 Josué Saiz-Pérez
Josué Saiz-Pérez Carmen Fenoll
Carmen Fenoll Montaña Mena
Montaña Mena- Facultad de Ciencias Ambientales y Bioquímica, Universidad de Castilla-La Mancha, Toledo, Spain
Stomata comprise two guard cells that function as microscopic valves in the plant epidermis, connecting mesophyll interstices to the atmosphere. Stomata regulate gas exchange and evapotranspiration, directly impacting photosynthesis and leaf temperature regulation, and their function is thus crucial for plant adaptability and fitness. In Arabidopsis, stomatal development is primarily driven by three basic helix-loop-helix transcription factors: SPEECHLESS (SPCH), MUTE, and FAMA, and occurs within the broader context of leaf development. During leaf development, a characteristic division-to-differentiation transition zone, marked by the first cell cycle arrest front (1st AF), progresses from the apex to the base of the leaf blade. The repeated division of meristemoids (M), self-renewing cells of stomatal lineages, is not halted during 1st AF, requiring a second arrest front, which is associated with activity of the PEAPOD (PPD) proteins, PEAPOD1 (PPD1) and PEAPOD2 (PPD2), which form a transcriptional repressor complex that halts M stem cell-like activity; however, the relationship between PPDs and stomatal development has not been fully elucidated. Here, we review data on PPD-mediated regulation of light signaling and the cell cycle and the influence of these factors on stomatal development.
1 Introduction
Multicellular organisms are generated through a spectrum of timely cell fate decisions, comprising both division and differentiation, that occur within developmental windows, ensuring functional organ development in mature organisms. For example, the plant leaf is an organ generated by an intricate molecular interplay that determines its shape, size, symmetry, and dorsoventrality. Investigation of the specific genetic determinants influencing leaf blade development (Kierzkowski et al., 2019) has identified gradients of morphogens (Ten Tusscher, 2020), known as mobile growth factors (MGFs) that, in combination with two cell cycle arrest fronts, drive a switch from leaf cell proliferation to differentiation (White, 2006; Kazama et al., 2010). Leaf development involves timely transcriptional regulation of a plethora of master regulatory genes in overlapping domains, to establish a specific pre-determined pattern (Vanhaeren et al., 2014, 2015; Castelán-Muñoz et al., 2019; Qiu et al., 2024).
Stomata are microscopic valve structures in the leaf epidermis comprising two guard cells (GCs) arising from meristemoids (Ms), cells with stem cell-like activity that contribute to leaf development. The function of stomata is to dynamically connect plant mesophyll to the atmosphere, and is indispensable for plant viability (Dittberner et al., 2018; Driesen et al., 2020). The opening and closing of stomata, along with their abundance and distribution, are regulated by physiological and environmental cues that modify gas exchange and evapotranspiration, directly influencing photosynthesis and leaf temperature regulation (Doheny-Adams et al., 2012; de Marcos et al., 2015; Pérez-Bueno et al., 2022). In addition to stomatal dynamics and function, a network of stomatal development genes controls their abundance and distribution. In Arabidopsis thaliana (Arabidopsis) stomatal development occurs through serial stereotypical division-differentiation events involving well-established cell types that are broadly distributed throughout the leaf epidermis (Bergmann and Sack, 2007), and is primarily driven by three basic helix-loop-helix transcription factors: SPEECHLESS (SPCH) (MacAlister et al., 2007), MUTE (Pillitteri et al., 2007), and FAMA (Ohashi-Ito and Bergmann, 2006; Jordan et al., 2015). These transcription factors require heterodimerization with SCREAM/SCREAM2 (SCRMs) to regulate stomatal-related genes (Kanaoka et al., 2008), and alteration of these key regulators modifies cell number and organ size, underlining their relevance in leaf development (Pérez-Bueno et al., 2022). In addition, the stomatal gene network comprises several positive and negative regulators, whose modulation alters stomata quantity and distribution (Saiz-Pérez et al., 2024). In the context of leaf development, Ms stem cell-like activity directly impinge on stomatal abundance in the mature organ, and therefore on physiological status (Dow et al., 2014), resulting in an adaptative trait with a strong genetic basis, as revealed by the broad natural variability observed in Arabidopsis (Delgado et al., 2011; Dittberner et al., 2018; Delgado et al., 2019).
Stomata are generated by self-renewing activity of Ms, broadly dispersed stomatal precursor cells which retain the capacity to divide several times without changes in cell-type fate, and are pivotal to epidermal development (Smit and Bergmann, 2023). Among the main characteristics of Ms is asymmetric cell division (ACD). Ms are generated from a meristemoid mother cell (MMCs) through an entry ACD that generates a M and a stomatal lineage ground cell (SLGC). Amplifying divisions allow M self-renewal, while generating neighboring SLGCs which subsequently differentiate into pavement cells (PCs) (Shpak et al., 2005). Alternatively, spacing ACDs generate satellite Ms from SLGCs, maintaining a one-cell spacing rule, which ensures proper stomata function (Hara et al., 2007; Dow et al., 2014). This remarkable ACD capacity generates around 65% to 82% of all leaf epidermal cells (Geisler et al., 2000). Interestingly, amplifying M division is absent in monocot plants, due to a lack of M self-renewal capacity (Vatén and Bergmann, 2012).
In Arabidopsis, leaf growth involves the interconnection of proliferation and expansion of leaf primordia cells (Asl et al., 2011), commencing with a group of founder cells flanking the shoot apical meristem (SAM) (Reinhardt et al., 2000; Kalve et al., 2014), which undergo profuse division, directly increasing cell numbers to form the mature organ; this proliferation phase is restricted to a specific time frame. Upon reaching a threshold, active mitotic cells exit the cell cycle and start to expand, while some PCs continue to increase in size through endocycles (Breuer et al., 2010; Magyar et al., 2012). Hence, several cellular processes determine final leaf shape and size, as follows: initial number of founder cells derived from the SAM; cell division rate; timing of cell division span and cell expansion; and extent of M proliferation (Hepworth et al., 2018). Alteration of any of these elements will modify mature leaf size (Gonzalez et al., 2012).
A crucial element controlling the division to differentiation switch in leaf blades is the 1st cell cycle arrest front (1st AF), which progresses to the base of the leaf blade from the apical region via MGFs (Kazama et al., 2010; Andriankaja et al., 2012). In Arabidopsis, stomatal proliferation and commitment also follow a tip-to-base direction; however, 1st AF does not stop self-renewing capacity of Ms, which requires the action of a 2nd arrest front (2ndAF) that has been linked to PEAPOD (PPD) proteins. There are two PPD paralogs, PEAPOD1 (PPD1) and PEAPOD2 (PPD2), which halt stomatal lineage stem cell-like activity, causing a switch of Ms from proliferative to committed states (White, 2006).
