 Roberto Miño1,2
Roberto Miño1,2 Gabriel I. Ballesteros1,3*
Gabriel I. Ballesteros1,3* Karina B. Ruiz4
Karina B. Ruiz4 Ian S. Acuña-Rodríguez1,3
Ian S. Acuña-Rodríguez1,3 Marco A. Molina-Montenegro1,2,5
Marco A. Molina-Montenegro1,2,5- 1Centro de Ecología Integrativa (CEI), Universidad de Talca, Talca, Chile
- 2Instituto de Ciencias Biológicas (ICB), Universidad de Talca, Talca, Chile
- 3Dirección de Investigación, Vicerrectoría Académica, Universidad de Talca, Talca, Chile
- 4Química y Farmacia, Facultad de Ciencias de la Salud, Universidad Arturo Prat, Iquique, Chile
- 5Centro de Investigación en Estudios Avanzados del Maule (CIEAM), Universidad Católica del Maule, Talca, Chile
Soil salinity threatens global food security, making salt tolerance a key agronomic trait. Quinoa (Chenopodium quinoa Willd.), a halophytic pseudo-cereal known for its high nutritional value, emerges as a promising candidate due to its inherent resilience to saline conditions. Although quinoa's physiological and morphological adaptations to salinity are documented, the role of native fungal endophytes in enhancing salinity tolerance remains largely unexplored, particularly across diverse genotypes. This study investigates the contributions of quinoa-associated endophytes to salinity tolerance and seed quality in different genotypes, thus contributing to understand ecological interactions bolstering crop resilience. To achieve this objective, five quinoa genotypes were selected based on their distribution along a 2,200 km latitudinal gradient (19°–39° S), representing a range of ecological niches. Plants with (E+) and without (E−) fungal endophytes were subjected to salinity treatments of 0, 200, and 400 mM NaCl. Salinity tolerance was assessed through photochemical efficiency, gene expression analysis of CqNHX1, and plant survival rates. Seed quality was evaluated by measuring seed weight and protein content, providing a comprehensive assessment of the endophytes' impact on quinoa under stress conditions. Our results reveal that native microbiomes significantly enhanced salinity tolerance and seed quality in a genotype-dependent manner. Notably, E+ plants demonstrated improved photochemical efficiency and higher expression levels of CqNHX1 under high salinity conditions, with survival rates increasing by up to 30% compared to E− plants. Seed weight and protein content were also positively affected, with E+ plants showing up to a 25% increase in protein content under 400 mM NaCl stress. Remarkably, E+ plants exhibited no negative effects under non-saline conditions. These findings suggest that fungal endophytes interactions shift from neutral to beneficial under salinity, with no trade-offs under normal conditions. This highlights the potential role of endophytes in enhancing quinoa resilience and nutritional value, reinforcing their importance for crop adaptation in the face of climate change. Future research should explore the molecular mechanisms underlying these beneficial interactions and assess their applicability to other crops, paving the way for innovative strategies in plant breeding and conservation.
1 Introduction
Climatic conditions significantly influence agricultural productivity, affecting agroecosystems globally (Bilalis et al., 2019). Yet, these systems face increased challenges from environmental stressors, such as drought, high temperatures, and soil salinity, which are exacerbated by climate change (Okon, 2019; Bao et al., 2023). These are projected to become more severe globally forecasted to intensify in most parts of the world, reducing crop yields, survival, and quality, thus impairing food insecurity (Choukr-Allah et al., 2016; Abdel Latef et al., 2021; Ballesteros et al., 2023; Filho et al., 2023). In parallel, the growing global population further pressures agricultural systems to enhance crop yields and resilience to climate change (Weil et al., 2021). Identifying naturally drought and salt-resistant plant species, as alternative sources of edible products, such as leaves, sprouts, or grains, is vital to mitigate the impacts of climate change (Burritt, 2019; Hlásná Čepková et al., 2022; Pathan and Siddiqui, 2022).
Quinoa (Chenopodium quinoa Willd.), an Andean grain crop, emerges as a promising food source due to its gluten-free seeds, high protein content, and resistance to multiple abiotic stresses (high salinity, drought and frost) (Bazile et al., 2014; Li et al., 2021). Indeed, quinoa is well adapted to grow under marginal or extreme environmental conditions, where conventional crops falter (Zurita-Silva et al., 2014; Choukr-Allah et al., 2016; Hinojosa et al., 2018). It has a wide natural variability and genetic diversity, which stems from 7,000 years of domestication and adaptation to a wide range of edapho-climatic conditions and latitudes (from 11°N in Colombia to 40°S in Chile), with five different ecotypes and thousands of accessions (García-Parra et al., 2020). In Chile, there are two ecotypes of quinoa (Salares/Salt flats, and Coastal/Lowlands) cultivated in three isolated, fragmented production zones along a latitudinal gradient (19.7°S to 40°S) (Fuentes et al., 2009; Cai and Gao, 2020; Patiranage et al., 2022). This unique distribution of quinoa along a clinal gradient showcases its remarkable ability to adapt to diverse environmental conditions; it exhibits broad plasticity in terms of morphology, and can adapt to altitude, drought and salinity (Bazile et al., 2014; Martínez et al., 2015). Therefore, quinoa offers to be a promising crop to endure the increasing drought and salinity conditions under the global climatic change scenario (Cai and Gao, 2020).
Quinoa is considered as a facultative halophyte, thriving in saline soils. It has a high salinity tolerance, from 150 mM NaCl to up to 750 mM NaCl (Orsini et al., 2011; Adolf et al., 2013; Pitzschke, 2016; Hussin et al., 2023). In contrast, salt-sensitive crops such as barley, wheat, and corn experience significant yield reduction when salinity exceeds 40 mM NaCl, limiting their cultivation in saline environments (Hinojosa et al., 2018). Its salt resistance is a complex trait involving multiple genes and a diverse array of physiological, morphological, and biochemical mechanisms operating at various levels (Flowers and Colmer, 2015; Cai and Gao, 2020; Hussin et al., 2023). These mechanisms include Na+ exclusion and K+ retention in the leaf mesophyll, production and accumulation of osmolytes (such as soluble sugars, proline, and polyamines), and compartmentalization of Na+ in vacuoles through vacuolar NHX antiporter proteins and epidermal bladder cells (Ruiz et al., 2016; Cai and Gao, 2020; Moog et al., 2022). In general, salt stress tolerance shows significant ecotypic and genotypic variation, with genotypes from the Salares ecotype generally exhibiting higher tolerance compared to Coastal/Lowlands, which are more similar to glycophytic plants, like wheat (Bazile et al., 2014; Morales et al., 2017; Ruiz et al., 2019). Differences among genotypes are evident in cytosolic Na+ and K+/Na+ ratios, CO2 assimilation, germination rate, organic solute accumulation, and seedling growth under salinity stress (Ruiz-Carrasco et al., 2011; Ruiz et al., 2019; Cai and Gao, 2020). However, geographical distribution does not strictly determine salinity tolerance, as some lowland varieties may have similar or greater salt tolerance than Salares varieties (Schmöckel et al., 2017; Ruiz et al., 2017). In terms of nutritional-related parameters, quinoa seeds have a low content of sugar and calories; they present a high protein content with excellent balances of essential aminoacids, as well as high contents of fiber, lipids, carbohydrates, micro- and macronutrients (Granado-Rodríguez et al., 2021). Importantly, quinoa also exhibits a strong genotypic variability in seed quality-related parameters, including size, protein and mineral contents, which are also influenced by the environment (Miranda et al., 2011; Vega-Gálvez et al., 2018; Granado-Rodríguez et al., 2021).
Hence, the diversity observed among quinoa genotypes would constitute the primary factor enabling quinoa’s adaptability and growth across various ecosystems, while maintaining its productivity and nutritional properties, traits that would exclusively depend on its genomic information (Redman et al., 2022; Souri Laki et al., 2024). However, this perspective neglects the contribution of indigenous endophytic microbes to plants’ resilience and adaptation against environmental stress in complex habitats (Pitzschke, 2016; Redman et al., 2022). In quinoa, this topic has been largely underexplored, particularly in terms salt stress responses and seed nutrient content (González-Teuber et al., 2017, 2018, 2022). This is crucial, because if we wrongly attribute plant plasticity to either the environment or the genome, or the interaction of both, overlooking the influence of the associated microbial ecology, then our predictions about plants’ adaptation to future stresses are likely to be incorrect (Bolin, 2025).
Endophytes, which form symbiotic relationship with plants, are posited to significantly enhance quinoa’s adaptability to adverse conditions (Olivieri et al., 2021; Aizaz et al., 2023; Rétif et al., 2023). Through a complex array of mechanisms, endophytes may boost plant growth by improving nutrient uptake, nitrogen fixation and phytohormone production, thereby increasing yield and nutritional quality under stress. These microbial interactions would also increase plant yield parameters, even despite being challenged by stressful environments and/or harsh conditions (Aslam et al., 2020; Kaul et al., 2021; Osman et al., 2021; Omer et al., 2022; Badran et al., 2023). In quinoa, research on exogenic co-inoculation with specific fungal endophytes (Talaromyces minioluteus and Penicillium murcianum) has demonstrated beneficial effects in mitigating the impacts of drought and salinity. Yet, these studies were conducted on endophyte-free plants and limited to a unique genotype obtained from salt flats (González-Teuber et al., 2018, 2022). Hence, while the role of endophytic communities as enhancers of quinoa’s resilience to environmental stressors is an emerging area of interest, and some of the interactions have begun to be elucidated (González-Teuber et al., 2017), much remains to be explored. For instance, the diversity of indigenous, native endophytic communities across different quinoa genotypes, and their contributions in enhancing stress tolerance are largely unexplored, despite the growing body of research highlighting their beneficial influence on plant growth, yield and seed nutritional quality, particularly under abiotic stress conditions (Ballesteros et al., 2023). This knowledge gap is critical, given the realization that quinoa accessions from diverse eco-regions may host unique endophytes (U’Ren et al., 2024) potentially transmitted across generations, aiding for plants’ survival in harsh conditions (Molina-Montenegro et al., 2023). Concomitantly, macroclimatic variations, such as those observed across latitudinal gradients, are hypothesized to act as a driving force in shaping plant life-history traits. Consequently, it is proposed that such variations may also influence the incidence and structuring of fungal endophytic assemblages, affecting their diversity, abundance, and the nature of their symbiotic interactions with host plants (Arnold and Lutzoni, 2007; De Frenne et al., 2013; Mishcherikova et al., 2023; Molina-Montenegro et al., 2023). Yet, these interactions might not follow a strict latitudinal gradient, but be influenced by soil microbiota, plant species, host biomass and nutrient availability (De Frenne et al., 2013; Mishcherikova et al., 2023; U’Ren et al., 2024). Therefore, studying these dynamics across latitudinal gradients could clarify how indigenous endophytes help quinoa cope with abiotic stressors like drought and salinity.
