 Rainer Schwacke
Rainer Schwacke Marie E. Bolger1
Marie E. Bolger1 Björn Usadel
Björn Usadel- 1Forschungszentrum Jülich, Institute of Bio- and Geosciences, IBG-4 Bioinformatics, CEPLAS, BioSC, Jülich, Germany
- 2Heinrich-Heine University Düsseldorf, Faculty of Mathematical and Natural Sciences, Institute for Biological Data Science, CEPLAS, Düsseldorf, Germany
Advances in next-generation sequencing technologies over the last decade have substantially reduced the cost and effort required to sequence plant genomes. Whereas early efforts focused primarily on economically important crops and model species, attention has now turned to a broader range of plants, including those with larger and more complex genomes. In 2024, the genomes of 500 plant species were published, including 370 sequenced for the first time. Tracking and providing access to published plant genomes (now covering more than 1800 species) is an invaluable service for plant researchers. PubPlant is an online resource that serves this purpose by cataloging published plant genome sequences and offering multiple visualizations (https://www.plabipd.de/pubplant_main.html). It includes a chronology of genome publications, and cladograms to display the phylogenetic relationships among the sequenced plants. An overview diagram for seed plants highlights taxonomic orders and families with sequenced species and reveals those that have been overlooked thus far. As a use case for PubPlant, we evaluated the status of sequenced food crops. We found that the five plant families featuring the most food crops were those containing the most sequenced plant species.
Introduction
The first plant genome to be sequenced was that of the laboratory model Arabidopsis thaliana, published in the year 2000 (The Arabidopsis Genome Initiative, 2000). It took 10 further years to achieve the milestone of 20 sequenced plant genomes, but only another 4 years to pass 100 genomes, and by the year 2020 the milestone of 500 plant genomes had been achieved. Remarkably, 500 additional plant genomes were sequenced in the next 2 years (one tenth of the time needed for the first 500). This progress is still accelerating, mainly due to the advent of third-generation long-read sequencing technologies (Jiao and Schneeberger, 2017) and their continual refinement (Dumschott et al., 2020; Pucker et al., 2022). Advances in sequencing technologies have gone hand in hand with the development of more powerful bioinformatics algorithms for the assembly and annotation of genomic data. Genome assembly tools such as hifiasm (Cheng et al., 2021a) and verkko (Rautiainen et al., 2023) can integrate data from the two most popular long-read sequencing technologies, namely nanopore sequencing developed by Oxford Nanopore Technologies and single-molecule real-time sequencing commercialized as the PacBio platform by Pacific Biosciences (van Rengs et al., 2022).
