 Beáta Piršelová
Beáta Piršelová Jana Jakubčinová
Jana Jakubčinová- Department of Botany and Genetics, Faculty of Natural Sciences and Informatics, Constantine the Philosopher University in Nitra, Nitra, Slovakia
Cyanogenic glycosides (CGs) represent an important group of secondary metabolites predominantly of plant origin, characterized by their ability to release hydrogen cyanide upon enzymatic hydrolysis. These compounds are widely distributed across the plant kingdom, where they play a crucial role in defense against herbivores and pathogens. In recent years, advanced analytical tools have greatly expanded our knowledge of CGs by enabling the identification of less abundant forms. Based on the latest data from published scientific studies, this review presents a comprehensive overview of CGs, with a focus on their structural variability, biosynthetic pathways, ecological functions, and inherent toxicity. Special attention is given to the quantity and distribution of significant CGs in plants, as the available data is often heterogeneous, fragmented, and dispersed across the literature. Furthermore, the review explores emerging evidence regarding the biomedical relevance of selected CGs, including their putative anticancer properties and broader therapeutic potential. The findings presented in this review may be applied in fields such as pharmacology, toxicology, food safety, and plant biotechnology - either to enhance CG content for crop protection or, conversely, to eliminate such content in order to improve food safety.
1 Introduction
Cyanogenic glycosides (cyanoglycosides, CGs) are secondary metabolites of predominantly plant origin and account for nearly 90% of the broader group of plant toxins known as cyanogens (Nampoothiri, 2017). Chemically, CGs are α-hydroxynitrile glucoside consisting of two main components: a sugar moiety - most commonly glucose - and an aglycone, the non-sugar part of the molecule that contains the cyanogenic group (CN). These components are linked through a glycosidic bond. Glycosylation plays a crucial role in determining the stability, solubility, and biological activity of CGs, including their potential antitumor properties (Mosayyebi et al., 2020). It also influences the interaction between the aglycone and cellular structures, such as receptors and proteins, thereby affecting a compound’s biological function (Pelley, 2012). The aglycone can vary in its chemical structure, most commonly appearing as aliphatic, cyclic, aromatic, or heterocyclic compounds. This part of the molecule largely determines the toxicity of CGs. Natural cyanogenic glycosides display considerable structural diversity in both their sugar and aglycone components (Vetter, 2017). Some naturally occurring CGs exist as stereoisomers, for example: (R)-lotaustralin/(S)-epilotaustralin, (R)- prunasin/(S)- sambunigrin, and (2R)-taxyphyllin/(2S)-dhurrin (Yulvianti and Zidorn, 2021). The general structure of CGs is illustrated in Figure 1, with the structures of the most significant compounds shown in Figure 2. The chemical diversity of plant CGs are described in more detail in article Yulvianti and Zidorn (2021).
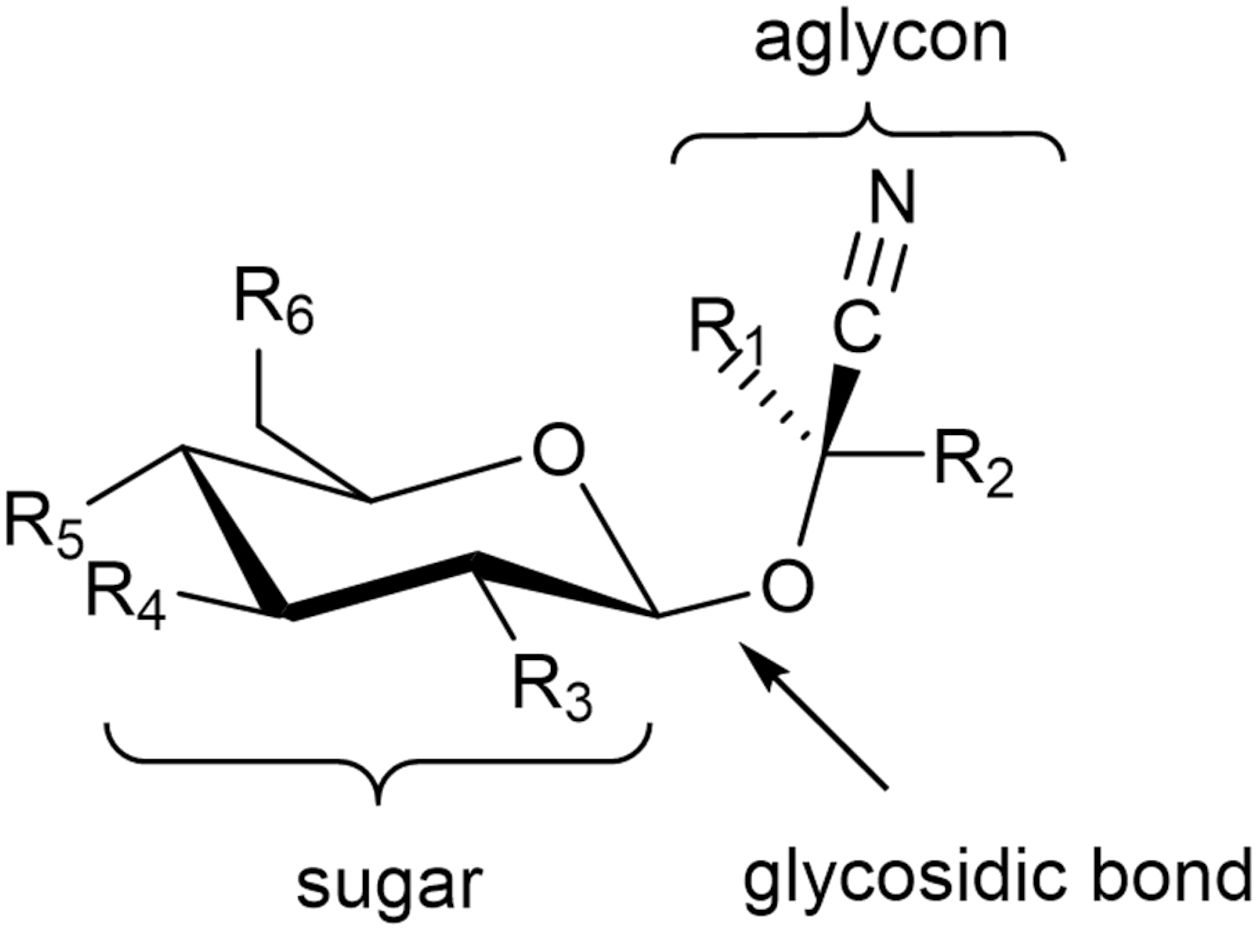
Figure 1. General structure of cyanogenic glycosides. R1 represents a proton for amygdalin, prunasin, and dhurrin and a methyl group for linamarin, while R2 is a variable organic group. R3–R6 represent variable inorganic (most commonly the hydroxyl group) or organic groups.
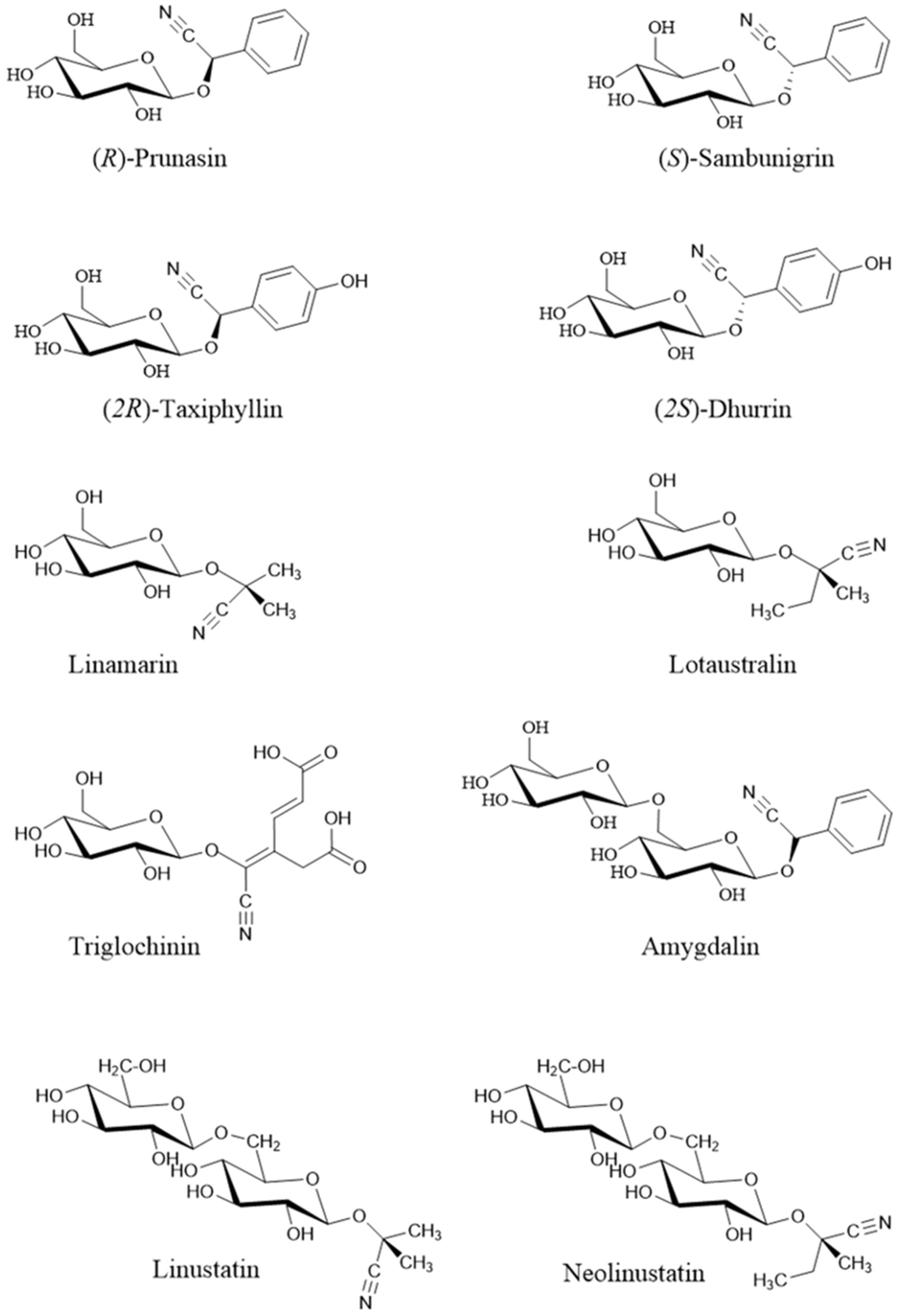
Figure 2. Structure of the most important cyanogenic glycosides (Rietjens and Eisenbrand, 2022). Stereoisomers: prunasin (R) / sambunigrin (S) and (2R)-taxyphyllin / (2S)-dhurrin.
Trivial names of CGs are usually derived from the Latin names of the plants from which they were first isolated (e.g. almond amygdalin - Prunus amygdalus). However, several isolated CGs do not have trivial names.
Currently, 112 distinct CGs are known from the plant kingdom (Yulvianti and Zidorn, 2021). For plants they are important as protection against being consumed by animals and also as protection against various microorganisms (Zagrobelny et al., 2018). But actually, this protection is not provided by CG itself, but rather by the toxic hydrogen cyanide (HCN) released from stored CGs, cyanolipids, or cyanohydrins (Lechtenberg, 2011). This process occurs in an acidic environment (at low pH) or under the influence of hydrolytic enzymes with the formation of free HCN after the mechanical disruption of tissues. Cyanogenesis occurs in two phases: Phase 1 - cleavage of the carbohydrate component, Phase 2 - cleavage of the aglycone to aldehyde or ketone and HCN (Figure 3).
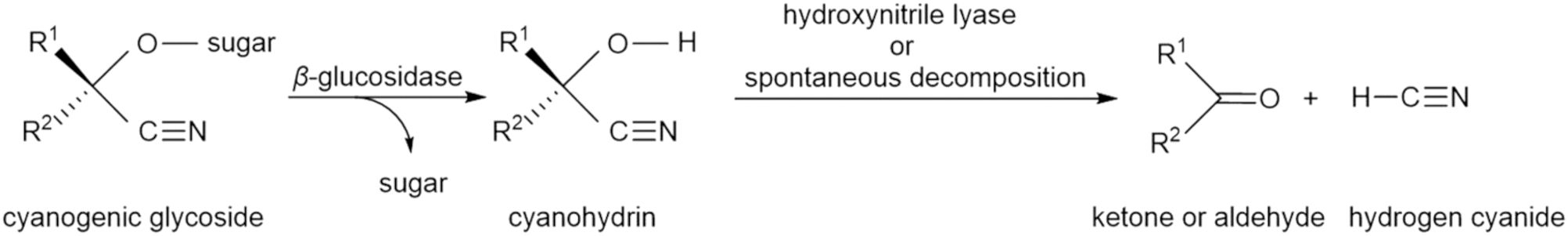
Figure 3. Glycoside cleavage – cyanogenesis.
While CGs are stored in vacuoles, β-glucosidases are localized in the apoplastic space, bound to cell walls in dicotyledonous plants, and in the cytoplasm and chloroplasts in monocotyledonous plants. Hydroxynitrile enzymes accumulate mainly in the cytoplasm and plasma membranes. When plant tissue is disrupted, CGs and enzymes come into contact, and the CGs degrade into cyanohydrins, HCN, and ketones. The different compartmentalization of CGs and enzymes helps prevent excessive HCN production and its toxicity in plants (Vetter, 2017). Yet the cause of the typical bitter odor in the mechanical disruption of seeds containing CGs is not HCN, but the released benzaldehyde (Griffin, 1974; Moertel et al., 1982). CGs are also a re-mobilizable reservoir of reduced nitrogen, and increase plant tolerance by reducing oxidative stress and may support seedling development (Sanchez-Perez et al., 2009; Pičmanová et al., 2015). Moreover, free cyanide, including that released from the CGs, may act as a signaling molecule (Siegień and Bogatek, 2006).
2 Biosynthesis of plant cyanogenic glycosides
Cyanogenic glycosides (CGs) are primarily derived from aliphatic amino acids (L-valine, L-isoleucine, L-leucine) and aromatic amino acids (L-phenylalanine, L-tyrosine). However, certain CGs - such as deidaclin, gynocardin, acalyphin, cycasin, and ranunculin - are synthesized from non-proteinogenic precursors (Nyirenda, 2020). While cyanogenic ferns and gymnosperm species predominantly produce aromatic CGs, angiosperms are known to synthesize both aliphatic and aromatic forms (Bak et al., 2006). To date, amino acid-derived cyanogenic glucoside pathways have been elucidated in various plant species. Despite species-specific variations, three conserved enzymatic steps have been identified across all CG biosynthetic pathways (Figure 4): 1. Amino acid hydroxylation – the conversion of α-amino acids to aldoximes via N-hydroxylated derivatives, mediated by membrane-bound enzymes from the cytochrome P450 (CYP) family. In gymnosperms and angiosperms, this is functionally conserved as the enzyme CYP79. 2. Cyanohydrin formation – the transformation of aldoximes into unstable cyanohydrins via further P450 cytochrome enzymes. In angiosperms, several more or less specific CYPs involved in this pathway have been characterized (CYP71, CYP706, CYP736). 3. Glycosylation - the attachment of a glucose unit, which stabilizes the cyanohydrins into cyanogenic glucosides. This step is catalyzed by the enzyme UDP-glucosyltransferase (in angiosperms, UGT85 and UGT94 have been characterized).