The current study aims to investigate, under semi-controlled greenhouse conditions, the trends and variations in the symbiotic roles of endophytes among five quinoa genotypes from a latitudinal gradient (19°S-38°S), encompassing different ecosystems. The central hypothesis posits that the relative effects, conferred by native endophytes to quinoa, would be following a clinal trend, which would be reflected in shifts on their contributions, from a higher salinity stress tolerance to an improvement in seed nutritional quality and productivity. Specifically, we expected that the relative contribution of native endophytes, in terms of salinity stress tolerance, would have a negative clinal variation, being proportionally higher at lower latitudes, which would be reflected on differences in terms of physiological, biochemical, and molecular responses to salinity stress conditions. Conversely, we expected that endophytes’ contributions in terms of quinoa productivity and seed nutritional properties would exhibit a positive clinal variation, gradually increasing in quinoa from higher latitudes, compared to quinoa from lower latitudes. To test these hypotheses, quinoa plants naturally retaining their endophytes (control, E+) were compared with endophyte-free plants (treatment, E-), under different NaCl concentrations: control (0 mM NaCl), mild (200 mM NaCl) and high salinity stress (400 mM NaCl). To assess the differential effects of endophytes, the following traits were measured: plants’ survival rate, photosynthetic efficiency (Fv/Fm), seed weight, seed protein content, and the expression of CqNHX1, a gene encoding for a vacuolar Na+/H+ antiporter involved in Na+ sequestration. Based on the measured parameters, a relative endophyte contribution index model (RECI) was constructed, and used to estimate the relative contribution of endophytes on the tested genotypes under salinity stress conditions. Overall, this study not only aims to elucidate the potential of habitat-adapted symbiosis along clinal variations but also seeks to explore endophytes’ contributions across different quinoa genotypes, thereby enhancing our understanding of their ecological and agricultural significance.
2 Methodology
2.1 Removal of endophytes from quinoa seeds
Quinoa (Chenopodium quinoa) seeds from five genotypes, distributed along a 2,200 km latitudinal gradient were selected for this study: Pandela (19°S) and Paihuano (29°S) (both belonging to the Salares ecotype), and PRP (34°S), UdeC9 (35°S), and BO78 (39°S) (corresponding to the Lowlands ecotype). Prior to sowing, seeds underwent surface sterilization by immersion in 1% v/v sodium hypochlorite for 3 minutes, followed by three consecutive washes in sterile distilled water (1 min each). 100 seeds from each genotype were individually sown in 50 cm³ pots filled with a sterile 4:1 mixture of sand and peat. After two weeks of growth, seedlings were randomly assigned to two experimental groups: one retaining its native endophytic fungal community (E+), and the other undergoing an endophyte removal treatment (E-). To achieve this, all individual seedlings (E+ and E-) were first treated with a broad-spectrum systemic antibiotic (rifampicin, 50 µg mL-1) for one week to eliminate endophytic bacteria (Barrera et al., 2020; Hereme et al., 2020). Five days after the antibiotic treatment, a treatment to eliminate fungal endophytes was administered only for E- seedlings, using the fungicide Benlate (2 g L-1; DuPont) at a concentration of 0.5 g L-1, for another week; the E+ group was treated with sterile water in equivalent volumes (Barrera et al., 2020; Hereme et al., 2020). After five days, and to verify the efficacy of the antibiotic and fungicide treatment, three seedlings from both groups were randomly selected, and five sections of their roots were cut using a razor blade. The effectiveness of the antibiotic treatment was assessed by checking for the absence of bacterial colony growth, through the culture of root samples on 10 petri dishes containing LB agar at 28°C for 5 days. In parallel, the antifungal treatment was verified using two different methods; half of the root samples assigned for antifungal evaluation were stained with trypan blue in an acid glycerol solution and inspected under microscope at 400x magnification (Motic BA410) observing only fungal hyphae in the E+ samples. The last set of root sections were incubated in potato dextrose agar plates (PDA, Difco) at 18°C for 2 weeks. Hence, after 6 weeks, E- quinoa plants were deemed as fungi endophyte-free only when there was no outgrowth of fungi nor bacteria in the agar plates, thus being suitable for their usage in subsequent experiments.
2.2 Plants, growth conditions, and salinity treatment
Seedlings were maintained in a greenhouse, under natural light and temperature conditions (1324 μmol m-2s-1 ± 243, 20°C ± 4), and watered daily with 20 mL of sterilized water per pot. A total of 180 E+ and 180 E- seedlings (N=360; 36 seedlings per genotype/endophyte condition) were transplanted into 10 L pots filled with a 4:1 mixture of sterile sand:peat and were randomly selected for salinity stress treatments, using the following concentrations: 0 mM NaCl (control), 200 mM NaCl, and 400 mM NaCl. The selection of these concentrations was based on the need to investigate the salinity response of Chenopodium quinoa across a range from non-saline to moderately and highly saline conditions relevant to its known tolerance (Jacobsen, 2003; Koyro and Eisa, 2008; Morales et al., 2011; Adolf et al., 2012; Ruiz et al., 2016). Salt stress treatment involved daily watering of E+ and E- plants with 200 mL of tap water. This was done using either plain tap water without NaCl (control, 0 mM), or tap water mixed with either 200 mM NaCl (for mild stress) or 400 mM NaCl (for severe stress). Therefore, for each genotype, the experiment included twelve replicates for each salinity level (0 mM, 200 mM, and 400 mM NaCl) and endophyte condition (E+ and E-) in a completely randomized block design. To minimize any potential block effect, all plants were rotated weekly.
2.3 Measurements of endophytes’ contributions to environmental tolerance traits
2.3.1 Chlorophyll fluorescence (Fv/Fm)
To assess the effects of symbiotic status on plant photosynthetic performance of quinoa plants upon different salinity stress conditions, the maximum quantum yield of photosystem II was estimated by leaf chlorophyll fluorescence measurements. This parameter has been widely used to characterize the response to stress in different plant species (Hereme et al., 2020; Balboa et al., 2020). Briefly, using a pulse-amplitude modulated fluorometer (Pocket PEA, Hansatech Instruments Ltd, Norfolk, United Kingdom), the chlorophyll fluorescence ratio Fv/Fm (where Fv refers to Fm – F0) was used to detect changes in PSII induced by salinity, on the tested genotypes with different symbiotic status (E+ and E). To ensure optimal photochemical efficiency, three fully expanded leaves per individual were dark-adapted for 30 minutes before being measured by means of a leaf clip. All measurements were conducted at noon.
2.3.2 RNA isolation, cDNA synthesis and quantitative RT-PCR analysis of CqNHX1 gene
Since NHX1 has been correlated with vacuolar compartmentation of Na+, we quantified the relative expression of CqNHX1 using quantitative real-time PCR (qRT-PCR). Briefly, total RNA was extracted from shoots employing the PureLink® Plant RNA Reagent (Invitrogen, USA) and remaining DNA was removed with the TURBO DNA-free™ Kit (Thermo Fisher Scientific, Waltham, MA, USA). Then, RNA integrity was assessed on agarose gels, and concentration measured with a NanoDrop One Microvolume UV-Vis Spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). cDNA was synthesized using the First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, Waltham, MA, USA). All kits were used according to the manufacturer’s protocols. Then, qRT-PCR was conducted using CqNHX1 primers previously published for C. quinoa (5’-GCACTTCTGTTGCTGTGAGTTCCA-3’ sense; 5’-TGTGCCCTGACCTCGTAAACTGAT-3’ antisense) and using as reference the Elongation Factor 1-a houskeeping gene (EF-1α): 5’-GTACGCATGGGTGCTTGACAAACTC-3’(sense); 5’- ATCAGCCTGGGAGGTACCAGTAAT-3’ (antisense) to normalize and estimate up- or down-regulation of the target genes under salinity stress (Ruiz-Carrasco et al., 2011; Zhu et al., 2021). PCRs were carried on an Mx3000P thermocycler (Agilent Technologies. with Brilliant SYBR Green Master Mix (Stratagene, USA) following manufacturer instructions. Cycle threshold (Ct) values were obtained and analyzed with the 2-ΔΔCt method (Livak and Schmittgen, 2001). Relative expression ratio (log2) and fold changes (FC) between CqNXH1 and EF-1α were calculated from the qRT-PCR efficiencies and the crossing point deviation using the mathematical model of Pfaffl (2001) and is presented as the means (± S.D) of five biological replicates per genotype on each experimental condition including symbiotic status and salinity.
2.4 Measurements of endophytes’ contributions on performance traits
2.4.1 Seed weight and protein content
In order to study the effects of symbiotic status on the improvement of quinoa genotypes’ performance upon different salinity stress conditions, the weight of one thousand seeds (SW in g) and the protein content were measured, as proxies of yield and nutritional seed quality, respectively (Granado-Rodríguez et al., 2021). Seed weight measurement was conducted using the standard procedure by FAO/ISTA (FAO & ISTA, 2023). First, the perigonium was removed by separating seeds from the chaff and straw. Then, four replicates were used, made up of 8 samples of 100 quinoa seeds with a diameter > 1.0mm. Seeds were weighed in a digital electronic scale (Boeco BBL-52; 0.01 g-precision) scale and multiplying the average by 10. These seeds were also used for estimation of soluble protein content (PC), using the method established by Bradford (1976) using bovine serum albumin (BSA, Sigma Aldrich) as a designated standard.
2.4.2 Plant final survival
For each experimental group (endophyte presence/cultivar/salinity treatment; N=12), plant survival was recorded at the end of the experiment as a binary parameter, considering that the plant was dead if it presented more than 90% of damaged tissue (Baldelomar et al., 2019). Survival percentage was calculated as (S/N) x 100, where S is the number of plants that survived until the end of the experiment and N was the total number of evaluated plants per experimental condition (N=12).
2.5 Integrated estimation of the relative microbiome contribution on quinoa salinity stress tolerance and performance
Using tolerance and performance parameters (Fv/Fm, plant survival, seed biomass, protein content) an integrative equation representing Plants’ Responses (PR) was used, as a proxy to estimate the relative endophyte contribution index calculation (RECI) for all quinoa genotypes under salinity stress conditions (Montecinos et al., 2012; Balboa et al., 2020). The PR equation uses as input Fv/Fm, survival, and seed biomass parameters weighted by 0.3, as these were deemed as relevant parameters for crop production, while 0.1 was used as weight for protein seed content. As the PR equation used a scale of 0 to 1 to each parameter (Equation 1), the results of survival, biomass and protein content were divided by 100 or 10, respectively. Then, PR with and without endophytes (PRE+; PRE-) were used to calculate a Relative Endophyte Contribution Index [RECI; Equation 2].
Where: PRE+ = Plant responses with symbiosis; PRE- = Plant responses without symbiosis
Hence, RECI values closer to 0 indicate that the native microbiome had a lesser contribution in plants’ performance under different conditions, while positive values indicate a higher contribution of the native microbiomes on these parameters.
