 Giuliano Bonanomi1,2
Giuliano Bonanomi1,2 Adriano Stinca3Giandomenico Amoroso1Emilia Allevato4
Adriano Stinca3Giandomenico Amoroso1Emilia Allevato4 Giuseppina Iacomino1
Giuseppina Iacomino1 Gianluigi Mauriello1
Gianluigi Mauriello1 Riccardo Motti1Alfredo Nicastri2Francesca Bellucci5Mohamed Rida Abelouah6
Riccardo Motti1Alfredo Nicastri2Francesca Bellucci5Mohamed Rida Abelouah6 Luigi Di Costanzo1
Luigi Di Costanzo1 Mohamed Idbella7*
Mohamed Idbella7*- 1Department of Agricultural Sciences, University of Naples Federico II, Portici, Italy
- 2Club Alpino Italiano, Presidente Comitato Scientifico Regionale and Sezione di Salerno, Salerno, Italy
- 3Department of Environmental, Biological and Pharmaceutical Sciences and Technologies, University of Campania Luigi Vanvitelli, Caserta, Italy
- 4Department of Biology, University of Naples “Federico II”, Naples, Italy
- 5Club Alpino Italiano, Presidente Gruppo Regionale and Sezione di Avellino, Avellino, Italy
- 6Laboratory of Aquatic Systems: Marine and Continental Environments (AQUAMAR), Faculty of Sciences, IbnZohr University, Agadir, Morocco
- 7AgroBioSciences (AgBS) Program, College of Agriculture and Environmental Sciences, Mohammed VI Polytechnic University, Ben Guerir, Morocco
Introduction: The treeline elevation in the Apennines is significantly lower than its climatic potential, often attributed to historical anthropogenic disturbances such as fires, logging, and grazing. However, the specific impacts of individual disturbance events, particularly fires, on treeline dynamics remain unclear. This study investigates the relationship between treeline elevation and historical fire events using Monte Cervati (1,899 m a.s.l.) as a model system.
Location: Monte Cervati, located in the Apennine Mountains, Italy.
Methods: The current treeline elevation was mapped, and vegetation was characterized. Pedo-anthracological analyses were conducted in a sinkhole near the summit to reconstruct past vegetation and fire history. Charcoal samples from a paleosol layer were identified and dated to determine past fire events and vegetation composition.
Results: The present treeline, composed exclusively of Fagus sylvatica, is situated at an average elevation of 1,710 m a.s.l., with higher elevations on northern slopes compared to southern ones. The vegetation above the treeline is dominated by small herbaceous species (Plantago subulata), with scattered shrubs such as Daphne oleoides and Juniperus communis. Notably, no F. sylvatica regeneration was observed above the treeline. Pedo-anthracological analysis revealed a charcoal-rich layer within a paleosol at 1,806 m a.s.l., dating back to approximately 4,800 BP. Charcoal analysis identified remains from herbaceous (Dactylis), shrubs (Daphne), and trees (F. sylvatica), indicating that past vegetation consisted of wooded grassland.
Conclusions: Historical fire events likely played a crucial role in shaping the current treeline structure by eliminating the arboreal component and promoting the establishment of a predominantly herbaceous landscape. These findings suggest that fire disturbances have long-term effects on treeline dynamics, potentially contributing to the treeline depression observed in the Apennines today.
1 Introduction
The upper limit of the closed forest is a key landscape feature that characterizes most of the mountain ranges worldwide that reach sufficient elevation. The trees that reach the highest elevation are found in the Himalayas where Juniperus tibetica Kom. forest reaches 4,900 m a.s.l (Miehe et al., 2007). In the Alps, some conifers including the Swiss pine (Pinus cembra L.) and the larch (Larix deciduas Mill.), can reach 2200–2300 meters of elevation (Körner and Paulsen, 2004). The upper limit of the forest is undoubtedly the result of the complex interplay of temperature on the physiology of trees: the lower the temperature, the lower the elevation of the treeline. Ideally moving from the equator toward the poles, the elevation of the upper treeline progressively decreases from about 4,000 m a.s.l. in the tropics to sea level at the highest latitudes of Norway and Canada in the northern hemisphere and Tierra del Fuego in the southern hemisphere.
The upper treeline is commonly associated with decreasing air and soil temperature as elevation increases. Early studies claimed that minimum winter temperatures limited tree growth but subsequent studies have identified an isotherm of approximately 6.7° that characterizes the upper treeline on a global scale (Körner, 2012). This thermal limit is manifested on mountain slopes, as an imaginary boundary that trees cannot cross. When temperatures fall below this threshold, only prostrate shrubs, cushion plants and some herbs, mostly perennial scans complete their life cycle. The limit of ~6.7°C is not arbitrary but has deep eco-physiological foundations. A few decades ago, it was thought that low temperatures limited the process of chlorophyll photosynthesis, but later it was widely demonstrated that trees living at the treeline can efficiently perform this process down to temperatures close to 0°C (Körner, 1998), as long as the water in the soil remains unfrozen. In fact low temperatures, below ~6.7°C, strongly slow down cell division, distension and elongation (Körner, 2012).
While the eco-physiological factors like temperature and moisture availability (Sigdel et al., 2018; Bailey et al., 2021) determining the altitudinal position of the treeline are well understood on a global scale, various disturbances can significantly influence it at regional and local level, altering the maximum elevation at which forest formations are observed. Avalanches (Walsh et al., 1994), biotic interactions (Liang et al., 2016; Sigdel et al., 2024) in addition to grazing and logging are the factors that tend to lower, therefore to “depress” the treeline elevation relative to the potential climatic limit of a given species (Körner, 1998; Harsch et al., 2009). On the European continent, human activity over thousands of years has radically altered the vegetation in hilly and mountainous regions of the Alps, the Pyrenees, the Carpathians, and the Spanish mountain ranges (Colombaroli et al., 2010). This human activity has been particularly intense and pervasive on the Apennines (Bonanomi et al., 2020).
Stretching over 1,000 km along peninsular Italy, the Apennines form a mountain range where Fagus sylvatica is the dominant species at the upper limit of the forest, an environment that in a few cases is shared with the Bosnian pine (Pinus heldreichii Christ) on the Pollino massif and with the mountain pine (Pinus mugo Turra) on the Majella. F. sylvatica is an Eurasiatic deciduous species with the widest range among European trees, spanning from the Scandinavian Peninsula and the British Isles to Central and Mediterranean Europe. It is a tree that suffers heat and summer drought lacking specific fire-adaptive traits (Stinca et al., 2021). Therefore, in the Mediterranean basin it finds refuge in mountain areas, taking advantage of optimal climatic conditions such as cool and rainy summers. A study by our research group has quantified the maximum elevation reached by beech on 302 Apennine peaks covering a total of 3,622 km of treeline (Bonanomi et al., 2018). On average, beeches have their elevation limit at 1,589 m a.s.l., although there is considerable variability across different Apennine Mountain ranges. This study also reveals that in Pollino Park we find continental beeches that grow at the highest elevation in the world, reaching 2,140 m a.s.l. on the slopes of Monte Serra del Prete. Throughout the Apennines, the millenary action of human being, aimed at opening clearings in the forest to favor cultivation and grazing or to obtain timber or produce charcoal, has led to a drastic lowering of the maximum elevation reached by the forest (Bonanomi et al., 2020).
In the last decades, the study of fossil pollen gradually accumulated in the lakes and in the few Apennines peat bogs have allowed us a detailed reconstruction of the alternation of the main plant species during the glaciation and, more recently, in the Holocene. In particular, pollen analysis has provided valuable insights into the response of beech to the alternation of glaciations and to human activity at both continental and regional scales (Watts, 1985; Magri et al., 2006). However, only a few studies are available on pollen profiles at high elevation in the Apennines. For example, Magri (2007) demonstrated that, at Campo Felice, a site located at ~1500 m a.s.l., in the last 90,000 years the main tree taxa (e.g. Abies, Fagus, Pinus, Quercus) have alternated with expansions and subsequent contractions of population. The use of pollen as a paleo-vegetation proxy, however, also has its limitations. Pollen analysis depends on anoxic conditions for preservation, limiting its presence to environments like lakes, peat bogs, and wet sediments. Moreover, pollen is dispersed by the wind over long distances, often tens or even hundreds of kilometers. As a result, this method reflects vegetation changes on a regional or district scale but cannot provide detailed information on local events, such as shifts in the treeline. In contrast, detailed spatial information that cannot be obtained through pollen analysis can be gathered from the study of fragments of carbonized wood. Charred remains from the past periods could be preserved in the soil horizons and their interpretation allows us to reveal part of the history of the former vegetation with great spatial precision. Furthermore, charcoal being chemically inert is preserved in the soil for several millennia (Carcaillet and Thinon, 1996; Talon et al., 1998; Mercuri et al., 2015). Overall, soil charcoal analysis, or pedo-anthracology, combined with radiocarbon dating (14C) of charred wood fragments, is an effective method for reconstructing past forest dynamics with high spatial resolution (Thinon, 1978). It provides coupled localized insights into fire history and past vegetation composition, and, most importantly, can offer robust and detailed information on past altitudinal treeline shifts, shedding light on the changes of this critical ecological boundary (Carcaillet and Thinon, 1996). For example, distinct black layers in soil profile rich of charcoal residues have recently been successfully studied for vegetation reconstruction in New Zealand (McWethy et al., 2010). Unfortunately, in the Apennine the information available to date is limited and fragmentary and does not allow us to provide an overall picture of the changes that have occurred in the high-altitude forests (Benatti et al., 2019).
