 Yali Li
Yali Li Guoxing He
Guoxing He Xiaoni Liu1,2*
Xiaoni Liu1,2* Tong Ji
Tong Ji- 1Key Laboratory of Grassland Ecosystem, Ministry of Education, Pratacultural College, Gansu Agricultural University, Lanzhou, Gansu, China
- 2Sino-U.S. Center for Grazing Land Ecosystem Sustainability, Lanzhou, Gansu, China
Introduction: Ecosystem multifunctionality (EMF) refers to the integrated capacity of an ecosystems to provide a variety of services and functions. It plays a crucial role in maintaining ecological balance and achieving sustainable grassland development. Grassland type is a classification unit based on dominant species within the grassland ecosystem. However, the response of EMF to grassland type is still unclear, and its influencing factors need to be studied in depth.
Methods: In this study, we investigated the distribution characteristics of EMF in the Longzhong Loess Plateau temperate desert under different grassland types (Kalidium foliatum type, Reaumuria songarica type, Salsola passerinum type, and Sympegma regelii type grasslands) and identified the key factors driving changes in EMF.
Results and discussion: The results indicated the K. foliatum type grassland exhibited higher EMF and belowground ecosystem multifunctionality (BEMF) indices (0.80 and 0.77, respectively), significantly greater compared to R. soongorica type, S. passerine type, and S. regelii type grasslands (P< 0.05). EMF and BEMF were significantly correlated with plant factors (coverage, root-shoot ratio), soil factors (mean weight diameter, MWD and geometric mean diameter, GMD) (P< 0.05), and showed strong positive relationships with Simpson diversity index. Hierarchical partitioning further indicated that MWD and GMD explained 29.86% and 38.21% of the variance in EMF, respectively. Structural equation modeling revealed that soil factors (MWD, GMD), plant factors (coverage, root-shoot ratio), and Simpson diversity index collectively explained 86% of the variation in EMF. Soil factors primarily exerted direct effects and indirectly promoted EMF by enhancing plant factors, with a total effect of 0.921. These findings suggest that K. foliatum type grassland can maintain EMF through soil factors, thereby supporting the sustainable development of temperate desert ecosystems in the Longzhong region of the Loess Plateau.
1 Introduction
Ecosystem multifunctionality (EMF) refers to the capacity of ecosystems to provide multiple services and functions (Yao et al., 2025). These functions such as net primary productivity, carbon sequestration, climate regulation, soil and water conservation, windbreaks and sand fixation, as well as species richness, among others (Hector and Bagchi, 2007). In recent years, global climate change and human activities have led to the degradation of grassland ecosystems, resulting in a decline in EMF (Li et al., 2024b; Yao et al., 2025). This decline disrupts ecosystem balance and poses a threat to the sustainable development of human societies (Bardgett et al., 2021; Wu et al., 2021). Against this background, scholars have begun to focus on the synergistic variations of functions such as soil retention, biodiversity maintenance, and material cycling within grassland ecosystems under different plant community conditions, conducting systematic investigations (Cheng et al., 2021; Yao et al., 2025). However, current research on the influencing factors of EMF under different grassland types remains insufficient, and the mechanisms by which different plant compositions affect EMF have not been systematically elucidated. This gap in knowledge limits a deeper understanding of the relationship between environmental factors and EMF, thereby constraining the optimization of ecosystem management strategies. Therefore, conducting systematic investigations into the influencing factors of EMF under various grassland types is of great significance for the sustainable utilization and conservation of grassland ecosystems.
As the fundamental units of grassland classification, grassland types are formed by different dominant plant species under similar habitat conditions and utilization modes (Li et al., 2024c). The species assembly, density, age, and growth stage of dominant plants vary among different grassland types, which significantly influences the plant community structure within each types (Bezemer et al., 2006). These factors, in turn, affect grassland EMF (Huang et al., 2024; Shu et al., 2023). Relevant studies have indicated that EMF varies significantly due to differences in dominant plant species assembly (Li et al., 2024a). Specifically, changes in the composition of dominant plants among different grassland types can regulate plant productivity and carbon sequestration capacities, thereby impacting ecosystem stability and functionality, ultimately influencing EMF (Shu et al., 2023; Wang et al., 2020; Novák et al., 2020). Therefore, understanding the effects of plant characteristics in different grassland types on EMF is crucial for the scientific conservation and sustainable management of grassland resources.
Soil, as a critical component of the ecosystem, directly influences EMF through its physicochemical properties. The dominant plant species in different grassland types can affect soil nutrient content and structure (Li et al., 2024c). Studies have shown that soil factors, such as pH and soil structure, play a more prominent role in regulating EMF (Han et al., 2021). Moreover, degradation of soil structure, such as erosion or compaction, can lead to nutrient loss and weaken ecosystem functions (Xiong et al., 2023). Therefore, an in-depth investigation into the regulatory mechanisms of soil factors across various grassland types would help enhance ecosystem stability and multifunctionality, providing scientific support for regional ecological restoration.
The Longzhong Loess Plateau is located in the arid and semi-arid regions of northern China, serves as an important ecological security barrier in northwest China (Li et al., 2024b). The area features complex topography, fragile ecological environments, and has been significantly affected by overexploitation and climate change (Liu et al., 2022). Due to environmental degradation, indicators such as vegetation coverage, plant diversity, and soil nutrients have declined, posing serious threats to ecosystem structure and function (Sun et al., 2015; Zhou et al., 2025; Jiang et al., 2016; Gang et al., 2018). As the main grassland type of the Longzhong Loess Plateau, temperate deserts are influenced by soil structural heterogeneity and climate variability, resulting in the formation of distinct grassland types dominated by Kalidium foliatum, Reaumuria songarica, Salsola passerinum, and Sympegma regelii (Li et al., 2024c; Xie et al., 2024). Accordingly, we used four grassland types (K. foliatum type, R. songarica type, S. passerinum type, and S. regelii type) in the Longzhong Loess Plateau temperate desert to assess the differences in EMF among different grassland types and to explore the main driving factors influencing EMF. Our hypotheses were that 1) EMF would be higher in K. foliatum type grasslands compared to the other three types. This is based on its root system adaptations to drought and saline-alkaline environments, which can improve soil structure and thereby enhance ecosystem functions (Li et al., 2024c). And 2) soil factors would have a more significant effect on EMF. By conducting this study, we hope to deepen our understanding of regulatory mechanisms of EMF in temperate desert grasslands and to provide a scientific basis for ecological conservation and grassland management.
2 Materials and methods
2.1 Study site
The research area is situated in the Loess Plateau of central Gansu Province (32°11′–42°57′N, 92°13′–108°46′E), administratively spanning Lanzhou, Dingxi, Baiyin, and Linxia Prefecture. Encompassing approximately 4.26×105 km2. Elevation ranges from 1300 to 4200 m. Annual precipitation is 240 mm, with a maximum daily rainfall of 40 mm. The absolute minimum temperature is -17.1°C, the maximum temperature is 33.2°C, the average annual temperature is 9°C, and the annual growing degree days amount to 3100°C. Soil dominated by sierozem soil (pH ≈7.7). The vegetation composition is predominantly grassland ecosystems, major plant species include K. foliatum, R. songarica, S. passerinum and S. regelii (Li et al., 2024b, 2024d).
2.2 Sample plot design
Grassland type data were obtained from the 1:1,000,000 Grassland Resource Map of China (available via the China Grassland and Ecological Network: http://www.ecograss.com.cn/). August 2021, four representative grassland types in the temperate desert zone of the Longzhong Loess Plateau were selected as experimental sites (Supplementary Figure S1):
1. K. foliatum type grassland (Kf): monodominant community with K. foliatum;
2. R. soongorica type grassland (Rs): characterized by R. soongorica as the keystone species;
3. S. passerina type grassland (Sp): featuring S. passerina as the predominant vegetation;
4. S. regelii type grassland (Sr): dominated by S. regelii communities.
Geospatial distributions and detailed site characteristics are illustrated in Figure 1. These sites were chosen based on their ecological representativeness and distinct dominant species assembly within the regional desert grassland biome. The area of each plot is 100 hm2. In each type of grassland plot, three independent sampling areas of 200 m × 200 m were established according to the principle of terrain homogeneity, with a spacing of ≥ 500 m between the sampling areas to avoid spatial interference.
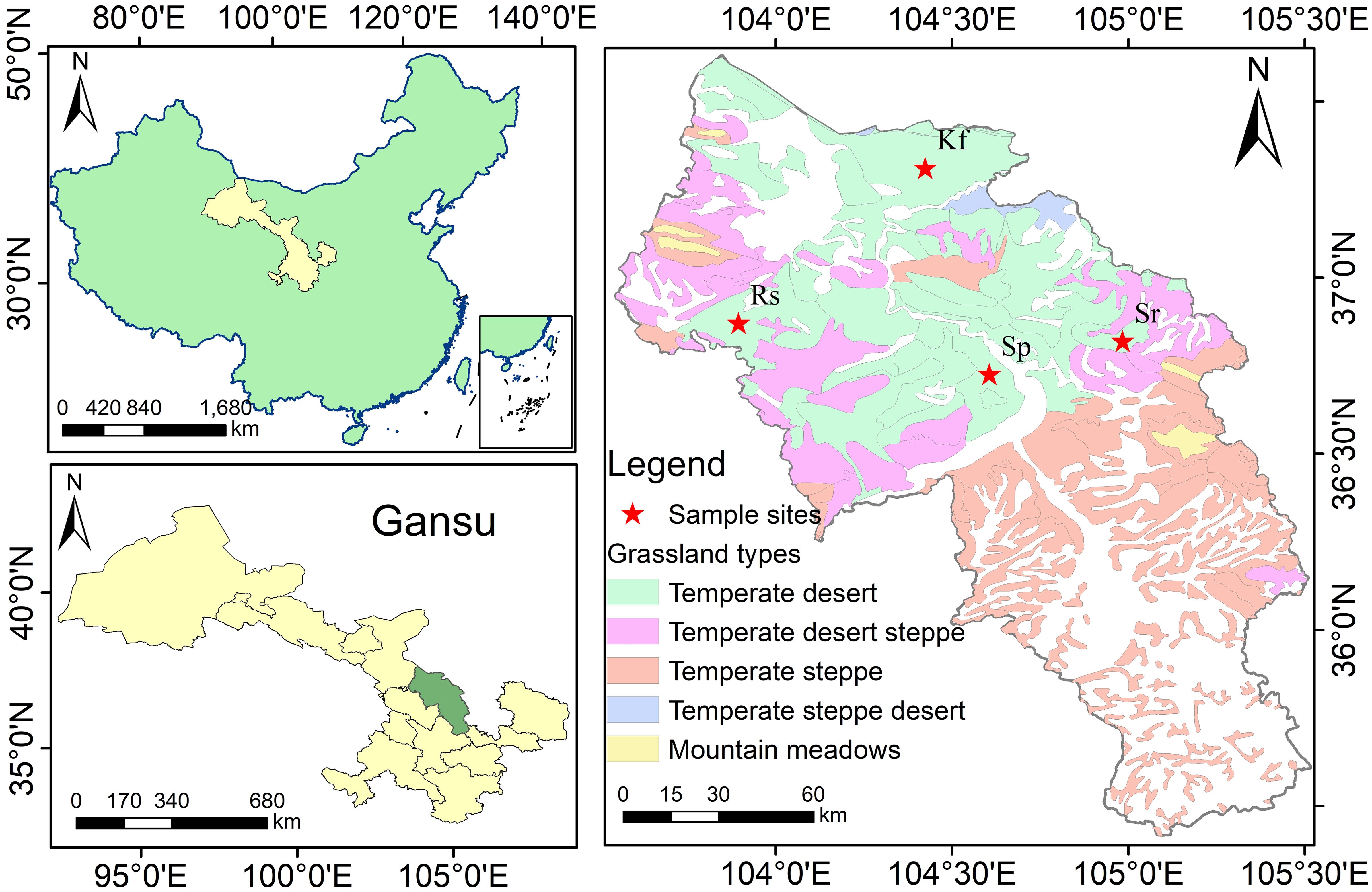
Figure 1. Distribution of sample. Kf, K. foliatum type grassland; Rs, R.soongorica type grassland; Sp, S. passerina type grassland; Sr, S. regelii type grassland.
