 Daicy Galvis-Tarazona
Daicy Galvis-Tarazona María Bohórquez-Quintero
María Bohórquez-Quintero Diana Arias-Moreno
Diana Arias-Moreno Zaida Ojeda-Pérez
Zaida Ojeda-Pérez- 1Research Group BIOPLASMA-UPTC, Faculty of Sciences, Universidad Pedagógica y Tecnológica de Colombia, Tunja, Colombia
- 2Plant Molecular Biotechnology Laboratory, Division of Molecular Biology, Instituto Potosino de Investigación Científica y Tecnológica AC, San Luis Potosí, Mexico
- 3Research Group in Biotechnology and Agricultural Production, Faculty of Agronomy Engineering, Plant Production and Health Department, Universidad del Tolima, Ibagué, Colombia
Introduction: Passiflora maliformis is a wild genotype of passionflower with antimicrobial activity and broad phytochemical properties, highlighting its potential as a genetic resource for crop improvement. Given that commercial passionflowers are severely affected by vascular wilt and stem rot caused by Fusarium oxysporum and F. solani, this study aimed to elucidate the resistance mechanisms of P. maliformis var. pubescens against these pathogens, with emphasis on histopathological aspects.
Methods: Pathogenicity tests were performed on seedlings germinated in vitro and ex vitro (SI and SE). Survival, disease incidence, and severity were evaluated in inoculated and non-inoculated plants. In addition, histopathological analyses of roots and stems were performed to characterize structural changes, defense responses, and pathogen colonization.
Results: The genotype exhibited high survival rates and reduced incidence and severity of Fusarium wilt and stem rot, with minimal damage to internal and external tissues. Comparable shoot and root development was observed between inoculated and control plants, indicating the effectiveness of defense mechanisms that maintained physiological performance. Histopathological examination revealed reinforced cell walls, limited pathogen penetration, and restricted vascular colonization.
Conclusion: P. maliformis var. pubescens demonstrated high responsiveness against F. oxysporum and F. solani, supporting their role as a potential resistant rootstock. These findings underscore the value of this wild genotype as a strategic genetic resource for breeding programs, integrated disease management, and the sustainable improvement of commercial passionflower crops.
1 Introduction
The genus Passiflora, comprising more than 520 recognized species, its a taxonomic group of substantial ethnobotanical, horticultural, medicinal, and economic importance (Moraes et al., 2020; Ożarowski and Karpiński, 2021). Its cultivation is predominantly concentrated in tropical and subtropical regions, with Brazil, Colombia, Peru, and Ecuador standing out as the main producers in South America (Fischer et al., 2021). Notably, Colombia is acknowledged as one of the principal centers of genetic diversity, as its heterogeneous agroecological conditions provide optimal environments for the cultivation and diversification of multiple species (Martínez et al., 2020).
In Colombia, the commercial cultivation of passionflowers encompasses taxa of significant agronomic, nutritional, and economic importance, including yellow passion fruit (Passiflora edulis f. flavicarpa Degener, locally known as maracuyá), purple passion fruit (P. edulis f. edulis Sims, gulupa), sweet granadilla (P. ligularis Juss., granadilla), banana passion fruit (P. tripartita var. mollissima [Kunth], curuba), and stone granadilla (P. maliformis L., cholupa) (Fischer et al., 2021). As of 2024, passion fruit cultivation in Colombia covered approximately 32,430 hectares across 28 departments, yielding an estimated production of 449,775 tons (Ministerio de Agricultura y Desarrollo Rural (MADR), 2025).
Passiflora maliformis is also a species of considerable agronomic interest. Based on morphological and genetic analyses, it is classified within the supersection Stipulata, section Granadillastrum, and series Tiliifolia (Yockteng and Nadot, 2004; Ramaiya et al., 2014). Native to the tropical regions of the Americas, this vigorous vine produces large, pendulous hermaphroditic flowers, which display high self-incompatibility (~95%), and fruits that mature within 50–60 days (Molano-Avellaneda et al., 2020). Its phytochemical profile, characterized by a high content of total phenols and strong antioxidant activity, confers extracts with antioxidant, cytotoxic, and antimicrobial properties (Sabogal-Palma et al., 2016).
Another Passiflora species of major agronomic importance in Colombia is purple passion fruit (P. edulis f. edulis), currently ranked as the third most exported fruit. Its profitability and strong demand in European and Canadian markets have driven a steady expansion of its cultivation (López Plazas, 2020). This product is consumed fresh or incorporated into various culinary preparations due to its characteristic aroma and flavor. Its fruits are rich in carbohydrates, essential minerals (Fe, Cu, P, K, Mg), vitamins (riboflavin, niacin, vitamin C), and bioactive compounds with recognized nutraceutical and industrial applications (Fonseca et al., 2022; Pereira et al., 2023). Altogether, these characteristics consolidate purple passion fruit as a strategic crop for Colombia’s agricultural sector and a benchmark for the development of other Passiflora species.
Despite their growing socioeconomic relevance, cultivated passionflower species are severely affected by soilborne pathogens, notably Fusarium oxysporum and F. solani, which cause vascular wilt and stem rot (Fischer and Rezende, 2008; Kiptui et al., 2020; Mukoye et al., 2022; Bernal-Moreno and Rodríguez, 2023; Wang et al., 2023). Under severe infections, these diseases may result in total crop losses (100%) (Torres et al., 1999; Salazar-Gonzalez et al., 2022). Given their persistence in soil and aggressive colonization strategy, these pathogens represent major constraints for purple passion fruit cultivation worldwide, raising significant concerns due to their economic impact (Fischer and Rezende, 2008; Yadeta and Thomma, 2013; Aiello et al., 2021; Kashyap et al., 2021; Gonçalves Barros Silva, 2023).
The infection cycle of Fusarium spp. comprises root adhesion, cell wall degradation through enzymatic activity, and xylem colonization by microconidia. These events trigger systemic dissemination that culminates in vascular blockage, physiological water stress, and host death (Ekwomadu and Mwanza, 2023). Fusarium wilt and stem rot progress acropetally and manifest through early chlorosis, wilting desiccation of aerial tissues, defoliation, vascular discoloration, and plant death (Ortiz et al., 2014). Stem rot is further characterized by necrosis at the root collar and development of perithecia, although its diagnosis is often complicated by overlapping symptoms with Fusarium wilt (Agrios, 2005; Ploetz, 2005).
Managing vascular wilt pathogens is particularly challenging due to their long persistence in soil and their capacity to infect plants at any developmental stage. Conventional control methods, including chemical, physical, and cultural practices, are often ineffective and economically unsustainable (Aiello et al., 2021; Bahadur, 2022; Ekwomadu and Mwanza, 2023). Consequently, increasing emphasis has been placed on genetic strategies such as breeding and grafting with resistant genotypes, which provide sustainable alternatives for the management of Fusarium infections (Panth et al., 2020; Carvalho et al., 2021). Grafting, in particular, has been extensively applied in agriculture to enhance crop tolerance to both biotic and abiotic stresses (Hurtado-Salazar et al., 2021; Williams et al., 2021).
While both grafting and breeding provide valuable opportunities for disease management, their application has not been fully extended to all commercial crops, partly due to genetic variability in the response of passion fruit species to pathogens such as Fusarium (Ferreira et al., 2023). In this context, studies conducted in producing countries such as Brazil and Colombia, have evaluated the resistance to F. oxysporum and F. solani of some passion fruit species grown in greenhouses or nurseries (Londoño, 2012; Silva Flores et al., 2012; Ortiz et al., 2014; Forero et al., 2015; Papadaki et al., 2017; Rios Ribeiro et al., 2018; Patiño Pacheco, 2020; Miguel-Wruck et al., 2021).
On the other hand, although P. maliformis has been proven to exhibit high compatibility and potential as a rootstock for purple passion fruit (P. edulis f. edulis), systematic evaluations of its resistance to Fusarium spp., particularly under controlled conditions, remain scarce (Betancourt and Muñóz, 2012; Miguel-Wruck et al., 2021; Roncatto et al., 2021). Therefore, the present study aimed to elucidate the resistance mechanisms of Passiflora maliformis var. pubescens against F. oxysporum (Fusarium wilt) and F. solani (stem rot), with emphasis on histopathological traits. The results provide a foundation for enhancing disease resistance and productivity in passionflower crops, supporting the validation of this wild genotype as a resistant rootstock and a valuable candidate for breeding programs.
2 Material and methods
This study was carried out at the BIOPLASMA-UPTC Plant Tissue Culture Laboratory in four main phases: (I) ex vitro and in vitro propagation of Passiflora maliformis var. pubescens seedlings, (II) obtaining pathogenic isolates of F. oxysporum and F. solani complexes, (III) inoculation of plant material and response evaluation, and (IV) histological analysis.
2.1 Plant and fungal material
Healthy seeds of Passiflora maliformis var. pubescens from the department of Boyacá were selected as source material. Such seeds were sown under ex vitro in peat and maintained under greenhouse conditions (SE) or germinated in vitro by zygotic embryo culture (SI) to obtain seedlings. When the SE seedlings were 3–4 cm tall, with development of cotyledonary leaves and a second pair of leaves, they were potted in 0.5 kg bags with a sterile substrate mixture composed of soil:capote:sand in a 3:2:1 ratio.
On the other hand, the SI seedlings were obtained with the methodology reported by Bohórquez et al., (2016). Thus, in vitro cultures were established from sexual embryos, to obtain 3–4 cm tall seedlings with developing cotyledonary leaves and a second pair of leaves that were gradually acclimatized in a greenhouse for about 20 days.
Meanwhile, the Fusarium oxysporum isolate used in this study was obtained from plants of Passiflora edulis Sims f. edulis (Gulupa) crops affected by Fusarium wilt, established in the municipality of Sutamarchán (Pedregal Bajo; altitude 2.200 m), department of Boyacá, Colombia. Isolated colonies were morphologically characterized from cultures grown on PDA (potato dextrose agar). Proliferated colonies were examined under a stereomicroscope. In addition, microscopic structures were observed on fresh samples stained with lactophenol cotton blue with an optical microscope (Nikon eclipse T2000). Qualitative descriptors, as proposed by Leslie and Summerell (2006) and Sañudo (2012) were used.
Molecular identification was performed by Corpogen Corporation by amplification of the ITS region using the universal primers ITS 4 and 5 (White et al., 1990). The sequence (Accession: PRJNA1216561) was analyzed with the NCBI database using BLAST software. The corresponding fungal isolate was assigned to the species name after comparison with representative sequences available in GenBank and UNITE. Molecular procedures allowed us to obtain fragments of 571 bp as a result of the amplification of PCR products of ITS regions. The taxonomic analysis of this sequence in relation to the NCBI and UNITE nr/nt database indicated a high similarity with Fusarium oxysporum (99% identity in 100% of its length).
On the other hand, the isolate of F. solani used here (MV184), was previously characterized and identified by Patiño Pacheco (2020) as a causal agent of Fusarium wilt in sweet granadilla and also recognized as highly pathogenic for 18 accessions of Passiflora.
