 Jéssica P. Ferreira
Jéssica P. Ferreira Márcia S. Vidal
Márcia S. Vidal José I. Baldani
José I. Baldani- 1Department of Crop Sciences, Crop Sciences Graduate Program, Federal Rural University of Rio de Janeiro (UFRRJ), Institute of Agronomy, Seropédica, Rio de Janeiro, Brazil
- 2Genetics and Biochemistry Laboratory, Embrapa Agrobiologia, Seropédica, Rio de Janeiro, Brazil
Plant growth-promoting bacteria (PGPB) possessing 1-aminocyclopropane-1-carboxylate (ACC) deaminase activity have the potential to enhance plant growth and development, particularly under adverse environmental conditions. This study aimed to identify bacterial strains with ACC deaminase activity able of mitigating the effects of water deficit stress and promoting the growth of Brachiaria genotypes. Bacterial strains isolated from Brachiaria genotypes were screened in vitro for ACC deaminase activity, and the presence of the acdS gene was confirmed via polymerase chain reaction (PCR) analysis. The bacterial isolates were screened for in vitro tolerance to water deficit stress, using 10% polyethylene glycol 8000 (PEG 8000) in association with B. ruziziensis and the effects of bacterial inoculation were assessed based on plant height and fresh biomass accumulation. Additionally, the association between endophytic bacterial strains and Brachiaria genotypes was evaluated using confocal laser microscope. The results showed that among the 213 strains tested, 32 demonstrate the ability to degrade ACC into α-ketobutyrate. ACC deaminase activity was detected in 17 strains, with values ranging from 1.98 to 102.52 μmol α-ketobutyrate mg-1 protein h-1. The presence of the acdS gene was confirmed in nine strains. The strains NRB142 (Paraburkholderia silvatlantica), NRB223 (Azospirillum melinis), and BR11790 (Herbaspirillum frisingense GSF30T) exhibited the most significant promotion of plant development in B. ruziziensis under water deficit stress mediated by 10% PEG 8000. Confocal microscopy analysis revealed the rhizospheric and inner root colonization of B. ruziziensis and B. brizantha cv. Paiaguás by the NRB142 mCherry-labeled strain. This study showed no predominance of a specific group of bacterial strains in terms of ACC deaminase activity. However, a subset of strains demonstrated the ability to colonize Brachiaria plants and mitigate the negative effects of water deficit stress. This study highlights the potential of ACC deaminase-producing bacteria in alleviating water deficit stress in Brachiaria plants supporting their use as a promising strategy for improving plant resilience under drought conditions.
1 Introduction
The Brazilian cattle herd is estimated to be the second largest in the world, with approximately 186.8 million head in 2025 (USDA, 2025). The herd is predominantly fed through free grazing on pastures, which remains the most economical and practical method of providing nutrition for livestock (Jank et al., 2014; Duarte et al., 2020). Among the pasture areas in Brazil, it is estimated that approximately 85% are occupied by plants of the Brachiaria genus (Jank et al., 2014).
Abiotic stresses are recognized as a primary influence impacting agricultural production globally (Shahid et al., 2023). Among abiotic stresses, water stress is the one that most frequently affects pasture productivity (Moore et al., 2020). Water is vital for plant development (Shao et al., 2008), and its scarcity can disrupt growth cycles, leading to yield reductions exceeding 50% (Boyer, 1982; Lisar et al., 2012). Water stress impacts various levels of plant organization (Yordanov et al., 2000), altering water potential, turgor pressure, nutrient transport, and gas exchange. Increased abscisic acid levels cause stomatal closure, inhibiting photosynthesis. This leads to the accumulation of Reactive Oxygen Species (ROS), causing cellular damage such as DNA lesions, protein synthesis inhibition, pigment oxidation, and membrane deterioration. The decline in chlorophyll content, often linked to oxidative stress, further hampers photosynthetic efficiency (Anjum et al., 2011; Taiz and Zeiger, 2017; Vurukonda et al., 2016). Additionally, water deficit compromises cell wall and membrane integrity, leading to cell death (Ali et al., 2025). These effects reduce plant growth, accelerate senescence, decrease dry matter production, and heighten susceptibility to diseases and pests, ultimately diminishing crop quality and yield (Li et al., 2009; Seleiman et al., 2021).
The application of microbial inoculants containing plant growth-promoting bacteria (PGPB) presents a cost-effective and environmentally sustainable solution to mitigate water deficit stress in crops (Poudel et al., 2021; Armanhi et al., 2021). PGPB enhance plant growth while offering protection against diseases and abiotic stresses, including drought, salinity, and nutrient imbalances (Dimkpa et al., 2009; Grover et al., 2011; Glick, 2012, 2015). In this context, the use of microorganisms is important due to their low cost and eco-friendly nature make them an attractive option for sustainable agriculture. Additionally, PGPB contribute to overall plant health, increase productivity and quality, and help maintain soil integrity (Zhang et al., 2019). Harnessing their potential can lead to more resilient crop systems, reducing the adverse impacts of water deficit stress while supporting sustainable farming practices.
Plant growth-promoting bacteria enhance plant tolerance to water stress through several key mechanisms. They improve nutrient availability via biological nitrogen fixation, phosphate solubilization, and siderophore production which aids iron uptake (Ali and Khan, 2021; Sati et al., 2023). Additionally, PGPB produce exopolysaccharides, which enhance soil structure, water retention, and root stability (Ali and Khan, 2021; Sati et al., 2023). They also regulate osmotic balance and activate antioxidant defenses, mitigating reactive oxygen species (ROS) damage and protecting biomolecules from oxidative stress (Grover et al., 2011; Sati et al., 2023). Furthermore, PGPB facilitates the synthesis of heat shock proteins (HSPs), dehydrins, and volatile organic compounds (VOCs), which assist in drought tolerance and adaptation (Kaushal and Wani, 2016). Their influence on phytohormone production, including auxins, gibberellins, cytokinins, abscisic acid, and ethylene, helps regulate plant growth under water deficit conditions (Kaushal and Wani, 2016).
Ethylene stress, triggered by abiotic and biotic factors, adversely affects plant growth, causing senescence, abscission, chlorophyll loss, and developmental inhibition (Pei et al., 2017; Glick et al., 2007a; Depaepe and van der Straeten, 2016; Kaushal and Wani, 2016). Plant growth-promoting bacteria (PGPB) expressing 1-aminocyclopropane-1-carboxylate (ACC) deaminase counteract excessive ethylene by cleaving ACC (the immediate precursor of ethylene) into ammonia and α-ketobutyrate (Glick et al., 1998). The acdS gene encoding ACC deaminase has been identified in diverse organisms within the Eukarya, Bacteria, and Archaea domains. However, it predominantly occurs in various bacterial species and some fungi (Singh et al., 2015; Soni et al., 2018). The products of enzymatic cleavage, ammonia and α-ketobutyrate, serve as carbon and nitrogen sources for bacterial metabolism (Glick and Nascimento, 2021). Additionally, α-ketobutyrate influences the transcriptional regulation of acdS, a gene involved in ethylene modulation, through its role in leucine biosynthesis (Soni et al., 2018). The acdS gene is regulated by AcdR (Lrp), a leucine-responsive protein, and co-regulated by AcdB, which forms a complex with ACC and Lrp, enabling acdS transcription (Glick et al., 2007a; Li and Glick, 2001; Cheng et al., 2008). Upon ACC deaminase synthesis, ACC is cleaved, generating ammonia and α-ketobutyrate, which is then used for leucine biosynthesis. As leucine accumulates, it binds to Lrp octamer, causing its dissociation into inactive dimers, ultimately blocking acdS transcription. This regulatory mechanism ensures that ACC deaminase is produced only when required (Cheng et al., 2008; Duan et al., 2013; Glick et al., 2007b; Grichko and Glick, 2000; Li and Glick, 2001). Therefore, this enzymatic action may help mitigate the stress effects of ethylene on plants colonized by these bacteria, enhancing growth by reducing its inhibitory influence. The fate of these byproducts seems to be important in microbial interactions with plants, supporting healthier development in challenging environments.
The increasing frequency of droughts due to climate change has intensified research on plant interactions with ACC deaminase-producing bacteria. Chandra et al. (2019, 2020) found that Variovorax paradoxus and a consortium of Ochrobactrum anthropi, Pseudomonas palleroniana, and Pseudomonas fluorescens improved millet and wheat growth under 35% water stress, increasing leaf nutrient concentrations and antioxidant properties. Similarly, Tahir et al. (2019) reported that corn inoculated with BN-5 and MD-23 strains showed enhanced grain yield, relative water content, and chlorophyll levels under 50% field capacity. Likewise, Tiwari et al. (2018) analyzed drought and saline stress mitigation in Panicum maximum inoculated with ACC deaminase-producing rhizobacteria, and observed improving membrane stability, solute accumulation, and photosynthetic pigments while Ojuederie and Babalola (2023) found that Pseudomonas sp. MRBP4, MRBP13, and Bacillus sp. MRBP10 strains enhanced maize drought tolerance by improving water retention and biomass production. Similarly, Jasso-Arreola et al. (2025) demonstrated that Pantoea sp. RCa62, isolated from Coffea arabica, increased leaf area, root development, and relative water content while reducing proline accumulation. Other studies have confirmed similar benefits in various crops, including tomato (Muñoz-Carvajal et al., 2024), soybean (Dubey et al., 2024), black gram (Chandwani and Amaresan, 2024), watermelon (Yavuz et al., 2023), and cluster bean (Jain and Saraf, 2023). These findings reinforce the potential of ACC deaminase-producing bacteria as a biological strategy for mitigating drought stress and promoting sustainable agriculture.
