions in which the availability of zinc is low. Previous studies revealed that the levels of expression of P. denitrificans ZnuA were higher than those of the SBP AztC in bacterial cultures grown under zinc-limiting conditions with redundant functional roles (Handali et al., 2015). In addition, in A. tumefaciens, the cooperation of TroCBA, ZnuABC, and two zinc chaperones present in this system (ZinT and YciC) are required for growth in zinc-depleted conditions (Chaoprasid et al., 2016). In K. pneumoniae, although ZniA is expressed at higher levels than ZnuA under zinc-limiting conditions, both SBPs were required for bacterial virulence and pathogenicity (Maunders et al., 2024).
The results show a host-dependent expression of both zinc transporter systems exerted at transcriptional levels. qRT-PCR analysis reveals that both transporters are expressed at lower levels in lentil bacteroids induced by the wild-type strain, whereas their expression increases in pea bacteroids, suggesting a higher restriction for zinc uptake into pea bacteroids. Similar results were obtained with the ZnuA-homologue in S. fredii CCBAU45436, which was shown to be differentially upregulated in different soybean species, with higher levels of expression in bacteroids from G. max than in G. soja (Jiao et al., 2018). Preliminary analysis by ICP-MS indicates that the zinc content of plant cytosol from pea and lentil nodules induced by the Rlv SPF25 strain is similar (data not shown). This suggests that the host-dependent pattern of both transporter systems might be explained not by differences in total zinc content but rather by the presence of zinc complexes that could be different in both symbioses, and this condition might be responsible for a higher restriction in providing zinc to the bacteroid in pea versus lentil pants. This is in line with our previous studies that showed that nickel is differentially chelated by malate and citrate in the cytoplasm of nodules induced by Rlv UPM791 in different legumes (Cacho et al., 2010; our unpublished results), an observation associated to the strong effects of the legume host on the Rlv UPM791 [NiFe]-hydrogenase enzyme, along with the different phenotypes of defective mutants in nickel transporters in pea and lentil plants (Brito et al., 2010). Analysis of the molecular basis of such host-dependent effect can allow further advances in the understanding of bacterial adaptation to this fascinating plant-microbe interaction.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
JS: Writing – original draft, Formal Analysis, Visualization, Data curation, Investigation. MB-G: Investigation, Writing – original draft, Data curation, Formal Analysis. LD-S: Formal Analysis, Writing – original draft, Investigation. ID-S: Writing – original draft, Investigation. JP: Conceptualization, Writing – review & editing, Funding acquisition, Supervision. MA: Writing – review & editing, Project administration, Writing – original draft, Investigation, Supervision, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by a grant from Agencia Estatal de Investigación, Ministerio de Ciencia e Investigación (PID2021-124344OB-I100) project to JMP., Spain. JNS was recipient of a Severo Ochoa Program FPI scholarship (PRE2019-090294) from the Ministerio de Ciencia e Investigación.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1598744/full#supplementary-material
References
Abramson, J., Adler, J., Dunger, J., Evans, R., Green, T., Pritzel, A., et al. (2024). Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 630, 493–500. doi: 10.1038/s41586-024-07487-w
Abreu, I., Mihelj, P., and Raimunda, D. (2019). Transition metal transporters in rhizobia: Tuning the inorganic micronutrient requirements to different living styles. Metallomics 11, 735–755. doi: 10.1039/c8mt00372f