2.3 Sample collection
2.3.1 Vegetation sampling
In each sampling area, one randomly positioned 20 m × 5 m shrub quadrats were established. Within each shrub quadrat, three 1 m × 1 m herbaceous subplots were randomly delineated for vegetation surveys (Supplementary Figure S2). Shrub communities were assessed for species composition, height, canopy diameter, litter and aboveground biomass within the 20 m × 5 m quadrats. Herbaceous communities were characterized by species composition, height, coverage, and aboveground biomass in the 1 m × 1 m subplots. Total vegetation coverage and biomass per quadrat were recorded. Total biomass was derived by summing the biomass of the shrub layer and herbaceous layer.
2.3.2 Soil physical properties sampling
Soil samples were collected within a 1 m × 1 m sampling plot after harvesting aboveground biomass. Using a stainless steel ring knife (volume: 100 cm3) and aluminum boxes, samples were taken at three depth intervals: 0–10 cm, 10–20 cm, and 20–30 cm. Three replicate samples were collected for each depth layer. Immediately after collection, the samples were sealed and transported to the laboratory for analysis of soil water content (using the 105°C drying method) and bulk density (using the ring knife method).
2.3.3 Chemical properties sampling
Soil samples were collected using a soil auger with a diameter of 5 cm, following an “S” shaped five-point sampling method. Samples were vertically extracted from three depth intervals: 0–10 cm, 10–20 cm, and 20–30 cm. Each depth layer consisted of five borehole samples, which were combined into one composite sample (n = 3 composite samples per layer). The samples were placed in resealable bags and stored in the dark. After air-drying in the laboratory, roots (>2 mm) and gravel were removed, and the soil was ground through a 2 mm nylon sieve. The prepared samples were stored in a desiccator until chemical analysis, which included measurements of pH, organic matter, total nitrogen, and other relevant properties.
2.3.4 Undisturbed soil sampling
Undisturbed soil columns (approximately 2 kg per layer) were collected using a flat-edged iron spade at three depth intervals: 0–10 cm, 10–20 cm, and 20–30 cm. The samples were placed in rigid plastic containers with cushioning pads and transported horizontally to minimize mechanical vibrations. After air-drying in the laboratory, the soil was manually crushed into 1 cm diameter fragments along natural fissures. These fragments were used for determining the composition of water-stable aggregates through wet sieving, with particle size classifications of >2 mm, 0.25–2 mm, 0.053 - 0.25 mm, and< 0.053 mm (Li et al., 2024c).
2.4 Vegetation characteristics
2.4.1 Coverage, height, and aboveground biomass measurement
Coverage (%) was quantified using a grid-based visual estimation method. Grass layer height (cm) was measured vertically from the ground surface to the apex of undisturbed foliage using a ruler. Shrub aboveground biomass (g·m-2) was determined via the standard branch method and placed in labeled bags for measurement. Herbaceous aboveground biomass was harvested at ground level, sorted by species, and processed similarly. All samples were dried in a 65°C oven until reaching a constant weight (approximately 48 hours) before weighing. Total aboveground biomass was calculated as the sum of the herbaceous biomass and shrub biomass per unit area (g·m2).
2.4.2 Belowground biomass measurement
Representative individual shrubs were selected, and the entire root system was excavated using a complete root extraction method. After removing the attached soil, the roots were placed in a 65°C oven to dry until reaching a constant weight. The dried root biomass was then weighed, and the biomass per unit area was calculated (g·m2). The belowground biomass was then obtained.
2.4.3 Litter biomass measurement
Within the study area, litter samples were systematically collected from 1 m × 1 m plots after plant collection in August 2021. During each sampling event, all undecomposed litter within the plot was manually collected using sterile polyethylene gloves. The collected samples were immediately transported to the laboratory for processing. After carefully removing any foreign materials, the litter samples were dried at 65°C until reaching a constant mass (≥48 hours). Subsequently, the weight of the dried samples was measured using a precision balance, accurate to 0.01 grams. The litter biomass was then obtained.
2.4.4 Leaf water content and root - shoot ratio
Leaves were collected from the selected shrubs and transported to the laboratory. After drying at 65°C for 48 hours, the water content of the leaves was measured.
The leaf water content (LWC) was calculated using the gravimetric method, expressed as (Wang et al., 2021):
where FW represents the fresh mass of leaves measured immediately after sampling, and DW denotes the constant mass obtained after oven-drying at 65°C for 48 hr.
Root-Shoot ratio, an indicator of biomass allocation between belowground and aboveground organs, was determined using (Qi et al., 2019):
2.4.5 Plant diversity assessment
Community-level plant diversity was evaluated using four diversity indices (Shannon - Wiener diversity index, H; Simpson diversity index, D; Margalef richness index, R; Pielou evenness index, E) (Peng et al., 2018; Yao et al., 2025). For the specific calculation formulas, please refer to the Supplementary Materials (Supplementary Table S1).
2.5 Soil physicochemical characterization
2.5.1 Physical properties
Soil bulk density (BD, g cm-3) was determined using the core method with stainless steel cutting rings (100 cm3 volume) (He et al., 2024). Soil water content (SWC, %) was calculated following gravimetric analysis (Yinglan et al., 2019):
Aggregate stability analysis was performed through wet sieving methodology. Air-dried soils (50 g) were fractionated using a nested sieve assembly (2 mm, 0.25 mm, 0.053 mm) in a mechanical sieve shaker. The protocol included: (i) 3-min saturation at controlled water level, (ii) vertical oscillation (3–4 cm amplitude) at 30 cycles min-1 for 5 min. Four aggregate fractions were obtained: > 2 mm, 0.25–2 mm, 0.053 - 0.25 mm, and< 0.053 mm (Cambardella and Elliott, 1993; Zhang et al., 2019). Each fraction was oven-dried at 65°C to constant mass and weighed (± 0.001 g).
Aggregate stability indices were computed as (Dou et al., 2020; Zeraatpisheh et al., 2021):
where Xi represents the mean diameter (mm) of the ith fraction, Wi is the mass percentage (%), MWD denotes mean weight diameter (mm), and GMD indicates geometric mean diameter.
2.5.2 Chemical properties
Soil pH was measured in 1:2.5 (w/v) soil-water suspension using a calibrated pH meter (Basic pH Meter PB-10, European). Electrical conductivity (EC, μS·cm-1) was determined in the same extract with a conductivity probe (Conductivity meter DDS-608, Sichuan, China).
The soil analysis methods include: Soil organic matter (SOM, g·kg-1) via potassium dichromate oxidation with heating (He et al., 2024). Soil total nitrogen (TN, g·kg-1) using an automatic carbon and nitrogen analyzer (Primacs SNC 100-IC-E; Sklar, Netherlands). Soil alkaline hydrolyzed nitrogen (AN, mg·kg-1) through alkaline diffusion. Soil total phosphorus (TP, g·kg-1) measured by acid digestion with molybdenum-blue spectrophotometry (Dual Beam UV-Vis Spectrophotometer TH-1901, Beijing Pulse Analytical General Instrument Co). Soil available phosphorus (AP, mg·kg-1) via NaHCO3 extraction with colorimetry. Soil total potassium (TK, g·kg-1) by NaOH fusion and flame photometry (Flame Photometer Model FP640, Shanghai). And soil available potassium (AK, mg·kg-1) by NH4OAc extraction with flame photometric quantification.
2.6 Ecosystem multifunctionality assessment
The ecosystem multifunctionality index (EMF) was quantified through systematic integration of 11 key functional indicators representing primary productivity and biogeochemical cycles (Hector and Bagchi, 2007; Maestre et al., 2012a). The selected indicators encompassed: aboveground biomass, belowground biomass, soil water content, bulk density, organic matter, total nitrogen, alkaline hydrolyzed nitrogen, total phosphorus, available phosphorus, total potassium, available potassium. The belowground ecosystem multifunctional index (BEMF) is calculated based on nine indicators: soil water content, bulk weight, organic matter, total nitrogen, alkaline hydrolyzed nitrogen, total phosphorus, available phosphorus content, total potassium and available potassium. All soil factors are represented as average values for the 0–30 cm depth. The measured indicators comprehensively consider primary productivity and soil nutrient cycling.
The EMF was calculated using the mean value method (Maestre et al., 2012a; Zhai et al., 2024; Yao et al., 2025).
In the first instance, data standardization was performed using min-max normalization (Soliveres et al., 2016; Delgado-Baquerizo et al., 2020):
where Yi denotes the standardized value (0–1 range) of the ith indicator, Xi represents the observed value, with Xmin and Xmax corresponding to the minimum and maximum values across all sampling plots, respectively.
The composite EMF index was calculated as:
where N represents the number of standardized indicators. This additive approach ensures equal weighting across all functions following established protocols.
2.7 Statistical analysis
The data were initially processed using Excel 2010. Normality and homogeneity of variance were used the shapiro.test and bartlett.test functions from the R “Stats” package. Subsequently, one-way ANOVA and Least Significant Difference (LSD) tests were performed using the aov function from the “Stats” package to evaluate the differences in vegetation and ecosystem functioning indicators among different grassland types. Statistical significance was set at P< 0.05 (Figures 2–4; Supplementary Figure S3).