In this context, the aim of this study is to investigate the distribution of Fagus sylvatica on Mount Cervati, as a representative model of the Southern Apennines because of the carbonate substrate and the depressed treeline in term of elevation compared to the climatic potential. The study first quantified the distribution of the current treeline as well as its current dynamics of regeneration and recolonization of the summit areas above the treeline. In addition, we studied a distinct black layer present in a sinkhole located close to the mountain peak (Figure 1). We hypothesized that such a black layer represents the traces of ancient fires that would transform the vegetation of the summit of Cervati. Consequently, we used the pedo-anthracological approach based on optical and scanning electron microscopy and energy dispersive spectroscopy (SEM–EDS) for taxonomic identification of the charcoal samples and to evaluate the long-term alterations of wood buried in the soil. Radiocarbon dating of charcoal samples was conducted to determine the period in which historical fires occurred. The specific hypotheses tested were as follows:
i. The F. sylvatica treeline is depressed in terms of elevation compared to the species’ potential.
ii. F. sylvatica is unable to recolonize open areas above the treeline.
iii. Past historical fires may have contributed to depressing the upper limit of the treeline.

Figure 1. Location of Cervati massif in Southern Italy (A); Fagus sylvatica treeline on the South slope at ~1,700 m a.s.l., with the transition between forest and the rocky grassland (B), view of the summit sinkhole whose bottom is at an elevation of 1806 m a.s.l., (black arrow - C), and detail of the exposed profile of the sinkhole with the black layer located at about 150 cm depth (yellow arrow - D).
2 Materials and methods
2.1 Study sites description
With 261 peaks rising above 2,000 m a.s.l., the 1,200 km long Apennine mountain range spans 38°N to 44°N. Pinus nigra J. F. Arnold subsp. nigra and Pinus heldreichii Christ subsp. Leucodermis (Antoine) E. Murray are rare and remnant plant populations are found in the middle and southern regions of the Apennines, respectively, while F. sylvatica dominates the treeline along the chain (Bonanomi et al., 2018). Limestone makes up most of the substrate, with sporadic arenaceous-peliticflysch in the Laga groups (https://www.pcn.minambiente.it/GN). With mean temperatures between 0 and 11°C in January and 24 and 28°C in July, the climate is a mountainous variation of the Mediterranean type. Total annual precipitation varies between 800 and 3,000 mm, with regular winter snowfall occurrences above 1,000 m a.s.l.
The present study was carried in the Mt. Cervati located in the National Park of Cilento, Vallo di Diano and Alburni, southern Italy (40°17’8.97”N, 15°29’11.18”E; 1,899 m a.s.l.). Study site also fall within the Site of Community Importance IT8050024 “Monte Cervati, Centaurino e Montagne di Laurino”, and Special Area of Conservation IT8050046 “Monte Cervati e dintorni”. The massif is composed of a mainly carbonates substrate with intense karst activity, ensuring a year-round water supply to neighboring territories. At lower elevations (0–800 m a.s.l.) annual precipitation averages are around 800 mm, while at higher elevations rainfall exceeds 1000 mm. The natural vegetation of the area is characterized by different forest coenosis that cover all slopes of the massif, while in the summit area there are rocky meadows often used for grazing. The forests, especially at lower elevations, are often coppice and composed of many broadleaf trees such as Castanea sativa Mill., Quercus cerris L., Quercus pubescens Willd. subsp. pubescens, Acer opalus Mill. subsp. obtusatum (Waldst. & Kit. ex Willd.) Gams, Ulmus minor Mill. subsp. minor, Ostrya carpinifolia Scop., etc. Beech forests appear from about 900 m a.s.l. and, in addition to F. sylvatica, only rarely show other tree species (e.g. Taxus baccata L., Ilex aquifolium L., etc.). This vegetation type can be attributed to the habitat “Apennine beech forests with Taxus and Ilex” (code 9210*, Habitats Directive 92/43/EEC), widespread in the Italian Peninsula and Sicily in the supratemperate bioclimatic plan and rarely in the upper mesotemperate (Zitti et al., 2016). In the plant communities of the summit area of the massif, perennial herbaceous species are very abundant, many of which are endemic and rare (e.g. Potentilla rigoana Th. Wolf, Cynoglossum magellense Ten., Pedicularis elegans Ten., Gentianella columnae (Ten.) Holub, Saxifraga exarataVill. subsp. ampullacea (Ten.) D.A. Webb (Santangelo et al., 1994). (Figure 1). Also noteworthy are the shrublands with Lavandula austroapennina N.G. Passal., Tundis & Upson which can be observed on some rocky slopes of the massif. This species is a recently described endemic of the southern Apennines (Gravina et al., 2023).
2.2 Current treeline assessment
Treeline assessment was conducted for Mt. Cervati following the approach of Bonanomi et al. (2018), that recorded treeline elevation of all peaks with an elevation above 1,500 m a.s.l. across the Apennines. In this study treeline elevation was carefully mapped using the tool Google Earth Pro™ (Google, Inc. Mountain View, CA, USA) using images spanning from 2004 to 2018. The treeline elevation was recorded by manually delineating the boundary between forest and grassland on the four different aspects (north, east, south and west) of mountain. Afterward, the digitized drawn lines were carefully measured for their maximum, average, and minimum elevations above sea level. The treeline elevation measured with satellite images was compared with ground measurements using GPS (Garmin Montana 750i) at forty points in total, ten for each exposure.
2.3 Current vegetation beyond the treeline and Fagus sylvatica regeneration
The current natural vegetation of the summit study site was analyzed through field surveys conducted in June 2024, followed by detailed taxonomic analyses in the laboratory. Sampling was performed in eight 1 × 1 m plots (i.e. 4 on the sinkhole and 4 on the surrounding slopes) recording all vascular taxa and visually estimating their absolute percentage cover. The taxonomic identification was carried out using Pignatti et al. (2017a, b, 2018, 2019), while nomenclature and taxa delimitation followed the checklist of Italian vascular flora (Bartolucci et al., 2024). The collected plant material was stored in the Herbarium Austroitalicum (IT).
The surveys concerning F. sylvatica regeneration was conducted at the treeline during the summer of 2024. Four rectangular transects (10 × 200 m), or belt transects, was established for each aspect for a total of sixteen transects. The transects were spaced at least 200 meters from the treeline to open grassland. First, we measured the overall height of F. sylvatica at the treeline using a telescopic graduation meter. In each transect, all beech seedlings and samplings were counted and the distance from the treeline was measured. In addition, the total cover (%) of shrubs, herbaceous species and outcrops was quantified in each transect.
2.4 Soil sampling in the sinkhole, anthracological analysis and radiocarbon dating
In June 2022, sampling activities were conducted in the summit sinkhole area, located at an elevation of 1,816 m a.s.l. (40°17’8.97”N, 15°29’11.18”E), just below the mountain peak (Figure 1). To investigate Holocene vegetation and fire history, we applied a soil charcoal analysis approach (Carcaillet and Thinon, 1996; Robin and Nelle, 2014). Soil samples were collected along an exposed profile of the sinkhole at four depths: 50 cm, 100 cm, 150 cm at the black layer suspected of being rich in charcoal residue due to its dark color, and near the bottom at 200 cm depth. Approximately 10 kg of soil was collected for each depth (Figure 2). The soil samples were placed in polyethylene bags and transported to the laboratories of the Department of Agriculture, where it was air-dried and then stored at room temperature.