The aim of this review was to integrate knowledge of PPDs functions, focusing on PPDs modulation of Ms self-renewal activity. We explore the relationship of PPDs with stomatal development, light-mediated regulation, and the cell cycle machinery, whilst placing them into molecular context.
2 Stomatal lineage meristemoids display stem cell-like activity after 1st AF
Progression of 1st AF in developing leaves causes transition from cell proliferation to differentiation, involving post-mitotic cell expansion directly related to maturation. There is dynamic interplay between 1st AF and MGF gradients, where maximum MGF concentrations occur at the leaf base, although AF exhibits different thresholds for MGFs depending on spatial coordinates, causing cell division to stop based on organ dynamics and developmental stage (Kazama et al., 2010). CYCB1;1 reporters, which mark a linear border of actively dividing cells, have been used as evidence of 1st AF progression in leaf primordia (Kazama et al., 2010; Baekelandt et al., 2018). In contrast to SAM and root apical meristem (RAM), which divide continuously, maintaining constant meristem size and constituent cell number (Miwa et al., 2009), the active dividing regions change dynamically in Arabidopsis leaves. Hence, unlike the constant activity of SAM and RAM, leaf blade 1st AF does not progress uniformly. During a specific period, 1st AF imposes a non-dividing zone in more distal regions of the leaf blade, while remaining unaltered close to blade base, generating a proliferation zone and consequent leaf morphogenesis; however, while 1st AF halts protodermal cells division, it does not restrict ACD activity of Ms. Prevention of stomatal lineage cell ACD activity requires a 2nd AF, driven by PPDs (Figure 1A), which also modify the shape of the 1st AF (Baekelandt et al., 2018).
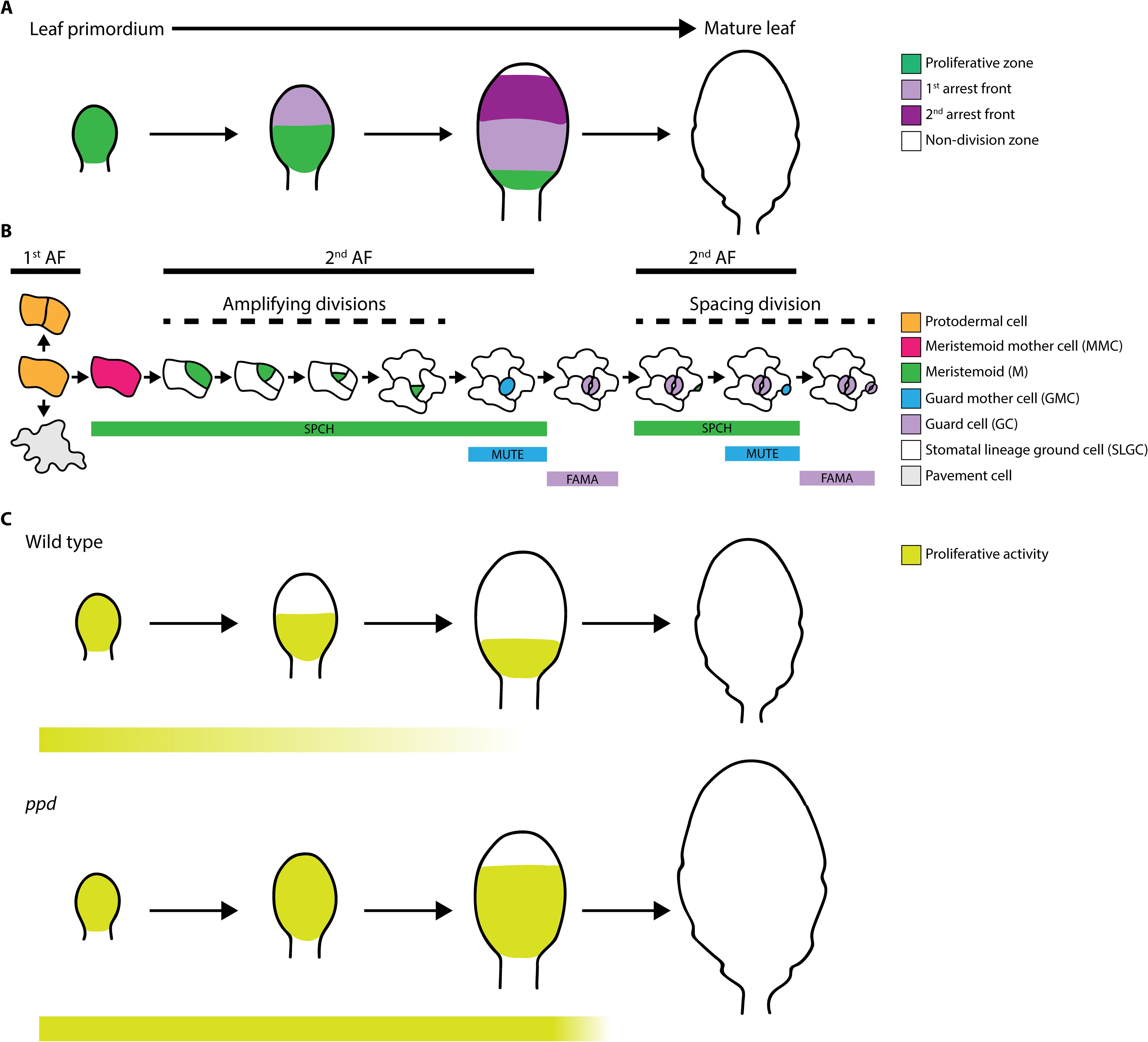
Figure 1. (A) Schematic overview of the proliferative zone, 1st cell cycle arrest front (1st arrest front), 2nd cell cycle arrest front (2nd arrest front), and non-division zone during development from leaf primordium to mature leaf in Arabidopsis. (B) Stomatal lineage cell types found in leaf epidermis: protodermal cell, meristemoid mother cell (MMC), meristemoid (M), guard mother cell (GMC) and guard cell (GC), stomatal lineage ground cell (SLGC) and pavement cell. Among them, only M exhibit proliferative activity. Cell types where 1st arrest front (1st AF) and 2nd arrest front (2nd AF) act are depicted with bold lines. Amplifying and spacing divisions in the stomatal lineages are indicated with dashed lines. (C) Comparison of proliferative activity zones during leaf development in wild type (above) and ppd (below) mutant plants. Extended M proliferative activity in ppd mutants is depicted in pale green bars below. Note that ppd mutation results in bigger leaves.
