 Lourdes Delgado-Aceves1
Lourdes Delgado-Aceves1 Santiago Corona2
Santiago Corona2 José Juvencio Castañeda-Nava1José Manuel Rodríguez-Domínguez1
José Juvencio Castañeda-Nava1José Manuel Rodríguez-Domínguez1 Antonia Gutiérrez-Mora1*
Antonia Gutiérrez-Mora1*- 1Unidad de Biotecnología Vegetal, Centro de Investigación y Asistencia en Tecnología y Diseño del Estado de Jalisco, Zapopan, Jalisco, Mexico
- 2Departamento de Botánica y Zoología, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, Zapopan, Jalisco, Mexico
Agave maximiliana Baker is a key species in the production of the traditional mexican beverage “Raicilla”. However, its slow reproductive cycle, lack of vegetative propagation, and increased commercial exploitation pose a threat to the conservation of its natural populations. This study presents, for the first time, a protocol for indirect somatic embryogenesis from zygotic embryos of A. maximiliana, as a biotechnological strategy for its mass propagation and conservation. Mature seeds were collected, and their viability was assessed using tetrazolium staining, revealing a significant loss of viability in seeds stored for more than two years. Embryonic axes were cultured on Murashige and Skoog medium supplemented with different concentrations of 2,4-D or picloram in combination with BAP. The highest embryogenic callus formation rates (up to 90%) were obtained with the treatment consisting of 36.24 µM 2,4-D, 33.12 µM picloram, and 4.44 µM BAP, resulting in an embryo-forming capacity (EFC) of 20%. Histological analyses confirmed the presence of proembryogenic cell masses and somatic embryos at different developmental stages, indicating an asynchronous regenerative system. The conversion rate of embryos into viable plantlets reached up to 90%, although some abnormalities were observed, associated with high auxin concentrations. This protocol represents a valuable tool for the conservation, reforestation, and sustainable production of this endemic and economically important species.
1 Introduction
Agave maximiliana Baker (commonly known as “Lechuguilla”) is a species of the Asparagaceae family with a geographic distribution in western Mexico. It primarily grows on rocky slopes within tropical deciduous oak and pine-oak forests, at elevations between 1500 and 2000 masl (Vázquez-García et al., 2007; Santacruz-Ruvalcaba et al., 2022). Due to its high sugar content and its traditional management practices, A. maximiliana is used in the production of the distilled beverage “Raicilla.” This beverage was recently granted a designation of appellation origin (Diario Oficial de la Federación, 2019), covering 16 municipalities in the state of Jalisco—Atengo, Chiquilistlán, Juchitlán, Tecolotlán, Tenamaxtlán, Puerto Vallarta, Cabo Corrientes, Tomatlán, Atenguillo, Ayutla, Cuautla, Guachinango, Mascota, Mixtlán, San Sebastián del Oeste, and Talpa de Allende—and one in the state of Nayarit: Bahía de Banderas.
Several Agave species found in the region have been used for Raicilla production, including A. inaequidens, A. valenciana (in the mountainous areas), and A. angustifolia and A. rhodacantha (in the coastal regions of Jalisco and Nayarit). However, A. maximiliana currently stands out as the principal species exploited for commercial production (Hernández-Cuevas et al., 2023). The recent rise in Raicilla’s popularity has led to increased consumption and the establishment of extensive agave plantations, often replacing traditional agroforestry systems and contributing to the overharvesting of wild populations (Hernández-Cuevas et al., 2023; Delgado-Aceves et al., 2024a). One example is A. valenciana, a cohabiting species with A. maximiliana, which has recently been classified as critically endangered by The IUCN Red List of Threatened Species™ due to a sharp decline in its natural populations (Torres-García et al., 2018).
In A. maximiliana, propagation is limited to seed germination from mature fruits, which develop approximately four months after pollination. The flowering stalk (scape) typically emerges after seven years (in early maturing individuals at 1500 masl) and up to ten years in others (personal communication, december 2023). This reproductive limitation, combined with the absence of asexual propagation (e.g., bulbils or rhizomatous shoots), makes the species highly vulnerable to degradation of its natural populations. Additionally, seeds produced on the scape are exposed to human extraction, natural degradation, predation by animals, and other biotic and abiotic factors, further reducing the species’ survival rate. Despite the progressive agroindustrial exploitation of A. maximiliana, studies have noted that plantations from various producers still maintain notable levels of genetic diversity. Morphological variability in monoculture plantations reflects the species’ recent domestication history. However, other commercially cultivated Agave species, such as A. tequilana Weber cv. Azul, A. cupreata, and A. potatorum, have undergone artificial selection pressures that have led to reduced genetic diversity, altered genetic structure, and signs of inbreeding (Cabrera-Toledo et al., 2020). To conserve the genetic variability of A. maximiliana without compromising Raicilla production, agroforestry practices should be promoted, and monoculture systems must be modified through agroecological approaches (Torres-García et al., 2019). These should be coupled with long-term conservation strategies using biotechnological tools (Hernández-Cuevas et al., 2023; Delgado-Aceves et al., 2024b). In vitro plant tissue culture can serve as a key component in integrated management strategies for threatened and endangered species, as it enables large-scale propagation of individuals for reforestation and commercial use, without further endangering wild populations (Chávez-Ortiz et al., 2021).
Recent reviews on somatic embryogenesis-based regeneration in Agave species highlight the development of micropropagation protocols in more than ten species of commercial interest. These include A. victoria-reginae (Rodríguez-Garay et al., 1996), A. sisalana (Nikam et al., 2003), A. vera-cruz Mill (Tejavathi et al., 2007), A. tequilana var. Azul (Portillo et al., 2007), A. angustifolia (Arzate-Fernández et al., 2016), A. fourcroydes (Monja-Mio and Robert, 2013), A. americana (Naziri et al., 2019), A. marmorata (Martínez-Martínez et al., 2021), A. tequilana cv. ‘Chato’ (Delgado-Aceves et al., 2022), and A. salmiana (Ángeles-Vázquez et al., 2023). These biotechnological advancements have been applied to optimize the production of diverse Agave-based products, including distilled and fermented beverages, fibers, ornamental and medicinal uses, as well as derivatives like syrups and agave honey (Bautista-Montes et al., 2022). However, a deeper understanding of the hormonal control mechanisms involved in these regeneration protocols is essential, as Agave species exhibit differential responses (Nic-Can and Loyola-Vargas, 2016). Somatic embryogenesis in Agave species has been most successfully induced using synthetic auxins such as 2,4-dichlorophenoxyacetic acid (2,4-D) and picloram, often in combination with cytokinins like kinetin or benzylaminopurine (BAP). These plant growth regulators play a key role in callus induction, maintenance, and embryo formation. For instance, 2,4-D has been widely used to initiate embryogenic calli in A. tequilana (Portillo et al., 2007) A. angustifolia (Arzate-Fernández et al., 2016) and A. salmiana (Ángeles-Vázquez et al., 2023, while picloram has shown higher efficiency A. fourcroydes and cultivars of A. tequilana (Delgado-Aceves et al., 2021). The concentration, duration of exposure, and interaction between auxins and cytokinins significantly affect the embryogenic response and are critical factors for successful large-scale propagation (Nikam et al., 2003; Tejavathi et al., 2007; Rodríguez-Garay, 2016).
In this study, we report for the first time the induction of somatic embryos in A. maximiliana through indirect somatic embryogenesis. This could support the sustainable supply of planting material for Raicilla production while preserving genetic diversity, contribute to the species medium- and long-term conservation, and generate large-scale plant material for reforestation and ecological restoration efforts.
2 Materials and methods
2.1 Seed collection
Mature fruits were collected in May 2023 and 2024 from five Agave maximiliana individuals aged 7 to 8 years, growing in a natural population located in San Jose del Mosco, Mascota, Jalisco, Mexico (20°33’10.7”N, 104°54’02.0”W; 20.552984, -104.900551). To assess seed viability, additional seeds from previous years’ collections—donated by local producers from the same region—were also used.
2.2 Seed germination and viability assessment
Germination tests were performed in accordance with the guidelines of the International Seed Testing Association (International Seed Testing Association, 2021). Given the limited availability of viable (black-colored) seeds, the number of seeds per experimental unit was adjusted accordingly. For each year, four replicates of 50 randomly selected black seeds were used. Seeds were imbibed in distilled water for 24 hours prior to sowing. Following imbibition, seeds were placed above moist paper towels within plastic containers. Germination was assessed ten days after establishment, and seeds were considered germinated when the radicle extended at least 5 mm beyond the seed coat. All tests were conducted under controlled conditions at 25 °C in complete darkness. Germination was expressed as a percentage of total seeds per replicate.