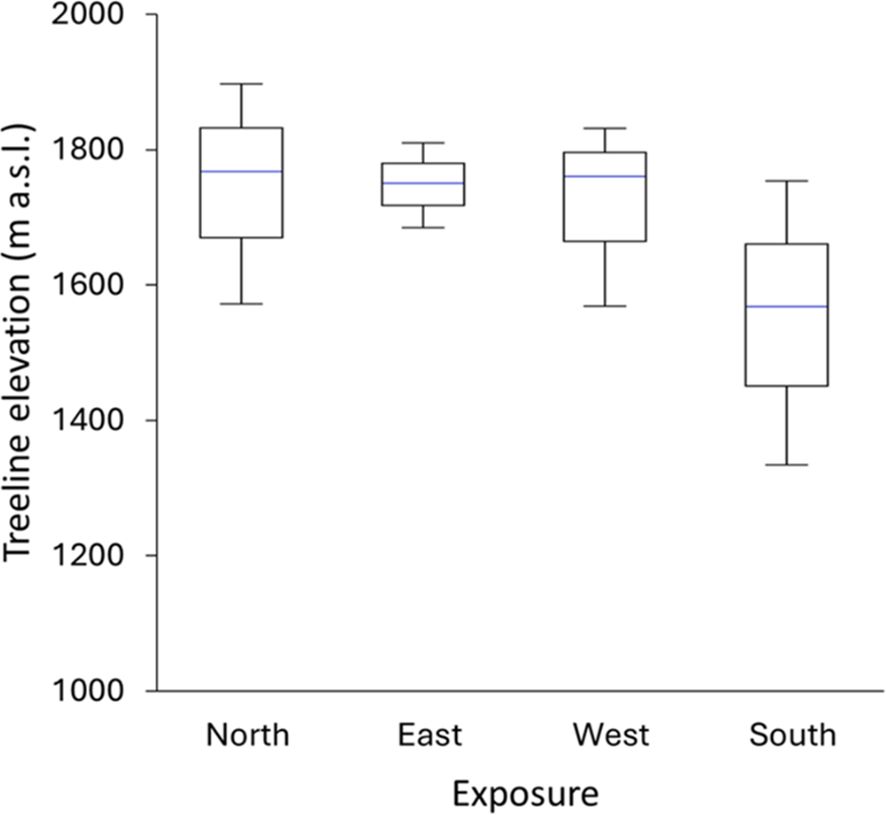
Figure 2. Elevation of F. sylvatica treeline according to exposition; each box represents the average value; upper and lower whiskers reach are minimum and maximum values observed.
The soil was subsequently washed with water over a column of sieves with calibrated mesh sizes of 4 mm, 2 mm, 1 mm, and 0.5 mm to remove fine sediment. Charcoal fragments were extracted under a binocular microscope and stored for further analysis. Identification of the charred wood fragments was performed by examining the three anatomical planes of the wood under a reflected light microscope, with magnifications ranging from 50x to 1000x. The identification was based on a comparison with wood anatomy atlases (Greguss, 1959; Schweingruber, 1990) and electronic identification keys (Heiss, 2002; Dallwitz et al., 1995; InsideWood, 2004-present). The Anthracological approach is often criticized because wind can carry charcoal particles from fires occurring at lower altitudes up to the treeline and beyond. However, experimental studies and trap-based observations summarized in Whitlock and Larsen (2001) have shown that macroscopic charcoal (>125–250 µm) is not transported far from the fire margin and typically deposited within a few meters from the fire source.
2.5 Radiocarbon dating
Three charcoal fragments previously identified as Dactylis were selected for radiocarbon dating at the laboratories of the Department of Mathematics and Physics, University of Campania “Luigi Vanvitelli.” We selected only charcoal fragments from herbaceous species to minimize potential discrepancies between the plant’s lifespan and the fire event, thereby avoiding the well-known “old wood” effect—where dating the inner part of a large, long-lived tree reflects the year that specific part of the wood was formed, rather than the year the tree actually burned (Geib, 2008).
Conventional 14C ages were calibrated using the OxCal 4.4 [v. 175] online program (Ramsey, 2009) and the IntCal20 calibration curve data (Reimer et al., 2020). To better constrain the burning event(s), the 14C dates were tentatively pooled into different groupings using the R_Combine function of OxCal, which statistically merges dates assumed to derive from the same event and applies a χ² test to evaluate their internal consistency, in order to verify the assumption that the burning corresponds to a single date or to multiple distinct events (Ramsey, 2009).
2.6 Charcoal scanning electron microscopy and energy dispersive spectroscopy
We employed scanning electron microscopy and energy dispersive spectroscopy (SEM–EDS) to analyze Fagus sylvatica wood and Dactylis spp. herbs. Samples, approximately 1 × 0.5 × 0.3 cm in dimension, were attached to SEM stubs using conductive carbon paint with double-sided adhesive tape. To achieve topographical mapping of the chemical elements within these samples, SEM imaging was conducted in tandem with energy-dispersive X-ray spectroscopy (EDS). EDS utilizes a focused beam of accelerated electrons to induce fluorescence emissions from the elements present in the samples. The energy of these emitted X-rays is unique to each element, enabling identification and analysis of the chemical composition within the outermost layers (spanning a depth of several hundred nanometers to approximately 2 μm) of the Fagus sylvatica wood and Dactylis spp. herbs. A field emission scanning electron microscope (SEM, FEI Nova NanoSEM 450) installed at the Centro di Microscopia within the Dipartimento di Scienze Chimiche, University of Napoli Federico II, was used to study the samples’ morphology. The images were acquired by collecting secondary electrons (SE) with a Bruker ETD detector, all operating at room temperature. The imaging parameters included a working distance of 4.9 mm and an acceleration voltage of 3.00 kV. Analysis of the SEM images was performed using the AZtecCrystal software package, which facilitates the processing of data obtained via electron backscatter diffraction (EBSD) (Wilkinson and Britton, 2012).
3 Results
3.1 Current treeline elevation
Overall, current mean treeline elevation in Cervati was 1,710 m a.s.l., although considerable variability among exposure was found with the highest values in northern and lowest in southern expositions (1,743 and 1,554 m a.s.l., respectively) (Figure 2). The absolute maximum treeline elevation was 1,891 m a.s.l., just few meter below the top of the mountain (i.e. 1,899 m a.s.l.), with the lowest values of only 1,342 m a.s.l. on the southern exposition. Finally, we found a notable Δ maximum – minim treeline elevation of 276 m, with a highest Δ for southern exposition (409 m) compared to eastern (122 m), western (256 m) and norther (317 m) expositions.
3.2 Current vegetation beyond the treeline and Fagus sylvatica regeneration
Our field surveys carried out above the treeline have highlighted the current presence of two vegetation types related to substrate conditions (Table 1). The periodically flooded and almost flat soil of the sinkhole shows a community with a low number of vascular species (average species richness per m2 of 5.25 ± 1.23 standard deviation) and dominated by Plantago subulata (average cover 58.75%). Instead, the rocky slopes surrounding the sinkhole are characterized by more biodiverse cenoses (average species richness per m2 of 9.25 ± 1.98 standard deviation) with a prevalence of Globularia cordifolia L. subsp. Bellidifolia (Nyman) Wettst. (average cover 41.25%). Both vegetation types are heliophilous, composed mostly of hemicryptophytes, and species of conservation interest (i.e. Italian endemic species such as Achillea tenorei Grande, Potentilla calabra Ten., and Potentilla rigoana Th. Wolf), and intensively grazed by horses and cattle.
