 Eliana Monteiro1,2
Eliana Monteiro1,2 Paula Baptista3
Paula Baptista3 Sofia Silva3
Sofia Silva3 Márcia Carvalho1,2,4
Márcia Carvalho1,2,4 Radek Bragança5
Radek Bragança5 Kieran J. Guinan6
Kieran J. Guinan6 Neerakkal Sujeeth6
Neerakkal Sujeeth6 Isabel Cortez1,2,7
Isabel Cortez1,2,7 Berta Gonçalves1,2,8
Berta Gonçalves1,2,8 Isaura Castro1,2,4*
Isaura Castro1,2,4*- 1Centre for the Research and Technology of Agro-Environmental and Biological Sciences (CITAB), University of Trás-os-Montes e Alto Douro (UTAD), Vila Real, Portugal
- 2Institute for Innovation, Capacity Building and Sustainability of Agri-food Production (Inov4Agro) (UTAD), Vila Real, Portugal
- 3Mountain Research Center (CIMO), Polytechnic Institute of Bragança, Bragança, Portugal
- 4Department of Genetics and Biotechnology, University of Trás-os-Montes e Alto Douro (UTAD), Vila Real, Portugal
- 5BioComposites Centre, Bangor University, Bangor, United Kingdom
- 6Clash Industrial Estate, BioAtlantis Ltd., Tralee, Ireland
- 7Department of Agronomy, University of Trás-os-Montes e Alto Douro (UTAD), Vila Real, Portugal
- 8Department of Biology and Environment, University of Trás-os-Montes e Alto Douro (UTAD), Vila Real, Portugal
Introduction: Grapevine is highly susceptible to fungal diseases such as downy mildew and powdery mildew, which are traditionally managed through the intensive use of chemical fungicides. However, in the context of increasingly sustainable viticulture, biofungicides derived from plant and yeast extracts are gaining attention. Despite this, their impact on the grapevine leaf microbiome, crucial for plant health and disease resilience, remains underexplored.
Material and methods: This study evaluated the effects of foliar applications of biofungicides (nettle extract, Japanese knotweed extract, and a yeast-based formulation - T66 and T90) in comparison with conventional chemical treatments and control (no treatment). Over two consecutive growing seasons, high-throughput sequencing was used to assess the diversity and composition of fungal and bacterial communities on grapevine leaves.
Results: Bacterial communities were more sensitive to treatments and interannual variability than fungal communities, which remained relatively stable. Conventional treatment (CT) showed the highest influence on fungal and bacterial composition, reducing the diversity of both. Some important fungal (Aureobasidium and Sporobolomyces) and bacterial (Pseudomonas and Sphingomonas) genera associated with the promotion of plant growth, health, and biocontrol were detected.
Discussion: These findings reinforce the potential of new treatments with putative fungicide effects to modulate the leaf microbiome, particularly bacterial communities, without disrupting the natural fungal balance. Thus highlight their relevance for developing sustainable viticultural practices aimed at improving plant protection.
1 Introduction
There is a growing awareness of the environmental and health impacts of indiscriminate chemical pesticide use in agriculture. In line with the Green Deal’s “Farm to Fork” strategy, the European Commission aims for ‘at least 50% of the EU’s agricultural land to be under organic farming by 2030’. It is expected that stricter controls on certain fungicides will lead to their potential withdrawal from the market in the near future. Challenges are also being faced in controlling certain fungal diseases, which may require more frequent spraying and repeated usage of the same active compound, potentially contributing to the emergence of pathogen resistance (Koledenkova et al., 2022; Monteiro et al., 2022). Furthermore, climate change characterized by dry and hot periods followed by unexpected rainfall, affects grapevine growth cycles and extends the window of susceptibility to fungal diseases, making its control more challenging particularly as they can develop more quickly and aggressively (Elad and Pertot, 2014; Monteiro et al., 2022).
The Demarcated Douro Region (DDR) is the largest and most heterogeneous mountainous wine region in the world and known for its unique terroir and Mediterranean climate (Fraga et al., 2017). Downy mildew (caused by Plasmopara viticola (Berk. & M.A. Curtis) Berl. & De Toni) and powdery mildew (caused by Erysiphe necator (Schwein.)), are obligate biotrophic pathogens and major contributors to some of the most problematic diseases in grape production (Karthick et al., 2019; Koledenkova et al., 2022). These diseases cause both quantitative and qualitative losses, affecting grape clusters (reducing harvest yields) and leaves (Karthick et al., 2019; Koledenkova et al., 2022). The increased difficulty in controlling these diseases combined with the need to maintain high productivity, has led farmers to adopt intensive crop protection practices resulting in high production costs (Ortega et al., 2023). In response, there is a growing need for sustainable farming systems that reduce chemical pesticides use. Plant- and yeast-based formulations offer promising alternatives. For instance, nettle extracts are rich in compounds such as saponins, flavonoids, tannins, proteins, and amino acids (Joshi et al., 2014), and Japanese knotweed extracts contain bioactive compounds like resveratrol and emodin (Oleszek et al., 2019; Borovaya et al., 2020). Both plant extracts demonstrated potential to improve the physiological and biochemical parameters of the grapevine (Monteiro et al., 2024a, 2024b) and presented antimicrobial effects (Rodino et al., 2018; Ghazal et al., 2019). Yeasts, commonly used in various industries, also exhibit plant-protective properties, including the production of volatile compounds, toxins, lytic enzymes, and the induction of plant resistance pathways (Puccioni et al., 2025). Studies have demonstrated their ability to enhance grape and wine quality and control downy mildew (Garde-Cerdán et al., 2017; Puccioni et al., 2025).
Despite these advances, the effect of biofungicides on the plant microbiome particularly in grapevines remains underexplored. The interactions between plants and their associated microbiomes, whether in the rhizosphere or phyllosphere, play a critical role in plant health, productivity, and stress resilience (Müller et al., 2016; Colla et al., 2017). Understanding the microbial community composition can provide valuable insights for managing disease pressure and improving grape quality (Leveau and Tech, 2011). High-throughput sequencing technologies have enabled a better understanding of the grapevine microbiome, revealing insights into both above- and below-ground microbial communities (Wei et al., 2018; Fadiji and Babalola, 2020).
Given the global importance of viticulture, and particularly in the DDR, this study aimed to investigate the effects of foliar applications of biofungicides derived from plant and yeast extracts in comparison with conventional chemical treatments. Over two consecutive years, we assessed their impact on the diversity and composition of fungal and bacterial communities on grapevine leaves, using metagenomic tools to analyze the grapevine leaf microbiome, which plays a key role in both grape production and wine quality.
2 Materials and methods
2.1 Plant material and sampling
Samples were obtained from the black skinned Vitis vinifera cv. ‘Touriga Franca’, grafted on 1103P rootstock, in two growing seasons (2020 and 2021). The trial was installed in an experimental vineyard located at University of Trás-os-Montes e Alto Douro (41°17’14.8”N 7°44’14.8”W, 500m above sea level), Baixo Corgo sub-region of the Douro Demarcated Region, Vila Real, northern Portugal. The topsoil (0–20 cm) exhibited a medium texture, 2.24% of organic matter, 47 mg/kg of P2O5, 104 mg/kg of K2O and a pH (KCl) 5.5. Vines were managed in rainfed conditions and grown using standard cultural practices commonly employed by commercial farmers. Vines were trained to bilateral Guyot and pruned to 12 buds per plant and spaced 2.20 m × 1 m between and along the rows. This area has Mediterranean climatic characteristics, with a warm-temperate climate with dry and hot summers, and higher precipitation during the winter months and very low during the summer. Monthly temperature and precipitation values were recorded by a weather station located near to the experimental site and are shown in Supplementary Figure 1.