2.2 Pathogenicity test
Two Fusarium isolates pathogenic to passion fruits were selected for this study. In vivo pathogenicity was tested using two inoculation methods: 1) root immersion and 2) soil infestation with rice grains. The conidial suspension was prepared following the methodology by Namiki et al. (1994) with a concentration of 1x106 con/ml. Seedlings were removed from the substrate, and their roots were washed with sterile distilled water. For inoculation, the root system of uniformly appearing plants was immersed in the conidial suspension for 5 minutes; the solution was agitated before and during the procedure. Then, seedlings were transplanted into a sterile substrate composed of soil, rice husk, and peat in a 3:1:1 ratio.
For soil infestation with the rice grain method, Chaff-Grain medium was prepared following the methodology described by Leslie and Summerell (2006). This suspension was homogenized in 1 kg of substrate as above and the seedlings were sown in the inoculated substrate.
In the pathogenicity assays, several SE and SI seedlings equivalent to those used in each inoculation treatment were included as controls. The roots of these control seedlings were initially immersed in sterile distilled water and then planted in the sterile substrate, following the same procedure described for the inoculated plants.
A completely randomized block design was used with three replications, and an equal number of seedlings was assigned to 10 treatments (Table 1). Each experimental unit consisted of six seedlings per replicate. Disease progression was assessed by randomly sampling plants at 3-day intervals, starting from the date of transplanting to pots and continuing for 27 days.
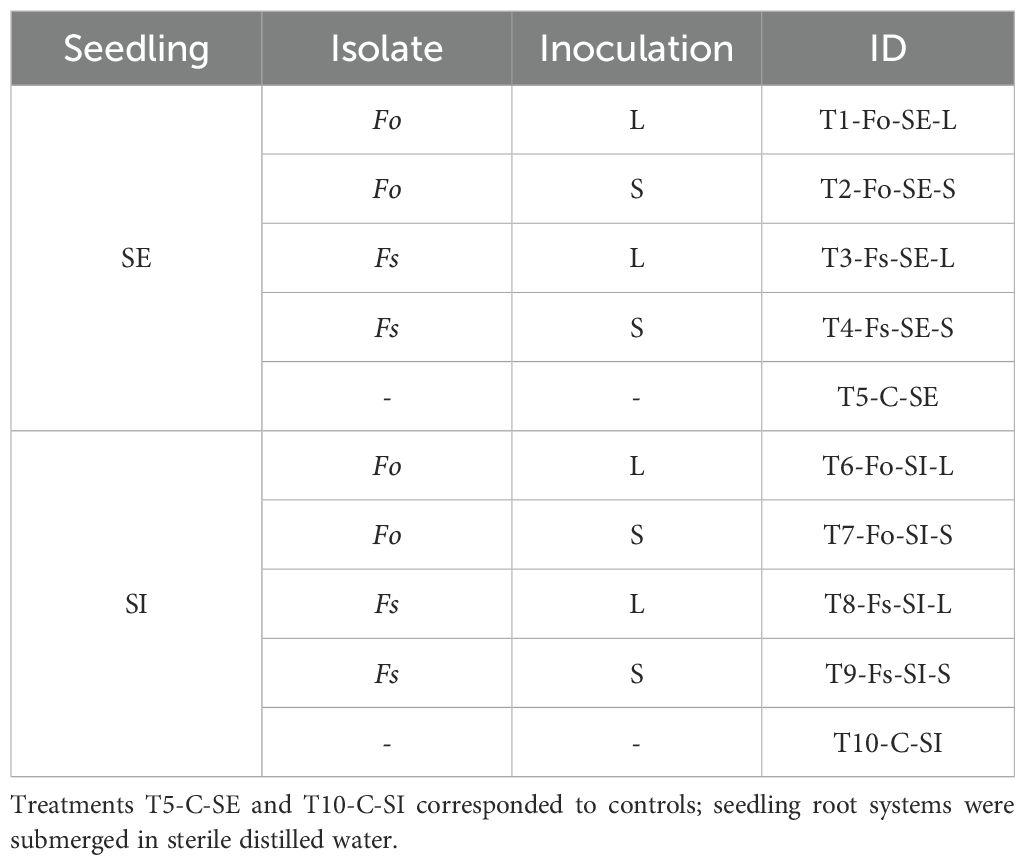
Table 1. Inoculation treatments on Passiflora maliformis var. pubescens germinated ex vitro (SE) and in vitro (SI), by liquid suspension (L) and infestation on solid substrate (S), with Fusarium oxysporum (Fo) and Fusarium solani (Fs).
2.3 Disease assessment.
The disease progression was assessed through qualitative and quantitative estimators, such as incidence, severity and mortality. Incubation period refers to time from inoculation to the expression of disease symptoms in the plants. Incidence of the disease was determined by formula: Incidence= (number of diseased individuals/total number of individuals) *100. Regarding disease severity was assessed using 0–9 scale based on the presence of discoloration, chlorosis, leaf defoliation, the appearance of the collar and root system, and the percentage of plant area affected (Figure 1), as follows: Grade 0 = no infection (0-10%), Grade 3 = 11-20%, Grade 5 = 21-50%, Grade 7 = 51-75%, Grade 9 = 76%- 100%, according to Rodriguez et al. (1999). Seedlings exhibiting severity grades higher than 7 were removed from the pots to further examine the collar and root system.
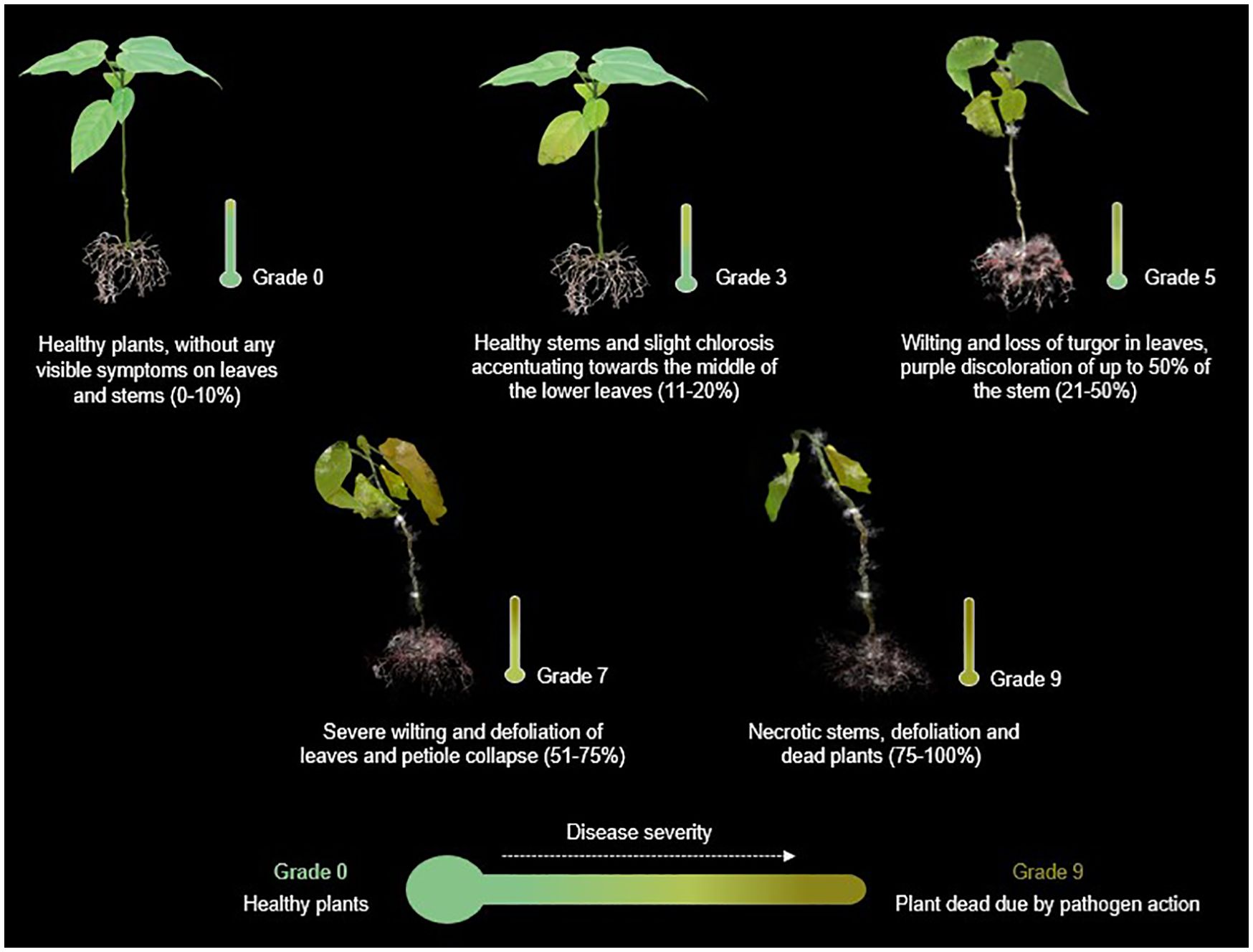
Figure 1. Representative illustration of the progress of Fusarium wilt based on symptomatic progression and percentage of the plant affected. Scheme devised and recreated by the authors with images illustrated by Camila Reyes and Angie Carvajal.
Moreover, disease intensity was quantified using the severity index, calculated according to the method proposed by Townsend and Heuberger (1943). These values were then used to compute the area under the disease progression curve (AUDPC), which integrates disease development over time for comparison among treatments (López-Santiago et al., 2008). Additionally, plant mortality (expressed as a percentage of dead plants), was recorded from each treatment 27 days after inoculation.
2.4 Histological analysis
A comparative study of the infection process by both pathogens in the plant tissues was carried out using light microscopy to understand the histological changes/mechanisms associated with the response of P. maliformis var. pubescens to inoculation with F. oxysporum and F. solani species complexes. After monitoring, stems and roots were isolated from inoculated plants and SE and SI controls. Tissue samples were sectioned and processed according to the protocol described by Ortiz et al., (2014) with some modifications. The sections were isolated and fixed for 24 h in FAA. Subsequently, these were dehydrated in a series of ethanol isopropanol and acetone. The samples were embedded in paraffin, and histological sections (8-10 µm) were obtained with a rotary microtome. Cross-sections were stained with Safranin-FastGreen and observed under a microscope LEICA DM750 binocular microscope.
2.5 Data analysis
Logistic regression analysis was selected as the primary method for data analysis, given the inclusion of multiple independent variables, which justified the use of a multivariate approach. Using a maximum likelihood estimator, the model identified the set of coefficients that maximized the probability of reproducing the observed data. Incidence and severity were coded as binary (“no” = 0, “yes” = 1) and ordinal (scale 0 to 9) variables, respectively, making probabilistic modeling appropriate. A binomial logistic regression model was applied for incidence, while a multinomial logistic regression model was used for severity.