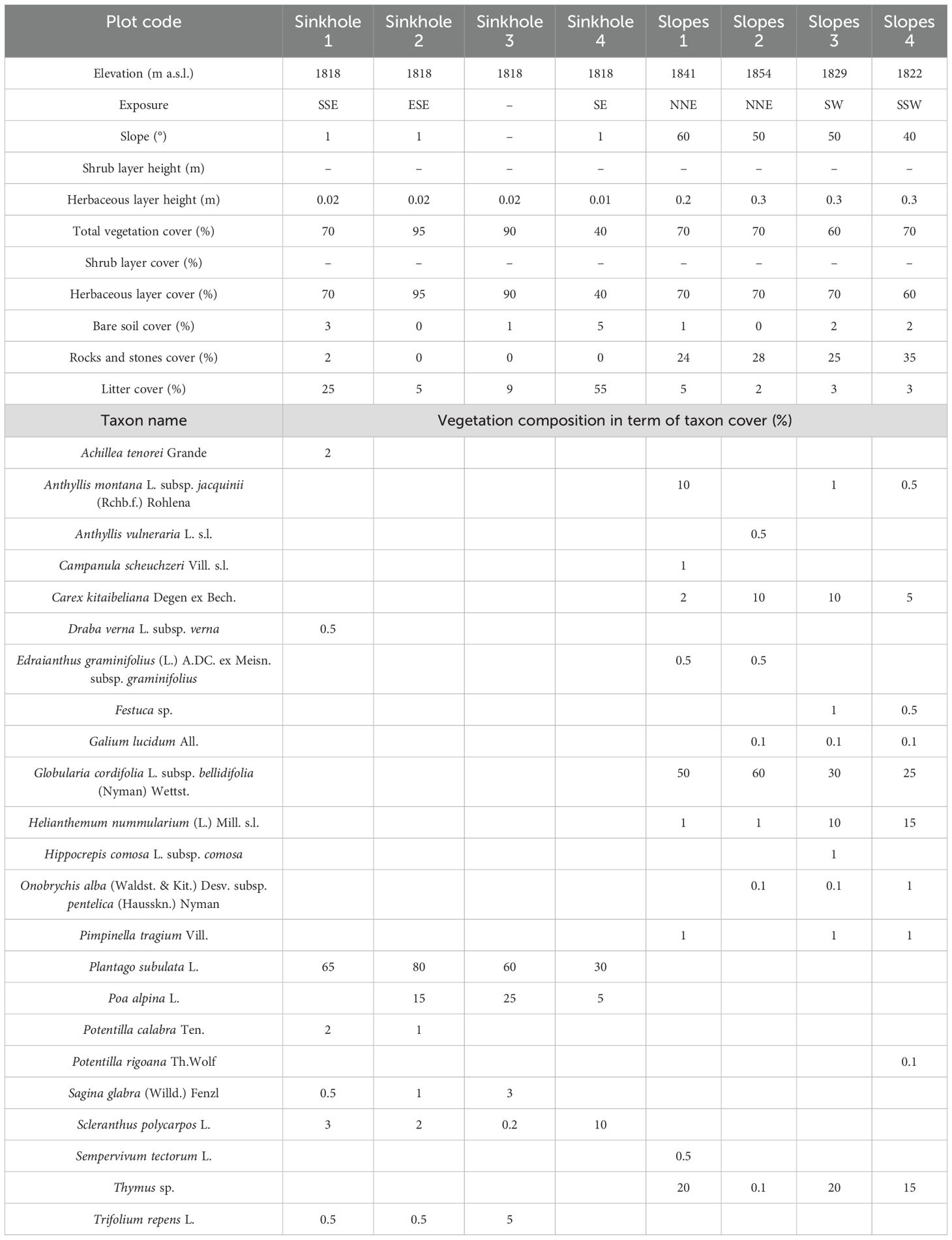
Table 1. Vegetation report of sinkhole and slope plant communities from Mt. Cervati.
Above treeline, shrub cover was very low (<5%) with scattered Juniperus communis L. And Daphne oleoides Schreb. subsp. oleoides. Concerning F. sylvatica regeneration, seedlings were completely absent above the treeline irrespectively of elevation and exposure of the transect. In fact, despite intensive searches in the transects arranged on the four sides of the mountain, no F. sylvatica seedlings or saplings were observed beyond the treeline.
3.3 Charcoal dating and identification
Concerning charcoal dating, the three single-fragment charcoal samples from the black layer, gave calendar year dates, after calibration at 2σ intervals, ranging from 4845 to 4529 cal BP (Table 2, Figure 3). The results obtained for all dated samples could not be combined, as the χ² test failed [χ² test: df = 2, p = 7.8 (5%: 6.0)]. However, when sample b was combined individually with sample a and then with sample c, good statistical significance was achieved [respectively: χ² test, df = 1, p = 0.2 (5%: 8.8) and χ² test, df = 1, p = 3.3 (5%: 3.8)]. This resulted in two combined date ranges: 4951 to 4653 (median = 4841) and 4831 to 4581 (median = 4710), both with a 95.4% probability (Figure 4).
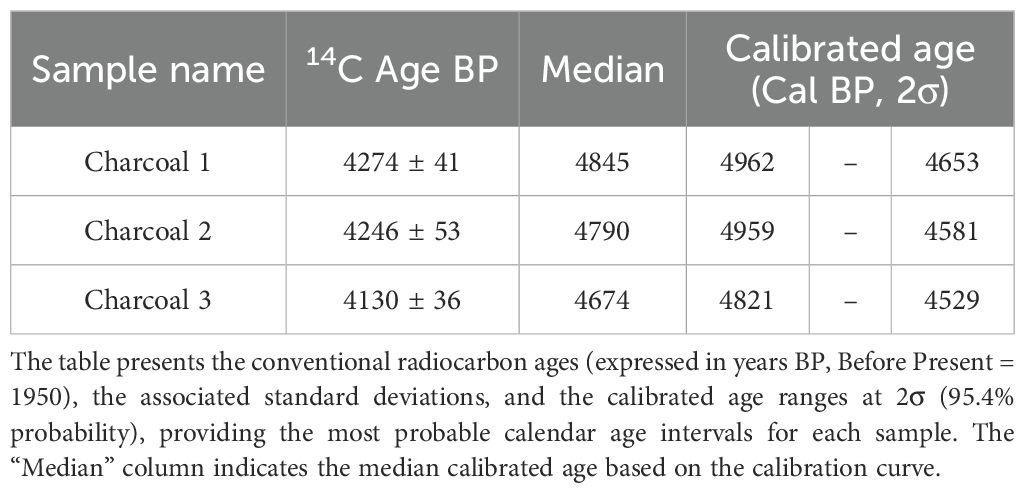
Table 2. Radiocarbon dating results of the three charcoal samples retrieved from the soil profile close the summit of Mt. Cervati.

Figure 3. Soil profile in the sink hole with the black layer at ~150 cm depth, note the ice axe for scale (A), detail of sampling point (B), and multi-plot of calibrated date of the three charcoal samples (C).
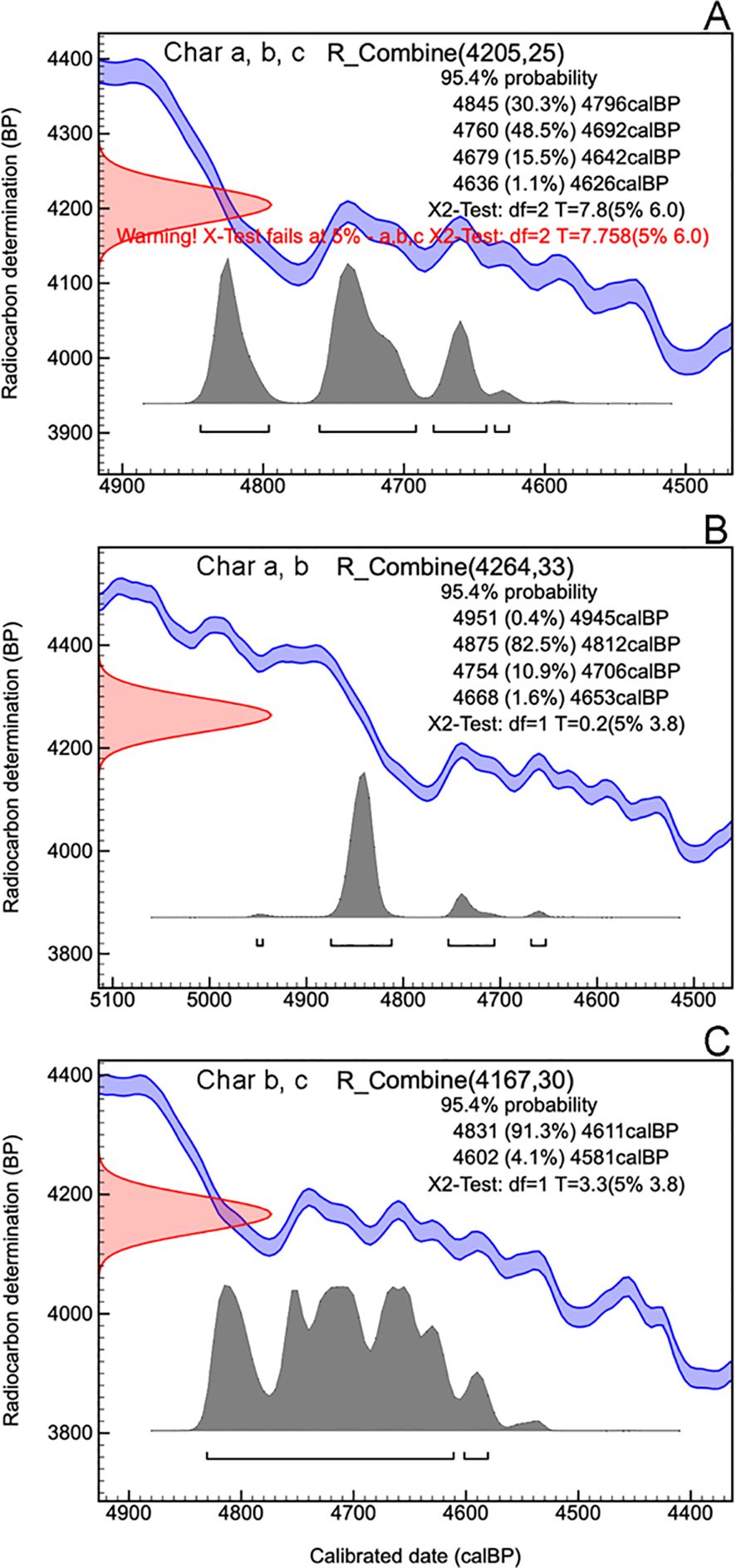
Figure 4. OxCal plot of the combined date of samples Charcoal a, Charcoal b and Charcoal c (A), of the combined date of charcoal samples Charcoal a and Charcoal b (B) and of the combined date of charcoal samples Charcoal b and Charcoal c (C).