Three replicates of six plants per treatment randomized along four vineyard rows of 36 plants were sprayed between leaves unfolded (BBCH 11) and veraison (BBCH 81) (Lorenz et al., 1995). Foliar sprayings were conducted during the morning, covering the whole canopy and according to disease incidence and weather conditions. Six different foliar treatments were tested, namely: i) nettle extract - NE (Urtica sp.; Bangor University) (3%); ii) Japanese knotweed extract - JKE (Reynoutria japonica; Bangor University) (4.5%); iii) T66 - Yeast-based formulation (1%; BioAtlantis Ltd.); iv) T90 - Yeast-based formulation (1%; BioAtlantis Ltd.); v) conventional treatment - CT (4.8% (p/p) cymoxanil + 40% (p/p) folpet + 8% (p/p) metalaxyl (SAPEC) or 10.1% (p/p) penconazole (SAPEC) or 50% (p/p) tebuconazole + 25% (p/p) trifloxystrobin (BAYER) or 8% (p/p) cymoxanil + 66% (p/p) folpet (ASCENZA) or 50% (p/p) kresoxim-methyl (BASF)) (prepared according to the manufacturer guidelines), and vi) control - C (water) (Supplementary Tables 1, 2). T66 and T90 are proprietary biological formulations derived from microbial fermentation, developed by BioAtlantis Ltd. These formulations contain proprietary strains of yeast: in formulation T66 exhibits moderate to low tolerance to salt, whereas the strain in formulation T90 demonstrates a high tolerance, capable of withstanding sodium chloride concentrations of up to 25%. The strains were suspended in water along with a stabilizing agent.
The applications were made in 2020 (NE20, JKE20, T6620, T9020, CT20, C20) (Supplementary Table 1) and in 2021 (NE21, JKE21, T6621, T9021, CT21, C21) (Supplementary Table 2). At harvest (BBCH 89), leaves were sampled directly to liquid nitrogen, taken from the three randomized blocks of six plants in each of the six treatment conditions. The samples were stored at –80°C until analysis.
2.2 DNA extraction
DNA was extracted from leaves using the DNeasy® Plant Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer’s protocol. Briefly, leaf material was macerated in liquid nitrogen, and about 100 mg of leaf powder was used. DNA concentration was determined by UV spectrometer (Nanodrop® ND-1000, Thermo Fisher Scientific, USA) and stored at -20°C until metagenomic analysis.
2.3 Metagenomic analysis
For microbial community profiling, DNA samples were sent to Novogene Co., Ltd. (Cambridge, UK) for high-throughput sequencing. The bacterial and fungal communities were characterized through 16S rRNA gene (V3-V4), ITS1 and ITS2 regions sequencing, respectively (Supplementary Table 3). Library preparation, sequencing, and initial quality control were performed by Novogene according to standard protocols. The sequencing was conducted on the Illumina platform, using 2 × 250 bp paired-end chemistry to generate raw reads. The resulting raw data were processed using QIIME1 for quality filtering, OTU clustering and taxonomic classification.
2.4 Data analysis
Diversity metrics, including observed species richness and the Shannon-Weiner diversity index, were calculated using PAST v4.03 (Hammer et al., 2001) to assess within-sample microbial diversity. The Krona charts were performed according to the method described by Ondov et al. (2013) for visualizing and exploring the taxonomic composition of metagenomic data. Differences in community composition among sample groups were evaluated using PERMANOVA (Permutational Multivariate Analysis of Variance), also performed in PAST, based on Bray–Curtis dissimilarity matrices (Anderson, 2001). Further multivariate analyses were carried out using the Community Analysis Package v3.01 (CAP) (Clarke, 1993). Non-metric multidimensional scaling (NMDS) was used to visualize patterns in microbial community composition across samples. The Kruskal’s stress value was calculated to assess the goodness-of-fit of the NMDS ordination, with lower values indicating a better representation of the original dissimilarity data in reduced dimensions (Kruskal, 1964). Similarity Percentage (SIMPER) analysis was applied to identify the taxa contributing most to the observed differences between groups. Analysis of Similarities (ANOSIM) was used to statistically test the dissimilarity in community structure among predefined groups. ANOSIM generates a p-value (with significance considered at p < 0.05) and an R2, which quantify the degree of separation between groups: values close to 0 indicate little to no separation, while values approaching 1 suggest complete dissimilarity (Clarke, 1993). All statistical analyses were performed with a significant level of p < 0.05.
To explore relationships between bacterial and fungal taxa, a Spearman correlation analysis was performed between the relative abundances of bacterial and fungal genera across all treatments. Only genera representing more than 0.5% of the total relative abundance in the complete dataset were included. All statistical analyses were performed with a significant level of p < 0.05.
3 Results and discussion
3.1 Diversity and composition of fungal community
The richness (number of identified taxa) and the Shannon-Weiner index (Figure 1) were calculated to estimate fungal diversity. The treatments NE, T90, and T66 presented higher richness in both years compared to C suggesting that these treatments increased fungal diversity. In contrast, JKE20, JKE21, and CT20 have shown lower richness values indicating a reduction in taxa diversity under these conditions. Among all treatments, T6620 showed the highest richness, whereas CT20 consistently displayed the lowest, with the differences between these two treatments being statistically significant. Significant differences between years for richness were also observed in the treatments JKE and C.
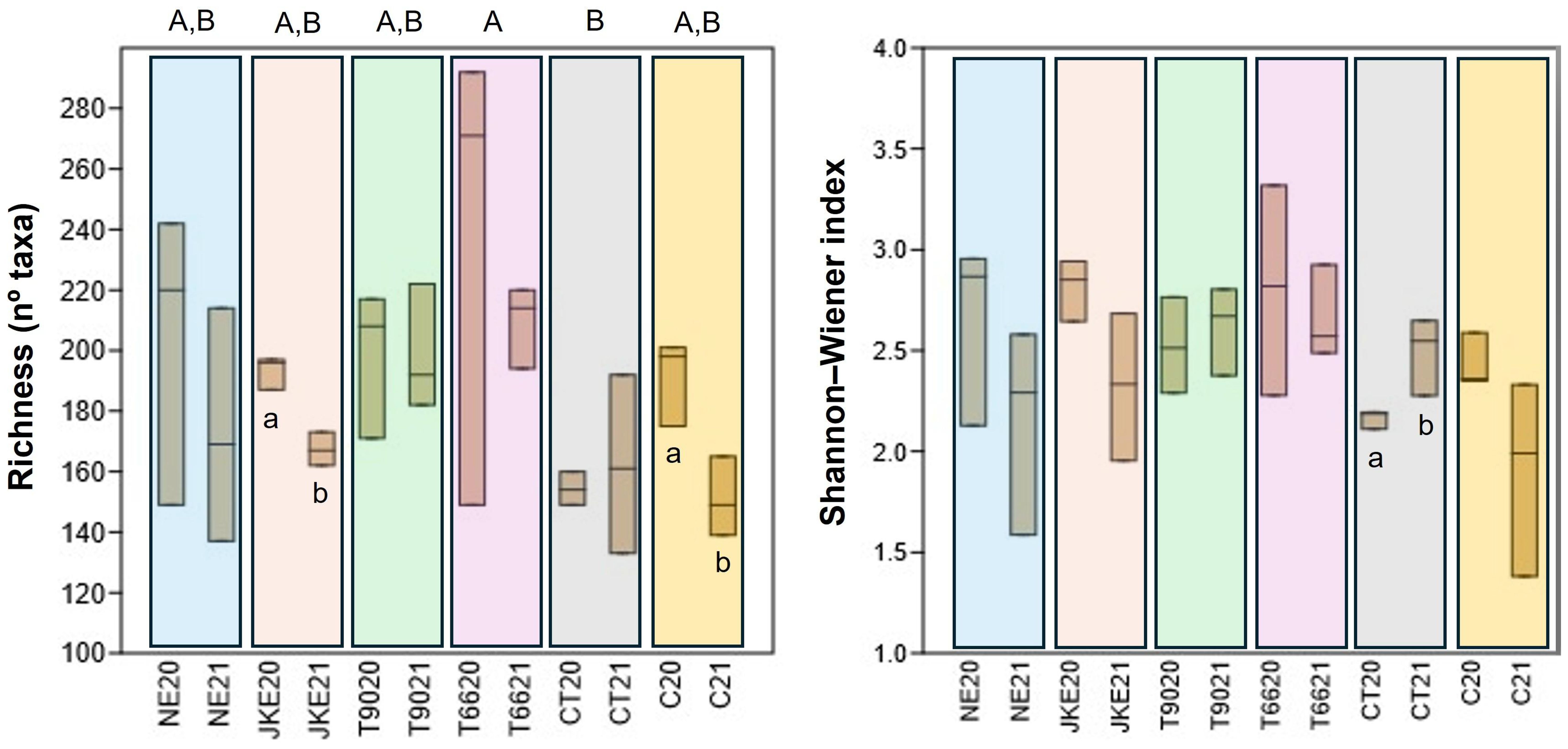
Figure 1. Boxplots showing the fungal community diversity (Richness and Shannon-Weiner index) of cv. ‘Touriga Franca’ under six different foliar treatments at harvest of 2020 and 2021. Different letters mean significant differences between treatments (uppercase) and differences between years for the same treatment (lowercase); no letters mean no-significant differences. NE, nettle extract; JKE, Japanese knotweed extract; T66, yeast extract; T99, yeast extract; CT, conventional treatment; C, control.
