 Vaida Sirgedaitė-Šėžienė1
Vaida Sirgedaitė-Šėžienė1 Greta Striganavičiūtė1
Greta Striganavičiūtė1 Milana Šilanskienė1
Milana Šilanskienė1 Inesa Kniuipytė2
Inesa Kniuipytė2 Marius Praspaliauskas2
Marius Praspaliauskas2 Irena Vaškevičienė2Egidijus Lemanas2
Irena Vaškevičienė2Egidijus Lemanas2 Dorotėja Vaitiekūnaitė1*
Dorotėja Vaitiekūnaitė1*- 1Laboratory of Forest Plant Biotechnology, Institute of Forestry, Lithuanian Research Centre for Agriculture and Forestry, Girionys, Lithuania
- 2Laboratory of Heat–Equipment Research and Testing, Lithuanian Energy Institute, Kaunas, Lithuania
This study investigates the phytoremediation potential of Populus tremula L. and Salix caprea L. in response to As, Cd, and Pb exposure using hydroponics. Seedlings were exposed to 5–50 µM Cd, 100–1000 µM As, and 50–200 µM Pb in P. tremula, and to 5–50 µM Cd, 25–100 µM As, and 200–600 µM Pb in S. caprea. By analyzing growth, heavy metal(loid) (hereafter referred to as ‘metals’) uptake, biochemical markers (phenolics, soluble sugars, lipid peroxidation, antioxidant enzymes), and shifts in elemental (P, S, Mg, K, Ca, Mn, Zn, Cu, Fe) composition, this study provides a comprehensive evaluation of these species response to metal contamination at the seedling stage. Distinct dose- and metal-specific responses were observed, with Pb exposure inducing enhancing growth effects (height increase up to 27%, total chlorophyll increase up to 67%) and hormesis at low to moderate concentrations (equivalent to 200 µM of Pb(NO3)2), while Cd and As reduced growth in both species. Biochemical analyses revealed significant impacts on the antioxidant activity in response to metal stress, with differences in the involvement of enzymatic vs. non-enzymatic defenses, i.e., an initial enzymatic response, and a shift towards secondary metabolite production under prolonged or severe stress. S. caprea exhibited higher translocation of Cd (0.77 at 5 µM), suggesting its potential for phytoextraction, while both species demonstrated strong phytostabilization capacity for Pb (up to 0.54% of Pb in root DW). Nutrient homeostasis disruptions were observed, with both species showing altered nutrient uptake and distribution, e.g., co-accumulation of Cd and Zn, with Zn increase up to 639% in Cd-treated S. caprea (50 µM). These results offer valuable insights into the biochemical mechanisms underlying heavy metal tolerance in P. tremula and S. caprea, while suggesting directions for future studies on the real-world applicability of phytoremediation strategies.
1 Introduction
Heavy metal(loid)s (HMs) (hereafter referred to as ‘metals’) such as cadmium (Cd), arsenic (As), and lead (Pb) are persistent environmental contaminants that cannot be metabolically degraded, and thus can accumulate in ecosystems, leading to detrimental effects on plants, animals, and ultimately human health (De Vries et al., 2022; Sharma et al., 2023). In Europe, efforts to reduce HM emissions have been implemented, resulting in notable decreases between 2005 and 2022: Pb emissions decreased by 44%, and Cd by 39% across the EU-27 Member States (De Vries et al., 2022). However, despite these reductions, HMs continue to accumulate in ecosystems, posing ongoing risks. As an example, the spatial distribution of Cd in the topsoil across the EU and the United Kingdom was analyzed using 21,682 soil samples from the LUCAS soil module, collected in 2009. Among these samples, 5.5% contained Cd concentrations exceeding 1 mg/kg (Ballabio et al., 2024) while a total of at least 10% (depending on the imposed limits, 19 or even 36%) of all tested agricultural soil samples exceeded the limits for the tested HMs (As and Pb included) (Yunta et al., 2024). Based on these results, the EU soil strategy states that by 2050, soil pollution should be reduced to levels no longer considered harmful to human health and the environment (European Commission (EC), 2021).
Phytoremediation has emerged as a sustainable and ecologically sound approach to mitigate HM contamination long term. This green technology utilizes the natural capabilities of plants to remove, stabilize, degrade, or sequester pollutants from contaminated soils. Compared to traditional physical and chemical methods, phytoremediation is generally more cost-effective, particularly in large-scale applications, as it requires minimal input once the plants are established. It’s also less environmentally disruptive, in fact, it can enhance the ecological balance by promoting plant growth and biodiversity (Kumar et al., 2022; Sharma et al., 2023).
Among the different phytoremediation strategies, phytoextraction - metal uptake and translocation to harvestable shoots - and phytostabilization - metal retention in roots or the rhizosphere - are particularly relevant to woody species. The efficiency of either mechanism depends on plant traits, pollutant type, and environmental conditions (Kumar et al., 2022), and directly informs the selection of species for either metal removal or containment. Despite the promise of phytoremediation, several challenges impede its widespread application. The process can be slow, often requiring multiple growing seasons to achieve meaningful pollutant removal. Additionally, the disposal of contaminated plant biomass must be managed carefully to prevent secondary contamination. The efficiency of phytoremediation is influenced by site-specific factors, such as soil and site properties, pH, contaminant type, moisture levels, and temperature, which vary widely across locations (Fendrich et al., 2024; Yunta et al., 2024). Moreover, it has been shown that the capacity for HM accumulation varies greatly among species and even among cultivars within the same species (Kumar et al., 2022; Sharma et al., 2023), hence selecting appropriate plant species with high contaminant uptake and tolerance is critical, yet only a limited number of species possess these capabilities.
A notable research gap exists in the identification and utilization of plant species for phytoremediation purposes. In September 2017, a global database cataloged 754 hyperaccumulator plant species, including 5 As hyperaccumulators, 7 for Cd, and 8 for Pb, among others (Reeves et al., 2017). Hyperaccumulators are plants capable of absorbing and accumulating exceptionally high concentrations of HMs or other contaminants in their tissues without suffering toxic effects. The majority of hyperaccumulator research has focused on herbaceous plants (Rascio and Navari-Izzo, 2011; Kumar et al., 2022), with limited studies on trees or other woody species. Trees, particularly fast-growing species, offer advantages such as extensive root systems and substantial biomass production, which could enhance phytoremediation efficiency. Few tree species have been studied for their phytoremediation potential, and even fewer have been identified as hyperaccumulators (Rascio and Navari-Izzo, 2011; Reeves et al., 2017). Nonetheless, several studies have already demonstrated the potential of Salix and Populus species for heavy-metal remediation (Konlechner et al., 2013; Salam et al., 2022). However, most of them focused on cuttings or field studies with a notable gap in researching early seedling establishment and response to contaminated environment.
To address this gap, the present study aims to evaluate the effectiveness of two fast-growing tree species, Populus tremula L. (European aspen) and Salix caprea L. (goat willow), in remediating Cd, Pb, and As contaminated substrates at an early developmental stage. This research provides a novel contribution by systematically comparing the responses of these two ecologically important Salicaceae species to three distinct HMs (Cd, Pb, As) during the critical early seedling stage, when foundational tolerance mechanisms are established and phytoremediation potential can be assessed. These species were selected based on their widespread distribution across temperate Europe, rapid biomass production, tolerance to poor soil conditions, and prior use in land reclamation. Additionally, the study seeks to determine whether either species functions as a hyperaccumulator of these metals, thereby contributing to the development of more efficient and sustainable phytoremediation strategies. While previous studies have typically focused on mature plants or cuttings, our comprehensive approach enables direct comparison of early-stage metal-specific responses across both species under standardized conditions. Based on the known physiological characteristics of Salicaceae species and preliminary studies on related taxa, we hypothesize that: (I) both species will demonstrate greater tolerance to Pb than to Cd or As, (II) S. caprea will exhibit higher translocation factors than P. tremula due to its documented metal accumulation capabilities, and (III) successful metal tolerance will be accompanied by upregulated antioxidant systems and altered nutrient homeostasis. Our systematic comparative approach using controlled hydroponic conditions will reveal metal-specific bioaccumulation patterns that inform optimal phytoremediation applications for each species and provide essential baseline data for early-intervention phytoremediation strategies.
2 Materials and methods
2.1 Seeds and initial growth environment
Populus tremula L. and Salix caprea L. seeds were collected in the spring of 2024 and stored at +4°C until the start of the experiment (2–3 months). The seeds were sprouted in the lab (+22°C) in glass Petri dishes on moist (tap water) filter paper and when roots emerged the seedlings were transferred to moist rockwool cubes (soaked in distilled water, pH 5.6, adjusted using HCl and NaOH) and grown in modified full-strength (100%) Hoagland’s nutrient medium, containing 6 mM KNO3, 2.32 mM Ca(NO3)2×4H2O, 1.86 mM MgSO4×7H2O, 1 mM NH4H2PO4, 46 μM H3BO3, 9 μM MnCl2×4H2O, 8.99 μM C12H12Fe2O18, 0.76 μM ZnSO4×7H2O, 0.5 μM CuSO4×5H2O, and 0.58 μM Na2MoO4×2H2O. The seedlings were incubated under semi-controlled conditions (day/night temperature of 25/20°C, natural light) for 5 weeks until the BBCH-14 growth stage (4 true leaves) was reached (Guilayn et al., 2020). The seedlings were watered as needed with half-strength (50%) Hoagland’s nutrient medium.
2.2 Hydroponic conditions and experimental groups
After the BBCH-14 growth stage was reached, the seedlings were transferred to the hydroponic system into 12 L non-see-through plastic containers (42×35×11 cm). The container covers had holes with plastic aquatic pots. Each container was filled with 8 L of full-strength Hoagland’s medium and the rockwool cubes were placed into the aquatic pots with the lower half fully submerged. For both P. tremula and S. caprea 10 experimental groups were studied: control group and 9 treatment groups. The medium in 9 containers was contaminated with HMs – As (AsH15Na2O11, 98% purity; Thermo Fisher, Germany), Pb (Pb(NO3)2, 99% purity; VWR International GmbH, Austria) and Cd (CdSO4, 99% purity; Thermo Fisher) right before transferring the seedlings. For P. tremula the 9 groups were 3 concentrations of As (100, 500 and 1000 µM), 3 concentrations of Pb (50, 100 and 200 µM) and 3 concentrations of Cd (5, 25, 50 µM). For S. caprea the 9 groups were treated with 3 concentrations of As (25, 50, 100 µM), 3 concentrations of Pb (200, 400 and 600 µM) and 3 concentrations of Cd (5, 25, 50 µM). Continuous aeration was provided by air pumps (14020; 220 V, 50 Hz, 5 W, China) connected to plastic tracheas, and an oxygen diffuser (18 cm) secured to the bottom of the container. The seedlings were grown under semi-controlled conditions (day/night temperature of 25/20°C, natural light) for 4 weeks in a greenhouse. During this time the seedlings were watered as needed with full-strength Hoagland’s medium.
2.3 Growth measurements and sampling
Immediately after transferring to the hydroponic conditions, seedling height was measured. This measurement was repeated after 4 weeks to determine height increase over the period. The longest root length was only measured at the end of the experiment, as root systems were embedded in rockwool cubes and could not be measured accurately without disrupting growth. Tolerance index (TI) was calculated based on modified formula from shoot height and shows a relative measure of a plant’s ability to withstand and grow in the presence of contaminants (Yongpisanphop et al., 2017). The aboveground and root biomass of the seedlings were collected separately for subsequent biochemical and elemental composition analyses. For the biochemical analyses, 6 fresh samples of 0.1 g were taken from the leaf and 6 from the root tissues from each experimental group. Each sample was comprised of smaller samples taken from the biomass of the whole group. The samples were frozen for subsequent analyses. For the elemental composition analyses, the root and shoot biomass was completely dried at 60°C in a forced-air ventilation oven (Binder FD 115, Germany) to a constant dry weight (DW) and weighed again (Mettler Toledo XP2003S DR, Switzerland). Three samples of ~0.2-0.4 g were taken from each experimental group.
2.4 Biochemical analyses
Seedling total chlorophyll, carotenoid (CAR), secondary metabolite (SM): total phenol (TPC) and total flavonoid (TFC), lipid peroxidation level, i.e., malondialdehyde (MDA), soluble sugar, (SS), antioxidant enzyme (catalase (CAT), superoxide dismutase (SOD), guaiacol peroxidase (POX), ascorbate peroxidase (APX), glutathione reductase (GR), glutathione S-transferase (GST)) levels were measured spectrophotometrically using a SpectroStar Nano microplate reader (BMG Labtech, Offenburg, Germany) with 96-well microplates. A detailed list of the formulas for biochemical analyses done are listed in the relevant references and Supplementary Table 1.
2.4.1 Extract preparation for photosynthetic pigment, secondary metabolite, MDA, and sugar analysis
For each experimental group, three 0.1 g samples were pulverized using a tissue homogenizer (Precellys 24, Bertin Technologies, France) at 1956×g for 30 seconds with the addition of two metal beads. Next, 1.5 mL of 80% ethanol (v/v in water, MV GROUP Production, Lithuania) was added, and the homogenization process was repeated under the same conditions. The resulting mixture was then centrifuged at 16,090×g for 30 minutes at 4°C using a Hettich Universal 32R centrifuge (Andreas Hettich GmbH & Co. KG, Germany). The supernatant was collected and used for the analysis of total chlorophyll, CAR, TPC, TFC, MDA, and SS.