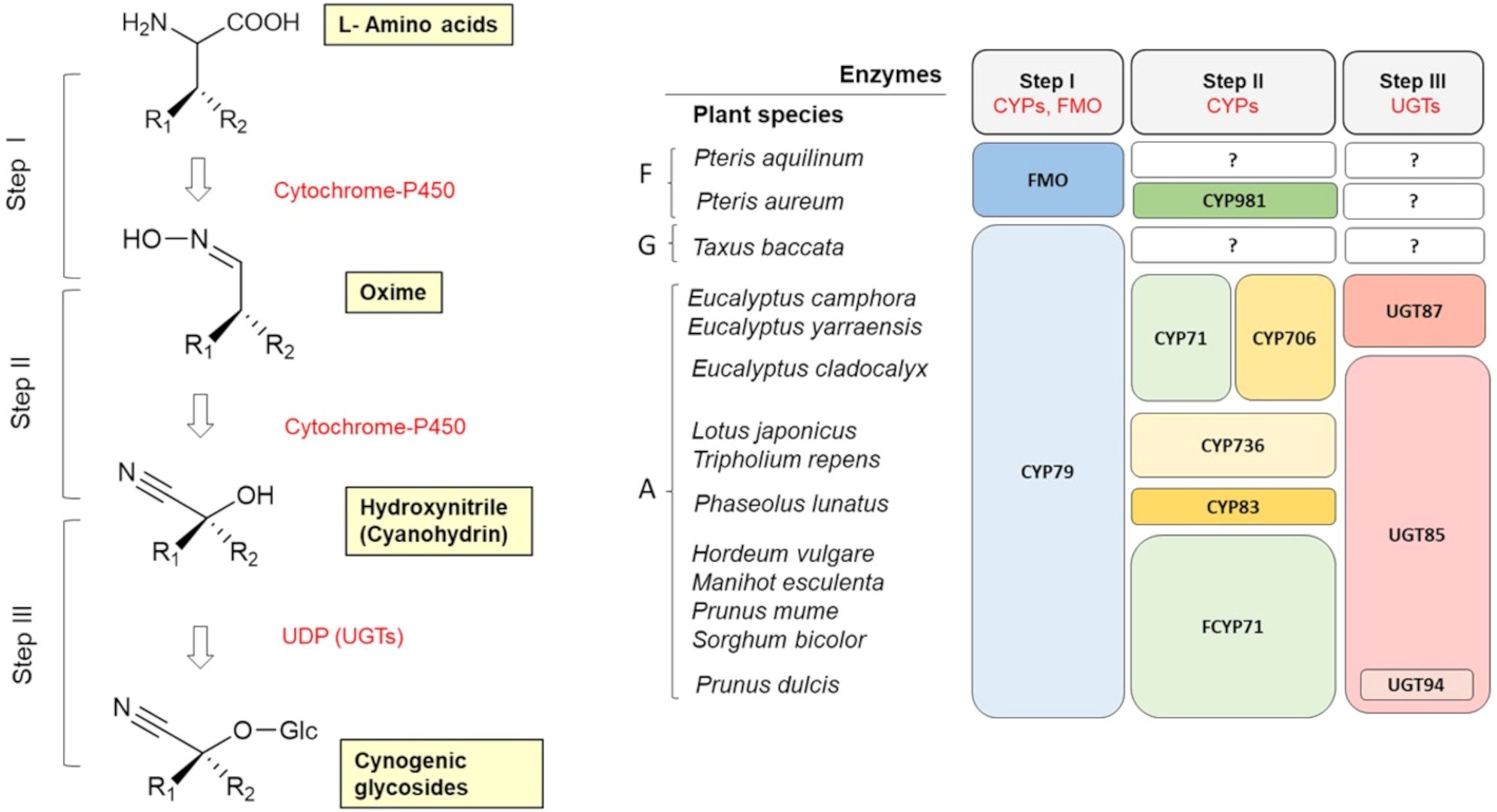
Figure 4. General scheme of the biosynthesis of cyanogenic glycosides in plants (adapted from Ganjewala et al., 2010). Evolution of key enzymes in cyanogenic glycoside biosynthesis in ferns (F), gymnosperms (G) and angiosperms (A) (adapted from Sánchez-Pérez and Neilson, 2024). CYP, cytochrome P450; FMO, flavin-containing monooxygenase; UGT, UDP-glucosyltransferase; ‘?’ denotes an unknown step.
Transcription factors of the basic helix-loop-helix (bHLH) type play a key role in the regulation of CGs biosynthesis (Harun and Mohamed-Hussein, 2024). The plasticity of CYP gene expression, combined with their catalytic versatility, has made them key drivers of evolutionary innovation in plant secondary metabolism, allowing plants to colonize new environments and co-evolve with herbivores and pathogens (Bak et al., 2006; Xu et al., 2015).
In recent decades, significant progress has been made in the study of CG biosynthetic pathways and their regulation, which has enabled a deeper understanding of plant adaptation mechanisms and their evolutionary processes. This topic has been explored in more detail in studies by Forslund et al. (2004); Bak et al. (2006); Morant et al. (2007); Sun et al. (2018); Thodberg et al. (2020); Yulvianti and Zidorn (2021); Boter and Diaz (2023); Harun and Mohamed-Hussein (2024).
3 Genetic and ecological aspects of cyanogenesis
Cyanogenesis was first described in white clover (Trifolium repens) (Mirande, 1912), and it soon became evident that this species is polymorphic in terms of cyanogenesis – that is, both cyanogenic and acyanogenic plants occur within the same population (Armstrong et al., 1913). It was shown that this form of ecological adaptation results from polymorphism (the presence or absence) of genes responsible for both the synthesis of CGs (Ac) and the synthesis of β-glucosidases, enzymes that break down CGs (Li) (Hughes, 1991). Plants that carry at least one dominant (functional) allele at both genes (Ac and Li) are cyanogenic, while the occurrence of two nonfunctional alleles (ac and li) at either gene confers the acyanogenic phenotype. The Ac gene corresponds to the gene encoding cytochrome P450 from the CYP79D protein subgroup (specifically CYP79D15). CYP79D orthologs catalyze the first step in the biosynthesis of cyanogenic glycosides (Figure 4) (Olsen et al., 2008, 2013).
This chemical defense polymorphism is among the most long-studied and best-documented examples of adaptive polymorphism in plants. More cyanogenic plants are found in warmer and more humid regions with higher herbivore activity. However, since cyanogenesis is quite energetically costly, cyanogenic plants exhibit slower growth and reproduction in these areas. This represents a classic example of an evolutionary trade-off between defense and growth. It should, however, be noted that not all cyanogenic plants exhibit adaptive polymorphism. In many species, cyanogenesis is genetically fixed - either all individuals are cyanogenic, or none are. Adaptive polymorphism, as thoroughly documented in Trifolium repens, represents a specific evolutionary phenomenon that occurs in only certain species where selective pressures maintain both the presence and absence of cyanogenic expression within the same population (Olsen et al., 2008).
4 Distribution and content of cyanogenic glycosides in plants
CG synthesis is relatively widespread in the plant kingdom. More than 3000 plant species belonging to 130 families are cyanogenic (Yadav et al., 2023), including ferns, gymnosperms and angiosperms. In the agricultural context, the main sources of CGs are seeds and by-products of crops such as flax (Linus usitatissimum), apricot (Prunus armeniaca), bitter almond (Prunus dulcis), sorghum (Sorghum vulgare), wheat (Triticum aestivum), barley (Hordeum vulgare), oat (Avena sativa), cassava (Manihot esculenta) and apple (Malus pumila) (Hegnauer, 1986; Jones, 1998). In general, CGs exhibit a highly specialized distribution, with a given type of CG typically occurring in only one or two plant families. Furthermore, individual plant species generally produce only one or two types of CGs, reflecting their metabolic specialization and ecological adaptations (Bolarinwa et al., 2014a; Süli et al., 2017). Amygdalin and prunasin, for example, are predominantly found in plants of the Rosaceae family (e.g. Prunus spp., Malus spp.), where it functions as a chemical defense against herbivores. Linamarin and lotaustralin are characteristic of tropical and subtropical plants from the Fabaceae and Euphorbiaceae families (e.g., Phaseolus lunatus, Manihot esculenta), primarily serving to protect these plants from insect herbivores and microbial pathogens. A representative cyanogenic glycoside of the Poaceae family is dhurrin, which is especially abundant in young leaves of Sorghum bicolor, where it enhances the plant’s resistance to herbivores during early developmental stages.
However, the defensive potential of CGs is also manifested in the process of plant adaptation to various abiotic stressors, such as drought, excessive moisture, mineral imbalance, frost, trampling, and herbicide exposure (Bolarinwa et al., 2014a). Moreover, the degree of HCN induction appears to differ depending on whether the stress is chronic or acute (Wheeler et al., 1990; Woodrow et al., 2002). In stressed plants, where photosynthetic rate is reduced, CGs may also provide a ready source of nitrogen, remobilized when the stress is alleviated (O’Donnell et al., 2013; Schmidt et al., 2018). Under stress conditions, they also reduce oxidative stress and regulate the transport of carbon and nitrogen in plants (Conn, 1980; Rosati et al., 2019). Younger plants contain CGs much more than older ones (Dreyer et al., 1981). Some plants are not completely cyanogenic, others are not cyanogenic throughout the growing season. Cereal leaves are cyanogenic for example, but the grains are not. Papaya and mango leaves are also cyanogenic, but the fruits are not. Drought, frost, and the use of nitrates and herbicides can increase their amount and thus their toxicity to animals (Busk and Møller, 2002). Seasonal changes in the cyanide content of some species have also been reported (Robakowski et al., 2016; Bartnik and Facey, 2017). The amount of the most significant CGs in plants (expressed as the equivalent amount of HCN) is given in Table 1. The absolute amounts of individual CGs are listed in Tables 2–9. However, the reported CG levels in plant tissues also depend on the method of extraction and determination, as well as on the genotype, plant age, soil condition, fertilizer application, climatic conditions, and other factors (Bolarinwa et al., 2014a; Tahir et al., 2024).
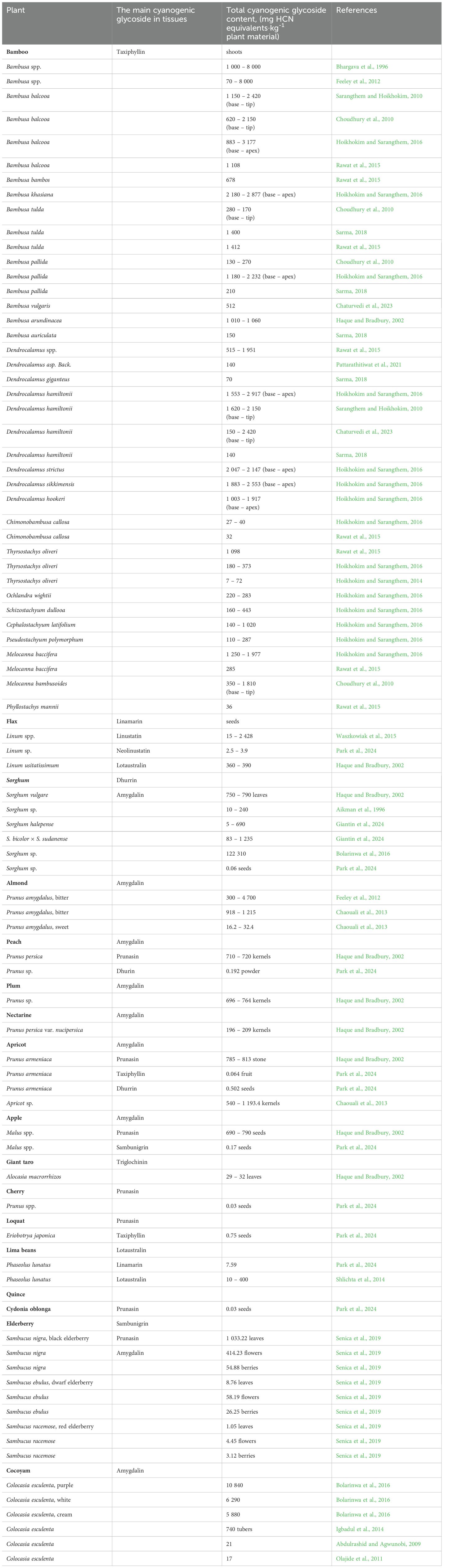
Table 1. Amount of cyanogenic glycosides in plants (mg HCN equivalents·kg-1 plant material).
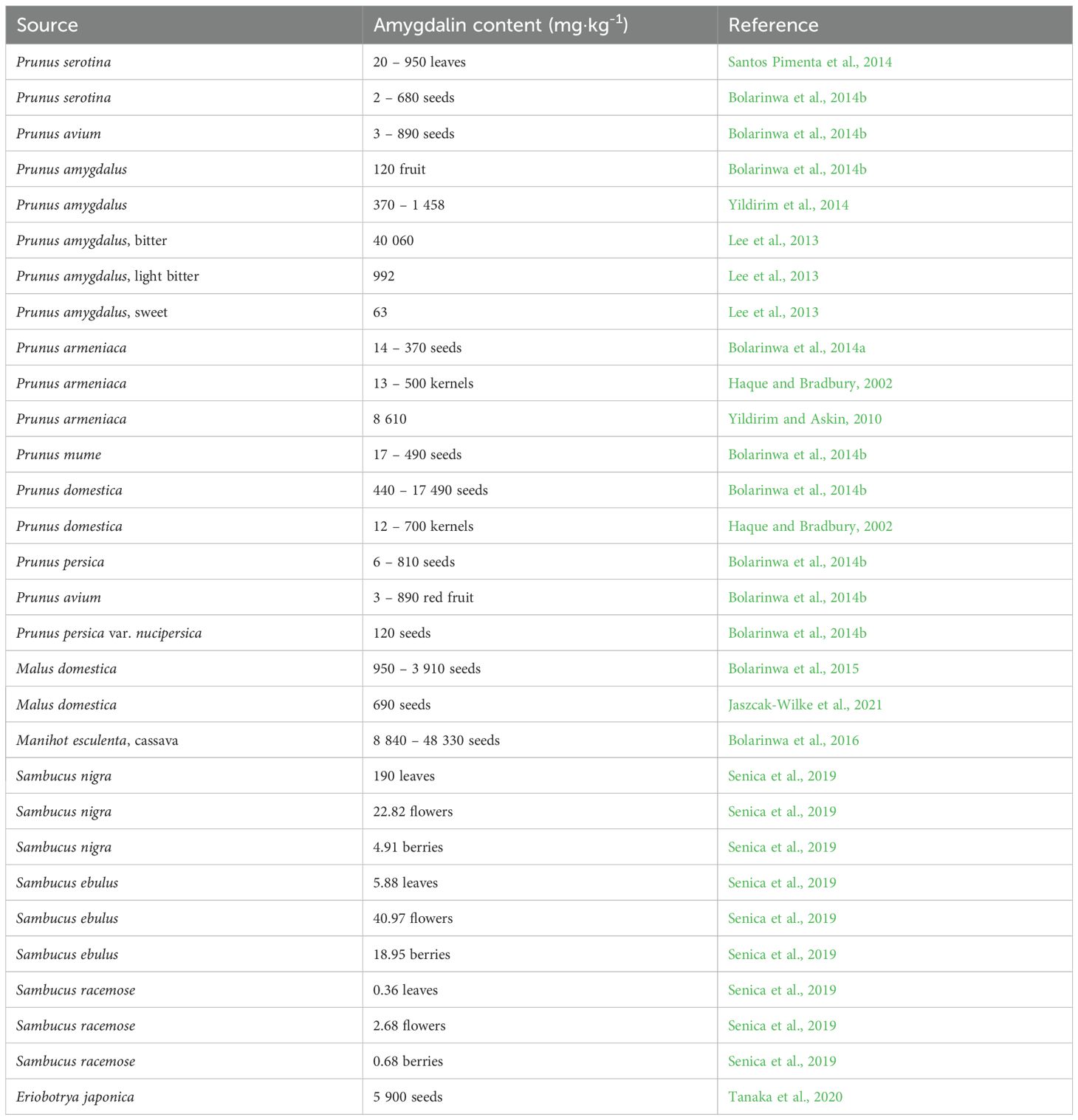
Table 2. Amygdalin content in plants.