No significant differences between treatments were detected for the Shannon-Weiner index. However, in terms of inter-annual variation in CT plants there was a significant increase in diversity from 2020 to 2021. The treatments CT and C presented lower Shannon-Weiner values compared to the other treatments, suggesting that plant- and yeast-based extracts contributed to enhance diversity of community balance.
To further investigate treatment and year effects on fungal community composition, Bray–Curtis dissimilarity analysis was applied and visualized via non-metric multidimensional scaling (NMDS) plots (Figure 2; Supplementary Figure 2), with PERMANOVA results shown in Supplementary Table 4. The PERMANOVA analysis revealed a significant effect of treatment (p = 0.0002), followed by year (p = 0.0015), explaining approximately 56% and 10% of the variation in fungal community composition, respectively (Supplementary Table 4). The interaction (treatment and year) had no influence in the fungal community composition (p = 0.998) (Supplementary Table 4). These findings were corroborated by NMDS plots, which clustered the samples according to treatment and year (Figure 2; Supplementary Figure 2).
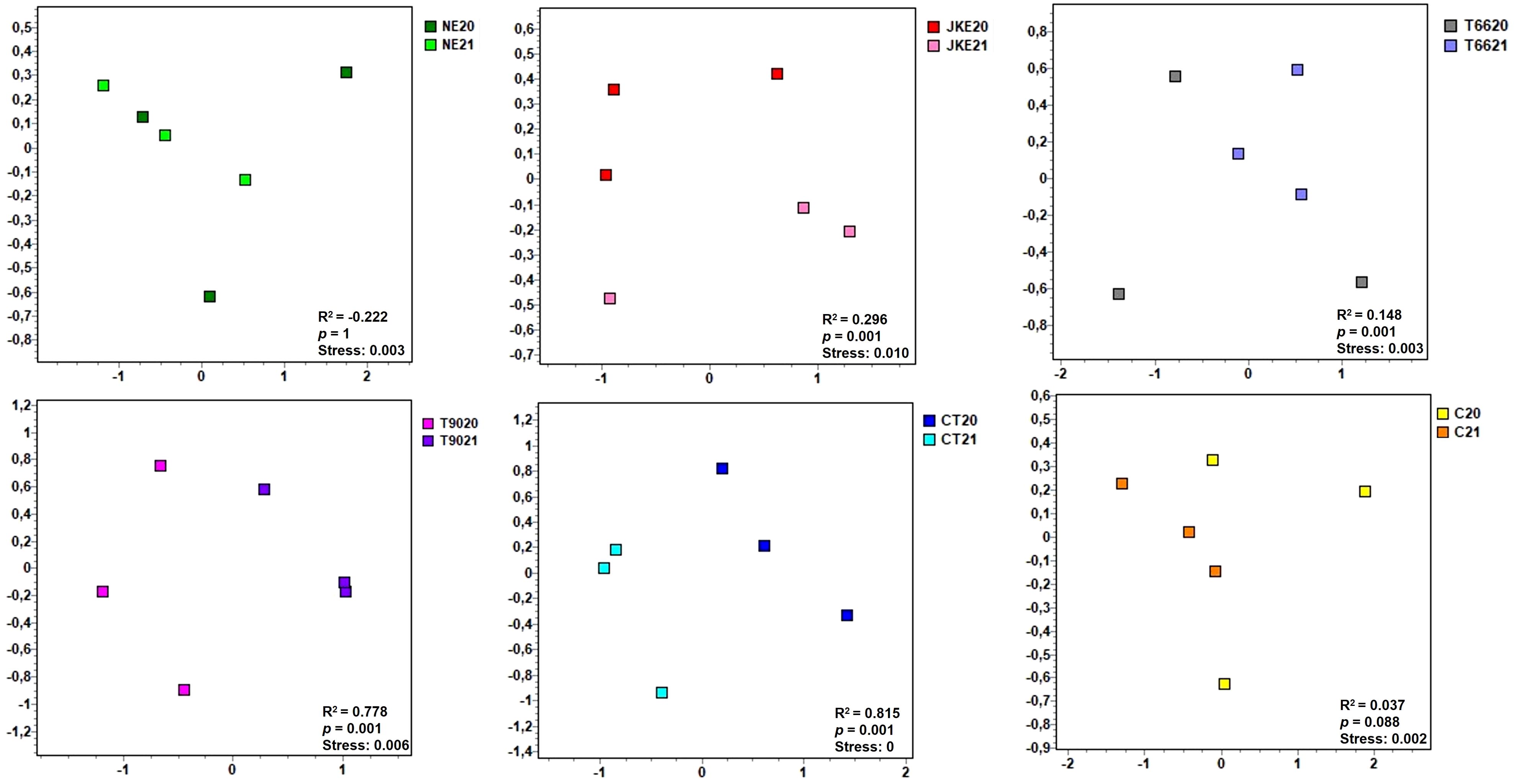
Figure 2. Non-metric multidimensional scaling (NMDS) plots of foliar fungal communities detected in cv. ‘Touriga Franca’ with six different foliar treatments at harvest of 2020 and 2021. Clustering analysis was performed with Bray-Curtis distance. Analysis of similarity (ANOSIM), based on Bray-Curtis distance and Kruskal’s stress values are displayed. NE, Nettle extract; JKE, Japanese knotweed extract; T66 and T90, Yeast extract; CT, Conventional treatment; C, Control.
In 2020 ANOSIM analysis indicated significant differences in fungal composition between treatments (R2 = 0.163, p = 0.004; Supplementary Figure 2A). The CT treatment differed significantly from all the others, while no significant differences were detected between the remaining treatments (Supplementary Table 5). In the SIMPER analysis, Erysiphe necator the causal agent of powdery mildew and one of the most damaging fungal diseases in grapevine (Wei et al., 2018; Gobbi et al., 2020), was identified as the species that most contributed to the differentiation between CT and treatments NE, T90, and C. For JKE and T66, Aureobasidium pullulans was the key species differentiating them from CT (Supplementary Table 6). A. pullulans has shown potential as a biocontrol agent against Botrytis cinerea the causal agent of bunch rot in grapevine (Karácsony et al., 2023).
In 2021, the differences between treatments were more pronounced (ANOSIM R2 = 0.418, p = 0.001; Supplementary Figure 2B). According to ANOSIM analysis treatments CT and C differed significantly from each other and from the other treatments, which revealed no significant difference among themselves (Supplementary Table 7). SIMPER analysis indicated as in 2020 that E. necator differentiated C from all the other treatments (JKE, T66, T90, CT) and also contributed to the differentiation of CT from NE, while Sporobolomyces roseus differentiated CT from T66 and T90 (Supplementary Table 8). S. roseus has been reported to reduce post-harvest diseases in apples (Janisiewicz et al., 1994), suggesting potential applications in grapevine protection.