2.4.2 Total phenol content
In brief, the extract was mixed with Folin–Ciocalteu reagent (VWR International GmbH) (1:9 v/v in water) and incubated for 5 minutes. Subsequently, 10% Na2CO3 was added, and the mixture was kept in the dark for 1 hour. Absorbance was then measured at 725 nm. The formulas and calibration for the analysis were described in detail previously (Beniušytė et al., 2023).
2.4.3 Total flavonoid content
Briefly, for TFC the extract was combined with a reaction buffer containing absolute ethyl alcohol (MERCK, Germany), aluminum chloride (Alfa Aesar, Germany), potassium acetate (Sigma Aldrich, USA), and distilled water (dH2O). The mixture was incubated in the dark for 30 minutes. Absorbance measurements were taken at 415 nm. The formulas and calibration for the analysis were as described in detail previously (Beniušytė et al., 2023).
2.4.4 Chlorophyll a and b, and carotenoid content
Absorbance of the extract was measured at 470, 648, and 664 nm. The formulas and calibration for the analyses were as described in detail previously (Beniušytė et al., 2023). Total chlorophyll content was calculated by adding chlorophyll a and b content.
2.4.5 Malondialdehyde content
In brief, MDA was quantified using a modified method. The supernatant was mixed with a reaction mixture containing trichloroacetic acid (Molar Chemicals Kft, Hungary) and thiobarbituric acid (Alfa Aesar). The mixture was incubated at 95°C for 30 minutes and then cooled on ice. Absorbance was recorded at 440, 532, and 600 nm. The formulas and further details for the analysis were described previously (Čėsnienė et al., 2023).
2.4.6 Soluble sugar content
Soluble sugars were quantified by mixing the sample with anthrone reagent (Carl Roth, Germany), prepared by dissolving anthrone in concentrated H2SO4 (Chempur, Poland) according to Čėsnienė et al (Čėsnienė et al., 2023). The mixture was incubated at 90°C for 1 hour, and absorbance was measured at 620 nm.
2.4.7 Potassium phosphate buffer preparation
K-phosphate buffers of varying pH values were prepared by mixing stock solutions of 1 M K2HPO4 (Carl Roth) and 1 M KH2PO4 (Chempur). Buffers with pH values of 6.5, 7.0, 7.4 and 7.8 were prepared as required.
2.4.8 Extract preparation for enzyme analysis
For enzyme activity assays, three 0.1 g samples of fresh biomass from each experimental group were pulverized in liquid nitrogen using a mortar and pestle and then mixed with an extraction buffer containing K-phosphate buffer (pH 7.8), Triton X (Carl Roth), polyvinylpolypyrrolidone (PVPP) (Sigma Aldrich), and ascorbic acid (ASC) (Chempur). The mixture was centrifuged at 14,020×g at −4°C for 1 hour, and the resulting supernatant was used to measure protein levels, as well as CAT and SOD activity. For APX, POX, GR, and GST assays, the supernatant was further purified using Sephadex G-25 columns (Column PD-10, Cytiva, UK), and the bulk filtrate was used for the latter analyses. All procedures were carried out on ice to preserve sample integrity. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.4.9 Total protein quantification
Protein concentration was determined by mixing the crude extract with Biuret reagent, composed of CuSO4, Na-K tartrate (Eurochemicals, Lithuania), and Na2CO3 in NaOH solution, followed by the addition of Folin–Ciocalteu reagent. After incubating for 50 minutes at room temperature, absorbance was measured at 660 nm. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.4.10 Catalase activity
Catalase activity was assessed by mixing the crude extract with K-phosphate buffer (pH 7) and 30% H2O2. Absorbance was measured at 240 nm at regular intervals to monitor the activity. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.4.11 Superoxide dismutase activity
SOD activity was assessed by mixing the crude extract with a reaction buffer containing K-phosphate buffer (pH 7.8), methionine (AppliChem, Germany), nitro blue tetrazolium (NBT) (VWR International GmbH), ethylenediaminetetraacetic acid (EDTA) (Chempur), and riboflavin (AppliChem). The reaction mixture was exposed to white light (irradiance 30 μmol m-² s-¹) until the samples darkened relative to the control. Absorbance was then measured at 550 nm to determine SOD activity. The formulas and further details for the analysis were described previously (Čėsnienė et al., 2023).
2.4.12 Guaiacol peroxidase activity
POX activity was evaluated by mixing the filtered extract with a reaction solution containing pyrogallol (Chempur), a 50 mM K-phosphate buffer (pH 6.5), and 30% H2O2. Absorbance at 430 nm was measured at regular intervals to monitor the activity. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.4.13 Ascorbate peroxidase activity
APX activity was measured by mixing the filtered extract with ascorbic acid (ASC) solution, 50 mM K-phosphate buffer (pH 7.0), and 30% H2O2. Absorbance at 290 nm was recorded at regular intervals to monitor the activity. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.4.14 Glutathione S-transferase activity
GST activity was measured by combining the filtered extract with a reaction buffer containing K-phosphate buffer (pH 6.5), 1-chloro-2,4-dinitrobenzene (CDNB) (Thermo Fisher), and reduced L-glutathione (GSH) solution (Sigma Aldrich). Absorbance at 340 nm was recorded at regular intervals to monitor the activity. The formulas and further details for the analysis were described previously (Sirgedaitė−Šėžienė et al., 2024).
2.4.15 Glutathione reductase activity
To measure GR enzyme activity, the filtered extract was combined with a reaction buffer consisting of HEPES buffer (pH 8) (Sigma Aldrich), EDTA, and NADPH (Carl Roth). After adding oxidized L-glutathione (Applichem) (GSSG), absorbance changes at 340 nm were monitored at regular intervals. The formulas and further details for the analysis were described previously (Beniušytė et al., 2023).
2.5 Elemental composition analyses
The previously dried samples were pulverized using an IKA MF 10 mill (IKA®-Werke GmbH & Co. KG, Germany). The pulverized samples were then digested with a mixture of 65% HNO3, 30% H2O2, and 48% HF in a volume ratio of 8:3:1. The digestion process was performed using a high-pressure microwave digester (Anton Paar, Multiwave GO Plus, Austria). The concentrations of Cd, As, Pb, P, S, Mg, K, Ca, Mn, Zn, Cu, and Fe in the digested shoot and root samples were determined using inductively coupled plasma optical emission spectrometry (ICP-OES, Optima 8000, PerkinElmer, USA). To ensure the accuracy of the analytical methods certified reference material (CRM) was used, specifically CRM-BCR-129 hay powder, approved by the Community Bureau of Reference (BCR). To monitor ICP-OES signal drift, a control solution (5 mg/L) was analyzed every 20 samples. Calibration was repeated whenever the measured value deviated by more than 10% from the predetermined boundaries. Each sample was analyzed in triplicate to ensure accuracy and reproducibility. Element calibration was conducted at four calibration points using PerkinElmer standard solutions: for Cd, As, Pb, Mg, Ca, Mn, Zn, and Fe, the Multi-Element Quality Control Standard 21 was used; for K, the Quality Control Standard 7A was used; and for P and S, Pure Standards were used. The accuracy of elemental analysis was evaluated using a linear correlation coefficient of at least 0.999, ensuring high precision in the results (Kniuipytė et al., 2023).
Additionally, in response to HM exposure, several indices were calculated following the formulas provided by Yongpisanphop et al (Yongpisanphop et al., 2017): bioconcentration factor (BCF) for tree leaves and roots showing how much HM was found in these tissues as compared to the nutrient medium and translocation factor (TF) showing how much of the HMs was transferred from roots to leaves. A detailed list of the formulas for the indices used are listed in the relevant references and Supplementary Table 1.
2.6 Statistical analysis
The data were compiled, and graphs were generated using Microsoft Office Excel. Statistical analyses were conducted with SPSS version 28.0.1.1 (IBM Inc.). Since the biochemical data did not meet normality or homogeneity assumptions, even after transformation, the Kruskal–Wallis H test was used for independent samples. Pairwise rank comparisons were performed using Dunn’s post hoc test (p<0.05). A 95% confidence level was maintained, and the results of Dunn’s test were reported with Bonferroni-adjusted p-values to identify specific groups with significant differences. The elemental composition and metal uptake data were expressed as a mean from 3 pooled samples due to constraints in the amount of biomass the plants produced. And thus lacks a robust statistical analysis.
3 Results
3.1 HM impact on Populus tremula
Out of the nine treatment groups for P. tremula, only seven survived until the end of the experiment. Exposure to 500 and 1000 µM of As resulted in complete shoot growth inhibition and severe chlorosis (Figure 1), preventing further analyses for these two groups due to lack of biomass. To assess the seedlings’ ability to tolerate heavy metals (HMs) in the nutrient medium, several indices were calculated for the remaining groups. In all Pb-treated groups, the TI (tolerance index) exceeded 100%, ranging from 113% to 127% (Table 1, Figure 2), indicating enhanced growth under contamination. The TF (translocation factor) in these groups ranged from 0.01 to 0.06 (Table 1). Under Pb exposure, the BCF (bioconcentration factor) for P. tremula shoots remained below 1 (0.42–0.84), while in the roots, it ranged from 15.92 to 66.22 (Table 1). The precise HM concentrations in leaves and roots for all experimental groups are presented in Table 1. Notably, all HM levels increased with rising concentrations in both tissues, with roots consistently exhibiting higher concentrations of the target HMs. Cd levels in leaves ranged from 59 to 538 µg/g DW, while root concentrations varied between 176 and 1713 µg/g DW. In Pb-treated Populus seedlings, leaf concentrations ranged from 9 to 17 µg/g DW, whereas root levels were between 165 and 2746 µg/g DW.
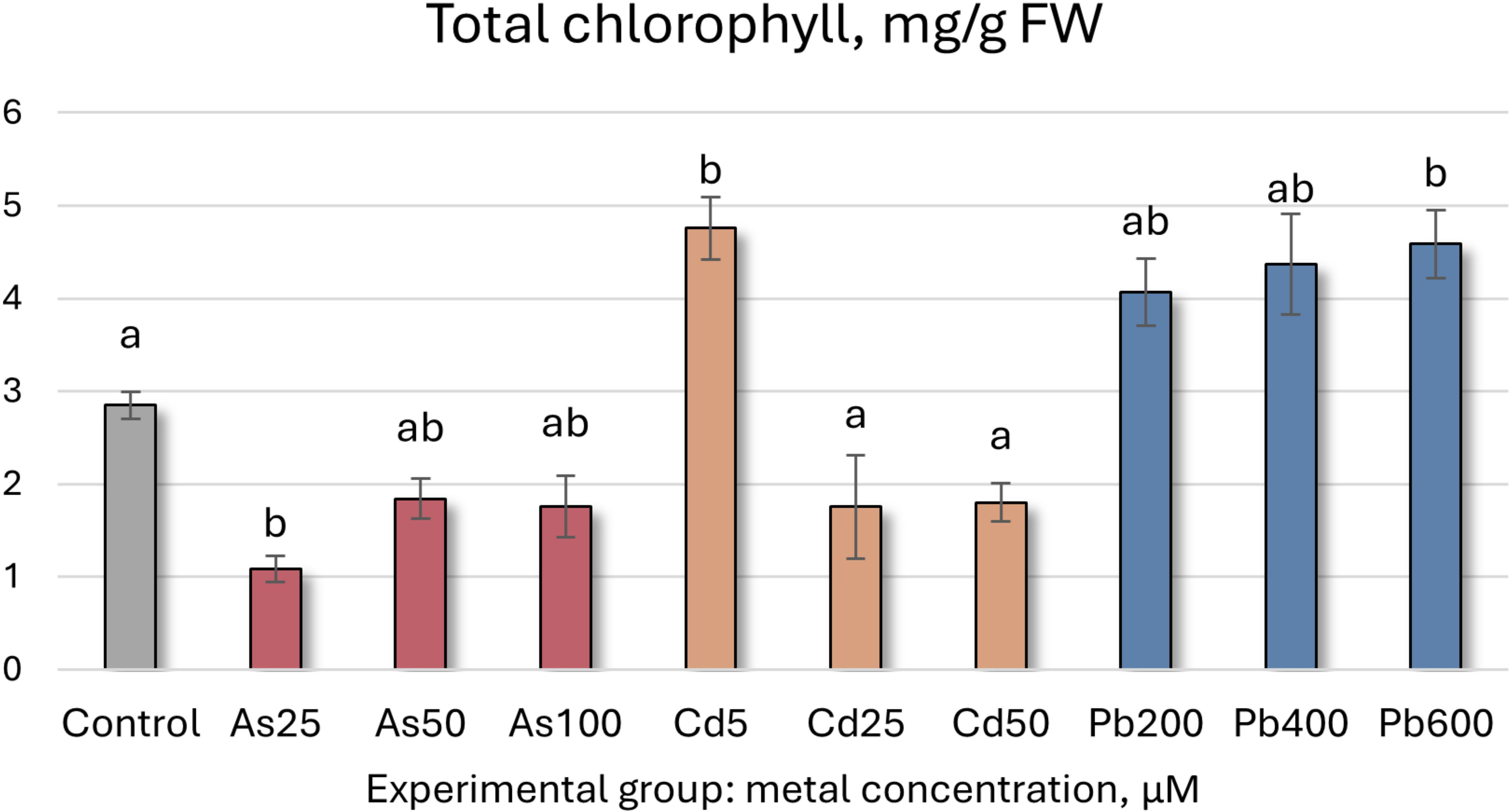
Figure 1. Total chlorophyll concentrations (mg/g FW ± SD) in Salix caprea after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 25, 50, 100 μM of As, 5, 25, 50 μM of Cd, and 200, 400 and 600 μM of Pb). Different letters next to the same color columns and the control denote a significant difference between concentrations based on Dunn’s test (p<0.05).