3 Proliferative capacity of meristemoids
Stomatal lineages begin with acquisition of meristemoid mother cell (MMC) identity by a protodermal cell via SPCH expression. SPCH activity triggers an entry ACD, generating a M retaining the ability to divide asymmetrically through reiterative amplifying divisions (Figure 1B), a process known as the proliferative stage of stomatal lineage, which relies on M self-renewal capacity (Han et al., 2022); whereas in SLGCs, SPCH activity causes spacing divisions generating satellite Ms (Figure 1B). Ms can go through up to three amplifying ACDs before differentiating into guard mother cells (GMCs) via activity of MUTE and its downstream genes, which orchestrate a symmetrical cell division to generate the GCs comprising mature functional stomata (Kim and Torii, 2024). Subsequently, FAMA prevents further GC divisions (Hachez et al., 2011). Unlike Ms, GMCs and GCs do not exhibit proliferative activity; nevertheless, SPCH activity is not restricted to M stages, and overlaps with MUTE expression domains (Lopez-Anido et al., 2021).
The prolonged stage of proliferative activity involving meristemoid ACDs is a key mechanism regulating leaf development. To date, the only molecules associated with this predefined state controlling ACDs are PPDs. Deletion of genes encoding PPDs (i.e., ppd mutants) results in larger leaves, due to PPDs role in repressing cell proliferation across leaf tissues, including mesophyll cells and stomatal lineage cells (Figure 1C; White, 2006). Similarly, artificial microRNA targeting genes encoding both PPDs (i.e., ami-ppd) led to similar phenotypes (Gonzalez et al., 2015). Nevertheless, although transcripts encoding SPCH, MUTE, TMM, and POLAR are up-regulated in ppd mutants, none of these genes are direct targets of PPD2 (Gonzalez et al., 2015), indicating that PPDs indirectly modulate stomatal development.
4 PEAPOD complex function in stomatal development
PPD1 and PPD2 proteins share 84% identity with the TIFY transcription factor protein family (Pérez et al., 2014) and are involved in diverse protein-protein interactions, mediated by their ZIM, JAZ, and PPD-specific domains (Vanholme et al., 2007; Chini et al., 2009; Pauwels et al., 2010; Pauwels and Goossens, 2011). PPDs require other protein adaptors to form a transcriptional repressor complex, which is widely conserved among plants, other than grasses (Schneider et al., 2021, 2024).
In stomatal development, PPDs interact with KINASE-INDUCIBLE DOMAIN INTERACTING8/9 (KIX8/9) proteins to repress M ACDs (Gonzalez et al., 2015). The phenotypes of ami-ppd, ppd2, and kix8kix9 mutants include extended M ACDs, dome-shaped leaves, and increased leaf size (Baekelandt et al., 2018; Li et al., 2018). Similar to PPDs, KIX8/9 are present in the vast majority of plants, with the exception of the Poaceae family (Gonzalez et al., 2015). PPD-KIX8/9 complex stability is regulated by 26S proteasome-dependent degradation controlled by STERILE APETALA (SAP/SOD3), an F-box protein that forms part of SKP1/Cullin/F-box E3 ubiquitin ligase complex, targeting KIX-PPD (Li et al., 2018). SAP physically associates with PPDs and KIX8/9. Consequently, up- or down-regulation of SAP alters M ACD and organ size (Wang et al., 2016; Li et al., 2018).
PPD2 and KIX8 have dominant roles in M ACD regulation. The phenotypes of ppd1 and kix9 mutants do not differ in cotyledon area to those of wild type, whereas kix8 and ppd2 mutants have increased areas. Further, these differences are enhanced in kix8kix9 and ppd1ppd2 double mutants, and even more pronounced in the quadruple mutant, kix8kix9ppd1ppd2; suggesting that PPD1 and KIX9 can modulate epidermal development, but require PPD2 and KIX8 to exert their effects (Liu et al., 2020).
PPDs expression and function are not restricted to stomatal lineage cells and leaf tissue, both are widely expressed and also modulate development of root, stem, inflorescence, flower, silique, and seed (Zhu et al., 2020). Hence, the complex functional plasticity of PPDs depends on the distinct molecular contexts imposed by tissue-dependent microenvironments.
5 PEAPOD integration of light regulation, cell cycle and stomatal development
PPDs are involved in light signaling, a process also linked to control of stomatal development, revealing an interplay among PPDs, stomatal genes, and key regulators of the light signaling network. Light is composed of distinct wavelengths, including the red and blue spectra, perceived by diverse plant photoreceptors. Red/far-red (FR) wavelengths are perceived by phytochromes (PHYs) (Chen and Chory, 2011), whereas blue/UV-A are sensed by cryptochromes (CRYs) (Cashmore et al., 1999). Both PHY and CRY light-receptors regulate photomorphogenesis via complex regulatory mechanisms (Jiao et al., 2007). In Arabidopsis, the PHY gene family comprises five members (PHYA to PHYE), where PHYA/B are the most prominent regulators of growth and development (Franklin and Quail, 2010). PHYA has a major role in FR perception and dark transition, whereas PHYB is important in red light detection, as it is stable under these conditions (Clough and Vierstra, 1997). The CRY gene family includes two receptors, CRY1 and CRY2, which regulate several developmental processes, including hypocotyl elongation, flowering time and stomatal development (Guo et al., 1998; Cao et al., 2021). The PHYTOCHROME-INTERACTING FACTOR (PIF) family comprises eight PIFs (PIF1–8) with distinct roles in development and modulation of light signaling, which connect both PHY and CRY light receptors (Kim et al., 2024). Amongst PIFs, PIF4 is a predominant factor that controls both light signaling and thermomorphogenesis (Xu and Zhu, 2021), with light-receptor and transduction signaling pathways partially converging on PIF4. Physical interaction between PIF4 and the active form of PHYB causes PIF4 ubiquitylation and subsequent degradation by the 26S proteasome (Figure 2A; Xu et al., 2015). CRY1 regulates PIF4 activity under blue light and warm-temperature, repressing its binding activity to target gene cis-regulatory elements (Ma et al., 2016; Pedmale et al., 2016).