To evaluate seed viability, the seed coat was removed, and the zygotic embryo was extracted from seeds collected in 2019, 2021, 2022, 2023, and 2024. The excised embryos were immersed in a 1% 2,3,5-triphenyltetrazolium chloride solution (0.5 g 2,3,5-triphenyltetrazolium chloride + 50 mL PBS) and incubated in Petri dishes for 2 hours in a NOVATECH® incubator at 40°C in the absence of light.
After incubation, embryos were removed from the 2,3,5-triphenyltetrazolium chloride solution, rinsed with distilled water, and placed on moist paper towels for evaluation. Observations were performed using a Leica® EZ4 HD stereomicroscope to assess staining patterns of the zygotic embryos. The evaluated variables included the number of viable and non-viable embryos, as well as viability and vigor percentages based on staining intensity, following the criteria of Hernández-Castro et al. (2021) in A. angustifolia. Embryos-stained deep red were classified as vigorous, pale red embryos were considered viable with low vigor, and unstained embryos were classified as non-viable.
For each seed collection year, four replicates of 50 seeds were analyzed. Statistical analysis was performed using a Kruskal–Wallis test followed by Dunn’s multiple comparisons test to identify differences in seed viability among years. Analyses were carried out using R version 4.4.1 (R Core Team, 2024), and the PMCMRplus (Pohlert, 2024) package.
2.3 Somatic embryogenesis induction
Seeds were initially washed with 300 µL of TWEEN® 20 and 500 µL of RIDOMIL GOLD® 480 in 250 mL of distilled water under continuous agitation for 1 hour. Following this, under a laminar flow hood, seeds were disinfected in 100 mL of a solution composed of 5% sodium hypochlorite and sterile distilled water in a 30:70 v/v ratio with 150 µL of TWEEN® 20 for 15 minutes. Seeds were then rinsed three times with sterile distilled water to remove residual disinfectant and left in the third rinse container to imbibe for 24 hours. Embryonic axes were extracted from the seeds under a stereomicroscope in aseptic conditions and used as initial explants. Somatic embryogenesis induction was performed using two auxins tested at different concentrations: 0.0, 9.06, 18.12, 27.18, and 36.24 μM of 2,4-dichlorophenoxyacetic acid (2,4-D), and 0.0, 8.28, 16.56, 24.84, and 33.12 μM of 4-amino-3,5,6-trichloro-2-pyridinecarboxylic acid (picloram) following Delgado-Aceves et al. (2021). Each auxin was independently combined with 0.0, 2.22, 3.33, or 4.44 μM of 6-benzylaminopurine (BAP). Embryonic axes were incubated under dark conditions for 40 days at 25 ± 2°C for induction.
Subsequently, the resulting calli were transferred to Petri dishes containing modified semi-solid MS (Murashige and Skoog, 1962) expression medium (with NH4NO3 reduced to 5 mM), supplemented with 500 mg L-1 glutamine and 250 mg L-1 casein hydrolysate, and cultured for an additional 60 days, following the protocol described by Delgado-Aceves et al. (2021). During this phase, cultures were exposed to a 16/8 h light/dark photoperiod at a light intensity of 27 mol m-2 s-1.
A completely randomized factorial design (CRD) was implemented to evaluate the effects of two plant growth regulators on callogenesis in Agave maximiliana. The experiment followed a 5 × 4 factorial scheme,involving: 2,4-D and picloram, each tested at five levels and combined with four levels of BAP during the induction phase. Each treatment included five replicates, and each experimental unit consisted of one Petri dish (60 x 15 mm) containing 10 mL of semi-solid medium and one zygotic embryo as the explant. Aligned Rank Transform ANOVA (Wobbrock et al., 2011) were carried out with the ARTool package (Kay et al., 2015) to evaluate if there was a difference in the percentage of callus formation among the different hormone concentrations, one for 2,4-D and one for Picloram. Estimated marginal means were obtained using the emmeans package (Lenth, 2024), and multiple comparisons were conducted with the multcompView package (Graves et al., 2019). Visualization was created with the ggplot2 package (Wickham, 2016). All of this was done in R version 4.4.1 (R Core Team, 2024).
The number of somatic embryos generated per callus was recorded after 60 days under a stereomicroscope (Leica® EZ4 W) at 10x magnification.
For all treatments, the mean percentage of callus formation was calculated. Friable and embryogenic calli (those showing somatic embryo formation) were selected and subjected to a second induction cycle on expression medium supplemented with 9.06 μM of 2,4-D. The Embryo-Forming Capacity (EFC) index was defined by the equation previously reported by Delgado-Aceves et al. (2021):
2.4 Histological analysis
Additionally, samples of calli and somatic embryos were collected in triplicate for each treatment. Samples were embedded in polyethylene glycol (PEG, molecular weight 1450) at a 1:4 ratio (PEG: deionized water) following the protocol described by Burger and Richter (1991). A rotary microtome was used to obtain 15 µm-thick sections from the PEG-embedded material. Subsequently, a double staining procedure was performed using 0.5% acetocarmine and 0.5% Evans blue in a 1:1 (w/v) ratio, as described by Gupta and Durzan (1987). The same staining protocol was applied to fresh samples. Observations were conducted using a light microscope (Leica® Microsystems EZ4 W).
3 Results and discussion
3.1 Capsule characterization and germination
In this study, the capsules exhibited average dimensions of 6.5 cm in length and 2.6 cm in width. A preliminary seed count per fruit revealed an average of 274 black seeds and 131 white seeds, based on a sample of 10 capsules in 2023 sample (Figure 1).

Figure 1. Biological material of Agave maximiliana. (a) Mature fruit. (b) Viable (black) and non-viable (white) seeds. (c) Tetrazolium-stained zygotic embryos showing full viability. (d) Seed germination.
Although the Agave maximiliana seeds analyzed in this study were collected from the same geographical region, the results show that the year of collection had a considerable effect on seed viability and germination. While seeds collected in 2023 and 2024 exhibited high germination rates (98.4% ± xxx and 90.0 ± 9.8%, respectively), no germination was observed in samples from 2019, 2021, and 2022, despite being evaluated under similar conditions. These findings suggest that factors associated with the year of collection—such as interannual climatic variability, the physiological status of the mother plants at the time of fruit development, or postharvest handling—can significantly influence seed viability, even within the same collection area. This highlights the importance of considering both environmental and temporal contexts in propagation studies.
Additionally, other factors such as seed size or comparisons across distinct geographical zones should also be considered in future germination studies. For instance, seed size has been shown to influence seedling emergence and development in A. salmiana (Vázquez Díaz et al., 2011), although in some cases, such as A. potatorum, germination has varied more strongly with collection date and treatment conditions rather than seed size itself (Villavicencio-Gutiérrez et al., 2024). These considerations highlight the need for a multifactorial approach to understanding germination behavior in Agave species.
3.2 Statistical analysis and viability evaluation
Statistical analyses revealed significant differences in seed viability among the evaluated years (chi-squared = 17.544, df = 4, p-value = 0.001515). Specifically, seeds from 2024 exhibited significantly higher viability compared to those from 2019, 2021, and 2022 (P< 0.05), but not compared to seeds from 2023.
From a qualitative perspective, although no significant differences were observed in the viability percentages between the 2023 and 2024 seeds, a clear difference in embryo vigor was evident. Embryos from 2024 exhibited a more intense coloration when stained with tetrazolium, in contrast to those from 2023 (Figure 2). These results suggest that A. maximiliana seeds experience a significant loss of viability over time, to the extent that after two years of storage at room temperature, they become practically unusable.

Figure 2. Agave maximiliana embryos stained with tetrazolium. (a) Embryos from 2019. (b) Embryos from 2021. (c) Embryos from 2022. (d) Embryos from 2023. (e) Embryos from 2024.
The viability of biological material plays a crucial role in the success of somatic embryogenesis induction (Ángeles-Vázquez et al., 2023). The viability of zygotic embryos is a critical factor in the successful induction of somatic embryogenesis, as it directly influences the cellular competence of the explant and the subsequent morphogenic response (Ángeles-Vázquez et al., 2023). Viable embryos provide metabolically active, totipotent cells capable of re-entering the embryogenic pathway under appropriate in vitro conditions (Merkle and Nairn, 2005). The physiological state of the embryo at the time of excision—particularly its developmental stage, hydration level, and cellular integrity—can markedly affect its responsiveness (Gaj, 2004). Several studies have demonstrated that immature and metabolically active zygotic embryos often exhibit higher embryogenic potential compared to mature or desiccated ones (Jiménez, 2005; Quiroz-Figueroa et al., 2006). Therefore, assessing and ensuring the viability of zygotic embryos prior to culture is essential to optimize somatic embryogenesis protocols, particularly in species with low regenerative capacity or high susceptibility to browning and necrosis.