Abreu, I., Saéz, Á., Castro-Rodríguez, R., Escudero, V., Rodríguez-Haas, B., Senovilla, M., et al. (2017). Medicago truncatula zinc-iron permease6 provides zinc to rhizobia-infected nodule cells. Plant Cell. Environ. 40, 2706–2719. doi: 10.1111/pce.13035
Akhtar, A. A. and Turner, D. P. (2022). The role of bacterial ATP-binding cassette (ABC) transporters in pathogenesis and virulence: Therapeutic and vaccine potential. Microb. Pathog. 171, 105734. doi: 10.1016/j.micpath.2022.105734
Ammendola, S., Pasquali, P., Pistoia, C., Petrucci, P., Petrarca, P., Rotilio, G., et al. (2007). High-affinity Zn2+ uptake system ZnuABC is required for bacterial zinc homeostasis in intracellular environments and contributes to the virulence of Salmonella enterica. Infect. Immun. 75, 5867–5876. doi: 10.1128/IAI.00559-07
Bailey, T. L., Williams, N., Misleh, C., and Li, W. W. (2006). MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 34, 586–589. doi: 10.1093/nar/gkl198
Ballesteros-Gutiérrez, M., Albareda, M., Barbas, C., López-Gonzálvez, Á., Dunn, M. F., and Palacios, J. M. (2023). A host-specific diaminobutyrate aminotransferase contributes to symbiotic performance, homoserine metabolism, and competitiveness in the Rhizobium leguminosarum/Pisum sativum system. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1182563
Banerjee, S., Wei, B., Bhattacharyya-Pakrasi, M., Pakrasi, H. B., and Smith, T. J. (2003). Structural determinants of metal specificity in the zinc transport protein ZnuA from Synechocystis 6803. J. Mol. Biol. 333, 1061–1069. doi: 10.1016/j.jmb.2003.09.008
Bayle, L., Chimalapati, S., Schoehn, G., Brown, J., Vernet, T., and Durmort, C. (2011). Zinc uptake by Streptococcus pneumoniae depends on both AdcA and AdcAII and is essential for normal bacterial morphology and virulence. Mol. Microbiol. 82, 904–916. doi: 10.1111/j.1365-2958.2011.07862.x
Beringer, J. E. (1974). R factor transfer in Rhizobium leguminosarum. J. Gen. Microbiol. 84, 188–198. doi: 10.1099/00221287-84-1-188
Berntsson, R. P. A., Smits, S. H. J., Schmitt, L., Slotboom, D. J., and Poolman, B. (2010). A structural classification of substrate-binding proteins. FEBS Lett. 584, 2606–2617. doi: 10.1016/j.febslet.2010.04.043
Bertani, G. (1951). Studies on lysogenesis I. J. Bacteriol. 62, 293–300. doi: 10.1128/jb.62.3.293-300.1951
Bhubhanil, S., Sittipo, P., Chaoprasid, P., Nookabkaew, S., Sukchawalit, R., and Mongkolsuk, S. (2014). Control of zinc homeostasis in Agrobacterium tumefaciens via zur and the zinc uptake genes znuABC and zinT. Microbiol. (United Kingdom) 160, 2452–2463. doi: 10.1099/mic.0.082446-0
Blindauer, C. A. (2015). Advances in the molecular understanding of biological zinc transport. Chem. Commun. 51, 4544–4563. doi: 10.1039/c4cc10174j
Brito, B., Palacios, J. M., Hidalgo, E., Imperial, J., and Ruiz-Argueso, T. (1994). Nickel availability to pea (Pisum sativum L.) plants limits hydrogenase activity of Rhizobium leguminosarum bv. viciae bacteroids by affecting the processing of the hydrogenase structural subunits. J. Bacteriol. 176, 5297–5303. doi: 10.1128/jb.176.17.5297-5303.1994
Brito, B., Prieto, R. I., Cabrera, E., Mandrand-Berthelot, M. A., Imperial, J., Ruiz-Argüeso, T., et al. (2010). Rhizobium leguminosarum hupE encodes a nickel transporter required for hydrogenase activity. J. Bacteriol. 192, 925–935. doi: 10.1128/JB.01045-09
Brito, B., Toffanin, A., Prieto, R. I., Imperial, J., Ruiz-Argüeso, T., and Palacios, J. M. (2008). Host-dependent expression of Rhizobium leguminosarum bv. viciae hydrogenase is controlled at transcriptional and post-transcriptional levels in legume nodules. Mol. Plant-Microbe Interact. 21, 597–604. doi: 10.1094/MPMI-21-5-0597
Bui, H. B. and Inaba, K. (2024). Structures, mechanisms, and physiological functions of zinc transporters in different biological kingdoms. Int. J. Mol. Sci. 25, 3045. doi: 10.3390/ijms25053045
Cacho, C., Brito, B., Palacios, J., Pérez-Conde, C., and Cámara, C. (2010). Speciation of nickel by HPLC-UV/MS in pea nodules. Talanta 83, 78–83. doi: 10.1016/j.talanta.2010.08.044