Backward stepwise selection was employed to assess the significance of the factors: day, plant material, fungal species, inoculation technique, and their interactions. Model significance, main effects, and interactions were evaluated using the Chi-square test. Factor levels were compared through odds ratios and predicted probabilities. The AUDPC was analyzed using analysis of variance (ANOVA) to detect significant treatment effects. All statistical analyses were performed using SAS software (SAS Institute Inc, 2019).
3 Results
3.1 Disease, incidence and severity
Symptoms of Fusarium wilt and stem rot after inoculation included yellowing of lower leaves. Then, chlorosis spread through the plant, and the tissues progressively became necrotic and collapsed, ultimately dying. In other cases, symptoms were observable until plants collapsed without prior occurrence of discoloration and disease severity varied among the seedlings evaluated. The incubation period of F. solani and F. oxysporum was nine days in SI, and 15 days in SE, with both types of inoculation (Figures 2A, B).
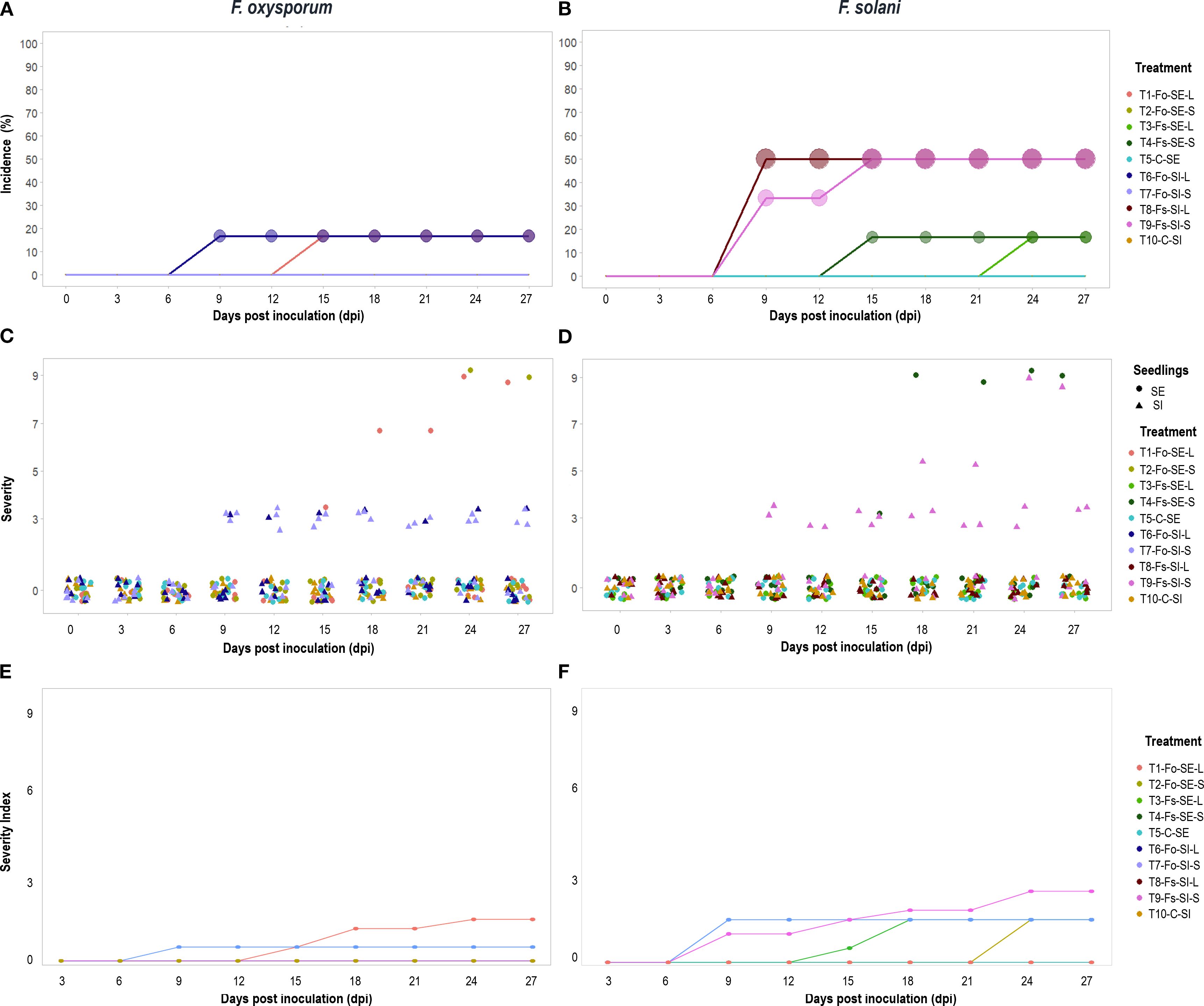
Figure 2. Response of Passiflora maliformis var. pubescens to F. oxysporum (right plots) and F. solani (left plots) inoculation during 27 dpi (days post inoculation). In terms of incidence (A, B), the percentage of affected plants coincides with the size of the circles, the higher the value, the larger the size of the figure. Severity (C, D), data set and follow-up to seedlings that integrated the sample and severity index (E, F).
The Fusarium species tested provoked visible symptoms on both SE and SI plants with both inoculation methods. The incidence of disease due to F. oxysporum after 27 dpi ranged from 0 to 16.67%, and the highest value in both SE and SI plants was recorded with liquid inoculum (Figure 2A). With F. solani, the percentage rates of infection at 27 dpi ranged between 16.67 and 50%, with the highest data for SI for both types of inoculation (Figure 2B).
In terms of severity, symptomatic and asymptomatic plants were identified at 27 dpi. Of the diseased plants 60% were slightly affected (grade 3) while the remaining 40% developed drastic symptoms (grade 9) (Figures 2C, D), with an F. solani: F. oxysporum ratio of 5:1 and 3:1 for each grade, respectively. Thus, in SI plants, there was a higher risk of developing the disease and with more severe symptoms under the action of F. solani (Figure 2D).
The analysis identified statistically significant differences in incidence and severity between fungal species and plant material (SE, SI) (p<0.0001; Supplementary Tables S1, S2). In addition, the probability of finding healthy plants of SE and SI was higher than 50%. Likewise, the occurrence of Fusarium wilt with medium-high severity (5-9) was less than 10% for both cases (Supplementary Figure S1).
The severity index obtained was 2.5 on the scale with a maximum value of 9. In plants inoculated with F. oxysporum the index was less than or equal to 1.5, while with F. solani it was 2.5 (Figure 2E, F). In addition, the analysis identified statistically significant differences in severity index between fungal species and plant material (SE, SI) (p<0.001; Supplementary Tables S3, S4).
The AUDPC obtained ranged from 0 for plants without symptoms to 32.75 for the most affected plants (Table 2). The results were not statistically different between treatments (p=0,3263); Supplementary Table S5), this can be associated, with low fungal affectation at SE and SI. With respect to mortality it was determined that Fusarium wilt is not a limiting factor for the viability of P. maliformis var. pubescens. Only 6.67% of the inoculated plants died by 27 dpi, of which 5.25% and 1.75% corresponded to SE and SI, respectively. In general, the inoculated plants remained viable (93.33%).
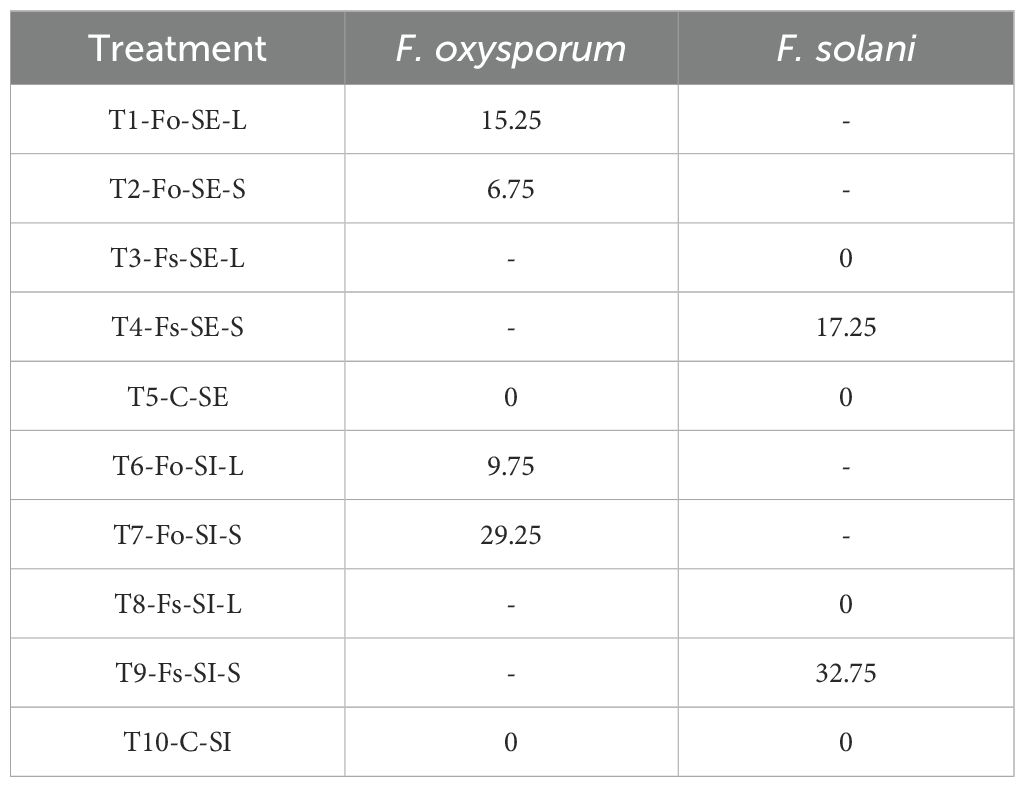
Table 2. Fusarium wilt AUDPC, 27dpi of Passiflora maliformis var. pubescens with Fusarium oxysporum (Fo) and Fusarium solani (Fs).
3.2 Histological analysis of Fusarium infection in P. maliformis var pubescens
In transverse microsections of stem and root of non-inoculated plants, monostratified epidermis, collenchyma, parenchyma, vascular cells, pith and sporadically druses were observed. Vascular cells were arranged in eustella, that is, distributed in a circle and separated by the interphasicular cambium. These tissues and characteristics were also apparent in plants inoculated with F. oxysporum and F. solani, with some changes (Figure 3).