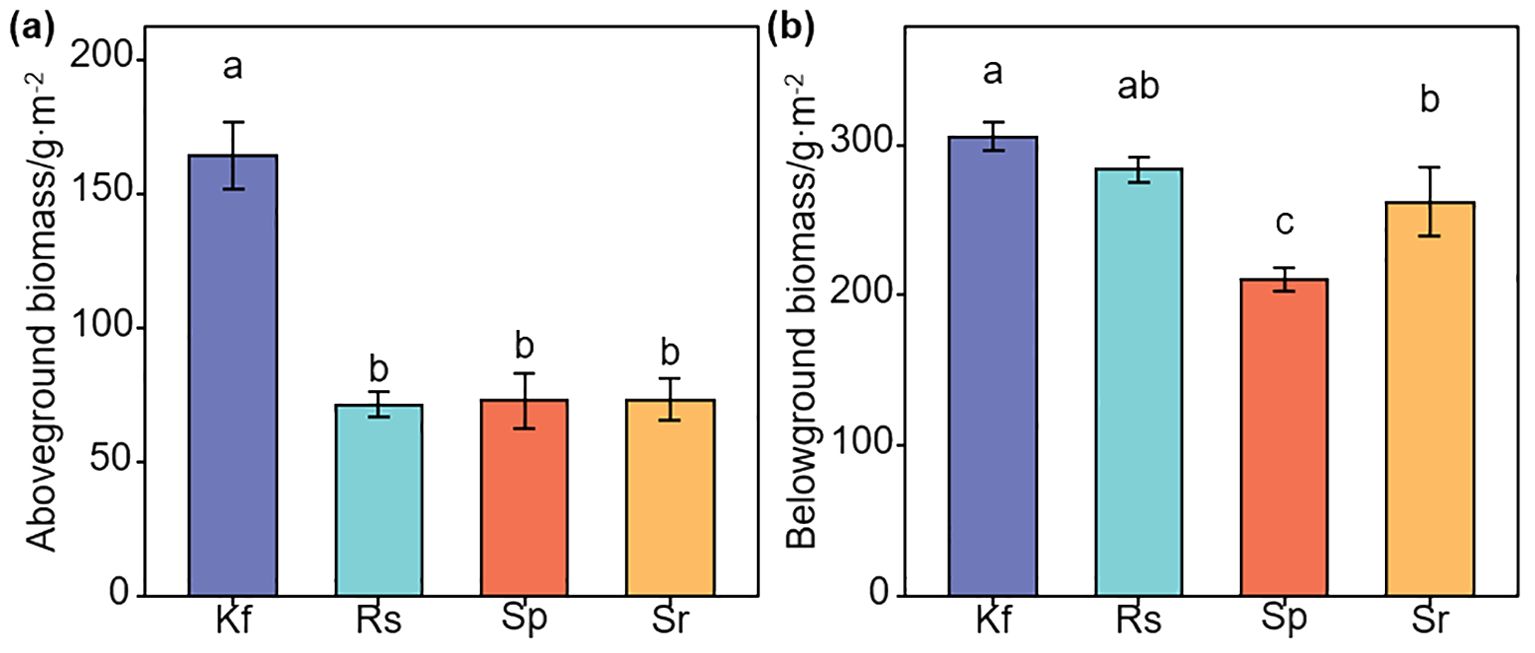
Figure 2. Effects of different grassland types on aboveground (a) and belowground (b) biomasses. Different lowercase letters indicate significant differences among the four grassland types (P< 0.05). Kf, K. foliatum type grassland; Rs, R. soongorica type grassland; Sp, S. passerina type grassland; Sr, S. regelii type grassland.
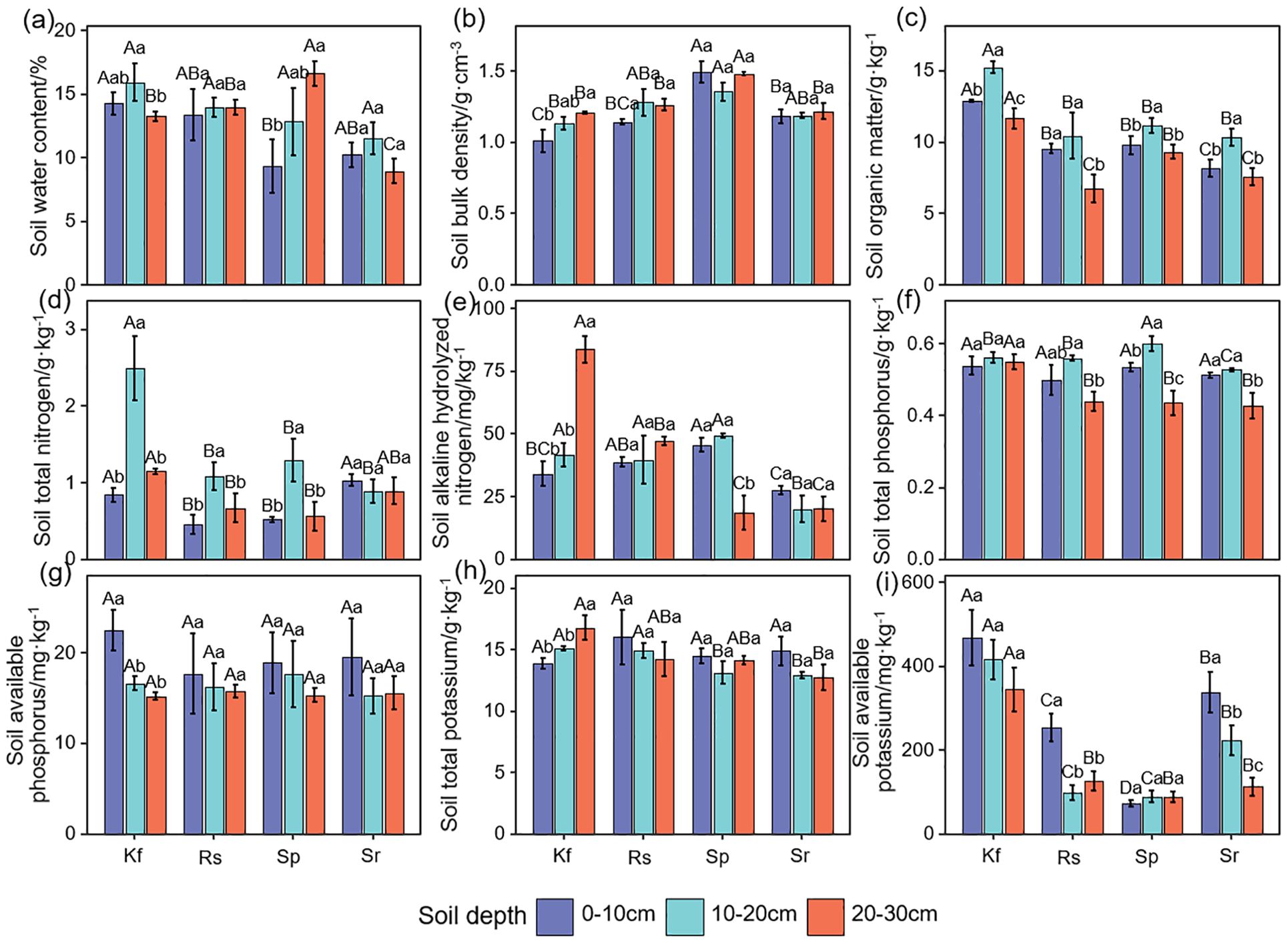
Figure 3. Effects of different grassland types on soil water content (a), bulk density (b), organic matter (c), total nitrogen (d), alkaline hydrolyzed nitrogen (e), total phosphorus (f), available phosphorus (g), total potassium (h), and available potassium (i). Different uppercase letters indicate significant differences among the four grassland types (P< 0.05); different lowercase letters indicate significant differences among different soil layers within the same grassland type (P< 0.05). Kf, K. foliatum type grassland; Rs, R. soongorica type grassland; Sp, S. passerina type grassland; Sr, S. regelii type grassland.
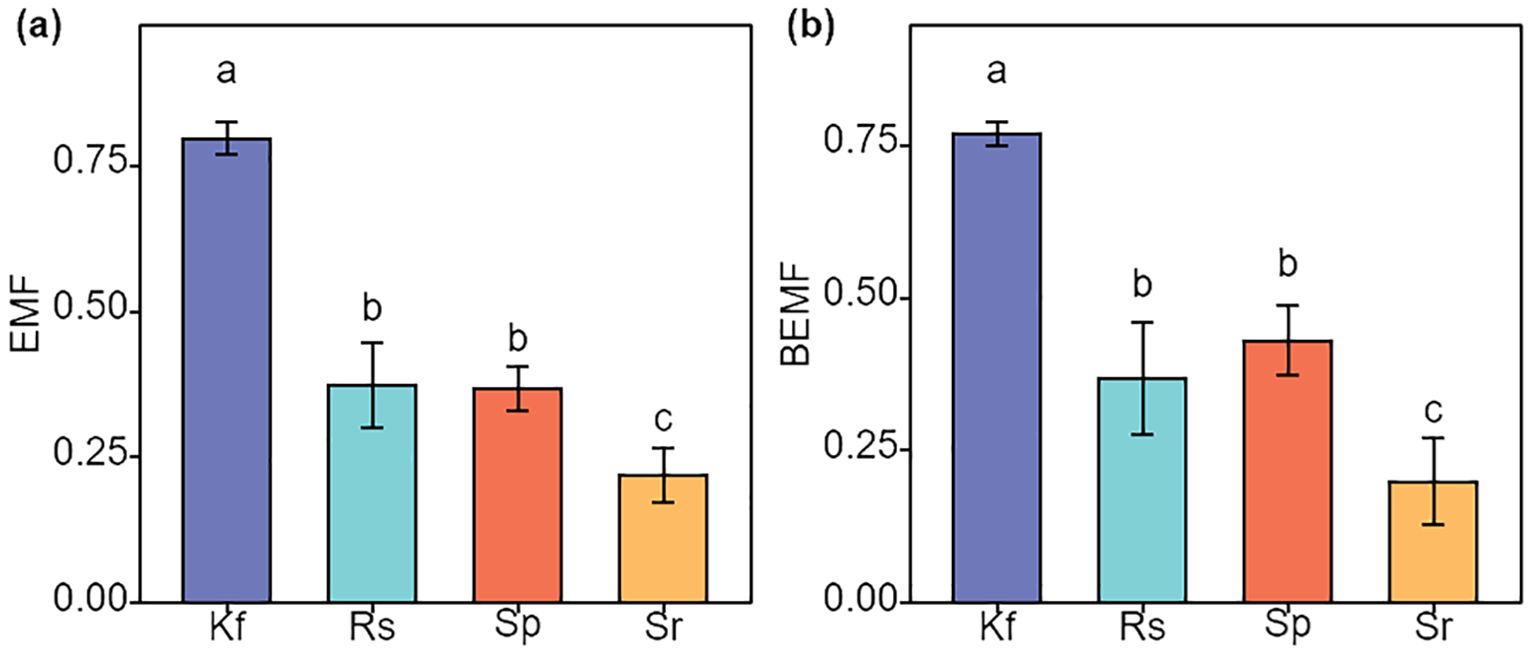
Figure 4. Effects of different grassland types on EMF, ecosystem multifunctionality (a), and BEMF, belowground ecosystem multifunctionality (b). Different lowercase letters indicate significant differences among the four grassland types (P< 0.05). Kf, K. foliatum type grassland; Rs, R. soongorica type grassland; Sp, S. passerina type grassland; Sr, S. regelii type grassland.