The advances in third-generation sequencing have reduced the cost and effort needed for genome sequencing to such a degree that it is now within the means of even moderately-funded research groups. It is therefore unsurprising that recent years have seen a dramatic increase in the number of plant whole-genome sequencing projects (Figure 1). By the end of 2024, more than 1800 plant species had been sequenced, more than 500 of which have been sequenced twice and more than 200 of which have been sequenced three or more times. The number of plant species that have been sequenced is increasing steadily, but the number of individual sequenced genomes is increasing at a much faster rate due to the re-sequencing of the same species multiple times. Re-sequencing is driven by the repetition of earlier sequencing efforts using more advanced technologies to obtain better and more complete genomes as well as intra-species pan-genome projects involving the sequencing of multiple individuals (such as different varieties, cultivars or ecotypes) within a species to explore the full genomic landscape (Golicz et al., 2016). Prominent examples of staple crop pan-genomes include maize (Hufford et al., 2021), barley (Jayakodi et al., 2020), wheat (Jiao et al., 2024), rice (Qin et al., 2021) and potato (Tang et al., 2022; Bozan et al., 2023), as well as fruit crops such as tomato (Gao et al., 2019; Zhou et al., 2022), and beverage crops such as tea (Chen et al., 2023; Tariq et al., 2024).
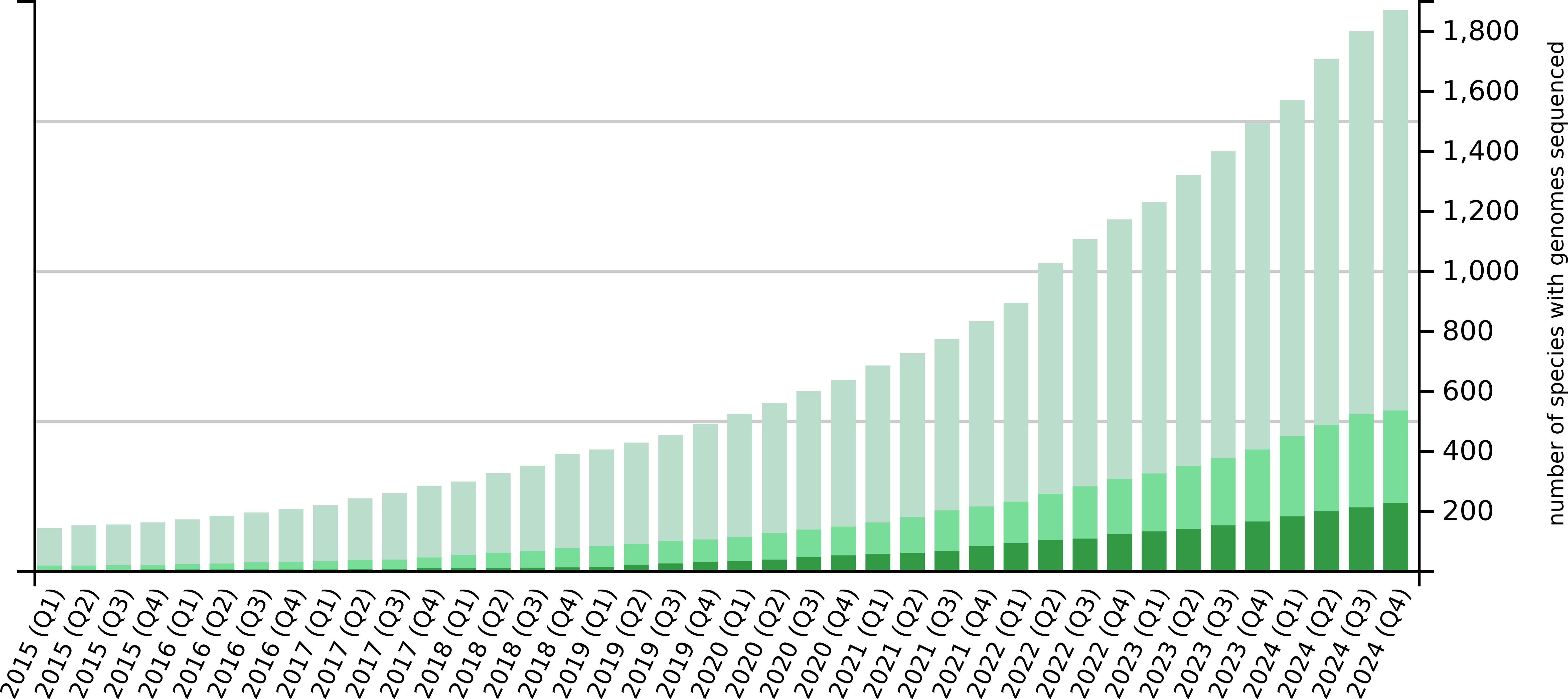
Figure 1. Number of plant species with sequenced and published genomes over time. The light green bars show the number of species that have been sequenced at least once, the medium green bars represent species that have been sequenced at least twice, and the dark green bars are those that have been sequenced three or more times. The bar chart shows quarterly data for the past decade.
The history of plant genome sequencing has been summarized at intervals to highlight the status of sequenced plant genomes at the time of publication, often focusing on particular technological advances (Michael and Jackson, 2013; Chen et al., 2019; Kersey, 2019; Shirasawa et al., 2021; Kress et al., 2022; Sun et al., 2022; Bernal-Gallardo and de Folter, 2024). But such is the pace of change that such review articles are often out of date by the time they are published. One attempt to present a more frequently updated resource is the Plants-Genomes-Technologies (N3) database (Xie et al., 2024). The current iteration is version 3.0 (accessed February 17, 2025), which was published on January 11, 2024. Here, we describe an additional online resource called PubPlant (https://www.plabipd.de/pubplant_main.html) that has tracked and continuously updated published plant genome sequences for almost a decade.
Methods
Tracking published plant genomes
PubPlant tracks published plant genomes using manual search and curation methods. Searches are conducted using the Google search engine, cited reference searches in the Web of Science database (Clarivate, 2025), and the NCBI PubMed citation database (PubMed, 2025). All search results are reviewed manually, and relevant information is extracted and entered into JavaScript Object Notation (JSON) files via copy and paste. These manually curated JSON files serve as the data source for the web-based diagrams that visualize the current status of sequenced plant genomes. The JSON source files also encode phylogenetic information, with taxonomic relationships based on the classifications listed in Table 1. Diagrams are generated client-side using a JavaScript script that employs the D3.js library (v4, https://d3js.org) for cladogram rendering, and the Vis.js library (v3.1, https://visjs.org) for the timeline visualizations.
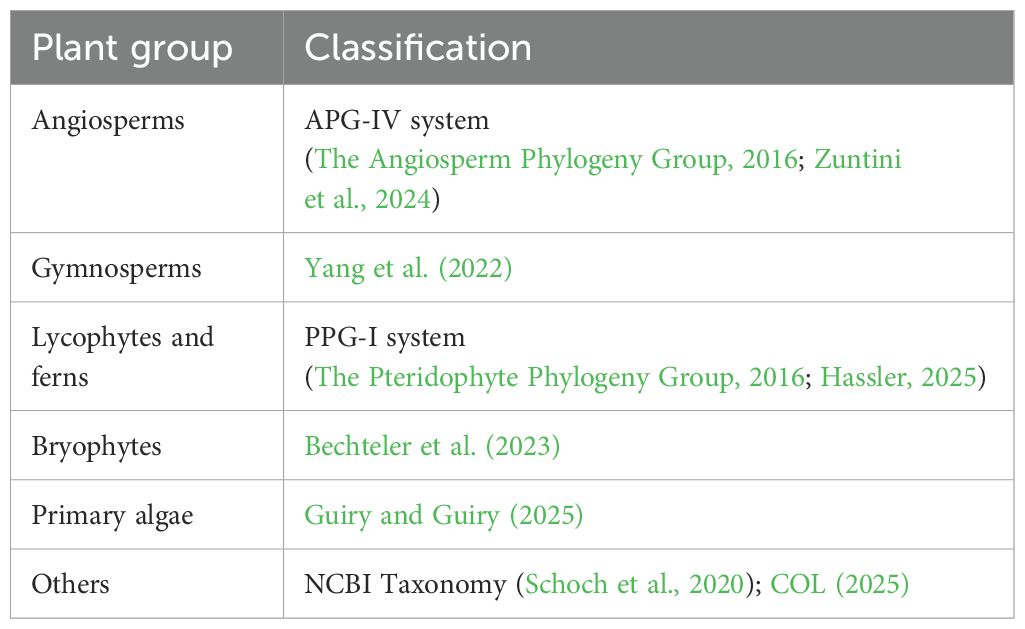
Table 1. Resources used for phylogenetic classification in the diagrams presented by PubPlant.