3.3 Comparative analysis of callogenesis induction
The induction of callogenesis in treatments with 2,4-dichlorophenoxyacetic acid (2,4-D) and 6-benzylaminopurine (BAP) did not exhibit statistically significant differences among the combinations tested (p > 0.05), as all treatments were assigned to the same statistical group (“a”). Nonetheless, a positive trend was observed in the estimated percentages of callus formation, ranging from approximately 30% in the control (0 µM 2,4-D + 0 µM BAP) to over 70% in the treatment containing 9.05 µM 2,4-D + 3.33 µM BAP. Although increasing concentrations of 2,4-D (up to 36.20 µM) appeared to promote a gradual enhancement in callogenesis, this trend was not supported by statistical significance. Likewise, the presence or absence of BAP within the range of 2.22–4.44 µM did not yield any meaningful variation in the formation of callus. These results suggest that, while 2,4-D may have a role in promoting callogenesis, its effect under the tested conditions was not sufficient to generate statistically distinguishable responses, and BAP showed no clear additive effect.
In contrast, the response of callogenesis to treatments combining picloram and BAP varied significantly. The control (0 µM picloram + 0 µM BAP) exhibited the lowest callus induction (≈22%) and was statistically different from the remaining treatments. A progressive increase in callogenesis was observed with higher concentrations of picloram, reaching a maximum response of approximately 85% in treatment 3 (16.56 µM picloram + 0 µM BAP). Overall, increasing picloram levels (up to 33.13 µM) were positively associated with enhanced callogenic potential. However, as observed with 2,4-D, the inclusion of BAP (2.22–4.44 µM) did not significantly improve the callogenic response, regardless of the auxin concentration. These findings indicate that picloram is essential for inducing callogenesis in this system, with the optimal response achieved at 16.56 µM, while BAP does not play a significant role in enhancing this effect.
Supporting these observations, the aligned rank transform (ART) ANOVA revealed a significant effect of 2,4-D concentration on callogenesis (p< 0.001). Post hoc comparisons indicated that the only statistically significant differences occurred between treatments lacking 2,4-D and those that included it. In the case of picloram, ART ANOVA revealed significant main effects of both auxin and cytokinin concentrations, as well as a significant interaction between the two factors (p< 0.001 in all cases). Multiple comparisons identified only treatment 3 (16.56 µM picloram + 0 µM BAP) as being significantly different from the control (p = 0.021), positioning it as the most effective treatment, although only marginally superior to other picloram-based combinations (Figure 3).
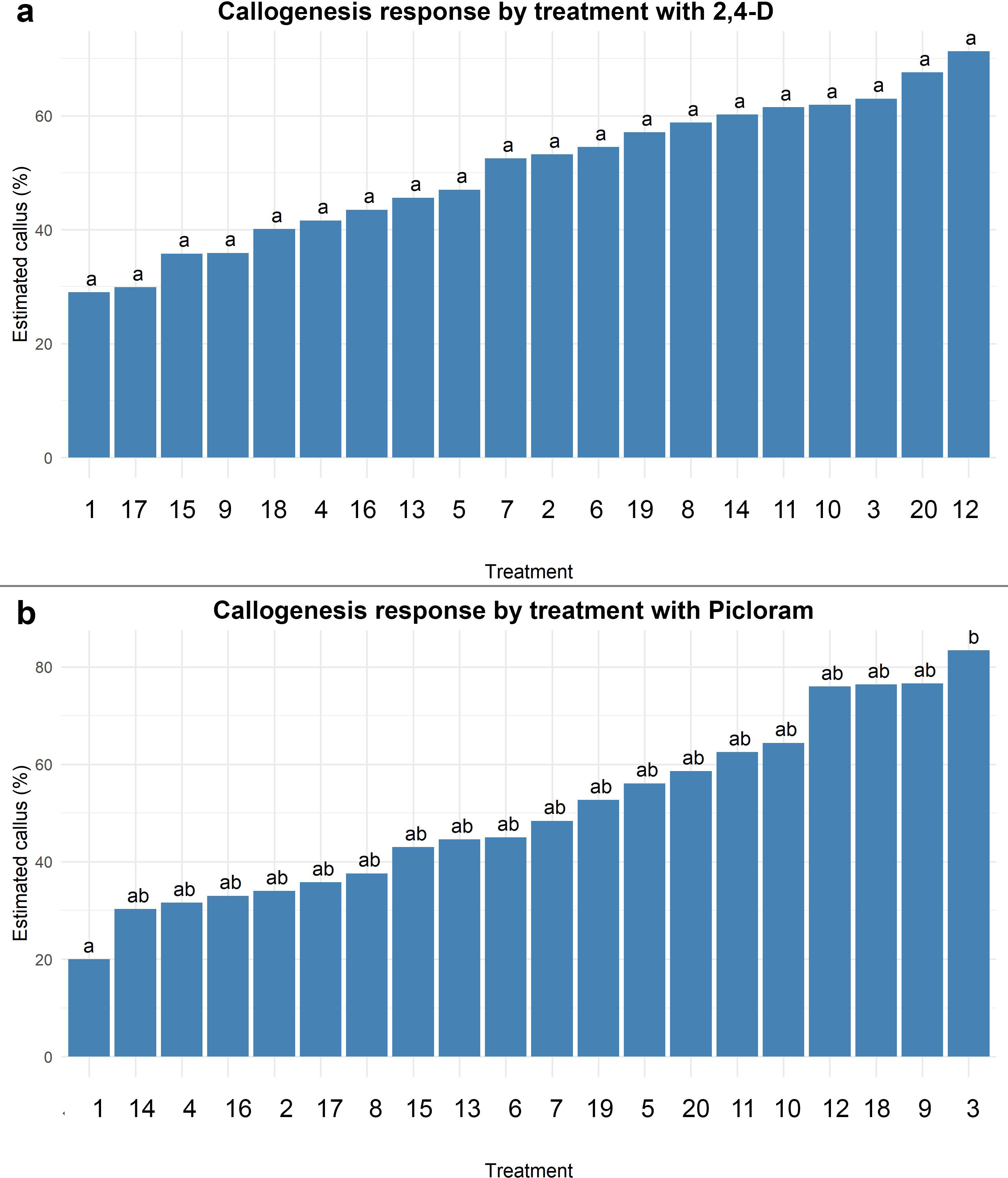
Figure 3. Estimated callogenesis response (%) in Agave maximiliana under different combinations of auxin and 6-Benzylaminopurine (BAP). (a) Treatments using 2,4-D, (b) Treatments using Picloram. Bars represent estimated marginal means (with 95% confidence intervals) from an ART ANOVA model. Treatments are ordered from lowest to highest response. Different letters above bars indicate statistically significant differences between treatments (Tukey-adjusted, p< 0.05).
Although the combination of picloram and BAP demonstrated high efficacy in inducing embryogenic calli, it is important to reiterate that such success is ultimately modulated by the genotype of the explant, which governs the cellular response to exogenous regulators. In Agave species, differences in endogenous hormone sensitivity, metabolic activity, and chromatin organization across genotypes influence both the rate and quality of somatic embryogenesis (Rodríguez-Garay, 2016; Monja-Mio and Robert, 2016). In this study, the use of A. maximiliana zygotic embryos from a single natural population helped control for genotypic variation, yet the observed variability across treatments further supports that embryogenic competence is inherently genotype-dependent, with hormonal combinations acting as modulators rather than primary drivers.
The auxin picloram has been reported as one of the most effective compounds for promoting extensive callus formation in Agave species (Monja-Mio and Robert, 2013; Delgado-Aceves et al., 2021). This high callogenic potential supports the generation of a larger number of dedifferentiated cells, which under appropriate conditions may become polarized and initiate embryo formation. However, the genotype and type of callus (particularly friable), among other abiotic factors, play a critical role in determining the embryogenic potential (Ramírez-Mosqueda, 2022).
3.4 Auxin efficacy and hormonal synergy growth regulators in somatic embryogenesis
The purpose of testing different concentrations and modes of action of auxins to evaluate their ability to induce callus formation in zygotic embryos was to assess the feasibility of using higher or lower concentrations for optimal protocol standardization. Treatments with 2,4-D and picloram resulted in high callus formation rates, callus formation was virtually absent in treatments containing only BAP (without auxins), confirming that BAP alone does not induce callogenesis. The most effective treatment, which combined 36.24 µM 2,4-D, 33.12 µM picloram, and 4.44 µM BAP respectively, achieved an EFC of 20%, suggesting a possible synergistic effect at higher concentrations of both auxins and cytokinins.