When comparing the inter-annual variation for each treatment NE (ANOSIM R2 = -0.222, p = 1) and C (ANOSIM R2 = -0.037, p = 0.088) showed no significant differences in fungal composition. T66 (ANOSIM R2 = 0.148, p = 0.001) displayed minimal differences (Figure 2). JKE (ANOSIM R2 = 0.296, p = 0.001) showed moderate inter-annual variation, while T90 (ANOSIM R2 = 0.778, p = 0.001) and CT (ANOSIM R2 = 0.815, p = 0.001) exhibited the highest variation (Figure 2). SIMPER analysis indicated that E. necator was the species that differentiated the years in JKE, T66, T90 and C, whereas in CT, the key differentiating species was A. pullulans (Supplementary Table 9).
A total of five fungal phyla were identified in both years (Supplementary Figure 3). The phylum Chytridiomycota was exclusive in 2020 and the Glomeromycota exclusive in 2021. Both years have four phyla in common. Ascomycota and Basidiomycota were the predominant phyla in all treatments in both years (Supplementary Figure 3), aligning with previous studies on grapevine leaves fungal diversity (Zhang et al., 2017; Miliordos et al., 2021). A total of 100 fungal families were found in 2020 and 98 in 2021 (Supplementary Figure 4). At the family level, treatments CT20, CT21, and C21 were distinct from the others, showing a different taxonomic composition compared to the remaining treatments (Supplementary Figure 4). Marked inter-annual variation in fungal family abundance was observed, particularly in NE, T90, CT and C (Supplementary Figure 4). At the genus level 90 genera were identified in 2020 and 71 in 2021 (Figure 3), with the top ten genera including Erysiphe, Cladosporium, Mycosphaerella and Sporobolomyces (Figure 3; Supplementary Figure 5) in both years. Erysiphe was notably dominant, containing important phytopathogens such as E. necator, extremely important in viticulture as previously mentioned. The treatments NE, T90 and C, in 2020, and NE, JKE and C, in 2021, exhibited the highest relative abundance of Erysiphe (Figure 3). It was also verified that this genus was more predominant in 2021 than in 2020, which may have been influenced by meteorological, since 2021 was rainier than 2020 (Supplementary Figure 1) and powdery mildew infections are promoted by humidity (Belhadj et al., 2006). According to ADVID (2021) the mild temperatures recorded in the spring/summer period of 2021 created good conditions for the development of powdery mildew, that remained active, requiring in some situations a curative protection strategy until the end of the cycle.
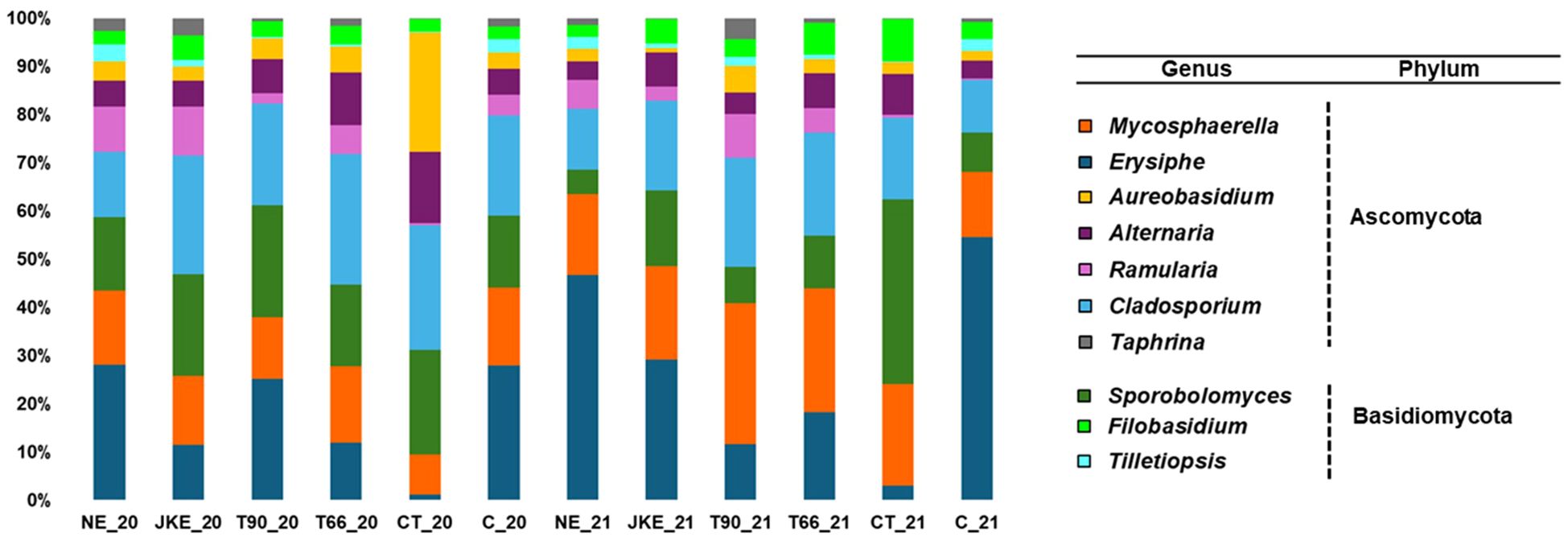
Figure 3. Relative abundance of fungal community (at genus level) detected in cv. ‘Touriga Franca’ with six different foliar treatments, at harvest of 2020 and 2021. Only the top 10 fungal genera are presented. NE, Nettle extract; JKE, Japanese knotweed extract; T66 and T90, Yeast extract; CT, Conventional treatment; C, Control.
3.2 Diversity and composition of bacterial community
To assess the influence of plant- and yeast- based extracts on bacterial diversity, richness (number of identified taxa) and Shannon-Weiner index were calculated (Figure 4). In 2020 richness values were generally lower across treatments, with CT and C showing the highest values. In 2021 richness was higher in NE, JKE, and T90, and lower in T66, CT, and C. Treatments T66 showed a significant decrease in richness from 2020 to 2021.
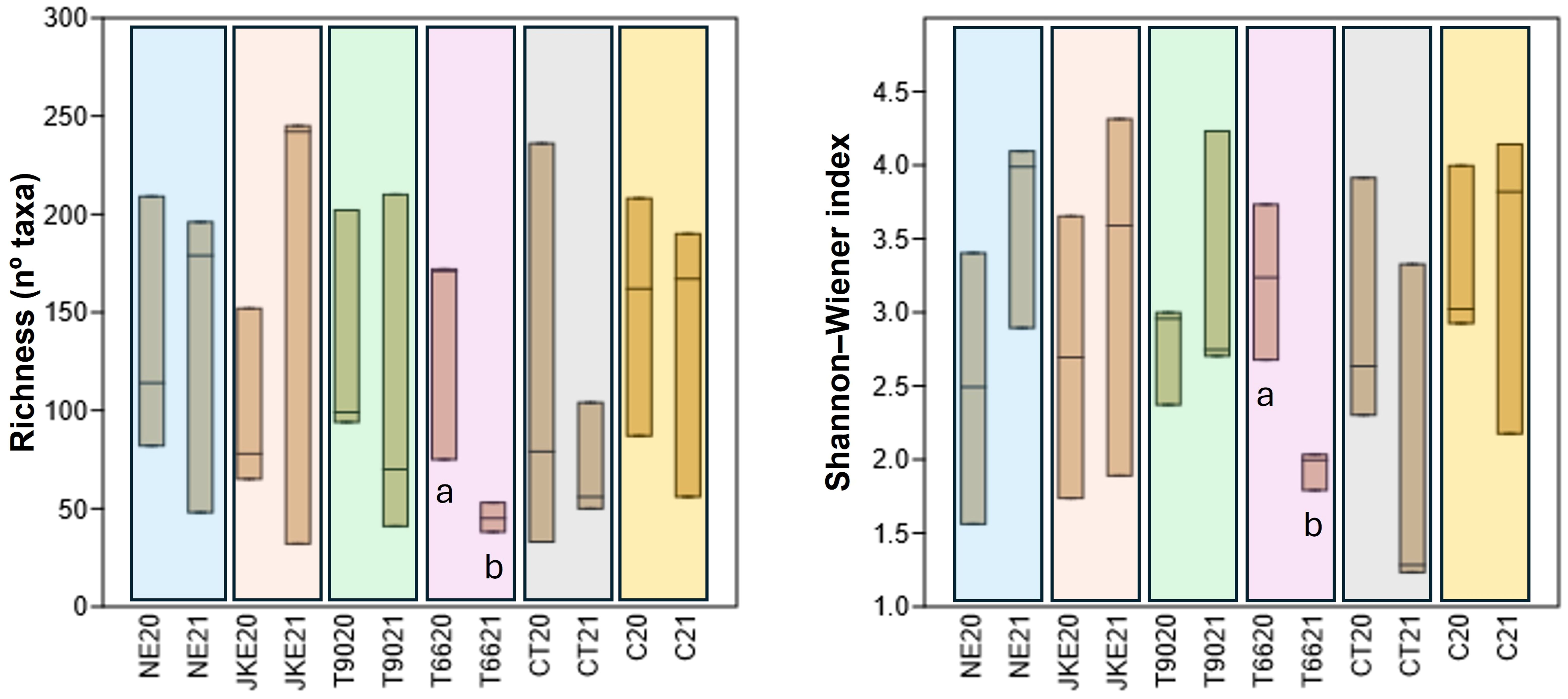
Figure 4. Boxplots showing the bacterial community diversity (Richness and Shannon-Weiner index) of cv. ‘Touriga Franca’ under six different foliar treatments at harvest of 2020 and 2021. Different letters mean significant differences between years for the same treatment; no letters mean no-significant differences. NE, Nettle extract; JKE, Japanese knotweed extract; T66 and T90, Yeast extract; CT, Conventional treatment; C, Control.