Cerasi, M., Ammendola, S., and Battistoni, A. (2013). Competition for zinc binding in the host-pathogen interaction. Front. Cell. Infect. Microbiol. 3. doi: 10.3389/fcimb.2013.00108
Cha-aim, K., Fukunaga, T., Hoshida, H., and Akada, R. (2009). Reliable fusion PCR mediated by GC-rich overlap sequences. Gene 434, 43–49. doi: 10.1016/j.gene.2008.12.014
Chaoprasid, P., Dokpikul, T., Johnrod, J., Sirirakphaisarn, S., Nookabkaew, S., Sukchawalit, R., et al. (2016). Agrobacterium tumefaciens Zur regulates the high-affinity zinc uptake system TroCBA and the putative metal chaperone YciC, along with ZinT and ZnuABC, for survival under zinc-limiting conditions. Appl. Environ. Microbiol. 82, 3503–3514. doi: 10.1128/AEM.00299-16
Chen, C.-Y. and Morse, S. A. (2001). Identification and characterization of a high-affinity zinc uptake system in Neisseria gonorrhoeae. FEMS Microbiol. Lett. 202, 67–71. doi: 10.1111/j.1574-6968.2001.tb10781.x
Chenna, R., Sugawara, H., Koike, T., Lopez, R., Gibson, T. J., Higgins, D. G., et al. (2003). Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 31, 3497–3500. doi: 10.1093/nar/gkg500
Choi, S. and Bird, A. J. (2014). Zinc’ing sensibly: Controlling zinc homeostasis at the transcriptional level. Metallomics 6, 1198–1215. doi: 10.1039/c4mt00064a
Claverys, J. P. (2001). A new family of high-affinity ABC manganese and zinc permeases. Res. Microbiol. 152, 231–243. doi: 10.1016/S0923-2508(01)01195-0
Corbett, D., Wang, J., Schuler, S., Gloria, L. C., Glenn, S., Brough, D., et al. (2012). Two zinc uptake systems contribute to the full virulence of Listeria monocytogenes during growth in vitro and in vivo. Infect. Immun. 80, 14–21. doi: 10.1128/IAI.05904-11
Den Herder, G. and Parniske, M. (2009). The unbearable naivety of legumes in symbiosis. Curr. Opin. Plant Biol. 12, 491–499. doi: 10.1016/j.pbi.2009.05.010
Desrosiers, D. C., Bearden, S. W., Mier, I., Abney, J., Paulley, J. T., Fetherston, J. D., et al. (2010). Znu is the predominant zinc importer in Yersinia pestis during in vitro growth but is not essential for virulence. Infect. Immun. 78, 5163–5177. doi: 10.1128/IAI.00732-10
Durán, D., Albareda, M., García, C., Marina, A. I., Ruiz-Argüeso, T., and Palacios, J. M. (2021). Proteome Analysis Reveals a Significant Host-Specific Response in Rhizobium leguminosarum bv. viciae Endosymbiotic Cells. Mol. Cell. Proteom. 20, e002276. doi: 10.1074/MCP.RA120.002276
Gaballa, A. and Helmann, J. D. (1998). Identification of a zinc-specific metalloregulatory protein, Zur, controlling zinc transport operons in Bacillus subtilis. J. Bacteriol. 180, 5815–5821. doi: 10.1128/jb.180.22.5815-5821.1998
González-Guerrero, M., Navarro-Gómez, C., Rosa-Núñez, E., Echávarri-Erasun, C., Imperial, J., and Escudero, V. (2023). Forging a symbiosis: transition metal delivery in symbiotic nitrogen fixation. New Phytol. 239, 2113–2125. doi: 10.1111/nph.19098
Grass, G., Franke, S., Taudte, N., Nies, D. H., Kucharski, L. M., Maguire, M. E., et al. (2005). The metal permease ZupT from Escherichia coli is a transporter with a broad substrate spectrum. J. Bacteriol. 187, 1604–1611. doi: 10.1128/JB.187.5.1604-1611.2005
Hanahan, D. (1983). Studies on transformation of Escherichia coli with plasmids. J. Mol. Biol. 166, 557–580. doi: 10.1016/S0022-2836(83)80284-8
Handali, M., Neupane, D. P., Roychowdhury, H., and Yukl, E. T. (2015). Transcriptional regulation, metal binding properties and structure of Pden1597, an unusual zinc transport protein from Paracoccus denitrificans. J. Biol. Chem. 290, 11878–11889. doi: 10.1074/jbc.M115.645853
Hantke, K. (2001). Bacterial zinc transporters and regulators. BioMetals 14, 239–249. doi: 10.1023/A:1012984713391
Hantke, K. (2005). Bacterial zinc uptake and regulators. Curr. Opin. Microbiol. 8, 196–202. doi: 10.1016/j.mib.2005.02.001