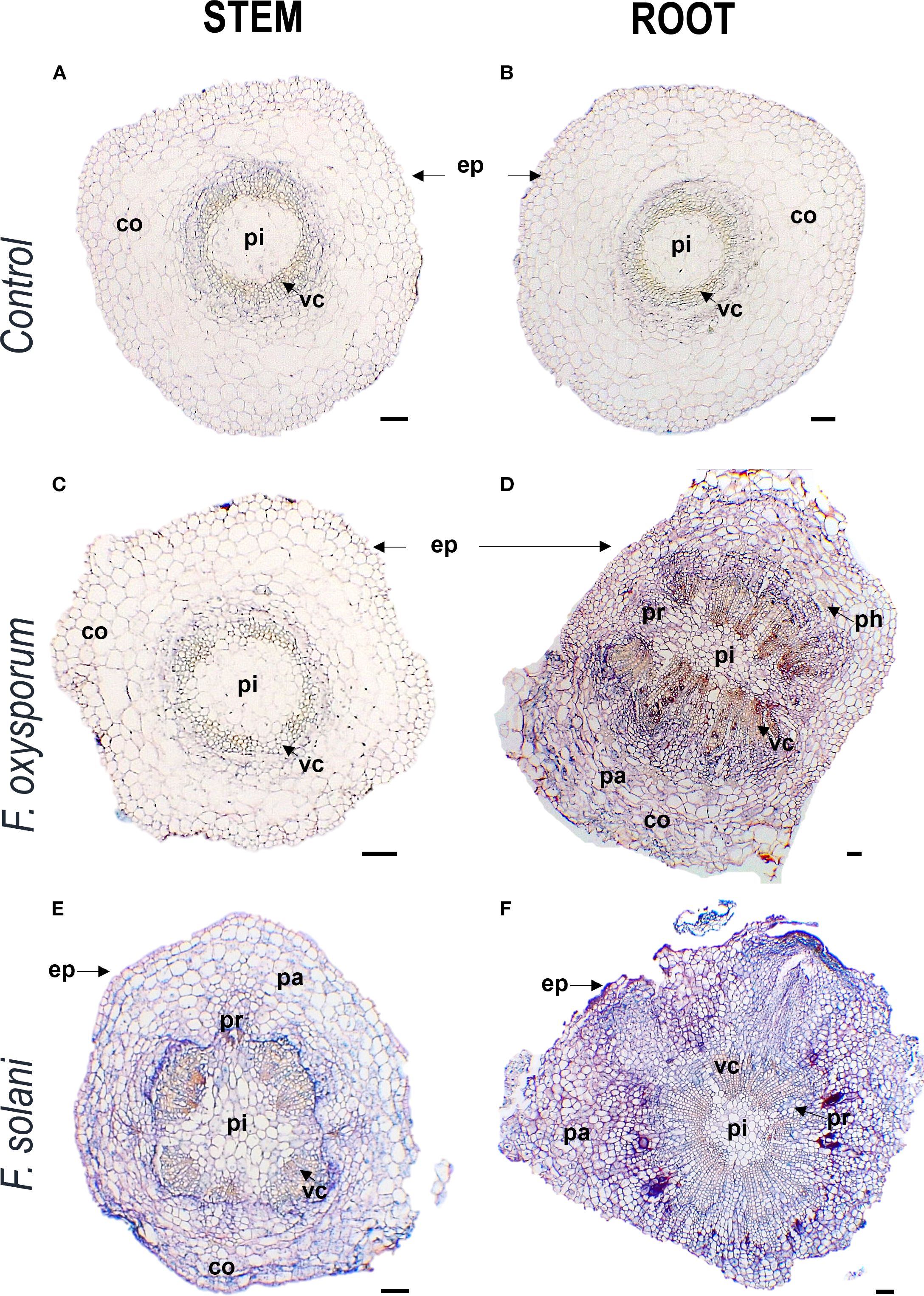
Figure 3. Photomicrographs of cross-cutting sections of stems and roots of Passiflora maliformis var. pubescens control (A, B) and under inoculation with Fusarium oxysporum (C, D) and Fusarium solani (E, F). co, cortex; ep, epidermis; pa, parenchyma; ph, phloem; pi, pith; pr, pith radial and vc, vascular cells. Scale bars, 100 µm. Image created by the authors. The coloration in each image reflects the affinity of the tissues to the Safranin-FastGreen staining method.
The anatomical response of the plant to contact with the pathogen was consistent with the degree of severity of the disease. In inoculated plants with mild to medium symptomatology (grades 0 - 5), xylem cells with dense secretions inside and with thickened and lignified cell walls were observed in the stem, showing histological changes compared to control individuals (Figure 4). In the root, these mechanisms were more frequent and were complemented by secretions in the intercellular spaces and and what appeared to be tyloses (Figure 5). On the other hand, in severely affected plants (grades 7 and 9), tissues gradually collapsed until they suffered complete degradation, while the abundance of fungal spores increased (Supplementary Figure S2). These cellular changes coincided with the appearance of noticeable symptoms of infection leading to plant death.
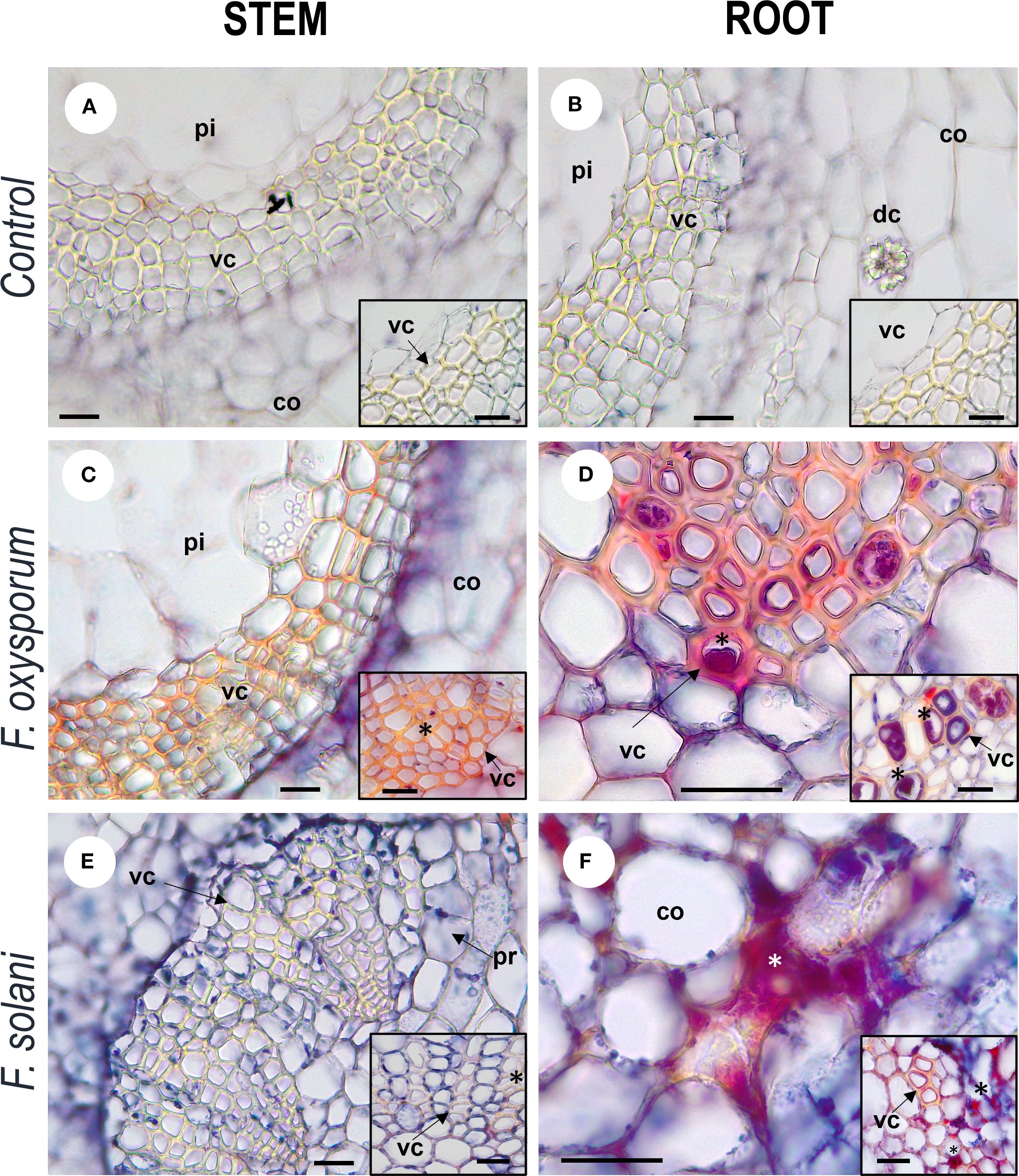
Figure 4. Photomicrographs of cross-cutting sections of stems and roots of Passiflora maliformis var. pubescens control (A, B) and under inoculation with Fusarium oxysporum (C, D) and Fusarium solani (E, F). co, cortex; dc, druse crystal; pi, pith; and vc, vascular cells. Scale bars, 25 µm. Image created by the authors. In inoculated tissues, inside and between the vascular cells there are red-violet dense substances (undetermined), marked with asterisks; absent in the micro-preparations of control plants.
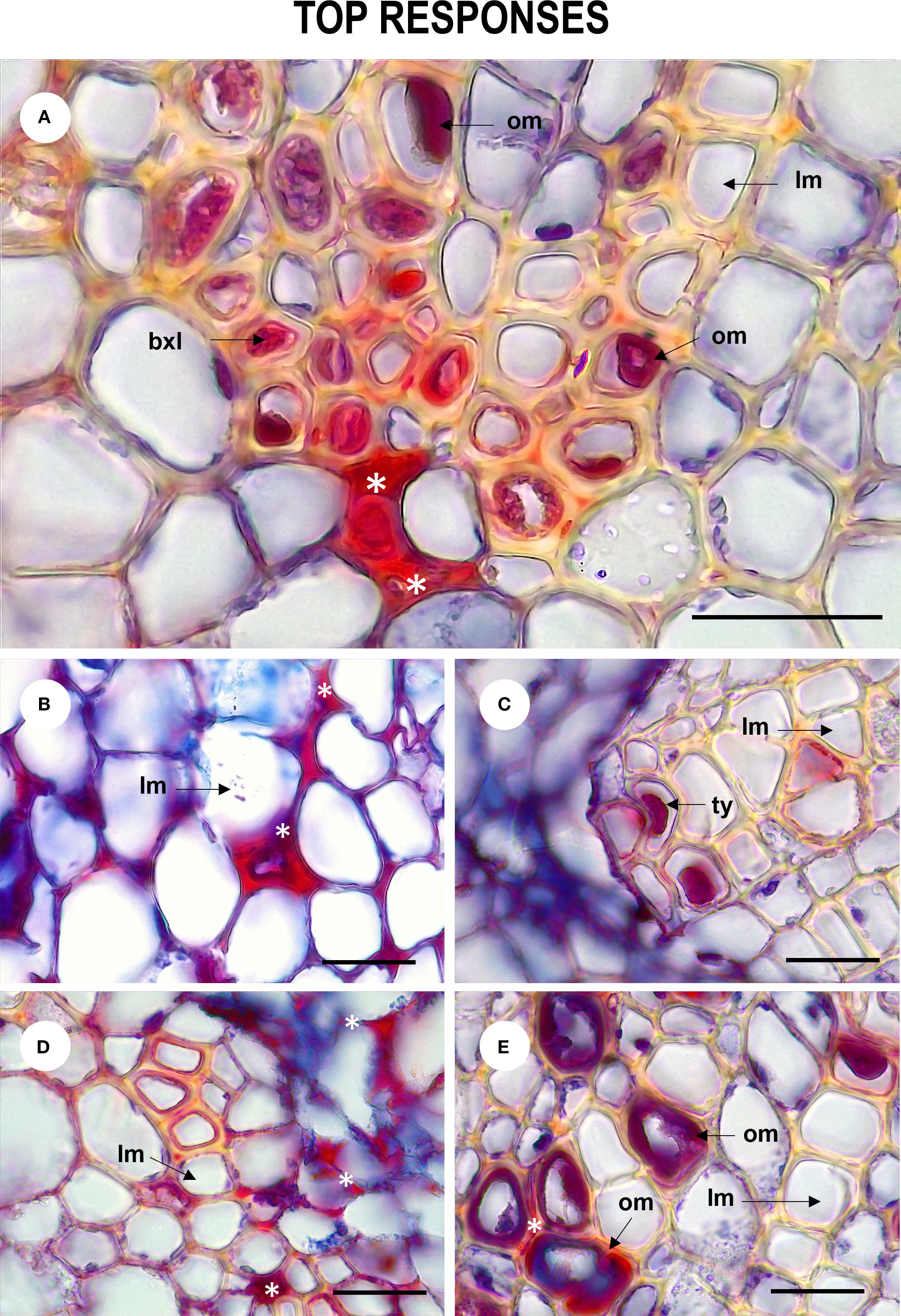
Figure 5. Tissue defense responses of Passiflora maliformis var. pubescens inoculated with Fusarium oxysporum and Fusarium solani denoted by the occurrence of dense substances (undetermined) (A, B, D), occluding material (A, E) and tyloses (C). om, occluding material; lm, lumen; bxl, blocked xylem lumen and ty, tyloses. Scale bars, 25 µm. Image created by the authors. 27 dpi showing accumulation of substances in the intercellular space (white asterisk).