In this study, principal component analysis (PCA) was performed to reduce the dimensionality of the ecosystem function indicators. Three principal components with eigenvalues greater than 1 were extracted using the “FactoMineR” package (Figure 5). The contribution weights of each indicator to the principal components were calculated to assess their relative contributions to the EMF (Xiao et al., 2024).
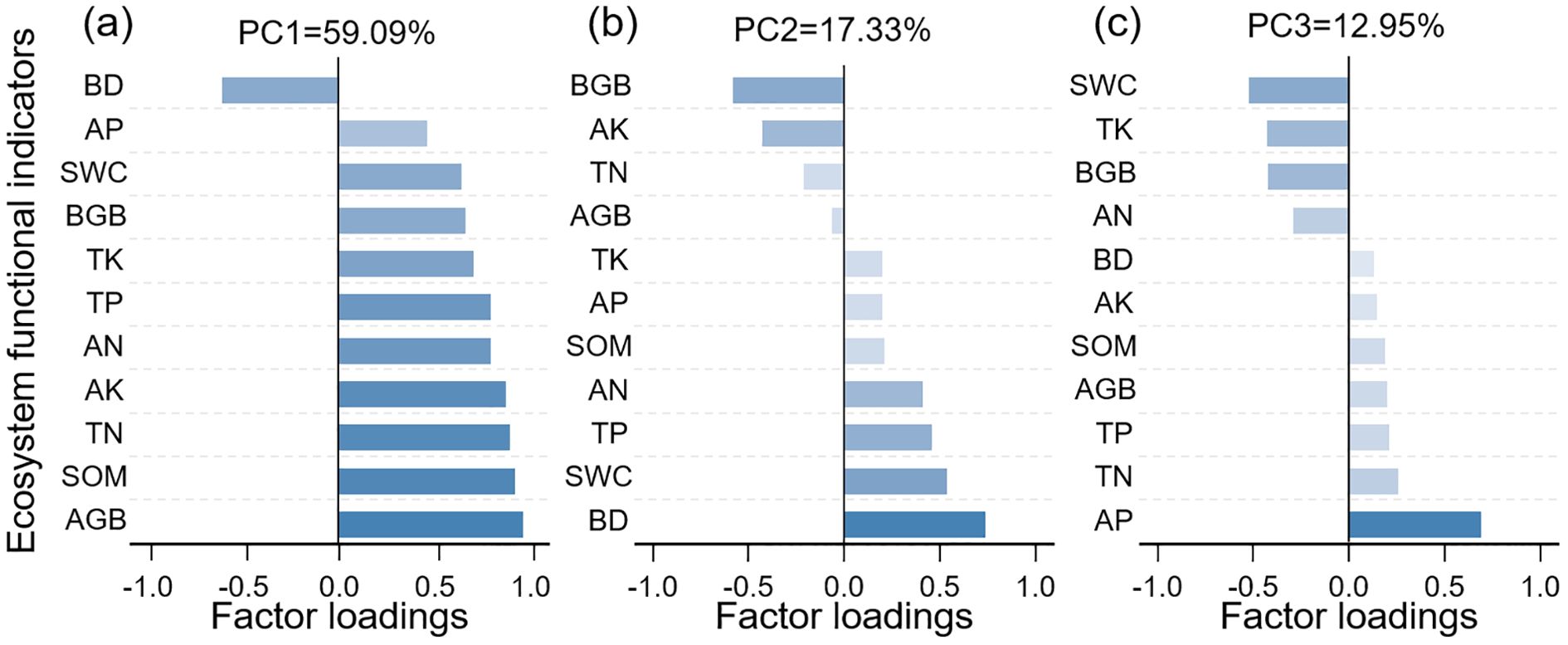
Figure 5. Relative contributions of single ecosystem functions to EMF. EMF: PC1 (a), PC2 (b), and PC3 (c), ecosystem multifunctionality; AGB, Aboveground biomass; BGB, Belowground biomass; SWC, Soil water content; BD, Soil bulk density; SOM, Soil organic matter; TN, Soil total nitrogen; AN, Soil alkaline hydrolyzed nitrogen; TP, Soil total phosphorus; AP, Soil available phosphorus; TK, Soil total potassium; AK, Soil available potassium.
Pearson correlation analysis was conducted using the “Hmisc” package to evaluate the relationships among ecosystem multifunctionality, plant community characteristics, soil properties, and ecosystem functional indicators (Figures 6; Supplementary Figure S4). To further investigate the data, hierarchical partitioning analysis using the “glmm.hp” package was performed to identify the key factors influencing the changes in EMF and BEMF (Figure 7) (Liu et al., 2025). Finally, a Partial Least Squares Path Model (PLS-PM) was utilized to explore the direct and indirect effects of soil factors, plant factors, and plant diversity index on EMF (Figure 8), based on a priori model (Supplementary Figure S5). The PLS-PM model was implemented using the R package “plspm” (He et al., 2025). Figures were produced using R version 4.3.2 and ArcGIS 10.2 software (Figure 1).
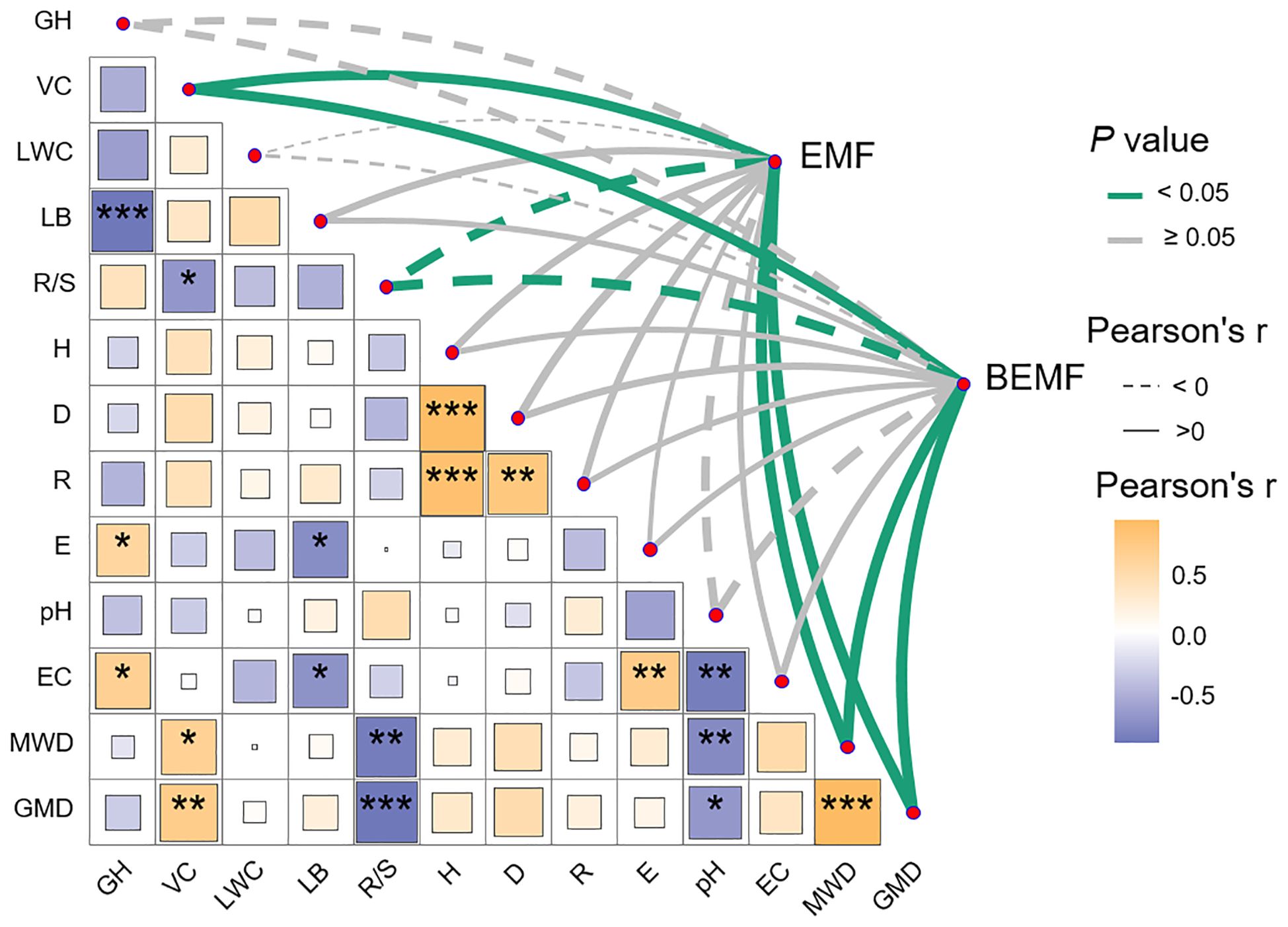
Figure 6. Pearson correlation analysis of ecosystem multifunctionality (EMF, BEMF), and plant and soil factors. EMF, Ecosystem multifunctionality; BEMF, Belowground ecosystem multifunctionality; GH, Height; VC, Coverage; LWC, Leaf water content; LB, Litter bionmass; R/S, Root-Shoot ratio; H, Shannon - Wiener index; D, Simpson index; R, Margalef richness index; E, Pielou evenness index; pH, Soil pH; EC, Soil electrical conductivity; MWD, Mean weight diameter; GMD, Geometric mean diameter. *p< 0.05; **p< 0.01; ***p< 0.001.