An increasing trend in EFC was observed with higher BAP concentrations, although greater variability in the data was also noted. Importantly, callus formation alone did not guarantee the subsequent formation of somatic embryos, indicating that the factors inducing callogenesis do not necessarily trigger somatic embryogenesis. Some treatments that produced a high percentage of callus yielded an EFC of zero, which may suggest the need to modify the type of auxin or its ratio with BAP (Table 1). Notably, treatment 15 (containing 3.33 µM BAP) also stood out, reaching an EFC of 31.92, indicating that this parameter could be associated with indirect regeneration or another embryogenic response.
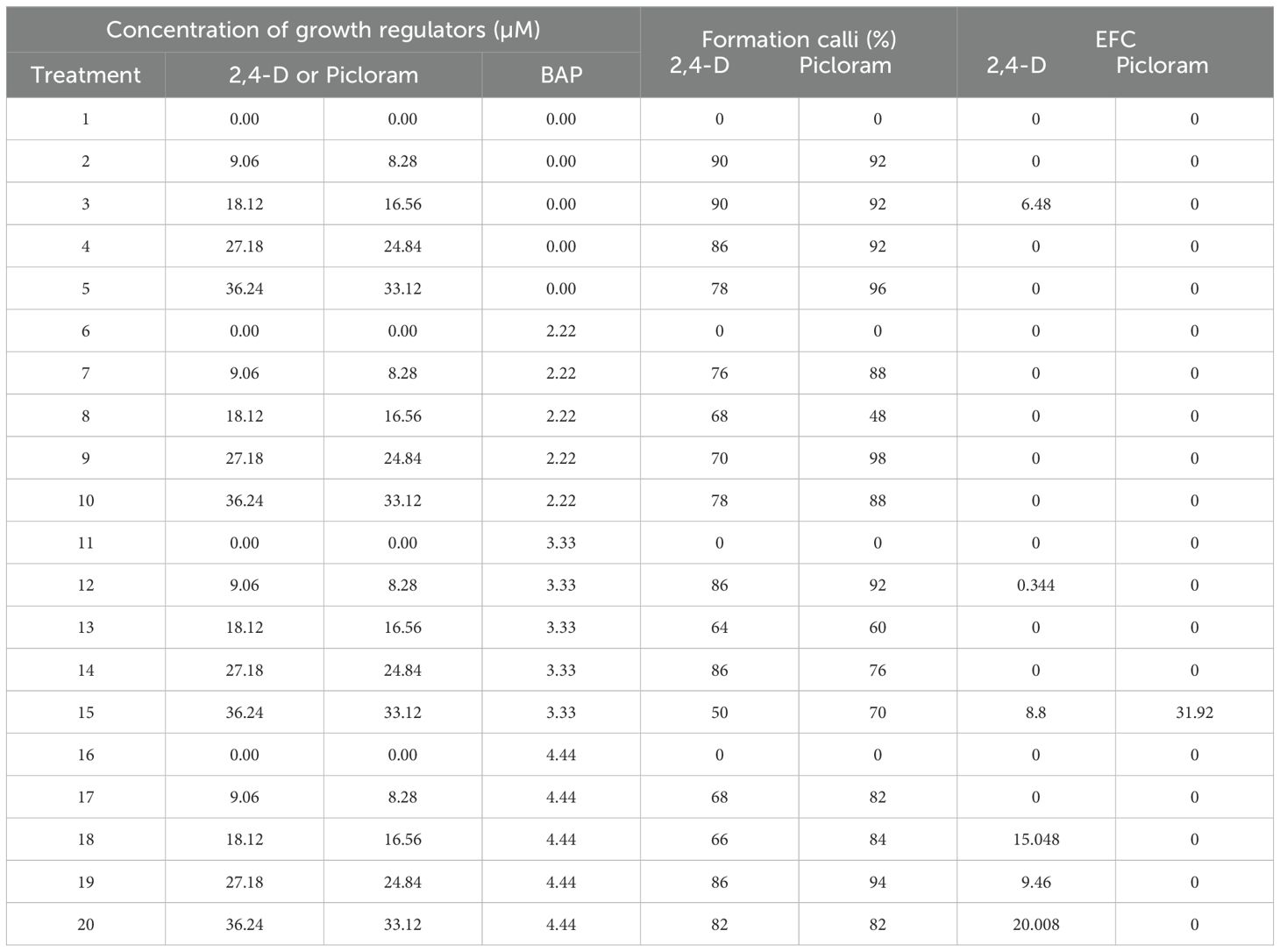
Table 1. Concentrations of 2,4-D and/or picloram on response of eficiency formation capacity (EFC) from zygotic embryos explant of Agave maximiliana after 100 d of culture.
Indirect somatic embryogenesis induced from zygotic embryos has proven to be a viable strategy, primarily due to the accessibility, quality, and physiological state of the embryos used as explants. Several studies have reported that the efficiency of this process is closely linked to the viability of zygotic embryos, making it a key determinant in the successful induction of embryogenic structures (Arzate-Fernández and Mejía-Franco, 2011; Ángeles-Vázquez et al., 2023). The choice of plant growth regulator must be strategic and based on three key factors: the plant species used, the specific stage of the protocol (induction or maturation), and the production goal—whether to maximize the number of embryos or prioritize their regenerative quality (Jiménez, 2005; Von Arnold, 2008). Making informed decisions in this regard enables the optimization of in vitro culture systems and enhances the overall efficiency of plant biofactories.
To better understand the role of auxins in the induction of indirect somatic embryogenesis, a comparative analysis of the behavior of the main auxins used in regeneration processes is presented (Table 2). Picloram, a synthetic auxin, has proven to be highly effective in inducing both callogenesis and somatic embryogenesis in various plant species, notable for its high capacity for cellular penetration and its strong potential to trigger dedifferentiation (Machakova et al., 2008). In contrast, 2,4-D, another widely used synthetic auxin, is well known for promoting cell division and maintaining totipotency in in vitro cultures, making it a key component in many somatic embryogenesis protocols.
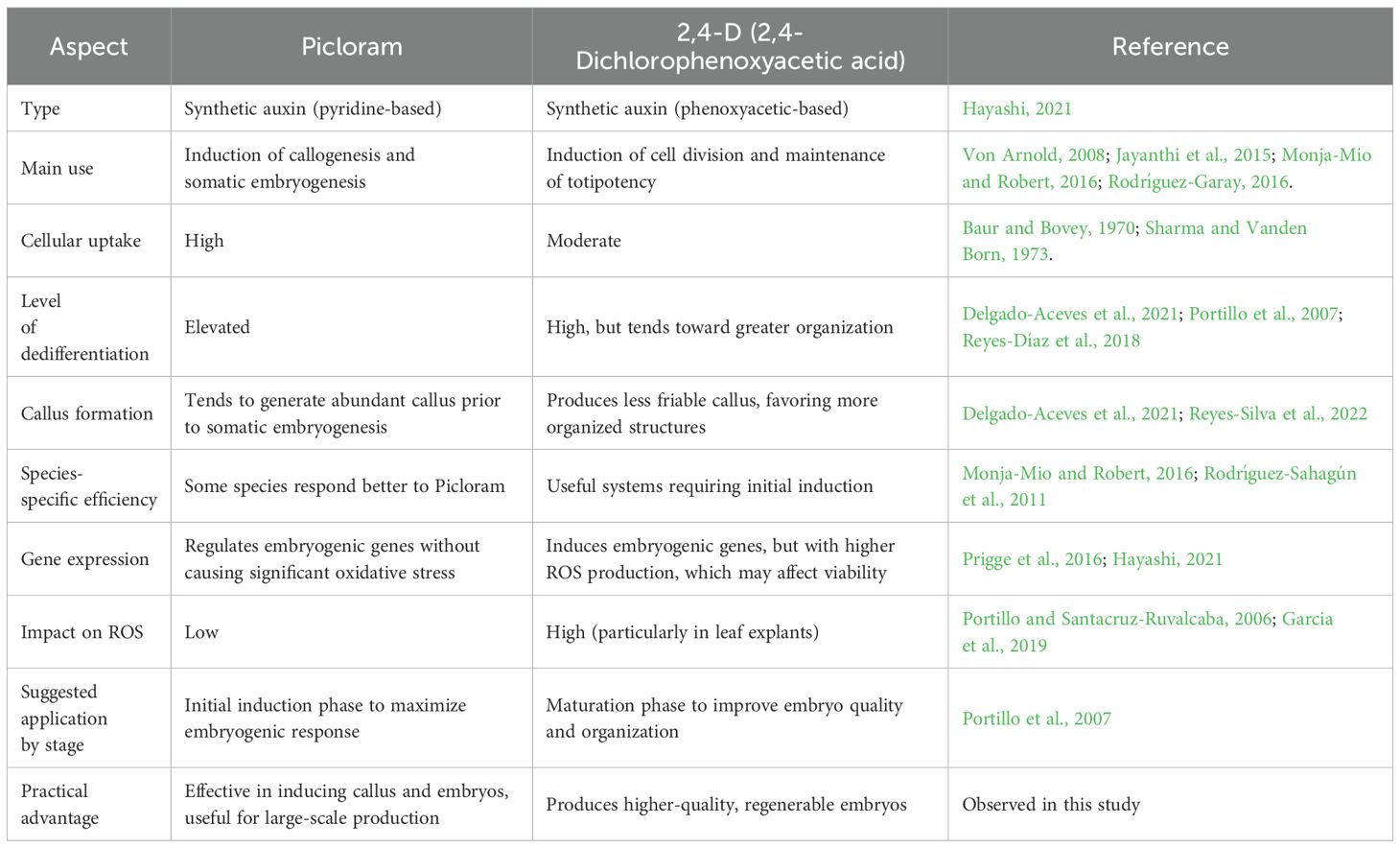
Table 2. Comparative analysis onembryogenic behavior of the auxins 2,4-D and Picloram.