To be included in PubPlant, the sequenced genome must belong to a plant (from the Archaeplastida group, which includes land plants, charophytes, green algae, glaucophytes, red algae and Rhodelphidophyta) and must be comprehensively described in a peer-reviewed journal. This means that the reads must be assembled into contigs, the contigs must have undergone scaffolding, and structural gene annotation must be completed. Accordingly, PubPlant excludes highly fragmented genomes or those with incomplete or missing structural annotations.
If a publication includes the genomes of multiple accessions representing a single species, such as an intra-species pan-genome, PubPlant counts the species as sequenced and published once. If a pan-genome is based on the genomes of different species – often described as a super pan-genome (Khan et al., 2020) – each species is counted individually. The accepted scientific names of vascular plant species are cross-checked against Plants of the World Online (POWO), 2025 (Govaerts et al., 2021). The lowest recognized taxonomic rank is the species. Accordingly, genomes of different subspecies or varieties are pooled.
Food crops
All statistical data concerning major food crop species were extracted from the Food and Agriculture Organization Corporate Statistical Database (FAOSTAT, 2024). The data provided by FAOSTAT refer to crop categories with an annual production of more than 5000 tons in 2022. Each of these categories contains one or several food crops. The NCBI Taxonomy Database (Schoch et al., 2020) was used to assign correct scientific names to individual crops.
Results and discussion
Chronology of published plant genomes
PubPlant’s timeline view displays sequenced plant genomes according to the chronology of their first publication dates (Figure 2, a full list of published plant genomes is presented in Supplementary Table 1). If there are two publications for the same plant species on the same date, the publication with the earliest acceptance date is listed. The timeline view groups the plant entries as belonging to dicotyledons, non-dicotyledons, non-angiosperms or algae (Figure 2).
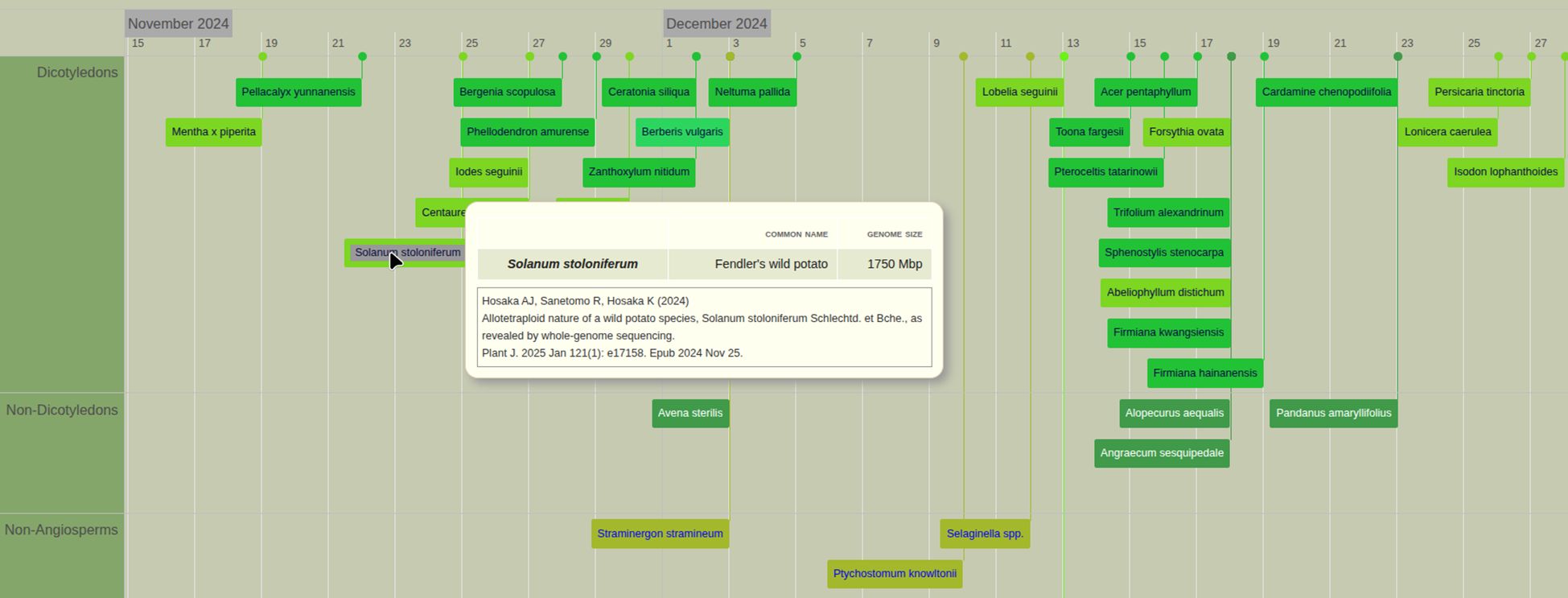
Figure 2. Timeline view of published plant genomes in PubPlant. The colored boxes refer to sequenced genomes of an individual species or a genus and multiple species (spp. for species pluralis) if the publication describes several species of the same genus. On the website, the mouse-over function displays the scientific name (or a list of scientific names if the box refers to several species of the same genus, as in the example shown for Selaginella spp.), the common plant name, the genome size, and the citation of the first publication. Clicking on the box links to the full-text publication in a new window.
Phylogeny of sequenced plant genomes
PubPlant’s cladogram view arranges published plant genomes according to the phylogenetic position of each species (Figure 3). The cladograms for flowering and non-flowering plants are displayed separately to improve legibility. Each entry in the cladogram provides a mouse-over function that displays a popup box containing the scientific name, common name and genome size of the plant. Publication details (one or more publications), including links to the corresponding full-text articles, are also provided in the popup box.