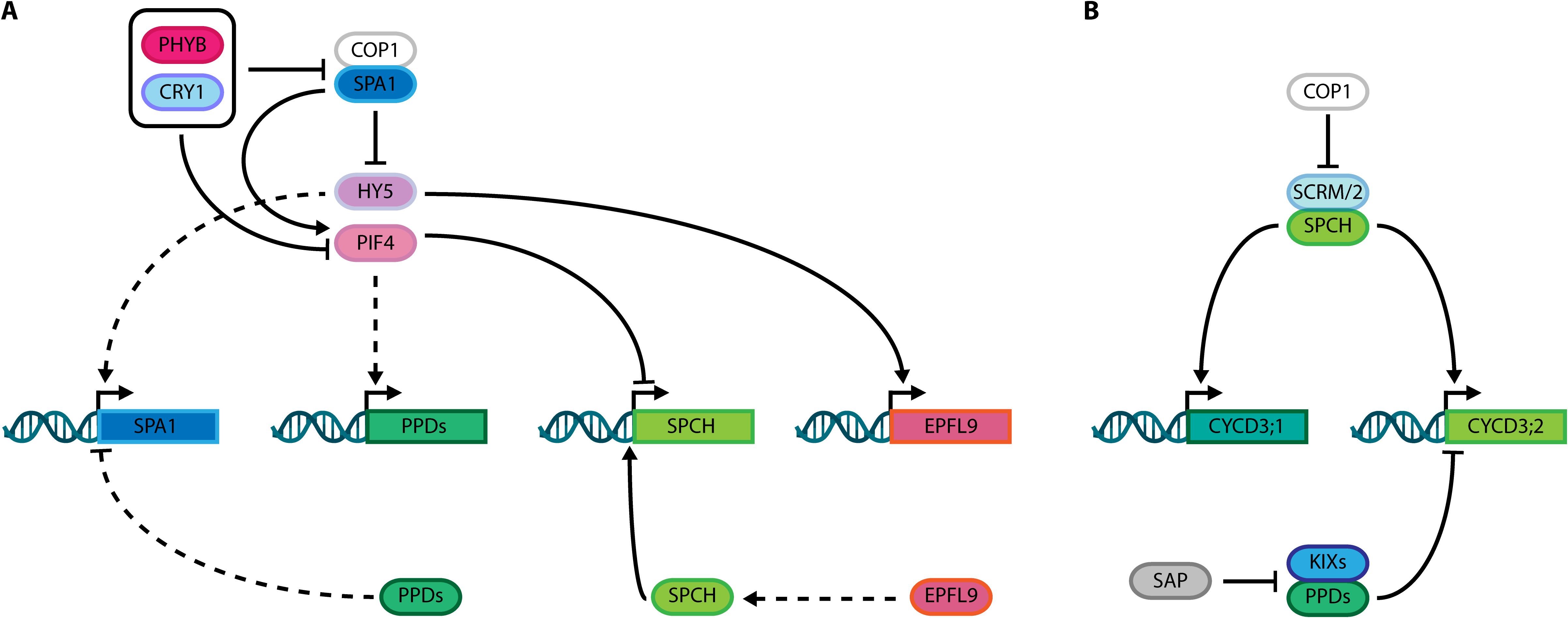
Figure 2. (A) Integration of PEAPODs (PPD) function with the control of light signaling networks comprising PHYB, CRY1, COP1, SPA1, HY5 and PIF4; and stomatal development SPCH-EPFL9 module. (B) Schematic overview showing interconnection of PPD complex, and its direct regulator SAP, with SPCH, SCRM/2 and the cell cycle machinery through their known targets, CYCD3;1 and CYCD3;2. Dashed lines: indirect activation/stabilization or repression/degradation. Solid lines: direct activation/stabilization or repression/degradation.
Regarding the role of PHYs in stomatal development, only PHYB is reported to modify stomatal development via PIF4 under fluctuating light intensity (Casson et al., 2009; Casson and Hetherington, 2014). Further, PIF4 directly represses SPCH transcription under supra-optimal temperature conditions (Lau et al., 2018). Among CRYs, CRY1 promotes stomatal development by blue light-dependent physical interaction with SPCH, which enhances its DNA-binding activity (Cao et al., 2021; Chen et al., 2025).
Upon light perception, PHYs and CRYs inactivate two classes of repressors that act in Arabidopsis light signaling and stomatal developmental networks, including complexes comprised of CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1) and SUPPRESSOR OF PHYTOCHROME A-105 (SPA) family proteins. The SPA gene family includes four genes (SPA1–4) that positively control COP1 E3 ubiquitin ligase activity (Laubinger et al., 2004; Hoecker, 2017). These molecules constitute an important hub connecting light signaling, stomatal development, and PPD gene networks. SPA1 phosphorylates and stabilizes PIF4 (Lee et al., 2020), whereas PPDs repress SPA1 transcription (White, 2022). Additionally, light intensity mediates indirect transcriptional activation of PPDs by PIF4 (White, 2022), while COP1 degrades SCRM/2 in a light-dependent manner, thereby modulating stomatal development (Kang et al., 2009; Lee et al., 2017). In parallel, ELONGATED HYPOCOTIL5 (HY5), a bZIP transcription factor whose accumulation promotes photomorphogenesis, is degraded in a light-dependent manner via the COP1/SPA1 complex (Saijo et al., 2003; Wang et al., 2021b). Strikingly, whereas HY5 indirectly activates SPA1, PPDs repress SPA1 transcription, connecting these two pathways (Figure 2A). Furthermore, HY5 directly binds and controls the expression of stomatal development genes in a light-dependent way, by modulating paracrine signaling mediated by EPIDERMAL PATTERNING FACTOR LIKE9 (EPFL9/STOMAGEN), a peptide that stabilizes SPCH, which self-regulates its expression (Hunt et al., 2010; Lau et al., 2014; Wang et al., 2021a). These roles of PPDs in modulation of light signaling suggest an indirect mechanism for controlling stomatal number in response to light intensity and might indicate an effect on stomatal fate acquisition by halting M ACD in a light-dependent manner. Nonetheless, PPD regulation of stomatal development is not solely related to the crosstalk between light signaling and stomatal gene network.
The cell cycle machinery is also directly linked to cell divisions in stomatal lineages, controlling timing and cell phase states during M-GMC-GC differentiation (Desvoyes and Gutierrez, 2020; Han et al., 2022; Zuch et al., 2023; Han and Torii, 2019; Xie et al., 2010). The CYCLIN (CYC) D gene family is among regulators promoting cell division, and includes the D-3 type Cyclin (CYCD3) genes. The three Arabidopsis CYCD3 proteins are: CYCLIN D3;1 (CYCD3;1), CYCLIN D3;2 (CYCD3;2), and CYCLIN D3;3 (CYCD3;3), which exhibit different expression patterns, but all promote cell division in Arabidopsis tissues (Menges et al., 2006), and influence cell quantity in leaves, as demonstrated by the reduced cell numbers in the triple cycD3;1–3 mutant (Dewitte et al., 2007). Together with KIX8/9, PPD2 directly represses CYCD3;2 and CYCD3;3 transcription (Gonzalez et al., 2015), while SPCH up-regulates CYCD3;1 and CYCD3;2 (Figure 2B), whose transcripts accumulate in Ms at early stages of lineage development and are associated with proliferative stages of stomatal lineages (Adrian et al., 2015; Vatén et al., 2018). Notably, CYCD3;2 is the only D-3 type Cyclin both directly bound and up-regulated by SPCH, and also found to be upregulated in the ami-ppd RNAseq dataset (Lau et al., 2014; Gonzalez et al., 2015). Further, CYCD3;2 overexpression phenocopies the dome-shaped leaves observed in ppd, ami-ppd, and ppd2 mutants, without altering leaf size. Conversely, CYCD3;1 and CYCD3;2 inactivation partially restores the ami-ppd phenotype (Baekelandt et al., 2018). Moreover, as repression of SPA1 transcription by PPDs influences the HY5-EPFL9 module, PPDs function might be affecting SPCH activity indirectly. This could putatively represent a mechanism of indirect modulation of stomatal development by light signaling via PPD-mediated cell cycle control.