Hawkins, J. P. and Oresnik, I. J. (2022). The rhizobium-legume symbiosis: co-opting successful stress management. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.796045
Holm, L., Laiho, A., Törönen, P., and Salgado, M. (2023). DALI shines a light on remote homologs: One hundred discoveries. Protein Sci. 32, e4519. doi: 10.1002/pro.4519
Ilari, A., Alaleona, F., Petrarca, P., Battistoni, A., and Chiancone, E. (2011). The X-ray structure of the zinc transporter ZnuA from Salmonella enterica discloses a unique triad of zinc-coordinating histidines. J. Mol. Biol. 409, 630–641. doi: 10.1016/j.jmb.2011.04.036
Jiao, J., Ni, M., Zhang, B., Zhang, Z., Young, J. P. W., Chan, T. F., et al. (2018). Coordinated regulation of core and accessory genes in the multipartite genome of Sinorhizobium fredii. PloS Genet. 14, 1007428. doi: 10.1371/journal.pgen.1007428
Jiao, J., Wu, L. J., Zhang, B., Hu, Y., Li, Y., Zhang, X. X., et al. (2016). MucR is required for transcriptional activation of conserved ion transporters to support nitrogen fixation of Sinorhizobium fredii in soybean nodules. Mol. Plant-Microbe Interact. 29, 352–361. doi: 10.1094/MPMI-01-16-0019-R
Kandari, D., Joshi, H., and Bhatnagar, R. (2021). Zur: Zinc-sensing transcriptional regulator in a diverse set of bacterial species. Pathogens 10, 344. doi: 10.3390/pathogens10030344
Kovach, M. E., Elzer, P. H., Hill, D. S., Robertson, G. T., Farris, M. A., Roop, R. M., et al. (1995). Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 166, 175–176. doi: 10.1016/0378-1119(95)00584-1
Laguerre, G., Louvrier, P., Allard, M. R., and Amarger, N. (2003). Compatibility of rhizobial genotypes within natural populations of Rhizobium leguminosarum biovar viciae for nodulation of host legumes. Appl. Environ. Microbiol. 69, 2276–2283. doi: 10.1128/AEM.69.4.2276-2283.2003
Lawrence, M. C., Pilling, P. A., Epa, V. C., Berry, A. M., Ogunniyi, A. D., and Paton, J. C. (1998). The crystal structure of pneumococcal surface antigen PsaA reveals a metal-binding site and a novel structure for a putative ABC-type binding protein. Structure 6, 1553–1561. doi: 10.1016/S0969-2126(98)00153-1
Ledermann, R., Schulte, C. C. M., and Poole, P. S. (2021). How rhizobia adapt to the nodule environment. J. Bacteriol. 203, e00539–e00520. doi: 10.1128/JB.00539-20
León-Mediavilla, J., Senovilla, M., Montiel, J., Gil-Díez, P., Saez, Á., Kryvoruchko, I. S., et al. (2018). MtMTP2-Facilitated zinc transport into intracellular compartments is essential for nodule development in Medicago truncatula. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.00990
Livak, K. J. and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lodwig, E., Kumar, S., Allaway, D., Bourdes, A., Prell, J., Priefer, U., et al. (2004). Regulation of L-alanine dehydrogenase in Rhizobium leguminosarum bv. viciae and its role in pea nodules. J. Bacteriol. 186, 842–849. doi: 10.1128/JB.186.3.842-849.2004
Maunders, E. A., Giles, M. W., Ganio, K., Cunningham, B. A., Bennett-Wood, V., Cole, G. B., et al. (2024). Zinc acquisition and its contribution to Klebsiella pneumoniae virulence. Front. Cell. Infect. Microbiol. 13. doi: 10.3389/fcimb.2023.1322973
Meléndez, A. B., Valencia, D., and Yukl, E. T. (2020). Specificity of interactions between components of two zinc ABC transporters in Paracoccus denitrificans. Int. J. Mol. Sci. 21, 1–12. doi: 10.3390/ijms21239098
Mikhaylina, A., Ksibe, A. Z., Scanlan, D. J., and Blindauer, C. A. (2018). Bacterial zinc uptake regulator proteins and their regulons. Biochem. Soc Trans. 46, 983–1001. doi: 10.1042/BST20170228
Moreau, S., Meyer, J. M., and Puppo, A. (1995). Uptake of iron by symbiosomes and bacteroids from soybean nodules. FEBS Lett. 361, 225–228. doi: 10.1016/0014-5793(95)00155-3
Neupane, D. P., Avalos, D., Fullam, S., Roychowdhury, H., and Yukl, E. T. (2017). Mechanisms of zinc binding to the solute-binding protein AztC and transfer from the metallochaperone AztD. J. Biol. Chem. 292, 17496–17505. doi: 10.1074/jbc.M117.804799