Microscopy allowed to identify and illustrate the main stages of the Fusarium wilt disease cycle (Figure 6). Thus, it was possible to confirm that the germination of chlamydospores, the development of hyphae that entered the root epidermis and subsequently colonized the cortex constitute the first stages of the disease. Upon colonization, the plant develops response mechanisms to prevent the spread of the fungus in the tissues. However, when fungal infection overcomes these barriers, internal and external tissue collapse occurs in the host, followed by the formation of pathogen reproductive spores (Figure 7). Therefore, the results of this study allowed us to relate the activation of defense mechanisms with a lower degree of disease severity.
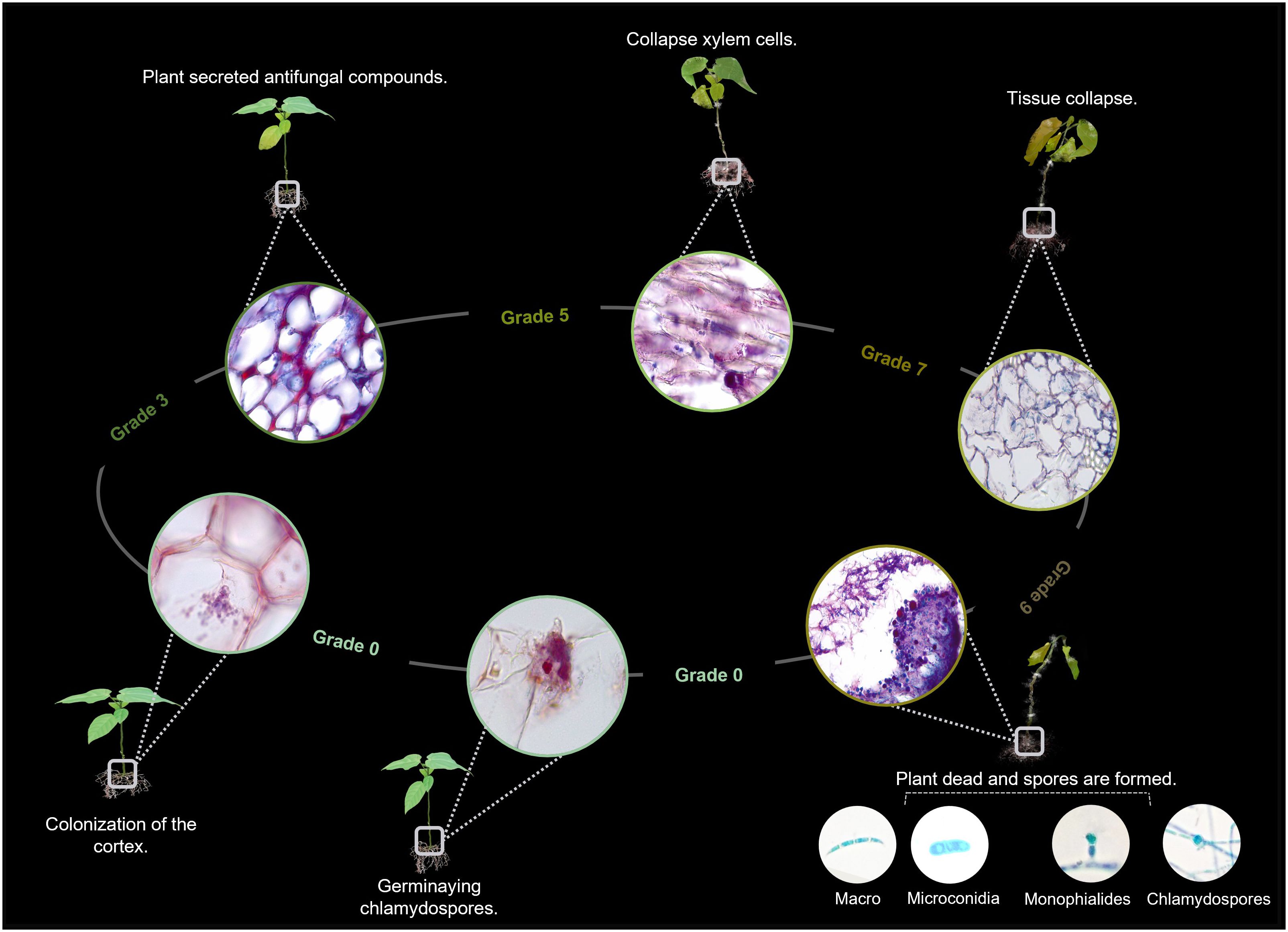
Figure 6. Disease cycle of Fusarium wilt in Passiflora maliformis var. pubescens based on the study conducted; idea adapted from Jangir et al. (2021). The illustration represents the main stages of interaction between the pathogen and the host (plant).
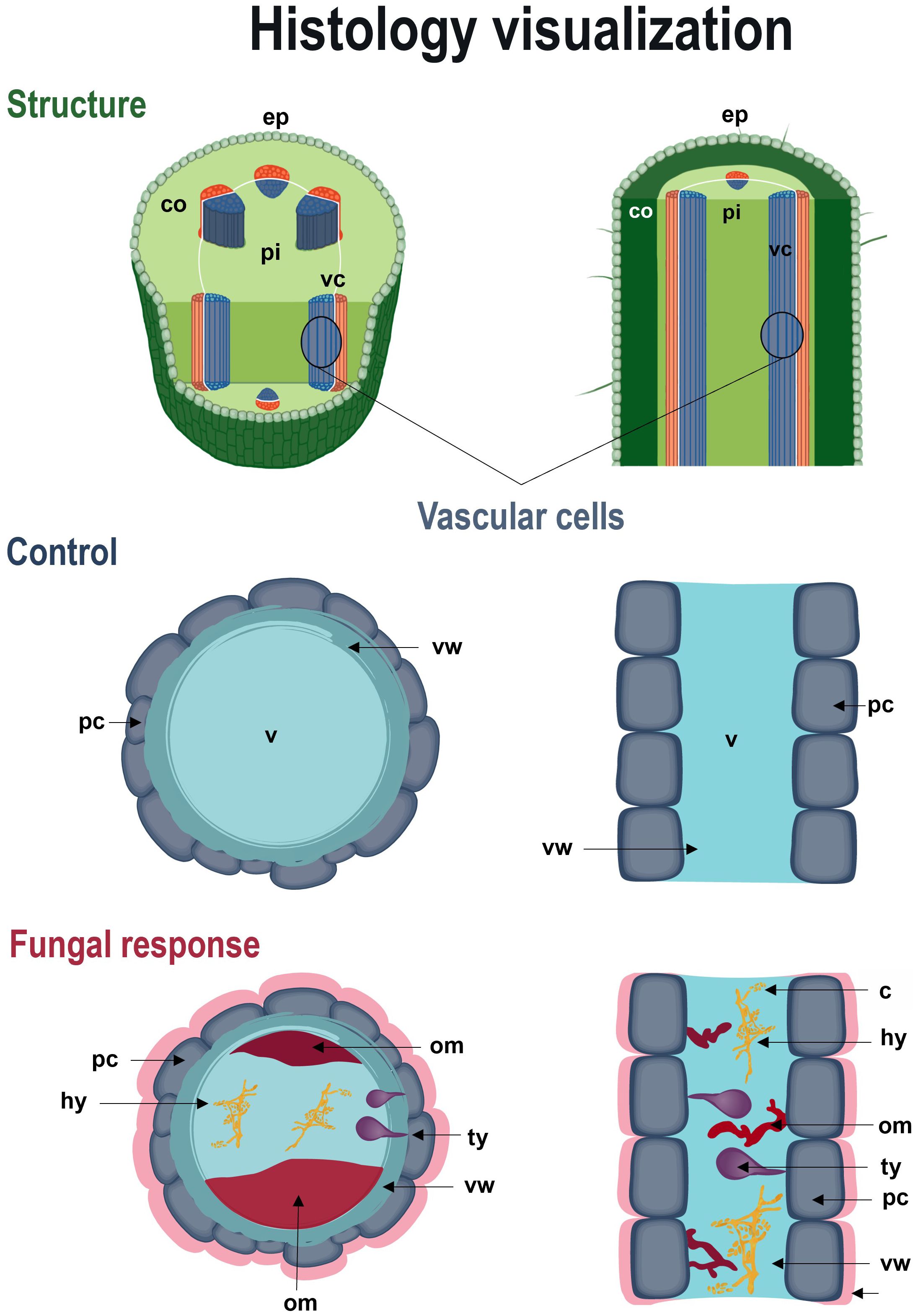
Figure 7. Schematic recreation of defense responses of Passiflora maliformis var. pubescens to Fusarium wilt. Longitudinal and cross-sectional view of the histological response of the host to pathogenic action. co, cortex; ep, epidermis; pi, pith; om, occluding material; sic, substances in the intercellular space; ty, tyloses and vc, vascular cells; v, vessel.