6 Conclusions and perspectives
In summary, the PPD complex acts as a molecular hub, integrating both light signaling and CYCD3-mediated cell cycle control, while restricting M self-renewing activity. Hence, stomatal phenotypes of PPD complex mutants may be partially explained by the crosstalk among these distinct hubs, although a direct connection between PPDs and stomatal development remains to be established. Additionally, the role of PPDs and their adaptor proteins KIX8/9 in limiting the M ACDs requires additional investigation, as ppd stomatal phenotypes have not been mechanistically explained. Further studies are also required to explore potential new roles of the PPD complex in hormonal and environmental regulation of stomatal development. Given the importance of stomatal development in leaf morphogenesis and the regulatory role of the PPD complex in organ growth, deepening our knowledge in this area may be instrumental for improving crop productivity through translational approaches.
Author contributions
JSP: Writing – original draft, Conceptualization, Writing – review & editing. CF: Funding acquisition, Supervision, Resources, Writing – review & editing. MM: Writing – review & editing, Funding acquisition, Resources, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by grants from the Spanish Government (PID2019-105362RB-I00 and PID2022-137606NB-I00) and the Castilla-La Mancha Government (SBPLY/21/180225/000058) to MM and CF. The laboratory received support in the form of UCLM intramural grant (2022-GRIN-34125) and EU FEDER funds. JSP received an exchange grant from EMBO.
Acknowledgments
We thank Ana Rapp for technical support and laboratory management.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adrian, J., Chang, J., Ballenger, C. E., Bargmann, B. O. R., Alassimone, J., Davies, K. A., et al. (2015). Transcriptome dynamics of the stomatal lineage: birth, amplification, and termination of a self-renewing population. Dev. Cell 33, 107–118. doi: 10.1016/j.devcel.2015.01.025
Andriankaja, M., Dhondt, S., De Bodt, S., Vanhaeren, H., Coppens, F., De Milde, L., et al. (2012). Exit from Proliferation during Leaf Development in Arabidopsis thaliana: A Not-So-Gradual Process. Dev. Cell 22, 64–78. doi: 10.1016/j.devcel.2011.11.011
Asl, L. K., Dhondt, S., Boudolf, V., Beemster, G. T. S., Beeckman, T., Inzé, D., et al. (2011). Model-based analysis of Arabidopsis leaf epidermal cells reveals distinct division and expansion patterns for pavement and guard cells. Plant Physiol. 156, 2172–2183. doi: 10.1104/pp.111.181180
Baekelandt, A., Pauwels, L., Wang, Z., Li, N., De Milde, L., Natran, A., et al. (2018). Arabidopsis leaf flatness is regulated by PPD2 and NINJA through repression of CYCLIN D3 genes. Plant Physiol. 178, 217–232. doi: 10.1104/pp.18.00327
Bergmann, D. C. and Sack, F. D. (2007). Stomatal development. Annu. Rev. Plant Biol. 58, 163–181. doi: 10.1146/annurev.arplant.58.032806.104023
Breuer, C., Ishida, T., and Sugimoto, K. (2010). Developmental control of endocycles and cell growth in plants. Curr. Opin. Plant Biol. 13, 654–660. doi: 10.1016/j.pbi.2010.10.006
Cao, X., Xu, P., Liu, Y., Yang, G., Liu, M., Chen, L., et al. (2021). Arabidopsis cryptochrome 1 promotes stomatal development through repression of AGB1 inhibition of SPEECHLESS DNA-binding activity. J. Integr. Plant Biol. 63, 1967–1981. doi: 10.1111/jipb.13168
Cashmore, A. R., Jarillo, J. A., Wu, Y.-J., and Liu, D. (1999). Cryptochromes: blue light receptors for plants and animals. Science 284, 760–765. doi: 10.1126/science.284.5415.760
Casson, S. A., Franklin, K. A., Gray, J. E., Grierson, C. S., Whitelam, G. C., and Hetherington, A. M. (2009). phytochrome B and PIF4 regulate stomatal development in response to light quantity. Curr. Biol. CB 19, 229–234. doi: 10.1016/j.cub.2008.12.046
Casson, S. A. and Hetherington, A. M. (2014). phytochrome B is required for light-mediated systemic control of stomatal development. Curr. Biol. 24, 1216–1221. doi: 10.1016/j.cub.2014.03.074
Castelán-Muñoz, N., Herrera, J., Cajero-Sánchez, W., Arrizubieta, M., Trejo, C., García-Ponce, B., et al. (2019). MADS-box genes are key components of genetic regulatory networks involved in abiotic stress and plastic developmental responses in plants. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00853
Chen, L., Cao, X., Li, Y., Liu, M., Liu, Y., Guan, Y., et al. (2025). Photoexcited cryptochrome 1 interacts with SPCHLESS to regulate stomatal development in Arabidopsis. Plant Cell Environ. 48, 286–296. doi: 10.1111/pce.15123
Chen, M. and Chory, J. (2011). Phytochrome signaling mechanisms and the control of plant development. Trends Cell Biol. 21, 664–671. doi: 10.1016/j.tcb.2011.07.002
Chini, A., Fonseca, S., Chico, J. M., Fernández-Calvo, P., and Solano, R. (2009). The ZIM domain mediates homo- and heteromeric interactions between Arabidopsis JAZ proteins. Plant J. Cell Mol. Biol. 59, 77–87. doi: 10.1111/j.1365-313X.2009.03852.x
Clough, R. C. and Vierstra, R. D. (1997). Phytochrome degradation. Plant Cell Environ. 20, 713–721. doi: 10.1046/j.1365-3040.1997.d01-107.x
Delgado, D., Alonso-Blanco, C., Fenoll, C., and Mena, M. (2011). Natural variation in stomatal abundance of Arabidopsis thaliana includes cryptic diversity for different developmental processes. Ann. Bot. 107, 1247–1258. doi: 10.1093/aob/mcr060