2.6 Statistical analysis
To evaluate whether the linear relationship between the latitude of origin of each cultivar and their responses to saline stress is influenced by the plant’s symbiotic status (E+ or E-), we fitted separate linear mixed models (LMMs) for each measured response variable: Fv/Fm, CqNHX1 relative fold expression, seed weight, seed protein content, and final plant survival. In these models, symbiotic status (E+: with symbiosis; E-: without symbiosis), saline stress level (control, 200 mM, and 400 mM), and latitude of origin (19°, 29°, 34°, 35°, and 39° south) were included as fixed factors. The models were implemented using the “lme” function from the nlme R package (Pinheiro et al., 2024), with population treated as a random factor in the error structure. For each response variable, we compared the slopes of the fitted models between experimental groups using the “emtrends” function from the emmeans package (Lenth, 2025). In order to deeply explore the specific role of the symbiotic condition within each level of saline stress, new independent LMM models were also fitted only with the data that corresponded to a given condition (i.e., 0, 200 or 400 mM). To ensure the validity of the models, in all cases the normality of residuals was assessed using the Shapiro-Wilk test (R Core Team, 2024).
3 Results
3.1 Chlorophyll fluorescence Fv/Fm
In northern-origin plants, E+ individuals maintained a higher mean photochemical efficiency compared to E- plants. This symbiotic advantage, however, diminished when comparing genotypes from southern latitudes, indicating that all experimental factors act as significant drivers of this response (Figure 1a, Supplementary Table 1). Under control conditions, the photosynthetic efficiency of the experimental plants, as described by Fv/Fm monitoring, revealed that both E+ and E- plants exhibited comparable photosynthetic performance (Figure 1a). This equivalence was consistent among genotypes from different latitudinal origins, as indicated by the non-significant slopes in both groups (black “a”). These results demonstrate that photochemical efficiency did not vary significantly with the latitudinal origin for either group in the absence of salt stress (Supplementary Table 2). However, at 200 mM NaCl, the influence of endophyte presence became evident, particularly among genotypes from northern latitudes. A similar pattern was observed under 400 mM NaCl, where E+ plants exhibited a negative and significant slope (red “c”), indicating differences in the capacity of genotypes from distinct latitudinal origins to tolerate saline stress (Figure 1a; Supplementary Table 3). In contrast, E- plants showed no significant relationship with latitude (black “a”), suggesting that the impact of salt stress was uniform across all genotypes (Figure 1a). These findings underscore the adaptive role of microbial symbionts, particularly among quinoa genotypes originating from northern latitudes.
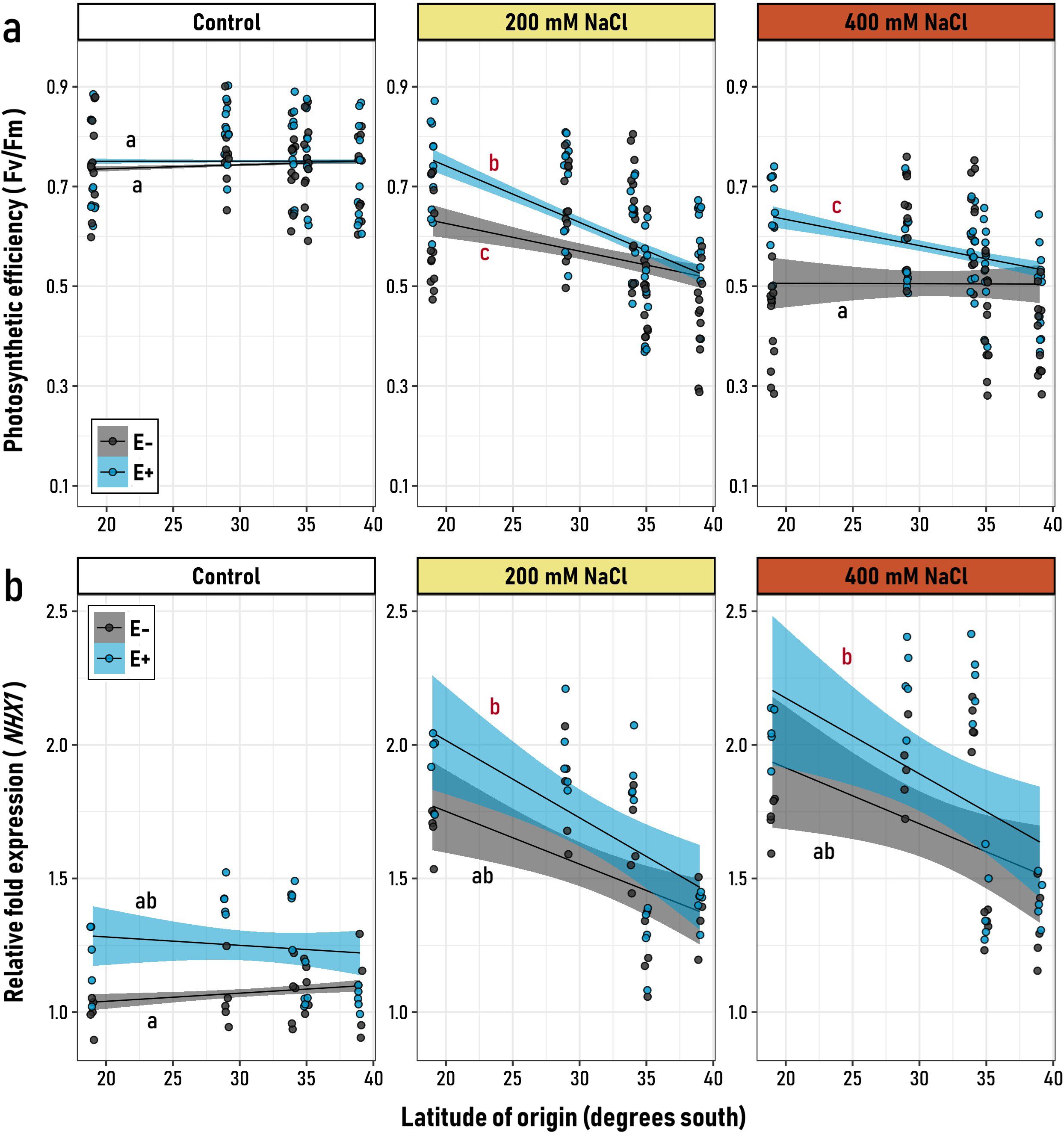
Figure 1. Relationship between the latitudinal origin of five Chenopodium quinoa genotypes (19° to 39° S) and (a) their photosynthetic efficiency (Fv/Fm), and (b) the relative expression of the NHX1 gene, in symbiotic (E+, blue) and non-symbiotic (E−, grey) plants under varying levels of saline stress (Control, 200 and, 400 mM NaCl). Black regression lines represent the best-fit LMM models, with blue and grey shaded areas indicating 95% confidence intervals for E+ and E− groups, respectively. Letters on the plot denote model slopes: red for slopes significantly different from zero, and black for non-significant slopes. Pairwise comparisons of trends among the six experimental groups were conducted, with different letters indicating significant differences (p< 0.05, Tukey-adjusted).
3.2 CqNHX1 relative fold expression
As expected, both symbiotic (E+) and non-symbiotic (E-) plants displayed an overall increase in the expression of salt tolerance mechanisms, such as those regulated by the CqNHX1 gene, when exposed to higher concentrations of NaCl (Figure 1b). Nevertheless, the non-significant triple interaction of the LMM model (Microbiome x Salt treatment x Latitudinal origin) suggests that the effect of the latitude was not detected in the overall general model (Supplementary Table 1). However, when plant responses were analyzed within each salt treatment, CqNHX1 expression levels were more pronounced among plants from northern latitudes. This is evidenced by the negative, significant slopes observed in both E+ and E- plants under 200 and 400 mM NaCl treatments (Figure 1b; Supplementary Table 3), which is similar to the trends observed in Fv/Fm monitoring. Interestingly, the largest differences between symbiotic groups occurred under control conditions. Independent LMMs within each salt treatment revealed that E+ and E- plants differed significantly in their mean responses only under control and 200 mM NaCl conditions (Supplementary Table 2). At 400 mM NaCl, the two groups were statistically indistinguishable (Supplementary Table 2). This suggests that, in the absence of stress, symbiotic quinoa plants (E+) exhibit a basal, constitutive state in which relative NHX1 expression levels consistently surpass those of non-symbiotic plants (E-) across all latitudinal origins (Supplementary Table 2). However, as the level of stress increases, the relative expression advantage of E+ plants over E- plants tends to diminish.
3.3 Seed weight and protein content
Both the microbiome and the salt treatment appeared as significant drivers of the seed weight and its protein content, however, contrary to seed weight, for the seed protein content the effect of these drivers was variable along the latitudinal gradient (Supplementary Table 1). Under control conditions, the latitudinal origin significantly influenced the seed weight of both symbiotic (E+) and non-symbiotic (E-) plants, with seeds from southern latitudes being ~30% heavier compared to the northern latitudes (Figure 2a). However, when plants were subjected to saline conditions, this pattern persisted only in E+ plants, as indicated by significant and similarly positive slopes (Figure 2). In contrast, E- plants exhibited equivalent seed weights across latitudinal origins, characterized by non-significant (flat) slopes (Figure 2; Supplementary Table 3). However, as salt stress increased, the maintenance of this trait depended on the presence of symbiotic endophytes. Interestingly, quinoa seeds from genotypes at 19° S showed no clear effect of endophyte treatment on seed weight (Figure 2a), suggesting that in this cultivar, genetic factors might play a more significant role in determining seed weight.