Figure 3. Cladogram view of plant species with sequenced genomes in PubPlant, showing the order Sapindales as an example. The cladogram view goes beyond the taxonomic rank of family, also showing subfamilies and species with sequenced genomes. On the website, the mouse-over function on the scientific name displays a popup box showing the genome size and listing publications (up to three for plants that have been sequenced multiple times) with links to the full-text article(s).
Overview diagram of sequenced seed plants
The two major groups of seed plants are the angiosperms and gymnosperms. Angiosperms are by far the most diverse extant plant group, comprising more than 350,000 known species assigned to more than 400 families in 64 orders. The gymnosperms are a much smaller group, containing only 1100 living species, which is comparable to some of the larger angiosperm genera (e.g., Solanum, Acacia and Rhododendron) in terms of species numbers. By the end of 2024, 1700 angiosperm species and 26 gymnosperm species had been sequenced.
While the cladogram diagrams in PubPlant focus at the individual species level, the overview diagram provides a view at the taxonomic family level using an embedded progress bar. The cladogram displays the phylogenetic position of each plant family while the progress bar depicts the number of sequenced species in that family (Figure 4). The overview diagram also includes families lacking any sequenced species thus far. This reveals that there are many plant families with no sequenced species (e.g., 25 of 38 families in the order Caryophyllales).

Figure 4. Overview diagram of sequenced angiosperms. The example shows the order Caryophyllales, which contains 38 families according to the APG-IV system. The family sizes are represented by light brown bars. The dark brown bars show how many of the species have been sequenced. All bars use a logarithmic scale. On the website, the mouse-over function on a dark bar shows the names of the genera and the number of species with sequenced genomes in a popup box. A yellow dot indicates recently published genomes. A mouse-over tooltip lists the individual species names and links to the full-text articles.
Use case: sequenced food crops
As a use case, we evaluated the current status of sequenced food crops. Economically important food crop plants have always been a major target for sequencing efforts because this provides information about the genes responsible for agronomically favorable traits. The enormous genome sizes of some of these crops, mainly due to polyploidy, the presence of repetitive DNA (Jackson et al., 2011) and very long introns (Xu et al., 2024), hindered progress until the advent of long-read sequencing (Pellicer and Leitch, 2020). For example, the sizes of the onion and broad bean genomes are 16 Gbp (Hao et al., 2023) and 13 Gbp (Jayakodi et al., 2023), respectively. But thanks to the advances mentioned earlier, almost all of the food crop species listed in the FAOSTAT database (FAOSTAT, 2024) have now been sequenced.
Further analysis of the most highly sequenced plant families (Poaceae, Fabaceae, Solanaceae, Rosaceae and Brassicaceae) indicated that these also contain the largest number of food crop species (Figure 5, listed in Table 2). Other families such as Salicaceae, which do not contain any food crops, also may have sequenced species, but this reflects the numerous poplar and willow species in that family, which are important to the timber industry. Similarly, many species of Orchidaceae have been sequenced (Figure 5) despite the presence of only one crop species (vanilla), due to the economic importance of many orchids as ornamental plants.
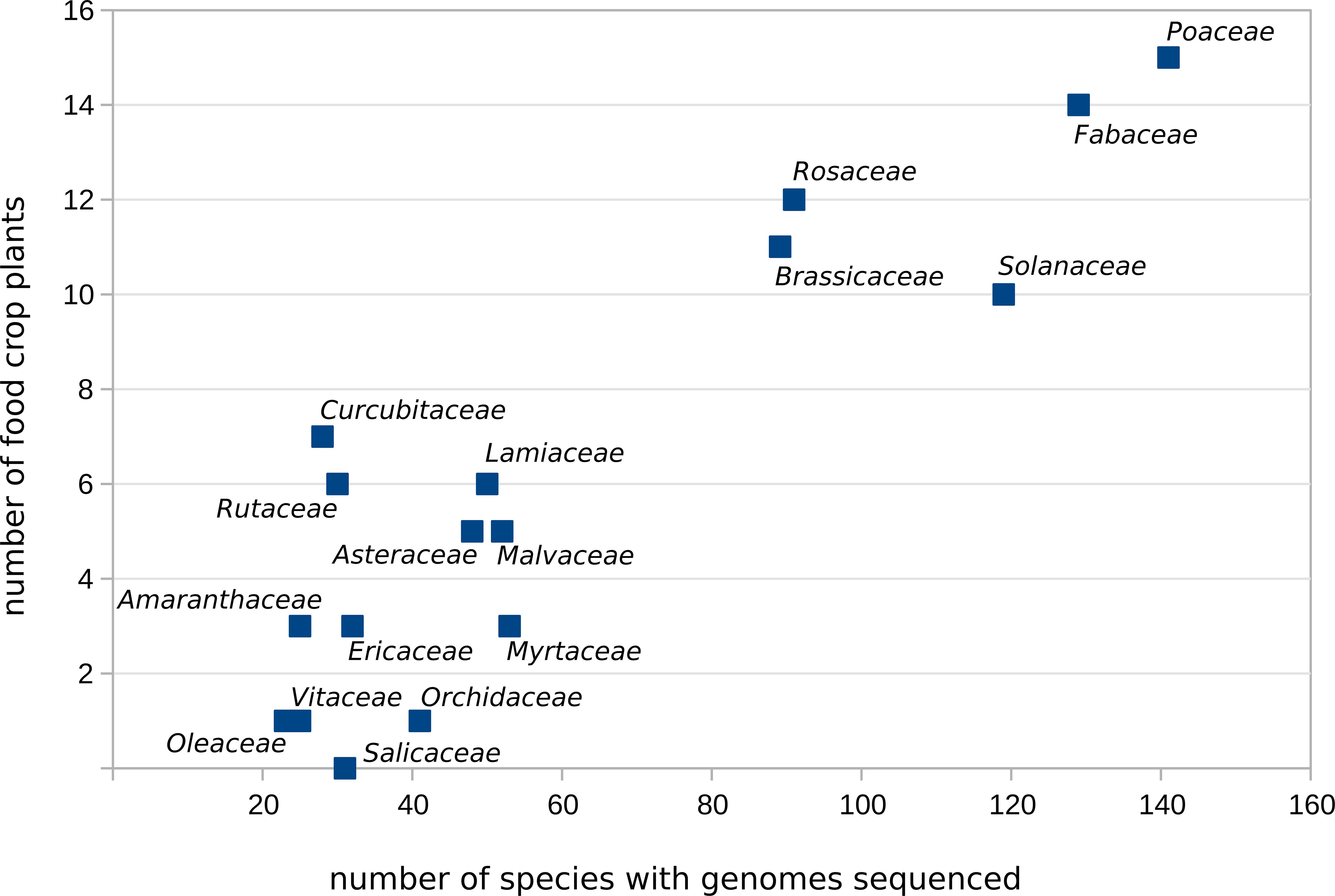
Figure 5. Scatter plot showing plant families in which more than 20 species have been sequenced. The x-axis indicates the number of species per family with sequenced genomes (evaluated February 2025) and the y-axis indicates the number of major food crops per family according to the FAOSTAT database (FAOSTAT, 2024), including those that have yet to be sequenced.