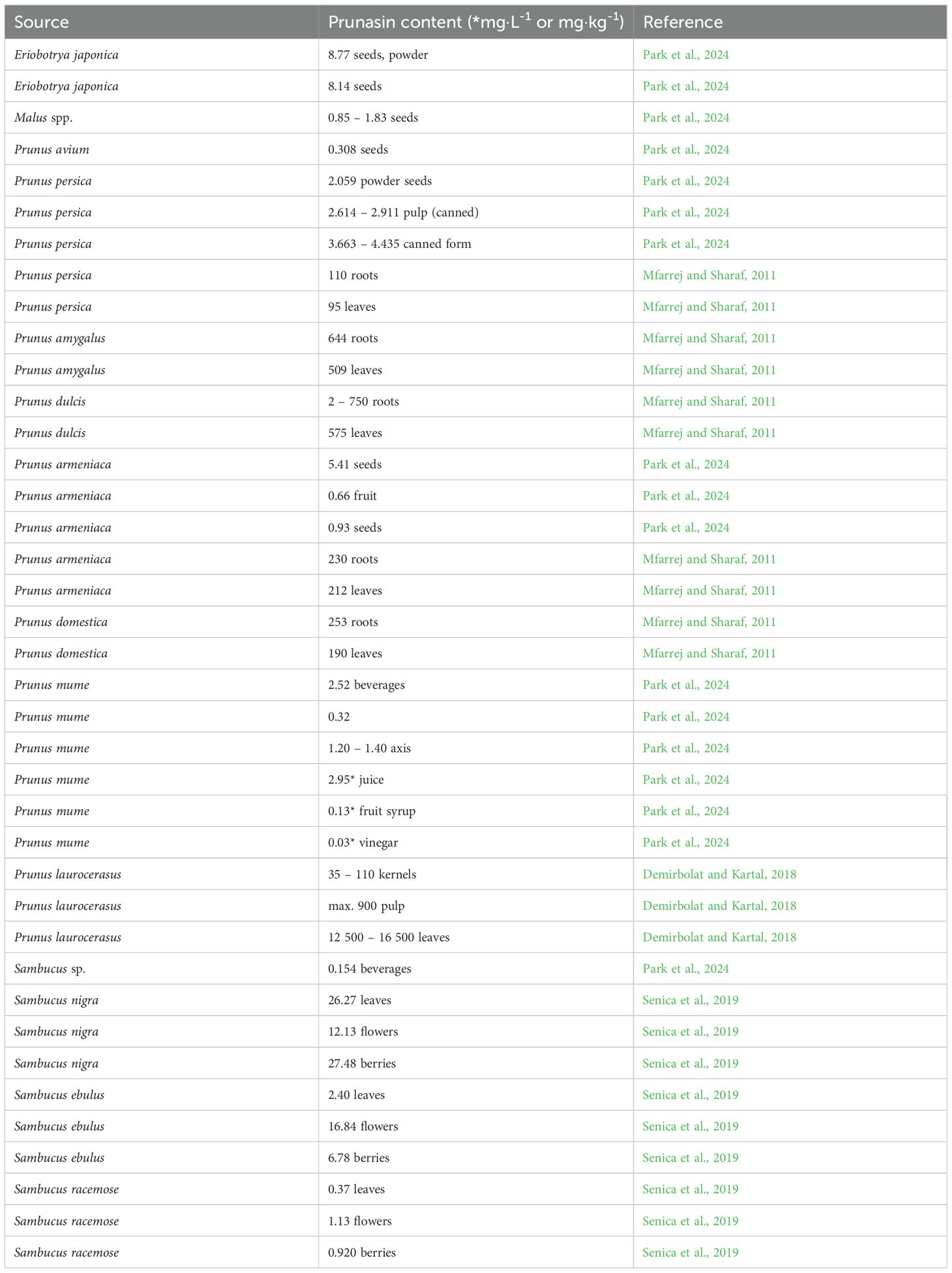
Table 3. Prunasin content in different plant species/foods.
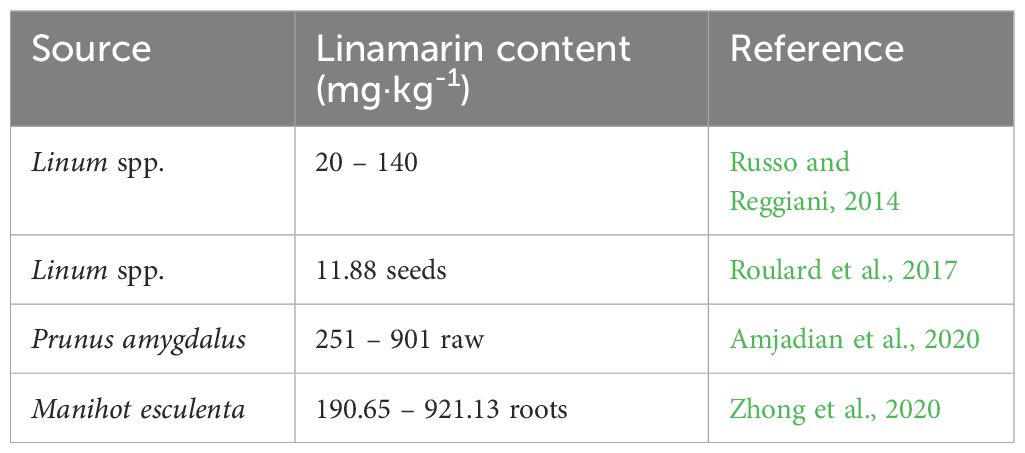
Table 4. Linamarin content in different plant species.
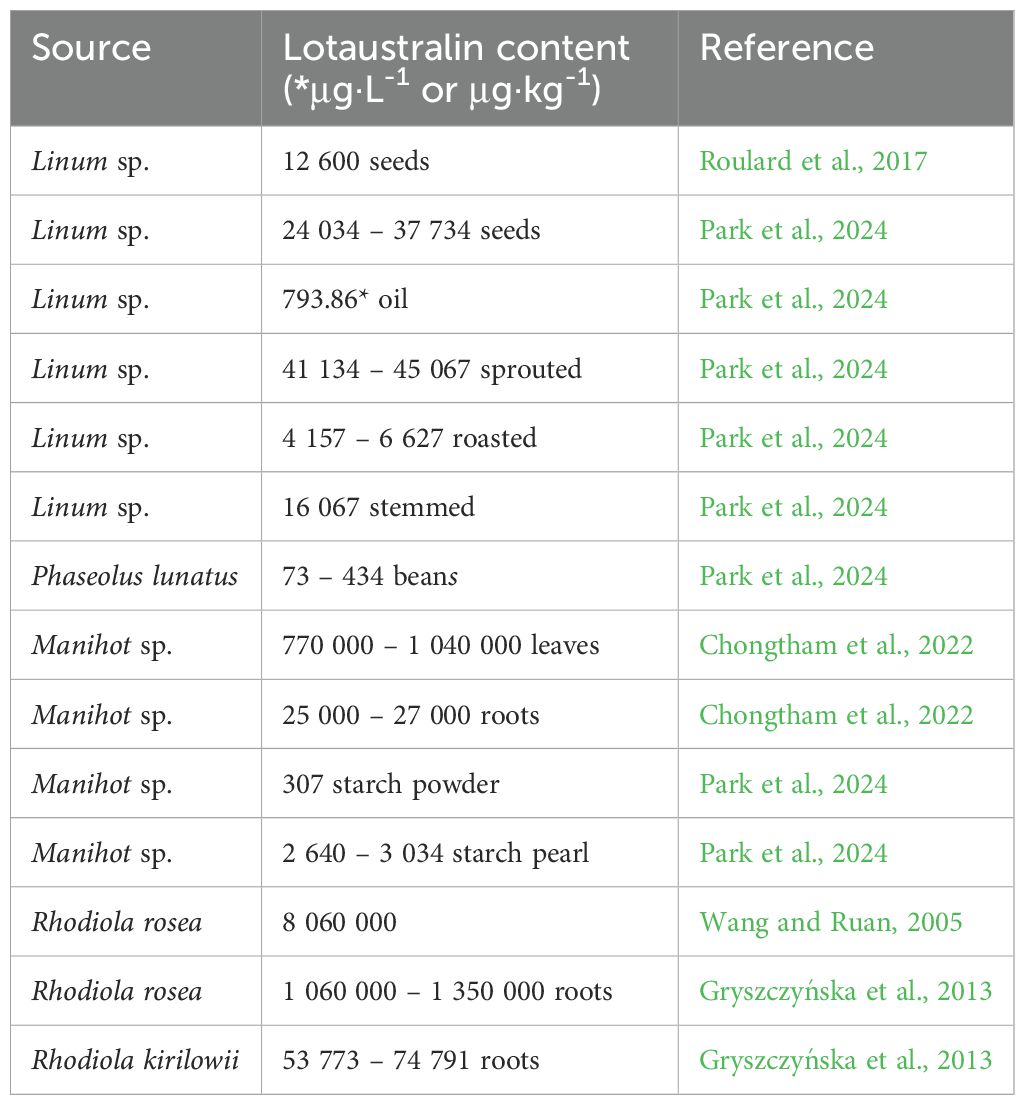
Table 5. Lotaustralin content in different plant species.
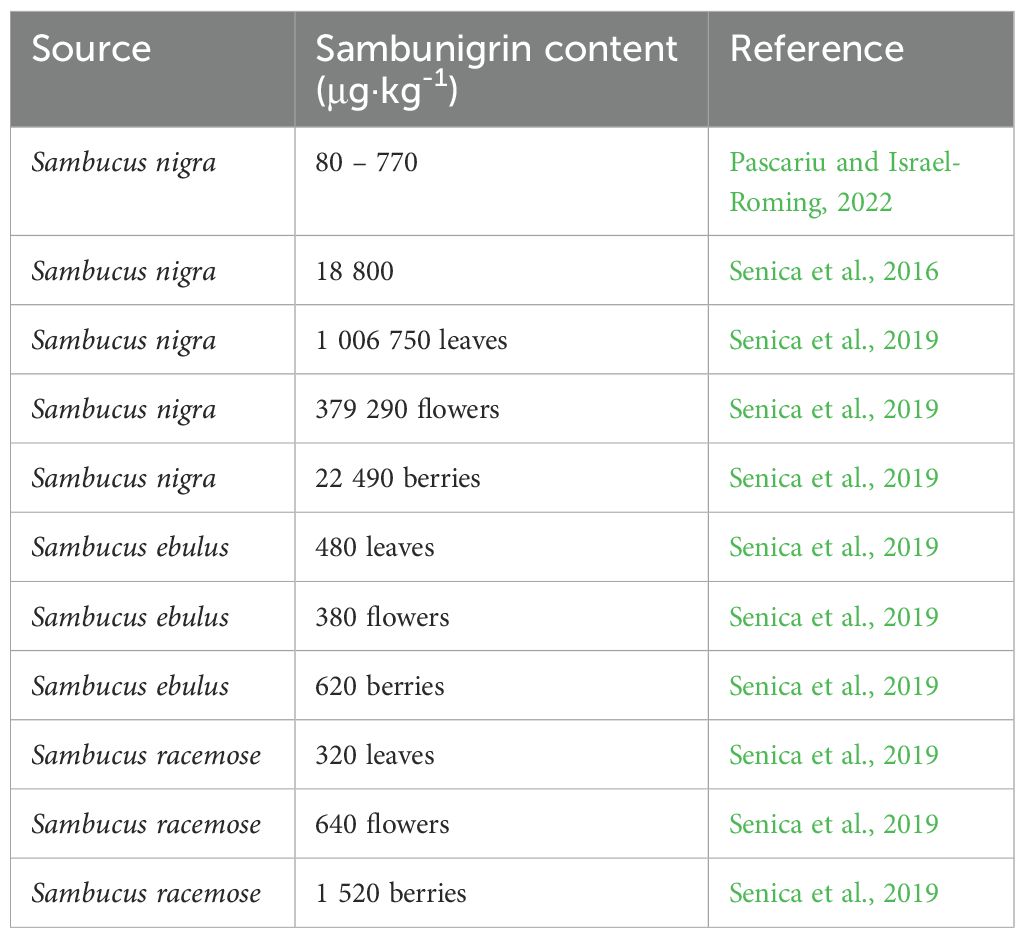
Table 6. Sambunigrin content in elderberries.
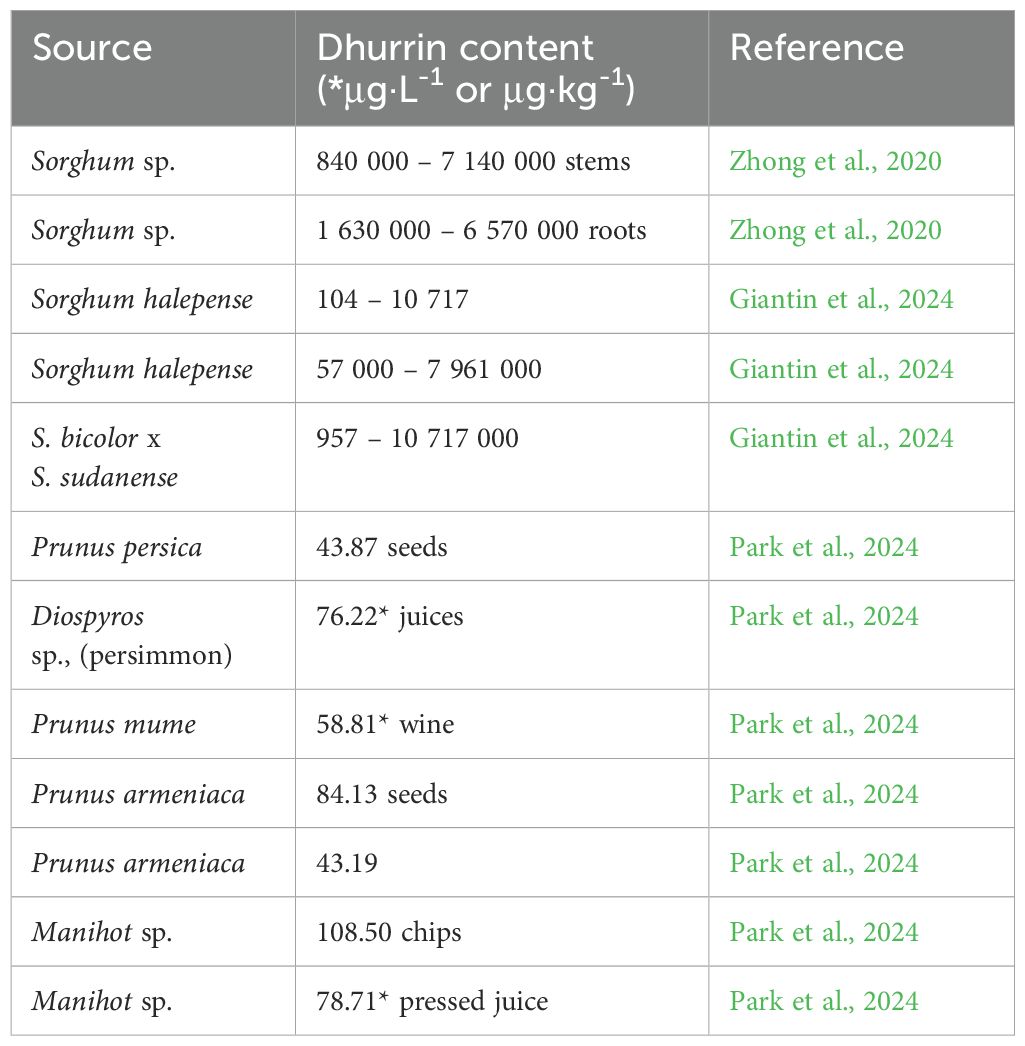
Table 7. Dhurrin content in different plant species.
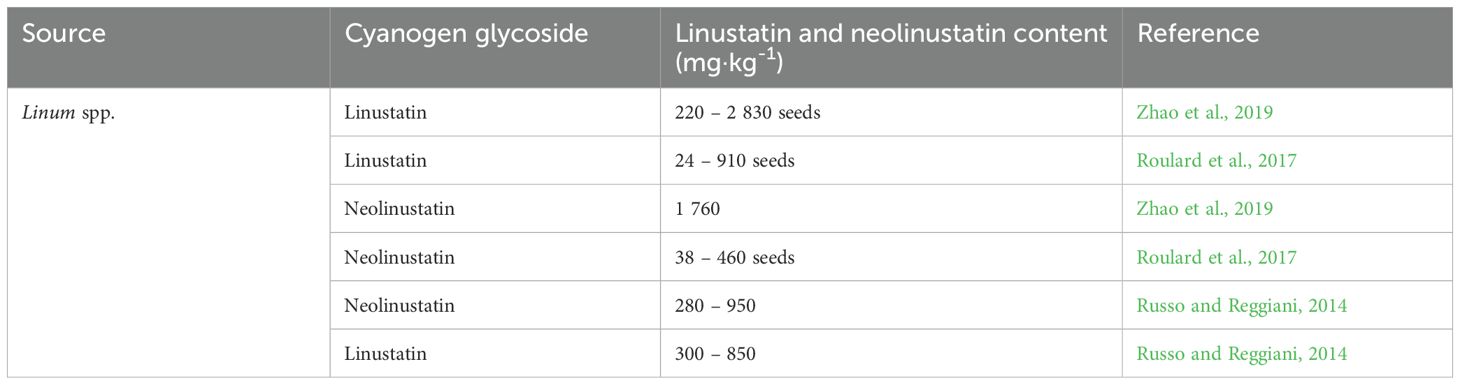
Table 8. Linustatin and neolinustatin content in flax.