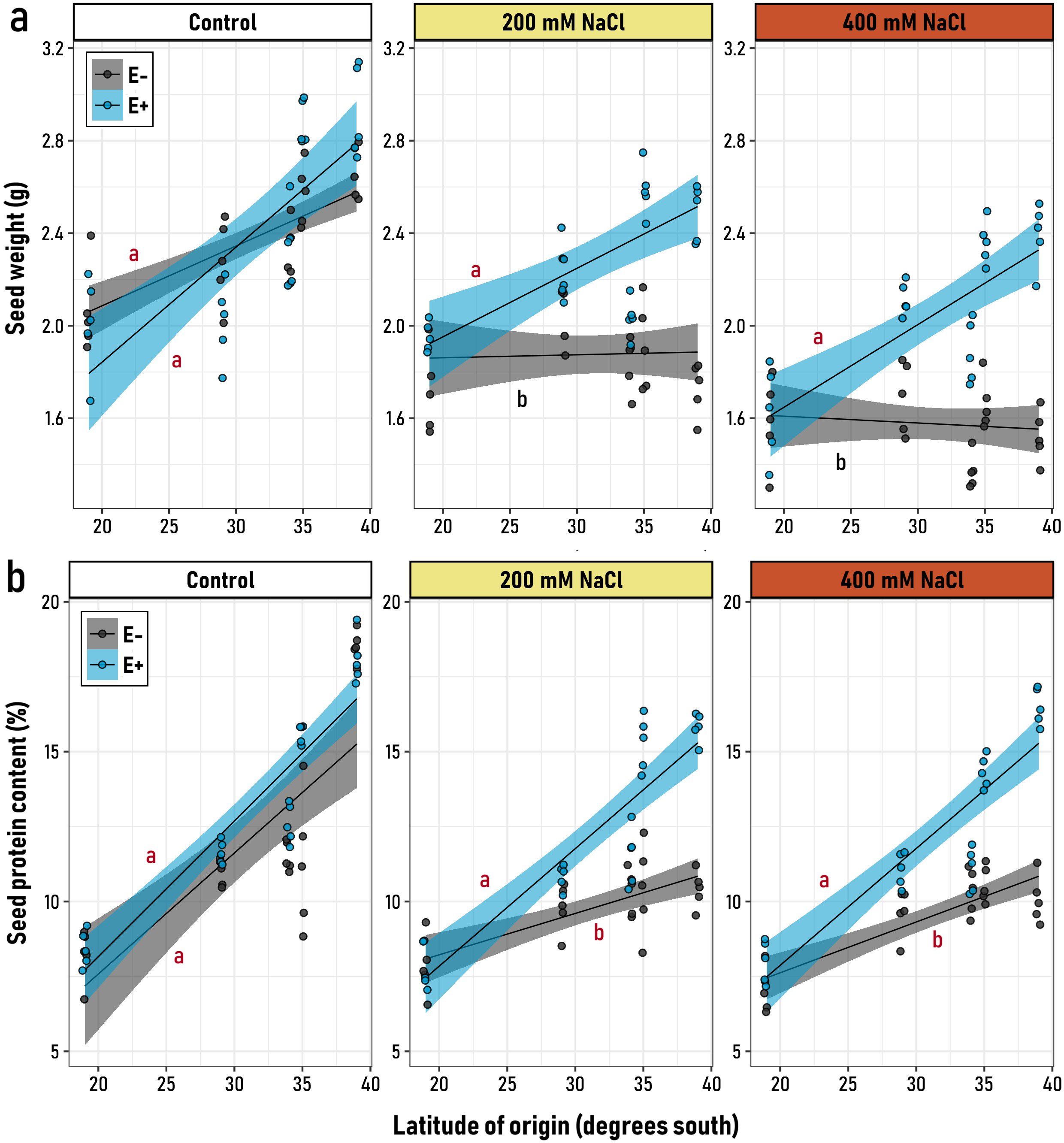
Figure 2. Relationship between the latitudinal origin of five Chenopodium quinoa genotypes (19° to 39° S) and (a) seed mean weight and (b) the seed protein content, in symbiotic (E+, blue) and non-symbiotic (E−, grey) plants under varying levels of saline stress (Control, 200 and, 400 mM NaCl). Black regression lines represent the best-fit LMM models, with blue and grey shaded areas indicating 95% confidence intervals for E+ and E− groups, respectively. Letters on the plot denote model slopes: red for slopes significantly different from zero, and black for non-significant slopes. Pairwise comparisons of trends among the six experimental groups were conducted, with different letters indicating significant differences (p< 0.05, Tukey-adjusted).
Similar to the trends observed for seed weight, both E+ (with endophytes) and E- (without endophytes) plants exhibited comparable positive slopes for seed protein content across the latitude of origin under control conditions (Supplementary Table 2). This indicates that, in the absence of stress, seed protein content was primarily determined by the cultivar rather than by the symbiotic status (Figure 2b). However, under saline conditions (200 and 400 mM NaCl), E+ plants maintained a strong, positive, and significant relationship between seed protein content and latitude. In contrast, E- plants exhibited a weaker positive slope, which was significantly different from that of E+ plants.
3.4 Plant final survival
As denoted by the significant interaction of microbiome x salt treatment x latitudinal origin, the effect of the microbiome on the plant response to salinity stress in terms of survival was variable along the latitudinal gradient (Supplementary Table 1). Under control conditions, both E+ and E- plants exhibited high survival rates, which were consistent across latitudes for both symbiotic groups, as evidenced by the non-significant (flat) slopes (Figure 3). Similarly, under moderate saline stress (200 mM), plants from both symbiotic treatments consistently showed high survival rates compared to control plants. However, it could be observed that the survival percentages of plants with (E+) and without (E-) microbial symbionts started to diverge at northern latitudes (19° S, Figure 3). This divergence became evident under severe saline stress (400 mM), where the higher differences among symbiotic and non-symbiotic plants can be observed in the northernmost originated cultivar (83.3% for E+ vs 25% for E-). Evidence of this is the significant negative slope of the symbiotic (E+) plant group at 400 mM of NaCl (Supplementary Tables 2, 3). Notably, the impact of microbial symbionts on survival varies among genotypes from different latitudinal origins, while becoming significantly different under highly stressful saline conditions (400 mM). Under these conditions, endophytes appear to be inconsequential for southern varieties, whereas they confer a survival advantage (and beneficial effect) to northern genotypes (Figure 3).
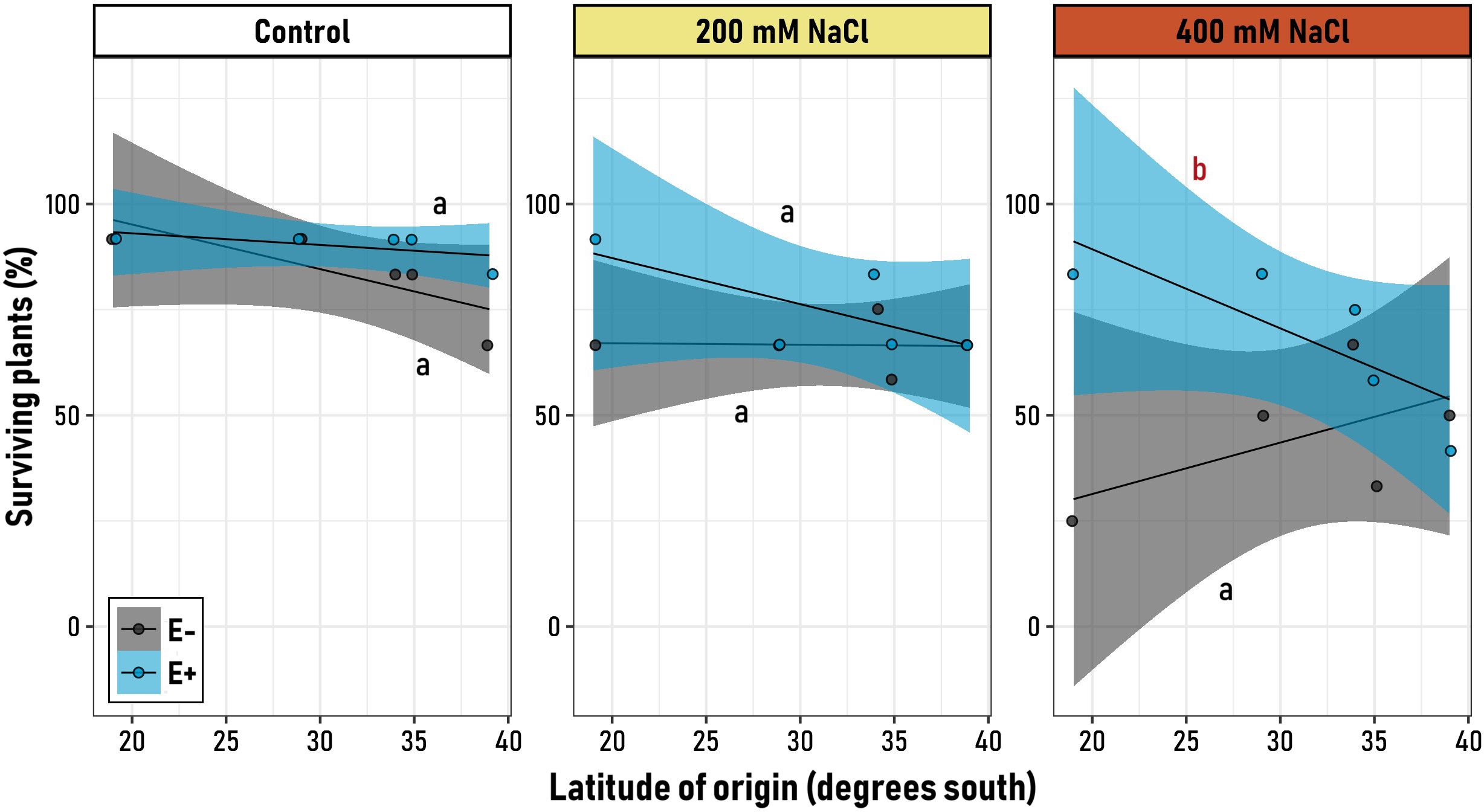
Figure 3. Relationship between the latitudinal origin of five Chenopodium quinoa genotypes (19° to 39° S) and the final proportion of survival plants, in symbiotic (E+, blue) and non-symbiotic (E−, grey) plants under varying levels of saline stress (Control, 200 mM NaCl, 400 mM NaCl). Black regression lines represent the best-fit LMM models, with blue and grey shaded areas indicating 95% confidence intervals for E+ and E− groups, respectively. Letters on the plot denote model slopes: red for slopes significantly different from zero, and black for non-significant slopes. Pairwise comparisons of trends among the six experimental groups were conducted, with different letters indicating significant differences (p< 0.05, Tukey-adjusted).
3.5 Integrated estimation of the microbiome contribution to quinoa performance upon salinity stress
Through the calculation of the Plant Response model (PR), a Relative Endophyte Contribution Index (RECI) was calculated for symbiotic (E+) and non-symbiotic (E-) plants under the three different salinity conditions (Figure 4; Supplementary Figure 1). According to this model, the presence of the native symbionts had a positive effect, albeit with different magnitudes, on all genotypes upon 200 mM and 400 mM NaCl. On the other hand, their contributions under control conditions would follow a positive relation with the clinal gradient. This suggests that the microbiome would play an important role in these parameters under salinity stress. When comparing genotypes, our results show that endophytes’ contribution would be similar for all treatments in PRP, but with a lesser contribution at 400 mM NaCl (4.36%), while at control condition, the observed endophytes’ contribution was similar to salinity stress conditions (3.69%). Therefore, PRP (coastal/lowlands ecotype) would be an average-performing genotype in control condition but would also be the best performing cultivar under the experimental salinity stress conditions used in this study. On the other hand, at 400 mM NaCl, Pandela (Salares ecotype) would be the genotype with the highest relative endophyte contribution (24.13%).