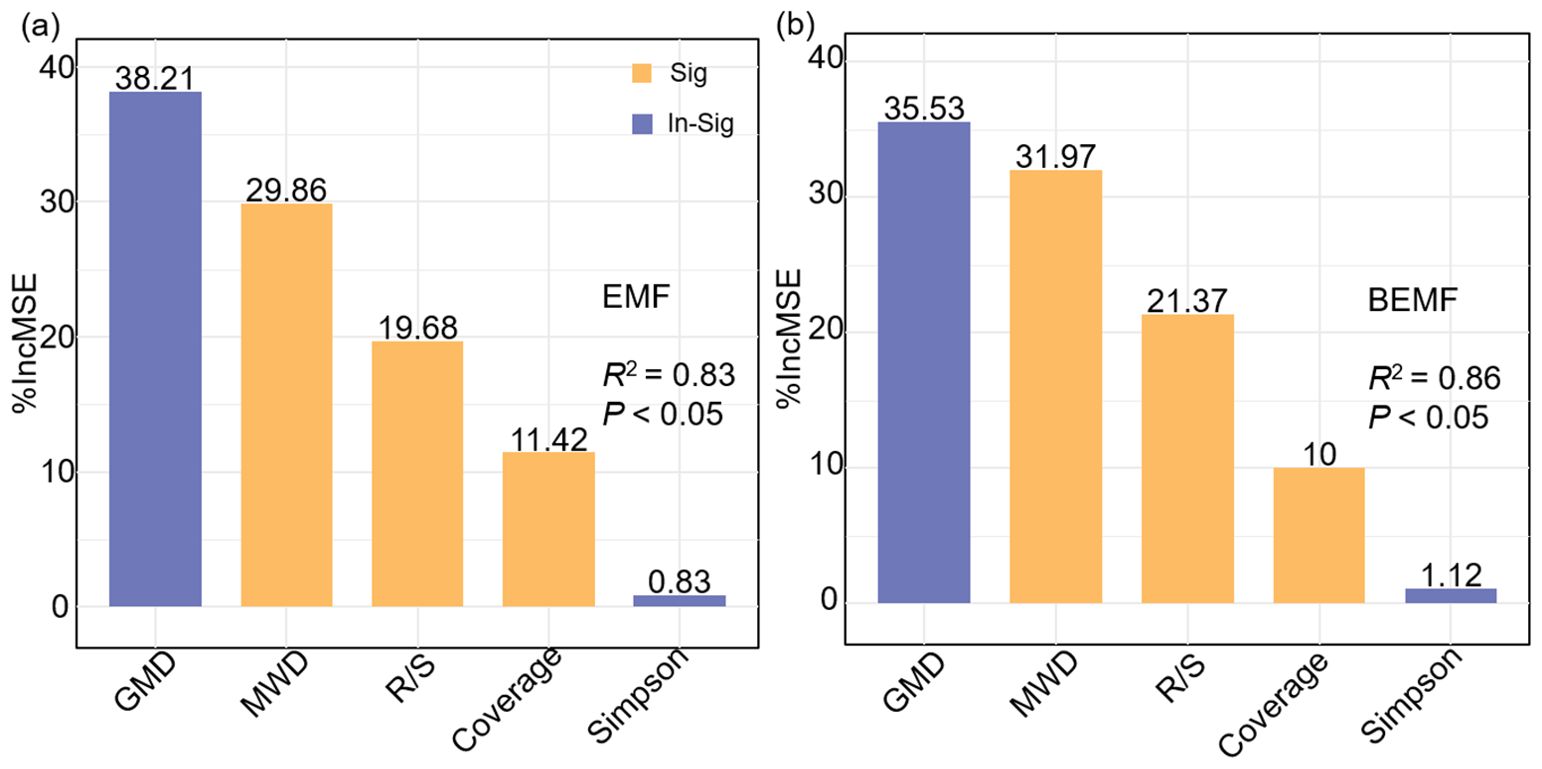
Figure 7. Importance of plant community characteristics and soil properties in EMF (a), and BEMF (b). EMF, Ecosystem multifunctionality; BEMF, Belowground ecosystem multifunctionality; R/S, Root-Shoot ratio; MWD, Mean weight diameter; GMD, Geometric mean diameter.
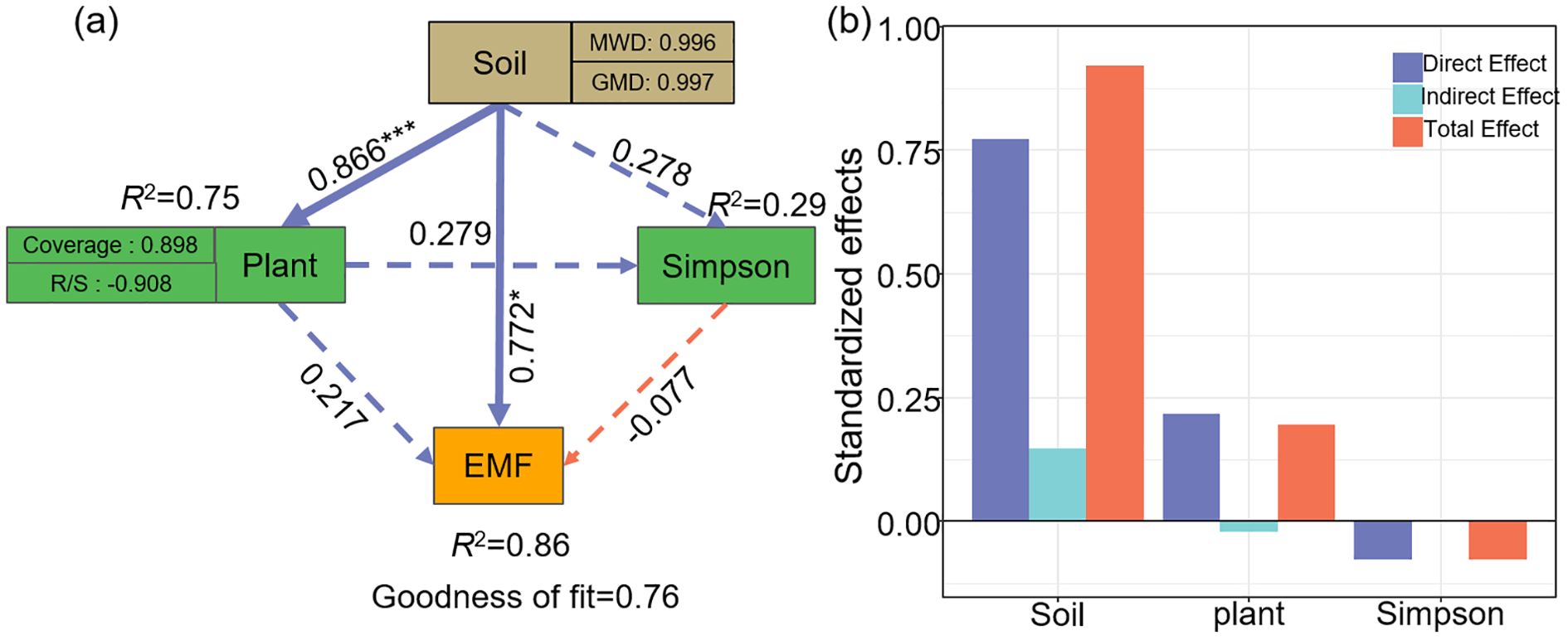
Figure 8. Effect of soil (MWD, GMD), plant (Coverage, Root-Shoot ratio), Simpson on EMF (a); Standardized effects of driving factors on EMF (b). Blue and orange arrows indicate positive and negative paths, respectively. Solid lines represent significant relationships, while dashed lines indicate insignificant relationships (*p< 0.05; ***p< 0.001). EMF, Ecosystem multifunctionality; R/S, Root-Shoot ratio; MWD, Mean weight diameter; GMD, Geometric mean diameter.
3 Results
3.1 Differences in plant community characteristics between grassland types
Plant community characteristics varied among grassland types. R. songarica and S. passerinum type grasslands had taller plants (P< 0.05, Supplementary Figure S3a), while coverage was highest in K. foliatum type grassland (P > 0.05; Supplementary Figure S3b). S. regelii type grassland had the highest leaf water content, and R. songarica type grassland the lowest (P< 0.05; Supplementary Figure S3c). Biomass was greatest in K. foliatum type grassland (Figures 2a, b; Supplementary Figure S3d), with root-shoot ratios highest in R. songarica type grassland (P< 0.05, Supplementary Figure S3e). Plant diversity indices showed the highest values in K. foliatum type grassland (Supplementary Figures S3f–h). S. passerinum type grassland had highest Pielou evenness index (Supplementary Figure S3i).
3.2 Differences in ecosystem functional indicators of different grassland types
As can be seen in Figure 3a, the soil water content was highest in the K. foliatum type grassland in both the 0–10 and 10–20 cm soil layers, while it was highest in the 20–30 cm in the S. passerina type grassland (P< 0.05). With the deepening of the soil layer, the soil water content of the K. foliatum type, R. soongorica type and S. regelii type grasslands showed a tendency of first increasing and then decreasing, and reached the maximum at 10–20 cm, while the S. passerina type grassland showed a tendency of gradually increasing. K. foliatum type grassland had the lowest soil bulk density in all soil horizons and was significantly lower (P< 0.05) than S. passerina type grassland, but not significantly different from R. soongorica type and S. regelii type grasslands (P > 0.05, Figure 3b). In the vertical direction of the soil layer, the soil bulk density of both K. foliatum type and S. regelii type grasslands showed a slow increasing trend with the depth of the soil layer.
There were significant differences in soil organic matter content between grassland types (Figure 3c). And the soil organic matter content showed a tendency of first increasing and then decreasing with the deepening of the soil layer. Soil organic matter content was highest (P< 0.05) in the K. foliatum type grassland in the 0–30 cm layer and lowest (P< 0.05) in the S. regelii type grassland.
Soil total nitrogen, alkaline hydrolyzed nitrogen, total phosphorus, available phosphorus, total potassium, and available potassium were overall highest in the K. foliatum type grassland for all soil horizons (Figures 3d–i). There was no significant difference in soil available phosphorus content among different grassland types (P > 0.05), and the rest of the ecosystem function indicators were significantly different (P< 0.05). With the deepening of the soil layer, the soil total nitrogen and total phosphorus content in each grassland type showed a trend of first increasing and then decreasing. Soil alkaline hydrolyzed nitrogen content showed that the K. foliatum type and R. soongorica type grassland gradually increased with the deepening of the soil layer, S. passerina type grassland firstly increasing and then decreasing, and S. regelii type grassland gradually decreasing. Soil available phosphorus and available potassium content tended to decrease with deepening of the soil layer. Soil total potassium content of the K. foliatum type grassland showed a gradual increase with deepening of the soil layer, the R. soongorica type and S. regelii type grassland showed a gradual decrease, and the S. passerina type grassland showed a first decreasing and then an increasing trend.
3.3 Differences in ecosystem multifunctionality of different grassland types
The mean method was used to calculate EMF, and BEMF and to compare the differences in EMF, and BEMF between the different grassland types. As can be seen from Figures 4a, b, the K. foliatum type grassland had EMF, and BEMF indices at 0.80 and 0.77, respectively, which were significantly higher (P< 0. 05) than in the R. soongorica type, S. passerina type and S. regelii type grassland. The R. soongorica type grassland did not have significantly difference in EMF and BEMF compared to S. passerina type grassland (P > 0.05), but were significantly higher (P< 0. 05) than S. regelii type grassland.
Three common factors were extracted from the 11 indicator variables (Figure 5), accounting for variance contributions of 59.09%, 17.33%, and 12.95%, respectively, with a cumulative contribution rate of 89.37%. Among them, aboveground biomass, soil organic matter, total nitrogen, and available potassium predominantly load on Factor 1 (Figure 5a). Soil bulk density, belowground biomass, and soil water content were primarily associated with Factor 2 (Figure 5b); and soil available phosphorus and soil water content mainly load on Factor 3 (Figure 5c). Regarding the contribution of individual ecosystem function indicators to EMF, soil available potassium, aboveground biomass, total nitrogen, and soil organic matter have relatively higher proportions (Figure 5).