4 Discussion
Research related to diseases affecting crops is essential to guide comprehensive prevention, management and control strategies (Rahut et al., 2022; Zhang et al., 2023). Also, being relevant for the production of safe and healthy food, it can contribute to food security (Li et al., 2023). In the Passiflora production sector, it is a priority to have cultivars that are resistant to Fusarium wilt and meet nutritional, productive and industrial demands (Amorim-Pereira et al., 2019). However, our current knowledge of the relationship between F. oxysporum and F. solani and wild genotypes of Passiflora is still incomplete. This study contributes to filling that gap, through the evaluation of the response of a wild genotype to the action of Fusarium wilt and stem rot causal agents (Mukoye et al., 2022; Nassimbwa et al., 2022). This analysis included aspects frequently reported to determine the response of Passiflora species/accessions to the action of Fusarium spp., in addition to a histopathological description.
F. oxysporum affected less than 17% of the seedlings, with a low symptomatic progression, a severity index of up to 0.5, and an average AUDPC of 6.25. Although inoculation with F. solani led to an incidence ≤50%, symptomatology was mild, the severity index was less than 1.75 and disease progress (AUDPC) had a mean value of 21.5. In addition, mortality after 27 dpi in inoculated plants was less than 7%, with similar records for F. oxysporum and F. solani. These results contrast with those reported for species susceptible to Fusarium wilt and stem rot such as Passiflora edulis f. flavicarpa Degener, P. edulis f. edulis Sims and P. ligularis Juss, where incidence generally ranges between 40-100% (Forero et al., 2015; Melo et al., 2020; Rocha et al., 2021) with medium-high affectation (Ortiz and Hoyos-Carvajal, 2016) and a severity index ranging from 5-7.5 (on a scale of 1-10) (Lima et al., 2019). Likewise, mortality of up to 70% and survival of less than 30% have been reported (Pires et al., 2022).
On the other hand, in the case of resistant accessions, previous studies have reported incidences between 0 and 50% (Londoño, 2012; Patiño-Pacheco and Pérez-Cardona, 2021), mortality rates below 50% (Amorim-Pereira et al., 2019; Melo et al., 2020) survival rates of 67-100% (Carvalho et al., 2021; Oliveira et al., 2022), and progression of severity (AUDPC) close or equal to zero (Pérez and Forbes, 2008); which would indicate a low level of infection in the genotype evaluated in this study.
The search for new sources of resistance against Fusarium wilt and stem rot requires efficient detection methods that provide information on physical and chemical response mechanisms (Sampaio et al., 2020). Upon an attack of pathogenic microorganisms plants activate complex immune networks to prevent or minimize colonization of their internal structures and deprivation of nutrients (Margaritopoulou et al., 2020; Duarte-Alvarado et al., 2021). In vascular wilt, the formation of structural barriers in and around the vascular bundles is one of the most important components of defense against the disease according to Kashyap et al. (2021).
Internal anatomical analysis revealed intercellular and intracellular secretion of dense substances, as well as possible lignification of cell walls and tyloses formation, as potential response mechanisms of P. maliformis var. pubescens to infection by isolates of the F. oxysporum and F. solani complexes. These reactions play a crucial role in limiting the spread of the pathogen, since physical structures neutralize or reduce further spread and chemicals can lead to the death or inhibition of pathogen growth (De Micco et al., 2016; Chakraborty et al., 2022).
In the initial steps of the response to Fusarium wilt, gels are formed within the xylem that are mainly composed of polysaccharides and phenolic compounds exuded from the parenchymal cells adjacent to the infection point. Once these compounds are fused to the cell wall, oxidation and polymerization reactions occur to form a durable barrier at the interface of the infected and healthy tissues (Ardila et al., 2014). Gels and dense substances fix conidia on vascular elements and localize the pathogen when F. oxysporum infection occurs (Muche and Yemata, 2022).
Other common barrier mechanisms against the disease are cell wall strengthening due to lignin deposition and the synthesis and accumulation of callose and phenolic compounds in inter- or intracellular spaces that block Fusarium progression in the host (Sampaio et al., 2020). Phenols have antifungal capacity by affecting the cell permeability of microorganisms, causing structural and functional changes in enzymes and membrane proteins, leading to alterations in the pH gradient and in the ATP production and conservation system (Carmona et al., 2020).
The results of this study suggested a high response capacity of the genotype since the defense mechanisms limited the pathogenic action of Fusarium spp. and low internal and external tissue damage was observed. Overall, both inoculated and non-inoculated SE and SI seedlings exhibited comparable shoot and root growth and development, which may indicate that the defense mechanisms of the genotype were effective in maintaining normal physiological performance. In this sense, the use of in vitro culture allowed the rapid and large-scale production of plant material, while providing a platform to evaluate resistance traits under controlled conditions. This dual functionality reinforces its potential application in breeding programs and integrated disease management strategies (Bernal- Moreno et al., 2018; Bernal-Moreno and Rodríguez, 2023). Therefore, these results are especially relevant, as this wild genotype and its genetic background may serve as a strategic resource to address key biotechnological and agronomic challenges currently faced by promising Passiflora species.
Given the severe limitations caused by Fusarium wilt and stem rot in passion fruits agricultural systems, management of these diseases often relies on synthetic fungicides (Liu et al., 2025). Such as benzimidazoles (e.g., carbendazim), triazoles (e.g., tebuconazole) and strobilurins (e.g., azoxystrobin), which alter fungal membrane integrity or interfere with mitochondrial respiration. However, their continued use has raised increasing concerns about high economic costs, limited systemic mobility, persistence in the environment, risks to human health, and the emergence of resistant pathogen populations (Mikaberidze et al., 2014).
Ecological alternatives for the biorational control of F. oxysporum and F. solani include fungal and bacterial antagonists such as Trichoderma harzianum, T. asperellum, T. virens, Bacillus subtilis, B. amyloliquefaciens, Pseudomonas fluorescens, P. putida, and P. protegens. These microorganisms have demonstrated inhibition rates of up to 94% in vitro and, in some cases, in the field, through mechanisms that include mycoparasitism, the production of hydrolytic enzymes (e.g., chitinase and β-1,3-glucanase), the emission of volatile organic compounds, and the induction of systemic resistance in host plants (Verma et al., 2007; Ongena and Jacques, 2008; Ben Khedher et al., 2015; Mehmood et al., 2023; Yao et al., 2023). However, the effectiveness of these strategies in the field remains variable, largely influenced by factors such as interactions with native soil microbiota, climate variability, and the limited persistence of inoculants in the rhizosphere (Awu et al., 2023; Wang et al., 2024).
In this context, the use of resistant rootstocks emerges as a complementary and sustainable solution. Resistant genotypes do not require repeated applications, do not generate toxic residues, and are less susceptible to environmental fluctuations (Ayala-Doñas et al., 2020; Suansia and Chandra Samal, 2021). Therefore, the identification of Passiflora rootstocks resistant to Fusarium wilt and stem rot represents a significant contribution to Colombia, one of the world’s leading producers and exporters. This strategy offers a low environmental impact alternative that can improve the productivity of commercial species, reduce the incidence and severity of Fusarium-related diseases, and promote adaptability in various growing conditions.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
DG-T: Investigation, Formal analysis, Resources, Writing – original draft, Writing – review & editing, Funding acquisition, Validation, Data curation, Visualization, Conceptualization, Methodology. MB-Q: Writing – original draft, Investigation, Formal analysis, Funding acquisition, Validation, Writing – review & editing, Conceptualization, Data curation, Methodology. DA-M: Writing – original draft, Funding acquisition, Validation, Writing – review & editing. ZO-P: Funding acquisition, Writing – review & editing, Writing – original draft, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by the Ministry of Science Technology and Innovation of Colombia and the Boyacá Government, through the “Convocation 865 of 2019 for the closure of gaps in the agricultural sector Boyacá”.
Acknowledgments
The authors thank the BIOPLASMA-UPTC research group and exalt the contribution of Dr. José Constantino Pacheco Maldonado who also conceptualized and designed the research. Additionally, the authors would like to thank Johan Sebastian Urquijo Ruiz, Lorena Reyes and Miguel Ángel Cáceres Hernández for the data analysis, and to the researcher Sergio Ochatt for the grammatical adjustment of the English and his scientific review at article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1635702/full#supplementary-material
References
Aiello, D., Fiorenza, A., Leonardi, G. R., and Vitale, A. (2021). Fusarium nirenbergiae (Fusarium oxysporum species complex) causing the wilting of passion fruit in Italy. Plants 10. doi: 10.3390/plants10102011
Amorim-Pereira, P. P., Silva-Lima, L. K., Soares, T. L., Ferraz-Laranjeira, F., Nunes de Jesus, O., and Girardi, E. A. (2019). Initial vegetative growth and survival analysis for the assessment of Fusarium wilt resistance in Passiflora spp. Crop Prot. 121, 107–131. doi: 10.1016/C2020-0-02752-0
Ardila, H. D., Torres, A. M., Martínez, S. T., and Higuera, B. L. (2014). Biochemical and molecular evidence for the role of class III peroxidases in the resistance of carnation (Dianthus caryophyllus L) to Fusarium oxysporum f. sp. dianthi. Physiol. Mol. Plant Pathol. 85, 42–52. doi: 10.1016/j.pmpp.2014.01.003
Awu, J. E., Nyaku, S. T., Amissah, J. N., Okorley, B. A., Agyapong, P. J. A., Doku, F. E., et al. (2023). Grafting for sustainable management of Fusarium wilt disease in tomato production in Ghana. J. Agric. Food Res. 14, 100710. doi: 10.1016/j.jafr.2023.100710
Ayala-Doñas, A., de Cara-García, M., Talavera-Rubia, M., and Verdejo-Lucas, S. (2020). Management of soil-borne fungi and root-knot nematodes in cucurbits through breeding for resistance and grafting. Agronomy 10. doi: 10.3390/agronomy10111641
Bahadur, A. (2022). “Current status of Fusarium and their management strategies,” in Fusarium - An Overview of the Genus. Ed. Mirmajlessi, S. M. (IntechOpen, London, United Kingdom), 41–60. doi: 10.5772/intechopen.95213
Ben Khedher, S., Kilani-Feki, O., Dammak, M., Jabnoun-Khiareddine, H., Daami-Remadi, M., and Tounsi, S. (2015). Efficacy of Bacillus subtilis V26 as a biological control agent against Rhizoctonia solani on potato. Comptes Rendus - Biol. 338, 784–792. doi: 10.1016/j.crvi.2015.09.005
Bernal- Moreno, S. C., Duarte- Rodríguez, L. C., Bohórquez- Quintero, M.D.l.Á., Araque, E. J., and Pacheco, J. C. (2018). Nonzygotic embryogenesis in Passiflora Maliformis. Rev. Peru. Biol. 25, 281–290. doi: 10.15381/rpb.v25i3.15211
Bernal-Moreno, J. P. and Rodríguez, N. (2023). Responses of landraces and commercial cultivars of yellow passion fruit to the prevalence of. Fusarium oxysporum. Agron. Colomb. 41, 1–14. doi: 10.15446/agron.colomb.v41n1.104450
Betancourt, Y. and Muñóz, C. (2012). Estudio de la compatibilidad histológica en microinjertos de Passiflora edulis Sims. con Passiflora Maliformis L. y Passiflora Maliformis var. pubescens (Tunja, Boyacá, Colombia: Universidad Pedagógica Y Tecnológica De Colombia).