4 Discussion
Quinoa is a crop that can grow and survive under extreme environmental conditions (Jacobsen, 2003; Ruiz et al., 2015). While it has been used as a model plant to understand salt tolerance mechanisms (Ruiz et al., 2015; Manaa et al., 2019), to our knowledge, no prior studies have examined the beneficial presence of its native microbiome on relevant agronomic traits. Our results provide novel evidence suggesting a relative, differential endophyte contribution on quinoa plants, as variations in the endophyte-mediated effects on salinity stress tolerance, seed yield, nutritional content, and plant survival were observed along the latitudinal gradient. Considering that this study assessed both symbiotic status and salinity stress conditions in five genotypes, a novel index was used to estimate the relative endophyte contribution for each genotype and salinity condition; similar approaches have been used in previous studies to account for such differences (Montecinos et al., 2012; Balboa et al., 2020).
Remarkably, a positive, higher contribution of microbiome in salinity stress tolerance and plant survival was observed in genotypes from the northern part of the gradient (19S°-29°S; Figure 3), while in the southern part, a higher, significant role in terms of quinoa grain quality and seed productivity was observed (Figure 2). One possible explanation for these differential endophytes’ activity could be that genotypes from two different ecotypes were compared in this study (Salt flats/Salares, and Coastal/Lowlands). These are naturally cultivated in three isolated, fragmented production zones, including the extreme arid Andean northern highlands (ca 19.7°S), the saline and semiarid soils along the central coast (ca 34.3°), to the rainy-temperate southern regions (ca. 39°S, up to ca. 43°S). Along these zones, a highly contrasting transition of edaphoclimatic conditions is displayed, such as decreasing soil salinity and increasing rainfall (Bazile et al., 2014; Ayala et al., 2020; Ruiz-Carrasco et al., 2011; Miranda et al., 2012). Therefore, it is likely that the relative endophytes’ contribution to environmental tolerance, seed quality and productivity under saline conditions would be following an “habitat-adapted symbiosis” pattern (sensu Rodriguez et al., 2008). Presumably, the differential responses observed for all tested quinoa ecotypes would arise from endophytes’ previous exposure, adaptation and coevolutionary dynamics to selective pressures of the different ecosystems on which quinoa is found (Garnica-Díaz et al., 2023; Ballesteros et al., 2023; Bolin, 2025). These results are supported by differential levels of salt tolerance observed for endophytes isolated from halophyte plants, that could be due to their genetic background, or their interplay with their hosts (Manjunatha et al., 2022). It is plausible that quinoa responses would be mediated by their native endophytic communities rather than solely induced by the stressful environment. The latter uncovers a cryptic yet crucial role of native symbiotic endophytes in plant phenotypic responses to environmental fluctuations (Bolin, 2025).
Photosynthesis is among the most severely affected processes during NaCl salt stress exposure in plants, due to the accumulation of high levels of toxic Na+ and osmotically induced stomatal closure (Shabala et al., 2012; Manaa et al., 2019). One way to study the negative effect of NaCl in plants is by measuring the decrease of maximum quantum yield of fluorescence (Fv/Fm). In the case of facultative halophyte plants, such as C. quinoa, previous reports have shown non-apparent negative effects, or a slight, albeit significant reduction on leaf photochemical performance (Fv/Fm) for several genotypes upon salinity stress (from 100 mM to 400 mM NaCl), while also showing positive growth rates (Hariadi et al., 2011; Shabala et al., 2012; Manaa et al., 2019). Surprisingly, this study provides contrasting evidence in terms of Fv/Fm, as significant differences were observed for genotypes upon mild (200 mM NaCl) and severe salinity stress (400 mM NaCl), and that removal of endophytes is linked to a significant reduction in Fv/Fm values, while also revealing a pattern aligned with the latitudinal gradient, upon mild and severe salinity stress, but not in control conditions (Figure 1a). These negative effects are likely explained by a decline of leaf pigments contents or instability in the pigment-protein complex (Badawy et al., 2021). Moreover, our results imply that endophytes would differentially mitigate the negative effects of saline stress on photosynthetic pigments, photosynthetic performance and PSII functionality, as these effects would be higher in genotypes from the northern part of the gradient (19° - 29°S) compared to genotypes from the southern part of the gradient.
Remarkably, in quinoa genotypes from the northern part of the gradient, symbiotic endophytes significantly induced the expression levels for the gene encoding for the proton-coupled Na+/H+ antiporter 1 (CqNHX1) compared to plants from the southern end, even under conditions of mild and severe salinity stress. CqNHX1 sequesters cytoplasmic Na+ into the vacuole or endosome, in exchange for H+ efflux to the cytosol, thus contributing to reduce Na+ toxicity, maintaining pH homeostasis in plant cells, and contributing to K+ uptake (Ruiz-Carrasco et al., 2011; Bassil and Blumwald, 2014). In quinoa, it has been shown that moderate salt stress (300 mM NaCl) elicits differential expression of CqNHX1, being higher on salt tolerant accessions, although transcriptional responses exhibit genotype-specific variations (Ruiz-Carrasco et al., 2011; Liu et al., 2024). Our study further elucidates the influence of native endophytes upon CqNHX1 expression, revealing a pattern aligned with the latitudinal gradient. Specifically, in the case of the Salares ecotype, endophytes enhanced the expression levels of CqNHX1 compared to their endophyte-free (E-) counterparts (Figure 1b). Conversely, in Coastal/Lowland genotypes subjected at severe salt stress conditions (400 mM NaCl), no significant differences in CqNHX1 expression were observed between E+ and E- plants (Figure 1b). Concomitantly, exogen inoculation using endophytes isolated from plants growing in saline environments showed an enhancement of NHX1 expression and conferred salinity stress tolerance across various salt-sensitive crops, including pepper, tomato, lettuce, and onions (Acuña-Rodríguez et al., 2019; Molina-Montenegro et al., 2020; Ballesteros et al., 2023). Taken together, these results suggest a correlation between the presence of symbiotic endophytes, the higher expression of CqNHX1, and Fv/Fm values. From an overall perspective, the observed differential contribution of endophytes in the enhancement of salinity stress tolerance are likely a result of co-evolutionary adaptations between endophytes and host plants, to cope with either dry or saline environments (González-Teuber et al., 2022).
Quinoa seed yield and nutritional quality are deemed as variable parameters, resulting from the interaction of genetic and multiple environmental factors (temperature, climatic condition, water status, soil nutrient content) (Chouhan et al., 2021; Granado-Rodríguez et al., 2021). While it has reported a relationship between agroecological conditions and environmental factors on nutritional traits of quinoa seeds (Granado-Rodríguez et al., 2021), there is still limited knowledge about the interaction between nutritional traits and symbiotic status. Despite this, it has been proposed that the associated microbiota would enhance plant growth under abiotic stress conditions, through different pathways, including higher production of primary and secondary metabolites, improved photosynthesis, better nutrient absorption, increases in antioxidant content, and greater antioxidant enzymatic activity (González-Teuber et al., 2022; Munir et al., 2022; Singh et al., 2023). Therefore, the negative effects derived from salinity stress would be counteracted by the presence of the native microbiome (Singh et al., 2023). In the case of severe salinity stress, Coastal/Lowland genotypes retaining their endophytes had a higher seed weight and protein content, compared to the endophyte-free condition, even despite having a reduction of ~18% in terms of photosynthetic performance (Supplementary Figure 1). On the other hand, the native endophytes from the Salares genotypes would not display a contribution in terms of quinoa productivity and nutritional quality. These results indicate that the role of endophytes in the symbiotic relationship turns from neutral to beneficial along the latitudinal gradient upon a higher level of salinity stress; this is reflected in the positive relation between seed weight and latitudinal gradient at 400 mM NaCl, suggesting that the native fungal endophytes would be contributing in a reduction of the metabolic costs involved in seed development (Carvalhais et al., 2013). One possible explanation to these observations is that genetic factors and morphological/physiological effects may cause significant differences in the endophytic communities during plant growth and development, particularly under high salinity conditions, thus leading to differential, beneficial effects (Wu et al., 2024). Remarkably, this plant-microbe interaction would not have a detrimental effect on seed weight and protein content, as similar values were observed for all genotypes under control conditions, indicating a neutral interaction (Hereme et al., 2020). Similarly, in wheat, inoculation of halophyte-isolated endophytes had a positive effect on these parameters compared to non-inoculated plants, while non-detrimental effects were observed (Manjunatha et al., 2022). Therefore, while the specific molecular mechanisms involved in these interactions remain to be elucidated, our results clearly point towards an increasingly positive contribution of endophytes under increasing salinity conditions along this latitudinal gradient.
Notably, while retaining endophytes provides differential advantages, these advantages would depend on the tested genotypes and/or their latitudinal origin. In the case of quinoa, this contribution would be higher in terms of salinity stress tolerance in the northern part of the gradient, and in terms of seed productivity from the southern part of the gradient (Figure 4). Hence, it is plausible that endophytes would be playing an important role in ecotypic differentiation, which is considered as one of the main strategies used by organisms to colonize and establish themselves in various environments (Molina-Montenegro et al., 2023). While geographic and environmental factors are known to favor ecotypic differentiation, microbes associated with plants may also play an important role in environmental responses (Liu et al., 2021; Bolin, 2025). This adaptive flexibility, which may promote plant-microorganism fitness alignments, not only enhances a plant’s capacity to navigate and thrive in various ecological niches but also underscores the pivotal role of the microbiome and its composition in the adaptation process. This interplay between genetic and molecular mechanisms of plants and microbiome composition is crucial, as it allows for a dynamic response to environmental challenges, thereby greatly influencing and elucidating the adaptation of certain ecotypes to their specific habitats (Fang et al., 2022; Ramandi et al., 2022). A recent study by Pang and colleagues (2020) further supports this perspective, revealing that both ecotype and environmental conditions can specifically restructure the microbiome, enriching certain hosts’ endophyte microbial taxa. This enrichment facilitates the microbiome’s significant contribution to the adaptation of various ecotypes to their respective environmental conditions. This is consistent with other studies, comparing microbial communities of ecotypes well-adapted to contrasting environmental conditions (Bowsher et al., 2020; Liu et al., 2021). Our study’s outcomes align with studies like those by Pang et al. (2020), suggesting that microbiome composition is integral to the adaptation of plant ecotypes to varying environmental conditions. Overall, and using the relative endophyte contribution index (RECI), a novel integrative model proposed in this study, we found an overall positive relationship between quinoa and its native endophytes. However, their contributions were distinctly influenced by their original habitat, which could be a consequence of differences in their putative, underlying associated molecular mechanisms and functional pathways encoded within their genomes (Partida-Martínez and Heil, 2011). This is especially true in the case of symbiotic endophytic relationships with quinoa plants from environments with higher salinity levels, such as the case of Pandela, on which endophytes contributions were higher compared to the other genotypes. On the other hand, and using the RECI model, we found slight, positive beneficial effects of endophytes for the PRP genotype, even under high salinity stress conditions. Moreover, and considering the plant response model, PRP would have a higher performance compared to the other genotypes, even without its endophytes (Supplementary Figure 1). This suggests that, in this genotype, plant-microbe interactions would be slightly positive or even neutral, as no negative effects were observed in the measured parameters.