3.4 Influence of plant community characteristics and soil properties on ecosystem multifunctionality
According to the correlation analysis, aboveground biomass, soil organic matter, and total nitrogen were significantly positively correlated with EMF, BEMF, mean weight diameter, and geometric mean diameter (P< 0.05), while showing a significant negative correlation with the root-shoot ratio (Supplementary Figure S4). Correlation analysis revealed that both EMF and BEMF were significantly positively correlated with coverage, mean weight diameter, and geometric mean diameter, while showing a significant negative correlation with the root-shoot ratio (P< 0.05). Furthermore, they exhibited a strong positive correlation with the Simpson index (Figure 6).
Based on the previous analysis, we selected vegetation factors strongly associated with ecosystem multifunctionality (Coverage and Root-Shoot ratio), Simpson index, and soil factors (mean weight diameter and geometric mean diameter) for hierarchical partitioning and the construction of a structural equation model. The results indicated that the most important explanatory factors for EMF were geometric mean diameter (38.21%) and mean weight diameter (29.86%). These explanatory factors collectively explained 83% (P< 0.05) of the EMF variation, with the most highly explained of the explanatory factors being soil geometric mean diameter (Figure 7a). The results obtained from screening the explanatory factors of BEMF were found to be more similar to those obtained from the screening of EMF. Among them, the soil geometric mean diameter (35.53%) and mean weight diameter (31.97%) showed a higher degree of explanation compared to other indicators of BEMF (Figure 7b). From the results in different hierarchical partitioning analysis, soil factors (mean weight diameter and geometric mean diameter) can better explain the variation in EMF.
Structural equation modelling (SEM) analysis further indicated that soil factors (mean weight diameter and geometric mean diameter), plant factors (coverage and root-shoot ratio), and Simpson diversity index collectively explained 86% of the variation in EMF (R2 = 0.86, Figure 8a). On the one hand, soil factors directly influenced EMF (standardized coefficient: β = 0.772), exhibiting a significant positive effect (P< 0.05). On the other hand, soil factors indirectly enhanced EMF by improving plant factors and Simpson diversity index. The total effects of the soil factors, plant factors, and Simpson diversity index on EMF were 0.921, 0.196, and -0.077, respectively (Figure 8b).
4 Discussion
4.1 Effects of different grassland types on plant community characteristics, ecological function indicators and ecosystem multifunctionality
This study selected four grassland types on the Loess Plateau (K. foliatum type, R. songarica type, S. passerinum type, and S. regelii type) to analyze plant community characteristics, ecological function indicators, and ecosystem multifunctionality. The study found that the height of plants in the research area ranged from 21 to 36 cm, and the vegetation coverage ranged from 28% to 36%. This indicates that the coverage in the temperate desert area is relatively low, which is consistent with the findings of Zhang and Zhao (2015) in the Hexi Corridor region of Gansu. Leaf water content reflects the drought resistance and environmental adaptability of desert plants (Wang et al., 2021). This study found that grassland types dominated by R. songarica exhibited the lowest leaf water content, while those dominated by S. regelii showed the highest. These findings are consistent with Li et al. (2020), who also reported that grasslands with R. songarica as the dominant species had lower leaf water content compared to other grassland types. The root - shoot ratio is used to evaluate the allocation characteristics of aboveground and belowground biomass in plants. A high root - shoot ratio indicates that plants allocate more resources to their root systems, while a low root - shoot ratio suggests that more resources are invested in the growth of the aboveground parts (Qi et al., 2019; Wang et al., 2023b). This study indicated that the root - shoot ratio is highest in the R. songarica type grassland, primarily because of the well-developed root systems of the dominant species, R. songarica, combined with a lower aboveground biomass (Figure 2a), which enables better adaptation to arid environments.
This study found that temperate desert grassland communities of different types exhibited relatively low biodiversity, with significant differences in diversity indices, although their overall variation patterns were largely consistent. The K. foliatum type grassland showed the highest values for the Shannon - Wiener, Simpson, and Margalef richness indices, whereas the community dominated by S. regelii exhibited the lowest values for all diversity indices. This is primarily because the grassland type dominated by S. regelii has a more homogeneous species assembly compared to other grassland types, with poor species distribution uniformity, resulting in a pronounced single-species community and poor performance in species diversity. The study by Dong et al. (2020) found that among eight typical shrub communities in the western section of the Hexi Corridor in Gansu, the diversity index of the K. foliatum community was the highest, while the vegetation communities had few species, relatively simple community structures, and very uneven species distribution, which is consistent with the results of this study. Osmonali et al. (2023) also found that the species assembly in desert plant communities in Kazakhstan is primarily dominated by a single dominant community. Additionally, the relatively single species assembly of the study area is closely related to the semi-arid climate of Longzhong. This has naturally led to the formation of a temperate desert community dominated by a single species, adapted to the arid and water-scarce habitat conditions (Li et al., 2024b). The grassland is primarily dominated by K. foliatum, R. songarica, S. passerinum, and S. regelii (Li et al., 2024c), which are minimally affected by external environmental influences, resulting in highly stable community structures. Moreover, temperate desert plants possess drought, cold, salt-alkali tolerance, and resilience to poor soils, enabling them to better adapt to the harsh, dry, and windy conditions with nutrient-deficient soils typical of Longzhong. In summary, the aboveground biomass of temperate desert vegetation in Longzhong is relatively low, while the belowground biomass accounts for a higher proportion. The litter biomass is the lowest, and overall vegetation coverage is low, resulting in relatively low plant diversity. The grassland dominated by K. foliatum generally exhibits higher ecological value. Thus, K. foliatum may play a positive role in the restoration of temperate desert vegetation.
This study analyzed the distribution characteristics of soil properties across different grassland types in the temperate desert of Longzhong. The results showed that the K. foliatum type grasslands had higher soil water content and lower bulk density, with bulk density increasing gradually with soil depth. This trend is attributed to the extensive root systems of K. foliatum, which primarily extend and intertwine within the surface soil, loosening the soil and thereby reducing the surface soil bulk density (Mueller et al., 2024). Significant differences in soil bulk density were observed among various grassland types within the same soil layer. This variation may be attributed to the different dominant plant species and their root distribution patterns across the grassland types, which consequently lead to differences in soil bulk density (Chen et al., 2024). Soil nutrients provide essential elements necessary for plant growth and development (Zeng et al., 2016; Li et al., 2024c). It is generally recognized that the primary sources of soil nutrients in grasslands mainly rely on plant residues and root secretions (Ochoa-Hueso et al., 2020). This study found that the overall nutrient content of soils in the temperate desert of the Longzhong region is relatively low, and the distribution characteristics among different soil layers and grassland types with different dominant plant species show a high consistency with previous research findings (Lu et al., 2023). Among them, the grassland type with K. foliatum as the dominant plant had the highest soil organic matter, total nitrogen and total phosphorus content. This is mainly due to the higher plant aboveground biomass, litter biomass and belowground biomass in the grassland type with K. foliatum as the dominant plant. Inputs of litter and root residues increase soil nutrient fixation in K. foliatum type grasslands (Li et al., 2021). At the same time, soil organic matter, total nitrogen and total phosphorus overall with the deepening of the soil layer showed a trend of first increasing and then decreasing, the highest in the 10–20 cm soil layer. This is due to the soil nutrients are mainly affected by the plant root system, the soil-forming parent material and other factors (Anderson, 1988). Together with the fact that the roots of the dominant plants of the temperate desert (K. foliatum, R. songarica, S. passerinum, and S. regelii) are mainly distributed in the sub-surface layer (10–20 cm), this results in a high level of nutrient content. Soil available nutrients, as key elements that plants can directly absorb, play a crucial role in plant growth and development (Li et al., 2024e). This study found that in the temperate desert of Longzhong, all four grassland types with K. foliatum as the dominant species exhibited higher levels of soil available nutrients. This is primarily attributed to the higher vegetation coverage, height, litter and belowground biomass, as well as elevated levels of total nutrients in these grasslands, which facilitate the accumulation of available nutrients and result in their higher content. In the vertical direction, the variation patterns of available phosphorus and available potassium are consistent, both showing a gradual decrease with increasing soil depth. This indicated that the temperate desert in Longzhong has a high efficiency in absorbing and utilizing nutrients from the soil surface layer. Conversely, the content of soil alkaline hydrolyzed nitrogen exhibits significant differences in its vertical distribution across different grassland types. These variations are influenced by factors such as vegetation coverage, dominant plant species, and plant characteristics.
The analysis of ecosystem multifunctionality indicated that the K. foliatum type grassland has the highest multifunctionality at both the overall and belowground ecosystem levels. The R. songarica type and S. passerinum type grasslands were ranked second, while the S. regelii type grassland has the lowest multifunctionality (Figure 4). This suggested that K. foliatum plants have a higher potential for restoring the multifunctionality of temperate desert ecosystems. The reasons for differences in ecosystem multifunctionality among the four grassland types may stem from significant variations in their soil physical and chemical properties. Such heterogeneity can influence key ecological processes, such as nutrient cycling and energy flow, thereby affecting the stability of the grassland ecosystem (Wang et al., 2023a). For example, the lower plant biomass in S. regelii type grasslands reduces the vegetation’s capacity to protect the soil.
4.2 Influence of plant and soil factors on ecosystem multifunctionality
Existing studies have identified plant community characteristics as the main factor regulating EMF (Xu et al., 2024; Yao et al., 2025). Among them, a significant positive correlation between plant diversity and EMF is now a widely shared view (Maestre et al., 2012b). Also studies on the Loess Plateau have shown that plant diversity can sometimes better explain EMF (Wang et al., 2022). This study also found that ecosystem multifunctionality showed an increasing trend with the rise in plant diversity indices (Figure 6). The relationship between plant diversity and EMF is altered by factors such as ecosystem type and disturbance factors (Dietrich et al., 2024). It has been demonstrated that the effect of species richness on EMF is dependent on the evenness of species in the community (Maestre et al., 2012b). In this study, plant root-shoot ratio showed a significant negative correlation with ecosystem multifunctionality, while vegetation coverage, mean weight diameter, and geometric mean diameter exhibited significant positive correlations. Additionally, the Simpson diversity index had a relatively strong positive correlation with ecosystem multifunctionality (Figure 6). It also showed that the plant community characteristics and soil properties influence ecosystem multifunctionality. It also showed that soil factors, such as mean weight diameter and geometric mean diameter, have a high explanatory rates for variations in ecosystem multifunctionality (Figure 7), indicating that soil aggregates play a key role in influencing ecosystem multifunctionality in the temperate desert regions of the Loess Plateau. Similar studies also found that soil conditions are primary factors determining ecosystem characteristics like plant community primary productivity (Van Breemen, 1993).