Therefore, this study aimed to identify diazotrophic bacteria with ACC deaminase activity that can alleviate water deficit stress and promote the growth of Brachiaria genotypes under stress conditions induced by polyethylene glycol 8000 (PEG 8000). Additionally, it assessed the ability of the selected bacterium to colonize and establish in Brachiaria ruziziensis plants grown in vitro under PEG 8000 treatment.
2 Materials and methods
2.1 Qualitative screening for bacterial strains with ACC deaminase activity
The screening for ACC deaminase activity in the strains was conducted following the approach described by Glick et al. (1995), with minor modifications. The study involved the analysis of 213 diazotrophic strains isolated from various Brachiaria genotypes as part of the Embrapa project (number 02.13.08.004.00.00). The isolation, the taxonomic and partial functional characterization of these 213 strains were previously reported by Ribeiro et al. (2020).
The strains were cultivated in 5 mL of DYGS medium (Rodrigues Neto et al., 1986) and incubated at 30°C, 180 rpm, for 24 or 48 h, depending on the bacterial growth rate. Following incubation, 100 µL of the culture was transferred to new tubes containing 5 mL of LGI or NFb medium (Baldani et al., 2014) supplemented with 1 g L⁻¹ of (NH4)2SO4 as a nitrogen source and incubated under the same conditions. Afterward, 100 µL of the second-round culture was transferred to fresh tubes containing 5 mL of LGI or NFB medium, but without the nitrogen source. The culture medium was then supplemented with 3 mmol L⁻¹ ACC and incubated under the previously described conditions.
Petri dishes containing Noble Agar (low nitrogen content) were supplemented with 3 mmol L⁻¹ ACC from a filtered sterilized stock solution (0.5 mol L⁻¹), which was evenly spread over the surface of the culture medium. Cultures were inoculated using a sterile cotton swab and incubated at 30°C for 48 or 60 h, depending on growth conditions. For the negative control, cultures were plated on LGI or NFb media without the addition of inorganic nitrogen or ACC substrate. A diazotrophic Herbaspirillum frisingense strain GSF30T, recognized for its ACC deaminase activity, was used as a positive control (Rothballer et al., 2008).
2.2 Quantification of ACC deaminase activity produced by the pre-selected strains
The activity of ACC deaminase was assessed using the method described by Penrose and Glick (2003), which quantifies the α-ketobutyrate produced through the cleaved by the enzyme ACC deaminase. The quantification process involved measuring the absorbance of bacterial sample at 540 nm and comparing the results to a standard α-ketobutyrate curve ranging from 10 to 1000 µmol. To determine the specific activity of the cultures, protein concentration was measured using the Bradford method (Bradford, 1976).
The pre-selected strains (qualitative assays) were cultured overnight in 5 mL of DYGS medium at 30°C, 180 rpm, for 24 h. After incubation, the cells were harvested by centrifugation at 5,000 xg for 10 min at 4°C, followed by washing with NFb or LGI medium (without a nitrogen source). The bacterial pellet was then resuspended in 5 mL of NFb or LGI medium supplemented with 3 mmol L⁻¹ ACC as the sole nitrogen source. The culture was incubated for 24 h with shaking at 180 rpm at 30°C. Subsequently, the bacterial cells were harvested again by centrifugation at 5,000 xg, 4°C, for 10 min. The cells were washed twice with 5 mL of 0.1 mol L⁻¹ Tris-HCl buffer (pH 7.6). Finally, the cell suspension was transferred to a microcentrifuge tube and centrifuged at 10,000 g for 1 min.
All the supernatant was carefully removed, and the cell pellet was utilized for the enzymatic assay. The pellets were resuspended in 400 μL of 0.1 mol L⁻¹ Tris-HCl buffer (pH 8.0), followed by the addition of 20 μL of toluene and vortexing for 30 s. Subsequently, 50 μL of the toluene-treated cells were incubated with 5 μL of 0.5 M ACC at 30°C for 30 min. After incubation, 500 µL of 0.56 M HCl was added, and the mixture was vortexed and centrifuged at 10,000 xg for 5 min at room temperature. The resulting supernatant (500 μL) was vortexed with 400 μL of 0.56 M HCl and 150 μL of 2,4-dinitrophenylhydrazine reagent (0.2% 2,4-dinitrophenylhydrazine in 2 M HCl). The mixture was incubated at 30°C for 30 min, followed by the addition of 1 mL of 2 M NaOH and thorough mixing. Absorbance at 540 nm was then measured using a spectrophotometer. The cell suspension without ACC served as the negative control. Specific activity of the cultures was determined by protein quantification following the Bradford method (Bradford, 1976). ACC deaminase activity was expressed as μmol of α-ketobutyrate per mg of protein per hour.
2.3 Detection of the acdS sequence in the bacterial strains
The positive strains in the qualitative screening and those that showed results in the quantification of ACC deaminase enzyme activity were used in the detection of the acdS sequence. Genomic DNA was extracted using the commercial Wizard® Genomic DNA Purification Kit (Promega, Madison, USA) following the manufacturer’s instructions. The concentration of genomic DNA was evaluated using a Nanodrop® 3300 spectrophotometer (Thermo Fisher Scientific Inc., Waltham, USA). PCR reactions were performed using two pairs of primers described by Li et al. (2015): acdSf3 (5′ – ATCGGCGGCATCCAGWSNAAYCANAC – 3′), acdSr3 (5′ – GTGCATCGACTTGCCCTCRTANACNGGRT – 3′), and acdSr4 (5′ – GGCACGCCGCCCARRTGNRCRTA – 3′). Each amplification reaction was conducted in a final volume of 25 μL, consisting of 20 ng µL⁻¹ genomic DNA, 1× Taq DNA polymerase buffer (1 mM Tris-HCl, pH 9.0, and 5 mM KCl), 0.5 mM of each dNTP, 3 mM MgCl2, 0.4 μM of each primer, and 0.1 U µL⁻¹ Taq DNA polymerase (Promega, Madison, USA). Negative control samples were prepared by replacing bacterial DNA with ultrapure water. Amplification reactions were carried out in a SureCycler 8800 thermocycler (Agilent Technologies, Santa Clara, USA) programmed for initial denaturation at 94°C for 4 min; 35 cycles at 94°C for 45 s, 53°C for 45 s, and 72°C for 1 min; followed by a final extension at 72°C for 10 min. After amplification, 2 μL of PCR product was analyzed through electrophoresis on a 1.5% (w/v) agarose gel at 90 volts (~5 V/cm) for 1 h and 30 min in 1× TAE buffer (40 mM Tris-acetate, pH 8.0, and 1 mM EDTA, pH 8.0). The gel was stained with ethidium bromide solution (0.5 μg mL⁻¹) and visualized under ultraviolet light using a KODAK® Gel Logic Cabinet 100 photoceller (Eastman Kodak Company, Rochester, USA).
2.4 In vitro response of B. ruziziensis to inoculation with diazotrophic ACC deaminase-producing strains under stress conditions
A preliminary experiment was conducted to determine the optimal PEG 8000 concentration for in vitro studies. The application of 20% PEG 8000 was highly detrimental, leading to the death of nearly all plants (data not shown). Based on these results, a subsequent experiment was performed using a reduced concentration of 10% PEG 8000.
The gnotobiotic inoculation experiment was conducted using disinfested seeds of Brachiaria ruziziensis, a genotype with low tolerance to water deficit stress. The seeds were peeled and sterilized by washing in 70% (v/v) ethanol for 3 min, followed by immersion in sodium hypochlorite (4–6% v/v free chlorine) with agitation for 10 min. After that, the seeds were rinsed three times with sterile distilled water and placed in Petri dishes containing an agar/water medium (0.5% agar supplemented with 500 mg L⁻¹ of yeast extract). The plates were initially incubated in the dark at 30°C for 24 h. Subsequently, they were transferred to a BOD incubator (model LB41, LABTEC, Londrina, Paraná, Brazil) and maintained at 30°C with a 12-h photoperiod for 4 days to ensure complete germination.
Meanwhile, the bacterial strains were inoculated in 50 mL of liquid DYGS medium and incubated under agitation at 180 rpm and 30°C, for 24 or 48 h, depending on the bacterial growth rate. Bacterial growth was quantified using the micro drop technique (Romeiro, 2007). The inoculum concentrations obtained were 104 CFU mL⁻¹ for strain BR11790 and 105 CFU mL⁻¹ for the other target diazotrophic bacteria. Non-contaminated seedlings were carefully removed from the agar/water medium and immersed for 1 h in the bacterial culture suspension of their respective strains: NRB032 (Stenotrophomonas maltophilia), NRB039 (Nitrospirillum amazonense), NRB058 (Pseudomonas cremoricolorata), NRB096 (Bacillus safensis), NRB123 (N. amazonense), NRB124 (Paraburkholderia silvatlantica), NRB127 (Herbaspirillum seropedicae), NRB138 (Gluconacetobacter diazotrophicus), NRB142 (P. silvatlantica), NRB223 (Azospirillum melinis) and BR11790 (H. frisingense GSF30T). These strains were employed as they showed positive results in the qualitative screening and demonstrated activity in the quantification of the ACC deaminase enzyme. A Herbaspirillum frisingense strain GSF30T was used as a positive control. The seedlings assigned to the control treatment were immersed in flasks containing the same volume of DYGS liquid medium for the same duration. Afterward, the inoculated seedlings were transferred to glass tubes containing 25 mL of MS medium (Murashige and Skoog, 1962) supplemented with 30 g L⁻¹ of sucrose, with the pH adjusted to 5.8. The medium was prepared both with and without 10% PEG 8000. The plants were then placed in a growth room and maintained for 30 days under a photoperiod of 16 h of light and 8 h of darkness, at a constant temperature of 25°C.
