 Zhixin Li†
Zhixin Li† Mingming Hu†Tao LiuYuan TangRuhongji LiuZhengbo PengCheng WangZhenglan PengZhonglin Wang
Mingming Hu†Tao LiuYuan TangRuhongji LiuZhengbo PengCheng WangZhenglan PengZhonglin Wang Zongkui ChenZhiyuan Yang
Zongkui ChenZhiyuan Yang Yongjian Sun
Yongjian Sun Jun Ma*
Jun Ma*- Rice Research Institute of Sichuan Agricultural University/Crop Ecophysiology and Cultivation Key Laboratory of Sichuan Province, Chengdu, China
Introduction: Imbalanced N–K ratios reduce nutrient uptake efficiency while increasing lodging susceptibility, thereby destabilizing yield potential. Optimizing N–K ratios is therefore crucial for improving nutrient efficiency, lodging resistance, and yield potential.
Methods: This study employed the hybrid indica rice cultivar F-you 498 as experimental material. Two K management strategies (basal:panicle = 10:0 and 5:5, denoted as K1 and K2) and three N application regimes (basal:tiller:panicle = 7:3:0, 5:3:2, and 3:3:4, denoted as N1, N2, and N3) were tested. Both fertilizers were applied at identical total rates of 150 kg ha-¹ for N and K to investigate N–K interactions on rice growth and nutrient utilization.
Results: N–K interactions significantly affected dry matter accumulation, nutrient uptake, lodging resistance, and yield. Split potassium application (K2) increased grain yield by 3.06% compared with basal-only application (K1), by increasing productive panicles, spikelets per panicle, total spikelets, and seed-setting rate. K2 enhanced post-heading dry matter translocation and improved N–K uptake, elevating panicle N and K accumulation by 5.01% and 13.70%, respectively. K2 also significantly improved lodging resistance. Under K2, the N3 treatment further increased yield by enhancing the number of effective panicles, grains per panicle, and total spikelets, with average yield increases of 12.17% and 4.77% compared with N1 and N2, respectively. Post-heading dry matter accumulation, remobilization ratio, and contribution rate in N3 were higher than in N1 and N2, with two-year average increases of 25.54%, 5.37%, and 7.42% compared with N1, and 12.68%, 2.76%, and 2.57% compared with N2. N3 also promoted the translocation of N and K. Compared with N1 and N2, N3 increased whole-plant N translocation and N transferred to the panicle by 38.09% and 27.45%, and by 14.53% and 12.45%, respectively; whole-plant K translocation and K transferred to the panicle increased by 11.46% and 28.26%, and by 13.35% and 18.35%, respectively. Additionally, N3 improved lodging resistance by thickening internodes and stem-sheath walls. The lodging index was significantly negatively correlated with N and K accumulation in stem-sheaths.
Discussion: Overall, the K2N3 combination enhances post-heading assimilate allocation and nutrient translocation in machine-transplanted rice, strengthens stem mechanical properties, optimizes panicle traits, and ultimately achieves stable and high yields.
1 Introduction
As a primary food crop, enhancing the cultivation and productivity of rice has long been a priority in agricultural research. Recent technological advancements and agricultural modernization have amplified the importance of mechanized planting in rice production. Compared with manual transplanting, machine-transplanted rice offers superior operational efficiency and reduced labor intensity (Bhatt et al., 2023), making it a key cultural practice in major rice-growing regions of China. Current research predominantly focuses on planting density (Begum et al., 2022), fertilization methods (Mohapatra et al., 2022), micronutrient supplementation (Jinjala et al., 2024), or single-nutrient management (Jiang et al., 2024). However, persistent misalignment between conventional fertilization practices and the nutrient supply–demand dynamics of mechanized rice systems continues to constrain yield potential. This highlights the urgent need to mitigate structural imbalances in nutrient supply–demand patterns under mechanized cultivation to further unlock rice productivity and safeguard food security.