Regarding Shannon-Weiner index (Figure 4) T66 also showed a significant decrease in diversity from 2020 to 2021, consistent with the richness results. In contrast, the remaining treatments (except CT) showed an opposite trend, with increases diversity in 2021, suggesting a possible effect of the year on the bacterial community structure.
PERMANOVA analysis based on Bray–Curtis dissimilarity revealed that treatment significantly influenced bacterial composition (p = 0.044), explaining about 52% of the variation (Supplementary Table 10). However, neither year (p = 0.080) nor the interaction between treatment and year (p = 0.920) significantly affected bacterial community composition (Supplementary Table 10).
The NMDS plot based on the Bray–Curtis (Figure 4; Supplementary Figure 6) showed that in 2020 there were no significant differences in bacterial composition between treatments (ANOSIM R2 = - 0.039, p = 0.791; Supplementary Figure 6A), as confirmed by ANOSIM (Supplementary Table 11). In 2021 significant differences between treatments were observed (ANOSIM R2 = 0.163, p = 0.004), with CT being the most dissimilar (Supplementary Figure 6B). ANOSIM results confirmed that CT differed significantly from the other treatments (NE, JKE, T66, T90 and C), while T66 was also different from NE, CT and C (Supplementary Table 12). SIMPER analysis indicated that the genus Pseudomonas contributed the most to the dissimilarity between CT and the other treatments (NE, T66, T90, C) (Supplementary Table 13). Pseudomonas spp. have demonstrated biocontrol potential against grapevine trunk diseases (Niem et al., 2020), downy mildew control (Sun et al., 2024), and Botrytis cinerea, by inducing plant defense mechanisms (Verhagen et al., 2010). For T66 the genus Lactococcus was responsible for its dissimilarity from treatments NE and C (Supplementary Table 13).
When comparing each treatment individually between years (Figure 5) no significant differences were found for JKE (ANOSIM R2 = -0.185, p = 1) and C (ANOSIM R2 = -0.333, p = 1). The NE treatment presented minor but statistically significant differences between years (ANOSIM R2 = 0.111, p = 0.001) (Figure 5). Moderate differences were observed for T90 (ANOSIM R2 = 0.444, p = 0.001) and CT (ANOSIM R2 = 0.704, p = 0.001) (Figure 5). The treatment T66 exhibited pronounced differences between years (ANOSIM R2 = 1.00, p = 0.001) (Figure 5). According to the SIMPER analysis (Supplementary Table 14) the main genera contributing to the differences between years were Escherichia-Shigella (NE), Sphingomonas (T66), Delftia (T90) and, Pseudomonas (CT). Notably, Sphingomonas and Pseudomonas are associated with plant health and productivity (Gobbi et al., 2020) and have been identified as consistent components of the grapevine microbiome, including in healthy, diseased, and recovered vines (Bulgari et al., 2014).
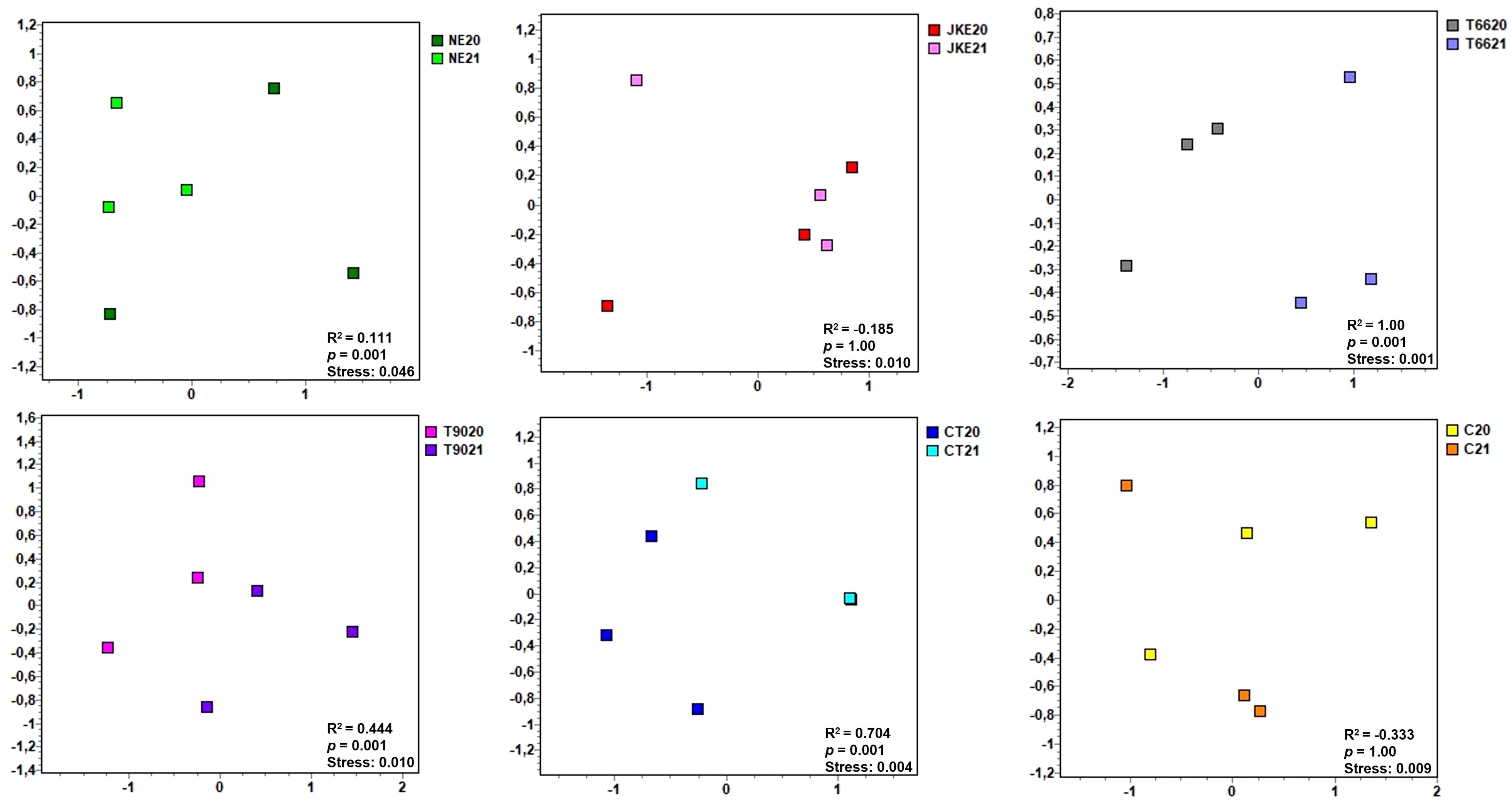
Figure 5. Non-metric multidimensional scaling (NMDS) plots of foliar bacterial communities detected in cv. ‘Touriga Franca’ with six different foliar treatments at harvest of 2020 and 2021. Clustering analysis was performed with Bray-Curtis distance. Analysis of similarity (ANOSIM), based on Bray-Curtis distance and Kruskal’s stress values are displayed. NE, Nettle extract; JKE, Japanese knotweed extract; T66 and T90, Yeast extract; CT, Conventional treatment; C, Control.