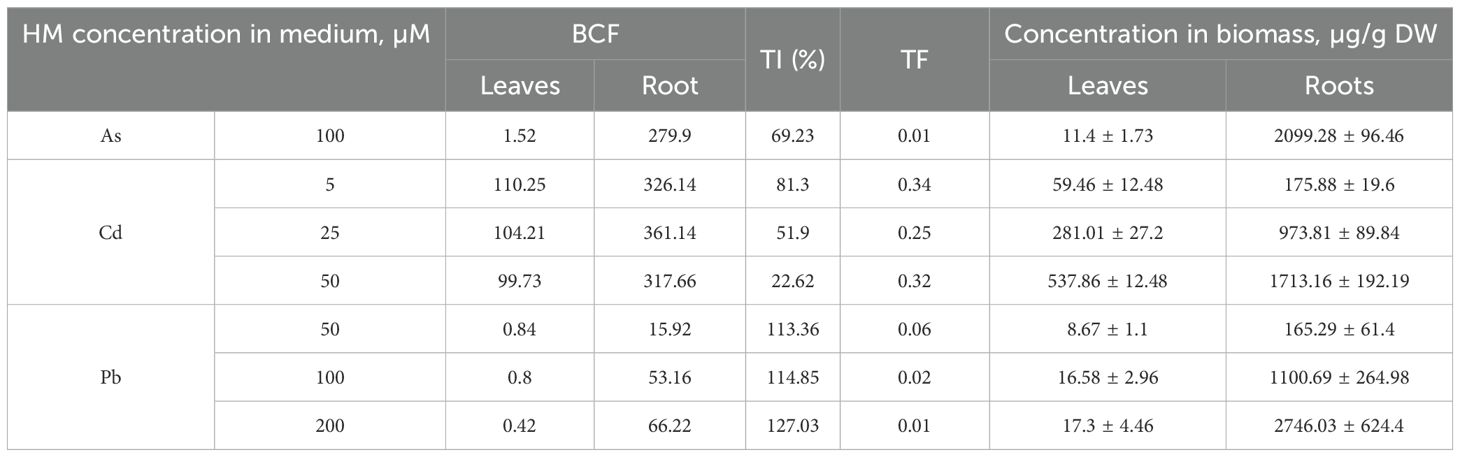
Table 1. Bioconcentration factor (BCF), tolerance index (TI), translocation factor (TF) and heavy metal (HM) concentrations in the leaves and roots of Populus tremula in different experimental groups (from 3 pooled replicates; control – untreated, treated with 100 µM of As, 5, 25, 50 µM of Cd, and 50, 100 and 200 µM of Pb).
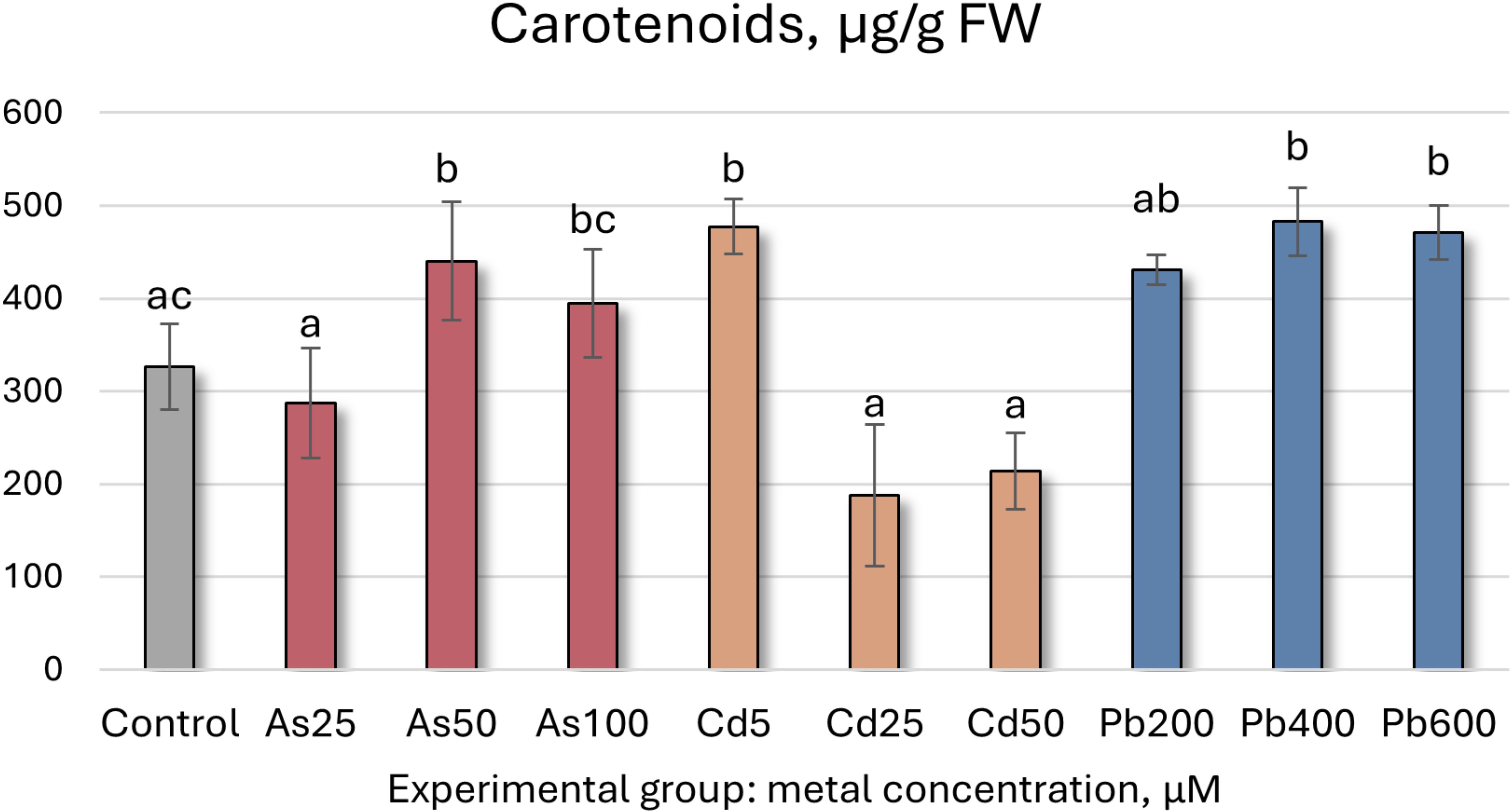
Figure 2. Carotenoid concentrations (μg/g FW ± SD) in Salix caprea after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 25, 50, 100 μM of As, 5, 25, 50 μM of Cd, and 200, 400 and 600 μM of Pb). Different letters next to the same color columns and the control denote a significant difference between concentrations based on Dunn’s test (p<0.05).
Under Cd exposure, P. tremula exhibited low TIs, ranging from 23% to 81% across all tested concentrations (5, 25, and 50 µM), indicating poor seedling health. TFs were relatively high, ranging from 0.25 to 0.34, while BCFs ranged from 99.73 to 110.25 in the leaves and 317.66 to 326.14 in the roots. Similarly, P. tremula showed low tolerance to As (100 µM), with a TI of 69% (Figure 1, Table 1), a TF of 0.01, a low leaf BCF (1.52), and a root BCF of 279.9. In both cases this suggests limited As and Cd translocation but significant root accumulation (Table 1).
HM exposure significantly altered all tested biochemical indicators. While the TI already reflects shoot growth, a closer analysis reveals that As exposure (100 µM) reduced shoot height and root length by 31% and 23%, respectively. In the same group, total chlorophyll concentration decreased by 52% (Figure 3), while MDA (indicator for lipid peroxidation and thus cellular damage) and TPC levels increased. TFC, CAR, and SS levels remained stable, whereas antioxidant enzyme activity generally increased, except for POX, which remained unchanged, and SOD, which declined by 34% (Table 2). Overall, the interpretation of secondary metabolite, SS, CAR, and antioxidant enzyme data is more complex and should be interpreted holistically, as they can both mean enhanced stress and tolerance.
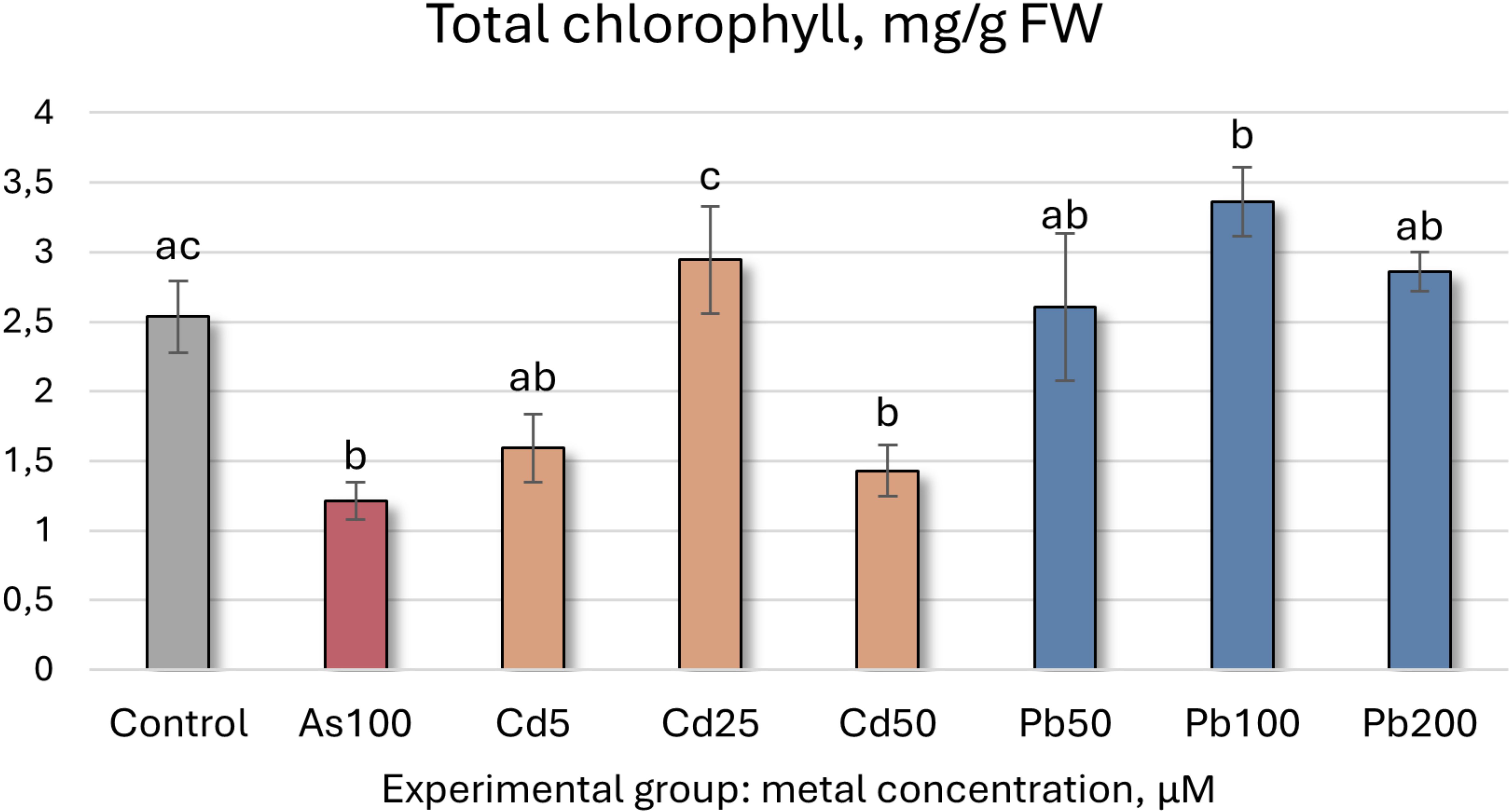
Figure 3. Total chlorophyll concentrations (mg/g FW ± SD) in Populus tremula after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 100 µM of As, 5, 25, 50 µM of Cd, and 50, 100 and 200 µM of Pb). Different letters next to the same color columns and the control denote a significant difference between concentrations based on Dunn’s test (p<0.05).