Neupane, D. P., Kumar, S., and Yukl, E. T. (2019). Two ABC transporters and a periplasmic metallochaperone participate in zinc acquisition in Paracoccus denitrificans. Biochemistry 58, 126–136. doi: 10.1021/acs.biochem.8b00854
Novichkov, P. S., Laikova, O. N., Novichkova, E. S., Gelfand, M. S., Arkin, A. P., Dubchak, I., et al. (2009). RegPrecise: A database of curated genomic inferences of transcriptional regulatory interactions in prokaryotes. Nucleic Acids Res. 38, 894. doi: 10.1093/nar/gkp894
O’Gara, F. and Shanmugam, K. T. (1976). Regulation of nitrogen fixation by Rhizobia export of fixed N2 as NH4+. Biochim. Biophys. Acta 437, 313–321. doi: 10.1016/0304-4165(76)90001-5
O’Hara, G. W. (2001). Nutritional constraints on root nodule bacteria affecting symbiotic nitrogen fixation: a review. Aust. J. Exp. Agric. 41, 417–433. doi: 10.1071/EA00087
Osman, D., Foster, A. W., Chen, J., Svedaite, K., Steed, J. W., Lurie-Luke, E., et al. (2017). Fine control of metal concentrations is necessary for cells to discern zinc from cobalt. Nat. Commun. 8, 2085. doi: 10.1038/s41467-017-02085-z
Outten, C. E. and O’Halloran, T. V. (2001). Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 292, 2488–2492. doi: 10.1126/science.1060331
Patzer, S. I. and Hantke, K. (1998). The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol. 28, 1199–1210. doi: 10.1046/j.1365-2958.1998.00883.x
Patzer, S. I. and Hantke, K. (2000). The zinc-responsive regulator Zur and its control of the znu gene cluster encoding the ZnuABC zinc uptake system in Escherichia coli. J. Biol. Chem. 275, 24321–24332. doi: 10.1074/jbc.M001775200
Pederick, V. G., Eijkelkamp, B. A., Begg, S. L., Ween, M. P., McAllister, L. J., Paton, J. C., et al. (2015). ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Sci. Rep. 5, 13139. doi: 10.1038/srep13139
Plumptre, C. D., Eijkelkamp, B. A., Morey, J. R., Behr, F., Couñago, R. M., Ogunniyi, A. D., et al. (2014). AdcA and AdcAII employ distinct zinc acquisition mechanisms and contribute additively to zinc homeostasis in Streptococcus pneumoniae. Mol. Microbiol. 91, 834–851. doi: 10.1111/mmi.12504
Poole, P., Ramachandran, V., and Terpolilli, J. (2018). Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 16, 291–303. doi: 10.1038/nrmicro.2017.171
Rukhman, V., Anati, R., Melamed-Frank, M., and Adir, N. (2005). The MntC crystal structure suggests that import of Mn2+ in cyanobacteria is redox controlled. J. Mol. Biol. 348, 961–969. doi: 10.1016/j.jmb.2005.03.006
Saitou, N. and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425. doi: 10.1093/oxfordjournals.molbev.a040454
Sambrook, J. and Russell, D. W. (2001). Molecular Cloning: a laboratory manual. 3th ed (New York: Cold Spring Harbor Laboratory Press).
Sánchez-Cañizares, C., Rey, L., Durán, D., Temprano, F., Sánchez-Jiménez, P., Navarro, A., et al. (2011). Endosymbiotic bacteria nodulating a new endemic lupine Lupinus mariae-josephi from alkaline soils in Eastern Spain represent a new lineage within the Bradyrhizobium genus. Syst. Appl. Microbiol. 34, 207–215. doi: 10.1016/j.syapm.2010.11.020
Schäfer, A., Tauch, A., Jäger, W., Kalinowski, J., Thierbach, G., and Pühler, A. (1994). Small mobilizable multi-purpose cloning vectors derived from the Escherichia coli plasmids pK18 and pK19: selection of defined deletions in the chromosome of Corynebacterium glutamicum. Gene 145, 90324–90327. doi: 10.1016/0378-1119(94)90324-7
Schröder, J., Jochmann, N., Rodionov, D. A., and Tauch, A. (2010). The Zur regulon of Corynebacterium glutamicum ATCC 13032. . BMC Genomics 11, 12. doi: 10.1186/1471-2164-11-12
Simon, R., Priefer, U. B., and Pühler., A. (1983). “Vector plasmids for in-Vivo and in-Vitro manipulations of Gram-Negative bacteria,” in Molecular genetics of the bacteria-plant interaction. Ed. Pühler, A. (Springer-Verlagg, Berlin), 98–106.