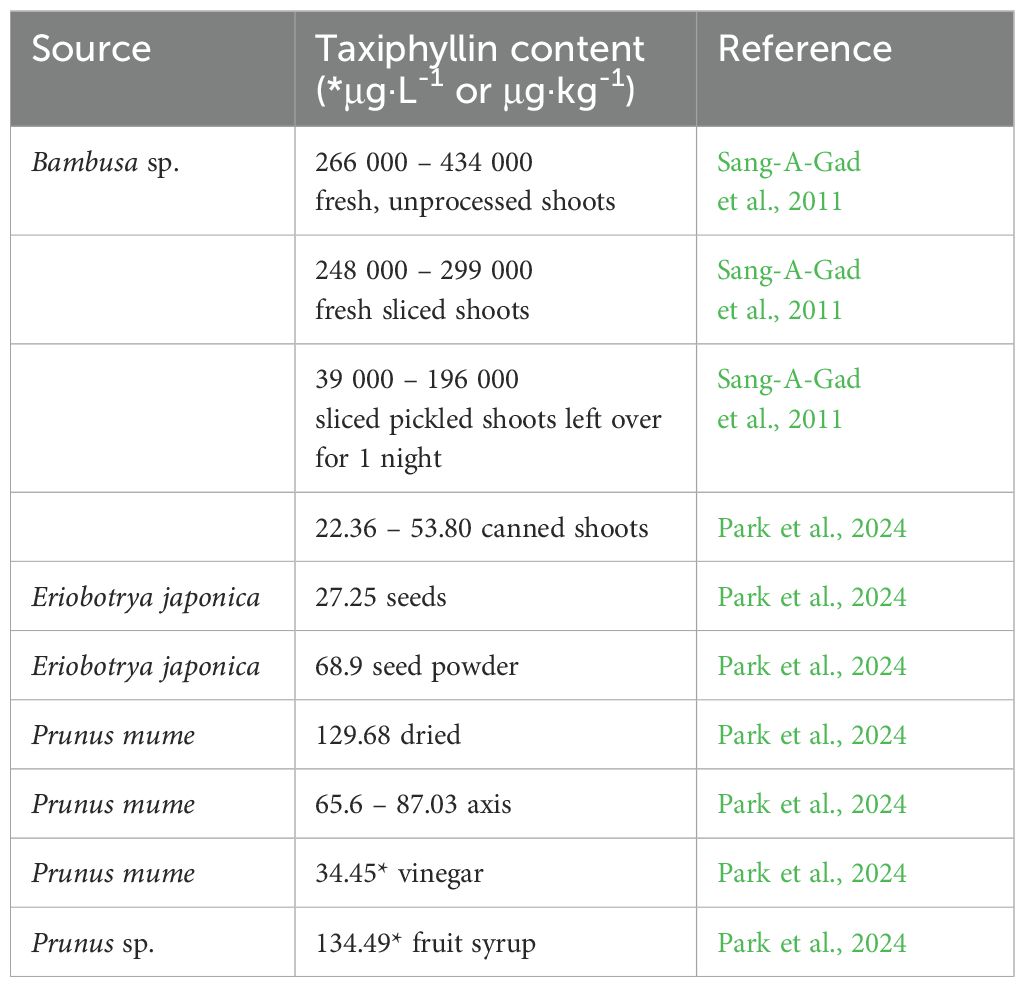
Table 9. Taxiphyllin content in different plant species.
Some specialized herbivores (mainly insects) preferentially feed on cyanogenic plants and use them as protection against predators. Several arthropod species (e.g., Diplopoda, Chilopoda, Insecta) can even synthesize CGs de novo. The unique plant-insect interaction based on CG is extensively discussed in the study by Zagrobelny et al. (2018).
5 Technologies for reducing cyanogenic glycoside content in foods
CGs are considered antinutrients that reduce the quality of feed and food, causing various health issues in animals, including humans. It is recommended that such plants be treated prior to consumption to minimize HCN content. Different types of processing methods are used to reduce CG content in plants. The most important processing methods include drying, grinding, dipping, peeling, ultrasound-assisted detoxification, autoclaving, soaking, boiling and fermentation (Bolarinwa et al., 2014a). The latter has proven to be highly effective, for example, in reducing CG content in bamboo shoots (Chongtham et al., 2022). In the process of acid fermentation of certain CGs, the bacteria Lactobacillus plantarum, Bacillus subtilis, Bacillus licheniformis, and Bacillus sonorensis proved to be effective (Abban et al., 2013; Menon et al., 2015). Sun drying after retting reduces cyanide content by 98.6%. Boiling/cooking can reduce free cyanide content by 96% within 15 minutes. After heating for 25 minutes, bound cyanide is reduced by 55% (Nampoothiri, 2017; Chongtham et al., 2022). A reduction of cyanides by 93% was also achieved by applying sodium bicarbonate (5 mL of a 0.4% NaHCO3 solution) to 1 g of cassava leaves (Latif et al., 2019). Conserved stone fruit must be peeled because cyanides also occur in the resulting infusion up to 33 mg·kg-1 HCN. However, the processing methods applied are not always sufficiently effective, and a certain amount of CG remains in plant products, thus posing potential health risks. Tables 2-9 also show varying amounts of CG in differently processed products. The issue of reducing cyanide content in plants and processed products is further explored by Rawat et al. (2015); Bolarinwa et al. (2016); Tahir et al. (2024) and others. Studies have also been developed to estimate the risks associated with the daily intake of CGs in food (Schrenk et al., 2019; Park et al., 2024).
6 Cyanogenesis, cyanide detoxification in plants and animals
When assessing the harmful effects of substances involved in cyanogenesis, the focus is mostly on the effects of released HCN; other components (intact glycosides and their hydrolysis products) do not appear to be serious in terms of acute toxicity. HCN is extremely toxic to animals, including humans. The lethal HCN dosage in most animal species is in the range of 2 mg·kg-1 to 2.5 mg·kg-1, with the exception of pandas (Clarke et al., 1981; Panter, 2018). The acute oral lethal dose of HCN for humans is reported to be 0.5 – 3.5 mg·kg-1 of body weight (Halstrom and Moiler, 1945). The permissible limit of cyanogen content in food is 500 mg·kg-1 (Food and Agriculture Organization, 2005).
HCN toxicity in animals, including humans, is due to blocking the release of energy from ATP (adenosine triphosphate) by inhibiting cytochrome oxidase activity in the respiratory chain (Figure 4). Hence the tissues and cells of the organisms are unable to utilize the oxygen that is transported by the blood, which can lead to internal suffocation (Gracia and Shepherd, 2004). The most important laboratory finding in cyanide poisoning is metabolic acidosis with dramatically increased lactate concentration (Baud et al., 2002) (Figure 4). The effects of HCN on the ability of the thyroid gland to store and process iodine are also documented (Erdogan, 2003). Clinical signs of acute poisoning include rapid breathing, decreased blood pressure and rapid pulse, dizziness, convulsions, vomiting, and blue discoloration of the skin due to lack of oxygen. As cellular hypoxia worsens, consciousness progresses to coma. Symptoms appear within seconds to minutes.
Cyanide detoxification in plants and animals is a critical biochemical process that helps mitigate the toxic effects of cyanogenic compounds. Both plants and animals have evolved mechanisms to detoxify or tolerate cyanide to survive in environments where these compounds are prevalent. The primary mechanism for cyanide detoxification in most plants is the β-cyanoalanine pathway. In this process, cyanide reacts with the amino acid L-cysteine to form β-cyanoalanine, catalyzed by the enzyme β-cyanoalanine synthase (CAS). This reaction occurs mainly in the mitochondria and is the major route by which plants detoxify endogenous cyanide. β-cyanoalanine can be further converted into asparagine, aspartate, and ammonia by β-cyanoalanine hydratase or nitrilase, integrating the cyanide-derived nitrogen into the plant’s nitrogen metabolism (Velišek and Hajšlova, 2009). The β-cyanoalanine synthase pathway is described in more detail by Machingura et al. (2016).
The most significant detoxification system in animals is the rhodanese enzyme system, which converts cyanide into thiocyanate, which is much less toxic and can be safely excreted through the urine (Gracia and Shepherd, 2004) (Figure 5). A further manner of detoxification is the binding of cyanide to hydroxocobalamin (vitamin B12), resulting in the formation of nontoxic cyanocobalamin.
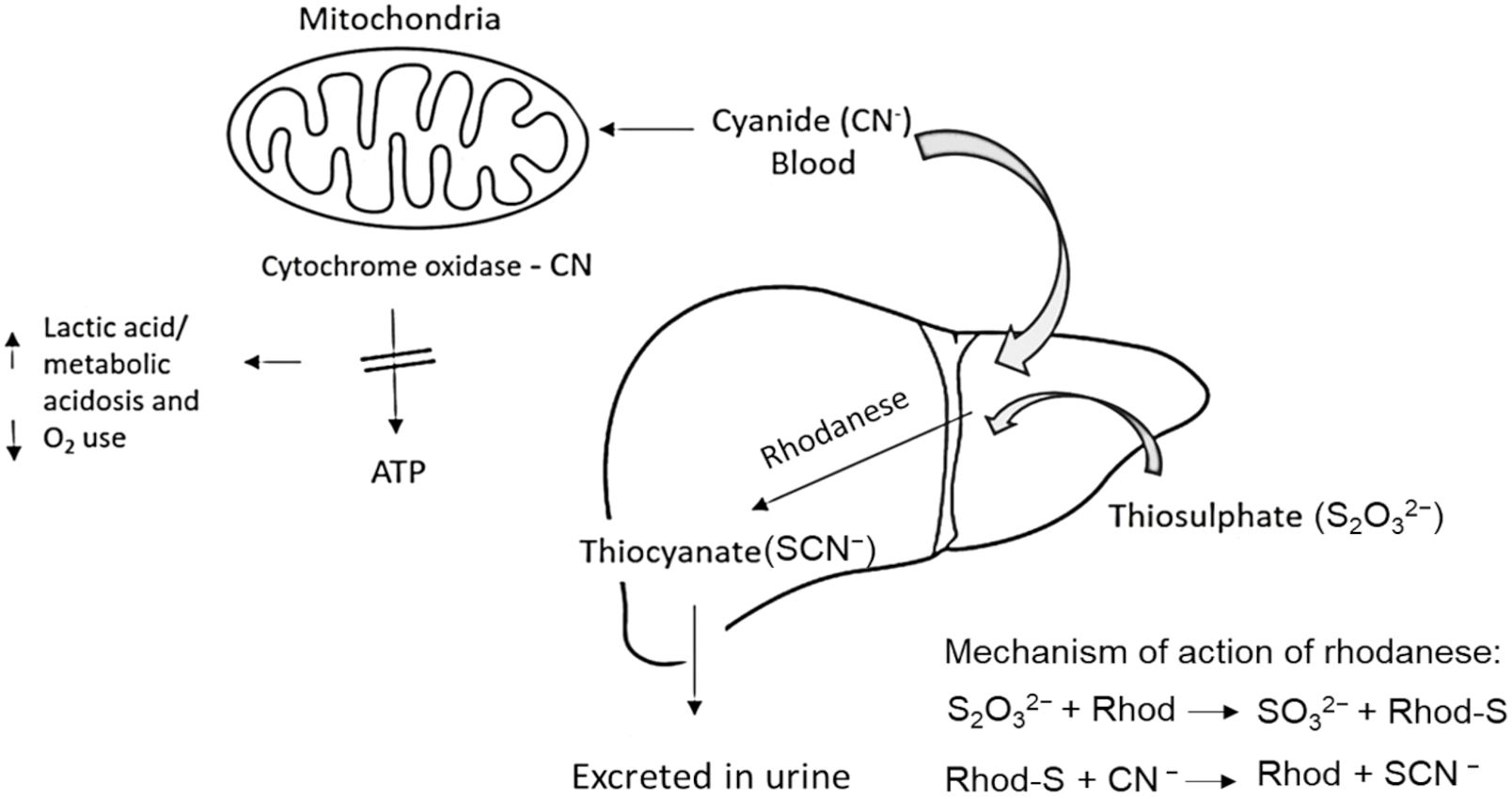
Figure 5. Detoxification of cyanide in animals. Cyanide, primarily absorbed through the skin and gastrointestinal tract, is relatively quickly converted in the liver by the enzyme rhodanese (Rhod) into the less toxic thiocyanate, which is excreted in the urine. Excess cyanide binds to cytochrome oxidase in mitochondria, leading to the inhibition of ATP production and the cessation of aerobic metabolism. The result is systemic hypoxia and the potentially death of the individual (adapted from: Gracia and Shepherd, 2004; Aussignargues et al., 2012).
The ability of an animal to tolerate certain doses of HCN also depends on the animal species, body weight, digestion rate, type of food, and the animal’s ability to detoxify the released HCN. The lethal dose for sheep is 2.4, cattle 2.0, mice 3.7, cats 2.0, 0, rats 0.5 – 10.0 and dogs 1.5 mg·kg-1 body weight (Jones, 1998). Ruminants are more sensitive to HCN poisoning because the enzymes that facilitate the release of HCN are destroyed by gastric HCl in these animals. Of this group, goats appear to be the most susceptible to cyanide (Patel et al., 2014). The specifics of CG poisoning in ruminants are described in detail by Gensa (2019). In non-ruminants, CGs are partially cleaved, and HCN is released only by the action of the colonic microflora where the pH is more suitable for the action of glycosides. But the hydrolysis is not complete, some glycosides are absorbed in their original form. In ruminants, many bacteria found in the rumen can hydrolyze CGs, with the degree of effectiveness depending on glycoside type and feed ration. The composition of gut microbiota also plays a significant role in the tolerance of mammals to the content of secondary metabolites in their diet. The gut microbiome of the giant panda and red panda contains a higher proportion of Pseudomonas bacteria compared to other mammals. Their microbiome is thus enriched with genes that encode the enzymes involved in the potential degradation or detoxification of HCN (Zhu et al., 2018). This is likely an evolutionary adaptation, that is not unique in the context of the plant kingdom, and can also be observed in some animals or microorganisms (Panter, 2018). Lemurs and gorillas also possess the unique ability to utilize high cyanide content in their diet without any acute or chronic harmful effects (Ballhorn et al., 2016). The metabolism of CGs in animals is described in more detail in the article by Cressey and Reeve (2019). However, estimating the health risks related to consuming CG-containing plants is often not straightforward, as sometimes the entire plant, including seeds, is consumed, while in other cases only the fruit or other parts are eaten. However, chronic intoxication can occur if plants containing CGs are part of the daily diet and consumed in larger quantities (Hartanti and Cahiani, 2020). More detailed data on the daily intake of CGs in selected foods is documented by Park et al. (2024).
7 Diseases caused by the toxic effects of cyanoglycosides
Chronic cyanide toxicity causes several diseases, especially in tropical areas where the main food is plant-based. Growth retardation, goiter, and cretinism are relatively common diseases in developing countries where people consume food with very low iodine content (<100 μg/day) and high cyanide content (Odo et al., 2014; Bolarinwa et al., 2016). High and sustained intake of cyanogens in sublethal concentrations of manioc (cassava) flour in combination with the low intake of sulfur amino acids also causes konzo - a disease of the upper motor neurons characterized by irreversible but non-progressive symmetric spastic para/tetraparesis which mostly affects children and women in the tropics (Howlett et al., 1990). Konzo disease is common during periods of drought, or when there is a shortage of alternative foods due to unfavorable social or environmental factors (Nzwalo and Cliff, 2011). Practically identical to konzo disease is mantakassa (Howlett et al., 1990; Bruyn and Poser, 2003). Tropical ataxic neuropathy (TAN) is another health problem associated with the continual consumption of improperly processed cassava products, especially in Africa (e.g. Nigeria). TAN is used to describe several neurological syndromes attributed to toxicolutricative causes. Symptoms of TAN include tongue pain, optic atrophy, neurosensory deafness, and sensory gait ataxia (Bolarinwa et al., 2016). Diseases caused by CGs are described in more detail in the study by Nyirenda (2020).
8 Significant cyanogenic glycosides
8.1 Amygdalin
Amygdalin ([(6-O-β-D-glucopyranosyl-β-D-glucopyranosyl)oxy](phenyl)acetonitrile, D-mandelonitrile β-D-gentiobioside) was isolated from apricot stones [Prunus dulcis (Mill.) D. Webb var. amara (DC.) H. Moore] by Pierre-Jean Robiquet and Antoine François Boutron Charlard in 1830 (Rosen and Shorr, 1979). As one of the most common CGs, it occurs in 1,200+ plant species. Amygdalin is highly concentrated in plants of the Rosaceae, Caprifoliaceae, and Oleaceae families (Hösel, 1981). Amygdalin is colorless with a melting point of 213°C, insoluble in non-polar solvents, and is highly soluble in ethanol and moderately soluble in water. Its highest concentrations are found in the seeds of fruits, which have a characteristic bitter taste due to the presence of amygdalin. Apricot seeds contain the highest amount of amygdalin, up to 2 – 2.5% by weight in most varieties of apricots (Table 2). Amygdalin content is significantly lower in seedless fruits (0.01 – 2.96 mg·g-1) and also in processed products (0.004 – 0.12 mg·g-1). In commercially available apple juices, the amygdalin content ranges from 0 to 0.007 mg·ml-1 (Bolarinwa et al., 2014b). A semisynthetic derivative of amygdalin is the structurally different laetrile (mandelonitrile-β-glucuronide) (He et al., 2020).