One experiment was conducted in a completely randomized design with four replications. The factors included stress induction mediated by PEG 8000 (present or absent), two seed treatments (inoculation with 11 diazotrophic strains exhibiting ACC deaminase activity and a control), and one Brachiaria genotype (B. ruziziensis). Each experimental unit consisted of a 100 mL glass tube containing 25 mL of MS medium. Analyses were performed 30 days post-inoculation by measuring plant height and the fresh biomass accumulation of leaves and roots. To compare treatment means, the Scott-Knott test was applied at a significant level of 0.05. All statistical analyses were conducted using the software ‘Sisvar’ version 5.3 (Ferreira, 2011).
2.5 Assessment of bacterial colonization in B. brizantha cv. Paiaguás and B. ruziziensis
To assess the Brachiaria plant colonization, a red-fluorescent derivative of NRB142 (P. silvatlantica) was constructed via transformation with plasmid pLMB426 applying the electroporation method. The transformed strain, designated NRB142 (mCherry), was cultivated in liquid or solid DYGS medium supplemented with gentamycin (80 μg mL⁻¹). Plants of B. brizantha cv. Paiaguás and B. ruziziensis with 5 days after germination were inoculated with NRB142 (mCherry). This strain was selected because it exhibited the highest performance in ACC deaminase activity quantification and showed beneficial effects in the in vitro test with PEG 8000. The gnotobiotic inoculation experiment utilized disinfected seeds of B. brizantha cv. Paiaguás and B. ruziziensis, as described in the previous section.
After germination, microorganism-free plants were removed from agar/water medium plates and transferred to glass tubes containing 25 mL of MS medium (Murashige and Skoog, 1962) supplemented with 30 g L⁻¹ sucrose and adjusted to pH 5.8 for rooting. Plants were maintained in MS medium for 30 days to promote root formation before being transferred to flasks containing 25 mL of Hoagland’s solution. Prior to transfer, inoculation with NRB142 (mCherry) was conducted in tubes designed for the inoculated treatment, using bacterial suspensions prepared at a concentration of 105 CFU mL⁻¹. Bacterial growth was quantified using a Neubauer chamber.
Control treatment tubes were inoculated with PBS buffer in volumes equal to the bacterial solution. The experiment followed a completely randomized design with four replications, considering two experimental factors: inoculation with or without NRB142 (mCherry) and two Brachiaria cultivars (B. brizantha cv. Paiaguás and B. ruziziensis). Each experimental unit consisted of a 100 mL glass tube containing 25 mL of MS medium (Murashige and Skoog, 1962).
Plants were maintained in a growth chamber for 30 days under a controlled photoperiod of 16 h of light and 8 h of darkness at 25°C. Harvests were performed at 3, 7, and 14 days after inoculation (dai). Endophytic and rhizospheric bacterial populations were quantified using the micro-drop technique (Romeiro, 2007). Confocal microscopy images were obtained using the LSM 700 microscope, AxioObserver (Carl Zeiss, Jena, Germany), and processed with Zen 2.3 software (Carl Zeiss, Jena, Germany).
3 Results
3.1 Screening for bacterial strains with ACC deaminase activity
The methodology adapted from Glick et al. (1995) was initially used to assess the presence of ACC deaminase activity in bacterial strains. Some strains exhibited growth in LGI or NFb agar plates with ACC as the sole nitrogen source, indicating positive ACC deaminase activity. The results indicated that bacterial growth relied on ACC as its sole nitrogen source, consistent with the methodology described by Glick et al. (1995). Screening of the 213 bacterial strains isolated from Brachiaria genotypes revealed that approximately 15% possessed ACC deaminase activity. Among these, 25% were isolated from rhizospheric soil, 25% from disinfected roots, and 50% from non-disinfected roots (Table 1).
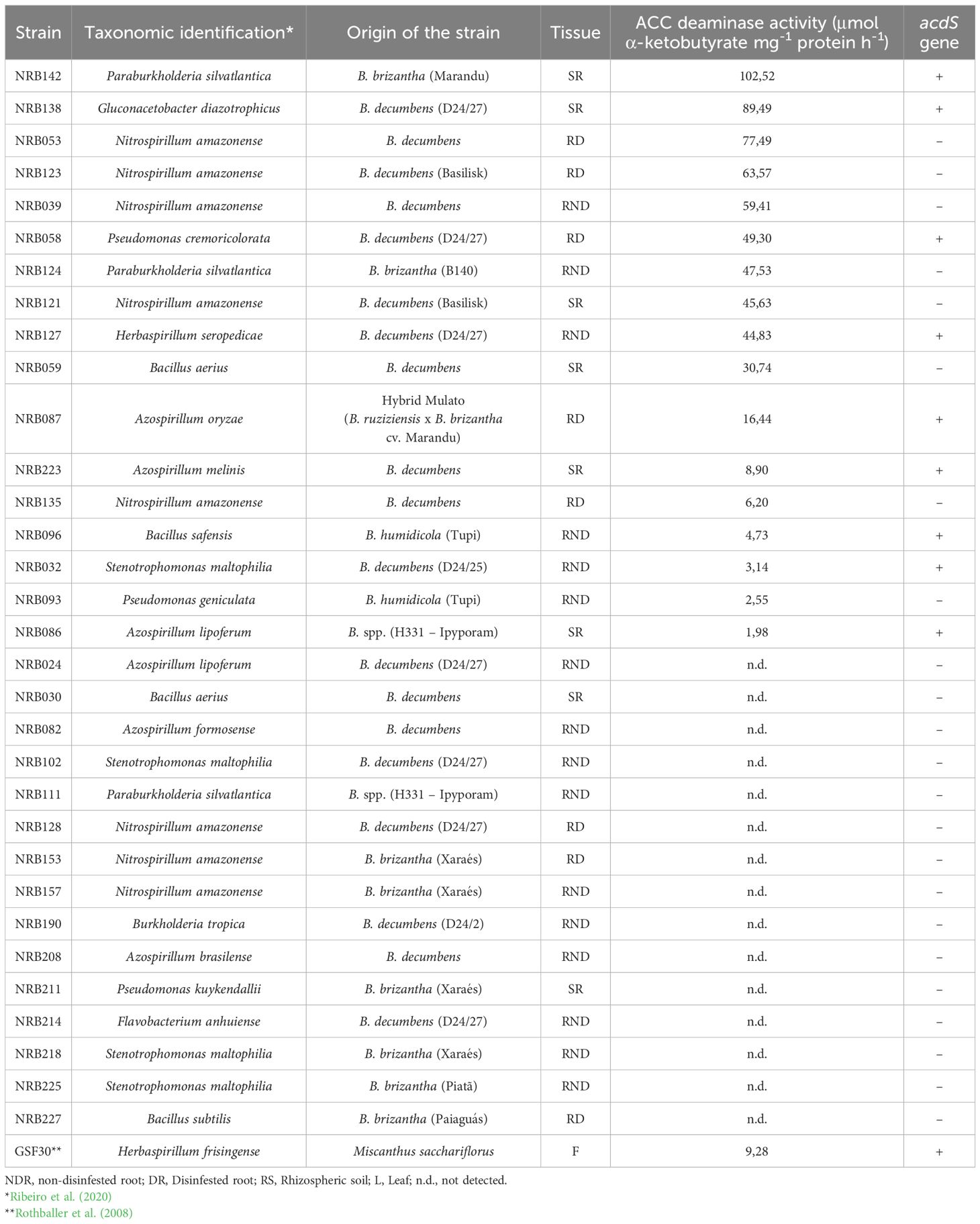
Table 1. Quantification of ACC deaminase activity produced by different strains originally isolated from Brachiaria genotypes.
3.2 Quantification of ACC deaminase activity
The activity of the enzyme ACC deaminase was determined by quantifying the α-ketobutyrate produced during the deamination of ACC by the enzyme. In this work, 32 strains that showed growth capacity in ACC culture medium as the sole nitrogen source were selected to quantify the activity of the ACC deaminase enzyme. The results indicate that 17 out of the 32 strains exhibited ACC deaminase activity in vitro (Table 1), while the remaining 15 strains tested negative for ACC deaminase activity. These findings suggest that growth in a medium with ACC as the sole nitrogen source is not sufficient to confirm that a bacterial strain possesses ACC deaminase activity. Therefore, it is essential to quantify ACC enzyme activity to verify its presence.
As expected, the positive control, H. frisingense GSF30T, exhibited ACC deaminase activity of 9.28 μmol α-ketobutyrate mg-1 protein h-1 in the present assay. Another species of the same genus, H. seropedicae (NRB127), showed higher activity than H. frisingense GSF30T, with a value of 44.83 μmol α-ketobutyrate mg-1 protein h-1. The species P. silvatlantica (NRB142) and G. diazotrophicus (NRB138) showed the highest ACC deaminase activities in vitro, with values of 102.52 and 89.49 μmol α-ketobutyrate mg-1 protein h-1, respectively. Among the results obtained, a group of bacteria presented intermediate ACC deaminase activity values, ranging from 49.0 to 16.0 μmol α-ketobutyrate mg-1 protein h-1. For instance, strain NRB058 (P. cremoricolorata) exhibited an activity of 49.30 μmol α-ketobutyrate mg-1 protein h-1 whereas the strain NRB087 (A. oryzae) showed a lower value of 16.44 μmol α-ketobutyrate mg-1 protein h-1. Regarding the lowest enzymatic activity values, results ranged from 8.90 to 1.98 μmol α-ketobutyrate mg⁻¹ protein h⁻¹. The lowest observed enzymatic activity was recorded for strain NRB086, with a value of 1.98 μmol α-ketobutyrate mg⁻¹ protein h⁻¹.