Bohórquez, M., Araque, E. J., and Pacheco, J. C. (2016). Germinative response and micropropagation of cóngolo (Tunja, Boyacá, Colombia: Passiflora Maliformis var. pubescens) (Tunja).
Carmona, S. L., Villarreal-Navarrete, A., Burbano-David, D., and Soto-Suárez, M. (2020). Cambios fisiológicos y mecanismos genéticos asociados a la marchitez vascular causada por Fusarium en tomate: una revisión actualizada. Temas Agrar. 25, 166–189. doi: 10.21897/rta.v25i2.2457
Carvalho, J. A., de Jesus, J. G., Araujo, K. L., Serafim, M. E., Gilio, T. A. S., and Neves, L. G. (2021). Passion fruit (Passiflora spp.) species as sources of resistance to soil phytopathogens Fusarium solani and Fusarium oxysporum f. sp. passiflorae complex. Rev. Bras. Frutic. 43, 1–9. doi: 10.1590/0100-29452021427
Chakraborty, N., Chandra, S., Sarkar, A., Ghosh, S., Dasgupta, A., and Acharya, K. (2022). An in planta approach for understanding defense responses in tomato plants against Fusarium oxysporum Schltdl. J. Plant Pathol., 129–136. doi: 10.1007/s42161-022-01224-w
De Micco, V., Balzano, A., Wheeler, E. A., and Baas, P. (2016). Tyloses and gums: a review of structure, function and occurrence of vessel occlusions. IAWA J. 37, 186–205. doi: 10.1163/22941932-20160130
Duarte-Alvarado, D., Lagos-Burbano, T. C., Lagos-Santander, L. K., and Benavides-Cardona, C. A. (2021). Evaluation of resistance to Fusarium oxysporum in genotypes of lulo (Solanum quitoense lam.). Rev. Fac. Nac. Agron. Medellin 74, 9603–9614. doi: 10.15446/rfnam.v74n2.90447
Ekwomadu, T. I. and Mwanza, M. (2023). Fusarium fungi pathogens, identification, adverse effects, disease management, and global food security: a review of the latest research. Agriculture 13. doi: 10.3390/agriculture13091810
Ferreira, L. G., Juliatti, F. C., De Moraes, M. F., Kobayasti, L., Roncatto, G., Juliatti, B. C. M., et al. (2023). Productivity and mortality, of Fusarium oxysporum f. sp. passiflorae in passionfruit vines brs rubi do cerrado, grafetd onto different rootstocks. Biosci. J. 39, 1–13. doi: 10.14393/BJ-v39n0a2023-61288
Fischer, G., Balaguera-López, H. E., and Magnitskiy, S. (2021). Review on the ecophysiology of important Andean fruits : Solanaceae. Rev. U.D.C.A Actual. Divulg. Científica 24, 1–12. doi: 10.31910/rudca.v24.n1.2021.1701
Fischer, I. H. and Rezende, J. A. M. (2008). Diseases of passion flower (Passiflora spp.). Pest Technol. 2, 1–19.
Fonseca, A. M. A., Geraldi, M. V., Junior, M. R. M., Silvestre, A. J. D., and Rocha, S. M. (2022). Purple passion fruit (Passiflora edulis f. edulis): A comprehensive review on the nutritional value, phytochemical profile and associated health effects. Food Res. Int. 160. doi: 10.1016/j.foodres.2022.111665
Forero, R., Ortiz, E., De León, W., Gómez, J. C., and Hoyos-Carvajal, L. (2015). Análisis de la resistencia a Fusarium oxysporum en plantas de Passiflora Maliformis L. Rev. Colomb. Cienc. Hortícolas 92, 197–208. doi: 10.17584/rcch.2015v9i2.4174
Gonçalves Barros Silva, A. M. (2023). Understanding the interaction between Meloidogyne incognita and Fusarium oxysporum f. sp. passiflorae in grafted passion fruit seedlings (Piracicaba, São Paulo, Brasil: University of São Paulo “Luiz de Queiroz).
Hurtado-Salazar, A., Ocampo-Pérez, J., Ceballos-Aguirre, N., García Jaramillo, D., and López, W. (2021). Passiflora: Genetic, Grafting and Biotechnology Approaches (Manizales, Caldas, Colombia: Investigac Nova Science Publishers).
Kashyap, A., Planas-Marquès, M., Capellades, M., Valls, M., and Coll, N. S. (2021). Blocking intruders: inducible physico-chemical barriers against plant vascular wilt pathogens. J. Exp. Bot. 72, 184–198. doi: 10.1093/jxb/eraa444
Kiptui, L. J., Toroitich, F. J., Kilalo, D. C., and Obonyo, M. (2020). Interaction between cowpea aphid-borne mosaic virus isolates and its effect on passion fruit woodiness disease on Passiflora edulis Sims and Passiflora ligularis Juss. Adv. Agric. 2020. doi: 10.1155/2020/8876498
Leslie, J. F. and Summerell, B. A. (2006). The Fusarium Laboratory Manual (USA: Wiley-Blackwell). First.
Li, J., He, K., Zhang, Q., Wu, X., Li, Z., Pan, X., et al. (2023). Draft genome and biological characteristics of Fusarium solani and Fusarium oxysporum causing black rot in Gastrodia elata. Int. J. Mol. Sci. 24. doi: 10.3390/ijms24054545
Lima, L. K. S., de Jesus, O. N., Soares, T. L., de Oliveira, S. A. S., Haddad, F., and Girardi, E. A. (2019). Water deficit increases the susceptibility of yellow passion fruit seedlings to Fusarium wilt in controlled conditions. Sci. Hortic. (Amsterdam). 243, 609–621. doi: 10.1016/j.scienta.2018.09.017
Liu, M., Guo, T., Yan, H., Yuan, Y., Xiao, Z., Liu, Y., et al. (2025). Pathogen identification and biological fungicides screening for Plumbago auriculata blight in China. Front. Microbiol. 16. doi: 10.3389/fmicb.2025.1609944
Londoño, J. (2012). Evaluación de la resistencia genética de especies de Passiflora spp a Fusarium spp, agente causal de la “secadera.” (Universidad Nacional de Colombia).
López Plazas, A. (2020). Native purple passion fruit is the key to exporting elite fruits. Period. Unal. Available online at: https://unperiodico.unal.edu.co/pages/detail/native-purple-passion-fruit-is-the-key-to-exporting-elite-fruits/ (Accessed December 12, 2024).
López-Santiago, J., Nieto-Ánge., R., Barrientos-Priego, A. F., Rodríguez-Pérez, E., Colinas-Leon, M. T., Borys, M. W., et al. (2008). Redalyc.Selección de variables morfológicas para la caracterización del tejocote (Crataegus spp.). Revista Chapingo Serie Horticultura. 14, 97–111. doi: 10.4454/JPP.V99I3.3951
Margaritopoulou, T., Toufexi, E., Kizis, D., Balayiannis, G., Anagnostopoulos, C., Theocharis, A., et al. (2020). Reynoutria sachalinensis extract elicits SA-dependent defense responses in courgette genotypes against powdery mildew caused by Podosphaera xanthii. Sci. Rep. 10, 3354. doi: 10.1038/s41598-020-60148-6
Martínez, M. A., Morillo, A. C., and Reyes-Ardila, W. (2020). Characterization of the genetic diversity in Passiflora spp. in the Boyacá department, Colombia. Chil. J. Agric. Res. 80, 342–351. doi: 10.4067/S0718-58392020000300342
Mehmood, N., Saeed, M., Zafarullah, S., Hyder, S., Rizvi, Z. F., Gondal, A. S., et al. (2023). Multifaceted impacts of plant-beneficial Pseudomonas spp. in managing various plant diseases and crop yield improvement. ACS Omega 8, 22296–22315. doi: 10.1021/acsomega.3c00870
Melo, N.J.D.A, Negreiros, A. M. P., Medeiros, H.L.D.S, and Sales Júnior, R. (2020). Evaluation of Fusarium wilt disease in passion fruit species inoculated with Fusarium oxysporum f.sp. passiflorae. J. Phytopathol. 168, 81–87. doi: 10.1111/jph.12871
Miguel-Wruck, D. S., Roncatto, G., Behling, M., Faleiro, V.D.O, Bonaldo, S. M., and Tardin, F. D. (2021). Identification of sources of resistance of passiflora rootstocks to fusariosis in areas with disease outbreaks in mato grosso state, Brazil. Rev. Bras. Frutic. 43, 1–9. doi: 10.1590/0100-29452021160
Mikaberidze, A., McDonald, B. A., and Bonhoeffer, S. (2014). Can high-risk fungicides be used in mixtures without selecting for fungicide resistance? Phytopathology 104, 324–331. doi: 10.1094/PHYTO-07-13-0204-R
Ministerio de Agricultura y Desarrollo Rural (MADR) (2025). Evaluaciones Agropecuarias Municipales 2024 (Bogotá D. C). Available online at: https://upra.gov.co/es-co/eva-2024 (Accessed April 14, 2025).