Amygdalin’s effects began to be studied from the late 1960s until the mid-1980s, whereby such research included tests to determine the chronic and acute toxicity and teratogenicity of amygdalin. The results showed that this substance’s toxicity depends on the manner of administration and dose. Adverse effects were shown to the least extent when administered intravenously and intramuscularly, while higher toxicity was recorded when administered orally (Beamer et al., 1983). With oral amygdalin intake, HCN poisoning from plant sources is not manifested until after a certain latency period, namely approximately 15–60 minutes (Kolesárová et al., 2021). The lethal dose for humans of intravenous injection of amygdalin is 5 g (Qadir and Fatima, 2017). The consumption of 50 bitter almonds is deadly for adults. However, for young children, 5–10 almonds are fatal (World Health Organisation, 2012). Higher doses of amygdalin causes symptoms of intoxication, nausea, and bluing of the skin. Regular use of amygdalin can cause nervous system problems and, ultimately, its disabling. Intoxication in humans is manifested by headache, dizziness and confusion, in severe cases by paralysis, coma, and death of the affected person (Sadoff et al., 1978). Omelka et al. (2021) focused their study on the effect of amygdalin on human osteoblast functions in vitro, and demonstrated that amygdalin at high concentrations (10 mg·ml-1) negatively affects osteoblasts, increases bone resorption, and reduces osteoblast viability. At high concentrations it also had a negative impact on the oxidative balance of male reproductive structures (Ďuračka et al., 2016).
Yet amygdalin is considered an important component of alternative medicine due to its wide range of healing effects (Kolesárová et al., 2021). At lower doses, it has positive effects in the treatment of asthma, bronchitis, diabetes, leprosy, vascular lesions, and sickle cell disease (Fukuda et al., 2003; Makarević et al., 2014; Song and Xu, 2014; Zhou et al., 2020). It can also relieve fevers, coughs, and thirst. Traces of released HCN and benzaldehyde from the amygdalin molecule can eliminate the occurrence of bacteria in the oral cavity, which is the cause of tooth decay and bad breath (Griffin, 1974). However, the anticancer effects of amygdalin have attracted the most attention. The use of bitter almond derivatives in the treatment of skin tumors is mentioned in 5,000-year-old Egyptian papyri. In the 1920s, apricot kernels were recognized in many states as a preventive and malignant inhibitor of cancer cell growth. The success of apricot kernels in cancer treatment was also supported by the American biochemist Ernst Theodore Krebs, who was the first to present amygdalin under the incorrect designation “vitamin B17” (Krebs, 1970) and believed that together with diet and vitamins, this substance could prevent cancer growth (Chandler et al., 1984). Amygdalin administration became one of the most popular and unconventional anti-cancer treatments in the 1970s, and has been used by 70% of American cancer patients since 1978 (Barakat et al., 2022). Howard and Miller (1984) report that while laetrile has shown little antitumor activity in animal studies, no antitumor activity has been reported in clinical trials in human populations. A similar conclusion was reached by Jaszcak-Wilke et al. (2021). Critics of amygdalin use warn that amygdalin is ineffective and even toxic, and say that its accumulation leads to severe poisoning (Blaheta et al., 2016). Side-effects associated with laetrile toxicity reflect symptoms of cyanide poisoning, including liver damage, difficulty walking, fever, subsequently coma, and eventually death. Amygdalin is still marketed as an anti-carcinogenic “vitamin B17” in many countries, and the United States (where this theory originated) has long proven its danger and ineffectiveness as a cancer treatment (Süli et al., 2017). But the results of many molecular biology studies have again highlighted the increased anti-tumor potential of amygdalin (Fukuda et al., 2003; Makarević et al., 2014; Song and Xu, 2014; Zhou et al., 2020). In the case of colon cancer, a decrease in the expression of many genes associated with growth functions, apoptosis and trafficking was observed at amygdalin concentrations of 0.25–5 mg·ml-1 (Kim et al., 2016). When tested on other cell lines with a concentration of 10 mg·ml-1 of amygdalin, the growth rate of breast cancer cells (MCF-7 and MDA-MB-231) was inhibited (Lee and Moon, 2016), sometimes leading to a decrease in cell motility or a reduction in the ability to synthesize collagen and fibronectin in the case of kidney cancer cells (Caki 1, A498, KTC-26) (Luo et al., 2016). The physiological and therapeutic effects of amygdalin are described in more detail in Kolesárová et al. (2021).
8.2 Prunasin
The cyanogenic monoglycoside prunasin (Prulaurasin, Laurocerasin, (2R)-(β-D-glucopyranosyloxy)(phenyl)acetonitrile, D-Mandelonitrile β-D-glucoside) is formed by removing one of the two β-D-glucopyranosyl groups from amygdalin with the enzyme β-glucosidase (Ellenhorn and Barceloux, 1997). Prunasin is a component of over 3,000 plant species, occurring mainly in plant tissues of the families Myrtaceae, Saxifragaceae, and Scrophulariaceae, and especially found in the families Rosaceae and Polypodiaceae (Vetter, 2017). Prunus (P.) species containing prunasin include e.g. P. armeniaca (apricot), P. dulcis (bitter almond), P. persica (peach), P. serotina (black/wild cherry), P. virginiana (red almond) and P. laurocerasus (cherry laurel) (Hodgson, 2012; Demirbolat and Kartal, 2018) (Table 3). Prunasin is synthesized by plants to protect seeds during maturation. Evident proof of this is provided by the study by Demirbolat and Kartal (2018), which highlighted the increasing content of prunasin in seeds (from an initial 3.5 mg·100 g-1 to 11 mg·100 g-1). During fruit maturation, such content decreases and eventually disappears. In the leaves, the average content is maintained (1250 to 1650 mg·100 g-1). To date, there is little data on the toxicokinetics of prunasin in humans. Prunasin intoxication due to the frequent consumption of some plants is common especially in tropical areas, and is associated with motor neuron diseases, such as console and mantakassa (Howlett et al., 1990; Bruyn and Poser, 2003). Toxic effects are particularly evident when fruits or preparations containing prunasin are taken concomitantly with foods containing high levels of β-glucosidase enzymes (such as apple and pear seeds).
In experiments performed in 2003 under in vivo and in vitro conditions, the antitumor activity of this compound was observed. It comprised the strong inhibition of the activation of the Epstein-Barr virus antigen induced by the tumor promoter. The researchers also observed a delay in the onset of skin cancer in mice (Fukuda et al., 2003). Prunasin can also be found in extracts from Prunus mume (Ume), which exhibit hepatoprotective, anti-inflammatory, antioxidant, antibacterial and anticancer properties. MK615 is a mixture of extracts containing Ume-derived hydrophobic substances (Morimoto-Yamashita et al., 2012). The antitumor properties of MK615, along with other extracts from Prunus mume, have been studied. Research has shown that MK615 inhibits proliferation and induces apoptotic cell death in a variety of cancer cells, including those from both solid and hematological tumors (Bailly, 2020). P. amygdalus var. amara treatment also significantly decreased cancer cell growth in most cancer cell lines, when doses and exposure time were taken into consideration (Shalayel et al., 2023).
8.3 Linamarin
Linamarin (formerly called phaseolunatin, 2-(β-D-glucopyranosyloxy)-2-methyl-propanenitrile, α-hydroxyisobutyronitrile β-D-glucose) is a derivative of valine and isoleucine. The data on linamarin content in plant tissues is very limited (Table 4). It is found in the leaves and roots of plants such as almond (Prunus amygdalus), flax (Linum usitatissimum) and manioc (Manihot esculenta). In manioc, linamarin represents more than 80% of all CGs (Kuete, 2014). Manioc, also known as cassava or yuca, is a major source of carbohydrates for some 500 million people worldwide, particularly in Africa, where it is the third most important food source. According to linamarin content, hot and sweet varieties of cassava are distinguished with both varieties being commonly consumed. But under certain circumstances, it becomes dangerous and even fatal for humans. Although cassava juice contains low protein content, it also contains a relatively large amount of CGs, especially linamarin and lotaustralin (Nassar and Dorea, 1982). These CGs are hydrolyzed in the presence of the enzyme linamarase (Hösel, 1981). Chronic linamarin poisoning is manifested by the occurrence of endemic tropical ataxic neuropathy (TAN), especially in the elderly, the development of console disease, and deteriorating health with a number of symptoms resulting from iodine deficiency (Howlett et al., 1990; Ernesto et al., 2002). Recent studies have highlighted the potential antineoplastic effect of linamarin, especially when administered with the activating enzyme linamarase. The application of linamarin together with linamarase shows cytotoxic effects against several cancer cell lines, including HT-29, MCF-7, Caov-3, and HeLa (Yusuf et al., 2006; Idibie et al., 2007; Mosayyebi et al., 2020). The toxicity of cyanide released during the action of linamarase is eliminated by using so-called ‘suicide gene therapy’, the principle of which involves introducing the desired gene into a cancer cell to convert non-toxic compounds into toxic substances at the tumor site (Zarogoulidis et al., 2013). The principle of this mixture’s cytotoxic effect is described in more detail by Liyanage et al. (2024). Song and Xu (2014) also mention the possible mechanism of linamarin, where the effect of HCN on the mitochondrial respiratory chain can lead to the death of cancer cells.
8.4 Lotaustralin
Lotaustralin (2-hydroxy-2-methylbutyronitrile-β-D-glucopyranoside or (2R)-2-(β-D-gluc opy- ranosyloxy)-2-methylbutanenitrile) is a CG found in plants of the families Linaceae (e.g. Linum usitatissimum), Euphorbiaceae (e.g. Manihot esculenta), Fabaceae (e.g. Phaseolus lunatus), and Crassulaceae (e.g. Rhodiola rosea) (Pulido and Gill, 2013) (Table 5). The methyl derivative of linamarin – lotaustralin and linamarin itself are the two main CG compounds in foods derived from manioc roots, while the content of HCN and cyanohydrins is generally low (Butter, 1965; Mlingi et al., 1995). The sweet type of cassava contains 50 times lower levels of this CG compared to its bitter version. The concentration of lotaustralin increases significantly, especially during the dry season (Bovell-Benjamin and Roberts, 2016). Compared to linamarin, as the main cyanogenic component (93%) present in manioc lotaustraline content is much lower (7%) (Liangcheng et al., 1995).
8.5 Sambunigrin
Sambunigrin (also known as L-prunasin or (2S)-(β-D-glucopyranosyloxy)-(phenyl)- acetonitrile) was isolated in 1905 from the leaves of black base (Sambucus nigra) by the French scientists, pharmacists and botanists J.L.L. Guignard and Dr. E. Bourquelot. It also occurs in the American species S. racemosa - S. calicarpa Greenea, S. microbotrys Rydberg (Hegnauer, 1989). Much less sambunigrin occurs in the North American elder (Sambucus canadensis) (Bohm and Glennie, 1971; Buhrmester et al., 2000). Sambunigrin content means that the unripe fruits of black elder must not be consumed directly. Likewise, all green parts of the plant contain toxic CGs. However, the flowers and ripe fruits no longer contain this substance. The (R) diastereomer of sambunigrin is R-prunasin. Heat treatment causes the decomposition of sambunigrin into compounds that are harmless to the human body (Młynarczyk et al., 2018).
In addition to sambunigrin, other CGs, such as amygdalin, dhurrin, prunasin, linamarin, zierin, and holocalin, have also been detected in the tissues of various elderberry species (Knudsen and Kaack, 2015; Appenteng et al., 2021). The most abundant include sambunigrin, amygdalin and prunasin (Senica et al., 2016, 2019; Pascariu and Israel-Roming, 2022), where sambunigrin predominates in red elderberry, prunasin in black elderberry, and amygdalin in dwarf elderberry (Senica et al., 2019).
8.6 Dhurrin
Dhurrin ((2S)-(β-D-glucopyranosyloxy)(4-hydroxyphenyl)acetonitrile, (S)-4-hydroxy- mandelonitrile β-D-glucoside) is a CG produced in many plants elonging to Poaceae, Rosaceae, Araliaceae, Proteaceae, Betulaceae, Chenopodiaceae, Proteaceae, Boraginaceae etc (Miller et al., 2006; Yadav et al., 2023). Dhurrin, discovered in several varieties of Sorghum in 1906 as being responsible for bovine poisoning by HCN, is most often associated with the species Sorghum bicolor (Poaceae) (Mao and Anderson, 1965). Although dhurrin provides plants with an effective defense against most herbivores; however, some beetles and aphids have developed mechanisms that allow them to resist its toxic effects (Pentzold et al., 2014). Dhurrin occurs in whole plants except mature seeds (grains) (Yadav et al., 2023), and it is toxic mainly to farm animals that consume it (Kojima et al., 1979). The biosynthesis, catabolism, and toxicity of dhurrin are described in more detail in Yadav et al. (2023). The concentration of dhurrin in tissues decreases with the age of plants, and is highest during seed germination when it reaches about 30% of shoots’ dry matter. Its content in tissues also increases due to various stress factors, mainly drought (Busk and Møller, 2002; Emendack et al., 2018). In addition, high concentrations of nitrate, also potentially toxic to ruminants, may accumulate during or shortly after periods of drought (Yadav et al., 2023). Sorghum malt contains a high amount of dhurrin (up to 1.400 mg·kg-1) that is insufficiently degraded in the malting process, so African beers may contain higher amounts of cyanide (about 11 mg·kg-1) (Tokpohozin et al., 2016). Dhurrin content in different plant species is shown in Table 7.
8.7 Linustatin and neolinustatin
Linustatin (2-{[6-O-(β-D-glucopyranosyl)-β-D-glucopyranosyl]oxy}-2-methylpropanenitrile) and neolinustanin [(2R)-2-{[6-O-(β-D-glucopyranosyl)-β-D-glucopyranosyl]oxy}-2-methyl-butane-nitrile)] are soluble in water and forms a weakly acidic solution. These CGs have so far been quantified only in flax (Table 8). Until recently, linamarin had been considered the main glycoside in flax seeds; however, much higher levels of the diglycosides linustatin and neolinustatin have been demonstrated (Russo and Reggiani, 2014). Therefore, consuming flax seeds in large quantities is not recommended. According to Daun et al. (2003), to reach acute cyanide toxicity, a person would need to consume eight cups (1 kg) of ground flaxseed. With the recommended daily intake of about one to two tablespoons, approximately 5–10 mg of HCN would be released (Rosling, 1994). When consuming ground flaxseed, the bioavailability of HCN and human exposure levels are higher than when consuming whole flaxseeds or when they are heat-treated. Cassava contains significantly more CGs than flaxseed (Touré and Xueming, 2010). Some sources (Smith et al., 1980) suggest that these CGs can protect rats from the toxic effects of selenium.
8.8 Taxiphyllin
Taxiphyllin is the (R)-enantiomer of dhurrin (2R)-(β-D-glucopyranosyloxy)(4-hydroxyphenyl)acetonitrile or (R)-4-hydroxymandelonitrile β-D-glucoside). Taxiphyllin is a CG found in bamboo shoots, Sorghum bicolor and Henriettella fascicularis (Calderón et al., 2003). Taxiphyllin is highly unstable and thermolabile. Although the taxiphyllin content in bamboo shoots is much higher than in cassava roots, the cyanide content in bamboo shoots decreases substantially following harvesting and processing. An approximately 80% reduction in CGs was achieved after vacuum freeze-drying for 24 hours at −50°C (Rawat et al., 2015). Taxiphyllin content in bamboo shoots is shown in Tables 1, 9. As mentioned earlier, pandas can withstand the toxic effects of bamboo CGs due to the composition of their gut microbiome. Taxiphyllin content in different plant species is shown in Table 9.
8.9 Triglochinin
Triglochinin ((2Z,4E)-4-[cyano(β-D-glucopyranosyloxy)methylene]-2-hexenedioic acid) was isolated from the flowers of the monocotyledonous plant Triglochin maritimum L (Eyjólfsson, 1970). Using chromatographic methods, two isomers of this compound were identified. This tyrosine-derived CG was later found in the tissues of Alocasia macrorrhiza, Thalictrum aquilegiifolium, and some plants from the Araliaceae family (Aralia spinosa) (Lechtenberg et al., 2022), as well as the Arecaceae family (Nahrstedt, 1975). The content of triglochinin in tissues is limited to certain periods of collection or developmental stages. In the case of A. spinosa, the flower buds collected in July showed the highest content of triglochinin, just below 0.2% dry weight. There is very little data on triglochinin content in plant tissues. Chongtham et al. (2022) measured 29–32 mg·kg-1 (as HCN equivalents) of this CG in the tissues of giant taro (Alocasia macrorrhizos).
9 Detection of cyanogenic glycosides
The detection of CGs in food is important for public health protection, as improper food processing can release toxic cyanide that is highly harmful to humans. Additionally, the detection of these substances plays an important role in complying with food regulations, which set maximum allowable concentrations of CGs in various foods (Cressey and Reeve, 2019; Vetter, 2000). Many countries have already introduced regulations to reduce the risk of cyanide exposure from consuming foods that contain these compounds.
The detection of CGs depends on several factors (Cressey and Reeve, 2019; Tahir et al., 2024), such as:
1. Type and concentration of CGs – different types of CGs may have varying abilities to release cyanide at different concentration.