No charcoal was detected at depths of 50 and 100 cm. In contrast, at a depth of 150 cm, a total of 76 charcoal fragments were recovered over the sieve 0.5 mm. Three taxa were identified: the tree Fagus sylvatica (n = 2), the shrub Daphne sp. (n = 6), and a perennial grass of the Poaceae family, Dactylis sp. (n = 25); 43 charcoal fragments remained unidentified. A few very (n= 10) small and poorly preserved charcoal fragments were found at the bottommost level (200 cm), but they could not be identified due to their extremely poor state of preservation coupled with their small size.
Scanning electron microscopy (SEM) revealed distinct anatomical features and taphonomic alterations in the ancient Fagus and Dactylis samples, as shown in Figure 5. Fagus (Figures 5A, B) displayed the expected thick-walled cells arranged in a regular pattern (Figure 5A), typical of hardwood trees, but with evidence of cell wall degradation (Figure 5A), consistent with an ancient sample. In contrast, Dactylis (Figures 5C, D) exhibited thinner cell walls and a more varied cellular arrangement (Figure 5C), characteristic of grasses. Notably, the Dactylis sample showed adhered depositions on cell surfaces (Figure 5D), giving it a rock-like solidified appearance, indicative of mineralization. The preservation of key cellular details in both samples, despite these alterations, highlights the resilience of plant tissues and the power of SEM analysis for investigating ancient plant remains.

Figure 5. Scanning electron microscope images of the anatomical structure in the longitudinal tangential plane of Fagus sylvatica wood, showing ray parenchyma cells in a multiseriate ray (A, B), and of vascular tissues in the longitudinal plane of Dactylis spp. herbaceous samples (C, D) at different magnifications. The images on the right provide a magnified view of the red-squared area from the corresponding left image.
SEM-EDS analysis provided crucial evidence linking the ancient Fagus and Dactylis samples to the Al2O3-rich suggesting that the charcoal is buried in a paleosoil (Sheldon and Tabor, 2009). As expected, the EDS mapping clearly showed a layer of Al2O3 covering both samples, consistent with the characteristic accumulation of this mineral in paleosols. The EDS maps and X-ray spectrum further confirmed the presence of aluminum (Al) in both the Fagus and Dactylis samples, as shown in Figure 6. Notably, a mapping consistent with uniform distribution of Fe2O3 was detected in Fagus but not in Dactylis, possibly indicating iron-related mineral formation within the degrading wood tissues. While both samples showed some silicate presence, the levels were higher in Dactylis, consistent with the expected silica accumulation in grasses, contributing to its rock-like appearance. This combined SEM-EDS analysis not only confirmed the presence of Al2O3, a hallmark of the paleosol environment, but also revealed how this mineral and other elements interacted with the plant tissues, influencing their preservation and taphonomic alteration.

Figure 6. Scanning electron microscope (SEM) images combined with Energy-Dispersive X-ray Spectroscopy (EDS) mapping and X-ray spectra emission showing the elemental composition of F sylvatica wood (left) and Dactylis spp. herbs (right). (a) Surfaces of F sylvatica charcoal, the elemental maps illustrate the accumulation of Al2O3. (b, f) Localized accumulation of aluminium (Al) on the surfaces of F sylvatica wood (b) and Dactylis spp. herbs (f). (c, g) EDS elemental mapping showing the distribution of oxygen (O) on the surfaces of F. sylvatica wood (c) and Dactylis spp. herbs (g). (d, h) EDS X-ray emission spectra from representative areas on F sylvatica wood (d) and Dactylis spp. herbs (h). (e) SEM image showing the carbon (C) distribution on a closer view of the surface of Dactylis spp. herbs. The localized accumulation of aluminium on both types of charcoal corroborates the radiodating data, as this process is particularly slow and occurs in paleosols that have undergone calcium leaching, which has not been found in large quantities as expected in recent soils of carbonate matrix. Sample scales are indicated in the respective images.
4 Discussion
4.1 Current treeline elevation and Fagus sylvatica regeneration
The treeline of Mt. Cervati is, on average, at 1,710 m a.s.l., which is higher than the average of the Apennine chain, which is 1,589 m a.s.l (Bonanomi et al., 2018). However, this value is decidedly lower than the estimated ecological potential of F. sylvatica at these latitudes, which is higher than 2,000 m a.s.l (Bonanomi et al., 2018). In fact, treelines of F. sylvatica higher than 2000 m a.s.l. have been described in the Apennines both further south, such as in Pollino (Rita et al., 2021), and further north, such as on Mount Argatone, Mount Greco and on Majella (Bonanomi et al., 2020). This implies that the Cervati treeline is depressed, on average and with respect to its climatic potential, by at least 300 m. The in-depth analysis by slope has also revealed that on the southern slope the treeline is on average 187 m lower than on the other slopes, where the minimum values are also found at only 1,342 m a.s.l., much lower than the absolute maximums located on some valleys on the northern slope that almost reach the top of the mountain at 1,891 m a.s.l. Our findings that the treeline elevation was consistently lower on warmer, south-facing slopes, is consistent with earlier research across the Apennines (Bonanomi et al., 2020). This pattern, though the underlying mechanisms remain unclear, may result from a synergistic interplay between human disturbance (such as logging and grazing pressure) and climatic restrictions (such as summer dryness), which could result in the loss of F. sylvatica canopy viability and regeneration capability.
When compared to the predicted climatic potential, the depression of the treeline elevation at Mont Cervati suggests a widespread anthropogenic influence. Our hypothesis states that within mountain groups, we found the coexistence of a highly depressed treeline, in several cases with elevation of only ~1,300 m a.s.l., alongside a very high treeline, reaching up to nearly 1,900 m a.s.l. The co-occurrences of such variability in treeline elevation across sites that are a few kilometers apart suggest that factors other than climate control this pattern. Given this, we propose that high elevation treeline exists in remote, inaccessible and steep valleys which have historically been shielded from anthropogenic disturbances, both past and present. Conversely, very depressed treeline is located in accessible areas that have been heavily exploited in previous centuries.