3.3 Detection of the acdS sequence in the genome of these bacterial strains
The predicted amplified PCR products (~ 680 bp with acdSf3/acdSr3 or ~ 760 bp with acdSf3/acdSr4) were successfully obtained for 9 bacterial strains exhibiting ACC deaminase: NRB032 (S. maltophilia), NRB058 (P. cremoricolorata), NRB086 (A. lipoferum), NRB087 (A. oryzae), NRB096 (B. safensis), NRB127 (H. seropedicae), NRB138 (G. diazotrophicus), NRB142 (P. silvatlantica) and NRB223 (A. melinis). Additionally, amplification was observed for the positive control H. frisigense GSF30T (BR11790). In contrast, no amplification of the acdS gene was detected in the negative control (blank sample). An agarose gel electrophoresis illustrating the respective amplified product is shown in Figure 1.
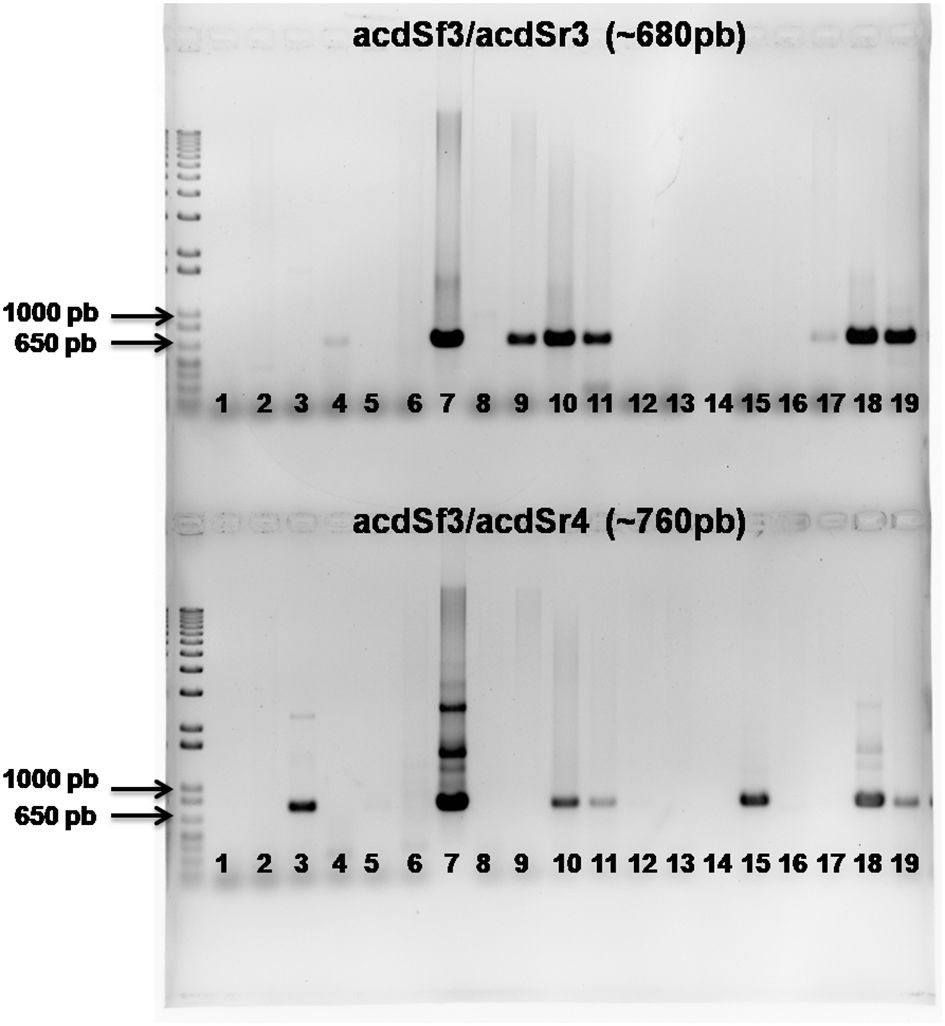
Figure 1. Amplification of the acdS gene from the chromosomal DNA of bacterial strains with ACC deaminase activity. M: Marker 1 kb Plus DNA ladder; Line 1: Water for PCR; Line 2: Negative control; Line 3: BR11790; Line 4: NRB032; Line 5: NRB039; Line 6: NRB053; Line 7: NRB058; Line 8: NRB059; Line 9: NRB086; Line 10: NRB087; Line 11: NRB096; Line 12: NRB121; Line 13: NRB123; Line 14: NRB124; Line 15: NRB127; Line 16: NRB135; Line 17: NRB138; Line 18: NRB142; Line 19: NRB223. The upper part of the gel shows the amplified product using the acdSf3/acdSr3 primers, while the bottom gel shows the combination of primers acdSf3/acdSr4.
3.4 ACC deaminase-producing bacteria mitigating water deficit stress in Brachiaria ruziziensis grown in vitro
The growth of B. ruziziensis in the presence of 10% PEG8000 was considerably decreased compared to control treatment (no PEG8000), which exhibited higher values for the analyzed variables (Table 2). There was a statistically significant difference (p <0.05) was observed in plant size between the control treatment and those subjected to water deficit stress. However, under water deficit stress, inoculation with strains NRB223 (A. melinis), BR11790 (H. frisigense GSF30T), NRB142 (P. silvatlantica), NRB032 (S. maltophilia) and NRB127 (H. seropedicae) lead to an increase in plant size, statistically differing from the other strains and the uninoculated plants. Among these plants inoculated with strain NRB223 (A. melinis) exhibited the greatest height (45.00 cm), while the shortest height (9.00 cm) was recorded in the uninoculated plants.
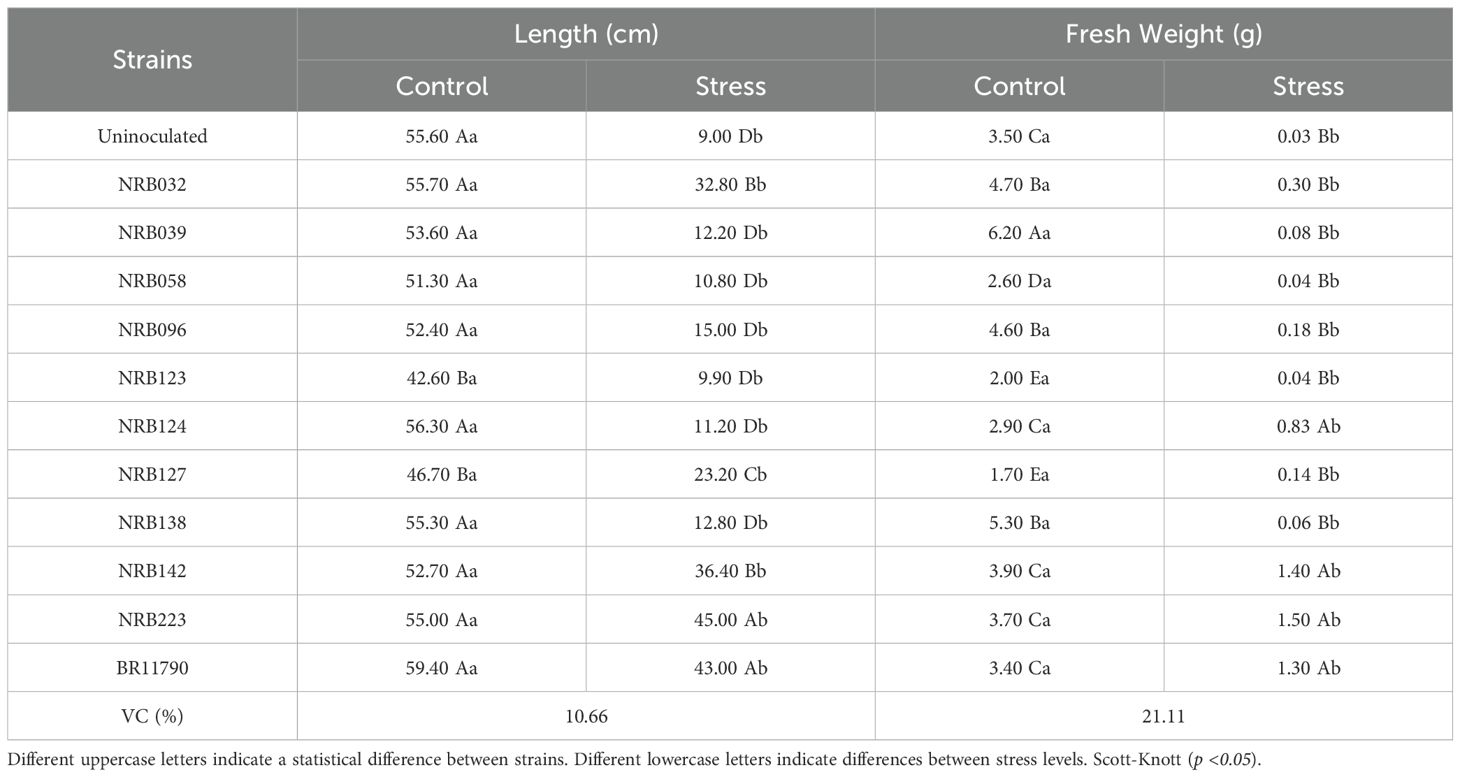
Table 2. Effect of ACC deaminase-producing bacteria inoculated in B. ruziziensis plants subjected to stress induced by polyethylene glycol 8000 - in vitro assay.