Auxin responses can be regulated through three main pathways: auxin metabolism, polar auxin transport, and signal transduction. Although synthetic auxins differ structurally, they are generally believed to interact with the TIR1/AFB-Aux/IAA co-receptor complex to activate auxin signaling. However, each synthetic auxin may trigger distinct biological responses due to differences in metabolic stability, polar transport dynamics within tissues, and specific affinities for various auxin receptor complexes (Hayashi, 2021). Indirect somatic embryogenesis was evidenced by asymmetric cell divisions, during which the apical–basal developmental axis of the forming embryo began to be established. Subsequently, the proembryogenic structures gave rise to the formation of the somatic embryo proper in the apical region (stained red) and the suspensor in the basal region (stained blue) (Figure 4a). The similarity between zygotic and somatic embryogenesis is further supported by the development of morphologically and functionally equivalent structures as well as by the differentiation of tissues characteristic of the embryo (Rodríguez-Garay, 2016). According to the observations, during the coleoptilar stage, the embryonic axis and radicle are formed independently from the originating callus (Figures 4b–f). In the foliar stage, the shoot and root meristems become defined, along with the differentiation of leaf primordia (Figure 4g) (Delgado-Aceves et al., 2021).
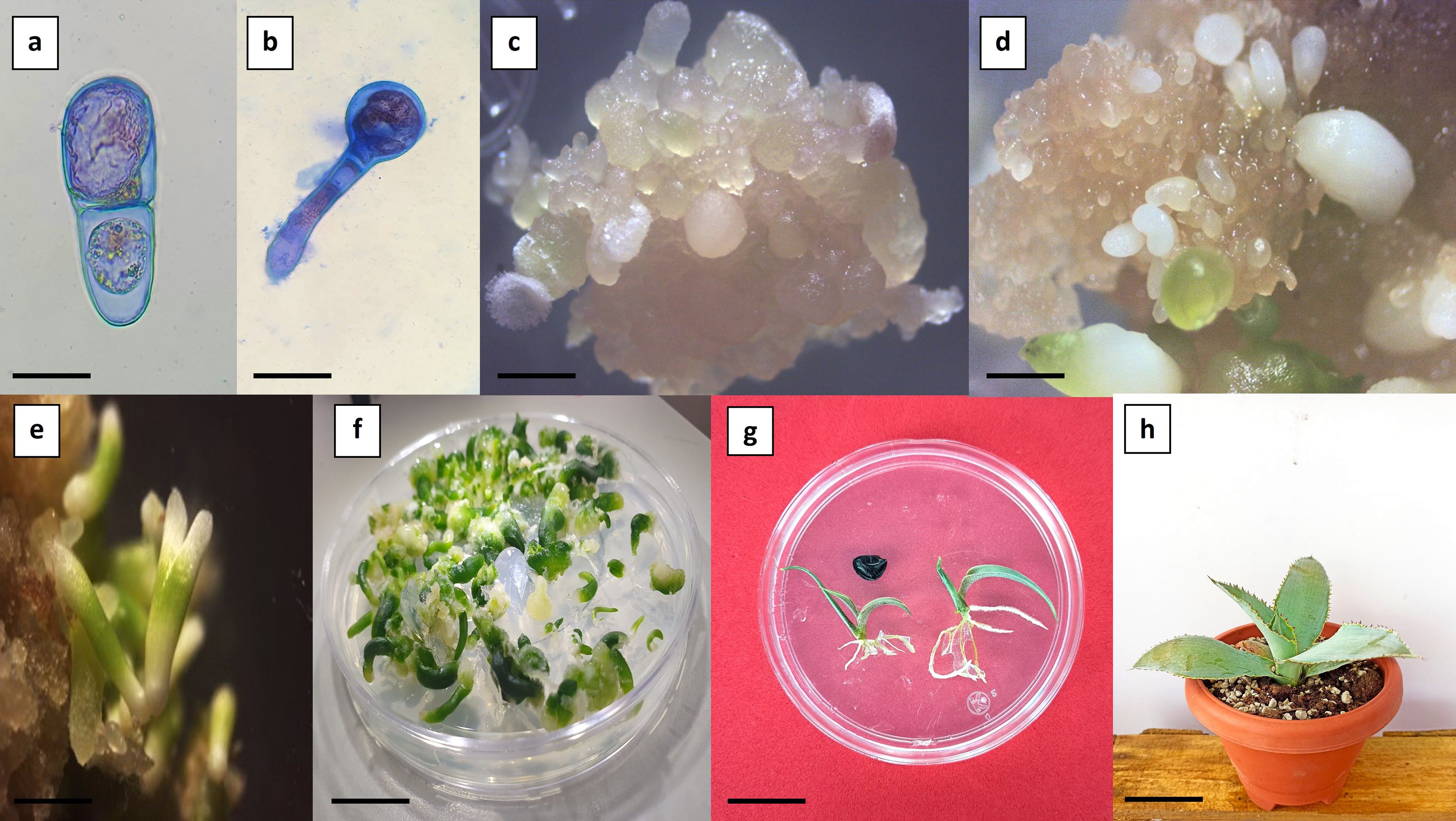
Figure 4. Indirect somatic embryogenesis in Agave maximiliana. (a) Asymmetric cell division evidenced by differential staining (bar = 25 µm). (b) Formation of proembryogenic structures (bar = 100 µm). (c) Friable callus with globular-stage somatic embryos (bar = 2 mm). (d) Friable callus with coleoptilar-stage somatic embryos (bar = 1.5 mm). (e) Presence of somatic embryos with dicot-like morphological characteristics (bar = 3 mm). (f) Germination of somatic embryos (bar = 5 mm). (g) Conversion and developmental comparison of seedlings: zygotic origin (left) and somatic origin (right) (bar = 1.5 cm). (h) Nine-month-old plant derived from somatic embryogenesis (bar = 5 cm).
The conversion rate of germinated somatic embryos into established plantlets reached up to 90% (Figure 4h). However, some somatic embryos displayed abnormal development, characterized by amorphous growth and/or arrested radicle formation. These abnormalities are likely associated with somaclonal variation induced by high concentrations of auxins (Garcia et al., 2019). Somatic embryos originate from a single somatic cell, which allows them to retain the genetic fidelity of the donor plants used in vitro propagation and conservation (Lambardi et al., 2008). However, the potential for somaclonal variation within the system must be considered.
3.5 Histological observations and asynchronous embryo development
Histological observations revealed that the differentiation and reorganization of embryogenic callus occur through a process of progressive cellular independence within the callus itself. It was possible to identify organized structures and characteristic tissues of somatic embryos, including embryos at globular and coleoptilar stages (Figure 5). The regeneration system exhibited asynchronous development, allowing the simultaneous observation of multiple embryogenic stages. Initial observations showed clusters of proembryogenic cells (PEMs) (Figures 5a, b) which represent a transitional stage toward embryogenesis in the presence of auxins. Within the callus tissue, appropriate hormonal gradients are established, leading to the formation of stem cell niches and their subsequent differentiation (Fehér, 2019). These embryo-competent cells display specific traits such as early activation of the cell division cycle, a more alkaline vacuolar pH, altered auxin metabolism, and non-functional chloroplasts (Zavattieri et al., 2010). The reorganization of the embryogenic callus demonstrated defined cell clusters and boundaries, giving rise to tissues characteristic of somatic embryos (Figures 5c, d). The globular and coleoptilar stages observed in this regeneration system (Figures 5e, f) are consistent with those reported in Agave cupreata (Hernández-Solís et al., 2023), A. salmiana (Ángeles-Vázquez et al., 2023), A. tequilana ‘Chato’ (Delgado-Aceves et al., 2021), A. angustifolia (Arzate-Fernández and Mejía-Franco, 2011), A. tequilana ‘Azul’ (Portillo et al., 2007), and A. sisalana (Nikam et al., 2003).