Despite intensive research, no seedling or sapling regeneration was detected above the treeline regardless of elevation and exposure. This observation suggests that the open grassland environment poses significant challenges to F. sylvatica to recolonization. According to Allegrezza et al. (2016), even at the treeline plants outside of the canopy cover on south-facing mountain slopes may face extremely high soil and air temperatures (up to 38°C) and severe summer droughts. These conditions significantly limit the ability of F. sylvatica to regenerate on open patches. In this approach, F. sylvatica may act as an ecosystem engineer by modifying forest microclimate through its canopy cover, thereby facilitating seedling establishment. As a result, we hypothesize that the treeline depression on southern mountain faces is a consequence of F. sylvatica reduced engineering potential following canopy removal. When left intact, this species exerts positive feedback on microclimate, reducing wind effects, buffering temperature extremes, and enhancing soil and air moisture (Hastings et al., 2007). A recent study (Bonanomi et al., 2021) demonstrated that in the presence of shrubs such as Juniperus communis and Pinus mugo, F. sylvatica can rapidly recolonize areas beyond the treeline. This is due to the facilitative effects of the microclimate modulated by the crowns of the nurse shrub, thereby improving the conditions for F. sylvatica establishment. We hypothesize that the buffering effect of plant canopies will be more significant in southern than in northern aspects, where air and soil temperature are already buffered by the lower solar radiation, because self-shading cushions local microclimate (Rita et al., 2021). The limited ability to recolonize open areas above the treeline has been reported in several areas of the globe and the combination of summer drought with the shortness of the growing season are formidable constraints for many species living at the upper treeline (Elliott et al., 2020; Lyu et al., 2019; Bar-On et al., 2022). Such factors undoubtedly limit the ability of the treeline to advance even where thermal limitations are less severe due to global warming (Harsch et al., 2009).
4.2 Past fire causes depression of current treeline?
European mountain ranges including the Alps, the Pyrenees and the Apennines have a history of intense disturbances such as recurrent fires, forest clearing, and intensive grazing that have profoundly altered vegetation across various elevation (Gobet et al., 2003; Bal et al., 2011). Human activity has been particularly intense along the Apennines as demonstrated by the pervasive presence of kiln platforms, observed from low elevation up to the treeline at approximately 2,000 m a.s.l (Carrari, 2015; Bonanomi et al., 2020). Studies based on pollen profiles, such as those conducted at Lake Monticchio located 73 km from Monte Cervati, have reconstructed vegetation fluctuations over the last 76,300 years (Watts et al., 1996), but with low resolution and therefore at a regional scale. Studies based on anthracological remains found in archaeological sites are more detailed. Near the study area, the Serratura cave (Di Pasquale et al., 2020) and the Maria Colombo cave (Belli, 2016) were studied. In both cases, the studies enable the reconstruction of the vegetation surrounding the cave area, although it cannot be ruled out a selection effect by populations that collected wood for cooking and heating. Furthermore, both caves are present at relatively low elevation, 830 m for Maria Colombo (Belli, 2016) and at sea level for the Serratura cave (Di Pasquale et al., 2020), therefore with limited relevance as regards the vegetation dynamics at the upper limit of the forest. In this context, the discovery of the black layer in the summit sinkhole of Monte Cervati provides a unique opportunity to reconstruct, with temporal precision, the changes in vegetation regarding the peaks of the Apennine mountains.
The localized accumulation of numerous charcoals remain within a single soil layer demonstrates that the summit of Monte Cervati experienced significant fire event(s). To estimate a probable calendar age for the burning event, we tentatively combined all ^14C dates using the OxCal function R_Combine. However, the χ² test indicated that this combination should be rejected at the 5% confidence level. Notably, the χ² test also failed when we attempted to combine samples a and c. A consistent result was obtained, however, by pooling sample b with sample a and sample c, one at a time. We therefore suggest that the black layer originated from multiple but temporally close fire events, with at least two separate occurrences at 4841 cal BP and 4710 cal BP, approximately 100 years apart. In partial agreement with our hypothesis, the vegetation affected by the fire was not a dense forest, but rather a plant community dominated by tall, likely perennial grass and few scattered trees, a vegetation particularly prone to fire. This suggests that around 4,800 years BP the vegetation of Cervati was more similar to a tall grassland or perhaps a wooded grassland. This event precedes by a few centuries the well-known 4200 BP event (Bini et al., 2019) which marked one of the most significant climate changes of Holocene. Starting from 4200 BP the climate in many regions of the globe has significantly dried out and in some areas even cooled. The event was of such magnitude that several researchers have hypothesized that it contributed to the decline or collapse of several ancient kingdoms (e.g., the Old Kingdom in Egypt, the Akkadian Empire in Mesopotamia, the Liangzhu culture in China) (Weiss, 2016). However, a recent analysis has downplayed the significance and impact of the 4200 BP event, also indicating that a particularly warm period during the Holocene occurred at 4800 BP (McKay et al., 2024), which coincides with the period in which the fires on the Cervati occurred. In our context, a plausible reconstruction is that a particularly warm period fostered the development of vegetation at high elevation, including a high herbaceous biomass that would create the conditions for wildfires. Such fire events, whose origin-natural or anthropogenic-remains uncertain, were likely extensive as it left significant traces in the soil profile. Subsequently, it is likely that the soil was eroded from the slopes of the sinkhole, creating a rocky environment visible today, and leading to the deposition of approximately 1.5 m of soil above the charcoal layer at the sinkhole’s center.
The charcoal, buried in the soil for 4800 years, then underwent a very advanced ageing process. This was clarified by the SEM-EDS analysis which highlighted the accumulation on the surfaces of Al2O3, known to be present in paleosol as it is not leachable (Sheldon and Tabor, 2009), unlike other cations such as Ca, Na and K. A recent study conducted on charcoal present in kilns in southern Italy forests over carbonate substrates similar to Cervati has shown that much more recent charcoal (aged about 100–200 years) show localized accumulations of Ca, Na and K (Iacomino et al., 2024).
Our study, however, cannot shed light on what happened between the end of the last glaciation and the initial phases of the Holocene. In fact, some evidence shows that during the last glacial maximum even on Mount Cervati there were some small glaciers (Palmentola et al., 1990). In the initial phase of the Holocene the climate was rather warm and humid along the Apennine chain (Giraudi et al., 2011), a factor that could have allowed the development of tree vegetation even at high elevations. To address these questions our research group is carrying out further investigations on Mount Cervati based on molecular methods of ancient DNA (Edwards, 2020) combined with pedo-anthracological analysis of soil layers older than the black layer investigated in this study.
5 Conclusions
At Monte Cervati, the remains of an ancient fire buried in a sinkhole were discovered, which we now know temporally preceded the great drought known as the 4200 BP event. This series of such fire events, likely favored by the warm and arid climate, changed the vegetation of the mountain contributing to giving it the appearance with rocky soil and dry grasslands, resembling the current appearance of the summit area of hundreds of mountains in the southern Apennines. Our study shows that in climates with summer drought, intense fire events can cause critical transition with consequent catastrophic shifts in ecosystems that enter an alternate stable state, i.e. woodland or dry grassland. Soil erosion following fire events makes summit areas no longer recolonizable by tree even in historical times. The current state is probably also maintained by the intense grazing still present in summit areas of most of the mountains in Appennines. The study, therefore, indicates that in mountain areas with arid climates in the summer months, global warming is unlikely to cause treeline advancement until other limiting factors like drought and grazing are relaxed. Future research will extend the combined approach of pedo-anthracology with the help of ancient DNA to other mountain groups of the Apennines such as Majella, Velino-Sirente, Sibillini and Pollino to better understand the vegetation history of these areas that experienced antropogenic pressure for millennia.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The manuscript is approved by all authors for publication. I would like to declare on behalf of my co-authors that the work described is original research that has not been published previously, and not under consideration for publication elsewhere.
Author contributions
GB: Methodology, Supervision, Conceptualization, Investigation, Writing – original draft, Formal Analysis, Resources. AS: Formal Analysis, Investigation, Writing – review & editing, Data curation, Validation. GA: Validation, Writing – review & editing, Investigation, Visualization. EA: Data curation, Methodology, Validation, Writing – review & editing, Investigation. GI: Validation, Data curation, Investigation, Writing – review & editing. GM: Investigation, Validation, Resources, Funding acquisition, Writing – review & editing. RM: Writing – review & editing, Methodology, Supervision, Investigation, Visualization. AN: Validation, Investigation, Writing – review & editing. FB: Writing – review & editing, Validation, Visualization, Investigation. MA: Investigation, Visualization, Writing – review & editing, Validation. LD: Writing – review & editing, Methodology, Visualization, Investigation, Supervision, Validation. MI: Resources, Supervision, Formal Analysis, Methodology, Data curation, Writing – original draft, Visualization, Investigation, Conceptualization, Validation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the Project “Rifugi montani sentinelle del clima e dell’ambiente” financed by Club Alpino Italiano.