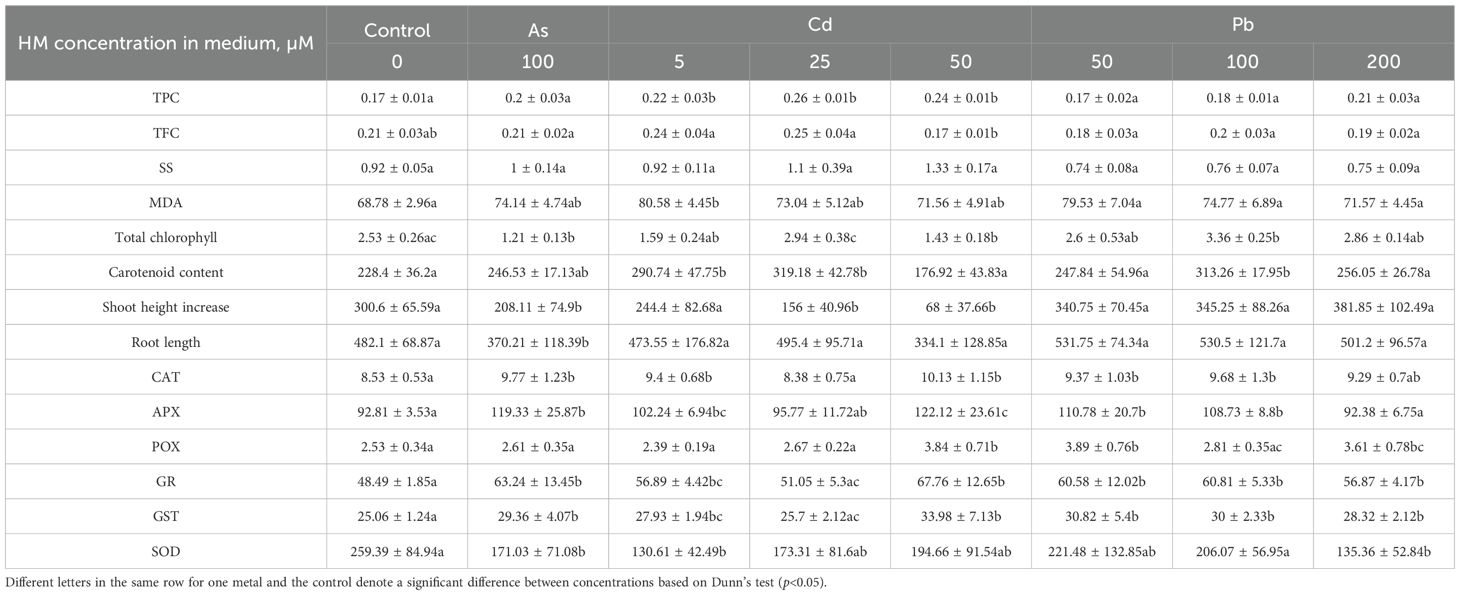
Table 2. Biochemical marker [total chlorophyll, carotenoid (CAR), total phenol (TPC) and total flavonoid (TFC) content, malondialdehyde (MDA), soluble sugar, (SS), antioxidant enzyme (catalase (CAT), superoxide dismutase (SOD), guaiacol peroxidase (POX), ascorbate peroxidase (APX), glutathione reductase (GR), glutathione S-transferase (GST)] levels in the leaves of Populus tremula in different experimental groups (control – untreated, treated with 100 µM of As, 5, 25, 50 µM of Cd, and 50, 100 and 200 µM of Pb).
The data for Cd exposure show similar trends (Table 2). Increasing Cd concentrations led to reduced growth, with shoot and root lengths decreasing by 77% and 31%, respectively, at 50 µM. MDA levels showed a slight increase across all Cd treatments. Total chlorophyll content decreased at 5 µM but exhibited a hormesis effect at 25 and 50 µM. Secondary metabolite (TPC and TFC) and CAR levels fluctuated, while antioxidant enzyme activity (CAT, POX, APX, GR, and GST) generally increased, except for SOD, which declined. These findings showcase that both As and Cd are phytotoxic to P. tremula at the tested concentrations.
Pb exposure yielded distinct results. Overall, growth remained statistically unchanged, with a slight upward trend as Pb concentrations increased, as can be noted from the TIs (Tables 1, 2; Figure 2). Total chlorophyll content significantly increased by 33% at 100 µM Pb, while remaining steady at 50 and 200 µM displaying a hormesis effect. SS, TFC, and MDA levels showed no significant changes, whereas TPC trended upward at 200 µM. CAR levels were slightly elevated at 100 µM (37%) and exhibited a hormesis effect across Pb concentrations. Antioxidant enzyme activity generally increased, except for SOD, which consistently decreased in all Pb-treated groups. These findings suggest that P. tremula responds positively to Pb contamination at these levels.
Furthermore, the elemental analysis of P, K, Ca, Mg, S, Fe, Mn, Cu, and Zn in P. tremula leaves and roots clearly indicates that HM contamination in the nutrient medium leads to significant metabolic shifts distinct in the leaves and roots. In all cases, both positive and negative shifts can be noted (Table 3). In the leaves, As exposure shows a significant increase in S and Zn, while most other elements decrease, especially Fe and Mn. In the roots, P, K, Mg, Mn and S are elevated, while Fe, Cu, Ca and Zn are substantially reduced. At low concentrations (50 µM), Pb generally increases P in the leaves and roots and Zn in the leaves, decreases Fe in the leaves and Zn in the roots. As the concentration increases (100 µM and 200 µM), the trend continues with increases in P in the leaves with decreases of S, Mn, Zn in the leaves. Increases of K, Ca and Mg and decreases of Cu and Zn in the roots can be noted, while other elements show more variability. Cd exposure shows a mix of increases and decreases in the leaves and roots. In leaves, S and Zn increase with increasing concentration, with notable reductions in K, Mg, Mn, and Fe. In the roots, P, and Mg show large increases, while Mn and Zn decrease with increasing concentration. As is particularly toxic to both the leaves and roots, showing marked decreases in essential elements such as Fe, Mn, and Cu, but large increases in S and Zn.
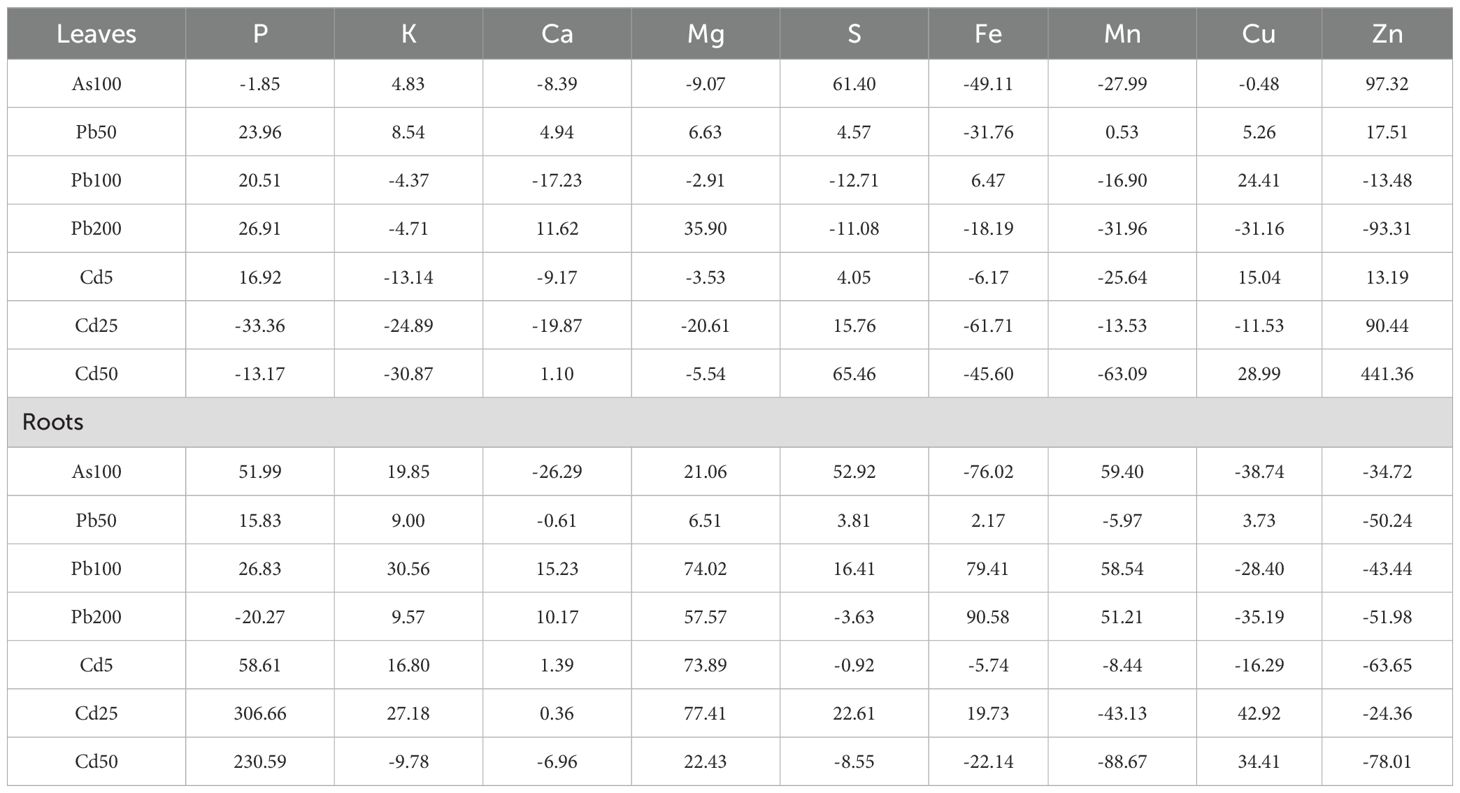
Table 3. Concentrations of various elements (P, K, Ca, Mg, S, Fe, Mn, Cu, Zn) in the leaves and roots of Populus tremula in different experimental groups (from 3 pooled replicates; treated with 100 µM of As, 5, 25, 50 µM of Cd, and 50, 100 and 200 µM of Pb) expressed as percent differences from control – untreated group.
Overall, Pb has a more positive effect on element uptake in both leaves and roots at lower concentrations (50 µM), but higher concentrations (100 µM and 200 µM) show more complex interactions in the leaves, yet with continued increased metal concentrations (e.g., Fe, K, Mg) in the roots. Cd displays a similar complex trend, with some elements (like Zn and S) showing considerable increases at higher concentrations in the leaves, while others (like K in the leaves and Mn in the roots) decrease significantly.
3.2 HM impact on Salix caprea
The severe chlorosis observed in P. tremula treated with 500 and 1000 µM of As, along with its strong positive response to Pb treatment, prompted an adjustment in the tested concentrations for S. caprea. As a result, S. caprea exposed to 25, 50, and 100 µM of As exhibited improved TIs, ranging from 91% to 108% (Table 4). In these groups, TF varied between 0.02 and 0.11, while the leaf BCF ranged from 1.28 to 15.6, and the root BCF spanned from 65.33 to 172.72. Comprehensive data on HM concentrations in Salix seedlings can be found in Table 4. As levels ranged from 10 to 29 µg/g DW in the leaves and 323 to 490 µg/g DW in the roots. Cd concentrations were 58 to 484 µg/g DW in the leaves and 80 to 622 µg/g DW in the roots, while Pb levels varied from 22 to 92 µg/g DW in the leaves and 500 to 5372 µg/g DW in the roots. These results follow a similar trend observed in Populus seedlings, where increasing HM concentrations in the medium corresponded to higher accumulation in plant tissues, with roots consistently exhibiting greater HM levels than leaves.
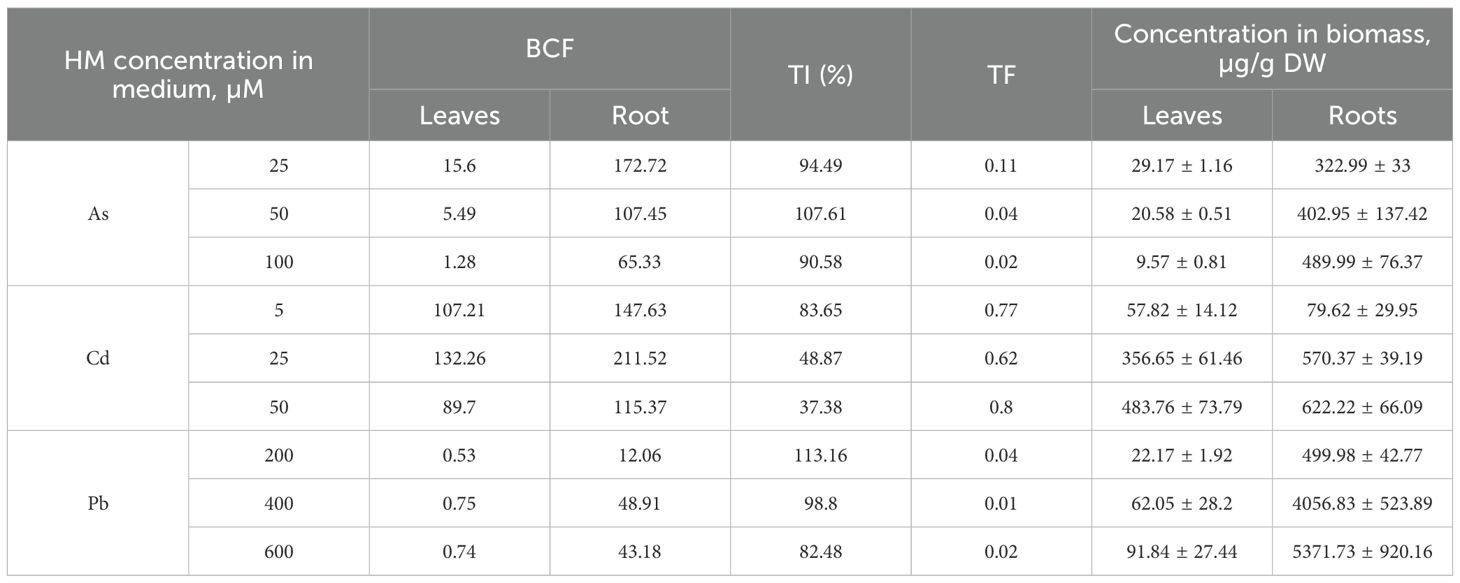
Table 4. Bioconcentration factor (BCF), tolerance index (TI), translocation factor (TF) and heavy metal concentrations in the leaves and roots of Salix caprea in different experimental groups (from 3 pooled replicates; control – untreated, treated with 25, 50, 100 µM of As, 5, 25, 50 µM of Cd, and 200, 400 and 600 µM of Pb).