In the absence of PEG8000, no statistically significant differences were observed among the inoculated treatments and the control, except for strains NRB123 (N. amazonense) and NRB127 (H. seropedicae), which showed smaller plant sizes (Table 2). Plants inoculated with strains NRB223 (A. melinis), NRB124 (P. silvatlantica), NRB032 (S. maltophilia) showed a higher increase in plant size, reaching up to 59.40 cm. In contrast, plants inoculated with strains NRB127 (H. seropedicae) and NRB123 (N. amazonense) displayed comparatively smaller sizes, measuring 46.70 and 42.60 cm, respectively.
A statistically significant difference (p <0.05) was observed in fresh biomass accumulation both under control (without PEG8000) and in treatments subjected to water deficit stress (with PEG 8000). Under control conditions, plants inoculated with strain NRB039 (N. amazonense) showed the highest fresh biomass accumulation (6.20 g), differing statistically from the other inoculated strains. In contrast, plants inoculated with the strain NRB127 (H. seropedicae) accumulated the lower fresh biomass accumulation (1.70 g). Under water deficit stress, plants inoculated with strain NRB223 (A. melinis) accumulated the highest fresh biomass (1.50 g), followed by those inoculated with strains NRB142 (P. silvatlantica), BR11790 (H. frisigense GSF30T), and NRB124 (P. silvatlantica). These inoculated plants differed statistically from plants inoculated with other strains and the uninoculated plants, which accumulated only 0.03g of biomass.
3.5 Colonization of Brachiaria genotypes by ACC-producing strain NRB142
The ability of strain NRB142 (P. silvatlantica) to colonize seedlings of B. brizantha cv. Paiaguás and B. ruziziensis was investigated under in vitro conditions. The NRB142 strain was selected for this experiment as it demonstrated the highest activity in the quantification of ACC deaminase enzyme activity. Additionally, this strain also contributed to plant tolerance against water deficit stress, induced by PEG in vitro. The strain was successfully labeled with the plasmid harboring the mCherry gene, which remained stable throughout the colonization study, as confirmed by bacterial counting and microscopy analysis conducted at 3, 7, and 14 days after inoculation (dai).
Bacterial counts showed that non-disinfected roots (NDR) presented a higher bacterial colonizing the root system compared to disinfested roots (DR) in both Brachiaria genotypes (Table 3). Differences in bacterial colonization were observed between both genotypes. At 3 dai, the Paiaguás genotype showed a greater bacterial population in both NDR or DR root system. By day 7, this pattern persisted for NDR, while that disinfected roots (DR) of the Ruziziensis genotype showed a higher bacterial population (4 × 104 cells g-1 fresh tissues). By 14 dai, a highest colony forming unit (CFU) g-1 fresh tissue was observed for NDR of the Ruziziensis genotype, whereas DR of the Paiaguás genotype exhibited the highest bacterial count. Despite these differences, results confirmed that strain NRB142 (P. silvatlantica) effectively colonizes both Brachiaria genotypes in numbers relatively high, including the root interior. Despite these differences, results confirm that strain NRB142 effectively colonizes both Brachiaria genotypes, including root interior colonization.
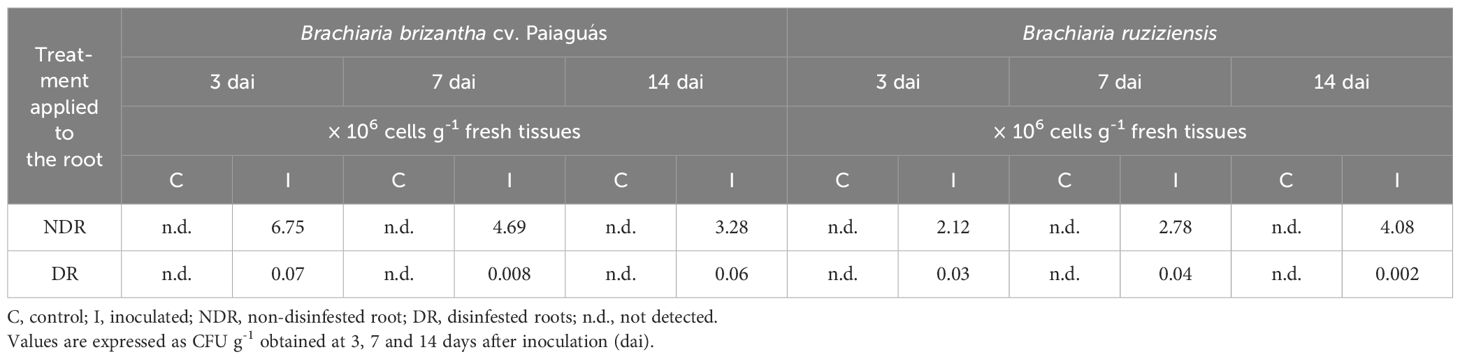
Table 3. Bacterial counting in roots of Brachiaria brizantha cv. Paiaguás and B. ruziziensis inoculated with NRB142(mCherry).
Microscopy analyses further validated these findings, showing significant bacterial aggregations attached to Brachiaria roots. Red fluorescent NRB142 (mCherry) cells were observed colonizing roots of B. brizantha cv. Paiaguás and B. ruziziensis (Figures 2B, D, F, H, J, L). In contrast, no fluorescent bacteria were detected in non-inoculated control plants (Figures 2A, C, E, G, I, K). The bacterial counting (CFU) analysis corroborated these observations, as no bacterial colonies developed on plates containing DYGS culture medium inoculated with dilutions from macerated roots of control plants.
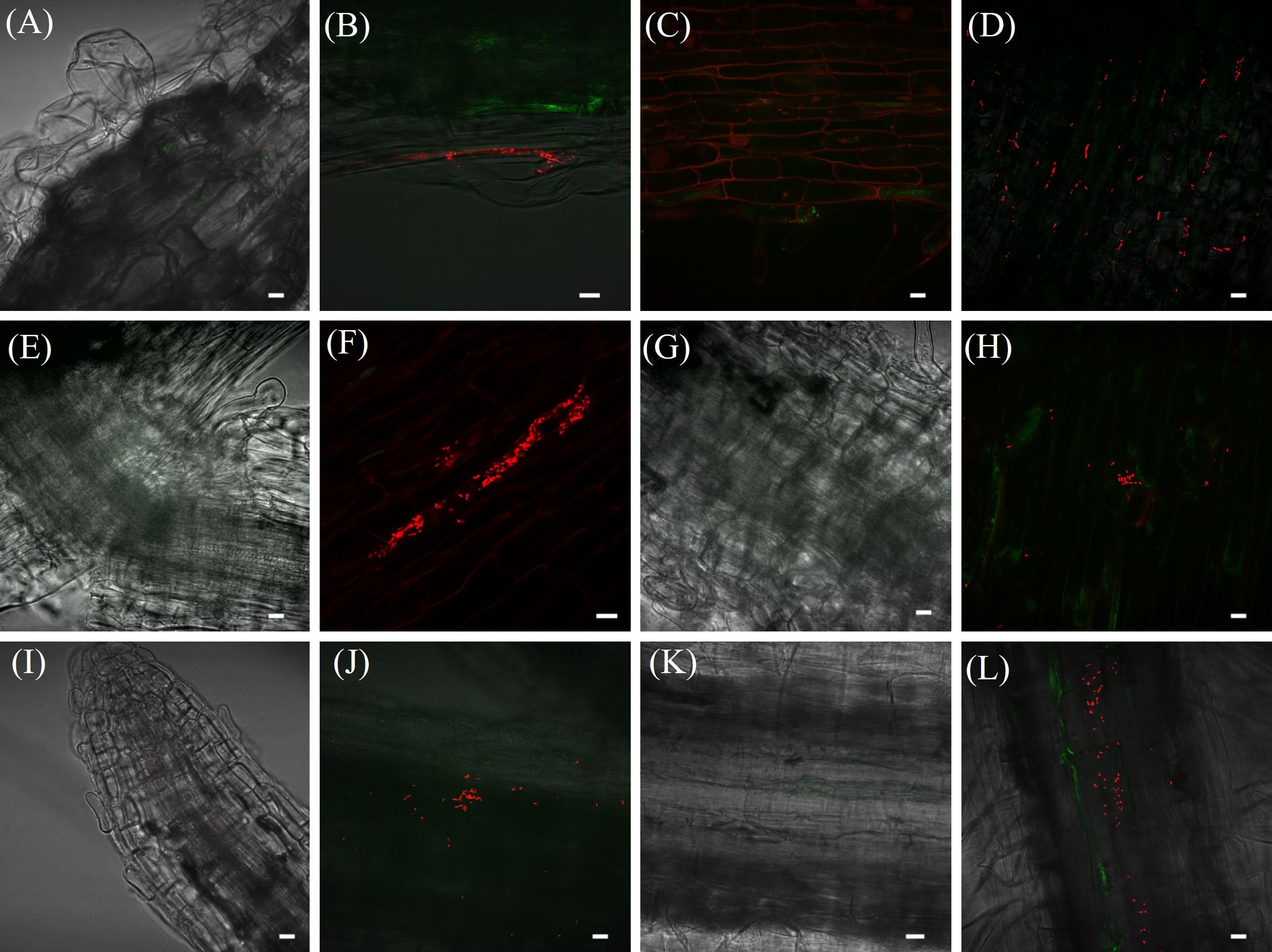
Figure 2. Microscopy images of NRB142 (mCherry) colonizing Brachiaria roots. Photos: uninoculated B. ruziziensis (A, E, I); inoculated B. ruziziensis (B, F, J); uninoculated B. brizantha cv. Paiaguás (C, G, K); inoculated B. brizantha cv. Paiaguás (D, H, L). At 3 [Line (A–D)]; 7 [Line (E–H)]; 14 [Line (I–L)] days after inoculation. Scale bars represent 10 μm.