Delgado, D., Sánchez-Bermejo, E., de Marcos, A., Martín-Jimenez, C., Fenoll, C., Alonso-Blanco, C., et al. (2019). A genetic dissection of natural variation for stomatal abundance traits in Arabidopsis. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01392
de Marcos, A., Triviño, M., Pérez-Bueno, M. L., Ballesteros, I., Barón, M., Mena, M., et al. (2015). Transcriptional profiles of Arabidopsis stomataless mutants reveal developmental and physiological features of life in the absence of stomata. Front. Plant Sci. 6, 456. doi: 10.3389/fpls.2015.00456
Desvoyes, B. and Gutierrez, C. (2020). Roles of plant retinoblastoma protein: cell cycle and beyond. EMBO J. 39, e105802. doi: 10.15252/embj.2020105802
Dewitte, W., Scofield, S., Alcasabas, A. A., Maughan, S. C., Menges, M., Braun, N., et al. (2007). Arabidopsis CYCD3 D-type cyclins link cell proliferation and endocycles and are rate-limiting for cytokinin responses. Proc. Natl. Acad. Sci. 104, 14537–14542. doi: 10.1073/pnas.0704166104
Dittberner, H., Korte, A., Mettler-Altmann, T., Weber, A. P. M., Monroe, G., and de Meaux, J. (2018). Natural variation in stomata size contributes to the local adaptation of water-use efficiency in Arabidopsis thaliana. Mol. Ecol. 27, 4052–4065. doi: 10.1111/mec.14838
Doheny-Adams, T., Hunt, L., Franks, P. J., Beerling, D. J., and Gray, J. E. (2012). Genetic manipulation of stomatal density influences stomatal size, plant growth and tolerance to restricted water supply across a growth carbon dioxide gradient. Philos. Trans. R. Soc B Biol. Sci. 367, 547–555. doi: 10.1098/rstb.2011.0272
Dow, G. J., Berry, J. A., and Bergmann, D. C. (2014). The physiological importance of developmental mechanisms that enforce proper stomatal spacing in Arabidopsis thaliana. New Phytol. 201, 1205–1217. doi: 10.1111/nph.12586
Driesen, E., Van den Ende, W., De Proft, M., and Saeys, W. (2020). Influence of environmental factors light, CO2, temperature, and relative humidity on stomatal opening and development: A review. Agronomy 10, 1975. doi: 10.3390/agronomy10121975
Franklin, K. A. and Quail, P. H. (2010). Phytochrome functions in Arabidopsis development. J. Exp. Bot. 61, 11–24. doi: 10.1093/jxb/erp304
Geisler, M., Nadeau, J., and Sack, F. D. (2000). Oriented Asymmetric Divisions That Generate the Stomatal Spacing Pattern in Arabidopsis Are Disrupted by the too many mouths Mutation. Plant Cell 12, 2075–2086.
Gonzalez, N., Pauwels, L., Baekelandt, A., De Milde, L., Van Leene, J., Besbrugge, N., et al. (2015). A repressor protein complex regulates leaf growth in Arabidopsis. Plant Cell 27, 2273–2287. doi: 10.1105/tpc.15.00006
Gonzalez, N., Vanhaeren, H., and Inzé, D. (2012). Leaf size control: complex coordination of cell division and expansion. Trends Plant Sci. 17, 332–340. doi: 10.1016/j.tplants.2012.02.003
Guo, H., Yang, H., Mockler, T. C., and Lin, C. (1998). Regulation of flowering time by Arabidopsis photoreceptors. Science 279, 1360–1363. doi: 10.1126/science.279.5355.1360
Hachez, C., Ohashi-Ito, K., Dong, J., and Bergmann, D. C. (2011). Differentiation of arabidopsis guard cells: analysis of the networks incorporating the basic helix-loop-helix transcription factor, FAMA. Plant Physiol. 155, 1458–1472. doi: 10.1104/pp.110.167718
Han, S.-K., Herrmann, A., Yang, J., Iwasaki, R., Sakamoto, T., Desvoyes, B., et al. (2022). Deceleration of the cell cycle underpins a switch from proliferative to terminal divisions in plant stomatal lineage. Dev. Cell 57, 569–582.e6. doi: 10.1016/j.devcel.2022.01.014
Han, S.-K. and Torii, K. U. (2019). Linking cell cycle to stomatal differentiation. Curr. Opin. Plant Biol. 51, 66–73. doi: 10.1016/j.pbi.2019.03.010
Hara, K., Kajita, R., Torii, K. U., Bergmann, D. C., and Kakimoto, T. (2007). The secretory peptide gene EPF1 enforces the stomatal one-cell-spacing rule. Genes Dev. 21, 1720–1725. doi: 10.1101/gad.1550707
Hepworth, C., Caine, R. S., Harrison, E. L., Sloan, J., and Gray, J. E. (2018). Stomatal development: focusing on the grasses. Curr. Opin. Plant Biol. 41, 1–7. doi: 10.1016/j.pbi.2017.07.009
Hoecker, U. (2017). The activities of the E3 ubiquitin ligase COP1/SPA, a key repressor in light signaling. Curr. Opin. Plant Biol. 37, 63–69. doi: 10.1016/j.pbi.2017.03.015
Hunt, L., Bailey, K. J., and Gray, J. E. (2010). The signalling peptide EPFL9 is a positive regulator of stomatal development. New Phytol. 186, 609–614. doi: 10.1111/j.1469-8137.2010.03200.x
Jiao, Y., Lau, O. S., and Deng, X. W. (2007). Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 8, 217–230. doi: 10.1038/nrg2049
Jordan, G. J., Carpenter, R. J., Koutoulis, A., Price, A., and Brodribb, T. J. (2015). Environmental adaptation in stomatal size independent of the effects of genome size. New Phytol. 205, 608–617. doi: 10.1111/nph.13076
Kalve, S., De Vos, D., and Beemster, G. T. S. (2014). Leaf development: a cellular perspective. Front. Plant Sci. 5. doi: 10.3389/fpls.2014.00362
Kanaoka, M. M., Pillitteri, L. J., Fujii, H., Yoshida, Y., Bogenschutz, N. L., Takabayashi, J., et al. (2008). SCREAM/ICE1 and SCREAM2 specify three cell-state transitional steps leading to Arabidopsis stomatal differentiation. Plant Cell 20, 1775–1785. doi: 10.1105/tpc.108.060848
Kang, C.-Y., Lian, H.-L., Wang, F.-F., Huang, J.-R., and Yang, H.-Q. (2009). Cryptochromes, phytochromes, and COP1 regulate light-controlled stomatal development in Arabidopsis. Plant Cell 21, 2624–2641. doi: 10.1105/tpc.109.069765