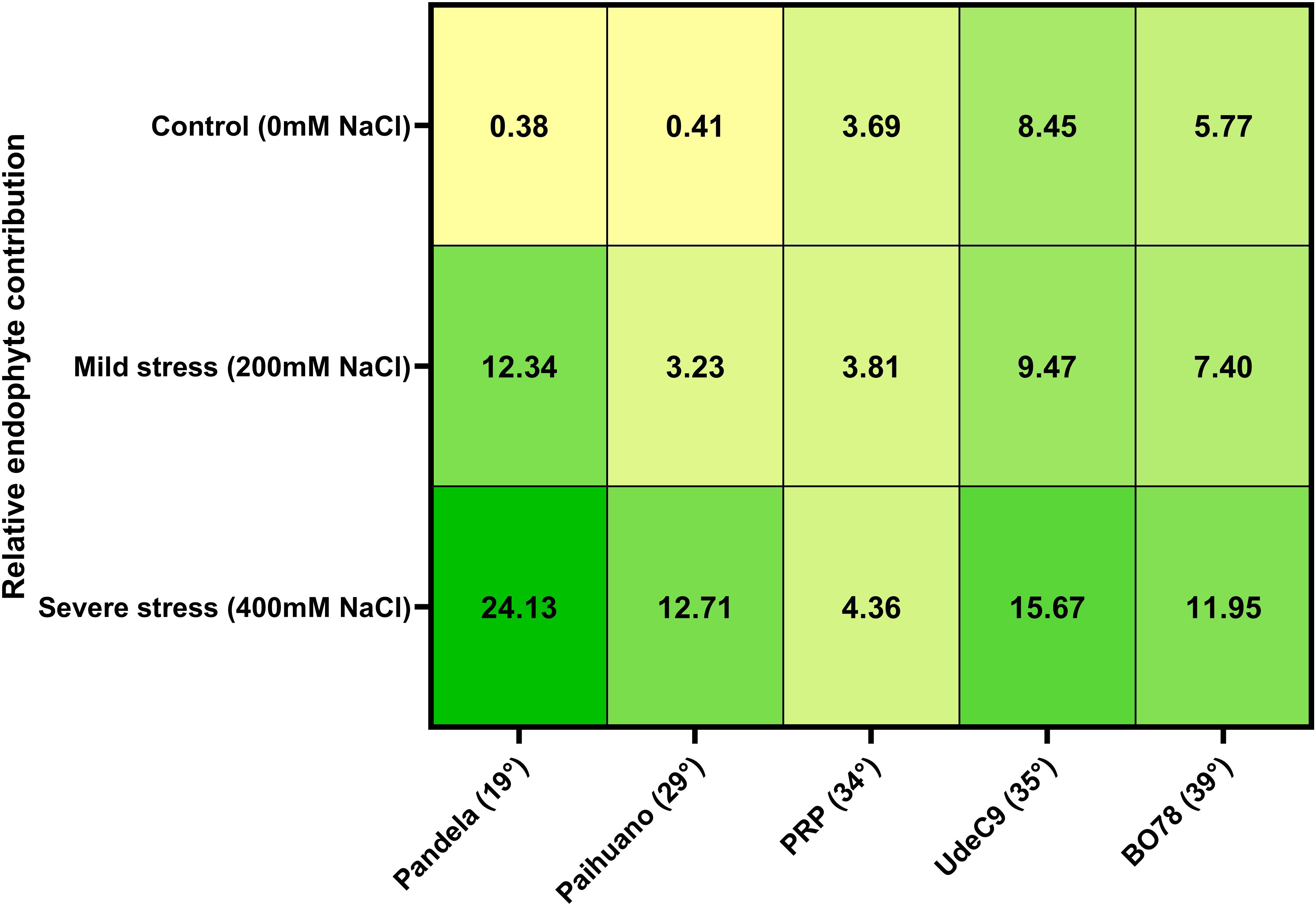
Figure 4. Relative endophyte contribution index (%) calculated using the RECI proposed model based on performance index (Equation 2). Higher numbers indicate a higher endophyte contribution.
5 Final remarks
Understanding and leveraging the role of plant-associated microbiomes is pivotal in enhancing crop productivity under environmental stress, a critical component in addressing global food security challenges. Quinoa, recognized as a superfood due to its exceptional nutritional quality and high stress tolerance, stands at the forefront of these efforts. Our study is pioneering in demonstrating differential contributions of native endophytes, from different genotypes across a latitudinal gradient, in terms of salinity tolerance and seed quality. These observations offer the possibility to conduct research on characterizing these native microbiomes in terms of their abundance, diversity and functional mechanisms, and to determine which would be key endophytes conferring these beneficial effects on both ecotypes. Moreover, these endophytes could also be isolated from these ecotypes, combined in novel bio-formulations or synthetic communities, and inoculated into quinoa and other crops, to further enhance their nutritional quality and tolerance to environmental stressors. This is a promising approach, particularly in the case of climate change scenarios, where productive ecosystems will become more arid and saline in future decades.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
RM: Methodology, Conceptualization, Investigation, Formal analysis, Writing – review & editing, Writing – original draft. GB: Writing – review & editing, Project administration, Funding acquisition, Formal analysis, Writing – original draft, Methodology, Investigation, Conceptualization. KR: Writing – review & editing, Methodology. IA: Methodology, Writing – original draft, Visualization. MM: Conceptualization, Writing – review & editing, Methodology, Resources, Investigation, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Chilean National Fund for Scientific and Technological Development (FONDECYT Iniciacion), grant number 11230888.
Acknowledgments
We acknowledge the valuable work of Ignacio Ascencio Balboa and María José Romero in the maintenance of plant material for this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1602553/full#supplementary-material
References
Abdel Latef, A. A. H., Omer, A. M., Badawy, A. A., Osman, M. S., and Ragaey, M. M. (2021). Strategy of Salt Tolerance and Interactive Impact of Azotobacter chroococcum and/or Alcaligenes faecalis Inoculation on Canola (Brassica napus L.) Plants Grown in Saline Soil. Plants 10, 110. doi: 10.3390/plants10010110
Acuña-Rodríguez, I. S., Hansen, H., Gallardo-Cerda, J., Atala, C., and Molina-Montenegro, M. A. (2019). Antarctic extremophiles: Biotechnological alternative to crop productivity in saline soils. Front. Bioengineering Biotechnol. 7, 22. doi: 10.3389/fbioe.2019.00022
Adolf, V. I., Shabala, S., Andersen, M. N., Razzaghi, F., Jacobsen, S. (2012). Varietal differences of quinoa’s tolerance to saline conditions. Plant Soil 357, 117–129. doi: 10.1007/s11104-012-1133-7
Adolf, V. I., Jacobsen, S. E., and Shabala, S. (2013). Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 92, 43–54. doi: 10.1016/j.envexpbot.2012.07.004
Aizaz, M., Ahmad, W., Asaf, S., Khan, I., Saad Jan, S., Alamri, S., et al. (2023). Characterization of the seed biopriming, plant growth-promoting, and salinity-ameliorating potential of halophilic fungi isolated from hypersaline habitats. Int. J. Mol. Sci. 24, 4904. doi: 10.3390/ijms24054904
Arnold, A. E. and Lutzoni, F. (2007). Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 88, 541–549. doi: 10.1890/05-1459
Aslam, M., Raza, M., Saleem, M., Waqas, M., Iqbal, R., Ahmad, S., et al. (2020). Improving strategic growth stage-based drought tolerance in quinoa by rhizobacterial inoculation. Commun. Soil Sci. Plant Anal. 51, 912–927. doi: 10.1080/00103624.2020.1744634
Ayala, C., Fuentes, F., and Contreras, S. (2020). Dormancy and cardinal temperatures for germination in seed from nine quinoa genotypes cultivated in Chile. Plant Genet. Resources: Characterization Utilization 18, 143–148. doi: 10.1017/S1479262120000209
Badawy, A. A., Alotaibi, M. O., Abdelaziz, A. M., Osman, M. S., Khalil, A. M. A., Saleh, A. M., et al. (2021). Enhancement of seawater stress tolerance in barley by the endophytic fungus Aspergillus ochraceus. Metabolites 11, 428. doi: 10.3390/metabo11070428
Badran, A., Eid, N. A., Hassan, A. R., and Mahmoudi, H. (2023). Differential responses in some quinoa genotypes of a consortium of beneficial endophytic bacteria against bacterial leaf spot disease. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1167250
Balboa, K., Ballesteros, G. I., and Molina-Montenegro, M. A. (2020). Integration of physiological and molecular traits would help to improve the insights of drought resistance in highbush blueberry cultivars. Plants 9, 1457. doi: 10.3390/plants9111457
Baldelomar, M., Atala, C., and Molina-Montenegro, M. A. (2019). Top-down and bottom-up effects deployed by a nurse shrub allow facilitating an endemic Mediterranean orchid. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00466
Ballesteros, G., Newsham, K., Acuña-Rodríguez, I., Atala, C., Torres-Díaz, C., and Molina-Montenegro, M. (2023). Extreme environments as sources of fungal endophytes mitigating climate change impacts on crops in Mediterranean-type ecosystems. Plants People Planet 5, e10415. doi: 10.1002/ppp3.10415
Bao, L., Yu, L., Li, Y., Yan, F., Lyne, V., and Ren, C. (2023). Climate change impacts on agroecosystems in China: Processes, mechanisms, and prospects. Chin. Geographical Sci. 33, 583–600. doi: 10.1007/s11769-023-1362-0
Barrera, A., Hereme, R., Ruiz-Lara, S., Larrondo, L. F., Gundel, P. E., Pollmann, S., et al. (2020). Fungal endophytes enhance the photoprotective mechanisms and photochemical efficiency in the Antarctic Colobanthus quitensis (Kunth) Bartl. exposed to UV-B radiation. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00122
Bassil, E. and Blumwald, E. (2014). The ins and outs of intracellular ion homeostasis: NHX-type cation/H(+) transporters. Curr. Opin. Plant Biol. 22, 1–6. doi: 10.1016/j.pbi.2014.08.002
Bazile, D., Martínez, E., and Fuentes, F. (2014). Diversity of quinoa in a biogeographical island: A review of constraints and potential from arid to temperate regions of Chile. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 42, 9733. doi: 10.15835/NBHA4229733
Bilalis, D., Roussis, I., Kakabouki, I., and Folina, A. (2019). Quinoa (Chenopodium quinoa Willd.) crop under Mediterranean conditions: A review. Ciencia e Investigación Agraria 46, 51–68. doi: 10.7764/rcia.v46i2.2151
Bolin, L. G. (2025). Soil microbes influence the ecology and evolution of plant plasticity. New Phytol. 245, 2224–2236. doi: 10.1111/nph.20383
Bowsher, A. W., Kearns, P. J., Popovic, D., Lowry, D. B., and Shade, A. (2020). Locally adapted Mimulus ecotypes differentially impact rhizosphere bacterial and archaeal communities in an environment-dependent manner. Phytobiomes J. 4, 53–63. doi: 10.1094/PBIOMES-05-19-0026-R
Bradford, M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Burritt, D. J. (2019). “Crop plant adaptation to climate change and extreme environments,” in Encyclopedia of Food Chemistry. Eds. Melton, L., Shahidi, F., and Varelis, P. (Elsevier, Amsterdam, Netherlands: Academic Press), 196–201.