Smith, P. K., Krohn, R. I., Hermanson, G. T., Mallia, A. K., Gartner, F. H., Provenzano, M. D., et al. (1985). Measurement of protein using bicinchoninic acid. Anal. Biochem. 150, 76–85. doi: 10.1016/0003-2697(85)90442-7
Sun, X., Baker, H. M., Ge, R., Sun, H., He, Q. Y., and Baker, E. N. (2009). Crystal structure and metal binding properties of the lipoprotein MtsA, responsible for iron transport in Streptococcus pyogenes. Biochemistry 48, 6184–6190. doi: 10.1021/bi900552c
Tamura, K., Stecher, G., and Kumar, S. (2021). MEGA11: molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 38, 3022–3027. doi: 10.1093/molbev/msab120
Tett, A. J., Rudder, S. J., Bourdès, A., Karunakaran, R., and Poole, P. S. (2012). Regulat able vectors for environmental gene expression in Alphaproteobacteria. Appl. Environ. Microbiol. 78, 01188–01112. doi: 10.1128/AEM.01188-12
Vahling-Armstrong, C. M., Zhou, H., Benyon, L., Morgan, J. K., and Duan, Y. (2012). Two plant bacteria, S. meliloti and Ca. Liberibacter asiaticus, share functional znuABC homologues that encode for a high affinity zinc uptake system. PloS One 7, 37340. doi: 10.1371/journal.pone.0037340
Vincent, J. M. (1970). A manual for the practical study of root-nodule bacteria, I.B.P. Handbook Vol. 8 (Oxford, UK: Blackwell scientific publisher), 164.
Wei, B., Randich, A. M., Bhattacharyya-Pakrasi, M., Pakrasi, H. B., and Smith, T. J. (2007). Possible regulatory role for the histidine-rich loop in the zinc transport protein, ZnuA. Biochemistry 46, 8734–8743. doi: 10.1021/bi700763w
Wheatley, R. M., Ramachandran, V. K., Geddes, B. A., Perry, B. J., Yost, C. K., and Poole, P. S. (2017). Role of O2 in the growth of Rhizobium leguminosarum bv. viciae 3841 on glucose and succinate. J. Bacteriol. 199, 00572–00516. doi: 10.1128/JB.00572-16
Yatsunyk, L. A., Easton, J. A., Kim, L. R., Sugarbaker, S. A., Bennett, B., Breece, R. M., et al. (2008). Structure and metal binding properties of ZnuA, a periplasmic zinc transporter from Escherichia coli. J. Biol. Inorg. Chem. 13, 271–288. doi: 10.1007/s00775-007-0320-0
Keywords: Rhizobium leguminosarum, nitrogen fixation, zinc uptake, symbiosis, legumes, pea, lentil
Citation: Soldek JN, Ballesteros-Gutiérrez M, Díaz-Sáez L, Delgado-Santamaría I, Palacios JM and Albareda M (2025) Two zinc ABC transporters contribute to Rhizobium leguminosarum symbiosis with Pisum sativum and Lens culinaris. Front. Plant Sci. 16:1598744. doi: 10.3389/fpls.2025.1598744
Received: 23 March 2025; Accepted: 19 May 2025;
Published: 09 June 2025.
Edited by:
Katharina Pawlowski, Stockholm University, SwedenReviewed by:
Emanuel Maltempi Souza, Federal University of Paraná, BrazilChang-Fu Tian, China Agricultural University, China
Copyright © 2025 Soldek, Ballesteros-Gutiérrez, Díaz-Sáez, Delgado-Santamaría, Palacios and Albareda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marta Albareda, bWFydGEuYWxiYXJlZGFAdXBtLmVz
†Present addresses: Marta Ballesteros-Gutiérrez, Departamento de Biología, Facultad de Ciencias, Universidad Autónoma de Madrid, Madrid, Spain
Ignacio Delgado-Santamaría, Faculty of Chemistry, Biotechnology and Food Science, Norwegian University of Life Sciences, As, Norway
‡ORCID: Joanna N. Soldek, orcid.org/0000-0002-4536-1606
Marta Ballesteros-Gutiérrez, orcid.org/0000-0002-8995-1739