Kazama, T., Ichihashi, Y., Murata, S., and Tsukaya, H. (2010). The mechanism of cell cycle arrest front progression explained by a KLUH/CYP78A5-dependent mobile growth factor in developing leaves of Arabidopsis thaliana. Plant Cell Physiol. 51, 1046–1054. doi: 10.1093/pcp/pcq051
Kierzkowski, D., Runions, A., Vuolo, F., Strauss, S., Lymbouridou, R., Routier-Kierzkowska, A.-L., et al. (2019). A growth-based framework for leaf shape development and diversity. Cell 177, 1405–1418.e17. doi: 10.1016/j.cell.2019.05.011
Kim, H., Lee, N., Kim, Y., and Choi, G. (2024). The phytochrome-interacting factor genes PIF1 and PIF4 are functionally diversified due to divergence of promoters and proteins. Plant Cell 36, 2778–2797. doi: 10.1093/plcell/koae110
Kim, E.-D. and Torii, K. U. (2024). Stomatal cell fate commitment via transcriptional and epigenetic control: Timing is crucial. Plant Cell Environ. 47, 3288–3298. doi: 10.1111/pce.14761
Lau, O. S., Davies, K. A., Chang, J., Adrian, J., Rowe, M. H., Ballenger, C. E., et al. (2014). Direct roles of SPEECHLESS in the specification of stomatal self-renewing cells. Science 345, 1605–1609. doi: 10.1126/science.1256888
Lau, O. S., Song, Z., Zhou, Z., Davies, K. A., Chang, J., Yang, X., et al. (2018). Direct control of SPEECHLESS by PIF4 in the high-temperature response of stomatal development. Curr. Biol. 28, 1273–1280.e3. doi: 10.1016/j.cub.2018.02.054
Laubinger, S., Fittinghoff, K., and Hoecker, U. (2004). The SPA quartet: a family of WD-repeat proteins with a central role in suppression of photomorphogenesis in arabidopsis. Plant Cell 16, 2293–2306. doi: 10.1105/tpc.104.024216
Lee, J.-H., Jung, J.-H., and Park, C.-M. (2017). Light inhibits COP1-mediated degradation of ICE transcription factors to induce stomatal development in Arabidopsis. Plant Cell 29, 2817–2830. doi: 10.1105/tpc.17.00371
Lee, S., Paik, I., and Huq, E. (2020). SPAs promote thermomorphogenesis by regulating the phyB-PIF4 module in Arabidopsis. Development 147, dev189233. doi: 10.1242/dev.189233
Li, N., Liu, Z., Wang, Z., Ru, L., Gonzalez, N., Baekelandt, A., et al. (2018). STERILE APETALA modulates the stability of a repressor protein complex to control organ size in Arabidopsis thaliana. PloS Genet. 14, e1007218. doi: 10.1371/journal.pgen.1007218
Liu, Z., Li, N., Zhang, Y., and Li, Y. (2020). Transcriptional repression of GIF1 by the KIX-PPD-MYC repressor complex controls seed size in Arabidopsis. Nat. Commun. 11, 1846. doi: 10.1038/s41467-020-15603-3
Lopez-Anido, C. B., Vatén, A., Smoot, N. K., Sharma, N., Guo, V., Gong, Y., et al. (2021). Single-cell resolution of lineage trajectories in the Arabidopsis stomatal lineage and developing leaf. Dev. Cell 56, 1043–1055.e4. doi: 10.1016/j.devcel.2021.03.014
Ma, D., Li, X., Guo, Y., Chu, J., Fang, S., Yan, C., et al. (2016). Cryptochrome 1 interacts with PIF4 to regulate high temperature-mediated hypocotyl elongation in response to blue light. Proc. Natl. Acad. Sci. 113, 224–229. doi: 10.1073/pnas.1511437113
MacAlister, C. A., Ohashi-Ito, K., and Bergmann, D. C. (2007). Transcription factor control of asymmetric cell divisions that establish the stomatal lineage. Nature 445, 537–540. doi: 10.1038/nature05491
Magyar, Z., Horváth, B., Khan, S., Mohammed, B., Henriques, R., De Veylder, L., et al. (2012). Arabidopsis E2FA stimulates proliferation and endocycle separately through RBR-bound and RBR-free complexes. EMBO J. 31, 1480–1493. doi: 10.1038/emboj.2012.13
Menges, M., Samland, A. K., Planchais, S., and Murray, J. A. H. (2006). The D-type cyclin CYCD3;1 is limiting for the G1-to-S-phase transition in Arabidopsis. Plant Cell 18, 893–906. doi: 10.1105/tpc.105.039636
Miwa, H., Kinoshita, A., Fukuda, H., and Sawa, S. (2009). Plant meristems: CLAVATA3/ESR-related signaling in the shoot apical meristem and the root apical meristem. J. Plant Res. 122, 31–39. doi: 10.1007/s10265-008-0207-3
Ohashi-Ito, K. and Bergmann, D. C. (2006). Arabidopsis FAMA controls the final proliferation/differentiation switch during stomatal development. Plant Cell 18, 2493–2505. doi: 10.1105/tpc.106.046136
Pauwels, L., Barbero, G. F., Geerinck, J., Tilleman, S., Grunewald, W., Pérez, A. C., et al. (2010). NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 464, 788–791. doi: 10.1038/nature08854
Pauwels, L. and Goossens, A. (2011). The JAZ proteins: A crucial interface in the jasmonate signaling cascade. Plant Cell 23, 3089–3100. doi: 10.1105/tpc.111.089300
Pedmale, U. V., Huang, S. C., Zander, M., Cole, B. J., Hetzel, J., Ljung, K., et al. (2016). Cryptochromes interact directly with PIFs to control plant growth in limiting blue light. Cell 164, 233–245. doi: 10.1016/j.cell.2015.12.018
Pérez, A. C., Durand, A. N., Bossche, R. V., Clercq, R. D., Persiau, G., Wees, S. C. M. V., et al. (2014). The non-JAZ TIFY protein TIFY8 from Arabidopsis thaliana is a transcriptional repressor. PloS One 9, e84891. doi: 10.1371/journal.pone.0084891
Pérez-Bueno, M. L., Illescas-Miranda, J., Martín-Forero, A. F., de Marcos, A., Barón, M., Fenoll, C., et al. (2022). An extremely low stomatal density mutant overcomes cooling limitations at supra-optimal temperature by adjusting stomatal size and leaf thickness. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.919299
Pillitteri, L. J., Sloan, D. B., Bogenschutz, N. L., and Torii, K. U. (2007). Termination of asymmetric cell division and differentiation of stomata. Nature 445, 501–505. doi: 10.1038/nature05467