4 Discussion
The screening for ACC deaminase-producing bacteria revealed a lower number of strains exhibiting positive activity in isolates from rhizospheric soil and disinfected roots compared to those from non-disinfected roots. These findings align with the study by Timmusk et al. (2011) that showed abundance of bacteria producing ACC deaminase in the rhizosphere of Hordeum spontaneum plants and almost null in soil samples. Despite the relatively low percentage of isolates displaying ACC deaminase activity, the results presented here are consistent with other studies. For example, Jalili et al. (2009) using a similar methodology to characterize Pseudomonads species, found that 14% of isolates exhibited ACC deaminase activity - 16% in Pseudomonas putida and 12% in Pseudomonas fluorescens. Likewise, Duraivadivel et al. (2020) observed a low percentage (22.4%) of ACC deaminase-producing bacteria within the total bacterial community associated with Eichhornia crassipes. Gupta and Pandey (2019) reported that approximately 30% of bacterial isolates from the rhizospheric soil of garlic (Allium sativum) exhibited ACC deaminase activity. Similarly, Zhang et al. (2020) identified a small fraction (9%) of bacterial isolates with ACC deaminase activity in the rhizosphere soil of jujube trees.
All tested strains exhibited ACC deaminase activity greater than 20 nmol α-ketobutyrate mg-1 protein h-1, a threshold identified by Penrose and Glick (2003) as sufficient for a bacterium to growth in medium ACC-containing media and potential plant growth-promotion. The highest detected activities ranged between 80 and 100 μmol α-ketobutyrate mg-1 protein h-1. However, Penrose and Glick (2003) also noted that bacteria with elevated ACC deaminase activity (300 to 400 nmol α-ketobutyrate mg-1 protein h-1) do not necessarily stimulate greater root elongation than those bacteria with lower enzyme activity. The results presented here agreed with those found by Li et al. (2011), where Pseudomonas and Herbaspirillum species exhibited ACC deaminase activity comparable to the values observed here. Furthermore, ACC deaminase activity has been increasingly reported in the genus Herbaspirillum (Blaha et al., 2006; Rothballer et al., 2008; Islam et al., 2009; Onofre-Lemus et al., 2009; Sun et al., 2010). Similarly, Niu et al. (2018) identified ACC deaminase activity in Pseudomonas species isolated from millet, though the activity levels (39.40 µmol) were lower than those observed for Pseudomonas cremoricolorata (49.30 μmol α-ketobutyrate mg-1 protein h-1). According to Li et al. (2015), bacterial growth in an ACC-containing medium and the detection of low ACC deaminase activity do not guarantee the presence of ACC deaminase-producing bacteria. Therefore, unambiguous detection of the acdS gene is crucial for predicting enzyme activity and confirming ACC deaminase-producing bacteria. In the present study, the acdS gene was detected in nine bacterial strains. Chandra et al. (2018) employed primers (acdSf3 and acdSr3), as described by Li et al. (2015) to characterize bacterial strains isolated from soil samples collected in India. They successfully amplified a ~680 bp fragment specific to the acdS gene from the genomic DNA of Pseudomonas sp. DPB13, DPB15, and DPB16.
Polyethylene glycol is widely used in assays to simulate water stress in plants by lowering the water potential of the rooting medium, subsequently affecting plant water potential (Lawlor, 1970). In this study, PEG 8000 was utilized to assess the effect of ACC deaminase-producing bacterial inoculation on Brachiaria ruziziensis under in vitro water stress conditions. The results demonstrated that bacterial inoculation significantly enhanced root and shoot development in plants treated with strains NRB142 (P. silvatlantica), NRB223 (A. melinis), and BR11790 (H. frisingense GSF30T). Similarly, Kang et al. (2014) reported the successful colonization of cucumber plants by Burkholderia cepacia SE4, Promicromonospora sp. SE188 and Acinetobacter calcoaceticus SE370. These rhizobacteria conferred protection to plants grown under PEG-mediated stress, promoting increased fresh biomass accumulation in both shoots and roots biomass compared to non-inoculated control plants. These findings agreed with the results observed in the present study with Brachiaria genotypes, reinforcing the potential of ACC deaminase-producing bacteria in enhancing plant tolerance to water deficit stress.
Barnawal et al. (2017) reported that wheat plants inoculated with Bacillus subtilis strain LDR2 produced significantly higher biomass production compared to non-inoculated plants, both subjected to PEG 10000-mediated stress. Similarly, Govindasamy et al. (2020) observed that Sorghum bicolor plants inoculated with plant growth-promoting bacteria displayed a notable increase in shoot and root length under PEG 8000-mediated stress conditions.
An essential factor in mitigating of the water deficit stress is the plant colonization by ACC deaminase-producing bacteria. In this study, we demonstrated that strain NRB142 (P. silvatlantica) successfully colonized Brachiaria plants endophytically, with bacterial counts showing indicating substantial colonization the inoculated Brachiaria roots. This finding corroborated with those of García et al. (2019), who documented the colonization of barley plants (Hordeum vulgare L.) by Paraburkholderia tropica MTo-293 through colony counting and confocal microscopy. Similarly, Ramirez-Mata et al. (2018) employed the mCherry reporter gene to monitor Azospirillum brasilense colonization in wheat plants (Triticum aestivum), while Ferreira et al. (2020) evaluated Rhizobium sp. BR 10268 colonization in sugarcane mini-setts. These studies highlight the effectiveness of molecular and microscopy-based techniques in assessing bacterial colonization. In addition, it further supported the use of such methodologies to confirm the colonization of Brachiaria genotypes by the ACC deaminase-producing strain NRB142 (P. silvatlantica).
5 Conclusion
Plant growth-promoting bacteria with ACC deaminase activity plays an important role in enhancing plant tolerance to water deficit stress, thereby improving biomass production and yield. Our study showed that some bacterial strains exhibited notable ACC deaminase activity in vitro and demonstrated the ability to protect Brachiaria plants under PEG 8000-mediated stress conditions. The results indicated that ACC deaminase-producing strains improved physiological and agronomic parameters of Brachiaria plants, including shoot and root length as well as enhanced biomass accumulation under water stress conditions.
These findings suggest that diazotrophic bacterial strains containing ACC deaminase could serve as effective inoculants to alleviate the negative impacts of water deficit stress on Brachiaria genotypes. However, further validation through greenhouse and field experiments is necessary to confirm the mitigation effects of these bacteria across different Brachiaria genotypes exposed to varying levels of water stress.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author/s.
Author contributions
JF: Data curation, Writing – review & editing, Conceptualization, Formal analysis, Investigation, Methodology, Project administration, Validation, Visualization, Writing – original draft. MV: Supervision, Writing – original draft, Writing – review & editing, Conceptualization, Investigation, Project administration, Visualization. JB: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was funded by the project Embrapa MP2 (n° 12.13.08.004.00.02.003). The first author was supported by a fellowship from the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001. JB was supported by fellowship from CNPq (process 317306/2021) and a bench fee from FAPERJ–CNE (E26/202.550/2019).