A total of 14 bacterial phyla were detected in 2020 and 13 in 2021, 12 in common in both years (Supplementary Figure 7). The phyla Chloroflexie and Latescibacterota were exclusive in 2020 and Methylomirabilota in 2021. In both years, Proteobacteria and Firmicutes were the dominant phyla (Supplementary Figure 7), which is consistent with previous studies on grapevine leaves from different regions and cultivars (Leveau and Tech, 2011; ZarraonaIndia et al., 2015; Wei et al., 2018; Gobbi et al., 2020). At family level 76 were found in 2020 and 73 in 2021, with 60 shared between both years (Supplementary Figure 8). At the genus level 164 bacteria genera were detected in 2020 and 160 in 2021, with 128 shared between the two years (Figure 6). Overall, the most abundant genera were Pseudomonas, followed by Sphingomonas (Figure 6; Supplementary Figure 9A), as previously reported in grapevine leaf microbiomes studies conducted in the field across different regions (California, Long Island and Xinjiang province) and cultivars (‘Chardonnay’, ‘Merlot’ and ‘Cabernet Sauvignon’) (Leveau and Tech, 2011; ZarraonaIndia et al., 2015; Wei et al., 2018), as well as in vitro studies (Cappelletti et al., 2016). According to the Krona charts, Sphingomonas was the most abundant genus in 2020, whereas in 2021 was Pseudomonas (Supplementary Figures 9B, C). NE was the treatment with the highest number of detected bacterial genera in 2020, while JKE presented the highest genera richness in 2021 (Figure 6).
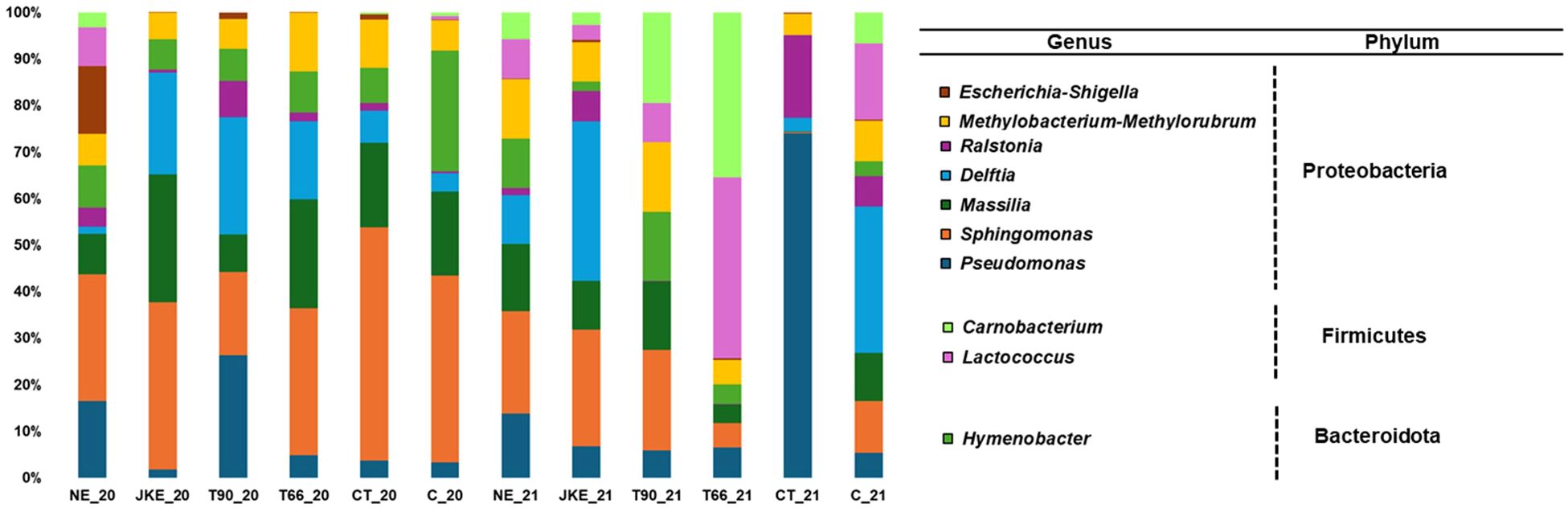
Figure 6. Relative abundance of bacterial community (at genus level) detected in cv. ‘Touriga Franca’ with different foliar treatments, at harvest of 2020 and 2021. Only the top 10 fungal genera are presented. NE, Nettle extract; JKE, Japanese knotweed extract; T66 and T90, Yeast extract; CT, Conventional treatment; C, Control.
The mechanisms by which plant- and yeast-based extracts modulate the grapevines natural defenses, as well as their general ecological impacts, particularly on the grapevine leaf microbiome, remain insufficiently explored. Therefore, in this study, we investigated the effect of biofungicides derived from plant (NE and JKE) and yeast (T66 and T90) extracts on the grapevine leaf microbiome. In this study it was possible to observe that both ‘treatment’ and ‘year’ influenced the composition of the fungal and bacterial leaves communities. Regarding the fungal community the differences can be seen in the Krona charts (Supplementary Figure 5) and in the relative abundance graphs (Figure 3; Supplementary Figures 3, 4). Among the treatments, CT appears to have the greatest influence showing the lowest richness and diversity compared to the treatments under study (NE, JKE, T66, T90) and also with C (Figure 1). Regarding the effect of the year, a larger variation was observed particularly in the NE, T90, CT, and C treatments, as evidenced by significant changes in family (Supplementary Figure 4) and genus abundance (Figure 3) between 2020 and 2021. A higher richness was observed in the bacteria community compared to the fungal community, consistent with the findings of Zhang et al. (2017) in a study on grapevine leaves (cv. ‘Kyoho’) in Beijing. The CT21 treatment showed the strongest impact on bacterial community composition, confirmed by NMDS plots, which show its clear separation from the other treatments and also supported by the ANOSIM test (Supplementary Figure 6B; Supplementary Table 12). This impact may be explained by the fact that conventional treatment (CT) is based on synthetic chemicals, and that the frequency of application of broad-spectrum fungicides may affect not only pathogens, but also beneficial or neutral microorganisms present on grapevine leaves. In contrast, the plant- and yeast-based treatments used in this study are generally considered more favorable to microbial communities. These formulations contain bioactive molecules with antifungal activity, including elicitors that can activate plant defense mechanisms. Thus, instead of acting directly on the microbiome, their effects are mainly related to the plant’s immune responses. The T6621 treatment presented a difference in bacterial composition (Figure 6) especially in relation to NE21 and C21 treatments, and it is possible to observe in NMDS plots its difference in relation to these two treatments (Supplementary Figure 6B), which is also confirmed by the ANOSIM and SIMPER (Supplementary Tables 11, 12). The influence of ‘treatment’ and ‘year’ on the bacterial community was also evident in the Krona charts (Supplementary Figure 9) and in the relative abundance graphs (Figure 6; Supplementary Figures 7, 8). As previously mentioned, Sphingomonas was the most abundant genus in 2020 and Pseudomonas in 2021 (Figure 6), and both genera are known for their roles in plant health and productivity. These findings suggest that bacterial community is more sensitive to the inter-annual variation than the fungal community which appears to be more resilient and does not modify its composition over the years. The decision to perform sampling over two consecutive years aimed to capture the effect of interannual environmental variability on the grapevine microbiome. Previous studies have shown that microbial community composition, abundance, and distribution, can be influenced by climate conditions, geographical location, host plant phenology, physical and chemical properties, phyllosphere or soil characteristics, vineyard management practices, and the grapevine cultivar (Azevedo-Silva et al., 2021; Castanera et al., 2024). In our study, we observed substantial differences in microbial composition, particularly in the control treatment (C), across the two years. These differences, evident at various taxonomic levels (Figures 3, 6; Supplementary Figures 3, 4, 7, 8), suggest that microbial communities may undergo reorganization in response to changing environmental conditions. Although the main focus of the study was not to track adaptation mechanisms, these results provide initial evidence that such adaptive ecological responses may occur. This aspect could be explored in more depth in future long-term studies.