2. Molecular structure of CGs – differences in chemical structure affect how these glycosides behave during detection, and what methods are most suitable for their extraction and identification.
3. Composition of an individual’s gut microbiome – microbial content in the digestive system can influence the metabolism of CGs and cyanide production.
4. Extraction method – the way CGs are extracted from the sample (e.g., using different solvents, temperatures, or extraction times) affects the efficiency and accuracy of detection (Vetter, 2000).
5. Quantification method used – various analytical techniques (e.g., HPLC – High-Performance Liquid Chromatography, GLC = Gas-Liquid Chromatography) may have different sensitivities and specificities when measuring CGs content.
6. Presence of other substances – other compounds in the sample may interfere with the detection process or affect measurement accuracy.
CGs are quantified using direct and indirect methods of determination. The direct method targets CGs as the molecules of interest, while the indirect method focuses on the released HCN after hydrolysis (Azmi, 2019; Hartanti and Cahiani, 2020) (Figure 6). One of the most well-known indirect methods of determination is the Guignard sodium picrate test (Hartanti and Cahiani, 2020).
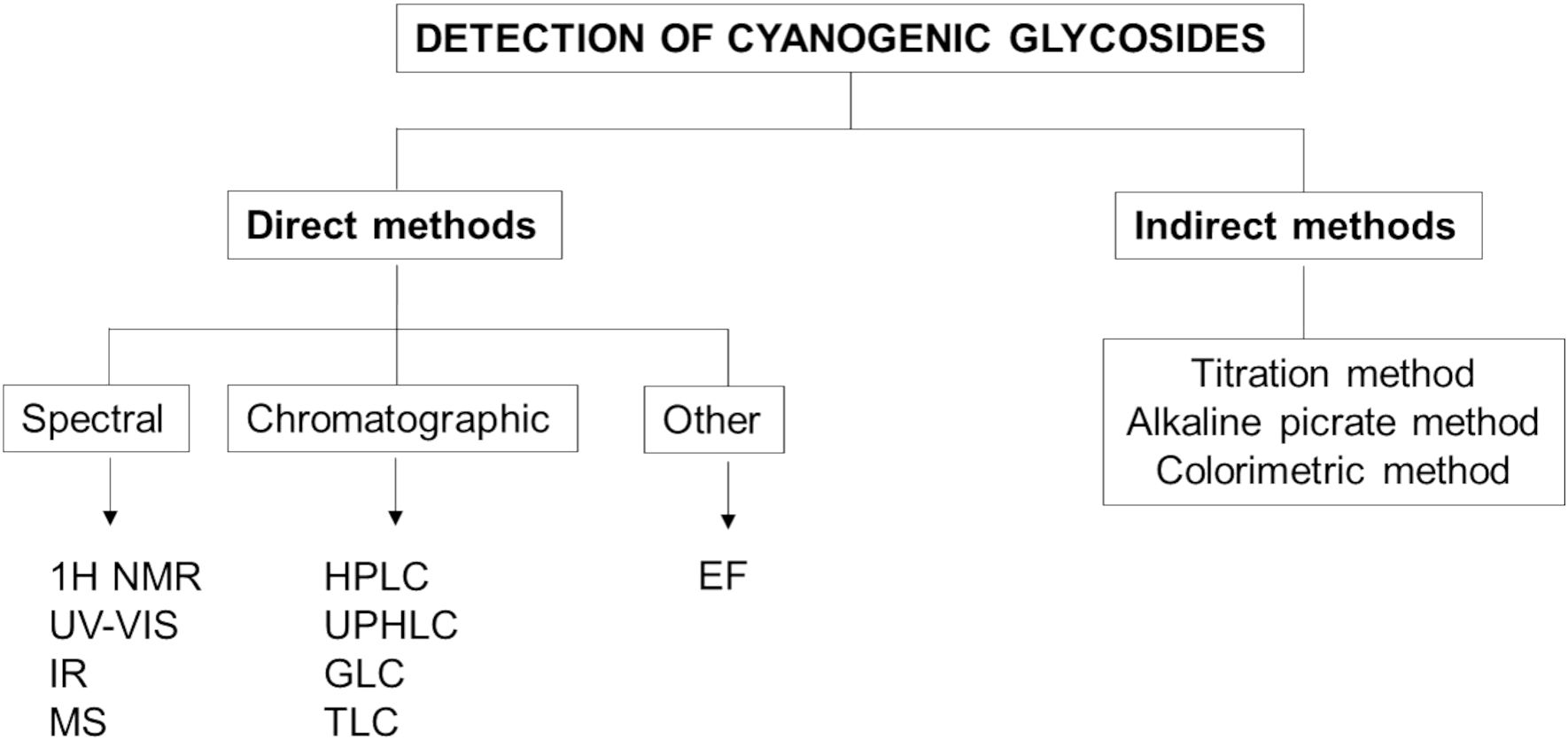
Figure 6. Methods used for the detection of cyanogenic glycosides (1H NMR, Proton Nuclear Magnetic Resonance; UV-Vis, Ultraviolet-Visible Spectroscopy; IR, Infrared Spectroscopy; MS, Mass Spectroscopy; HPLC, High-Performance Liquid Chromatography; UPHLC, Ultra-Performance Liquid Chromatography; GLC, Gas-Liquid Chromatography; TLC, Thin Layer Chromatography; EF, Electrophoresis).
Many reviews summarize this issue. Analytical methods for the determination of amygdalin are clearly presented by Popa et al. (2021) who highlight various analytical methods with detailed parameters. Zhao et al. (2024) focus on the issue in the comparison of HPLC/UPLC methods for the determination of CGs. Risk assessment of food safety associated with foods containing CGs was addressed by Cressey et al., with a focus on rural New Zealand (Cressey et al., 2022).
10 Conclusion
Cyanogenic glycosides represent a broad group of structurally differing compounds with various biochemical properties. Some organisms use cyanogenic acids as protection against predators. These compounds are also present in many plants, which in some countries form an important part of the diet for local populations. The harmful effects of CGs on the human body are fairly well researched, and there is a vast database of scientific studies on their toxic properties. The risks associated with the consumption of processed and unprocessed plant parts containing these substances can now be more accurately estimated. Although cyanide itself is extremely toxic and can cause severe poisoning, some plants containing CGs are the subject of intensive research, especially for their potential in therapeutic applications. Current studies are focusing on the synthesis of derivatives of these compounds that have enhanced anti-tumor effects, which opens up new opportunities for cancer treatment. However, it is essential that the risks associated with the release of cyanide, which remains highly toxic, are not overlooked in this research. As a result, much research is focused on developing technologies and methods that allow the breakdown of cyanide compounds to be controlled or minimized with the aim to avoid adverse health effects.
Author contributions
BP: Project administration, Formal analysis, Conceptualization, Supervision, Writing – original draft, Writing – review & editing, Funding acquisition. JJ: Visualization, Formal analysis, Writing – original draft, Software, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Ministry of Education, Research, Development and Youth of the Slovak Republic, grant numbers VEGA 2/0034/25.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abban, S., Brimer, L., Abdelgadir, W. S., Jakobsen, M., and Thorsen, L. (2013). Screening for Bacillus subtilis group isolates that degrade cyanogens at pH 4.5–5.0. Int. J. Food Microbiol. 161, 31–35. doi: 10.1016/j.ijfoodmicro.2012.11.019
Abdulrashid, M. and Agwunobi, L. N. (2009). Taro cocoyam (Colocasia esculenta) meal as feed ingredient in poultry. Pak. J. Nutr. 8, 668–673. doi: 10.3923/pjn.2009.668.673
Aikman, K., Bergman, D., Ebinger, J., and Seigler, D. (1996). Variation of cyanogenesis in some plant species of the midwestern United States. Biochem. System Ecol. 24, 637–645. doi: 10.1016/S0305-1978(96)00069-5
Amjadian, O. A., Arji, I., Changizi, M., Khaghani, S., and Salehi, H. R. (2020). Determination of cyanogenic glycosides in endemic species of wild almond seeds in the Zagros Mountains. Rev. Bras. Bot. 43, 697–704. doi: 10.1007/s40415-020-00668-7
Appenteng, M. K., Krueger, R., Johnson, M. C., Ingold, H., Bell, R., Thomas, A. L., et al. (2021). Cyanogenic glycoside analysis in American elderberry. Molecules 26, 1384. doi: 10.3390/molecules26051384
Armstrong, H. E., Armstrong, E. F., and Andhorton, E. (1913). Herbagestudies II – Variation in Lotus corniculatus and Trifolium repens (cyanophoric plants). R. Soc Proc. B. 86, 262–269. doi: 10.1098/rspb.1913.0021
Aussignargues, C., Giuliani, M. C., Infossi, P., Lojou, E., Guiral, N., Giudici-Orticoni, M. T., et al. (2012). Rhodanese functions as sulfur supplier for key enzymes in sulfur energy metabolism. J. Biol. Chem. 287, 19936–19948. doi: 10.1016/j.molp.2023.09.014
Azmi, A. F. M. (2019). Cyanogenic Glycosides in Cassava (The University of Leeds School of Food Science and Nutrition), 1–243. Available online at: https://etheses.whiterose.ac.uk/ (Accessed April 25, 2025).
Bailly, C. (2020). Anticancer properties of Prunus mume extracts (Chinese plum, Japanese apricot). J. Ethnopharmacol. 246, 112215. doi: 10.1016/j.jep.2019.112215
Bak, S., Paquette, S., Morant, M., Morant, A. V., Saito, S., Bjarnholt, N., et al. (2006). Cyanogenic glucosides: a case study for evolution and application of cytochromes P450. Phytochem. Rev. 5, 309–329. doi: 10.1007/s11101-006-9033-1
Ballhorn, D. J., Rakotoarivelo, F. P., and Kautz, S. (2016). Co-evolution of cyanogenic bamboos and bamboo lemurs on Madagascar. PloS One 11, e0158935. doi: 10.1371/journal.pone.0158935
Barakat, H., Aljutaily, T., Almujaydil, M. S., Algheshairy, R. M., Alhomaid, R. M., Almutairi, A. S., et al. (2022). Amygdalin: A review on its characteristics, antioxidant potential, gastrointestinal microbiota intervention, anticancer therapeutic and mechanisms, toxicity, and encapsulation. Biomolecul 12, 1514. doi: 10.3390/biom12101514
Bartnik, M. and Facey, P. C. (2017). “Glycosides,” in Pharmacognosy. Eds. McCreath, S. B. and Delgoda, R. (London-San Diego: Academic press), 101–161.
Baud, F. J., Borron, S. W., Megarbane, B., Trout, H., Lapostolle, F., Vicaut, E., et al. (2002). Value of lactic acidosis in the assessment of the severity of acute cyanide poisoning. Crit. Care Med. 30, 2044–2050. doi: 10.1097/00003246-200209000-00015
Beamer, W. C., Shealy, R. M., and Prough, D. S. (1983). Acute cyanide poisoning from laetrile ingestion. Ann. Emerg. Med. 12, 449–451. doi: 10.1016/s0196-0644(83)80345-x
Bhargava, A., Kumbhare, V., Srivastava, A., and Sahai, A. (1996). Bamboo parts and seeds for additional source nutrition. J. Food Sci. Technol. 32, 145–146.
Blaheta, R. A., Nelson, K., Haferkamp, A., and Juengel, E. (2016). Amygdalin, quackery or cure? Phytomedicine 23, 367–376. doi: 10.1016/j.phymed.2016.02.004
Bohm, B. A. and Glennie, C. W. (1971). A chemosystematic study of caprifoliaceae. Can. J. Bot. 49, 1799–1808. doi: 10.1139/b71-254
Bolarinwa, I. F., Olaniyan, S. A., Olatunde, S. J., Ayandokun, F. T., and Olaifa, I. A. (2016). Effect of processing on amygdalin and cyanide contents of some Nigerian Foods. J. Chem. Pharm. Res. 8, 106–113.
Bolarinwa, I. F., Orfila, C., and Morgan, M. R. A. (2014a). Natural Plant Toxins – Cyanogenic glycosides in plant foods (Saarbrücken, Germany: LAMBERT Academic Publishing).
Bolarinwa, I. F., Orfila, C., and Morgan, M. R. A. (2015). Determination of amygdalin in apple seeds, fresh apples, and processed apple juices. Food Chem. 170, 437–442. doi: 10.1016/j.foodchem.2014.08.083
Bolarinwa, I. F., Orila, C., and Morgan, M. R. A. (2014b). Amygdalin content of seeds kernels and food products commercially - available in the UK. Food Chem. 152, 133–139. doi: 10.1016/j.foodchem.2013.11.002
Boter, M. and Diaz, I. (2023). Cyanogenesis, a plant defence strategy against herbivores. Int. J. Mol. Sci. 24, 6982. doi: 10.3390/ijms24086982
Bovell-Benjamin, A. C. and Roberts, J. (2016). “Naturally occurring toxicants: presence in selected commonly consumed fruits,” in Regulating Safety of Traditional and Ethnic Foods. Eds. Prakash, V., Martín-Belloso, O., Keener, L., Astley, S., Braun, S., McMahon, H., and Lelieveld, H. (London, UK: Elsevier), 247–282.
Bruyn, G. W. and Poser, C. H. M. (2003). The history of tropical neurology (USA: Watson Publishing International).
Buhrmester, R. A., Ebinger, J. E., and Seigler, D. S. (2000). Sambunigrin and cyanogenic variability in populations of Sambucus canadensis L. (Caprifoliaceae). Biochem. Syst. Ecol. 28, 689–695. doi: 10.1016/S0305-1978(99)00105-2
Busk, P. K. and Møller, B. L. (2002). Dhurrin synthesis in sorghum is regulated at the transcriptional level and induced by nitrogen fertilization in older plant. Plant Physiol. 129, 1222–1231. doi: 10.1104/pp.000687
Butter, G. W. (1965). The distribution of the cyanoglycoside linamarin and lotaustralin in higher plants. Phytochem 4, 127–131.
Calderón, A. I., Terreaux, C., Gupta, M. P., Hostettmann, K., and Schenk, K. J. (2003). Taxiphyllin from henriettella fascicularis. Acta Crystallogr. C.: Struct. Chem. 34, 174–176. doi: 10.1002/chin.200330215
Chandler, R., Anderson, L., and Philipson, J. (1984). Laetrile in perspective. Can. Pharm. J. 117, 517–520.
Chaouali, N., Gana, I., Dorra, A., Khelifi, F., Nouioui, A., Masri, W., et al. (2013). Potential toxic levels of cyanide in almonds (Prunus amygdalus), apricot kernels (Prunus Armeniaca), and almond syrup. Int. Sch. Res. Notices 2013, 610648. doi: 10.1155/2013/610648
Chaturvedi, N., Sahrawat, N., Dua, S., and Banjola, P. (2023). Cyanogenic glycoside reduction of bamboo shoots species and their prospective as food victuals. Int. J. Sci. Res. Rev. 12, 42–52. doi: 10.37794/IJSRR.2023.12405
Chongtham, N., Bisht, M. S., Premlata, T., Bajwa, H. K., Sharma, V., and Santosh, O. (2022). Quality improvement of bamboo shoots by removal of antinutrients using different processing techniques: A review. J. Food Sci. Technol. 59, 1–11. doi: 10.1007/s13197-021-04987-9
Choudhury, D., Sahu, J. K., and Sharma, G. D. (2010). Biochemistry of bitterness in bamboo shoots. Assam Univ. J. Sci. Technol.: Phys. Sci. Technol. 6, 105–111.
Clarke, M., Harvey, D., and Humphreys, D. (1981). Veterinary Toxicology (London: ELBS and Bailliers, Tindal).
Conn, E. E. (1980). Cyanogenic compounds. Annu. Rev. Plant Physiol. 31, 433–451. doi: 10.1146/annurev.pp.31.060180.002245
Cressey, P. and Reeve, J. (2019). Metabolism of cyanogenic glycosides: A review. Food Chem. Toxicol. 125, 225–232. doi: 10.1016/j.fct.2019.01.002
Cressey, P., Saunders, D., and Auapaau, F. (2022). Evaluation of food safety risks associated with foods containing cyanogenic glycosides. New Zealand Food Safety. Institute of environmental science and research limited. ISBN 978-1-99-106234-5 (New Zealand: Christchurch).
Daun, J., Barthet, V., Chornick, T., Duguid, S., Thompson, L., and Cunnane, S. (2003). Structure, Composition, and Variety Development of Flaxseed (Champaign, Illinois, USA: AOCS Press).
Demirbolat, I. and Kartal, M. (2018). Prulaurasin content of leaves, kernels and pulps of Prunus lauracerasus L. (Cherry Laurel) during ripening. J. Res. Pharm. 22, 69–75. doi: 10.12991/jrp.2018.110
Dreyer, D. L., Reese, J. C., and Jones, K. C. (1981). Aphid feeding deterrents in sorghum: Bioassay isolation and characterization. J. Chem. Ecol. 7, 273–284. doi: 10.1007/BF00995750
Ďuračka, M., Tvrdá, E., Halenár, M., Zbynovská, K., Kolesár, E., Lukac, N., et al. (2016). The impact of amygdalin on the oxidative profile of rabbit testicular tissue. Mendel Net, 770–775.