Cai, Z. Q. and Gao, Q. (2020). Comparative physiological and biochemical mechanisms of salt tolerance in five contrasting highland quinoa cultivars. BMC Plant Biol. 20, 70. doi: 10.1186/s12870-020-2279-8
Carvalhais, L. C., Dennis, P. G., Fan, B., Fedoseyenko, D., Kierul, K., Becker, A., et al. (2013). Linking plant nutritional status to plant-microbe interactions. PLoS One. 8 (7), e68555. doi: 10.1371/journal.pone.0068555
Chouhan, D. K., Jaiswal, A. K., Gaurav, A., Mukherjee, J. P., and Verma, J. P. (2021). “PGPM as a potential bioinoculant for enhancing crop productivity under sustainable agriculture,” in Biofertilizers. Eds. Rakshit, A., Meena, V. S., Parihar, M., Singh, H. B., and Singh, A. K. (Cambridge, United Kingdom: Woodhead Publishing), 221–237.
Choukr-Allah, R., Rao, N. K., Hirich, A., Shahid, M., Alshankiti, A., Toderich, K., et al. (2016). Quinoa for marginal environments: Toward future food and nutritional security in MENA and Central Asia regions. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.00346
De Frenne, P., Graae, B. J., Rodríguez-Sánchez, F., Kolb, A., Chabrerie, O., Decocq, G., et al. (2013). Latitudinal gradients as natural laboratories to infer species’ responses to temperature. J. Ecol. 101, 784–795. doi: 10.1111/jec.2013.101.issue-3
Fang, T., Zhou, S., Qian, C., Yan, X., Yin, X., Fan, X., et al. (2022). Integrated metabolomics and transcriptomics insights on flavonoid biosynthesis of a medicinal functional forage, Agriophyllum squarrosum (L.), based on a common garden trial covering six ecotypes. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.985572
FAO & ISTA (2023). Guidelines for the establishment and management of seed testing laboratories – Joint FAO and ISTA Handbook (Rome, Italy: Food and Agriculture Organization of the United Nations). doi: 10.4060/cc6103en
Filho, W. L., Nagy, G. J., Setti, A. F. F., Sharifi, A., Donkor, F. K., Batista, K., et al. (2023). Handling the impacts of climate change on soil biodiversity. Sci. Total Environ. 869, 161671. doi: 10.1016/j.scitotenv.2023.161671
Flowers, T. J. and Colmer, T. D. (2015). Plant salt tolerance: Adaptations in halophytes. Ann. Bot. 115, 327–331. doi: 10.1093/aob/mcu
Fuentes, F. F., Martinez, E. A., Hinrichsen, P. V., Jellen, E. N., and Maughan, P. J. (2009). Assessment of genetic diversity patterns in Chilean quinoa (Chenopodium quinoa Willd.) germplasm using multiplex fluorescent microsatellite markers. Conserv. Genet. 10, 369–377. doi: 10.1007/s10592-008-9604-3
García-Parra, M., Zurita-Silva, A., Stechauner-Rohringer, R., Roa-Acosta, D., and Jacobsen, S.-E. (2020). Quinoa (Chenopodium quinoa Willd.) and its relationship with agroclimatic characteristics: A Colombian perspective. Chilean J. Agric. Res. 80, 290–302. doi: 10.4067/S0718-58392020000200290
Garnica-Díaz, C., Berazaín Iturralde, R., Cabrera, B., Calderón-Morales, E., Felipe, F. L., García, R., et al. (2023). Global plant ecology of tropical ultramafic ecosystems. Botanical Rev. 89, 115–157. doi: 10.1007/s12229-022-09278-2
González-Teuber, M., Contreras, R. A., Zúñiga, G., Barrera, D., and Bascuñán-Godoy, L. (2022). Synergistic association with root endophytic fungi improves morpho-physiological and biochemical responses of Chenopodium quinoa to salt stress. Front. Ecol. Evol. 9. doi: 10.3389/fevo.2021.787318
González-Teuber, M., Urzúa, A., Plaza, P., and Bascuñán-Godoy, L. (2018). Effects of root endophytic fungi on response of Chenopodium quinoa to drought stress. Plant Ecol. 219, 231–240. doi: 10.1007/s11258-017-0791-1
González-Teuber, M., Vilo, C., and Bascuñán-Godoy, L. (2017). Molecular characterization of endophytic fungi associated with the roots of Chenopodium quinoa inhabiting the Atacama Desert, Chile. Genomics Data 11, 109–112. doi: 10.1016/j.gdata.2016.12.015
Granado-Rodríguez, S., Aparicio, N., Matías, J., Pérez-Romero, L. F., Maestro, I., Gracés, I., et al. (2021). Studying the impact of different field environmental conditions on seed quality of quinoa: The case of three different years changing seed nutritional traits in southern Europe. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.649132
Hariadi, Y., Marandon, K., Tian, Y., Jacobsen, S. E., and Shabala, S. (2011). Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 62, 185–193. doi: 10.1093/jxb/erq257
Hereme, R., Morales-Navarro, S., Ballesteros, G., Barrera, A., Ramos, P., Gundel, P. E., et al. (2020). Fungal endophytes exert positive effects on Colobanthus quitensis under water stress but neutral under a projected climate change scenario in Antarctica. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.00264
Hinojosa, L., González, J. A., Barrios-Masias, F. H., Fuentes, F., and Murphy, K. M. (2018). Quinoa abiotic stress responses: A review. Plants 7, 106. doi: 10.3390/plants7040106
Hlásná Čepková, P., Dostalíková, L., Viehmannová, I., Jágr, M., and Janovská, D. (2022). Diversity of quinoa genetic resources for sustainable production: A survey on nutritive characteristics as influenced by environmental conditions. Front. Sustain. Food Syst. 6. doi: 10.3389/fsufs.2022.960159
Hussin, S. A., Ali, S. H., Lotfy, M. E., El-Samad, E. H. A., Eid, M. A., Abd-Elkader, A. M., et al. (2023). Morpho-physiological mechanisms of two different quinoa ecotypes to resist salt stress. BMC Plant Biol. 23, 374. doi: 10.1186/s12870-023-04342-4
Jacobsen, S. E. (2003). The Worldwide Potential for Quinoa (Chenopodium quinoa Willd.). Food Rev. Int. 19 (1–2), 167–177. doi: 10.1081/FRI-120018883
Kaul, S., Choudhary, M., Gupta, S., and Dhar, M. (2021). Engineering host microbiome for crop improvement and sustainable agriculture. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.635917
Koyro, H. W. and Eisa, S. S. (2008). Effect of salinity on composition, viability and germination of seeds of Chenopodium quinoa Willd.. Plant Soil 302, 79–90. doi: 10.1007/s11104-007-9457-4
Lenth, R. (2025). emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.10.7-100001. Available online at: https://rvlenth.github.io/emmeans/ (Accessed February 01, 2025).
Li, L., Lietz, G., and Seal, C. J. (2021). Phenolic, apparent antioxidant, and nutritional composition of quinoa (Chenopodium quinoa Willd.) seeds. Int. J. Food Sci. Technol. 56, 3245–3254. doi: 10.1111/ijfs.14962
Liu, H., Liu, Z., Zhou, Y., Qin, A., Li, C., Liu, Y., et al. (2024). Single-cell transcriptomic analysis reveals the developmental trajectory and transcriptional regulatory networks of quinoa salt bladders. Stress Biol. 4, 47. doi: 10.1007/s44154-024-00189-3
Liu, Y., Wang, H., Peng, Z., Li, D., Chen, W., Jiao, S., et al. (2021). Regulation of root secondary metabolites by partial root-associated microbiotas under the shaping of licorice ecotypic differentiation in northwest China. J. Integr. Plant Biol. 63, 2093–2109. doi: 10.1111/jipb.v63.12
Livak, K. J. and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Manaa, A., Goussi, R., Derbali, W., Cantamessa, S., Abdelly, C., and Barbato, R. (2019). Salinity tolerance of quinoa (Chenopodium quinoa Willd) as assessed by chloroplast ultrastructure and photosynthetic performance. Environ. Exp. Bot. 162, 103–114. doi: 10.1016/j.envexpbot.2019.02.012
Manjunatha, N., Li Hua, S., Sivasithamparam, K., Jones, M. G. K., Edwards, I., Wylie, S. J., et al. (2022). Fungal endophytes from salt-adapted plants confer salt tolerance and promote growth in wheat (Triticum aestivum L.) at early seedling stage. Microbiology 168 (8), 1–15. doi: 10.1099/mic.0.001225
Martínez, E., Fuentes, F., and Bazile, D. (2015). “History of quinoa: Its origin, domestication, diversification, and cultivation with particular reference to the Chilean context,” in Quinoa: Improvement and Sustainable Production (Hoboken, New Jersey, United States of America: John Wiley & Sons), Ch. 2. doi: 10.1002/9781118628041.CH2
Miranda, M., Vega-Gálvez, A., Quispe-Fuentes, I., Rodriguez, M. J., Maureira, H., and Martínez, E. (2012). Nutritional aspects of six quinoa (Chenopodium quinoa Willd.) ecotypes from three geographical areas of Chile. Chilean J. Agric. Res. 72 (2), 175–181. doi: 10.4067/S0718-58392012000200002
Miranda, M., Vega-Gálvez, A., Uribe, E., López, J., Martínez, E., Rodriguez, M. J., et al. (2011). Physico-chemical analysis, antioxidant capacity, and vitamins of six ecotypes of Chilean quinoa (Chenopodium quinoa Willd.). Proc. Food Sci. 1, 1439–1446. doi: 10.1016/J.PROFOO.2011.09.213
Mishcherikova, V., Lynikienė, J., Marčiulynas, A., Gedminas, A., Prylutskyi, O., Marčiulynienė, D., et al. (2023). Biogeography of fungal communities associated with Pinus sylvestris L. and Picea abies (L.) H. Karst. along the latitudinal gradient in Europe. J. Fungi (Basel Switzerland) 9, 829. doi: 10.3390/jof9080829
Molina-Montenegro, M. A., Acuña-Rodríguez, I. S., Torres-Díaz, C., Gundel, P. E., and Dreyer, I. (2020). Antarctic root endophytes improve physiological performance and yield in crops under salt stress by enhanced energy production and Na+ sequestration. Sci. Rep. 10, 1–10. doi: 10.1038/s41598-020-62544-4
Molina-Montenegro, M. A., Ballesteros, G. I., Acuña-Rodríguez, I. S., Pertierra, L. R., Greve, M., Richardson, D. M., et al. (2023). The “Trojan horse” strategy: Seed fungal endophyte symbiosis helps to explain the invasion success of the grass, Poa annua, in Maritime Antarctica. Diversity Distributions 29, 1432–1444. doi: 10.1111/ddi.13768