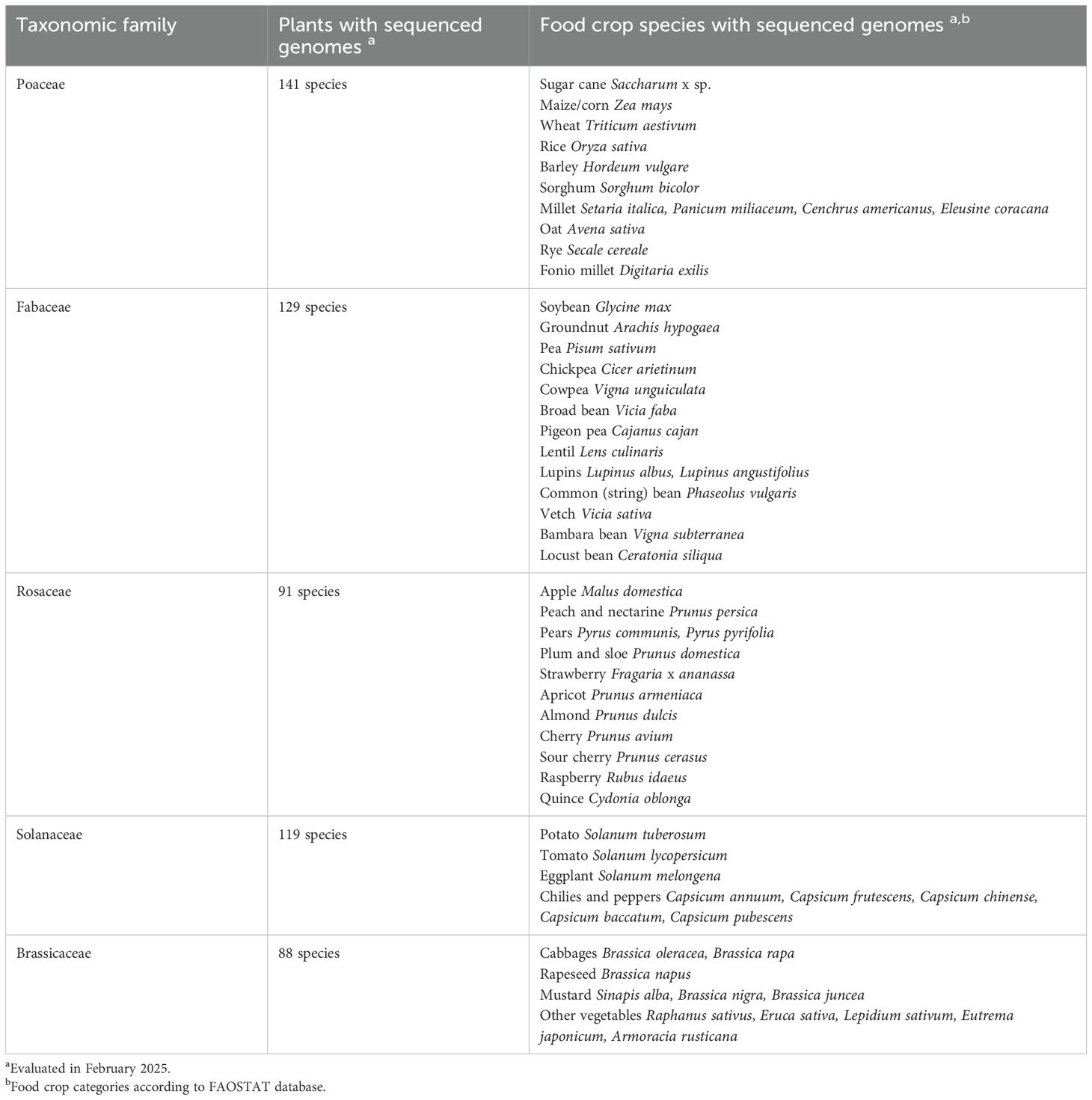
Table 2. Plant families with the highest numbers of sequenced genomes.