Qiu, Y., Li, Z., and Köhler, C. (2024). Ancestral duplication of MADS-box genes in land plants empowered the functional divergence between sporophytes and gametophytes. New Phytologist 244, 358–363. doi: 10.1111/nph.20065
Reinhardt, D., Mandel, T., and Kuhlemeier, C. (2000). Auxin regulates the initiation and radial position of plant lateral organs. Plant Cell 12, 507–518. doi: 10.1105/tpc.12.4.507
Saijo, Y., Sullivan, J. A., Wang, H., Yang, J., Shen, Y., Rubio, V., et al. (2003). The COP1–SPA1 interaction defines a critical step in phytochrome A-mediated regulation of HY5 activity. Genes Dev. 17, 2642–2647. doi: 10.1101/gad.1122903
Saiz-Pérez, J., Fenoll, C., and Mena, M. (2024). “Molecular genetics of stomatal development in Arabidopsis,” in Progress in Botany, vol. 85 . Eds. Lüttge, U., Cánovas, F. M., Risueño Almeida, M. C., Leuschner, C., and Pretzsch, H. (Springer Nature Switzerland, Cham), 105–138. doi: 10.1007/124_2024_82
Schneider, M., Gonzalez, N., Pauwels, L., Inzé, D., and Baekelandt, A. (2021). The PEAPOD pathway and its potential to improve crop yield. Trends Plant Sci. 26, 220–236. doi: 10.1016/j.tplants.2020.10.012
Schneider, M., Van Bel, M., Inzé, D., and Baekelandt, A. (2024). Leaf growth - complex regulation of a seemingly simple process. Plant J. Cell Mol. Biol. 117, 1018–1051. doi: 10.1111/tpj.16558
Shpak, E. D., McAbee, J. M., Pillitteri, L. J., and Torii, K. U. (2005). Stomatal patterning and differentiation by synergistic interactions of receptor kinases. Science 309, 290–293. doi: 10.1126/science.1109710
Smit, M. E. and Bergmann, D. C. (2023). The stomatal fates: Understanding initiation and enforcement of stomatal cell fate transitions. Curr. Opin. Plant Biol. 76, 102449. doi: 10.1016/j.pbi.2023.102449
Ten Tusscher, K. (2020). Of mice and plants: Comparative developmental systems biology. Dev. Biol. 460, 32–39. doi: 10.1016/j.ydbio.2018.10.024
Vanhaeren, H., Gonzalez, N., Coppens, F., De Milde, L., Van Daele, T., Vermeersch, M., et al. (2014). Combining growth-promoting genes leads to positive epistasis in Arabidopsis thaliana. eLife 3, e02252. doi: 10.7554/eLife.02252
Vanhaeren, H., Gonzalez, N., and Inzé, D. (2015). A journey through a leaf: phenomics analysis of leaf growth in Arabidopsis thaliana. Arab. Book 13, e0181. doi: 10.1199/tab.0181
Vanholme, B., Grunewald, W., Bateman, A., Kohchi, T., and Gheysen, G. (2007). The tify family previously known as ZIM. Trends Plant Sci. 12, 239–244. doi: 10.1016/j.tplants.2007.04.004
Vatén, A. and Bergmann, D. C. (2012). Mechanisms of stomatal development: an evolutionary view. EvoDevo 3, 11. doi: 10.1186/2041-9139-3-11
Vatén, A., Soyars, C. L., Tarr, P. T., Nimchuk, Z. L., and Bergmann, D. C. (2018). Modulation of asymmetric division diversity through cytokinin and SPEECHLESS regulatory interactions in the Arabidopsis stomatal lineage. Dev. Cell 47, 53–66.e5. doi: 10.1016/j.devcel.2018.08.007
Wang, Z., Li, N., Jiang, S., Gonzalez, N., Huang, X., Wang, Y., et al. (2016). SCFSAP controls organ size by targeting PPD proteins for degradation in Arabidopsis thaliana. Nat. Commun. 7, 11192. doi: 10.1038/ncomms11192
Wang, W., Paik, I., Kim, J., Hou, X., Sung, S., and Huq, E. (2021b). Direct phosphorylation of HY5 by SPA kinases to regulate photomorphogenesis in Arabidopsis. New Phytol. 230, 2311–2326. doi: 10.1111/nph.17332
Wang, S., Zhou, Z., Rahiman, R., Lee, G. S. Y., Yeo, Y. K., Yang, X., et al. (2021a). Light regulates stomatal development by modulating paracrine signaling from inner tissues. Nat. Commun. 12, 3403. doi: 10.1038/s41467-021-23728-2
White, D. W. R. (2006). PEAPOD regulates lamina size and curvature in Arabidopsis. Proc. Natl. Acad. Sci. 103, 13238–13243. doi: 10.1073/pnas.0604349103
White, D. W. R. (2022). PEAPOD repressors modulate and coordinate developmental responses to light intensity in Arabidopsis. New Phytol. 235, 1470–1485. doi: 10.1111/nph.18198
Xie, Z., Lee, E., Lucas, J. R., Morohashi, K., Li, D., Murray, J. A. H., et al. (2010). Regulation of cell proliferation in the stomatal lineage by the Arabidopsis MYB FOUR LIPS via direct targeting of core cell cycle genes. Plant Cell 22, 2306–2321. doi: 10.1105/tpc.110.074609
Xu, X., Paik, I., Zhu, L., and Huq, E. (2015). Illuminating progress in phytochrome-mediated light signaling pathways. Trends Plant Sci. 20, 641–650. doi: 10.1016/j.tplants.2015.06.010
Xu, Y. and Zhu, Z. (2021). PIF4 and PIF4-interacting proteins: at the nexus of plant light, temperature and hormone signal integrations. Int. J. Mol. Sci. 22, 10304. doi: 10.3390/ijms221910304
Zhu, Y., Luo, X., Liu, X., Wu, W., Cui, X., He, Y., et al. (2020). Arabidopsis PEAPODs function with LIKE HETEROCHROMATIN PROTEIN1 to regulate lateral organ growth. J. Integr. Plant Biol. 62, 812–831. doi: 10.1111/jipb.12841
Keywords: stomatal development, asymmetric cell division, PEAPOD, leaf development, light signaling, cell cycle
Citation: Saiz-Pérez J, Fenoll C and Mena M (2025) The PEAPOD repressor complex in Arabidopsis stomatal development. Front. Plant Sci. 16:1641102. doi: 10.3389/fpls.2025.1641102
Received: 04 June 2025; Accepted: 08 July 2025;
Published: 23 July 2025.
Edited by:
Stefan de Folter, National Polytechnic Institute of Mexico (CINVESTAV), MexicoReviewed by:
Shuka Ikematsu, Kyoto Sangyo University, JapanCopyright © 2025 Saiz-Pérez, Fenoll and Mena. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josué Saiz-Pérez, am9zdWUuc2FpenBlcmV6QHVjbG0uZXM=