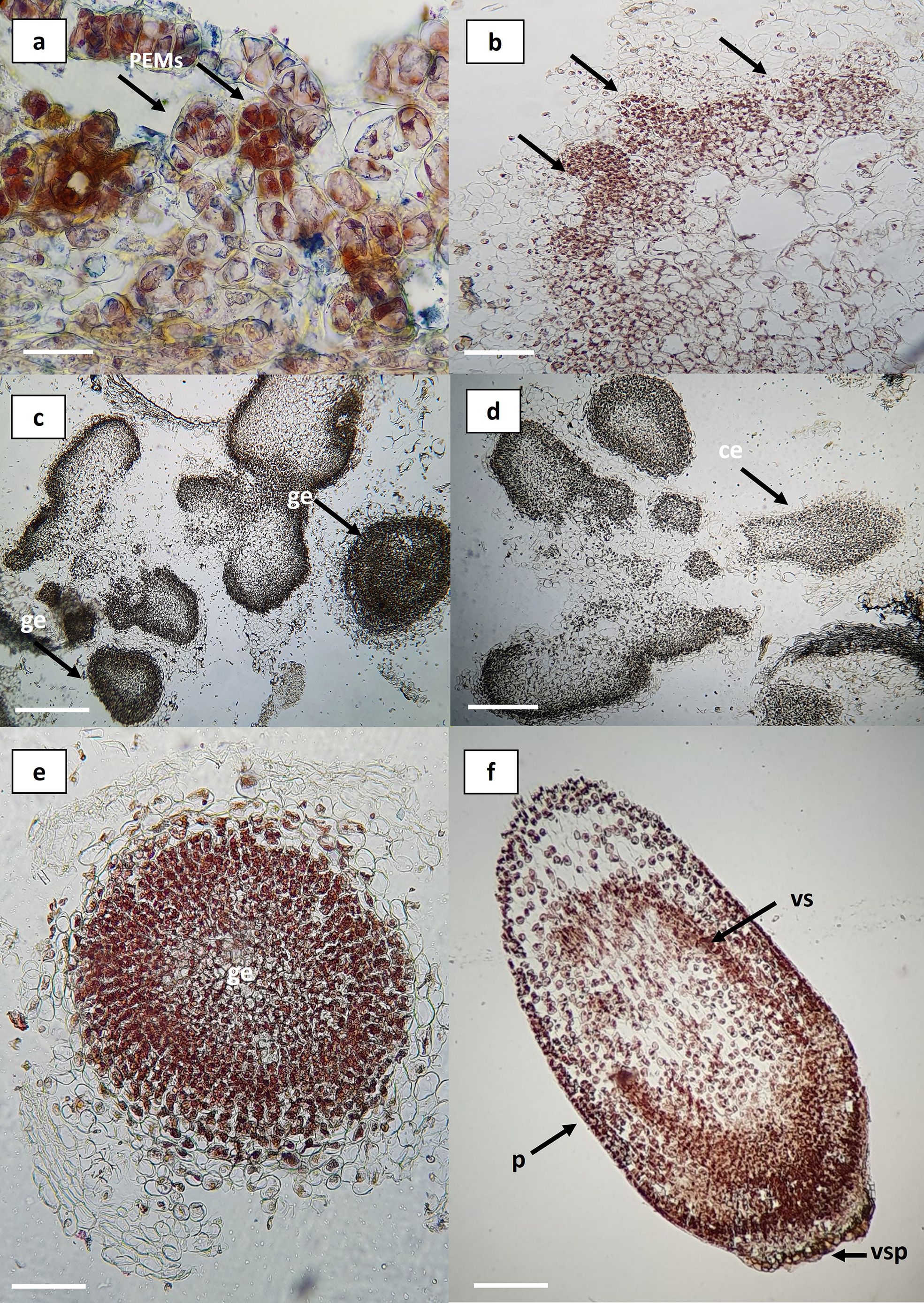
Figure 5. Histological sections of embryogenic callus in Agave maximiliana. (a) Cluster of pro-embryogenic cells (PEMs) in friable callus (bar = 100 µm). (b) Differentiation at the periphery of friable callus (black arrows) (bar = 200 µm). (c, d) Organized structures within the callus (bar = 1 mm). (e) Somatic embryo at the globular stage (bar = 100 µm). (f) Somatic embryo at the coleoptilar stage (bar = 100 µm). PEMs, pro-embryogenic masses; ge, globular embryo; ce, coleoptilar embryo; p, protoderm; vs, vascular strands; vsp, suspensor vestiges.
Therefore, the combination of picloram with the cytokinin BAP provides an effective method for the induction of indirect somatic embryogenesis.
4 Conclusion
This study presents, for the first time, a successful protocol for indirect somatic embryogenesis in Agave maximiliana using zygotic embryos as initial explants. The results demonstrate that both 2,4-D and picloram are effective in inducing embryogenic callus formation, with picloram showing higher efficiency and dose-dependent behavior. The optimal combination of auxins and cytokinins (36.24 µM 2,4-D, 33.12 µM picloram, and 4.44 µM BAP) yielded an EFC of up to 20%, while embryo-to-plantlet conversion rates reached 90%. Histological analyses confirmed the asynchronous development of somatic embryos and their structural and functional similarity to zygotic embryos. The progressive loss of seed viability within two years of storage, along with the species’ reproductive limitations, highlights the urgent need for alternative propagation methods. This protocol offers a reliable and scalable biotechnological tool for the conservation, reforestation, and sustainable production of A. maximiliana, contributing to the preservation of its genetic diversity and supporting the long-term viability of production systems. Furthermore, integrating this system into agroforestry schemes could support both the conservation and sustainable production of A. maximiliana in regions under pressure from Raicilla commercialization.
It is recommended to continue experiments using auxin and cytokinin concentrations near those that promoted somatic embryo formation. Additionally, it is essential to conduct genetic validation studies to ensure clonal stability, as well as to evaluate the agronomic performance of the regenerated plants under field conditions.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
LD: Writing – review & editing, Visualization, Methodology, Investigation, Software, Formal analysis, Writing – original draft, Data curation, Supervision, Conceptualization, Resources. SC: Data curation, Writing – original draft, Formal analysis, Visualization, Software, Conceptualization, Writing – review & editing, Investigation. JC: Supervision, Writing – review & editing, Visualization, Validation. JR: Validation, Visualization, Writing – review & editing, Supervision. AG: Validation, Visualization, Supervision, Investigation, Writing – review & editing, Funding acquisition, Project administration.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We express our deepest gratitude to the producers for their generosity, knowledge, and commitment—fundamental elements in the development of this work. We would like to thank Mr. Benito Salcedo and Manuel Salcedo whose collaboration and openness were especially valuable to this project. We also extend our sincere appreciation to Secretaría de Ciencia, Humanidades, Tecnología e Innovación (SECIHTI) for the financial support provided through the postdoctoral fellowship program, which enabled the consolidation of this initiative.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ángeles-Vázquez, B. V., Alvarez-Cervantes, J., Tovar-Jiménez, X., and Rodríguez-Garay, B. (2023). Plant Regeneration from Indirect Somatic Embryogenesis of Agave salmiana Otto ex Salm-Dyck Subsp. salmiana Using Zygotic Embryo Obtained by In-casa Pollination as Explants. Polibotánica 56, pp 171–182. doi: 10.18387/polibotanica.56.9
Arzate-Fernández, A. M. and Mejía-Franco, R. (2011). Capacidad Embriogénica de Callos Inducidos en Ejes Embrionarios Cigóticos de Agave angustifolia Haw. Rev. Fitotecnia Mexicana. 34, 2. doi: 10.35196/rfm.2011.2.101
Arzate-Fernández, A. M., Piña-Escutia, J. L., Norman-Mondragón, T. H., Reyes-Díaz, J. I., Guevara-Suárez, K. L., and Vázquez-García, L. M. (2016). Regeneración de Agave Mezcalero (Agave angustifolia Haw.) a Partir de Embriones Somáticos Encapsulados. Rev. Fitotecnia Mexicana. 39, 4. Available online at: https://www.redalyc.org/articulo.oa?id=61049142004 (Accessed July 11, 2025).