Cd contamination yielded intriguing results (Table 4). Salix seedlings treated with Cd displayed significantly higher translocation factors compared to other tested groups, ranging from 0.62 to 0.8. TIs varied between 37% and 84% (Figure 4, Table 4), while the leaf BCF ranged from 89.7 to 132.26, and the root BCF spanned from 115.37 to 211.52. Pb treatment yielded TIs of 82-133%, TFs of 0.01-0.04, leaf BCF of below 1 (0.53-0.75) and root BCFs ranging from 12.06 to 48.91. Notably, all Cd concentrations resulted in a measurable reduction in growth. However, data from the lowest tested concentration (5 µM) indicate that S. caprea exhibits hyperaccumulator potential for Cd.

Figure 4. Populus tremula L. after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 50, 100 and 200 µM of Pb).
Further analysis of S. caprea seedling biochemical health and growth markers reveals that the higher As concentrations (50 and 100 µM) induce significant shifts in the photosynthetic apparatus (Table 5, Figures 5, 6). Total chlorophyll levels decline across all As-treated groups by 35–62% (Figure 5). Similarly, carotenoid levels initially decrease before increasing, with a notable 35% rise at 50 µM of As (Table 5, Figure 6). Growth, MDA, SS, antioxidant enzyme activity, TFC, and TPC levels remain relatively stable at the lower concentrations (25 and 50 µM). However, at 100 µM of As, antioxidant enzyme activity decreases across all tested enzymes except for SOD.
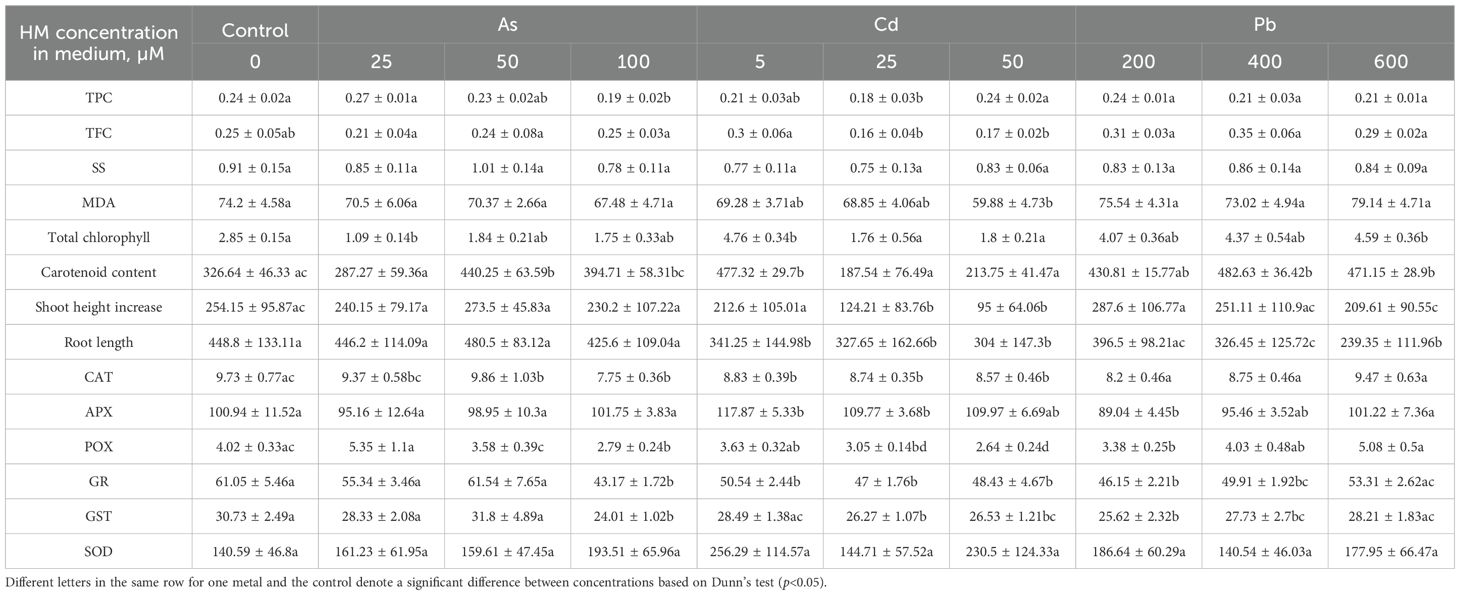
Table 5. Biochemical marker [total chlorophyll, carotenoid (CAR), total phenol (TPC) and total flavonoid (TFC) content, malondialdehyde (MDA), soluble sugar, (SS), antioxidant enzyme (catalase (CAT), superoxide dismutase (SOD), guaiacol peroxidase (POX), ascorbate peroxidase (APX), glutathione reductase (GR), glutathione S-transferase (GST)] levels in the leaves of Salix caprea in different experimental groups (control – untreated, treated with 25, 50, 100 µM of As, 5, 25, 50 µM of Cd, and 200, 400 and 600 µM of Pb).

Figure 5. Populus tremula L. after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 100, 500 and 1000 µM of As).

Figure 6. Salix caprea after 4 weeks grown in hydroponic conditions in different experimental groups (control – untreated, treated with 5, 25 and 50 µM of Cd).
In contrast, the lowest Cd concentration (5 µM) led to a significant increase in both total chlorophyll concentration and carotenoid levels, rising by 67% and 46%, respectively. However, higher Cd concentrations resulted in declines for both parameters (Figures 5, 6, Table 5). MDA and SS levels remained largely stable, but growth was negatively affected, particularly in the roots, with increasing Cd concentrations. Root length decreased by 24–32%, while shoot height increase was reduced by 16–63% (Figure 4). Antioxidant enzyme activity declined at the higher Cd concentrations (25 and 50 µM), except for SOD, which remained unchanged, and APX, which increased. At 5 µM Cd, only CAT, APX, and GR were affected, with APX activity rising by 17%, while CAT and GR levels decreased by 9% and 17%, respectively. Cd-treated seedlings also exhibited variability in both TPC and TFC.
In the Pb-treated group, growth parameters remained stable at the lowest concentration (200 µM) but declined at the higher concentrations (400 and 600 µM) (Table 5). Pb-treated seedlings also exhibited increased total chlorophyll levels, with a 61% rise at the highest concentration (Figure 5), along with elevated carotenoid levels (ranging from 32% to 44%) (Figure 6). TPC, SS, MDA, and SOD enzyme levels remained relatively unchanged, while TFC increased by 15-39%. At 200 µM Pb, most antioxidant enzyme activities were reduced, whereas at 400 µM, they exhibited some variability. Overall, these findings suggest that S. caprea has potential as a Cd hyperaccumulator at low concentrations. Additionally, this representative of the Salicaceae family, along with P. tremula, may be suitable for Pb phytostabilization.
Nutrient homeostasis plays a critical role in plant stress responses under HM exposure. Each metal demonstrates distinct patterns of uptake and distribution of nutrients across the Salix plant, with the roots and leaves responding differently based on the concentration of contamination (Table 6). As-treated groups exhibited a severe Ca and Mg depletion across roots and leaves, P depletion in the leaves, and Fe, Mn, Cu and Zn depletion in the roots. The declines in root nutrients were more pronounced, while leaves showed sporadic increases in metals. S accumulation was high, especially in roots. Fe, Mn and Cu showed elevated levels in the leaves. Pb-treated groups displayed a steady depletion of Ca and Mg in both roots and leaves. Zn accumulated in leaves but decreased in roots. Roots were less affected in P and K levels compared to leaves. Cu levels increased in the leaves, but decreased in the roots, while the other element response to Pb was variable. Under Cd contamination Cu and Zn levels were high in leaves but severely depleted in roots. The seedlings showed nutrient stress, with a general depletion of Ca, Mg, Fe, and Zn. Cd-treated roots accumulated P, K and S but showed losses in other nutrients.
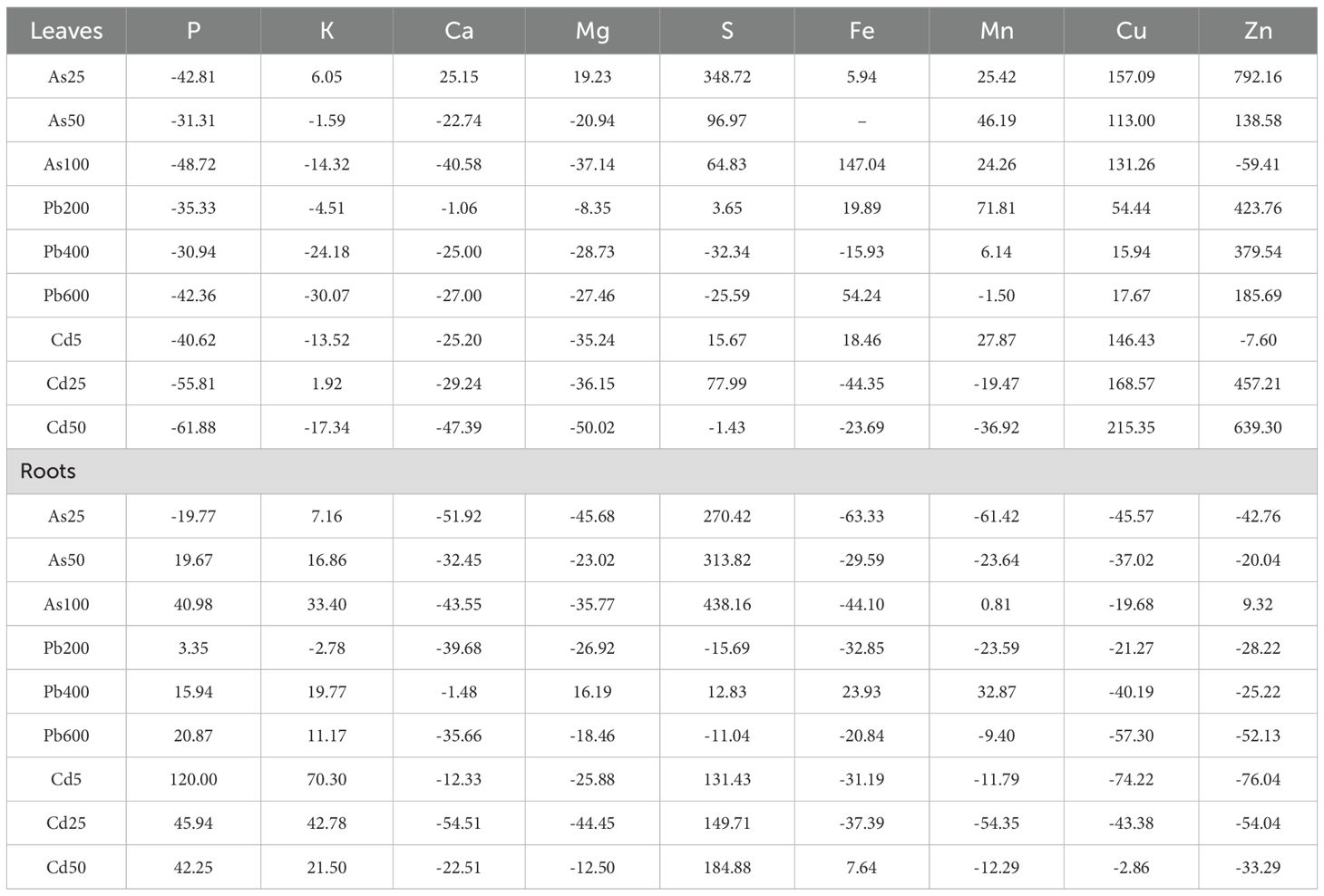
Table 6. Concentrations of various elements (P, K, Ca, Mg, S, Fe, Mn, Cu, Zn) in the leaves and roots of Salix caprea in different experimental groups (from 3 pooled replicates; treated with 25, 50, 100 µM of As, 5, 25, 50 µM of Cd, and 200, 400 and 600 µM of Pb) expressed as percent differences from control – untreated group.
Overall, Cd caused severe nutrient disruptions while Pb had a more moderate effect, allowing some Zn accumulation in leaves. Roots were more negatively affected across all metals.