Only a few food crops remain to be sequenced (Table 3), including several culinary spices belonging to the Apiaceae (e.g. anise), leek (Amaryllidaceae), gooseberry and currants (Grossulariaceae). Notably, the blackcurrant (Grossulariaceae) is one of the most recently sequenced crop plants (Ziegler et al., 2024). Almost all of the food crops yet to be sequenced rank in the bottom half of the top 120 food crops ordered by production quantity (Table 3). Although all important food crop plants have now been sequenced, the remaining unsequenced species tend to be important in countries that contribute less to plant genome sequencing (Table 3). The countries that have contributed the most to the publication of crop plant genomes are China, the USA, Japan, Germany and Australia (Xie et al., 2024).

Table 3. Food crop plants that have not been sequenced, or have been sequenced very recently.
Conclusion
In recent years, the publication of newly sequenced plant genomes has increased to such an extent that it has become a weekly event (Figure 1). Periodic review articles provide a snapshot of the situation at the time the manuscripts were written but are quickly outdated. We therefore seek to highlight the online resource PubPlant, which provides up-to-date information on published plant genomes. PubPlant features a timeline view, where sequenced plant genomes are arranged in chronological order by the date of first publication (Figure 2), and a cladogram view, showing all sequenced plant species arranged according to their phylogeny (Figure 3). Separate cladograms are provided to display the phylogenetic positions of sequenced flowering and non-flowering plants. A summary diagram shows the phylogenetic position of all sequenced seed plants down to the taxonomic family rank, including those without any species sequenced thus far. It shows the total number of species for each family, the number of sequenced species, and the sequenced genera, while highlighting recently published genomes (Figure 4).
As a use case for PubPlant, we evaluated the status of sequenced food crop plants, which tend to be prioritized for genome sequencing. Unsurprisingly, almost all major food crop species (FAOSTAT food crop categories ranked by production quantity, full list in Supplementary Table 2) have already been sequenced. In addition, the five plant families with the greatest number of sequenced plant genomes (Poaceae, Fabaceae, Rosaceae, Solanaceae and Brassicaceae) are also those containing the most major food crop species (Table 2).
PubPlant has been available online for more than 9 years and has been widely used, including as a resource to prepare review articles summarizing plant genome sequencing progress (Jiao and Schneeberger, 2017; Hao et al., 2022; Bernal-Gallardo and de Folter, 2024) and to evaluate the status of sequenced medicinal plants (Cheng et al., 2021b). Whereas other plant genome resources release irregular updates, PubPlant is updated on a monthly basis, providing a simple and intuitive source of current and historical information on sequenced plant genomes for the benefit of the entire plant research community.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
RS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review & editing, Resources. MB: Conceptualization, Methodology, Supervision, Validation, Writing – original draft, Writing – review & editing, Funding acquisition. BU: Project administration, Supervision, Writing – original draft, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. RS, MB and BU are supported by the German Federal Ministry of Research, Technology and Space (BMFTR) in the frame of the German Network for Bioinformatics Infrastructure (de.NBI).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1603547/full#supplementary-material
Supplementary Table 1 | List of published sequenced plant genomes in chronological order.
Supplementary Table 2 | List of food crop categories (FAOSTAT) ranked by production quantity.
References
Akutsu, M., Shinozawa, A., Nishiyama, T., Sakata, Y., and Hiwatashi, Y. (2024). De novo sequencing allows genome-wide identification of genes involved in galactomannan synthesis in locust bean (Ceratonia siliqua). DNA Res. 31, dsae033. doi: 10.1093/dnares/dsae033
COL. (2025). Catalogue of life. (Amsterdam, The Netherlands). Available online at: https://www.catalogueoflife.org (Accessed February 10, 2025).
Bechteler, J., Peñaloza-Bojacá, G., Bell, D., Gordon Burleigh, J., McDaniel, S. F., Christine Davis, E., et al. (2023). Comprehensive phylogenomic time tree of bryophytes reveals deep relationships and uncovers gene incongruences in the last 500 million years of diversification. Am. J. Bot. 110, e16249. doi: 10.1002/ajb2.v110.11
Bernal-Gallardo, J. J. and de Folter, S. (2024). Plant genome information facilitates plant functional genomics. Planta 259, 117. doi: 10.1007/s00425-024-04397-z
Bozan, I., Achakkagari, S. R., Anglin, N. L., Ellis, D., Tai, H. H., and Strömvik, M. V. (2023). Pan-genome analyses reveal impact of transposable elements and ploidy on the evolution of potato species. Proc. Natl. Acad. Sci. U. S. A. 120, e2211117120. doi: 10.1073/pnas.2211117120
Chen, F., Song, Y., Li, X., Chen, J., Mo, L., Zhang, X., et al. (2019). Genome sequences of horticultural plants: past, present, and future. Hortic. Res. 6, 112. doi: 10.1038/s41438-019-0195-6
Chen, S., Wang, P., Kong, W., Chai, K., Zhang, S., Yu, J., et al. (2023). Gene mining and genomics-assisted breeding empowered by the pangenome of tea plant Camellia sinensis. Nat. Plants 9, 1986–1999. doi: 10.1038/s41477-023-01565-z
Cheng, H., Concepcion, G. T., Feng, X., Zhang, H., and Li, H. (2021a). Haplotype-resolved de novo assembly using phased assembly graphs with hifiasm. Nat. Methods 18, 170–175. doi: 10.1038/s41592-020-01056-5
Cheng, Q. Q., Ouyang, Y., Tang, Z. Y., Lao, C. C., Zhang, Y. Y., Cheng, C. S., et al. (2021b). Review on the development and applications of medicinal plant genomes. Front. Plant Sci. 12, 791219. doi: 10.3389/fpls.2021.791219
Clarivate (2025). Web of Science database. Available online at: https://www.webofscience.com (Accessed February 10, 2024).