The results of the structural equation model indicated that environmental factors explain up to 86% of the variation in ecosystem multifunctionality (Figure 8). Soil factors, such as mean weight diameter and geometric mean diameter, have a direct positive effect on ecosystem multifunctionality. The productivity of arid and semi-arid ecosystems is significantly influenced by soil structure (Khasi et al., 2024). Han et al. (2021) found that soil aggregate structure is an important driver in shaping the grassland ecosystem multifunctionality. Our study also found that soil aggregates not only directly influence ecosystem multifunctionality, but also indirectly influence ecosystem multifunctionality by influencing plant factors and Simpson diversity index. This is mainly due to the fact that the mean weight diameter and geometric mean diameter, as indicators of soil aggregate stability, not only reflect the resistance of the soil structure to maintain its original form when exposed to external forces and environmental changes, but are also an important indicator for assessing the soil quality (Dou et al., 2020; Xia et al., 2022). Due to the low soil water content in the top layer of temperate deserts, there is a greater contribution to grassland nutrient content from the soil aggregate formation and stabilization of desert plants during their growth process (Zhang et al., 2024). This in turn affects plant growth and development, leading to the production of large amounts of plant residues, and this increased organic matter input affects plant diversity index, further affecting the grassland EMF (Wang et al., 2023a; Pichon et al., 2024). Improvements in soil structure may increase soil water and nutrient content, which may make it more sensitive to changes in EMF (Tian et al., 2024). In recent years, the temperate deserts of the Loess Plateau have become warmer and drier as a result of rising temperatures and decreasing precipitation (Miao et al., 2016). Although as reported in our study, K. foliatum desert plants favor the improvement of EMF indices, prolonged aridification in these areas may also greatly diminish their positive impact on ecosystem functions. Overall, K. foliatum desert plants can serve as a practical tool for enhancing ecosystem function in less arid grasslands, but it limits the ability of plant diversity to support multiple ecosystem functions. Our results provide valuable information for managing grasslands for biodiversity conservation.
5 Conclusion
Our study demonstrated that EMF, and BEMF showed the highest values in K. foliatum type grassland. Furthermore, soil factors (mean weight diameter, and geometric mean diameter) were more explanatory of EMF, suggesting that soil factors can more effectively predict EMF. Soil factors not only directly influence EMF, but may also indirectly drive changes in EMF through plant factors (coverage, root-shoot ratio) or Simpson diversity index. Therefore, in the process of ecological restoration, it should pay attention to the restoration of the interaction relationship between soil factors and plant factors. Meanwhile, K. foliatum plants have an important and positive role in achieving the restoration of desert grasslands on the Loess Plateau.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author contributions
YL: Data curation, Methodology, Investigation, Software, Writing – review & editing, Writing – original draft. GH: Investigation, Writing – review & editing, Methodology. XL: Supervision, Funding acquisition, Writing – review & editing. TJ: Methodology, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Open Competition Projects to Select the Best Candidates for Leading Key Initiatives of the Key Laboratory of Grassland Ecosystems, Gansu Agricultural University, Ministry of Education (KLGE-2024-01), Grassland Monitoring and Evaluation in Gansu Province (GSZYTC-ZCJC-21010), Self-Listed Provincial Forestry and Grassland Science and Technology Project in 2021 (2021kj071), A New Round of Grassland Reward and Subsidy Benefit Evaluation and Grassland Ecological Evaluation in Gansu Province (XZ20191225), 2023 Early Batch of Central Financial Forestry Reform and Development Funds Grassland Technology Support Project - Grassland Ecological Health Assessment in the Fragile Ecological Zone of the Hexi Corridor” (2023211); Research on the scope of desertification prevention and ecological restoration in the Shage Desert New Energy Base, sand transport, new materials and technologies, and technology integration demonstration (GSAU-JSFW-2024-116).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1619948/full#supplementary-material
References
Anderson, D. W. (1988). The effect of parent material and soil development on nutrient cycling in temperate ecosystems. Biogeochemistry 5, 71–97. doi: 10.1007/BF02180318
Bardgett, R. D., Bullock, J. M., Lavorel, S., Manning, P., Schaffner, U., Ostle, N., et al. (2021). Combatting global grassland degradation. Nat. Rev. Earth Environ. 2, 720–735. doi: 10.1038/s43017-021-00207-2
Bezemer, T. M., Lawson, C. S., Hedlund, K., Edwards, A. R., Brook, A. J., Igual, J. M., et al. (2006). Plant species and functional group effects on abiotic and microbial soil properties and plant–soil feedback responses in two grasslands. J. Ecol. 94, 893–904. doi: 10.1111/j.1365-2745.2006.01158.x
Cambardella, C. A. and Elliott, E. T. (1993). Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci. Soc Am. J. 57, 1071–1076. doi: 10.2136/sssaj1993.03615995005700040032x
Chen, L., Ding, M., Zhang, H., Wang, N., Zhang, Y., Wan, J., et al. (2024). Effects of dominant plant species change on soil multifunctionality in alpine meadows of the three-river source region. J. Cleaner Prod. 434, 139888. doi: 10.1016/j.jclepro.2023.139888
Cheng, Z., Cui, Z., Shi, J., Liu, Y., La Pierre, K. J., and Wu, G. (2021). Plant functional types drive differential responses of grassland ecosystem functions along a precipitation gradient. Ecol. Indic. 133, 108433. doi: 10.1016/j.ecolind.2021.108433
Delgado-Baquerizo, M., Reich, P. B., Trivedi, C., Eldridge, D. J., Abades, S., Alfaro, F. D., et al. (2020). Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 4, 210–220. doi: 10.1038/s41559-019-1084-y
Dietrich, P., Ebeling, A., Meyer, S. T., Asato, A. E. B., Bröcher, M., Gleixner, G., et al. (2024). Plant diversity and community age stabilize ecosystem multifunctionality. Global Change Biol. 30, e17225. doi: 10.1111/gcb.17225
Dong, X., Li, Y., Xin, Z., Duan, R., Yao, B., Bao, Y., et al. (2020). Gobi shrub species diversity and its distribution pattern in west Hexi Corridor. Arid. Land. Geogr. 43, 1514–1522. doi: 10.12118/j.issn.1000-6060.2020.06.12
Dou, Y., Yang, Y., An, S., and Zhu, Z. (2020). Effects of different vegetation restoration measures on soil aggregate stability and erodibility on the Loess Plateau, China. Catena 185, 104294. doi: 10.1016/j.catena.2019.104294
Gang, C., Zhao, W., Zhao, T., Zhang, Y., Gao, X., and Wen, Z. (2018). The impacts of land conversion and management measures on the grassland net primary productivity over the Loess Plateau, Northern China. Sci. Total Environ. 645, 827–836. doi: 10.1016/j.scitotenv.2018.07.161
Han, S., Delgado-Baquerizo, M., Luo, X., Liu, Y., Van Nostrand, J. D., Chen, W., et al. (2021). Soil aggregate size-dependent relationships between microbial functional diversity and multifunctionality. Soil Biol. Biochem. 154, 108143. doi: 10.1016/j.soilbio.2021.108143
He, G., Liu, X., Li, Y., and Ji, T. (2025). Response of plant life forms and soil physical properties to near-natural restoration measures in alpine grassland, Tibetan plateau. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1504754
He, G., Liu, X., Li, Y., Xu, H., Ji, T., Yang, Z., et al. (2024). Recovery in soil carbon stocks but reduced carbon stabilization after near-natural restoration in degraded alpine meadows. Sci. Rep. 14, 31124. doi: 10.1038/s41598-024-82434-3
Hector, A. and Bagchi, R. (2007). Biodiversity and ecosystem multifunctionality. Nature 448, 188–190. doi: 10.1038/nature05947
Huang, W., Zhu, Y., Yu, H., He, Y., Zhao, X., Wang, H., et al. (2024). Biodiversity drives ecosystem multifunctionality in sandy grasslands? Sci. Total Environ. 925, 171765. doi: 10.1016/j.scitotenv.2024.171765
Jiang, C., Wang, F., Zhang, H., and Dong, X. (2016). Quantifying changes in multiple ecosystem services during 2000–2012 on the Loess Plateau, China, as a result of climate variability and ecological restoration. Ecol. Eng. 97, 258–271. doi: 10.1016/j.ecoleng.2016.10.030
Khasi, Z., Askari, M. S., Amanifar, S., and Moravej, K. (2024). Assessing soil structural quality as an indicator of productivity under semi-arid climate. Soil Tillage Res. 236, 105945. doi: 10.1016/j.still.2023.105945
Li, Y., He, G., Liu, X., Xu, H., Ji, T., Lin, D., et al. (2024b). Distribution characteristics of carbon density in Plant–Soil system of Temperate Steppe and Temperate Desert in the Longzhong Loess Plateau. Agriculture 14, 177. doi: 10.3390/agriculture14020177
Li, Y., He, G., Liu, X., Zhang, D., Xu, H., Ji, T., et al. (2024c). Distribution characteristics of soil aggregate stability and organic carbon of different grassland types in the temperate desert of Longzhong Loess Plateau. Chin. J. Envir. Sci. 45, 5431–5440. doi: 10.13227/j.hjkx.202309164
Li, Y., He, G., Liu, X., Zhang, D., Xu, H., Ji, T., et al. (2024d). Variation characteristics of soil organic carbon fraction and carbon pool activity in four grassland types in the temperate desert of Longzhong. Acta Agrestia Sin. 32, 1489–1499. doi: 10.11733/j.issn.1007-0435.2024.05.019
Li, Y., Li, Y., Zhang, Q., Xu, G., Liang, G., Kim, D.-G., et al. (2024e). Enhancing soil carbon and nitrogen through grassland conversion from degraded croplands in China: Assessing magnitudes and identifying key drivers of phosphorus reduction. Soil Tillage Res. 236, 105943. doi: 10.1016/j.still.2023.105943
Li, X., Liu, Y., Wu, G., Lie, Z., Sheng, H., Aguila, L. C. R., et al. (2024a). Mixed plantations do not necessarily provide higher ecosystem multifunctionality than monoculture plantations. Sci. Total Environ. 914, 170156. doi: 10.1016/j.scitotenv.2024.170156