Ellenhorn, M. J. and Barceloux, D. G. (1997). Ellenhorn’s medical toxicology: Diagnosis and treatment of human poisoning (New York: Elsevier Science Publishing Company).
Emendack, Y., Burke, J., Laza, H., Sanchez, J., and Hayes, C. (2018). Abiotic stress effects on sorghum leaf dhurrin and soluble sugar contents throughout plant development. Crop Sci. 58, 1706–1716. doi: 10.2135/cropsci2018.01.0059
Erdogan, M. F. (2003). Thiocyanate overload and thyroid disease. Biofactors 19, 107–111. doi: 10.1002/biof.5520190302
Ernesto, M., Cardoso, A. P., Nicala, D., Mirione, E., Massaza, F., Cliff, J., et al. (2002). Persistent konzo and cyanogen toxicity from cassava in northern Mozambique. Acta Trop. 82, 357–362. doi: 10.1016/s0001-706x(02)00042-6
Eyjólfsson, R. (1970). Isolation and structure determination of triglochinin, a new cyanogenic glucoside from Triglochin maritimum. Phytochem 9, 845–851. doi: 10.1016/S0031-9422(00)85191-2
Feeley, M., Agudo, A., Bronson, R., Edgar, J., Grant, D., and Hambridge, T. (2012). Contaminants: cyanogenic glycosides. WHO Food Additives Series: 65. Safety Evaluation of Certain Food Additives and Contaminants (Geneva, Switzerland: World Health Organization), 171–325.
Food and Agriculture Organization (2005). National Mission on Bamboo Applications. FAO Recommended Nutritional Composition, 3 Aug (Rome, Italy: Food and Agricultural Organization).
Forslund, K., Morant, M., Jørgensen, B., Olsen, C. E., Asamizu, E., Sato, S., et al. (2004). Biosynthesis of the nitrile glucosides rhodiocyanoside A and D and the cyanogenic glucosides lotaustralin and linamarin in Lotus japonicus. Plant Physiol. 135, 71–84. doi: 10.1104/pp.103.038059
Fukuda, T., Ito, H., Mukainaka, T., Tokuda, H., Nishino, H., and Yoshida, T. (2003). Anti-tumour promoting effect of glycosides from Prunus persica seeds. Biol. Pharm. Bull. 26, 271–273. doi: 10.1248/bpb.26.271
Ganjewala, D., Kumar, S., Asha, D. S., and Ambika, K. (2010). Advances in cyanogenic glycosides biosynthesis and analyses in plants: A review. Acta Biol. Szeged. 54, 1–14.
Gensa, U. (2019). Review on cyanide poisoning in ruminants. J. Biol. Agric. Healthc. 9, 1–12. doi: 10.7176/JBAH
Giantin, S., Franzin, A., Brusa, F., Montemurro, V., Bozzetta, E., Caprai, E., et al. (2024). Overview of cyanide poisoning in cattle from Sorghum halepense and S. bicolor cultivars in Northwest Italy. Animals 14, 743. doi: 10.3390/ani14050743
Gracia, R. and Shepherd, G. (2004). Cyanide poisoning and its treatment. Pharmacother 24, 1358–1365. doi: 10.1592/phco.24.14.1358.43149
Gryszczyńska, A., Łowicki, Z., Opala, B., Krajewska-Patan, A., Buchwald, W., Czerny, B., et al. (2013). Determination of lotaustralin in Rhodiola species. Herba Pol. 59, 13–23. doi: 10.2478/hepo-2013-0008
Halstrom, F. and Moiler, K. D. (1945). Content of cyanide in human organs from cases of poisoning with cyanide taken by mouth with contribution to toxicology cyanides. Acta Pharmacol. Toxicol. 1, 18–28. doi: 10.1111/J.1600-0773.1945.TB02560.X
Haque, M. R. and Bradbury, J. (2002). Total cyanide determination of plants and foods using the picrate and acid hydrolysis methods. Food Chem. 77, 107–114. doi: 10.1016/S0308-8146(01)00313-2
Hartanti, D. and Cahiani, A. N. (2020). Plant cyanogenic glycosides: an overview. Farmasains. J. Farm. Il. Kes. 5, 1–6. doi: 10.22219/farmasains.v5i1.10047
Harun, S. and Mohamed-Hussein, Z. A. (2024). “Glucosinolates and cyanogenic glycosides, biosynthesis and mechanism of action leading to plant defense,” in Plant Specialized Metabolites. Reference Series in Phytochemistry. Eds. Mérillon, J. M. and Ramawat, K. G. (Springer, Cham). doi: 10.1007/978-3-031-30037-0_53-1
He, X. Y., Wu, L. J., Wang, W. W., Xie, P. J., Chen, Y. H., and Wang, F. (2020). Amygdalin - A pharmacological and toxicological review. J. Ethnopharmacol. 254, 112717. doi: 10.1016/j.jep.2020.112717
Hodgson, E. (2012). Toxins and venoms. Prog. Mol. Biol. Transl. Sci. 112, 373–415. doi: 10.1016/B978-0-12-415813-9.00014-3
Hoikhokim, N. G. A. and Sarangthem, K. (2014). Effects of culm height and growth on the cyanogenic glycosides content of Thyrsostachys oliveri Gamble. Int. J. Curr. Res. Aca. Rev. 2, 105–113.
Hoikhokim, N. G. A. and Sarangthem, K. (2016). Cyanogenic glycosides in edible succulent bamboo shoots of Manipur, India. Int. J. Curr. Res. Aca. Rev. 4, 64–72. doi: 10.20546/ijcrar.2016.408.006
Hösel, W. (1981). “Glycosylation and glycosidases,” in The biochemistry of plants. Eds. Stumpf, P. K. and Conn, E. E. (Academic Press, New York), 14–17.
Howard, R. and Miller, N. (1984). Unproven methods of cancer management. Oncol. Nurs. Forum. 11, 67–73.
Howlett, W. P., Brubaker, G. R., Mlingi, N., and Rosling, H. (1990). Konzo, an epidemic upper motor neuron disease studied in Tanzania. Brain 113, 223–235. doi: 10.1146/annurev.pp.31.060180.002245
Hughes, M. A. (1991). The cyanogenic polymorphism in Trifolium repens L. (white clover). Heredity 66, 105–115. doi: 10.1038/hdy.1991.13
Idibie, C. A., Davids, H., and Iyuke, S. E. (2007). Cytotoxicity of purified cassava linamarin to a selected cancer cell lines. Bioproc. Biosyst. Eng. 30, 261–269. doi: 10.1007/s00449-007-0122-3
Igbadul, B. D., Amoye, J., and Twadue, I. (2014). Effect of fermentation on the proximate composition, antinutritional factors and functional properties of cocoyam (Colocasia esculenta) flour. Afr. J. Food Sci. Technol. 5, 67–74. doi: 10.14303/ajfst.2014.016
Jaszcak-Wilke, E., Polkowska, Ż., Koprowski, M., Owsianik, K., Mietchel, A. E., and Bałczewski, P. (2021). Amygdalin: toxicity, anticancer activity and analytical procedures for its determination in plant seeds. Molecules 26, 2253. doi: 10.3390/molecules26082253
Jones, D. A. (1998). Why are so many food plants cyanogenic? Phytochem 47, 155–162. doi: 10.1016/s0031-9422(97)00425-1
Kim, C. S., Jo, K., Lee, I. S., and Kim, J. (2016). Topical application of apricot kernel extract improves dry eye symptoms in a unilateral exorbital lacrimal gland excision mouse. Nutrients 8, 750. doi: 10.3390/nu8110750
Knudsen, B. F. and Kaack, K. V. (2015). A review of human health and disease claims for elderberry (Sambucus nigra) fruit. Acta Hortic. 1061, 121–131. doi: 10.17660/ActaHortic.2015.1061.12
Kojima, M., Poulton, J. E., Thayer, S. S., and Conn, E. E. (1979). Tissue distributions of dhurrin and enzymes involved in its metabolism in leaves of Sorghum bicolor. Plant Physiol. 63, 1022–1028. doi: 10.1104/pp.63.6.1022
Kolesárová, A., Baldovská, S., and Roychoudhury, S. (2021). The multiple actions of amygdalin on cellular processes with an emphasis on female reproduction. Pharmaceuticals 14, 881. doi: 10.3390/ph14090881
Krebs, E. T. (1970). The nitriloside (Vitamin B-17): Their nature, occurrence and metabolic significance. J. Appl. Nutr. 22, 75–86.
Kuete, V. (2014). “Health effects of alkaloids from african medicinal plants,” in Toxicological Survey of African Medicinal Plants (Elsevier, USA), 611–634.
Latif, S., Zimmermann, S., Barati, Z., and Müller, J. (2019). Detoxification of cassava leaves by thermal, sodium bicarbonate, enzymatic, and ultrasonic treatments. J. Food Sci. 84, 1986–1991. doi: 10.1111/1750-3841.14658
Lechtenberg, M. (2011). Cyanogenesis in Higher Plants and Animals (Chichester, UK: John Wiley & Sons).
Lechtenberg, M., Sendker, J., Kastner, L., and Hensel, A. (2022). Cyanogenesis in aralia spinosa (Araliaceae). Planta Med. 88, 1209–1222. doi: 10.1055/a-1671-5525
Lee, H. M. and Moon, A. (2016). Amygdalin regulates apoptosis and adhesion in Hs578T triple-negative breast cancer cells. Biomol. Ther. 24, 62–66. doi: 10.4062/biomolther.2015.172
Lee, J., Zhang, G., Wood, E., Castillo, C. R., and E Mitchell, A. (2013). Quantification of amygdalin in nonbitter, semibitter, and bitter almonds (Prunus dulcis) by UHPLC-(ESI)QqQ MS/MS. J. Agric. Food Chem. 61, 7754–7759. doi: 10.1021/jf402295u
Liangcheng, D., Mpoko, B., Birger, L. M., and Barbara, A. H. (1995). The biosynthensis of cyanogenic glucosides in roots of cassava. Phytochem 39, 323–326.
Liyanage, S. D., Gunasekera, D., and Ratnaweera, C. N. (2024). Harnessing the anti-cancer potential of linamarin: A computational study on design and hydrolysis mechanisms of its derivatives. J. Mol. Graph. Model. 128, 108716. doi: 10.1016/j.jmgm.2024.108716
Luo, H., Li, L., Tang, J., Zhang, F., Zhao, F., Sun, D., et al. (2016). Amygdalin inhibits HSC-T6 cell proliferation and fibrosis through the regulation of TGF-β/CTGF. Mol. Cell. Toxicol. 12, 265–271. doi: 10.1007/s13273-016-0031-0
Machingura, M., Salomon, E., Jez, J. M., and Ebb, S. D. (2016). The β-cyanoalanine synthase pathway: beyond cyanide detoxification. Plant Cell Environ. 39, 2329–2341. doi: 10.1111/pce.12755
Makarević, J., Rutz, J., Juengel, E., Kaulfuss, S., Reiter, M., Tsaur, I., et al. (2014). Amygdalin blocks bladder cancer cell growth in vitro by diminishing cyclin A and Cdk2. PloS One 9, e105590. doi: 10.1371/journal.pone.0105590
Mao, C. H. and Anderson, L. (1965). Cyanogenesis in Sorghum vulgare. II. Mechanism of the alkaline hydrolysis of dhurrin (p-hydroxymandelonitrile glucoside). J. Org. Chem. 30, 603–607. doi: 10.1021/jo01013a073
Menon, R., Munjal, N., and Sturino, J. M. (2015). Characterization of amygdalin-degrading Lactobacillus species. J. Appl. Microbiol. 118, 443–453. doi: 10.1111/jam.12704
Mfarrej, M. F. B. and Sharaf, N. S. (2011). Host Selection of peach rootborer Capnodis tenebrionis L. (Coleoptera: Buprestidae) to stone-fruit trees in Jordan. Jordan J. Agric. Sci. 7, 682–689.
Miller, R. E., Jensen, R., and Woodrow, I. E. (2006). Frequency of cyanogenesis in tropical rainforests of far north Queensland, Australia. Ann. Bot. 97, 1017–1044. doi: 10.1093/aob/mcl048
Mirande, M. (1912). Sur la presence de l’acide cyanohydrique dans letrefle rampant (Trifolium repens L.). Compt Rend. Acad. Sci. (Paris) 155, 651–653.
Mlingi, N. L., Bainbridge, Z. A., and Poulter, N. H. (1995). Critical stages in cyanogen removal during cassava processing in southern Tanzania. Food Chem. 53, 29–33. doi: 10.1016/0308-8146(95)95782-2
Młynarczyk, K., Walkowiak-Tomczak, D., and Łysiak, G. P. (2018). Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 40, 377–390. doi: 10.1016/j.jff.2017.11.025
Moertel, C., Fleming, T., Rubin, J., Kvols, L., Sarna, G., Koch, R., et al. (1982). A clinical trial of amygdalin (Laetrile) in the treatment of human cancer. N. Engl. J. Med. 206, 201–206. doi: 10.1056/NEJM198201283060403
Morant, A. V., Jørgensen, K., Jørgensen, B., Dam, W., Olsen, C. E., Møller, B. L., et al. (2007). Lessons learned from metabolic engineering of cyanogenic glucosides. Metabolomics 3, 383–398. doi: 10.1007/s11306-007-0079-x
Morimoto-Yamashita, Y., Tokuda, M., Ito, T., Kikuchi, K., Maruyama, I., Torii, M., et al. (2012). “Japanese apricot (ume): A novel therapeutic approach for the treatment of periodontitis,” in pathogenesis and treatment of periodontitis. Ed. Buduneli, N. (London, UK: IntechOpen), 145–156.
Mosayyebi, B., Imani, M., Mohammadi, L., Akbarzadeh, A., Zarghami, N., Edalati, M., et al. (2020). An update on the toxicity of cyanogenic glycosides bioactive compounds: Possible clinical application in targeted cancer therapy. Mater. Chem. Phys. 246, 122841. doi: 10.1016/j.matchemphys.2020.122841
Nahrstedt, A. (1975). Triglochinin in araceen. Phytochem 14, 2627–2628. doi: 10.1016/0031-9422(75)85237-X
Nassar, M. A. and Dorea, J. G. (1982). Protein in cassava cultivars and its hybrid with wild manihot species. Turrialba 32, 429–432.
Nyirenda, K. (2020). Toxicity potential of cyanogenic glycosides in edible plants. Med. Toxicol. 1–19. doi: 10.5772/intechopen.91408
Nzwalo, H. and Cliff, J. (2011). Konzo: from poverty, cassava, and cyanogen intake to toxico-nutritional neurological disease. PloS Negl. Trop. Dis. 5, e1051. doi: 10.1371/journal.pntd.0001051
O’Donnell, N. H., Møller, B. L., Neale, A. D., Hamill, J. D., Blomstedt, C. K., and Gleadow, R. M. (2013). Effects of PEG-induced osmotic stress on growth and dhurrin levels of forage sorghum. Plant Physiol. Biochem. 73, 83–92. doi: 10.1016/j.plaphy.2013.09.001
Odo, C. E., Enechi, O. C., and Oburu, C. S. (2014). Concentrations of anti - nutritional factors in raw edible cocoyam (Colocasia esculenta) leaves. J. Pharm. Res. 8, 38–40.
Olajide, R., Akinsoyinu, A. O., Babayemi, O. J., Omojola, A. B., Abu, A. O., and Afolabi, K. D. (2011). Effect of processing on energy values, nutrient and anti-nutrient components of wild cocoyam [Colocasia esculenta (L.) Schott] Corm. Pak. J. Nutr. 10, 29–34. doi: 10.3923/pjn.2011.29.34
Olsen, K. M., Hsu, S. C., and Small, L. L. (2008). Evidence on the molecular basis of the Ac/ac adaptive cyanogenesis polymorphism in white clover (Trifolium repens L). Genetics 179, 517–526. doi: 10.1534/genetics.107.080366
Olsen, K. M., Kooyers, N. J., and Small, L. L. (2013). Recurrent gene deletions and the evolution of adaptive cyanogenesis polymorphisms in white clover (Trifolium repens L.). Mol. Ecol. 22, 724–738. doi: 10.1111/j.1365-294X.2012.05667.x
Omelka, R., Kováčová, V., Mondočková, V., Grosskopf, B., Kolesárová, A., and Martiniaková, M. (2021). Cyanogenic glycoside amygdalin influences functions of human osteoblasts. vitro. J. Environ. Sci. Health B. 56, 109–116. doi: 10.1080/03601234.2020.1852054
Panter, K. E. (2018). “Cyanogenic glycoside - containing plants,” in Veterinary Toxicology: Basic and Clinical Principles. Ed. Gupta, R. C. (Elsevier Inc, Houston, TX), 935–940.