4 Discussion
This study provides new insights into the phytoremediation potential of Populus tremula and Salix caprea, with a particular focus on seedling responses to As, Cd, and Pb exposure in a hydroponic system. Our findings demonstrate that both species exhibit distinct metal-specific and dose-specific responses during early development, with several key discoveries emerging from this research: (I) integrated assessment of antioxidant enzyme systems, secondary metabolites, and nutrient dynamics revealed species-specific tolerance mechanisms, and (II) species-specific bioaccumulation and translocation patterns were identified that inform targeted phytoremediation strategies. By analyzing growth, HM uptake efficiency, biochemical status, stress markers, and elemental composition shifts, this study offers a comprehensive evaluation of how these species respond to HM contamination at the critical seedling establishment phase. The results provide essential baseline data for understanding early-stage metal tolerance mechanisms and support the development of more targeted phytoremediation approaches using these economically and ecologically important species.
Most phytoremediation studies have focused on the ability of P. tremula and S. caprea to accumulate HMs when propagated from cuttings and/or grown in soil (Konlechner et al., 2013; Salam et al., 2022). In contrast, our study utilized seedlings and a hydroponic approach, eliminating soil-related confounding factors to allow precise quantification of metal uptake and physiological responses. This methodological choice offers a clearer understanding of metal stress adaptation at the early seedling stage and highlights previously underexplored biochemical and metabolic adjustments (Pietrini et al., 2010). Building upon these methodological advantages, our findings revealed distinct metal- and dose-specific responses that highlight the complexity of phytoremediation mechanisms. Pb exposure had minimal effects on growth and often induced hormesis, suggesting that low to moderate Pb levels may not severely impair physiological function in these species and in fact both species exhibited higher TI (based on shoot growth) ranging from 113% (50 µM) to 127% (200 µM) for P. tremula and 113% for S. caprea (200 µM of Pb). Conversely, Cd and As reduced TIs, and both shoot and root growth. These findings correspond with previous data, that Pb is less toxic to plants than Cd or As, possibly due to relatively reduced Pb mobility in the plant (Reddy and Patrick, 1977; Fritz et al., 2022). Furthermore, the observed hormetic effect has also been reported in previous studies on HMs. For instance, the potential Cd hyperaccumulator Bidens pilosa L. exhibited a 13.8% increase in shoot biomass (Sun et al., 2009), while Pb was found to enhance root system length in wheat (Erofeeva, 2014). Similar effects were later confirmed in corn for both Cd and Pb (Małkowski et al., 2020). However, growth reduction remains a widely documented response to HM exposure in plants (Schützendübel et al., 2001; Pietrini et al., 2010; Nahvi et al., 2024).
Complementing these growth responses, photosynthetic pigment analysis revealed further evidence of metal-specific adaptation mechanisms. Both Pb-treated trees showcased an increase in total chlorophyll levels and S. caprea also exhibited higher chlorophyll levels in seedlings treated with the lowest concentration of Cd (5 µM), while other Cd- and As-treated groups exhibited lower concentrations with carotenoid levels generally expressing the same tendencies. The increase in pigment content, specifically chlorophyll levels, has been previously observed, though not often, in other species (Agathokleous et al., 2020; Małkowski et al., 2020) with several potential mechanistic explanations behind this. This could be the cumulative result of a robust antioxidant system, which can mitigate oxidative stress and protect chlorophyll from degradation and an upregulation of biosynthetic pathways. For example, some HMs at low concentrations could potentially act as micronutrients [e.g., Cd mimicking Zn (Pongrac et al., 2009; Muszyńska and Labudda, 2019)]. While this is not very common for the non-essential elements like Cd, As and Pb, as opposed to higher doses of essential elements like Fe, Zn, etc., studies do mention the phenomenon (Muszyńska and Labudda, 2019). Some HMs can also induce phytohormone-like effects. This trait is not only linked with stimulating photosynthetic pigment production but may be responsible for slight lean toward shoot proliferation and simultaneously reduced root elongation, which was noted in Pb-treated groups in the current study. This was previously observed in corn treated with Pb. The authors discovered that the hormetic stimulation of shoot growth by HMs was consistently associated with increased auxin and flavonol levels, while maintaining hydrogen peroxide concentrations comparable to those in control plants (Małkowski et al., 2020).
On the other hand, HM toxicity often has a detrimental effect on the photosynthetic apparatus through a variety of pathways, affecting multiple components at different levels (Pietrini et al., 2010; Nahvi et al., 2024). Toxicity increases reactive oxygen species (ROS) and this can lead to peroxidation of thylakoid membranes, reducing electron transport efficiency (Bashir et al., 2015; Małkowski et al., 2020). HMs can directly inhibit crucial enzymes (like ALA dehydratase (Stobart et al., 1985; Sarangthem et al., 2011) and protochlorophyllide reductase (Stobart et al., 1985)), bind to key proteins (like D1) (Geiken et al., 1998) within the photosynthetic pathways. Furthermore, inhibition of oxygen-evolving complex disrupts water photolysis, reducing O2 evolution and ATP synthesis (Azmat et al., 2022). HMs can induce stomatal closure, reducing CO2 uptake and directly affecting RuBisCO activity too (Perfus-Barbeoch et al., 2002; Wang et al., 2009). This was shown in a hydroponic experiment with Cd-treated Arabidopsis thaliana L., Vicia faba L. and Commelina communis L (Perfus-Barbeoch et al., 2002). Pb can affect PEP carboxylase, and other key enzymes lowering CO2 assimilation (Wang et al., 2009), while As can interfere with phosphate metabolism, limiting ATP production (Finnegan and Chen, 2012).
To understand the mechanisms underlying these photosynthetic responses, we conducted detailed biochemical profiling that represents a major strength of the current study, which offers mechanistic insights into the antioxidant responses and metabolic shifts induced by HMs. The use of multiple biochemical markers, including lipid peroxidation (MDA), soluble sugars, flavonoids, and phenolic compounds, provides a holistic view of the stress response mechanisms in the tested species. Interestingly, in the current study while growth and pigment concentrations were affected, HM treatment minimally altered biochemical markers such as MDA and SS. In both trees, these indicators remained relatively steady, with the exception being Cd-treated Populus showing elevated MDA at the lowest concentration and upward trending SS levels. As MDA directly indicates lipid peroxidation levels, it gives a clear image of the HM caused ROS damage (Sun et al., 2009; Erofeeva, 2014). However, if antioxidant defences are highly active, lipid peroxidation (and thus MDA levels) may remain low despite HM exposure (Pang et al., 2002). In contrast SS levels can indicate several things; a direct increase in glucose production – carbon metabolism, and osmotic adjustment, which relates to stress levels (Rosa et al., 2009). Also, instead of accumulating soluble sugars, the plant might be diverting carbon into secondary metabolites (e.g. phenols) to enhance the antioxidative response (Caretto et al., 2015). Increased sugar transport to roots may also prevent SS buildup in leaves (Du et al., 2020).
As in both cases of HM impact on the photosynthesis system, and stress indicator levels, ROS plays a huge role, it’s imperative to look into the enzymatic and non-enzymatic antioxidant systems, i.e., secondary metabolites (TPC, TFC) and antioxidant enzymes (SOD, CAT, POX, APX, GR, GST) (Asare et al., 2023). These two systems work together but differ in their mechanism, speed, energy cost, and efficiency. Their deployment depends on the type, intensity, and duration of stress. Overall, the enzymes represent the first line of defense, i.e., rapid response, lower cost, while the non-enzymatic antioxidants represent the secondary or long-term defense (higher cost), which is triggered when ROS levels persist, or enzymatic defenses are insufficient (Gill and Tuteja, 2010). The slightly elevated MDA and SS levels noted in the Populus trial (Cd at 5 µM) can be attributed to induced stress, however, MDA levels decreased in the two higher concentration groups. This could be the result of the accompanying impact of slightly elevated SS and increased TPC (33-53%) levels. Furthermore, the decrease in antioxidant enzyme levels is evident in the same group. This further showcases the shift towards SMs rather than enzymatic response, which mitigated lipid damage. A similar response was noted in Cd-treated Pinus sylvestris L. Initially the treatment caused an increase in the enzymatic response, but it later shifted towards the accumulation of soluble phenolics (Schützendübel et al., 2001). Opposingly, P. tremula group treated with 50 µM of Cd demonstrates an increase in enzymatic activity alongside elevated TPC. The activation of both systems suggests an increase in stress levels. Again, this was previously noted in trials with other HMs in maize (AbdElgawad et al., 2020).
Cd-treated S. caprea showcases yet another response variation – reduced enzyme production (except for steady SOD and slightly upregulated APX) and reduced TFC production (33-35%) in the two higher Cd concentrations, and reduced TPC in the 25 µM treated group. At the same time growth rates were declining, as were photosynthesis pigment content. This indicates severe oxidative stress and metabolic disruption. Cd and Pb toxicity is known to interfere with phenylalanine ammonia-lyase (PAL), a key enzyme in flavonoid biosynthesis, in both a positive and negative way (Jańczak-Pieniążek et al., 2023). Additionally, Cd might have damaged chloroplasts and plastids (Lee and Back, 2017), which can impact phenol synthesis, or flavonoids and phenolics could have been depleted too quickly due to high ROS levels, exceeding the plant’s capacity to replenish them. Cd may also inhibit key enzyme production directly through cofactor replacement, protein binding or glutathione metabolism (Stobart et al., 1985; Geiken et al., 1998; Paradiso et al., 2008; Sarangthem et al., 2011).
A similar response can be noted in As-treated S. caprea. The group treated with 100 µM was mainly affected with reduced levels of CAT, POX, GR, GST and TPC. However, as the growth rate is similar to the control group, it could mean that S. caprea relies less on enzymatic detoxification and more on metal exclusion. The plant might be sequestering As into vacuoles or binding it to cell wall components (Rascio and Navari-Izzo, 2011). HM exclusion strategy is also a high likelihood as HM accumulation data suggests that the plant may be blocking As uptake at the root level (high levels in the roots vs. the leaves), reducing the need for detoxification compounds. Moreover, As is known to interfere with enzyme activity by binding to thiol groups (-SH) in proteins (January et al., 2008), as does Cd (Stobart et al., 1985; January et al., 2008), possibly leading to inhibited GR and GST (which require thiol-based glutathione to function) (Gill and Tuteja, 2010). Overall, this indicates that S. caprea employs As avoidance strategies at the concentrations tested rather than detoxification. Furthermore, elevated antioxidant enzyme activity (except for SOD) suggests an active detoxification mechanism that mitigates oxidative damage in Populus treated with As (100 µM). This suggests H2O2 accumulation and increased reliance on the ascorbate-glutathione (ASC-GSH) cycle to mitigate oxidative stress and As toxicity. This was previously observed in wheat with Cd toxicity, whereby ASC-GSH cycle enzymes showed increased activity in roots, correlating with Cd accumulation. Supplementing ASC biosynthesis alleviated Cd-induced oxidative stress in roots, without affecting Cd uptake (Paradiso et al., 2008). These findings reiterate that the ASC-GSH cycle is playing a crucial role in systemic antioxidant defense.
Pb treated S. caprea also exhibits efficient enzymatic detoxification mechanism with the activities of 5 out of 6 enzymes elevated (except for SOD), with other parameters steady at 200 µM. This changes with increasing concentrations – the metabolic shift towards increased enzyme and carotenoid (carotenoids may act as ROS scavengers too (Gill and Tuteja, 2010)) production is evident. Moreover, at the lower Pb concentrations studied in P. tremula, an increase in the enzymatic response was observed, but no corresponding SM production or other stress indicators. This, in part, corresponds with a review on Pb impact on plant metabolism. Collin et al. note that Pb treatment impact antioxidant enzyme activity positively, but they don’t mention SMs (Collin et al., 2022).
These biochemical adaptations directly influence metal accumulation patterns, with HM concentrations in both species being more concentrated in the roots for all three metals tested. The highest concentrations in the roots were noted in S. caprea for all three metals (490, 622, and 5372 µg/mg DW respectively for As, Cd and Pb), while the highest concentrations in the leaves varied – Pb and As were also highest in S. caprea (29 and 92 µg/mg DW respectively for As and Pb), but Cd concentrations were highest in P. tremula (538 µg/mg DW). Generally, the concentrations of HMs in both leaves and roots were higher with the increasing HM concentrations in the substrate. These differences reinforce the idea that different genotype/species plays a significant role in determining a plant’s bioaccumulation potential.
BCFs in both vegetative organs varied, with root numbers being higher. A BCF>1 signifies efficient metal accumulation, supporting phytoextraction, whereas BCF<1 suggests exclusion or limited uptake, favoring phytostabilization. Pb BCFs were around 1 in the leaves in both species, while they ranged from 12 to 49 in S. caprea and 16 to 66 in P. tremula roots. Cd BCFs were 90–132 in Salix and 100–110 in Populus leaves, while they ranged from 115 to 212 in S. caprea and 318 to 361 in P. tremula roots. As BCFs were 1–16 in Salix and were ~2 in Populus leaves, while they ranged from 65 to 173 in S. caprea and were 280 in P. tremula roots. In general, the TFs in both species were low. A TF>1 indicates efficient HM translocation to shoots, favoring phytoextraction, whereas TF<1 suggests metal retention in the roots, making the plant more suitable for phytostabilization. For Pb they ranged from 0.01 to 0.06 in both species, for As they were 0.01-0.11. These TFs along with the BCFs indicate that both species demonstrate strong phytostabilization capacity for Pb, suggesting their potential use in contaminated environments. Phytostabilization is a remediation technique whereby plants reduce the mobility and bioavailability of contaminants in the substrate. This process helps prevent the spread of pollutants, thus minimizing their uptake by other plants or leaching. Various mechanisms can be employed by the plant to achieve this goal, however in the case of the current study root absorption and binding is the most likely. Plant roots absorb contaminants but do not translocate them to aerial parts, keeping them contained in the rhizosphere (Rascio and Navari-Izzo, 2011). Furthermore, on the subject of Pb hyperaccumulation, Egendorf et al. note that while universal standards for Pb hyperaccumulation are not set, up until 2020 no species was shown to have BCF and TF over 1 and accumulate 1 g/kg of Pb in the roots, i.e., 0.1% (Egendorf et al., 2020). The current data is in line with their assessment, however S. caprea did notably exhibit >0.5% accumulation rate in the roots, with a high BCF in Pb trials.