Li, S., Wang, H., Gou, W., Wu, G., and Su, P. (2020). Relationship between leaf functional traits of mixed desert plants and microbial diversity in rhizosphere. Ecol. Environ. Sci. 29, 1713–1722. doi: 10.16258/j.cnki.1674-5906.2020.09.001
Li, M., Wang, J., Yao, T., Zhang, T., Zhou, Q., and Mihray·, M. (2021). Bacterial diversity and community structure in the rhizosphere of four halophytes. Curr. Microbiol. 78, 2720–2732. doi: 10.1007/s00284-021-02536-3
Liu, Y., Li, C., Zhao, X., Liu, W., Yang, X., Lv, W., et al. (2025). Fungal diversity shapes ecosystem multifunctionality in alpine grasslands under different herbivore assemblages: A case study from the Qinghai-Tibetan Plateau. Catena 256, 109109. doi: 10.1016/j.catena.2025.109109
Liu, S., Wu, J., Li, G., Yang, C., Yuan, J., and Xie, M. (2022). Seasonal freeze-thaw characteristics of soil carbon pools under different vegetation restoration types on the Longzhong Loess Plateau. Front. Ecol. Evol. 10. doi: 10.3389/fevo.2022.1019627
Lu, J., Feng, S., Wang, S., Zhang, B., Ning, Z., Wang, R., et al. (2023). Patterns and driving mechanism of soil organic carbon, nitrogen, and phosphorus stoichiometry across northern China’s desert-grassland transition zone. Catena 220, 106695. doi: 10.1016/j.catena.2022.106695
Maestre, F. T., Castillo-Monroy, A. P., Bowker, M. A., and Ochoa-Hueso, R. (2012a). Species richness effects on ecosystem multifunctionality depend on evenness, composition and spatial pattern. J. Ecol. 100, 317–330. doi: 10.1111/j.1365-2745.2011.01918.x
Maestre, F. T., Quero, J. L., Gotelli, N. J., Escudero, A., Ochoa, V., Delgado-Baquerizo, M., et al. (2012b). Plant species richness and ecosystem multifunctionality in global drylands. Science 335, 214–218. doi: 10.1126/science.1215442
Miao, C., Sun, Q., Duan, Q., and Wang, Y. (2016). Joint analysis of changes in temperature and precipitation on the Loess Plateau during the period 1961–2011. Clim. Dyn. 47, 3221–3234. doi: 10.1007/s00382-016-3022-x
Mueller, C. W., Baumert, V., Carminati, A., Germon, A., Holz, M., Kögel-Knabner, I., et al. (2024). From rhizosphere to detritusphere – Soil structure formation driven by plant roots and the interactions with soil biota. Soil Biol. Biochem. 193, 109396. doi: 10.1016/j.soilbio.2024.109396
Novák, J., Jankowski, K., Sosnowski, J., Malinowska, E., and Wiśniewska-Kadżajan, B. (2020). Influence of plant species and grasslands quality on sequestration of soil organic carbon. Ekol. (Bratislava) 39, 289–300. doi: 10.2478/eko-2020-0023
Ochoa-Hueso, R., Borer, E. T., Seabloom, E. W., Hobbie, S. E., Risch, A. C., Collins, S. L., et al. (2020). Microbial processing of plant remains is co-limited by multiple nutrients in global grasslands. Global Change Biol. 26, 4572–4582. doi: 10.1111/gcb.15146
Osmonali, B. B., Vesselova, P. V., Kudabayeva, G. M., Skaptsov, M. V., Shmakov, A. I., and Friesen, N. (2023). Phylogeny and flow cytometry of the genus Kalidium Moq. (Amaranthaceae s.l.) in Kazakhstan. Plants 12, 2619. doi: 10.3390/plants12142619
Peng, Y., Fan, M., Song, J., Cui, T., and Li, R. (2018). Assessment of plant species diversity based on hyperspectral indices at a fine scale. Sci. Rep. 8, 4776. doi: 10.1038/s41598-018-23136-5
Pichon, N. A., Cappelli, S. L., Soliveres, S., Mannall, T., Nwe, T. Z., Hölzel, N., et al. (2024). Nitrogen availability and plant functional composition modify biodiversity-multifunctionality relationships. Ecol. Lett. 27, e14361. doi: 10.1111/ele.14361
Qi, Y., Wei, W., Chen, C., and Chen, L. (2019). Plant root-shoot biomass allocation over diverse biomes: A global synthesis. Global Ecol. Conserv. 18, e00606. doi: 10.1016/j.gecco.2019.e00606
Shu, X., Liu, W., Hu, Y., Xia, L., Fan, K., Zhang, Y., et al. (2023). Ecosystem multifunctionality and soil microbial communities in response to ecological restoration in an alpine degraded grassland. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1173962
Soliveres, S., van der Plas, F., Manning, P., Prati, D., Gossner, M. M., Renner, S. C., et al. (2016). Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality. Nature 536, 456–459. doi: 10.1038/nature19092
Sun, W., Song, X., Mu, X., Gao, P., Wang, F., and Zhao, G. (2015). Spatiotemporal vegetation cover variations associated with climate change and ecological restoration in the Loess Plateau. Agric. For. Meteorol. 209–210, 87–99. doi: 10.1016/j.agrformet.2015.05.002
Tian, J., Ni, T., Miao, Z., Hao, Y., Ma, A., Dong, L., et al. (2024). Effects of biotic and abiotic factors on ecosystem multifunctionality of plantations. Ecol. Process 13, 44. doi: 10.1186/s13717-024-00525-5
Van Breemen, N. (1993). Soils as biotic constructs favouring net primary productivity. Geoderma 57, 183–211. doi: 10.1016/0016-7061(93)90002-3
Wang, X., Chen, X., Xu, J., Ji, Y., Du, X., and Gao, J. (2023b). Precipitation dominates the allocation strategy of above- and belowground biomass in plants on macro scales. Plants 12, 2843. doi: 10.3390/plants12152843
Wang, R., He, N., Li, S., Xu, L., and Li, M. (2021). Variation and adaptation of leaf water content among species, communities, and biomes. Environ. Res. Lett. 16, 124038. doi: 10.1088/1748-9326/ac38da
Wang, M., Lu, N., An, N., and Fu, B. (2022). Plant functional and phylogenetic diversity regulate ecosystem multifunctionality in Semi-Arid grassland during succession. Front. Environ. Sci. 9. doi: 10.3389/fenvs.2021.791801
Wang, L., Pang, X., Li, N., Qi, K., Huang, J., and Yin, C. (2020). Effects of vegetation type, fine and coarse roots on soil microbial communities and enzyme activities in eastern Tibetan plateau. Catena 194, 104694. doi: 10.1016/j.catena.2020.104694
Wang, C., Yu, W., Ma, L., Ye, X., Erdenebileg, E., Wang, R., et al. (2023a). Biotic and abiotic drivers of ecosystem multifunctionality: Evidence from the semi-arid grasslands of northern China. Sci. Total Environ. 887, 164158. doi: 10.1016/j.scitotenv.2023.164158
Wu, G., Cheng, Z., Alatalo, J. M., Zhao, J., and Liu, Y. (2021). Climate warming consistently reduces grassland ecosystem productivity. Earths Future 9, e2020EF001837. doi: 10.1029/2020EF001837
Xia, R., Shi, D., Ni, S., Wang, R., Zhang, J., and Song, G. (2022). Effects of soil erosion and soil amendment on soil aggregate stability in the cultivated-layer of sloping farmland in the Three Gorges Reservoir area. Soil Tillage Res. 223, 105447. doi: 10.1016/j.still.2022.105447
Xiao, Y., Liu, M., Miao, L., Yang, C., Wang, Q., and Wang, M. (2024). Plant diversity and ecosystem multifunctionality of different degraded grasslands in alpine meadows of Maqu. Ecol. Res. 39, 54–71. doi: 10.1111/1440-1703.12417
Xie, M., Yuan, J., Liu, S., Xu, G., Lu, Y., Yan, L., et al. (2024). Soil carbon and nitrogen pools and their storage characteristics under different vegetation restoration types on the Loess Plateau of Longzhong, China. Forests 15, 173. doi: 10.3390/f15010173
Xiong, M., Li, F., Liu, X., Liu, J., Luo, X., Xing, L., et al. (2023). Characterization of ecosystem services and their Trade-Off and synergistic relationships under different Land-Use scenarios on the Loess Plateau. Land 12, 2087. doi: 10.3390/land12122087
Xu, Z., Guo, X., Allen, W. J., Yu, X., Hu, Y., Wang, J., et al. (2024). Plant community diversity alters the response of ecosystem multifunctionality to multiple global change factors. Global Change Biol. 30, e17182. doi: 10.1111/gcb.17182
Yao, Z., Hu, M., Shi, L., Wu, Q., Zhang, D., Liu, G., et al. (2025). Driving factors of plant and soil properties on ecosystem multifunctionality vary among grassland types in the Qinghai-Tibetan Plateau. Plant Soil. doi: 10.1007/s11104-025-07229-5
Yinglan, A., Wang, G., Liu, T., Shrestha, S., Xue, B., and Tan, Z. (2019). Vertical variations of soil water and its controlling factors based on the structural equation model in a semi-arid grassland. Sci. Total Environ. 691, 1016–1026. doi: 10.1016/j.scitotenv.2019.07.181
Zeng, Q., Li, X., Dong, Y., An, S., and Darboux, F. (2016). Soil and plant components ecological stoichiometry in four steppe communities in the Loess Plateau of China. Catena 147, 481–488. doi: 10.1016/j.catena.2016.07.047
Zeraatpisheh, M., Ayoubi, S., Mirbagheri, Z., Mosaddeghi, M. R., and Xu, M. (2021). Spatial prediction of soil aggregate stability and soil organic carbon in aggregate fractions using machine learning algorithms and environmental variables. Geoderma Reg. 27, e00440. doi: 10.1016/j.geodrs.2021.e00440
Zhai, C., Han, L., Xiong, C., Ge, A., Yue, X., Li, Y., et al. (2024). Soil microbial diversity and network complexity drive the ecosystem multifunctionality of temperate grasslands under changing precipitation. Sci. Total Environ. 906, 167217. doi: 10.1016/j.scitotenv.2023.167217
Zhang, J., Chi, F., Wei, D., Zhou, B., Cai, S., Li, Y., et al. (2019). Impacts of Long-term fertilization on the molecular structure of humic acid and organic carbon content in soil aggregates in black soil. Sci. Rep. 9, 11908. doi: 10.1038/s41598-019-48406-8
Zhang, Y. and Zhao, W. (2015). Vegetation and soil property response of short-time fencing in temperate desert of the Hexi Corridor, northwestern China. Catena 133, 43–51. doi: 10.1016/j.catena.2015.04.019
Zhang, D., Zhao, Y., Qi, H., Shan, L., Chen, G., and Ning, T. (2024). Effects of Micro-Topography and vegetation on soil moisture on fixed sand dunes in Tengger Desert, China. Plants 13, 1571. doi: 10.3390/plants13111571
Keywords: grassland type, ecosystem multifunctionality, plant community characteristics, plant diversity, soil factors
Citation: Li Y, He G, Liu X and Ji T (2025) Characteristics of ecosystem multifunctionality and influencing factors of different grassland types in temperate desert of Longzhong Loess Plateau. Front. Plant Sci. 16:1619948. doi: 10.3389/fpls.2025.1619948
Received: 29 April 2025; Accepted: 25 June 2025;
Published: 17 July 2025.
Edited by:
Hao Zhang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Lei Xie, Nanjing Forestry University, ChinaAkram Salah, Lasbela University of Agriculture, Water and Marine Sciences, Pakistan
Copyright © 2025 Li, He, Liu and Ji. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiaoni Liu, bGl1eG5AZ3NhdS5lZHUuY24=