Dumschott, K., Schmidt, M. H.-W., Chawla, H. S., Snowdon, R., and Usadel, B. (2020). Oxford Nanopore Sequencing: New opportunities for plant genomics? J. Exp. Bot. 71, 5313–5322. doi: 10.1093/jxb/eraa263
FAOSTAT (2024). Food and agriculture organization of the united nations. Available online at: https://www.fao.org/faostat (Accessed March 9, 2024).
Gao, L., Gonda, I., Sun, H., Ma, Q., Bao, K., Tieman, D. M., et al. (2019). The tomato pan-genome uncovers new genes and a rare allele regulating fruit flavor. Nat. Genet. 51, 1044–1051. doi: 10.1038/s41588-019-0410-2
Golicz, A. A., Batley, J., and Edwards, D. (2016). Towards plant pangenomics. Plant Biotechnol. J. 14, 1099–1105. doi: 10.1111/pbi.2016.14.issue-4
Govaerts, R., Nic Lughadha, E., Black, N., Turner, R., and Paton, A. (2021). The World Checklist of Vascular Plants, a continuously updated resource for exploring global plant diversity. Sci. Data 8, 215. doi: 10.1038/s41597-021-00997-6
Guiry, M. D. and Guiry, G. M. (2025). AlgaeBase (University of Galway, Galway, Ireland). Available at: https://www.algaebase.org (Accessed March 09, 2025).
Hao, F., Liu, X., Zhou, B., Tian, Z., Zhou, L., Zong, H., et al. (2023). Chromosome-level genomes of three key Allium crops and their trait evolution. Nat. Genet. 55, 1976–1986. doi: 10.1038/s41588-023-01546-0
Hao, L., Xu, W., Qi, G., Xin, T., Xu, Z., Lei, H., et al. (2022). GAGE is a method for identification of plant species based on whole genome analysis and genome editing. Commun. Biol. 5, 947. doi: 10.1038/s42003-022-03894-9
Hassler, M. (2025). “Checklist of ferns and lycophytes of the world,” in Catalogue of life. Eds. Bánki, O., Roskov, Y., Döring, M., Ower, G., Hernández Robles, D. R., Plata Corredor, C. A., et al. (Amsterdam, Netherlands: Catalogue of life partners).
Hufford, M. B., Seetharam, A. S., Woodhouse, M. R., Chougule, K. M., Ou, S., Liu, J., et al. (2021). De novo assembly, annotation, and comparative analysis of 26 diverse maize genomes. Science 373, 655–662. doi: 10.1126/science.abg5289
Jackson, S. A., Iwata, A., Lee, S.-H., Schmutz, J., and Shoemaker, R. (2011). Sequencing crop genomes: approaches and applications: Tansley review. New Phytol. 191, 915–925. doi: 10.1111/j.1469-8137.2011.03804.x
Jayakodi, M., Golicz, A. A., Kreplak, J., Fechete, L. I., Angra, D., Bednář, P., et al. (2023). The giant diploid faba genome unlocks variation in a global protein crop. Nature 615, 652–659. doi: 10.1038/s41586-023-05791-5
Jayakodi, M., Padmarasu, S., Haberer, G., Bonthala, V. S., Gundlach, H., Monat, C., et al. (2020). The barley pan-genome reveals the hidden legacy of mutation breeding. Nature 588, 284–289. doi: 10.1038/s41586-020-2947-8
Jiao, W.-B. and Schneeberger, K. (2017). The impact of third generation genomic technologies on plant genome assembly. Curr. Opin. Plant Biol. 36, 64–70. doi: 10.1016/j.pbi.2017.02.002
Jiao, C., Xie, X., Hao, C., Chen, L., Xie, Y., Garg, V., et al. (2024). Pan-genome bridges wheat structural variations with habitat and breeding. Nature 637, 384–393. doi: 10.1038/s41586-024-08277-0
Kersey, P. J. (2019). Plant genome sequences: past, present, future. Curr. Opin. Plant Biol. 48, 1–8. doi: 10.1016/j.pbi.2018.11.001
Khan, A. W., Garg, V., Roorkiwal, M., Golicz, A. A., Edwards, D., and Varshney, R. K. (2020). Super-pan genome by integrating the wild side of a species for accelerated crop improvement. Trends Plant Sci. 25, 148–158. doi: 10.1016/j.tplants.2019.10.012
Kress, W. J., Soltis, D. E., Kersey, P. J., Wegrzyn, J. L., Leebens-Mack, J. H., Gostel, M. R., et al. (2022). Green plant genomes: What we know in an era of rapidly expanding opportunities. Proc. Natl. Acad. Sci. U.S.A. 119, e2115640118. doi: 10.1073/pnas.2115640118
Michael, T. P. and Jackson, S. (2013). The first 50 plant genomes. Plant Genome 6, eplantgenome2013.03.0001in. doi: 10.3835/plantgenome2013.03.0001in
Pellicer, J. and Leitch, I. J. (2020). The Plant DNA C-values database (release 7.1): an updated online repository of plant genome size data for comparative studies. New Phytol. 226, 301–305. doi: 10.1111/nph.v226.2
Plants of the World Online (POWO). (2025). Facilitated by the royal botanic gardens (Kew, London, UK). Available at: https://powo.science.kew.org/.