The TFs were higher for Cd, ranging 0.25-0.34 in Populus and with Cd-treated S. caprea displaying increased translocation potentials (TF 0.62-0.8) in Salix. This aligns with prior studies suggesting that Salix species possess strong Cd accumulation capabilities (Xu et al., 2019; Kong et al., 2020). Notably, plants with TF values >1 are considered hyperaccumulators, however even when TF values remain below 1, as in the case of S. caprea and Cd, the rapid growth and substantial biomass production of these trees could likely compensate for lower translocation efficiency, making them effective candidates for phytoremediation in the form of phytoextraction (Pulford and Watson, 2003). Unlike phytostabilization, phytoextraction focuses on long-term removal of contaminants by accumulating them in harvestable biomass. HMs are stored in vacuoles or bound to cell wall components in leaves and stems to minimize toxicity. Interestingly, some plants have adapted to sequester and detoxify HMs in cell components, which has been linked to reduced herbivory, pest resistance, and pathogen deterrence in plants.
The observed bioaccumulation patterns are intrinsically linked to broader physiological adjustments, as evidenced by the differential nutrient uptake patterns that suggest complex ion-homeostasis mechanisms. HMs can interfere with nutrient absorption at the root level, altering the availability and mobility of elements in plant tissues through competition for transporters or damage to the roots (Muszyńska and Labudda, 2019). A trend of reduced levels of Ca and Mg was noted in all experimental groups of tested metals in S. caprea in both leaves and roots. It has long been known that HMs can mimic Mg and thus take its place in the chlorophyll complex – this being one of the key mechanisms behind the damage to the photosynthesis apparatus by HMs (Riyazuddin et al., 2022). Additionally, Ca reduction with increasing Cd concentrations has been reported for barley (Veltrup, 1981). Similarly, nutrient leaching is also a likely scenario. For example, membrane lipid peroxidation causes K and Mg leakage from leaves. Murphy et al. reported that K leakage from Arabidopsis leaves did increase short term under Cu treatment (Murphy et al., 1999). In the current study the general trend was towards P, Ca, and Mg reduction in the leaves of both species, with the additional reduction of K in Salix. As mentioned, HMs can interfere with P assimilation (Finnegan and Chen, 2012). Additionally, oxidative stress can trigger Fe and Mn remobilization (noted in P. tremula), as both Fe and Mn are components in the antioxidative mechanism (Briat et al., 2010; Chandra et al., 2020). HMs may alter the redistribution and translocation patterns, i.e., xylem and phloem loading of nutrients, leading to imbalances in leaves and roots (Page and Feller, 2015). This was especially notable in both tested species in P and K patterns. Furthermore, “co-accumulation” is also a possibility, e.g., both Cd²+ and Zn²+ use the ZIP family transporters, and Zn accumulation was shown to positively correlate with increased Cd accumulation (Pongrac et al., 2009). In the current study, this phenomenon was noted in both species leaves with Salix exhibiting enhanced Zn levels of up to 639% and Populus expressing enhanced Zn levels of up to 441% (in both cases Zn levels and Cd levels increased with increasing concentrations of Cd). Moreover, S accumulation is a key protective strategy against HM toxicity (Dixit et al., 2016). In rice, high S treatment has been shown to reduce As accumulation in shoots by altering As transporters and enhancing antioxidant enzyme activities (Dixit et al., 2016). In the current study S accumulation was noted in both species in both leaves and roots.
Integrating our nutrient dynamics findings with the photosynthetic responses provides mechanistic insights into adaptive strategies. The relationship between nutrient levels and enhanced total chlorophyll levels in specific treatment groups reveals the intricate nature of metal tolerance mechanisms. For example, in Cd- and Pb-treated Salix (5 µM of Cd, 600 µM of Pb) and Populus (25 µM of Cd, 100 µM of Pb). In both cases of Cd, a steady increase in S levels was observed. Enhanced levels of Cu and Zn can be noted in Pb-treated Salix (600 µM), while P and Cu levels increased in Populus (100 µM of Pb). Various nutrients (P, K, Ca, Zn, Cu) were shown to enhance chlorophyll content by contributing indirectly by supporting photosynthesis, enzyme activity, and chlorophyll stability (Widjaja et al., 2024). These findings highlight the intricate interplay between nutrient dynamics and chlorophyll accumulation, suggesting a potential adaptive response to heavy metal exposure in Salix and Populus.
5 Conclusions
This study underscores the potential of P. tremula and S. caprea in phytoremediation applications, particularly for Pb phytostabilization (P. tremula and S. caprea) (concentrations equivalent up to 200 µM of Pb(NO3)2) and Cd phytoextraction (S. caprea) (concentrations equivalent up to 5 µM of CdSO4). At these levels a hormetic effect for both Pb-treated species is evident, while TF of 0.77 was noted in Cd-treated S. caprea (5 µM). A deep dive into the biochemical and physiological responses provides a clear understanding of how P. tremula and S. caprea react to heavy metal stress, highlighting the antioxidant response hierarchy, i.e., an initial enzymatic response, and a shift towards secondary metabolite production should the stressful conditions continue or are too severe, and shifts in elemental composition (Cd and Zn co-accumulation standing out). Future research should focus on long-term field studies to validate these findings in soil environments, explore microbial interactions that may enhance metal uptake and plant health, and assess strategies for biomass management post-harvest. Additionally, future research should focus on long-term effects, particularly concerning the sustainability of phytoremediation, which remain to be explored. Overall, this study contributes to a growing body of evidence supporting the use of fast-growing tree species in sustainable phytoremediation strategies, providing a foundation for further research into optimizing their efficiency for large-scale applications.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
VS-Š: Project administration, Formal Analysis, Writing – original draft, Resources, Conceptualization, Investigation, Validation, Writing – review & editing, Supervision, Funding acquisition, Methodology. GS: Conceptualization, Methodology, Investigation, Writing – review & editing, Validation, Formal Analysis, Visualization, Software, Data curation, Writing – original draft. MŠ: Writing – original draft, Formal Analysis, Visualization, Investigation, Writing – review & editing, Validation. IK: Writing – original draft, Formal Analysis, Software, Validation, Writing – review & editing, Investigation, Data curation. MP: Writing – review & editing, Writing – original draft, Investigation, Formal Analysis, Conceptualization, Validation, Methodology. IV: Validation, Supervision, Methodology, Formal Analysis, Conceptualization, Writing – original draft, Writing – review & editing, Investigation. EL: Writing – review & editing, Investigation, Conceptualization, Validation, Writing – original draft, Formal Analysis, Methodology. DV: Methodology, Data curation, Investigation, Software, Conceptualization, Writing – original draft, Validation, Visualization, Formal Analysis, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the European Union’s Horizon Europe research and innovation programme under grant agreement No. 101138432.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. During the preparation of this work the author(s) used ChatGPT (OpenAI) in order to improve the readability and language of the manuscript. After using this tool, the author(s) reviewed and edited the content as needed and take(s) full responsibility for the content of the published article.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1617432/full#supplementary-material
Abbreviations
APX, ascorbate peroxidase; CAR, carotenoid content; CAT, catalase; GR, glutathione reductase; GST, glutathione S-transferase; MDA, malondialdehyde; POX, guaiacol peroxidase; SOD, superoxide dismutase; SS, soluble sugars; TFC, total flavonoid content; TPC, total phenolic content; HM, heavy metals; SM, secondary metabolites; BCF, bioconcentration factor; TI, tolerance index; TF, translocation factor; DW, dry weight; ROS, reactive oxygen species; ALA, δ-aminolevulinic acid; RuBisCO, ribulose-1,5-bisphosphate carboxylase/oxygenase; PEP, phosphoenolpyruvate; PAL, phenylalanine ammonia-lyase; ASC-GSH, ascorbate-glutathione.
References
AbdElgawad, H., Zinta, G., Hamed, B. A., Selim, S., Beemster, G., Hozzein, W. N., et al. (2020). Maize roots and shoots show distinct profiles of oxidative stress and antioxidant defense under heavy metal toxicity. Environ. pollut. 258, 113705. doi: 10.1016/j.envpol.2019.113705
Agathokleous, E., Feng, Z. Z., and Peñuelas, J. (2020). Chlorophyll hormesis: Are chlorophylls major components of stress biology in higher plants? Sci. Total Environ. 726, 138697. doi: 10.1016/j.scitotenv.2020.138637
Asare, M. O., Száková, J., and Tlustoš, P. (2023). The fate of secondary metabolites in plants growing on Cd −, As −, and Pb − contaminated soils — a comprehensive review. Environ. Sci. pollut. Res. 30, 11378–11398. doi: 10.1007/s11356-022-24776-x
Azmat, R., Saleem, A., Ahmed, W., Qayyum, A., El-Serehy, H. A., and Ali, S. (2022). The investigation of the impact of toxicity of metals on oxygen-evolving complex in Spinacia oleracea. Antioxidants 11, 1–12. doi: 10.3390/antiox11091802
Ballabio, C., Jones, A., and Panagos, P. (2024). Cadmium in topsoils of the European Union – An analysis based on LUCAS topsoil database. Sci. Total Environ. 912, 168710. doi: 10.1016/j.scitotenv.2023.168710
Bashir, H., Qureshi, M. I., Ibrahim, M. M., and Iqbal, M. (2015). Chloroplast and photosystems: Impact of cadmium and iron deficiency. Photosynthetica 53, 321–335. doi: 10.1007/s11099-015-0152-z
Beniušytė, E., Čėsnienė, I., Sirgedaitė-Šėžienė, V., and Vaitiekūnaitė, D. (2023). Genotype-dependent jasmonic acid effect on Pinus sylvestris L. growth and induced systemic resistance indicators. Plants 12, 255. doi: 10.3390/plants12020255
Briat, J. F., Ravet, K., Arnaud, N., Duc, C., Boucherez, J., Touraine, B., et al. (2010). New insights into ferritin synthesis and function highlight a link between iron homeostasis and oxidative stress in plants. Ann. Bot. 105, 811–822. doi: 10.1093/aob/mcp128
Čėsnienė, I., Miškelytė, D., Novickij, V., Mildažienė, V., and Sirgedaitė−Šėžienė, V. (2023). Seed treatment with electromagnetic field induces different effects on emergence, growth and profiles of biochemical compounds in seven half-sib families of silver birch. Plants 12, 3048. doi: 10.3390/plants12173048
Caretto, S., Linsalata, V., Colella, G., Mita, G., and Lattanzio, V. (2015). Carbon fluxes between primary metabolism and phenolic pathway in plant tissues under stress. Int. J. Mol. Sci. 16, 26378–26394. doi: 10.3390/ijms161125967
Chandra, S., Roychoudhury, A., Roychoudhury, A., and Tripathi, D. K. (2020). Role of selenium and manganese in mitigating oxidative damages. Prot. Chem. Agents Amelior. Plant Abiotic Stress Biochem. Mol. Perspect., 597–621. doi: 10.1002/9781119552154.ch30
Collin, S., Baskar, A., Geevarghese, D. M., Ali, M. N. V. S., Bahubali, P., Choudhary, R., et al. (2022). Bioaccumulation of lead (Pb) and its effects in plants: A review. J. Hazard. Mater. Lett. 3, 100064. doi: 10.1016/j.hazl.2022.100064
De Vries, W., Römkens, P. F. A. M., Kros, J., Voogd, J. C., and Schulte-Uebbing, L. F. (2022). “Impacts of nutrients and heavy metals in European agriculture,” in Current and critical inputs in relation to air, soil and water quality, ETC-DI, p. 72
Dixit, G., Singh, A. P., Kumar, A., Mishra, S., Dwivedi, S., Kumar, S., et al. (2016). Reduced arsenic accumulation in rice (Oryza sativa L.) shoot involves sulfur mediated improved thiol metabolism, antioxidant system and altered arsenic transporters. Plant Physiol. Biochem. 99, 86–96. doi: 10.1016/j.plaphy.2015.11.005
Du, Y., Zhao, Q., Chen, L., Yao, X., Zhang, W., Zhang, B., et al. (2020). Effect of drought stress on sugar metabolism in leaves and roots of soybean seedlings. Plant Physiol. Biochem. 146, 1–12. doi: 10.1016/j.plaphy.2019.11.003
Egendorf, S. P., Groffman, P., Moore, G., and Cheng, Z. (2020). The limits of lead (Pb) phytoextraction and possibilities of phytostabilization in contaminated soil: A critical review. Int. J. Phytoremediation 22, 916–930. doi: 10.1080/15226514.2020.1774501
Erofeeva, E. A. (2014). Hormesis and paradoxical effects of wheat seedling (Triticum aestivum L.) parameters upon exposure to different pollutants in a wide range of doses. Dose-Response 12, 121–135. doi: 10.2203/dose-response.13-017.Erofeeva
European Commission (EC) (2021). “EU soil strategy for 2030,” in Reaping the benefits of healthy soils for people, food, nature and climate (Brussels: COM). Available online at: https://ec.europa.eu/environment/publications/eu-soil-strategy-2030_en. (Accessed July 09, 2025).