Nitrogen (N) is one of the essential nutrients required for rice growth. It enhances the synthesis of photosynthates, promotes the differentiation of tillers into productive panicles and florets (Song et al., 2025; Chen et al., 2018), and facilitates N remobilization to grains by regulating the activities of nitrate reductase (NR) and glutamine synthetase (GS) (Wu et al., 2025). An appropriate N application rate (180–240 kg ha-1), with a basal-to-panicle fertilizer ratio of 6:4, can help balance source–sink dynamics and improve the efficiency of assimilate translocation. In machine-transplanted rice production, excessive reliance on N fertilizers for yield pursuit, while neglecting balanced potassium (K) application, often results in structural imbalance in fertilization. K plays a critical role in rice growth by acting as an activator of various enzymes, enhancing CO2 assimilation, and promoting the translocation of photosynthates to developing grains. Additionally, K regulates N metabolism through its effects on enzymes such as NR and GS, thereby improving N remobilization to the grain and increasing the N harvest index (Gao et al., 2025; Tian et al., 2014). Studies have shown that balanced application of N and K not only improves nutrient use efficiency and reduces nutrient losses that contribute to environmental pollution, but also enhances lodging resistance by promoting lignin biosynthesis and thickening of stem cell walls (Chen et al., 2024; Lei et al., 2025).
In recent years, progress has been made in nutrient management research for machine-transplanted rice. Studies have shown that machine-transplanted rice requires more N (with an optimal N application rate of 225 kg ha-1) than conventional cultivation (160–200 kg ha-1) (Yao et al., 2021). Therefore, quantifying N demand is crucial for diagnosing N nutritional status and improving N use efficiency in mechanized systems. Further research has revealed that combined N and K application can balance the number of effective panicles and the length of basal internodes in machine-transplanted rice. Specifically, when 180 kg ha-¹ of N and 120 kg ha-¹ of K were applied, dry matter accumulation and sheath density increased, while bending moment decreased and breaking resistance improved, resulting in significantly higher grain yields (Sun et al., 2017). In addition, increasing K by 30% could offset the adverse effects of a 30% N reduction, significantly improving spikelet number and seed-setting rate, thereby enhancing rice productivity (Deng et al., 2024). However, previous studies have predominantly focused on single-nutrient input strategies or comparisons of fertilization rates, lacking a comprehensive analysis of N–K interactions and their optimal ratios in machine-transplanted rice cultivation. Addressing this gap is essential to resolve the current dilemma in mechanized rice production—high yields without efficiency, and increased output without proportional economic returns. Therefore, this study integrates the agronomic advantages of potted nursery mat transplanting with the physiological characteristics of nutrient demand in rice. The objective is to develop an optimal N–K application pattern that maximizes dry matter accumulation and translocation, nutrient uptake and utilization, lodging resistance, and grain yield. This work aims to provide a comprehensive theoretical foundation and a refined technical framework for achieving high-yield, resource-efficient, and environmentally sustainable rice production under mechanized cultivation systems.
2 Materials and methods
2.1 Experimental materials
The test variety was F You 498, a hybrid indica rice cultivar with a total growth duration of 155 days. The preceding crop was wheat. Field experiments were conducted during the 2018 and 2019 growing seasons at the Modern Agricultural Science and Technology Park of Sichuan Agricultural University, located in Chongzhou, Chengdu, Sichuan Province (30°42′ N, 103°28′ E). The surface soil layer (0–20 cm) was classified as sandy loam, and baseline soil fertility parameters are shown in Table 1.
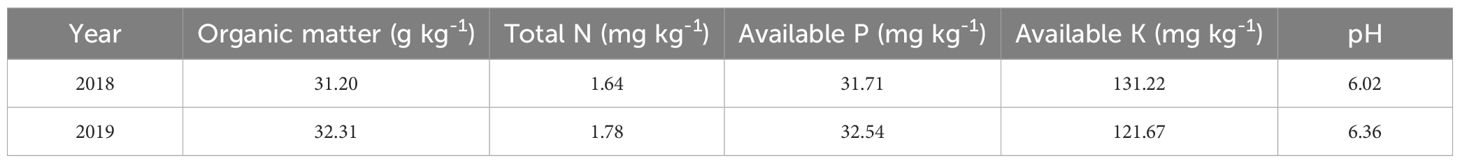
Table 1. Chemical properties of topsoil in the test field from 2018 to 2019.
Meteorological data for the rice growth periods in 2018 and 2019 were obtained from the Sichuan Meteorological Bureau (Figure 1).

Figure 1. Average temperature and rainfall during the rice growth period in the experimental area from 2018 to 2019.