Acknowledgments
The authors extend their appreciation to Fabio Marzaioli, Gaia Isoldi, Luigi Saulino for technical and logistic support during the field work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Allegrezza, M., Corti, G., Cocco, S., Pesaresi, S., Chirico, G. B., Saracino, A., et al. (2016). Microclimate buffering and fertility island formation during Juniperus communis ontogenesis modulate competition–facilitation balance. J. Veg. Sci. 27, 616–627. doi: 10.1111/jvs.12386
Bailey, S. N., Elliott, G. P., and Schliep, E. M. (2021). Seasonal temperature-moisture interactions limit seedling establishment at upper treeline in the Southern Rockies. Ecosphere 12, e03568. doi: 10.1002/ecs2.3568
Bal, M. C., Pelachs, A., Perez-Obiol, R., Julia, R., and Cunill, R. (2011). Fire history and human activities during the last 3300 calyr BP in Spain’s Central Pyrenees: the case of the Estany de Burg. Palaeogeogr. Palaeoclimatol. Palaeoecol. 300, 179–190. doi: 10.1016/j.palaeo.2010.12.023
Bar-On, P., Yaakobi, A., Moran, U., Rozenstein, O., Kopler, I., and Klein, T. (2022). A montane species treeline is defined by both temperature and drought effects on growth season length. Tree Physiol. 1 42, 1700–1719. doi: 10.1093/treephys/tpac070
Bartolucci, L., Peruzzi, G., Galasso, A., Alessandrini, A., Ardenghi, N. M. G., Bacchetta, G., et al. (2024). A second update to the checklist of the vascular flora native to Italy. Plant Biosyst. 158, 219–296. doi: 10.1080/11263504.2024.2320126
Belli, C. (2016). Caratteristiche paleoambientali e agricole del Vallo di Diano (SA). Integrazione di metodologie archeobotaniche. University of Salento, Lecce, Italy.
Benatti, A., Bal, M., Allée, P., Bosi, G., and Mercuri, A. M. (2019). Plant landscape reconstruction above the current timberline at the Monte Cimone and Cornoalle Scale mountain areas (Northern Apennines, Italy) during the Late Holocene: The evidence from soil charcoal. Holocene 29, 1767–1781. doi: 10.1177/0959683619862033
Bini, M., Zanchetta, G., Perşoiu, A., Cartier, R., Català, A., Cacho, I., et al. (2019). The 4.2 ka BP Event in the Mediterranean region: an overview. Clim. Past 15, 555–577. doi: 10.5194/cp-15-555-2019
Bonanomi, G., Mogavero, V., Rita, A., Zotti, M., Saulino, L., Tesei, G., et al. (2021). Shrub facilitation promotes advancing of the Fagus sylvatica treeline across the Apennines (Italy). J. Veg. Sci. 32, e13054. doi: 10.1111/jvs.13054
Bonanomi, G., Rita, A., Allevato, E., Cesarano, G., Saulino, L., Di Pasquale, G., et al. (2018). Anthropogenic and environmental factors affect the tree line position of Fagus sylvatica along the Apennines (Italy). J. Biogeogr. 45, 2595–2608. doi: 10.1111/jbi.13408
Bonanomi, G., Zotti, M., Mogavero, V., Cesarano, G., Saulino, L., Rita, A., et al. (2020). Climatic and anthropogenic factors explain the variability of Fagus sylvatica treeline elevation in fifteen mountain groups across the Apennines. For. Ecosyst. 7, 1–13. doi: 10.1186/s40663-020-0217-8
Carcaillet, C. and Thinon, M. (1996). Pedoanthracological contribution to the study of the evolution of the upper treeline in the Maurienne Valley (North French Alps): methodology and preliminary data. Rev. Palaeobot. Palynol. 91, 399–416. doi: 10.1016/0034-6667(95)00060-7
Carrari, E. (2015). Legacy effects of former charcoal kiln sites on the forest vegetation of a Mediterranean area. University of Florence, Florence, Italy.
Colombaroli, D., Henne, P. D., Kaltenrieder, P., Gobet, E., and Tinner, W. (2010). Species responses to fire, climate and human impact at tree line in the Alps as evidenced by palaeo-environmental records and a dynamic simulation model. J. Ecol. 98, 1346–1357. doi: 10.1111/j.1365-2745.2010.01723.x
Dallwitz, M. J., Paine, T. A., and Zurcher, E. J. (1995).User’s guide to Intkey: a program for interactive identification and information retrieval. Available online at: http://delta-intkey.com (Accessed June 20, 2025).
Di Pasquale, G., Saracino, A., Bosso, L., Russo, D., Moroni, A., Bonanomi, G., et al. (2020). Coastal pine-oak glacial refugia in the Mediterranean basin: A biogeographic approach based on charcoal analysis and spatial modelling. Forests 11, 673. doi: 10.3390/f11060673
Edwards, M. E. (2020). The maturing relationship between quaternary paleoecology and ancient sedimentary DNA. Quat. Res. 96, 39–47. doi: 10.1017/qua.2020.52
Elliott, G. P., Bailey, S. N., and Cardinal, S. J. (2020). Hotter drought as a disturbance at upper treeline in the southern Rocky Mountains. Ann. Am. Assoc. Geogr. 111, 756–770. doi: 10.1080/24694452.2020.1805292
Geib, P. R. (2008). Age discrepancies with the radiocarbon dating of sagebrush (Artemisia tridentata Nutt.). Radiocarbon 50, 347–357. doi: 10.1017/S0033822200053480
Giraudi, C., Magny, M., Zanchetta, G., and Drysdale, R. N. (2011). The Holocene climatic evolution of Mediterranean Italy: A review of the continental geological data. Holocene 21, 105–115. doi: 10.1177/0959683610377529
Gobet, E., Tinner, W., Hochuli, P. A., Van Leeuwen, J. F. N., and Ammann, B. (2003). Middle to Late Holocene vegetation history of the Upper Engadine (Swiss Alps): the role of man and fire. Veg. Hist. Archaeobot. 12, 143–163. doi: 10.1007/s00334-003-0017-4
Gravina, C., Formato, M., Piccolella, S., Fiorentino, M., Stinca, A., Pacifico, S., et al. (2023). Lavandulaaustroapennina (Lamiaceae): getting insights into bioactive polyphenols of a rare Italian endemic vascular plant. Int. J. Mol. Sci. 24, 8038. doi: 10.3390/ijms24098038
Greguss, P. (1959). Holzanatomie der Europäischen laubhölzer und sträucher (Budapest: AkademialKiado).
Harsch, M. A., Hulme, P. E., McGlone, M. S., and Duncan, R. P. (2009). Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 12, 1040–1049. doi: 10.1111/j.1461-0248.2009.01355.x
Hastings, A., Byers, J. E., Crooks, J. A., Cuddington, K., Jones, C. G., Lambrinos, J. G., et al. (2007). Ecosystem engineering in space and time. Ecol. Lett. 10, 153–164. doi: 10.1111/j.1461-0248.2006.00997.x
Heiss, A. G. (2002). Anatomy of European and North American woods – an interactive identification key. Available online at: https://ucrisportal.univie.ac.at/en/publications/anatomy-of-european-and-north-american-woods-an-interactive-ident (Accessed June 20, 2025).
Iacomino, G., Idbella, M., di Costanzo, L., Amoroso, G., Allevato, E., Abd-ElGawad, A. M., et al. (2024). Biochar aging, soil microbiota and chemistry of charcoal kilns in Mediterranean forests. Biochar 6, 1–20. doi: 10.1007/s42773-024-00378-3
InsideWood (2004). Available online at: http://insidewood/lib.ncsu.edu/search (Accessed June 20, 2025).
Körner, C. (1998). A re-assessment of high elevation treeline positions and their explanation. Oecologia 115, 445–459. doi: 10.1007/s004420050540
Körner, C. (2012). Alpine treelines. Functional ecology of the global high elevation tree limits (Basel, Switzerland: Springer Basel, Springer Science and Business Media).