Molano-Avellaneda, Z., Miranda-Lasprilla, D., and Ocampo-Pérez, J. (2020). Progress in the study of cholupa (Passiflora Maliformis L.) phenology in producing areas of Colombia. Rev. Colomb. Cienc. Hortic. 14, 32–43. doi: 10.17584/rcch.2020v14i1.11251
Moraes, A. M., Milward-de-Azevedo, M. A., Menini Neto, L., and de Faria, A. P. G. (2020). Distribution patterns of Passiflora L. (Passifloraceae s.s.) in the Serra da Mantiqueira, Southeast Brazil. Braz. J. Bot. 43, 999–1012. doi: 10.1007/s40415-020-00665-w
Muche, M. and Yemata, G. (2022). Epidemiology and pathogenicity of vascular wilt of chickpea (Cicer arietinum L.) caused by Fusarium oxysporum f. sp. ciceris, and the host defense responses. South Afr. J. Bot. 151, 339–348. doi: 10.1016/j.sajb.2022.10.008
Mukoye, B., Macharia, I., and Avedi, E. (2022). Distribution of passion fruit (Passiflora spp.) pests in Kenya. Afr. Phytosanitary J. 3, 47–55. doi: 10.52855/quyk4062
Namiki, F., Shiomi, T., Kayamura, T., and Tsuge, T. (1994). Characterization of the formae speciales of Fusarium oxysporum causing wilts of cucurbits by DNA fingerprinting with nuclear repetitive DNA sequences. Appl. Environ. Microbiol. 60, 2684–2691. doi: 10.1128/aem.60.8.2684-2691.1994
Nassimbwa, F., Matovu, E., Kiggundu, A., Changa, C., and Sseremba, G. (2022). Identity of Fusarium species associated with collar rot and wilt in passion fruit (Passiflora edulis). Student’s J. Heal. Res. Afr. 3, 10. doi: 10.51168/sjhrafrica.v3i12.246
Ożarowski, M. and Karpiński, T. (2021). Extracts and flavonoids of Passiflora species as promising anti-inflammatory and antioxidant substances. Curr. Pharm. Des. 27, 2582–2604. doi: 10.2174/1381612826666200526150113
Oliveira, D., Sobreira, R., Heinrich, L., Alvarez, P., Aparecida, P., Lima, M., et al. (2022). Passiflora mucronata, a passion fruit wild species resistant to fusariosis and a potential rootstock for commercial varieties. Sci. Hortic. (Amsterdam). 302. doi: 10.1016/j.scienta.2022.111174
Ongena, M. and Jacques, P. (2008). Bacillus lipopeptides: versatile weapons for plant disease biocontrol. Trends Microbiol. 16, 115–125. doi: 10.1016/j.tim.2007.12.009
Ortiz, E., Cruz, M., Melgarejo, L. M., Marquínez, X., and Hoyos-Carvajal, L. (2014). Histopathological features of infections caused by Fusarium oxysporum and F. solani in purple passionfruit plants (Passiflora edulis Sims). Summa Phytopathol. 40, 134–140. doi: 10.1590/0100-5405/1910
Ortiz, E. and Hoyos-Carvajal, L. (2016). Standard methods for inoculations of F. oxysporum and F. solani in Passiflora. Afr. J. Agric. Res. 11, 1569–1575. doi: 10.5897/AJAR2015.10448
Panth, M., Hassler, S. C., and Baysal-Gurel, F. (2020). Methods for management of soilborne diseases in crop production. Agriculture 10. doi: 10.3390/agriculture10010016
Papadaki, A., Bletsos, F., Menexes, G., Moustafa, A., and Lagopodi, A. (2017). Efectiveness of six rootstocks for fusarium wilt in Cucumber and their influence on growth, yield and fruit quality characteristics. J. Plant Pathol. 99, 643–650.
Patiño Pacheco, M. J. (2020). Evaluación de la resistencia de introducciones de Passifloras spp. a la “secadera” causada por Fusarium spp. de zonas productoras (Bogotá D.C., Colombia: Universidad Nacional de Colombia).
Patiño-Pacheco, M. J. and Pérez-Cardona, O. Y. (2021). Evaluación de la resistencia de genotipos de passifloras a Fusarium solani f. sp. passiflorae en granadilla. Entramado 17, 256–267. doi: 10.18041/1900-3803/entramado.2.6820
Pereira, Z. C., Cruz, J. M., dos, A., Corrêa, R. F., Sanches, E. A., Campelo, P. H., et al. (2023). Passion fruit (Passiflora spp.) pulp: a review on bioactive properties, health benefits and technological potential. Food Res. Int. 166. doi: 10.1016/j.foodres.2023.112626
Pérez, W. and Forbes, G. (2008). Manual Técnico: El tizón tardío de la papa (Lima, Perú: Comercial Gráfica Sucre Tiraje).
Pires, R. A., Jesus, O. N., Lima, L. K. S., Silva, L. N., and Laranjeira, F. F. (2022). Fusarium oxysporum f. sp. passiflorae isolates display variable virulence in Passiflora edulis Sims seedlings. Eur. J. Plant Pathol. 162, 465–476. doi: 10.1007/s10658-021-02417-2
Ploetz, R. C. (2005). Fusarium -induced diseases of tropical perennial crops. Phytopathology 96, 648–652. doi: 10.1094/phyto-96-0648
Rahut, D. B., Aryal, J. P., Manchanda, N., and Sonobe, T. (2022). “Expectations for household food security in the coming decades: A global scenario,” in Future Foods. Ed. Bhat, R. (Tartu, Estonia: Academic Press), 107–131. doi: 10.1016/B978-0-323-91001-9.00002-5
Ramaiya, S. D., Bujang, J. S., and Zakaria, M. H. (2014). Genetic diversity in Passiflora species assessed by morphological and ITS sequence analysis. Sci. World J. 2014. doi: 10.1155/2014/598313
Rios Ribeiro, L., De Oliveira E Silva, S., Santos De Oliveira, S. A., Amorim, E. P., Almeida, J., Serejo, S., et al. (2018). Sources of resistance to Fusarium oxysporum f. sp. cubense in banana germplasm. Rev. Bras. Frutic. Jaboticabal 40, 1–8. doi: 10.1590/0100-29452018202
Rocha, L. D. S., Xavier, A. A., and Ribeiro, R. C. F. (2021). Reaction of passion fruit genotypes to the complex meloidogyne incognita and Fusarium solani. Rev. Caatinga 34, 605–613. doi: 10.1590/1983-21252021v34n312rc
Rodriguez, H., Gomez, E., and Marmolejo (1999). Alternativas de manejo de la enfermedad pudrición seca o secadera del maracuyá Passiflora edulis var. flavicarpa en la zona norte del Valle y Cauca (Sede Palmira: Universidad Nacional de Colombia).
Roncatto, G., Wruck., D.S.M., Bothelho, S.D.C.C, Oliveira, S. S., and Romano, M. R. (2021). Fruit production by non-grafted and grafted Passiflora edulis brs gigante amarelo in Terra Nova Do Norte, Mato Grosso, Brazil. Int. J. Dev. Res. 11, 50251–50254. doi: 10.37118/ijdr.22780.09.2021
Sabogal-Palma, A. C., Chávez-Marín, J., Oliveros-Gómez, D. F., Murillo-Perea, E., and Méndez-Arteaga, J. J. (2016). Biological characteristics of Passiflora Maliformis from south Colombian massif. Bioagro 28, 3–12. Available online at: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84960406325&partnerID=40&md5=03e87d4ef6d0bfa14d4a5893ec4f3fc5 (Accessed January 25, 2024).
Salazar-Gonzalez, C., Yela-Caicedo, O., and Gomez-Espinoza, B. (2022). Molecular characterization of Fusarium spp. associated vascular wilt in passion fruit (Passiflora ligularis JUSS). Rev. Cienc. Agrícolas 39, 33–46. doi: 10.22267/rcia.223902.180
Sampaio, A. M., De Sousa Araújo, S., Rubiales, D., and Patto, M. C. V. (2020). Fusarium wilt management in legume crops. Agronomy 10, 1–25. doi: 10.3390/agronomy10081073
Sañudo, B. (2012). Introducción al manejo de frutales andino en la zona triguera baja de Nariño (Pasto, Colombia: Universidad de Nariño).
Silva Flores, P., Campos Otoni, W., Dev Dhingra, O., Severo de Souza Diniz, S. P., Miranda dos Santos, T., and Horst Bruckner, C. (2012). In vitro selection of yellow passion fruit genotypes for resistance to Fusarium vascular wilt. Plant Cell. Tissue Organ Cult. 108, 37–45. doi: 10.1007/s11240-011-0009-5
Suansia, A. and Chandra Samal, K. (2021). Vegetable grafting: a sustainable and eco-friendly strategy for soil-borne pest and disease management. J. Pharmacogn. Phytochem. 10, 1634–1642.
Torres, C., Sánchez de P, M., Bravo Otero, N., Marmolejo de la Torre, F., and Gómez, E. D. (1999). Enfermedades fungosas y bacterianas en el cultivo del maracuyá Passiflora edulis Sims var. flavicarpa (Cali, Colombia: Universidad de Colombia, Universidad del Valle).
Townsend, G. R. and Heuberger, J. W. (1943). Methods for estimating losses caused by diseases in fungicide experi ments. Plant Dis. Rep. XXVII, 340–343.
Verma, M., Brar, S. K., Tyagi, R. D., Surampalli, R. Y., and Valéro, J. R. (2007). Antagonistic fungi, Trichoderma spp.: panoply of biological control. Biochem. Eng. J. 37, 1–20. doi: 10.1016/j.bej.2007.05.012
Wang, Y., Chen, C., Li, J., Zhang, X., Teng, Y., Zhang, Z., et al. (2024). Seed characteristics of passion fruit (Passiflora edulis f. flavicarpa) rootstocks with different genotypes and responses of their seedlings to drought stress. Hortic. Sci. Technol. 42, 312–325. doi: 10.7235/HORT.20240027
Wang, Y., Teng, Y., Zhang, J., Zhang, Z., Wang, C., Wu, X., et al. (2023). Passion fruit plants alter the soil microbial community with continuous cropping and improve plant disease resistance by recruiting beneficial microorganisms. PloS One 18, e0281854. doi: 10.1371/journal.pone.0281854
White, T. J., Bruns, T., Lee, S., and Taylor, J. (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 105, 315–322. doi: 10.1016/b978-0-12-372180-8.50042-1
Williams, B., Ahsan, M. U., and Frank, M. H. (2021). Getting to the root of grafting-induced traits. Curr. Opin. Plant Biol. 59, 101988. doi: 10.1016/j.pbi.2020.101988
Yadeta, K. A. and Thomma, B. P. H. (2013). The xylem as battleground for plant hosts and vascular wilt pathogens. Front. Plant Sci. 4. doi: 10.3389/fpls.2013.00097
Yao, X., Guo, H., Zhang, K., Zhao, M., Ruan, J., and Chen, J. (2023). Trichoderma and its role in biological control of plant fungal and nematode disease. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1160551
Yockteng, R. and Nadot, S. (2004). Phylogenetic relationships among Passiflora species based on the glutamine synthetase nuclear gene expressed in chloroplast (ncpGS). Mol. Phylogenet. Evol. 31, 379–396. doi: 10.1016/S1055-7903(03)00277-X
Keywords: Fusarium wilt, stem rot, Passiflora maliformis, histopathology, disease resistance, rootstock potential
Citation: Galvis-Tarazona D, Bohórquez-Quintero M, Arias-Moreno D and Ojeda-Pérez Z (2025) Deciphering the resistance mechanism to Fusarium wilt and stem rot of Passiflora maliformis var. pubescens using histopathology aspects. Front. Plant Sci. 16:1635702. doi: 10.3389/fpls.2025.1635702
Received: 26 May 2025; Accepted: 15 September 2025;
Published: 07 October 2025.
Edited by:
Solomon Abate Mekonnen, Ethiopian Institute of Agricultural Research, EthiopiaReviewed by:
Malkhan Singh Gurjar, Indian Agricultural Research Institute (ICAR), IndiaSergii Krysenko, Valent BioSciences LLC, United States
Afeez Adesina Adedayo, Western Illinois University, United States
Copyright © 2025 Galvis-Tarazona, Bohórquez-Quintero, Arias-Moreno and Ojeda-Pérez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Diana Arias-Moreno, ZG1hcmlhc21AdXQuZWR1LmNv
†ORCID: Daicy Galvis-Tarazona, orcid.org/0000-0002-2251-0015
María Bohórquez-Quintero, orcid.org/0000-0003-1919-4346
Diana Arias-Moreno, orcid.org/0000-0001-6171-0549
Zaida Ojeda-Pérez, orcid.org/0000-0003-1630-6699