Montecinos, S., Salinas, P., Oses, R., González-Silvestre, L., and Molina-Montenegro, M. (2012). Linking climatic variability with spatial performance in two varieties of quinoa distributed in a semi-arid zone. Am. J. Plant Sci. 3, 1682–1687. doi: 10.4236/ajps.2012.312205
Moog, M. W., Trinh, M. D. L., Nørrevang, A. F., Bendtsen, A. K., Wang, C., Østerberg, J. T., et al. (2022). The epidermal bladder cell-free mutant of the salt-tolerant quinoa challenges our understanding of halophyte crop salinity tolerance. New Phytol. 236, 1409–1421. doi: 10.1111/nph.18420
Morales, A. J., Bajgain, P., Garver, Z., Maughan, P. J., and Udall, J. A. (2011). Physiological responses of Chenopodium quinoa to salt stress. Int. J. Plant Physiol. Biochem. 3, 219–232. doi: 10.5897/IJPPB11.026
Morales, A., Zurita-Silva, A., Maldonado, J., and Silva, H. (2017). Transcriptional responses of Chilean quinoa (Chenopodium quinoa Willd.) under water deficit conditions uncovers ABA-independent expression patterns. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00216
Munir, N., Hanif, M., Abideen, Z., Sohail, M., El-Keblawy, A., Radicetti, E., et al. (2022). Mechanisms and strategies of plant microbiome interactions to mitigate abiotic stresses. Agronomy 12, 2069. doi: 10.3390/agronomy12092069
Okon, O. G. (2019). “Effect of salinity on physiological processes in plants,” in Microorganisms in saline environments: Strategies and functions, vol. 56 . Eds. Giri, B. and Varma, A. (Springer, Cham), 10. Soil Biology. doi: 10.1007/978-3-030-18975-4_10
Olivieri, L., Saville, R., Gange, A., and Xu, X.-m. (2021). Apple endophyte community in relation to location, scion and rootstock genotypes, and susceptibility to European canker. FEMS Microbiol. Ecol. 97, fiab131. doi: 10.1093/femsec/fiab131
Omer, A. M., Osman, M. S., and Badawy, A. A. (2022). Inoculation with Azospirillum brasilense and/or Pseudomonas geniculata reinforces flax (Linum usitatissimum) growth by improving physiological activities under saline soil conditions. Bot. Stud. 63, 15. doi: 10.1186/s40529-022-00345-w
Orsini, F., Accorsi, M., Gianquinto, G., Dinelli, G., Antognoni, F., Carrasco, K. B. R., et al. (2011). Beyond the ionic and osmotic response to salinity in Chenopodium quinoa: Functional elements of successful halophytism. Funct. Plant Biol. 38, 818–831. doi: 10.1071/FP11088
Osman, M. S., Badawy, A. A., Osman, A. I., and Abdel Latef, A. A. H. (2021). Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 40, 1245–1256. doi: 10.1007/s00344-020-10185-2
Pang, Z., Xu, P., and Yu, D. (2020). Environmental adaptation of the root microbiome in two rice ecotypes. Microbiol. Res. 241, 126588. doi: 10.1016/j.micres.2020.126588
Partida-Martínez, L. P. and Heil, M. (2011). The microbe-free plant: Fact or artifact? Front. Plant Sci. 2. doi: 10.3389/fpls.2011.00100
Pathan, S. and Siddiqui, R. A. (2022). Nutritional composition and bioactive components in quinoa (Chenopodium quinoa Willd.) greens: A review. Nutrients 14, 558. doi: 10.3390/nu14030558
Patiranage, D. S. R., Rey, E., Emrani, N., Wellman, G., Schmid, K., and Schmöckel, S. M. (2022). Genome-wide association study in quinoa reveals selection pattern typical for crops with a short breeding history. Elife. 11, e66873. doi: 10.7554/eLife.66873
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, e45. doi: 10.1093/nar/29.9.e45
Pinheiro, J., Bates, D., R Core, Team. (2024). nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-166. Available online at: https://CRAN.R-project.org/package=nlme.
Pitzschke, A. (2016). Developmental peculiarities and seed-borne endophytes in quinoa: Omnipresent, robust Bacilli contribute to plant fitness. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00002
Ramandi, A., Nourashrafeddin, S. M., Marashi, S. H., and Seifi, A. (2022). Microbiome contributes to phenotypic plasticity in saffron crocus. World J. Microbiol. Biotechnol. 39 (1), 9. doi: 10.1007/s11274-022-03450-x
R Core Team (2024). R: A language and environment for statistical computing (R package version 1.10.7-100001) (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/ (Accessed November 02, 2024).
Redman, R. S., Anderson, J. A., Biaggi, T. M., Malmberg, K. E. L., Rienstra, M. N., Weaver, J. L., et al. (2022). Symbiotic modulation as a driver of niche expansion of coastal plants in the San Juan Archipelago of Washington State. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.868081
Rétif, F., Kunz, C., Calabro, K., Duval, C., Prado, S., Bailly, C., et al. (2023). Seed fungal endophytes as biostimulants and biocontrol agents to improve seed performance. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1260292
Rodriguez, R. J., Henson, J., Van Volkenburgh, E., Hoy, M., Wright, L., Beckwith, F., et al. (2008). Stress tolerance in plants via habitat-adapted symbiosis. ISME J. 2, 404–416. doi: 10.1038/ismej.2007.106
Ruiz, K. B., Aloisi, I., Del Duca, S., Canelo, V., Torrigiani, P., Silva, H., et al. (2016). Salares versus coastal ecotypes of quinoa: Salinity responses in Chilean landraces from contrasting habitats. Plant Physiol. Biochem. 100, 1–8. doi: 10.1016/j.plaphy.2016.01.010
Ruiz, K. B., Biondi, S., Martínez, E. A., Orsini, F., Antognoni, F., and Jacobsen, S. E. (2015). Quinoa – A model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosyst. 150, 357–371. doi: 10.1080/11263504.2015.1027317
Ruiz, K. B., Maldonado, J., Biondi, S., and Silva, H. (2019). RNA-seq analysis of salt-stressed versus non-salt-stressed transcriptomes of Chenopodium quinoa landrace R49. Genes (Basel) 10, 1042. doi: 10.3390/genes10121042
Ruiz, K. B., Rapparini, F., Bertazza, G., Silva, H., Torrigiani, P., and Biondi, S. (2017). Comparing salt-induced responses at the transcript level in a salares and coastal-lowlands landrace of quinoa (Chenopodium quinoa Willd). Environ. Exp. Bot. 139, 127–142. doi: 10.1016/j.envexpbot.2017.05.003
Ruiz-Carrasco, K., Antognoni, F., Coulibaly, A., Lizardi, S., Covarrubias, A., Martínez, E., et al. (2011). Variation in salinity tolerance of four lowland genotypes of quinoa (Chenopodium quinoa Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. 49, 1333–1341. doi: 10.1016/j.plaphy.2011.08.005
Schmöckel, S. M., Lightfoot, D. J., Razali, R., Tester, M., and Jarvis, D. E. (2017). Identification of putative transmembrane proteins involved in salinity tolerance in Chenopodium quinoa by integrating physiological data, RNAseq, and SNP analyses. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01023
Shabala, L., Mackay, A., Tian, Y., Jacobsen, S.-E., Zhou, D., and Shabala, S. (2012). Oxidative stress protection and stomatal patterning as components of salinity tolerance mechanism in Chenopodium quinoa. Physiologia Plantarum 146, 26–38. doi: 10.1111/j.1399-3054.2012.01599.x
Singh, A., Mazahar, S., Chapadgaonkar, S. S., Giri, P., and Shourie, A. (2023). Phyto-microbiome to mitigate abiotic stress in crop plants. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1210890
Souri Laki, E., Rabiei, B., Marashi, H., Jokarfard, V., and Börner, A. (2024). Association study of morpho-phenological traits in Chenopodium quinoa Willd. using SSR markers. Sci. Rep. 14, 5991. doi: 10.1038/s41598-024-56587-0
U’Ren, J. M., Oita, S., Lutzoni, F., Miadlikowska, J., Ball, B., Carbone, I., et al. (2024). Environmental drivers and cryptic biodiversity hotspots define endophytes in Earth’s largest terrestrial biome. Curr. Biol. 34, 1148–1156.e7. doi: 10.1016/j.cub.2024.01.063
Vega-Gálvez, A., Zura, L., Lutz, M., Jagus, R., Agüero, M. V., Pastén, A., et al. (2018). Assessment of dietary fiber, isoflavones and phenolic compounds with antioxidant and antimicrobial properties of Chenopodium quinoa Willd. Chilean J. Agric. Anim. Sci. 34, 57–67. doi: 10.4067/S0719-38902018005000101
Weil, C., Johnson, J., Caristan, R., Brauman, K., Bodirsky, B., Jalava, M., et al. (2021). Resilience of food sufficiency to future climate and societal changes. Res. Square. doi: 10.21203/RS.3.RS-135121/V1
Wu, C. D., Fan, Y. B., Chen, X., Cao, J. W., Ye, J. Y., Feng, M. L., et al. (2024). Analysis of endophytic bacterial diversity in seeds of different genotypes of cotton and the suppression of Verticillium wilt pathogen infection by a synthetic microbial community. BMC Plant Biol. 24, 263. doi: 10.1186/s12870-024-04910-2
Zhu, X., Wang, B., Wang, X., and Wei, X. (2021). Screening of stable internal reference gene of quinoa under hormone treatment and abiotic stress. Physiologia Plantarum 27, 2459–2470. doi: 10.1007/s12298-021-01094-z
Keywords: salt stress, protein content, halophyte, fungal endophytes, native microbiomes, latitudinal gradients, Chenopodium quinoa
Citation: Miño R, Ballesteros GI, Ruiz KB, Acuña-Rodríguez IS and Molina-Montenegro MA (2025) Fungal endophytes boost salt tolerance and seed quality in quinoa ecotypes along a latitudinal gradient. Front. Plant Sci. 16:1602553. doi: 10.3389/fpls.2025.1602553
Received: 29 March 2025; Accepted: 15 May 2025;
Published: 09 June 2025.
Edited by:
Marie-Joelle Virolle, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Dalia A. Gaber Mahmoud, University of Applied Sciences Erfurt, GermanyAli A. Badawy, Al-Azhar University, Egypt
Adrián González Ortega-Villaizán, Polytechnic University of Madrid, Spain
Copyright © 2025 Miño, Ballesteros, Ruiz, Acuña-Rodríguez and Molina-Montenegro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriel I. Ballesteros, Z2JhbGxlc3Rlcm9zQHV0YWxjYS5jbA==