Körner, C. and Paulsen, J. (2004). A world-wide study of high altitude treeline temperatures. J. Biogeogr. 31, 713–732. doi: 10.1111/j.1365-2699.2003.01043.x
Liang, E., Wang, Y., Piao, S., Lu, X., Camarero, J. J., Zhu, H., et al. (2016). Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. U.S.A. 113, 4380–4385. doi: 10.1073/pnas.1520582113
Lyu, L., Zhang, Q. B., Pellatt, M. G., Büntgen, U., Li, M. H., and Cherubini, P. (2019). Drought limitation on tree growth at the Northern Hemisphere’s highest tree line. Dendrochronologia 53, 40–47. doi: 10.1016/j.dendro.2018.11.006
Magri, D. (2007). Advances in Italian palynological studies: late Pleistocene and Holocene records. GFF 129, 337–344. doi: 10.1080/11035890701294337
Magri, D., Vendramin, G. G., Comps, B., Dupanloup, I., Geburek, T., Gömöry, D., et al. (2006). A new scenario for the Quaternary history of European beech populations: palaeobotanical evidence and 1 genetic consequences. New Phytol. 171, 199–221. doi: 10.1111/j.1469-8137.2006.01740.x
McKay, N. P., Kaufman, D. S., Arcusa, S. H., Kolus, H. R., Edge, D. C., Erb, M. P., et al. (2024). The 4.2 ka event is not remarkable in the context of Holocene climate variability. Nat. Commun. 15, 6555. doi: 10.1038/s41467-024-50886-w
McWethy, D. B., Whitlock, C., Wilmshurst, J. M., McGlone, M. S., Fromont, M., Li, X., et al. (2010). Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement. Proc. Natl. Acad. Sci. U.S.A. 107, 21343–21348. doi: 10.1073/pnas.1011801107
Mercuri, A. M., Allevato, E., Arobba, D., Bandini Mazzanti, M., Bosi, G., Caramiello, R., et al. (2015). Pollen and macroremains from Holocene archaeological sites: A dataset for the understanding of the bio-cultural diversity of the Italian landscape. Rev. Palaeobot. Palynol. 218, 250–266. doi: 10.1016/j.revpalbo.2014.05.010
Miehe, G., Miehe, S., Vogel, J., and La, D. (2007). Highest treeline in the northern hemisphere found in southern Tibet. Mt. Res. Dev. 27, 169–173. doi: 10.1659/mrd.0792
Palmentola, G., Acquafredda, P., and Fiore, S. (1990). A new correlation of the glacial moraines in the Southern Apennines, Italy. Geomorphology 3, 1–8. doi: 10.1016/0169-555X(90)90028-O
Pignatti, S., Guarino, R., and La Rosa, M. (2017a). Flora d’Italia Vol. 1 (Bologna, Italy: Edagricole), 1064.
Pignatti, S., Guarino, R., and La Rosa, M. (2017b). Flora d’Italia Vol. 2 (Bologna, Italy: Edagricole), 1178.
Pignatti, S., Guarino, R., and La Rosa, M. (2018). Flora d’Italia Vol. 3 (Bologna, Italy: Edagricole), 1288.
Pignatti, S., Guarino, R., and La Rosa, M. (2019). Flora d’Italia Vol. 4 (Bologna, Italy: Edagricole), 1054.
Ramsey, C. B. (2009). Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360. doi: 10.1017/S0033822200033865
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Ramsey, C. B., et al. (2020). The IntCal20 Northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757. doi: 10.1017/RDC.2020.41
Rita, A., Bonanomi, G., Allevato, E., Borghetti, M., Cesarano, G., Mogavero, V., et al. (2021). Topography modulates near-ground microclimate in the Mediterranean Fagus sylvatica treeline. Sci. Rep. 11, 8122. doi: 10.1038/s41598-021-87661-6
Robin, V. and Nelle, O. (2014). Contribution to the reconstruction of central European fire history, based on the soil charcoal analysis of study sites in northern and central Germany. Veg. Hist. Archaeobot. 23, 51–65. doi: 10.1007/s00334-014-0438-2
Santangelo, A., La Valva, V., Di Novella, N., and Caputo, G. (1994). La flora cacuminale del Monte Cervati (Appennino campano). Delpinoa n.s. 31-32, 99–139.
Schweingruber, F. H. (1990). “Anatomy of Europeanwoods,” in Amphoras from the 1970 excavations at Colchester Sheepen, British archaeological reports, vol. 142 . Ed. Sealey, P. R.British series (Stuttgart, Paul Haupt Berne and Stuttgart Publishers, BAR, Oxford).
Sheldon, N. D. and Tabor, N. J. (2009). Quantitative paleoenvironmental and paleoclimatic reconstruction using paleosols. Earth-Sci. Rev. 95, 1–52. doi: 10.1016/j.earscirev.2009.03.004
Sigdel, S. R., Wang, Y., Camarero, J. J., Liang, E., Zhu, H., and Peñuelas, J. (2018). Moisture-mediated responsiveness of treeline shifts to global warming in the Himalayas. Glob. Change Biol. 24, 5549–5559. doi: 10.1111/gcb.14428
Sigdel, S. R., Zheng, X. Y., Babst, F., Camarero, J. J., Gao, S., Li, X. X., et al. (2024). Accelerated succession in Himalayan alpine treelines under climatic warming. Nat. Plants 10, 1909–1918. doi: 10.1038/s41477-024-01855-0
Stinca, A., Musarella, C. M., Rosati, L., Laface, V. L. A., Licht, W., Fanfarillo, E., et al (2021). Italian vascular flora: new findings, updates and exploration of floristic similarities between regions. Diversity 13, 600. doi: 10.3390/d13110600
Talon, B., Carcaillet, C., and Thinon, M. (1998). Études pédoanthracologiques des variations de la limite supérieure des arbres au cours de l’Holocene dans les Alpes Francaises. Geogr. Phys. Quater. 52, 195–208. doi: 10.7202/004875ar
Thinon, M. (1978). La pédoanthracologie: Une nouvelle méthode d’analyse phytochronologique depuis le Néolithique. C.R. Acad. Sc. Paris 287, 1203–1206.
Walsh, S. J., Butler, D. R., Allen, T. R., and Malanson, G. P. (1994). Influence of snow patterns and snow avalanches on the alpine treeline ecotone. J. Veg. Sci. 5, 657–672. doi: 10.2307/3235881
Watts, W. A. (1985). A long pollen record from Laghi di Monticchio, southern Italy: A preliminary account. J. Geol. Soc 142, 491–499. doi: 10.1144/gsjgs.142.3.0491
Watts, W. A., Allen, J. R. M., and Huntley, B. (1996). Vegetation history and palaeoclimate of the last glacial period at Lago Grande di Monticchio, southern Italy. Quat. Sci. Rev. 15, 133–153. doi: 10.1016/0277-3791(95)00093-3
Weiss, H. (2016). Global megadrought, societal collapse and resilience at 4.2–3.9 ka BP across the Mediterranean and west Asia. Pages Mag. 24, 62–63. doi: 10.22498/pages.24.2.62
Whitlock, C. and Larsen, C. (2001). “Charcoal as a fire proxy.” in Tracking Environmental Change Using Lake Sediments, eds. Smol, J. P., Birks, H. J. B., and Last, W. M. (Dordrecht: Springer), 75–97.
Wilkinson, A. J. and Britton, T. B. (2012). Strains, planes, and EBSD in materials science. Mater. Today 15, 366–376. doi: 10.1016/S1369-7021(12)70163-3
Zitti, S., Frattaroli, A. R., Carli, E., and Cutini, M. (2016). “9210 *Faggeti degli Appennini con Taxus e Ilex,” in Manuali per il Monitoraggio di Specie e Habitat di Interesse Comunitario (Direttiva 92/43/CEE) in Italia: Habitat. Eds. Angelini, P., Casella, L., Grignetti, A., and Genovesi, P. (ISPRA, Roma, Italy), 234–235.
Keywords: pedo-anthracology, charcoal, vegetation history, fire, black layer, SEM-EDS
Citation: Bonanomi G, Stinca A, Amoroso G, Allevato E, Iacomino G, Mauriello G, Motti R, Nicastri A, Bellucci F, Abelouah MR, Di Costanzo L and Idbella M (2025) Fire history and treeline elevation in the Apennines: insights from pedo-anthracological analysis on Monte Cervati, Southern Italy. Front. Plant Sci. 16:1617687. doi: 10.3389/fpls.2025.1617687
Received: 24 April 2025; Accepted: 12 June 2025;
Published: 01 July 2025.
Edited by:
Feng Chen, Yunnan University, ChinaReviewed by:
Rubén Pardo Martínez, University of Granada, SpainEleonora Cagliero, University of Padua, Italy
José Manuel Zuñiga-Vásquez, National Institute of Forestry and Agricultural Research (INIFAP), Mexico
Copyright © 2025 Bonanomi, Stinca, Amoroso, Allevato, Iacomino, Mauriello, Motti, Nicastri, Bellucci, Abelouah, Di Costanzo and Idbella. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mohamed Idbella, bW9oYW1lZC5pZGJlbGxhQHVtNnAubWE=