Pucker, B., Irisarri, I., de Vries, J., and Xu, B. (2022). Plant genome sequence assembly in the era of long reads: Progress, challenges and future directions. Quant. Plant Biol. 3, e5. doi: 10.1017/qpb.2021.18
Qin, P., Lu, H., Du, H., Wang, H., Chen, W., Chen, Z., et al. (2021). Pan-genome analysis of 33 genetically diverse rice accessions reveals hidden genomic variations. Cell 184, 3542–3558. doi: 10.1016/j.cell.2021.04.046
Rautiainen, M., Nurk, S., Walenz, B. P., Logsdon, G. A., Porubsky, D., Rhie, A., et al. (2023). Telomere-to-telomere assembly of diploid chromosomes with Verkko. Nat. Biotechnol. 41, 1474–1482. doi: 10.1038/s41587-023-01662-6
Schoch, C. L., Ciufo, S., Domrachev, M., Hotton, C. L., Kannan, S., Khovanskaya, R., et al. (2020). NCBI Taxonomy: a comprehensive update on curation, resources and tools. Database (Oxford) 2020, baaa062. doi: 10.1093/database/baaa062
Shirasawa, K., Harada, D., Hirakawa, H., Isobe, S., and Kole, C. (2021). Chromosome-level de novo genome assemblies of over 100 plant species. Breed. Sci. 71, 117–124. doi: 10.1270/jsbbs.20146
Sun, Y., Shang, L., Zhu, Q.-H., Fan, L., and Guo, L. (2022). Twenty years of plant genome sequencing: achievements and challenges. Trends Plant Sci. 27, 391–401. doi: 10.1016/j.tplants.2021.10.006
Talbot, S. C., Pandelova, I., Lange, B. M., and Vining, K. J. (2024). A first look at the genome structure of hexaploid “Mitcham” peppermint (Mentha × piperita L.). G3 (Bethesda) 14, jkae195. doi: 10.1093/g3journal/jkae195
Tang, D., Jia, Y., Zhang, J., Li, H., Cheng, L., Wang, P., et al. (2022). Genome evolution and diversity of wild and cultivated potatoes. Nature 606, 535–541. doi: 10.1038/s41586-022-04822-x
Tariq, A., Meng, M., Jiang, X., Bolger, A., Beier, S., Buchmann, J. P., et al. (2024). In-depth exploration of the genomic diversity in tea varieties based on a newly constructed pangenome of Camellia sinensis. Plant J. 119, 2096–2115. doi: 10.1111/tpj.v119.4
The Angiosperm Phylogeny Group. (2016). An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc 181, 1–20. doi: 10.1111/boj.2016.181.issue-1
The Arabidopsis Genome Initiative (2000). Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408, 796–815. doi: 10.1038/35048692
The Pteridophyte Phylogeny Group. (2016). A community-derived classification for extant lycophytes and ferns: PPG I. J. Syst. Evol. 54, 563–603. doi: 10.1111/jse.12229
van Rengs, W. M. J., Schmidt, M. H.-W., Effgen, S., Le, D. B., Wang, Y., Zaidan, M. W. A. M., et al. (2022). A chromosome scale tomato genome built from complementary PacBio and Nanopore sequences alone reveals extensive linkage drag during breeding. Plant J. 110, 572–588. doi: 10.1111/tpj.v110.2
Vignale, F. A., Hernadez Garcia, A., Modenutti, C. P., Sosa, E. J., Defelipe, L. A., Oliveira, R., et al. (2025). Yerba mate (Ilex paraguariensis) genome provides new insights into convergent evolution of caffeine biosynthesis. Elife 14, e104759. doi: 10.7554/eLife.104759
Wang, H., Caetano-Andrade, V., Boivin, N., Clement, C. R., Ayala, W. E., Melinski, R. D., et al. (2025). Long-term human influence on the demography and genetic diversity of the hyperdominant Bertholletia excelsa in the Amazon Basin. Curr. Biol. 35, 629–639. doi: 10.1016/j.cub.2024.12.023
Xie, L., Gong, X., Yang, K., Huang, Y., Zhang, S., Shen, L., et al. (2024). Technology-enabled great leap in deciphering plant genomes. Nat. Plants 10, 551–566. doi: 10.1038/s41477-024-01655-6
Xu, S., Chen, R., Zhang, X., Wu, Y., Yang, L., Sun, Z., et al. (2024). The evolutionary tale of lilies: Giant genomes derived from transposon insertions and polyploidization. Innovation (Camb.) 5, 100726. doi: 10.1016/j.xinn.2024.100726
Yang, Y., Ferguson, D. K., Liu, B., Mao, K. S., Gao, L. M., Zhang, S. Z., et al. (2022). Recent advances on phylogenomics of gymnosperms and a new classification. Plant Divers. 44, 340–350. doi: 10.1016/j.pld.2022.05.003
Zhou, Y., Zhang, Z., Bao, Z., Li, H., Lyu, Y., Zan, Y., et al. (2022). Graph pangenome captures missing heritability and empowers tomato breeding. Nature 606, 527–534. doi: 10.1038/s41586-022-04808-9
Ziegler, F. M. R., Rosenthal, V., Vallarino, J. G., Genzel, F., Spettmann, S., Seliga, Ł., et al. (2024). A full genome assembly reveals drought stress effects on gene expression and metabolite profiles in blackcurrant (Ribes nigrum L.). Hortic. Res. 12, uhae313. doi: 10.1093/hr/uhae313
Keywords: sequenced plant genomes, plant genome publication, timeline, cladogram, Archaeplastida
Citation: Schwacke R, Bolger ME and Usadel B (2025) PubPlant – a continuously updated online resource for sequenced and published plant genomes. Front. Plant Sci. 16:1603547. doi: 10.3389/fpls.2025.1603547
Received: 31 March 2025; Accepted: 02 June 2025;
Published: 24 June 2025.
Edited by:
Guang-Long Wang, Huaiyin Institute of Technology, ChinaReviewed by:
Aureliano Bombarely, Polytechnic University of Valencia, SpainKang Zhang, Chinese Academy of Agricultural Sciences, China
Copyright © 2025 Schwacke, Bolger and Usadel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rainer Schwacke, ci5zY2h3YWNrZUBmei1qdWVsaWNoLmRl