Some studies also verified that alternative treatments to conventional fungicides can modulate the bacterial community on grapevine leaves without significantly altering the fungal community, which appears to be more resilient and mainly influenced by seasonality (Gobbi et al., 2020; Miliordos et al., 2021). These findings are consistent with the results of the present study, where bacterial communities responded more markedly to treatment and year variation, particularly under CT and T6621, while fungal communities showed greater stability across years. This reinforces the notion that eco-friendly treatments may shape the bacterial communities on the leaf surface without disrupting the natural fungal balance, a key factor when designing sustainable plant protection strategies.
In this study, Spearman correlation analysis between the relative abundances of bacterial and fungal genera was performed for each treatment (Supplementary Figure 10). In the NE treatment, a negative correlation was observed between the fungal genus Stemphylium and several bacterial genera (Supplementary Figure 10A). In the JKE treatment, the genus Tillietiopsis showed negative correlations with a significant number of bacterial genera (Supplementary Figure 10B). For the T66 treatment, Symmetrospora exhibited positive correlations with some bacteria genera, whereas Filobasidium showed negative correlations (Supplementary Figure 10C). In the case of the T90 treatment, the fungal genera Alternaria, Mycosphaerella, and Erysiphe showed predominantly positive correlations with multiple bacterial genera, while Taphrina, Ramularia, Neosetophoma, and Cladosporium showed mainly negative correlations with various bacterial genera (Supplementary Figure 10D). In contrast, only a few correlations were detected in the C and CT treatments, suggesting a more limited interaction between bacterial and fungal genera in these treatments (Supplementary Figure 10E, F).
4 Conclusion
Understanding microbial communities is crucial due to their role in plant protection and growth. The structure and dynamics of these communities are known to be influenced by environmental factors such as geography, grapevine cultivar, and climate. In this study, we demonstrated that alternative treatments to synthetic fungicides could modulate the microbial composition of grapevine leaves, particularly the bacterial community. These shifts may contribute to enhanced disease control and increased plant resilience, highlighting the potential of sustainable strategies in viticulture.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
EM: Formal Analysis, Writing – original draft, Writing – review & editing, Investigation. PB: Writing – review & editing, Formal Analysis, Investigation. SS: Investigation, Formal Analysis, Writing – review & editing. MC: Writing – review & editing, Investigation. RB: Investigation, Writing – review & editing. KG: Investigation, Writing – review & editing. NS: Investigation, Writing – review & editing. ICo: Conceptualization, Supervision, Writing – review & editing. BG: Supervision, Writing – review & editing, Conceptualization. ICa: Conceptualization, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by the European Regional Development Fund through the INTERREG Atlantic Area Program, under the NASPA-natural fungicides against air and soil borne pathogens in the Atlantic Area project (EAPA-451/2016) and by the National Funds by FCT -Portuguese Foundation for Science and Technology, under the projects UID/04033/2023: Centre for the Research and Technology of Agro-Environmental and Biological Sciences and LA/P/0126/2020 (https://doi.org/10.54499/LA/P/0126/2020).
Acknowledgments
The author Eliana Monteiro acknowledges the financial support provided by the Portuguese Foundation for Science and Technology (FCT) (PD/BD/150261/2019) under the Doctoral Program “Agricultural Production Chains—from fork to farm” (PD/00122/2012). The authors gratefully acknowledge the support of Miguel Baltazar, Sandra Pereira, and Sofia Correia in leaves sampling, whose dedication and collaboration greatly contributed to this research.
Conflict of interest
KG and NS are scientific researchers employed by BioAtlantis Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1637143/full#supplementary-material
References
ADVID (2021). Boletim final do ano vitícola 2021 (Vila Real, Portugal: ADVID). Available online at: www.advid.pt.
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Azevedo-Silva, D., Rasmussen, J. A., Carneiro, M., and Azevedo, H. (2021). Feasibility of applying shotgun metagenomic analyses to grapevine leaf, rhizosphere and soil microbiome characterisation. Aust. J. ofGrape Wine Res. 47, 519–526. doi: 10.1111/ajgw.12508
Belhadj, A., Saigne, C., Telef, N., Cluzet, S., Bouscaut, J., Corio-Costet, M. F., et al. (2006). Methyl jasmonate induces defense responses in grapevine and triggers protection against Erysiphe necator. J. Agric. Food Chem. 54, 9119–9125. doi: 10.1021/jf0618022
Borovaya, S., Lukyanchuk, L., Manyakhin, A., and Zorikova, O. (2020). Effect of Reynoutria japonica extract upon germination and upon resistance of its seeds against phytopathogenic fungi Triticum aestivum L., Hordeum vulgare L., and Glycine max (L.). Merr. Org. Agric. 10, 89–95. doi: 10.1007/s13165-019-00254-6
Bulgari, D., Casati, P., Quaglino, F., and Bianco, P. A. (2014). Endophytic bacterial community of grapevine leaves influenced by sampling date and phytoplasma infection process. BMC Microbiol. 14, 1–11. doi: 10.1186/1471-2180-14-198
Cappelletti, M., Perazzolli, M., Antonielli, L., Nesler, A., Torboli, E., Bianchedi, P. L., et al. (2016). Leaf treatments with a protein-based resistance inducer partially modify phyllosphere microbial communities of grapevine. Front. Plant Sci. 7. doi: 10.3389/fpls.2016.01053
Castanera, R., González-Miguel, V. M., Escolà, G., Olivé, M., Teixidó, N., Savé, R., et al. (2024). Cultivar-dependent variations in the microbiome of grapevine leaves. Horticulturae 10. doi: 10.3390/horticulturae10121333
Clarke, K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
Colla, G., Hoagland, L., Ruzzi, M., Cardarelli, M., Bonini, P., Canaguier, R., et al. (2017). Biostimulant action of protein hydrolysates: Unraveling their effects on plant physiology and microbiome. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02202
Elad, Y. and Pertot, I. (2014). Climate change impacts on plant pathogens and plant diseases. Crop Improv. 28, 37–41. doi: 10.1080/15427528.2014.865412
Fadiji, A. E. and Babalola, O. O. (2020). Metagenomics methods for the study of plant-associated microbial communities: A review. J. Microbiol. Methods 170, 105860. doi: 10.1016/j.mimet.2020.105860
Fraga, H., De Cortázar Atauri, I. G., Malheiro, A. C., Moutinho-Pereira, J., and Santos, J. A. (2017). Viticulture in Portugal: A review of recent trends and climate change projections. OENOOne 51, 61–69. doi: 10.20870/oeno-one.2016.0.0.1621
Garde-Cerdán, T., Mancini, V., Carrasco-Quiroz, M., Servili, A., Gutiérrez-Gamboa, G., Foglia, R., et al. (2017). Chitosan and Laminarin as alternatives to copper for Plasmopara viticola control: Effect on grape amino acid. J. Agric. Food Chem. 65, 7379–7386. doi: 10.1021/acs.jafc.7b02352
Ghazal, H. N., Al-Shahwany, A. W., and Al-Dulaimy, F. T. (2019). Control of gray mold on tomato plants by spraying Piper nigrum and Urtica dioica extracts under greenhouse condition. Iraqi J. Sci. 60, 961–971. doi: 10.24996/ijs.2019.60.5.4
Gobbi, A., Kyrkou, I., Filippi, E., Ellegaard-jensen, L., and Hansen, L. H. (2020). Seasonal epiphytic microbial dynamics on grapevine leaves under biocontrol and copper fungicide treatments. Sci. Rep. 10, 1–13. doi: 10.1038/s41598-019-56741-z
Hammer, Ø., Harper, D. A., and Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4 (1), 9. Available onlline at: http://palaeo-electronica.org/2001_1/past/issue1_01.htm.