Baur, J. R. and Bovey, R. W. (1970). The uptake of picloram by potato tuber tissue. Weed Science. 18, 1. doi: 10.1017/S0043174500077262
Bautista-Montes, E., Hernández-Soriano, L., and Simpson, J. (2022). Advances in the micropropagation and genetic transformation of Agave species. Plants 11, 13. doi: 10.3390/plants11131757
Burger, L. M. and Richter, H. G. (1991). Anatomia da Madeira (São Paulo, Brazil: Livraria Nobel S. A), 105.
Cabrera-Toledo, D., Vargas-Ponce, O., Ascencio-Ramírez, S., Valadez-Sandoval, L. M., Pérez-Alquicira, J., Morales-Saavedra, J., et al. (2020). Morphological and genetic variation in monocultures, forestry systems and wild populations of Agave maximiliana of Western Mexico: implications for its conservation. Front. Plant Science. 11. doi: 10.3389/fpls.2020.00817
Chávez-Ortiz, L. I., Morales-Domínguez, J. F., Rodríguez-Sahagún, A., and Pérez-Molphe-Balch., E. (2021). In vitro propagation of Agave guiengola gentry using semisolid medium and temporary immersion bioreactors. Phyton – Int. J. Exp. Botany. 90, 3. doi: 10.32604/phyton.2021.012862
Delgado-Aceves, L., Corona, S., Marin-Castro, U. R., Rascón-Díaz, M. P., Portillo, L., Gutiérrez-Mora, A., et al. (2024b). Comparative studies for cryopreservation of agave shoot tips by droplet-vitrification. Plants 13, 2609. doi: 10.3390/plants13182609
Delgado-Aceves, L., González-Arnao, M. T., Santacruz-Ruvalcaba, F., Folgado, R., and Portillo., L. (2021). Indirect somatic embryogenesis and cryopreservation of Agave tequilana weber cultivar ‘Chato’. Plants 10, 2. doi: 10.3390/plants10020249
Delgado-Aceves, L., Gutiérrez-Mora, A., and B. Salcedo-Ríos and Corona, P. (2024a). Manejo Agrobiotecnológico para la producción y conservación de Agave maximiliana Baker (Jalisco: Consejo editorial de CIATEJ).
Delgado-Aceves, M. L., Portillo, L., Folgado, R., Romo-Paz, F. J., and González-Arnao., M. T. (2022). New approaches for micropropagation and cryopreservation of Agave peacockii, an endangered species. Plant Cell Tiss Organ Cult 150. 85–95. doi: 10.1007/s11240-022-02299-2
Diario Oficial de la Federación (2019).PROYECTO de Norma Oficial Mexicana PROY-NOM-257-SE-2021, Bebidas alcohólicas-Raicilla-Denominación, especificaciones, información comercial y métodos de prueba. Available online at: https://www.dof.gob.mx/nota_detalle_popup.php?codigo=5650295 (Accessed May 01, 2025).
Fehér, A. (2019). Callus, dedifferentiation, totipotency, somatic embryogenesis: what these terms mean in the era of molecular plant biology? Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00536
Gaj, M. D. (2004). Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 43, 27–47. doi: 10.1023/B:GROW.0000038275.29262.fb
Garcia, C., Furtado de Almeida, A. A., Costa, M., Britto, D., Valle, R., Royaert, S., et al. (2019). “Abnormalities in somatic embryogenesis caused by 2,4-D: an overview,” in Plant Cell Tiss Organ Cult. 137, 193–212. doi: 10.1007/s11240-019-01569-8
Graves, S., Piepho, H. P., Selzer, L., and Dorai-Raj, S. (2019). multcompView: Visualizations of Paired Comparisons. R package version 0.1-9. Available online at: https://cran.r-project.org/package=multcompView (Accessed July 11, 2025).
Gupta, P. and Durzan, D. (1987). Biotechnology of somatic polyembryogenesis and plantlet regeneration in loblolly pine. Bio/Technology 5, pp 147–151. doi: 10.1038/nbt0287-147
Hayashi, K. I. (2021). Chemical biology in Auxin research. Cold Spring Harbor Perspect. Biol. 13, 5. doi: 10.1101/cshperspect.a040105
Hernández-Castro, E., López-Sandoval, Y. Y., Escobar-Álvarez, J. L., Ramírez-Reynoso, O., Maldonado-Peralta, M. A., and Valenzuela-Lagarda, J. L. (2021). Análisis Morfométrico de Semilla y Desarrollo de Plántulas de Maguey Sacatoro (Agave angustifolia Haw.). Ecosistemas y Recursos Agropecuarios. 8, 3. doi: 10.19136/era.a8n3.2964
Hernández-Cuevas, L. V., Salinas-Escobar, L. A., Segura-Castruita, M. Á., Palmeros-Suárez, P. A., and Gómez-Leyva, J. F. (2023). Physiological responses of Agave maximiliana to inoculation with autochthonous and allochthonous arbuscular mycorrhizal fungi. Plants (Basel). 12, 3. doi: 10.3390/plants12030535
Hernández-Solís, M., Arzate-Fernández, A. M., Martínez-Martínez, S. Y., and Acosta-Villagrán., L. (2023). Effect of promoting compounds of indirect somatic embryogenesis in three Agave species. Agrociencia 57, 1. doi: 10.47163/agrociencia.v57i1.2858
International Seed Testing Association. (2021). International rules for seed testing 2021. Wallisellen, Switzerland: ISTA.
Jayanthi, M., Susanthi, B., Murali Mohan, N., and Mandal, P. K. (2015). In vitro somatic embryogenesis and plantlet regeneration from immature male inflorescence of adult Dura and Tenera palms of Elaeis guineensis (Jacq.). SpringerPlus 4. doi: 10.1186/s40064-015-1025-4
Jiménez, V. M. (2005). Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul. 47, 91–110. doi: 10.1007/s10725-005-3478-x
Kay, M., Wobbrock, J. O., and Laviolette, S. R. (2015). ARTool: Aligned Rank Transform for nonparametric factorial analyses. R package version 0.10.7. Available online at: https://cran.r-project.org/package=ARTool (Accessed July 11, 2025).
Lambardi, M., Ozudogru, A., and Benelli, C. (2008). “Cryopreservation of embryogenic cultures,” in Plant Cryopreservation: A Practical Guide. Ed. Reed, B. M. (Springer, New York, EUA), 177–210.
Lenth, R. V. (2024). emmeans: Estimated Marginal Means, aka Least-Squares Means. R package version 1.10.0. Available online at: https://cran.r-project.org/package=emmeans (Accessed July 11, 2025).
Machakova, I., Zazimalova, E., and George, E. (2008). “Plant growth regulators I: introduction; auxins their analogues and inhibtors,” in Plant Propagation by Tissue Culture, 3rd Edition. Eds. George, E. F., Hall, M. A., and De Klerk, G. (Springer, The Netherlands), 175–204.
Martínez-Martínez, S., Arzate-Fernández, A., Álvarez-Aragón, C., Martínez-Velasco, I., and Norman-Mondragón, T. (2021). Regeneration of Agave marmorata Roelz plants in temporary immersion systems, via organogenesis and somatic embryogenesis. Trop. Subtrop Agroecosyst 24. doi: 10.56369/tsaes.3472
Merkle, S. A. and Nairn, C. J. (2005). Hardwood tree biotechnology. In Vitro Cell Dev Biol - Plant 41, 602–619. doi: 10.1079/IVP2005687
Monja-Mio, K. M. and Robert, M. L. (2013). Direct somatic embryogenesis of Agave fourcroydes Lem. through thin cell layer culture. In Vitro Cell Dev Biol - Plant 49, 541–549. doi: 10.1007/s11627-013-9535-7
Monja-Mio, K. M. and Robert, M. L. (2016). “Somatic embryogenesis in Agave: an overview,” in Somatic Embryogenesis: Fundamental Aspects and Applications. Eds. Loyola-Vargas, V. M. and Ochoa-Alejo, N. (Springer, Cham). doi: 10.1007/978-3-319-33705-0_17
Murashige, T. and Skoog, F. (1962). A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiologia Plantarum. 15, 3. doi: 10.1111/j.1399-3054.1962.tb08052.x
Naziri, M., Sadat, S., and Soltani Howyzeh, M. (2019). The effect of different hormone combinations on direct and indirect somatic embryogenesis in Agave americana. Iranian J. Plant Physiol. 9, 2739–2747. doi: 10.22034/IJPP.2019.664579
Nic-Can, G. I. and Loyola-Vargas, V. M. (2016). “The role of the auxins during somatic embryogenesis,” in Somatic Embryogenesis: Fundamental Aspects and Applications. Eds. Loyola-Vargas, V. M. and Ochoa-Alejo, N. (Springer, Cham), 171–190. doi: 10.1007/978-3-319-33705-0_10
Nikam, T. D., Bansude, G. M., and Aneesh Kumar., K. C. (2003). Somatic Embryogenesis in Sisal (Agave sisalana Perr. ex. Engelm). Plant Cell Rep. 22, 3. doi: 10.1007/s00299-003-0675-9
Pohlert, T. (2024). PMCMRplus: Calculate Pairwise Multiple Comparisons of Mean Rank Sums Extended (Version 1.9.12). Available online at: https://CRAN.R-project.org/package=PMCMRplus (accessed July 11, 2025).