Acknowledgments
The first author JF thanks Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES - Code 001) for the fellowship.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ali, S. and Khan, N. (2021). Delineation of mechanistic approaches employed by plant growth-promoting microorganisms for improving drought stress tolerance in plants. Microbiol. Res. 249, 126771. doi: 10.1016/j.micres.2021.126771
Ali, S., Mir, R. A., Haque, M. A., Danishuddin, Almalki, M. A., Alfredan, M., et al. (2025). Exploring physiological and molecular dynamics of drought stress responses in plants: challenges and future directions. Front. Plant Sci. 16. doi: 10.3389/fpls.2025.1565635
Anjum, S., Xie, X., and Wang, L. (2011). Morphological, physiological, and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 6, 2026–2032. doi: 10.5897/AJAR10.027
Armanhi, J. S. L., Souza, R. S. C., Biazotti, B. B., Yassitepe, J. E. D. C. T., and Arruda, P. (2021). Modulating drought stress response of maize by a synthetic bacterial community. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.747541
Baldani, J. I., Reis, V. M., Videira, S. S., Boddey, L. H., and Baldani, V. L. D. (2014). The art of isolating nitrogen-fixing bacteria from non-leguminous plants using N-free semi-solid media: a practical guide for microbiologists. Plant Soil. 384, 413–431. doi: 10.1007/s11104-014-2186-6
Barnawal, D., Bharti, N., Pandey, S. S., Pandey, A., Chanotiya, C. S., and Kalra, A. (2017). Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant 161, 502–514. doi: 10.1111/ppl.12614
Blaha, D., Prigent-Combaret, C., Mirza, M. S., and Moënne-Loccoz, Y. (2006). Phylogeny of the 1-aminocyclopropane-1-carboxylic acid deaminase-encoding gene acdS in phytobeneficial and pathogenic Proteobacteria and relation with strain biogeography. FEMS Microbiol. Ecol. 56, 455–470. doi: 10.1111/j.1574-6941.2006.00082.x
Boyer, J. S. (1982). Plant productivity and environment. Science. 218, 443–448. doi: 10.1126/science.218.4571.443
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1016/0003-2697(76)90527-3
Chandra, D., Srivastava, R., Glick, B. R., and Sharma, A. K. (2018). Drought-tolerant Pseudomonas spp. improve the growth performance of finger millet (Eleusine coracana (L.) Gaertn.) under non-stressed and drought-stressed conditions. Pedosphere 28, 227–240. doi: 10.1016/S1002-0160(18)60013-X
Chandra, D., Srivastava, R., Glick, B. R., and Sharma, A. K. (2020). Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech. 10, 1–15. doi: 10.1007/s13205-019-2046-4
Chandra, D., Srivastava, R., Gupta, V. V. S. R., Franco, C. M. M., Paasricha, N., Saifi, S. K., et al. (2019). Field performance of bacterial inoculants to alleviate water stress effects in wheat (Triticum aestivum L.). Plant Soil. 441, 261–281. doi: 10.1007/s11104-019-04115-9
Chandwani, S. and Amaresan, N. (2024). ACC deaminase producing bacteria alleviate the polyethylene glycol induced drought stress in black gram (Vigna mungo L.) by enhancing nutrient uptake and soil respiration activity. Sci. Hortic. 331, 113111. doi: 10.1016/j.scienta.2024.113111
Cheng, Z., Duncker, B. P., McConkey, B. J., and Glick, B. R. (2008). Transcriptional regulation of ACC deaminase gene expression in Pseudomonas putida UW4. Can. J. Microbiol. 54, 128–136. doi: 10.1139/W07-128
Depaepe, T. and van der Straeten, D. (2016). Ethylene. Encycl. Appl. Plant Sci. 1, 403–410. doi: 10.1016/B978-0-12-394807-6.00103-9
Dimkpa, C., Weinand, T., and Asch, F. (2009). Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 32, 1682–1694. doi: 10.1111/j.1365-3040.2009.02028.x
Duan, J., Jiang, W., Cheng, Z., Heikkila, J. J., and Glick, B. R. (2013). The complete genome sequence of the plant growth-promoting bacterium Pseudomonas sp. UW4. PLoS One 8, e58640. doi: 10.1371/journal.pone.0058640
Duarte, C. F. D., Cecato, U., Hungria, M., Fernandes, H. J., Biserra, T. T., Mamédio, D., et al. (2020). Inoculação de bactérias promotoras do crescimento vegetal em Urochloa ruziziensis. Res. Soc Dev. 9, e630985978. doi: 10.33448/rsd-v9i8.5978
Dubey, A., Malla, M. A., Kumar, A., Khan, M. L., and Kumari, S. (2024). Seed bio-priming with ACC deaminase-producing bacterial strains alleviates impact of drought stress in soybean (Glycine max (L.) Merr.). Rhizosphere 30, 100873. doi: 10.1016/j.rhisph.2024.100873
Duraivadivel, P., Gowtham, H. G., and Hariprasad, P. (2020). Co-occurrence of functionally diverse bacterial community as biofilm on the root surface of Eichhornia crassipes (Mart.) Solms-Laub. Sci. Total Environ. 714, 136683. doi: 10.1016/j.scitotenv.2020.136683
Ferreira, D. F. (2011). Sisvar: a computer statistical analysis system. Ciênc. Agrotec. 35, 1039–1042. doi: 10.1590/s1413-70542011000600001
Ferreira, N. S., Matos, G. F., Meneses, C. H. S. G., Reis, V. M., Rouws, J. R. C., Schwab, S., et al. (2020). Interaction of phytohormone-producing rhizobia with sugarcane mini-setts and their effect on plant development. Plant Soil. 451, 221–238. doi: 10.1007/s11104-019-04388-0
García, S. S., Bernabeu, P. R., Vio, S. A., Cattelan, N., García, J. E., Puente, M. L., et al. (2019). Paraburkholderia tropica as a plant-growth–promoting bacterium in barley: characterization of tissues colonization by culture-dependent and -independent techniques for use as an agronomic bioinput. Plant Soil. 451, 89–106. doi: 10.1007/s11104-019-04174-y
Glick, B. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012, 1–15. doi: 10.6064/2012/963401
Glick, B. R. (2015). “Introduction to plant growth-promoting bacteria,” in Beneficial Plant-Bacterial Interactions. Ed. Glick, B. R. (Springer Cham, Switzerland), 1–28.
Glick, B. R., Cheng, Z., Czarny, J., and Duan, J. (2007b). “Promotion of plant growth by ACC deaminase-producing soil bacteria,” in New Perspect. Approaches Plant Growth-Promot. Rhizobacteria Res. Eds. Bakker, P. A. H. M., Raaijmakers, J. M., Bloemberg, G., Höfte, M., Lemanceau, P., and Cooke, B. M. (Springer, Dordrecht). doi: 10.1007/978-1-4020-6776-1_8
Glick, B. R., Karaturovic, D. M., and Newell, P. C. (1995). A novel procedure for rapid isolation of plant growth promoting pseudomonads. Can. J. Microbiol. 41, 533–536. doi: 10.1139/m95-070
Glick, B. R. and Nascimento, F. X. (2021). Pseudomonas 1-aminocyclopropane-1-carboxylate (ACC) deaminase and its role in beneficial plant-microbe interactions. Microorganisms 9, 2467. doi: 10.3390/microorganisms9122467
Glick, B. R., Penrose, D. M., and Li, J. (1998). A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 190, 63–68. doi: 10.1006/jtbi.1997.0532
Glick, B. R., Todorovic, B., Czarny, J., Cheng, Z., Duan, J., and McConkey, B. (2007a). Promotion of plant growth by bacterial ACC deaminase. Crit. Rev. Plant Sci. 26, 227–242. doi: 10.1080/07352680701572966
Govindasamy, V., George, P., Kumar, M., Aher, L., Raina, S. K., Rane, J., et al. (2020). Multi-trait PGP rhizobacterial endophytes alleviate drought stress in a senescent genotype of sorghum (Sorghum bicolor L. Moench). 3 Biotech. 10, 1–14. doi: 10.1007/s13205-019-2001-4
Grichko, V. P. and Glick, B. R. (2000). Identification of DNA sequences that regulate the expression of the Enterobacter cloacae UW4 1-aminocyclopropane-1-carboxylic acid deaminase gene. Can. J. Microbiol. 46, 1159–1165. doi: 10.1139/cjm-46-12-1159
Grover, M., Ali, S. Z., Sandhya, V., Rasul, A., and Venkateswarlu, B. (2011). Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 27, 1231–1240. doi: 10.1007/s11274-010-0572-7
Gupta, S. and Pandey, S. (2019). ACC deaminase producing bacteria with multifarious plant growth promoting traits alleviates salinity stress in French Bean (Phaseolus vulgaris) plants. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.01506
Islam, M. R., Madhaiyan, M., Deka Boruah, H. P., Yim, W., Lee, G., Saravanan, V. S., et al. (2009). Characterization of plant growth-promoting traits of free-living diazotrophic bacteria and their inoculation effects on growth and nitrogen uptake of crop plants. J. Microbiol. Biotechnol. 19, 1213–1222. doi: 10.4014/jmb.0903.03028
Jain, R. and Saraf, M. (2023). ACC deaminase producing PGPR modulates nutrients uptake, soil properties and growth of cluster bean (Cyamopsis tetragonoloba L.) under deficit irrigation. Biologia 78, 2303–2316. doi: 10.1007/s11756-023-01376-9
Jalili, F., Khavazi, K., Pazira, E., Nejati, A., Rahmani, H. A., Sadaghiani, H. R., et al. (2009). Isolation and characterization of ACC deaminase-producing fluorescent pseudomonads, to alleviate salinity stress on canola (Brassica napus L.) growth. J. Plant Physiol. 166, 667–674. doi: 10.1016/j.jplph.2008.08.004
Jank, L. B., Sanzio, C., Valle, C. B., Simeão, R. M., and Alves, G. F. (2014). The value of improved pastures to Brazilian beef production. Crop Pasture Sci. 65, 1132–1137. doi: 10.1071/CP13319
Jasso-Arreola, Y., Ibarra, J. A., Rosas-Cárdenas, F. D. F., and and Estrada-de Los Santos, P. (2025). Beneficial effects of ACC deaminase-producing rhizobacteria on the drought stress resistance of coffea arabica L. Plants 14, 1084. doi: 10.3390/plants14071084
Kang, S. M., Khan, A. L., Waqas, M., You, Y. H., Kim, J. H., Kim, J. G., et al. (2014). Plant growth-promoting rhizobacteria reduce adverse effects of salinity and osmotic stress by regulating phytohormones and antioxidants in Cucumis sativus. J. Plant Interact. 9, 673–682. doi: 10.1080/17429145.2014.894587
Kaushal, M. and Wani, S. P. (2016). Plant-growth-promoting rhizobacteria: drought stress alleviators to ameliorate crop production in drylands. Ann. Microbiol. 66, 35–42. doi: 10.1007/s13213-015-1112-3
Lawlor, D. W. (1970). Absorption of polyethylene glycols by plants and their effects on plant growth. New Phytol. 69, 501–513. doi: 10.1111/j.1469-8137.1970.tb02446.x
Li, Z., Chang, S., Lin, L., Li, Y., and An, Q. (2011). A colorimetric assay of 1-aminocyclopropane-1-carboxylate (ACC) based on ninhydrin reaction for rapid screening of bacteria containing ACC deaminase. Lett. Appl. Microbiol. 53, 178–185. doi: 10.1111/j.1472-765X.2011.03088.x
Li, Z., Chang, S., Ye, S., Chen, M., Lin, L., Li, Y., et al. (2015). Differentiation of 1-aminocyclopropane-1-carboxylate (ACC) deaminase from its homologs is the key for identifying bacteria containing ACC deaminase. FEMS Microbiol. Ecol. 91, fiv112. doi: 10.1093/femsec/fiv112
Li, J. and Glick, B. R. (2001). Transcriptional regulation of the Enterobacter cloacae UW4 1-aminocyclopropane-1-carboxylate (ACC) deaminase gene (acdS). Can. J. Microbiol. 47, 359–367. doi: 10.1139/w01-009
Li, Y. P., Ye, W., Wang, M., and Yan, X. D. (2009). Climate change and drought: A risk assessment of crop-yield impacts. Clim. Res. 39, 31–46. doi: 10.3354/cr00797
Lisar, S. Y. S., Motafakkerazad, R., Hossain, M. M., and Rahman, I. M. M. (2012). “Water stress in plants: causes, effects, and responses,” in Water stress. Eds. Rahman, I. M. M. and Hasegawa, H. (InTech, Croatia), 1–14.