Janisiewicz, W. J., Peterson, D. L., and Bors, R. (1994). Control of storage decay of apples with sporobolomyces roseus. Plant Dis. doi: 10.1094/PD-78-0466
Joshi, B. C., Mukhija, M., and Kalia, A. N. (2014). Pharmacognostical review of Urtica dioica L. Int. J. Green Pharm. 8, 201. doi: 10.4103/0973-8258.142669
Karácsony, Z., Mondello, V., Fontaine, F., and Váczy, K. Z. (2023). The potential role of Aureobasidium pullulans in the development. Oeno One 57, 189–203. doi: 10.20870/oeno-one.2023.57.3.7463
Karthick, M., Kamalakannan, A., Malathi, V. G., Paranidharan, V., Sivakumar, U., Kavino, M., et al. (2019). Phenotypic characterization and molecular phylogenetic relationship of Erysiphe necator infecting grapes (Vitis vinifera). Curr. J. Appl. Sci. Technol. 37, 1–10. doi: 10.9734/cjast/2019/v37i330291
Koledenkova, K., Esmaeel, Q., Jacquard, C., Nowak, J., Clément, C., and Ait Barka, E. (2022). Plasmopara viticola the causal agent of downy mildew of grapevine : From its taxonomy to disease management. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.889472
Kruskal, J. B. (1964). Nonmetric multidimensional scaling: A numerical method. Psychometrika 29, 115–129. doi: 10.1007/BF02289694
Leveau, J. H. J. and Tech, J. J. (2011). Grapevine microbiomics: Bacterial diversity on grape leaves and berries revealed by high-throughput sequence analysis of 16S rRNA amplicons. Acta Hortic. 905, 31–42. doi: 10.17660/ActaHortic.2011.905.2
Lorenz, D. H., Eichhorn, K. W., Bleiholder, H., Klose, R., Meier, U., and Weber, E. (1995). Growth stages of the grapevine: Phenological growth stages of the grapevine (Vitis vinifera L. ssp. vinifera)—Codes and descriptions according to the extended BBCH scale. Aust. J. Grape Wine Res. 1, 100–103. doi: 10.1111/j.1755-0238.1995.tb00085.x
Miliordos, D. E., Tsiknia, M., Kontoudakis, N., Dimopoulou, M., Bouyioukos, C., and Kotseridis, Y. (2021). Impact of application of abscisic acid, benzothiadiazole and chitosan on berry quality characteristics and plant associated microbial communities of Vitis vinifera l var. Mouhtaro plants. Sustain. 13. doi: 10.3390/su13115802
Monteiro, E., Baltazar, M., Pereira, S., Correia, S., Ferreira, H., Bragança, R., et al. (2024a). Foliar application of nettle and Japanese knotweed extracts on Vitis vinifera: impact on phenylpropanoid biosynthesis and antioxidant activity during veraison and harvest of cv. Touriga Franca. J. Sci. Food Agric. 104, 4260–4267. doi: 10.1002/jsfa.13310
Monteiro, E., Correia, S., Baltazar, M., Pereira, S., Ferreira, H., Bragança, R., et al. (2024b). Foliar application of nettle and Japanese knotweed extracts on Vitis vinifera: Consequences for plant physiology, biochemical parameters, and yield. Horticulturae 10. doi: 10.3390/horticulturae10121275
Monteiro, E., Gonçalves, B., Cortez, I., and Castro, I. (2022). The role of biostimulants as alleviators of biotic and abiotic stresses in grapevine: A review. Plants 11, 396. doi: 10.3390/plants11030396
Müller, D. B., Vogel, C., Bai, Y., and Vorholt, J. A. (2016). The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 50, 211–234. doi: 10.1146/annurev-genet-120215-034952
Niem, J. M., Billones-Baaijens, R., Stodart, B., and Savocchia, S. (2020). Diversity profiling of grapevine microbial endosphere and antagonistic potential of endophytic pseudomonas against grapevine trunk diseases. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.00477
Oleszek, M., Kowalska, I., and Oleszek, W. (2019). Phytochemicals in bioenergy crops. Phytochem. Rev. 18, 893–927. doi: 10.1007/s11101-019-09639-7
Ondov, B. D., Bergman, N. H., and Phillippy, A. M. (2013). “Krona: Interactive metagenomic visualization in aweb browser,” in Encyclopedia of metagenomics. Ed. Nelson, K. (New York, NY, USA: Springer).
Ortega, P., Salcedo, R., Sánchez, E., and Gil, E. (2023). Biopesticides as alternatives to reduce the use of copper in Spanish and Portuguese viticulture: Main trends in adoption. Eur. J. Agron. 151, 126996. doi: 10.1016/j.eja.2023.126996
Puccioni, S., Biselli, C., Perria, R., Zanella, G., and D’Arcangelo, M. E. M. (2025). Alternative effects yeast-based biostimulants against downy mildew in Vitis vinifera cv Cabernet Sauvignon. Horticulturae 11. doi: 10.3390/horticulturae11020203
Rodino, S., Butu, M., and Butu, A. (2018). Alternative antimicrobial formula for plant protection. Bull. USAMV Ser. Agric. 75, 32–37. doi: 10.15835/buasvmcn-agr:002017
Sun, Z. B., Song, H. J., Liu, Y. Q., Ren, Q., Wang, Q. Y., Li, X. F., et al. (2024). The potential of microorganisms for the control of grape downy mildew—A review. J. Fungi 10. doi: 10.3390/jof10100702
Verhagen, B. W. M., Trotel-Aziz, P., Couderchet, M., Höfte, M., and Aziz, A. (2010). Pseudomonas spp.-induced systemic resistance to Botrytis cinerea is associated with induction and priming of defence responses in grapevine. J. Exp. Bot. 61, 249–260. doi: 10.1093/jxb/erp295
Wei, Y. J., Wu, Y., Yan, Y. Z., Zou, W., Xue, J., Ma, W. R., et al. (2018). High-throughput sequencing of microbial community diversity in soil, grapes, leaves, grape juice and wine of grapevine from China. PloS One 13, 1–17. doi: 10.1371/journal.pone.0193097
ZarraonaIndia, I., Owens, S. M., Weisenhorn, P., West, K., Hampton-Marcell, J., Lax, S., et al. (2015). The soil microbiome influences grapevine-associated microbiota. MBio 6. doi: 10.1128/mBio.02527-14
Keywords: leaves microbiome, microbial community diversity, metabarcoding, plant extracts, Vitis vinifera L., yeast-based formulations
Citation: Monteiro E, Baptista P, Silva S, Carvalho M, Bragança R, Guinan KJ, Sujeeth N, Cortez I, Gonçalves B and Castro I (2025) Metagenomic analysis of the effects of plant- and yeast-based formulations on the grapevine leaf microbiome of cv. ‘Touriga Franca’. Front. Plant Sci. 16:1637143. doi: 10.3389/fpls.2025.1637143
Received: 28 May 2025; Accepted: 28 July 2025;
Published: 14 August 2025.
Edited by:
Lucia Vieira Hoffmann, Brazilian Agricultural Research Corporation (EMBRAPA), BrazilReviewed by:
Subramanian Babu, VIT University, IndiaJorge Maurício Mondego, Instituto Agronômico de Campinas (IAC), Brazil
Copyright © 2025 Monteiro, Baptista, Silva, Carvalho, Bragança, Guinan, Sujeeth, Cortez, Gonçalves and Castro. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isaura Castro, aWNhc3Ryb0B1dGFkLnB0