2.2 Experimental design
A split-plot design with two factors was implemented: main plots were assigned to two K application regimes (basal:panicle = 10:0 [K1] and 5:5 [K2]), and subplots to three N splitting regimes (basal:tillering:panicle = 7:3:0 [N1], 5:3:2 [N2], and 3:3:4 [N3]). Fertilizers included urea (N≥46%), calcium superphosphate (P2O5≥12%), and potassium chloride (K2O≥60%). Total N and K inputs were uniformly 150 kg ha-1. Basal N and K were applied pre-transplanting, tillering fertilizer at the 5-leaf stage, and panicle fertilizer split equally at the 4th and 2nd leaf tip emergence stages. Phosphorus (P2O5–75 kg ha-1) was broadcast entirely as basal fertilizer.
Pot-mat seedlings were machine-transplanted on May 16, 2018, and May 20, 2019, at a hill spacing of 30 cm × 18 cm with 2–3 seedlings per hill. The experiment comprised 18 plots (three replicates), each covering 15 m2. A 30-cm-wide and 20-cm-high soil ridge wrapped in plastic film separated the plots to enable independent water and fertilizer management. All other practices followed conventional field management protocols.
2.3 Experimental measurement items and methods
2.3.1 Soil basic fertility determination
Prior to land preparation, soil samples were collected using a five-point sampling method. After natural air-drying and sieving, soil pH, organic matter, total N, available N, available P, and available K were measured using standard analytical methods.
2.3.2 Yield and its components
At maturity, 30 representative hills were selected from each plot to record the number of effective panicles. Five hills per plot were harvested for indoor analysis of spikelet number per panicle, seed-setting rate, and 1,000-grain weight. Grain yield was calculated based on the actual number of hills, excluding border rows.
2.3.3 Aboveground dry matter translocation and accumulation
At rice tillering peak, jointing, heading, and maturity stages, three representative hills from each plot were sampled and separated into stem, leaf, and panicle components. Samples were deactivated at 105°C for 30 min and dried at 80°C to constant weight. The dry weight was recorded and used to calculate the following parameters:
All abbreviations in equations follow conventional agronomic terminology: DW (dry weight), TDM (total dry matter), PDA (post-heading dry matter accumulation), and GDM (grain dry matter) represent biomass measurements. Subscripts specify plant organs (stem–sheath, leaf, panicle) and growth stages (heading, maturity). Units comply with SI standards (t ha-¹ for yield, % for ratios).
2.3.4 Nitrogen and potassium uptake and utilization characteristics
Plant samples from Section 1.3.3 were pulverized through an 80-mesh sieve. After wet digestion with concentrated H2SO4 total N content in organs was determined by micro-Kjeldahl distillation and titration, while total K content was measured via flame photometry. N and K uptake and utilization efficiencies were defined according to Wu et al. (2008) as follows:
All equations follow standard agronomic conventions with clear subscript notation: DW (dry weight), TDM (total dry matter), and N(K) (nitrogen/potassium content) are key parameters. Subscripts indicate plant organs (stem–sheath, leaf, panicle) and growth stages (heading, maturity).
2.3.5 Lodging resistance determination
At 20 days post-heading, five uniform plants per plot were selected. For each plant, three stems were measured for internode lengths (S2, S3, S4), fresh weights, and breaking strength of internodes. Breaking force was tested using a 0.001 N digital dynamometer with a 5-cm spacing between support points (Lei et al., 2013; Yang et al., 2013). After testing, stem diameters and wall thickness were measured with a vernier caliper.
2.4 Statistical analysis
Microsoft Excel 2016, SPSS 25.0 (SPSS Institute Inc., Chicago, USA), and Origin Pro 2020 (OriginLab, Northampton, MA, USA) were used for data analysis and visualization. Differences among treatments were tested using the least significant difference method at P < 0.05.
3 Results and analysis
3.1 Yield and its components
Year, K application, and N application rate significantly or highly significantly affected the number of effective panicles, grains per panicle, and grain yield (Table 2). The year × K interaction significantly influenced the number of effective panicles, while the year × N interaction significantly affected grains per panicle. The K × N interaction highly significantly impacted the number of effective panicles and grain yield.