Park, H., Chung, H., Choi, S., Bahn, Y. S., and Son, J. (2024). Evaluation of exposure to cyanogenic glycosides and potential hydrogen cyanide release in commercially available foods among the Korean population. Food Chem. 456, 139872. doi: 10.1016/j.foodchem.2024.139872
Pascariu, O. E. and Israel-Roming, F. (2022). Bioactive compounds from elderberry: extraction, health benefits, and food applications. Processes 10, 2288. doi: 10.3390/pr10112288
Patel, H., Singh, R., Mody, S., Modi, C., and Kamani, S. (2014). Cyanide poisoning in animals. Department of pharmacology and toxicology college of veterinary science and animal husbandry. Int. e-J. 3, 202–216.
Pattarathitiwat, P., Chinvongamorn, C., and Sansenya, S. (2021). Evaluation of cyanide content, volatile compounds profile, and biological properties of fresh and boiled sliced thai bamboo shoot (Dendrocalamus asper Back.). Prev. Nutr. Food Sci. 26, 92–99. doi: 10.3746/pnf.2021.26.1.92
Pelley, J. W. (2012). “Structure and properties of biologic molecules,” in Elsevier’s Integrated Review Biochemistry. Ed. Saunders, W. B. (Philadelphia: Saunders, Saunders Elsevier), 7–18.
Pentzold, S., Zagrobelny, M., Roelsgaard, P. S., Møller, B. L., and Bak, S. (2014). The multiple strategies of an insect herbivore to overcome plant cyanogenic glucoside defence. PloS One 9, e91337. doi: 10.1371/journal.pone.0091337
Pičmanová, M., Neilson, E. H., Motawia, M. S., Olsen, C. E., Agerbirk, N., and Gray, C. J. (2015). A recycling pathway for cyanogenic glycosides evidenced by the comparative metabolic profiling in three cyanogenic plant species. Biochem. J. 469, 375–389. doi: 10.1042/BJ20150390
Popa, V. M., Raba, D. N., Moldovan, C., Poiana, M. A., Misca, C. D., Dumbrava, D. G., et al. (2021). A review of amygdalin characteristics. J. Agroaliment. Process. Technol. 27, 409–415.
Pulido, O. M. and Gill, S. (2013). “Food and toxicologic pathology: an overview,” in Haschek and Rousseaux’s Handbook of Toxicologic Pathology. Eds. Haschek-Hock, W. M., Rousseaux, C. C., Walling, M. A., Bolon, B., Ochoa, R., and Mahler, B. W. (Amsterdam: Academic press), 1051–1076.
Qadir, M. and Fatima, K. (2017). Review on pharmacological activity of amygdalin. Arch. Cancer Res. 5, 10–12. doi: 10.21767/2254-6081.100160
Rawat, K., Nirmala, C., and Bisht, M. S. (2015). Processing techniques for reduction of cyanogenic glycosides from bamboo shoots (Damyang, South Korea: World Bamboo Organization).
Rietjens, I. M. C. M. and Eisenbrand, G. (2022). “Natural toxicants in plant-based foods, including herbs and spices and herbal food supplements, and accompanying risks,” in Present Knowledge in Food Safety. Eds. Knowles, M. E., Anelich, L. E., Boobis, A. R., and Popping, B. (London, UK), 2–25.
Robakowski, P., Bielinis, E., Stachowiak, J., Mejza, I., and Bułaj, B. (2016). Seasonal changes affect root prunasin concentration in Prunus serotina and override species interactions between P. serotina and. Quercus Petraea. J. Chem. Ecol. 9, 202–214. doi: 10.1007/s10886-016-0678-y
Rosati, V. C., Blomstedt, C. K., Møller, B. L., Garnett, T., and Gleadow, R. (2019). The interplay between water limitation, dhurrin, and nitrate in the low-cyanogenic sorghum mutant adult cyanide deficient class 1. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01458
Rosen, G. and Shorr, R. (1979). Laetrile: end play around the FDA. A Rev. Legal Developments. Ann. Intern. Med. 90, 418–423. doi: 10.7326/0003-4819-90-3-418
Rosling, H. (1994). Measuring effect in humans of dietary cyanide exposure from cassava. Acta Hortic. 375, 271–283. doi: 10.17660/ACTAHORTIC.1994.375.27
Roulard, R., Fontaine, J. X., Jamali, A., Cailleu, D., Tavernier, R., Guillot, X., et al. (2017). Use of qNMR for speciation of flaxseeds (Linum usitatissimum) and quantification of cyanogenic glycosides. Anal. Bioanal. Chem. 409, 7011–7026. doi: 10.1007/s00216-017-0637-7
Russo, R. and Reggiani, R. (2014). Variation in the content of cyanogenic glycosides in flaxseed meal from twenty-one varieties. Food Nutr. Sci. 5, 1456–1462. doi: 10.4236/fns.2014.515159
Sadoff, L., Fuchs, J., and Hollander, J. (1978). Rapid death associated with laetrile ingestion. JAMA 15, 1532. doi: 10.1001/jama.1978.03280420068022
Sanchez-Perez, R., Jorgensen, K., Motawia, M. S., Dicenta, F., and Møller, B. L. (2009). Tissue and cellular localization of individual beta-glycosidases using a substrate specific sugar reducing assay. Plant J. 60, 894–906. doi: 10.1111/j.1365-313X.2009.03997.x
Sánchez-Pérez, R. and Neilson, E. H. J. (2024). The case for sporadic cyanogenic glycoside evolution in plants. Curr. Opin. Plant Biol. 81, 102608. doi: 10.1016/j.pbi.2024.102608
Sang-A-Gad, P., Guharat, S., and Wananukul, W. (2011). A mass cyanide poisoning from pickling bamboo shoots. Clin. Toxicol. 49, 834–839. doi: 10.3109/15563650.2011.618456
Santos Pimenta, L. P., Schilthuizen, M., Verpoorte, R., and Choi, Y. H. (2014). Quantitative analysis of amygdalin and prunasin in Prunus serotina Ehrh. using 1H-NMR spectroscopy. Phytochem. Anal. 25, 122–126. doi: 10.1002/pca.2476
Sarangthem, K. and Hoikhokim, K. S. (2010). Cyanogen content in bamboo plants. Asian J. Bio Sci. 5, 178–180.
Sarma, M. P. (2018). Analysis of cyanide concentration in five selected bamboo shoots consumed in North East India. Bioequiv. Bioavailab. Int. J. 2, 1–3. doi: 10.23880/BEBA16000127
Schmidt, F. B., Cho, S. K., Olsen, C. E., Yang, S. W., Møller, B. L., and Jørgensen, K. (2018). Diurnal regulation of cyanogenic glucoside biosynthesis and endogenous turnover in cassava. Plant Direct 2, 1–9. doi: 10.1002/pld3.38
Schrenk, D., BIgnami, M., Bodin, L., Chipman, J. K., Del Mazo, J., Grasl-Kraupp, B., et al. (2019). Evaluation of the health risks related to the presence of cyanogenic glycosides in foods other than raw apricot kernels. EFSA J. 17, e05662:1–78. doi: 10.2903/j.efsa.2019.5662
Senica, M., Stampar, F., and Mikulic-Petkovsek, M. (2019). Harmful (cyanogenic glycoside) and beneficial (phenolic) compounds in different Sambucus species. J. Berry Res. 9, 395–409. doi: 10.3233/JBR-180369
Senica, M., Stampar, F., Veberic, R., and Mikulic-Petkovsek, M. (2016). Processed elderberry (Sambucus nigra L.) products: A beneficial or harmful food alternative? LWT Food Sci. Technol. 72, 182–188. doi: 10.1016/j.lwt.2016.04.056
Shalayel, M. H. F., Al-Mazaideh, G. M., Alanezi, A. A., Almuqati, A. F., and Alotaibi, M. (2023). The potential anti-cancerous activity of Prunus amygdalus var. amara extract. Processes 11, 1277. doi: 10.3390/pr11041277
Shlichta, J. G., Glauser, G., and Benrey, B. (2014). Variation in cyanogenic glycosides across populations of wild lima beans (Phaseolus lunatus) has no apparent effect on bruchid beetle performance. J. Chem. Ecol. 40, 468–475. doi: 10.1007/s10886-014-0434-0
Siegień, I. and Bogatek, R. (2006). Cyanide action in plants–from toxic to regulatory. Acta Physiol. Plant 28, 483–497. doi: 10.1007/BF02706632
Smith, J. R., C., R., Weisleder, D., Miller, R. W., Palmer, I. S., and Olson, O. E. J. (1980). Linustatin and neolinustatin: cyanogenic glycosides of linseed meal that protect animals against selenium toxicity. J. Org. Chem. 45, 3, 507–3, 510. doi: 10.1021/jo01291a027
Song, Z. and Xu, X. (2014). Advanced research on anti-tumor effects of amygdalin. J. Cancer Res. Ther. 10, 3–7. doi: 10.4103/0973-1482.139743
Süli, J., Sobeková, A., and Bujdošová, Z. (2017). Anti-nutritional substances in legumes. Chem. Sheets 111, 789–797.
Sun, Z., Zhang, K., Chen, C., Wu, Y., Tang, Y., Georgiev, M. I., et al. (2018). Biosynthesis and regulation of cyanogenic glycoside production in forage plants. Appl. Microbiol. Biotechnol. 102, 9–16. doi: 10.1007/s00253-017-8559-z
Tahir, F., Ali, E., Hassan, S. A., Bhat, Z. F., Walayat, N., Nawaz, A., et al. (2024). Cyanogenic glucosides in plant-based foods: Occurrence, detection methods, and detoxification strategies – A comprehensive review. Microchem. J. 199, 110065. doi: 10.1016/j.microc.2024.110065
Tanaka, T., Kimura, K., Kan, K., Katori, Y., Michishita, K., Nakano, H., et al. (2020). Quantification of amygdalin, prunasin, total cyanide and free cyanide in powdered loquat seeds. Food Addit. Contam. Part A 37, 1503–1509. doi: 10.1080/19440049.2020.1778186
Thodberg, S., Sørensen, M., Bellucci, M., Crocoll, C., Bendtsen, A. K., Nelson, D. R., et al. (2020). A flavin-dependent monooxygenase catalyzes the initial step in cyanogenic glycoside synthesis in ferns. Commun. Biol. 3, 507. doi: 10.1038/s42003-020-01224-5
Tokpohozin, S. E., Fischer, S., Sacher, B., and Becker, T. (2016). β-D-Glucosidase as “key enzyme” for sorghum cyanogenic glucoside (dhurrin) removal and beer bio-flavouring. Food Chem. Toxicol. 97, 217–223. doi: 10.1016/j.fct.2016.09.015
Touré, A. and Xueming, X. (2010). Flaxseed lignans: Source, biosynthesis, metabolism, antioxidant activity, bio-active components, and health benefits. Compr. Rev. Food Sci. Food Saf. 9, 261–269. doi: 10.1111/j.1541-4337.2009.00105.x
Vetter, J. (2000). Plant cyanogenic glycosides. Toxicon 38, 11–36. doi: 10.1016/s0041-0101(99)00128-2
Vetter, J. (2017). “Plant cyanogenic glycosides,” in Plant Toxins. Eds. Gopalakrishnakone, P., Carlini, C. R., and Ligabue-Braun, R. (Springer Science+ Business Media, Berlin, Germany), 288–315.
Wang, Q. and Ruan, X. (2005). Quantitative analysis of salidroside, tyrosol and lotaustralin in roots of Rhodiola L. plants grown at Tianshan Mountain. Chin. J. Pharm. Anal. 25, 1026–1029. doi: 10.1038/srep08337
Waszkowiak, K., Gliszczynska-Swiglo, A., Barthet, V., and Skrety, J. (2015). Effect of extraction method on the phenolic and cyanogenic glucoside profile of Flaxseed extracts and their antioxidant capacity. J. Am. Oil Chem. Soc 92, 1609–1619. doi: 10.1007/s11746-015-2729-x
Wheeler, J., Mulcahy, C., Walcott, J., and Rapp, G. (1990). Factors affecting the hydrogen cyanide potential of forage sorghum. Crop Pasture Sci. 41, 1093–1100. doi: 10.1071/AR9901093
Woodrow, I. E., Slocum, D. J., and Gleadow, R. M. (2002). Influence of water stress on cyanogenic capacity in Eucalyptus cladocalyx. Funct. Plant Biol. 29, 103–110. doi: 10.1071/PP01116
World Health Organisation (2012). Cyanogenic glycosides (Who Food Additives Series 30). Available online at: http://www.inchem.org.
Xu, J., Wang, X. Y., and Guo, W. Z. (2015). The cytochrome P450 superfamily: Key players in plant development and defense. J. Integr. Agric. 14, 1673–1686. doi: 10.1016/S2095-3119(14)60980-1
Yadav, M., Singh, I. K., and Singh, A. (2023). Dhurrin: A naturally occurring phytochemical as a weapon against insect herbivores Archana Singh. Phytochem 205, 113483. doi: 10.1016/j.phytochem.2022.113483
Yildirim, A. N., Akinci-Yildirim, F., Polat, M., and Sxan, B. (2014). Amygdalin content in kernels of several almond cultivars grown in Turkey. Hort. Sci. 49, 1268–1270. doi: 10.21273/HORTSCI.49.10.1268
Yildirim, F. A. and Askin, M. A. (2010). Variability of amygdalin content in seeds of sweet and bitter apricot cultivars in Turkey. Afr. J. Biotechnol. 9, 6522–6524. doi: 10.5897/AJB10.884
Yulvianti, M. and Zidorn, C. (2021). Chemical diversity of plant cyanogenic glycosides: An overview of reported natural products. Molecules 26, 719. doi: 10.3390/molecules26030719
Yusuf, U., Fakhru’l-Razi, A., Rosli, R., Iyuke, S., Billa, N., Nurhafizah, A., et al. (2006). An in vitro inhibition of human Malignant cell growth of crude water extract of cassava (Manihot esculenta Crantz) and commercial linamarin. Songklanakarin J. Sci. Technol. 28, 145–155.
Zagrobelny, M., De Castro, E. C. P., Moller, B. L., and Bak, S. (2018). Cyanogenesis in arthropods: from chemical warfare to nuptial gifts. Insects 9, 51. doi: 10.3390/insects9020051
Zarogoulidis, P., Darwiche, K., Sakkas, A., Yarmus, L., Haidong, H., Li, Q., et al. (2013). Suicide gene therapy for cancer - current strategies. J. Genet. Syndr. Gene Ther. 4, 16849. doi: 10.4172/2157-7412.1000139
Zhao, M., Bergaentzlé, M., Flieller, A., and Marchioni, E. (2019). Development and validation of an ultra-high performance liquid chromatography-high resolution mass spectrometry method for simultaneous quantification of cyanogenic glycosides and secoisolariciresinol diglucoside in flaxseed (Linum usitatissimum L.). J. Chromatogr. A. 1601, 214–223. doi: 10.1016/j.chroma.2019.04.072
Zhao, Y., Wen, S., Wang, Y., Zhang, W., Xu, X., and Mou, Y. (2024). A Review of recent advances in chromatographic quantification methods for cyanogenic glycosides. Molecules 29, 4801. doi: 10.3390/molecules29204801
Zhong, Y., Xu, T., Chen, Q., Li, K., Zhang, Z., Song, H., et al. (2020). Development and validation of eight cyanogenic glucosides via ultra-high-performance liquid chromatography-tandem mass spectrometry in agri-food. Food Chem. 331, 127305. doi: 10.1016/j.foodchem.2020.127305
Zhou, J., Hou, J., Rao, J., Zhou, C., Liu, Y., and Gao, W. (2020). Magnetically directed enzyme/prodrug prostate cancer therapy based on β-Glucosidase/Amygdalin. Int. J. Nanomed. 15, 4639–4657. doi: 10.2147/IJN.S242359
Keywords: cyanogenic glycosides, structure and biosynthesis, biological function, content and distribution, toxicity, anticancer potential
Citation: Piršelová B and Jakubčinová J (2025) Plant cyanogenic glycosides: from structure to properties and potential applications. Front. Plant Sci. 16:1612132. doi: 10.3389/fpls.2025.1612132
Received: 15 April 2025; Accepted: 26 June 2025;
Published: 31 July 2025.
Edited by:
Ping Su, China Academy of Chinese Medical Sciences, ChinaReviewed by:
Maria Valeria Lara, Universidad Nacional de Rosario, ArgentinaLourin Malak, Assiut University, Egypt
Copyright © 2025 Piršelová and Jakubčinová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Beáta Piršelová, YnBpcnNlbG92YUB1a2Yuc2s=