Fendrich, A. N., Van Eynde, E., Stasinopoulos, D. M., Rigby, R. A., Mezquita, F. Y., and Panagos, P. (2024). Modeling arsenic in European topsoils with a coupled semiparametric (GAMLSS-RF) model for censored data. Environ. Int. 185, 108544. doi: 10.1016/j.envint.2024.108544
Finnegan, P. M. and Chen, W. (2012). Arsenic toxicity: The effects on plant metabolism, Front. Physiol 3. doi: 10.3389/fphys.2012.00182
Fritz, B. G., Appriou, D., Counts, J. R., Sample, B. E., Bunn, A. L., Dimson, J. F., et al. (2022). Effects of lead and arsenic in soils from former orchards on growth of three plant species. Environ. Toxicol. Chem. 41, 1459–1465. doi: 10.1002/etc.5325
Geiken, B., Masojídek, J., Rizzuto, M., Pompili, M. L., and Giardi, M. T. (1998). Incorporation of [35S]methionine in higher plants reveals that stimulation of the D1 reaction centre II protein turnover accompanies tolerance to heavy metal stress. Plant Cell Environ. 21, 1265–1273. doi: 10.1046/j.1365-3040.1998.00361.x
Gill, S. S. and Tuteja, N. (2010). Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 48, 909–930. doi: 10.1016/j.plaphy.2010.08.016
Guilayn, F., Benbrahim, M., Rouez, M., Crest, M., Patureau, D., and Jimenez, J. (2020). Humic-like substances extracted from different digestates: First trials of lettuce biostimulation in hydroponic culture. Waste Manage. 104, 239–245. doi: 10.1016/j.wasman.2020.01.025
Jańczak-Pieniążek, M., Cichoński, J., Michalik, P., and Chrzanowski, G. (2023). Effect of heavy metal stress on phenolic compounds accumulation in winter wheat plants. Molecules 28. doi: 10.3390/molecules28010241
January, M. C., Cutright, T. J., Van Keulen, H., and Wei, R. (2008). Hydroponic phytoremediation of Cd, Cr, Ni, As, and Fe: Can Helianthus annuus hyperaccumulate multiple heavy metals? Chemosphere 70, 531–537. doi: 10.1016/j.chemosphere.2007.06.066
Kniuipytė, I., Dikšaitytė, A., Praspaliauskas, M., Pedišius, N., and Žaltauskaitė, J. (2023). Oilseed rape (Brassica napus L.) potential to remediate Cd contaminated soil under different soil water content. J. Environ. Manage. 325, 116627. doi: 10.1016/j.jenvman.2022.116627
Kong, X., Zhao, Y., and Tian, K. (2020). Insight into nitrogen and phosphorus enrichment on cadmium phytoextraction of hydroponically grown Salix matSudana Koidz cuttings. Environ. Sci. Pollut. Res. 27, 8406–8417. doi: 10.1007/s11356-019-07499-4
Konlechner, C., Türktas, M., Langer, I., Vaculík, M., Wenzel, W. W., Puschenreiter, M., et al. (2013). Expression of zinc and cadmium responsive genes in leaves of willow (Salix caprea L.) genotypes with different accumulation characteristics. Environ. pollut. 178, 121–127. doi: 10.1016/j.envpol.2013.02.033
Kumar, A., Tripti, D., Maiti, S. K., Maleva, M., and Borisova, G. (2022). Soil pollution and plant efficiency indices for phytoremediation of heavy metal(loid)s: two-decade study (2002–2021). Metals (Basel). 12, 1–36. doi: 10.3390/met12081330
Lee, H. Y. and Back, K. (2017). Cadmium disrupts subcellular organelles, including chloroplasts, resulting in melatonin induction in plants. Molecules 22. doi: 10.3390/molecules22101791
Małkowski, E., Sitko, K., Szopiński, M., Gieroń, Ż., Pogrzeba, M., Kalaji, H. M., et al. (2020). Hormesis in plants: The role of oxidative stress, auxins and photosynthesis in corn treated with Cd or Pb. Int. J. Mol. Sci. 21, 2099. doi: 10.3390/ijms21062099
Murphy, A. S., Eisinger, W. R., Shaff, J. E., Kochian, L. V., and Taiz, L. (1999). Early copper-induced leakage of K+ from Arabidopsis seedlings is mediated by ion channels and coupled to citrate efflux. Plant Physiol. 121, 1375–1382. doi: 10.1104/pp.121.4.1375
Muszyńska, E. and Labudda, M. (2019). Dual role of metallic trace elements in stress biology—from negative to beneficial impact on plants, Int. J. Mol. Sci. 20, 3117. doi: 10.3390/ijms20133117
Nahvi, H., Torabian, S., Hashemi, S. A., and Payam, H. (2024). Alnus glutinosa (Alder) sapling as a phytoremediator for cadmium in contaminated soil of industrial park. Results Eng. 22, 102317. doi: 10.1016/j.rineng.2024.102317
Page, V. and Feller, U. (2015). Heavy metals in crop plants: Transport and redistribution processes on the whole plant level. Agronomy 5, 447–463. doi: 10.3390/agronomy5030447
Pang, X., Wang, D., Xing, X., Peng, A., Zhang, F., and Li, C. (2002). Effect of La3+ on activities of antioxidant enzymes in wheat seedlings under lead stress in solution culture. Chemosphere 47, 1033–1039. doi: 10.1016/S0045-6535(02)00022-X
Paradiso, A., Berardino, R., De Pinto, M. C., Sanità Di Toppi, L., Storelli, M. M., Tommasi, F., et al. (2008). Increase in ascorbate-glutathione metabolism as local and precocious systemic responses induced by cadmium in durum wheat plants. Plant Cell Physiol. 49, 362–374. doi: 10.1093/pcp/pcn013
Perfus-Barbeoch, L., Leonhardt, N., Vavasseur, A., and Forestier, C. (2002). Heavy metal toxicity: Cadmium permeates through calcium channels and disturbs the plant water status. Plant J. 32, 539–548. doi: 10.1046/j.1365-313X.2002.01442.x
Pietrini, F., Zacchini, M., Iori, V., Pietrosanti, L., Bianconi, D., and Massacci, A. (2010). Screening of poplar clones for cadmium phytoremediation using photosynthesis, biomass and cadmium content analyses. Int. J. Phytoremediation 12, 105–120. doi: 10.1080/15226510902767163
Pongrac, P., Brvar, E., and Regvar, M. (2009). Impact of simultaneous Cd and Zn substrate amendments on metal accumulation in two Cd/Zn hyperaccumulating Thlaspi species. Acta Biol. Slov. 52, 61–71. doi: 10.14720/abs.52.2.15374
Pulford, I. D. and Watson, C. (2003). Phytoremediation of heavy metal-contaminated land by trees — a review. Environ. Int. 29, 529–540. doi: 10.1016/S0160-4120(02)00152-6
Rascio, N. and Navari-Izzo, F. (2011). Heavy metal hyperaccumulating plants: How and why do they do it? And what makes them so interesting? Plant Sci. 180, 169–181. doi: 10.1016/j.plantsci.2010.08.016
Reddy, C. N. and Patrick, W. H. (1977). Effect of Redox potential and pH on the uptake of cadmium and lead by rice plants. J. Environ. Qual 6, 259–262. doi: 10.2134/jeq1977.00472425000600030005x
Reeves, R. D., Baker, A. J. M., Jaffré, T., Erskine, P. D., Echevarria, G., and van der Ent, A. (2017). A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 218, 407–411. doi: 10.1111/nph.14907
Riyazuddin, R., Nisha, N., Ejaz, B., Khan, M. I. R., Kumar, M., Ramteke, P. W., et al. (2022). A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 12, 1–26. doi: 10.3390/biom12010043
Rosa, M., Prado, C., Podazza, G., Interdonato, R., González, J. A., Hilal, M., et al. (2009). Soluble sugars-metabolism, sensing and abiotic stress a complex network in the life of plants. Plant Signal. Behav. 4, 388–393. doi: 10.4161/psb.4.5.8294
Salam, M. M. A., Ruhui, W., Sinkkonen, A., Pappinen, A., and Pulkkinen, P. (2022). Effects of contaminated soil on the survival and growth performance of European (Populus tremula L.) and Hybrid Aspen (Populus tremula L. × Populus tremuloides Michx.) clones based on stand density. Plants 11, 1970. doi: 10.3390/plants11151970
Sarangthem, J., Jain, M., and Gadre, R. (2011). Inhibition of δ-aminolevulinic acid dehydratase activity by cadmium in excised etiolated maize leaf segments during greening. Plant Soil Environ. 57, 332–337. doi: 10.17221/45/2011-pse
Schützendübel, A., Schwanz, P., Teichmann, T., Gross, K., Langenfeld-Heyser, R., Godbold, D. L., et al. (2001). Cadmium-induced changes in antioxidative systems, hydrogen peroxide content, and differentiation in Scots pine roots. Plant Physiol. 127, 887–898. doi: 10.1104/pp.010318
Sharma, J. K., Kumar, N., Singh, N. P., and Santal, A. R. (2023). Phytoremediation technologies and their mechanism for removal of heavy metal from contaminated soil: An approach for a sustainable environment. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1076876
Sirgedaitė−Šėžienė, V., Vaitiekūnaitė, D., Šilanskienė, M., Čėsnienė, I., Striganavičiūtė, G., and Jaeger, A. (2024). Assessing the phytotoxic effects of class A foam application on northern Europe tree species: A short-term study on implications for wildfire management and ecosystem health. Ecol. Indic 166, 112393. doi: 10.1016/j.ecolind.2024.112393
Stobart, A. K., Griffiths, W. T., Ameen-Bukthari, I., and Sherwood, R. P. (1985). The effect of Cd2+ on the biosynthesis of chlorophyll in leaves of barley. Physiol. Plant 63, 293–298. doi: 10.1111/j.1399-3054.1985.tb04268.x
Sun, Y., Zhou, Q., Wang, L., and Liu, W. (2009). Cadmium tolerance and accumulation characteristics of Bidens pilosa L. as a potential Cd-hyperaccumulator. J. Hazard. Mater 161, 808–814. doi: 10.1016/j.jhazmat.2008.04.030
Veltrup, W. (1981). Effect of heavy metals on the calciuyl absorption by intact barley roots. J. Plant Nutr. 3, 225–231. doi: 10.1080/01904168109362831
Wang, X., Ze, Y., Wu, X., Chen, L., Huang, H., Liu, J., et al. (2009). Effect of Pb2+ on the kinetic and spectral characterization of ribulose-1,5-bisphosphate carboxylase/oxygenase. Chin. J. Chem. 27, 727–731. doi: 10.1002/cjoc.200990120
Widjaja, H., Rianto, I. A. D., Trisasongko, B. H., and Randrikasari, O. (2024). Modelling chlorophyll and nutrient contents of Peperomia obtusifolia “Green Gold” Using MAPIR RGN and RGB sensors. IOP Conf. Ser. Earth Environ. Sci. 1359, 012003. doi: 10.1088/1755-1315/1359/1/012003
Xu, X., Yang, B., Qin, G., Wang, H., Zhu, Y., Zhang, K., et al. (2019). Growth, accumulation, and antioxidative responses of two Salix genotypes exposed to cadmium and lead in hydroponic culture. Environ. Sci. pollut. Res. 26, 19770–19784. doi: 10.1007/s11356-019-05331-7
Yongpisanphop, J., Babel, S., Kruatrachue, M., and Pokethitiyook, P. (2017). Hydroponic screening of fast-growing tree species for lead phytoremediation potential, Bull. Environ. Contam. Toxicol. 99, 518–523. doi: 10.1007/s00128-017-2157-8
Keywords: aspen, willow, heavy metals, hydroponics, hormesis
Citation: Sirgedaitė-Šėžienė V, Striganavičiūtė G, Šilanskienė M, Kniuipytė I, Praspaliauskas M, Vaškevičienė I, Lemanas E and Vaitiekūnaitė D (2025) Evaluating Populus tremula L. and Salix caprea L. for phytoremediation: growth, metal uptake, and biochemical responses under arsenic, cadmium, and lead stress. Front. Plant Sci. 16:1617432. doi: 10.3389/fpls.2025.1617432
Received: 24 April 2025; Accepted: 18 July 2025;
Published: 01 August 2025.
Edited by:
Aamir Hamid Khan, University of Łódź, PolandReviewed by:
Rui Li, Chongqing University of Science and Technology, ChinaPadma Sharma, Panjab University, India
Amal Bouzidi, UMR6283 Institut des Molécules et Matériaux du Mans (IMMM), France
Copyright © 2025 Sirgedaitė-Šėžienė, Striganavičiūtė, Šilanskienė, Kniuipytė, Praspaliauskas, Vaškevičienė, Lemanas and Vaitiekūnaitė. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dorotėja Vaitiekūnaitė, ZG9yb3RlamEudmFpdGlla3VuYWl0ZUBsYW1tYy5sdA==