Portillo, L. and Santacruz-Ruvalcaba, F. (2006). Obtención de Embrioides de Agave tequilana Weber a Partir de Explantes de Raíz. Zonas Áridas, 11–19.
Portillo, L., Santacruz-Ruvalcaba, F., Gutiérrez-Mora, A., and Rodríguez-Garay., B. (2007). Somatic embryogenesis in Agave tequilana weber cultivar Azul. In Vitro Cell Dev Biol - Plant 43, 569–575. doi: 10.1007/s11627-007-9046-5
Prigge, M. J., Greenham, K., Zhang, Y., Santner, A., Castillejo, C., Mutka, A. M., et al. (2016). The Arabidopsis auxin receptor F-box proteins AFB4 and AFB5 are required for response to the synthetic auxin picloram. G3: Genes Genomes Genet. 6, 5. doi: 10.1534/g3.115.025585
Quiroz-Figueroa, F. R., Rojas-Herrera, R., Galaz-Ávalos, R. M., and Loyola-Vargas, V. M. (2006). Embryo production through somatic embryogenesis can be used to study cell differentiation in plants. Plant Cell, Tissue and Organ Culture 86, 285–301. doi: 10.1007/s11240-006-9139-6
Ramírez-Mosqueda, M. A. (2022). “Perspectives of somatic embryogenesis: concluding remarks,” in Somatic Embryogenesis: Methods and Protocols, Methods in Molecular Biology, vol. 2527 . Eds. German-Taramasco, M. and A., M. (Humana, New York, NY), 267–270. doi: 10.1007/978-1-0716-2485-2_19
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna: R Foundation for Statistical Computing). Available online at: https://www.R-project.org/ (Accessed July 11, 2025).
Reyes-Díaz, J. I., Arzate-Fernández, A. M., and Piña-Escutia, J. L. (2018). Sucrose and organic nitrogen sources have an influence in Agave angustifolia somatic embryogenesis. Rev. Mexicana Cienc. Agrícolas. 9, 7. doi: 10.29312/remexca.v9i7.1676
Reyes-Silva, A. I., Nuñez-Palenius, H. G., Ocampo, G., and Pérez-Molphe-Balch, E. (2022). Regeneración in vitro de Agave wocomahi Gentry (Asparagaceae). Rev. Fitotecnia Mexicana. 45, 4. doi: 10.35196/rfm.2022.4.493
Rodríguez-Garay, B. (2016). “Somatic embryogenesis in Agave spp,” in Somatic Embryogenesis: Fundamental Aspects and Applications. Eds. Loyola-Vargas, V. M. and Ochoa-Alejo, N. (Springer, Cham). doi: 10.1007/978-3-319-33705-0_16
Rodríguez-Garay, B., Gutiérrez-Mora, A., and Acosta-Dueñas, B. (1996). Somatic embryogenesis of Agave victoria-reginae Moore. Plant Cell Tiss Organ Cult. 46, 85–7. doi: 10.1007/BF00039700
Rodríguez-Sahagún, A., Acevedo-Hernández, G., Rodríguez-Domínguez, J. M., Rodríguez-Garay, B., Cervantes-Martínez, J., and Castellanos-Hernández, O. A. (2011). Effect of Light Quality and Culture Medium on Somatic Embryogenesis of Agave tequilana Weber var. Azul. Plant Cell Tissue Organ Culture. 104, 2. doi: 10.1007/s11240-010-9815-4
Santacruz-Ruvalcaba, F., Castañeda-Nava, J. J., Villanueva-Gónzalez, J. P., García-Sahagún, M. L., Portillo, L., and Contreras-Pacheco, M. L. (2022). Micropropagation of Agave maximiliana baker by axillary shoot proliferation. Polibotánica 54, pp 139–151. doi: 10.18387/polibotanica.54.9
Sharma, M. P. and Vanden Born, W. H. (1973). Uptake, cellular distribution, and metabolism of 14C-picloram by excised plant tissues. Physiologia Plantarum. 29, 1. doi: 10.1111/j.1399-3054.1973.tb04801.x
Tejavathi, D. H., Rajanna, M. D., Sowmya, R., and Gayathramma, K. (2007). Induction of somatic embryos from cultures of Agave vera-cruz Mill. In Vitro Cell Dev Biol - Plant 43, 423–428. doi: 10.1007/s11627-007-9088-8
Torres-García, I., García-Mendoza, A. J., and Casas, A. (2019a). Agave valenciana. The IUCN Red list of threatened species 2019: e.T115698769A116354608. doi: 10.2305/IUCN.UK.2019-3.RLTS.T115698769A116354608.en
Torres-García, I., Rendón-Sandoval, F. J., Blancas, J., Casas, A., and Moreno-Calles, A. I. (2019). The genus agave in agroforestry systems of Mexico. Botanical Sciences 97, 263–290. doi: 10.17129/botsci.2201
Vázquez Díaz, E., García Núñez, J. R., Peña Vásquez, C. B., Ramírez Tovar, H. M., and Morales Rodríguez, V. (2011). Tamaño de semilla, emergencia y desarrollo de la plántula de maguey (Agave salmiana Otto ex Salm-Dyck). Rev. Fitotecnia Mexicana 34, 167–173. doi: 10.35196/rfm.2011.3.167
Vázquez-García, J. A., Cházaro-Basáñez, M., Hernández-Vera, G., Vargas-Rodríguez, Y., and Pilar-Zamora, T. (2007). “Taxonomía del género Agave en el occidente de México: una panorámica preliminar,” in Agaves del Occidente de México. Eds. Vázquez-García, J. A., Cházaro-Basáñez, M., Hernández-Vera, G., and Zamora, P. (Universidad de Guadalajara, CUCEI-CUCBA press, Guadalajara, Mexico), 38–82.
Villavicencio-Gutiérrez, E. E., Torres-Tapia, M. A., Méndez González, J., Curiel-López, C., and Sánchez Pérez, F. (2024). Germinación de semillas en Agave potatorum Zucc. Rev. Mexicana Cienc. Forestales 15, 31–51. doi: 10.29298/rmcf.v15i86.1403
Von Arnold, S. (2008). “Somatic embryogenesis,” in Plant Propagation by Tissue Culture, 3rd ed, vol. 1 . Eds. George, E. F., Hall, M. A., and De Klerk, G. J. (Springer, Dordrecht, The Netherlands), 335–354.
Wickham, H. (2016). ggplot2: Elegant Graphics for Data Analysis (New York: Springer). Available online at: https://ggplot2.tidyverse.org (Accessed July 11, 2025).
Wobbrock, J. O., Findlater, L., Gergle, D., and Higgins, J. J. (2011). The aligned rank transform for nonparametric factorial analyses using only ANOVA procedures. Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, 143–146. doi: 10.1145/1978942.1978963
Keywords: zygotic embryo, viability, callus, somatic embryo, auxins, Raicilla
Citation: Delgado-Aceves L, Corona S, Castañeda-Nava JJ, Rodríguez-Domínguez JM and Gutiérrez-Mora A (2025) Indirect somatic embryogenesis of Agave maximiliana Baker. Front. Plant Sci. 16:1648362. doi: 10.3389/fpls.2025.1648362
Received: 18 June 2025; Accepted: 01 August 2025;
Published: 28 August 2025.
Edited by:
Elena Corredoira, Spanish National Research Council (CSIC), SpainReviewed by:
Valerie Pence, Cincinnati Zoo and Botanical Garden, United StatesRené García Martínez, Tecnologico de Estudios Superiores de Valle de Bravo, Mexico
Maria Teresa Gonzalez Arnao, Universidad Veracruzana, Mexico
Copyright © 2025 Delgado-Aceves, Corona, Castañeda-Nava, Rodríguez-Domínguez and Gutiérrez-Mora. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Antonia Gutiérrez-Mora, YWd1dGllcnJlekBjaWF0ZWoubXg=