Moore, K. J., Lenssen, A. W., and Fales, S. L. (2020). “Factors affecting forage quality,” in Forages. Eds. Moore, K. J., Collins, M., Nelson, C. J., and Redfearn, D. D. (Wiley, Hoboken, NJ), ch. 39. doi: 10.1002/9781119436669.ch39
Muñoz-Carvajal, E., González, M., Fuentes, Y., Oetiker, N., Giordano, A., and Stoll, A. (2024). Rhizobacteria Enterobacter sp. LHB11 and Bacillus sp. PIXIE Induced Systemic Tolerance Against Drought Stress in Tomato (Solanum lycopersicum). Agronomy 14, 3013. doi: 10.3390/agronomy14123013
Murashige, T. and Skoog, F. (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant 15, 473–497. doi: 10.1111/j.1399-3054.1962.tb08052.x
Niu, X., Song, L., Xiao, Y., and Ge, W. (2018). Drought-tolerant plant growth-promoting rhizobacteria associated with foxtail millet in a semi-arid and their potential in alleviating drought stress. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02580
Ojuederie, O. B. and Babalola, O. O. (2023). Growth enhancement and extenuation of drought stress in maize inoculated with multifaceted ACC deaminase producing rhizobacteria. Front. Sustain. Food Syst. 6, 1076844. doi: 10.3389/fsufs.2022.1076844
Onofre-Lemus, J., Hernández-Lucas, I., Girard, L., and Caballero-Mellado, J. (2009). ACC (1-aminocyclopropane-1-carboxylate) deaminase activity, a widespread trait in Burkholderia species, and its growth-promoting effect on tomato plants. Appl. Environ. Microbiol. 75, 6581–6590. doi: 10.1128/AEM.01240-09
Pei, H., Wang, H., Wang, L., Zheng, F., and Dong, C.-H. (2017). “Regulatory function of ethylene in plant responses to drought, cold, and salt stresses,” in Mechanism of Plant Hormone Signaling Under Stress. Ed. Pandey, G. K. (John Wiley & Sons, USA), 327–344. doi: 10.1002/9781118889022.ch13
Penrose, D. M. and Glick, B. R. (2003). Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant 118, 10–15. doi: 10.1034/j.1399-3054.2003.00086.x
Poudel, M., Mendes, R., Costa, L. A. S., Bueno, C. G., Meng, Y., Folimonova, S. Y., et al. (2021). The role of plant-associated bacteria, fungi, and viruses in drought stress mitigation. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.743512
Ramirez-Mata, A., Pacheco, M. R., Moreno, S. J., Xiqui-Vazquez, M. L., and Baca, B. E. (2018). Versatile use of Azospirillum brasilense strains tagged with egfp and mCherry genes for the visualization of biofilms associated with wheat roots. Microbiol. Res. 215, 155–163. doi: 10.1016/j.micres.2018.07.007
Ribeiro, N. V. D. S., Vidal, M. S., Barrios, S. C. L., Baldani, V. L. D., and Baldani, J. I. (2020). Genetic diversity and growth promoting characteristics of diazotrophic bacteria isolated from 20 genotypes of Brachiaria spp. Plant Soil 451, 187–205. doi: 10.1007/s11104-019-04263-y
Rodrigues Neto, J., Malavolta, V. A., Jr., and Victor, O. (1986). Meio simples para o isolamento e cultivo de Xanthomonas campestris pv. citri tipo B. Summa Phytopathol. 12, 16.
Romeiro, R. S. (2007). Técnica de microgota para contagem de células bacterianas viáveis em uma suspensão. Laboratório de Bacteriologia de Plantas (Viçosa: UFV).
Rothballer, M., Eckert, B., Schmid, M., Fekete, A., Schloter, M., Lehner, A., et al. (2008). Endophytic root colonization of gramineous plants by Herbaspirillum frisingense. FEMS Microbiol. Ecol. 66, 85–95. doi: 10.1111/j.1574-6941.2008.00582.x
Sati, D., Pande, V., Pandey, S. C., and Samant, M. (2023). Recent advances in PGPR and molecular mechanisms involved in drought stress resistance. J. Soil Sci. Plant Nutr. 23, 106–124. doi: 10.1007/s42729-021-00724-5
Seleiman, M. F., Al-Suhaibani, N., Ali, N., Akmal, M., Alotaibi, M., Refay, Y., et al. (2021). Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 10, 259. doi: 10.3390/plants10020259
Shahid, M., Singh, U. B., Khan, M. S., Singh, P., Kumar, R., Singh, R. N., et al. (2023). Bacterial ACC deaminase: Insights into enzymology, biochemistry, genetics, and potential role in amelioration of environmental stress in crop plants. Front. Microbiol. 14, 1132770. doi: 10.3389/fmicb.2023.1132770
Shao, H. B., Chu, L. Y., Jaleel, C. A., and Zhao, C. X. (2008). Water-deficit stress-induced anatomical changes in higher plants. Comptes Rendus Biologies 331, 215–225. doi: 10.1016/j.crvi.2008.01.002
Singh, R. P., Shelke, G. M., Kumar, A., and Jha, P. N. (2015). Biochemistry and genetics of ACC deaminase: A weapon to ‘stress ethylene’ produced in plants. Front. Microbiol. 6. doi: 10.3389/fmicb.2015.00937
Soni, R., Yadav, S. K., and Rajput, A. S. (2018). “ACC-deaminase producing rhizobacteria: prospects and application as stress busters for stressed agriculture,” in Microorganisms for Green Revolution. Microorganisms for Sustainability, vol. 7 . Eds. Panpatte, D., Jhala, Y., Shelat, H., and Vyas, R. (Springer, Singapore), 161–175.
Sun, L. N., Zhang, Y. F., He, L. Y., Chen, Z. J., Wang, Q. Y., Qian, M., et al. (2010). Genetic diversity and characterization of heavy metal-resistant-endophytic bacteria from two copper-tolerant plant species on copper mine wasteland. Bioresour. Technol. 101, 501–509. doi: 10.1016/j.biortech.2009.08.011
Tahir, M., Khalid, U., Khan, M. B., Shahid, M., Ahmad, I., Akram, M., et al. (2019). Auxin and 1-aminocyclopropane-1-carboxylate deaminase activity exhibiting rhizobacteria improved maize quality and productivity under drought conditions. Int. J. Agric. Biol. 21, 943–954. doi: 10.17957/IJAB/15.0979
Taiz, L. and Zeiger, E. (2017). Fisiologia e Desenvolvimento Vegetal. 6th edn (Porto Alegre: Artmed), 888 p.
Timmusk, S., Paalme, V., Pavlicek, T., Bergquist, J., Vangala, A., Danilas, T., et al. (2011). Bacterial distribution in the rhizosphere of wild barley under contrasting microclimates. PLoS One 6, 1–7. doi: 10.1371/journal.pone.0017968
Tiwari, G., Duraivadivel, P., Sharma, S., and Hariprasad, P. (2018). 1-aminocyclopropane-1-carboxylic acid deaminase producing beneficial rhizobacteria ameliorate the biomass characters of Panicum maximum Jacq. by mitigating drought and salt stress. Sci. Rep. 8, 1–12. doi: 10.1038/s41598-018-35565-3
USDA (2025). Livestock and poultry: world markets and trade. Available online at: https://apps.fas.usda.gov/psdonline/circulars/livestock_poultry.pdf (Accessed 2 June, 2025).
Vurukonda, S. S. K. P., Vardharajula, S., Shrivastava, M., and SkZ, A. (2016). Enhancement of drought stress tolerance in crops by plant growth-promoting rhizobacteria. Microbiol. Res. 184, 13–24. doi: 10.1016/j.micres.2015.12.003
Yavuz, D., Baştaş, K. K., Seymen, M., Yavuz, N., Kurtar, E. S., Süheri, S., et al. (2023). Role of ACC deaminase-producing rhizobacteria in alleviation of water stress in watermelon. Sci. Hortic. 321, 112288. doi: 10.1016/j.scienta.2023.112288
Yordanov, I., Velikova, V., and Tsonev, T. (2000). Plant responses to drought, acclimation, and stress tolerance. Photosynthetica. 38, 171–186. doi: 10.1023/A:1007201411474
Zhang, M., Yang, L., Hao, R., Bai, X., Wang, Y., and Yu, X. (2020). Drought-tolerant plant growth-promoting rhizobacteria isolated from jujube (Ziziphus jujuba) and their potential to enhance drought tolerance. Plant Soil. 452, 423–440. doi: 10.1007/s11104-020-04582-5
Keywords: plant growth promoting bacteria, pasture, 1-Aminocyclopropane-1-carboxylic acid, ethylene stress, inoculation, plant colonization, mCherry reporter gene, polyethylene glycol
Citation: Ferreira JP, Vidal MS and Baldani JI (2025) Exploring ACC deaminase-producing bacteria for drought stress mitigation in Brachiaria. Front. Plant Sci. 16:1607697. doi: 10.3389/fpls.2025.1607697
Received: 08 April 2025; Accepted: 18 July 2025;
Published: 19 August 2025.
Edited by:
Marie-Joelle Françoise Virolle, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
German Andres Estrada-Bonilla, Corporación Colombiana de Investigación Agropecuaria (Agrosavia)-Tibaitatá, ColombiaMohammad Shahid, Aligarh Muslim University, India
Copyright © 2025 Ferreira, Vidal and Baldani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José I. Baldani, aXZvLmJhbGRhbmlAZW1icmFwYS5icg==