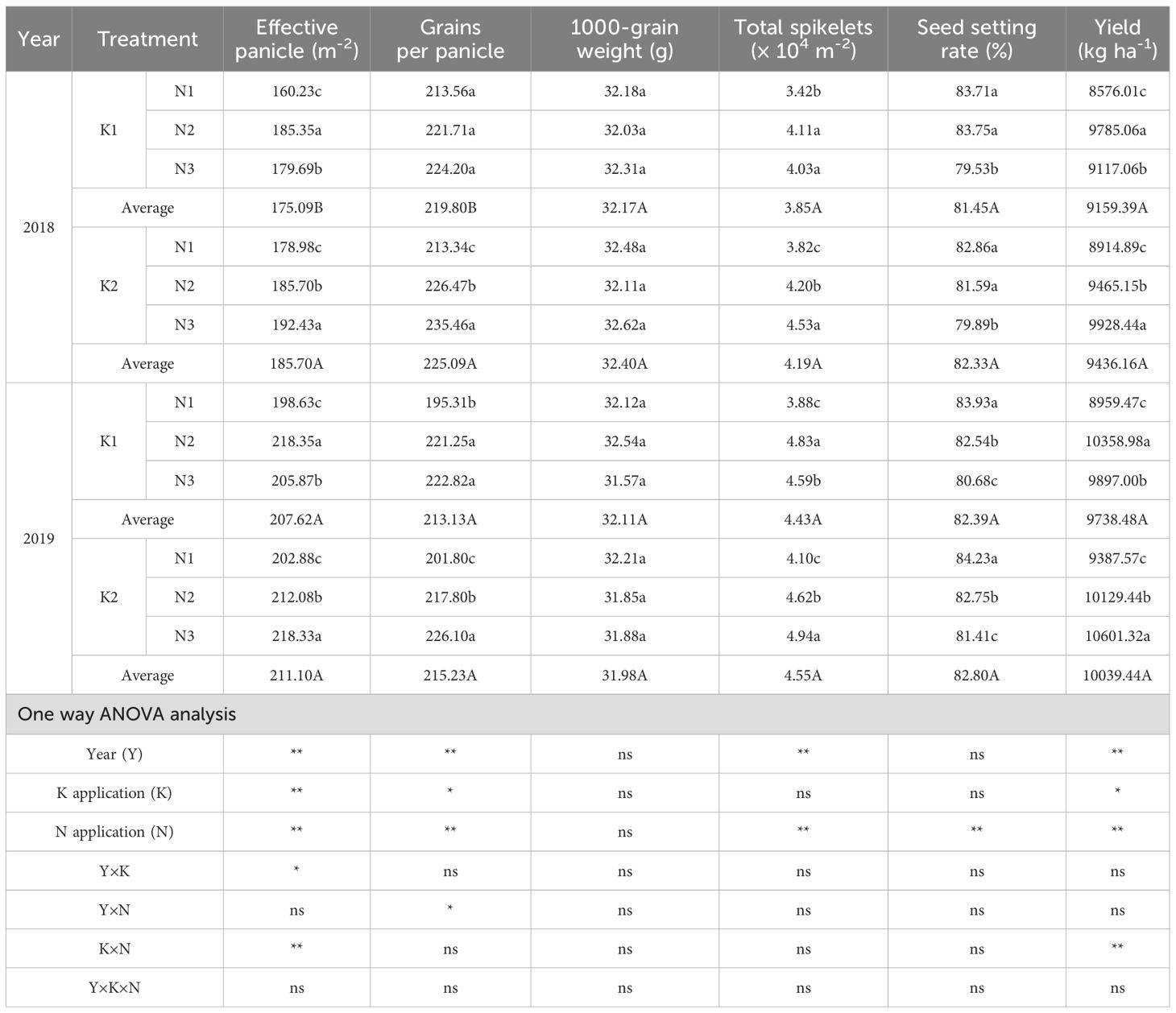
Table 2. Effects of N-K ratio on yield and yield components of mechanically inserted rice.
Across K treatments, grain yield was higher in K2 than in K1, with an average increase of 3.06% over two years. Under K2, yield increased progressively with panicle N application (N3 > N2 > N1), showing significant differences: N3 increased yield by 12.17% compared with N1 and by 4.77% compared with N2.
Regarding yield components, the number of effective panicles, grains per panicle, total spikelets, and grain-filling percentage were generally higher in K2 than in K1, though most differences were not statistically significant. Under K2, the number of effective panicles, grains per panicle, and total spikelets peaked in N3, significantly exceeding N1 by 7.57%, 11.18%, and 19.57%, and N2 by 3.26%, 3.89%, and 7.37%, respectively. Grain-filling percentage showed a declining trend, with relatively significant differences among treatments.
Different small letters after the data in the same column indicate significant differences between treatments under the same potash treatment and the same year, and the capital letters after the average data indicate significant differences between K1 and K2 (P<0.05) * and ** indicate significant effects at the 0.05 and 0.01 probability levels, respectively, and ns indicates no significant effect.
3.2 Dry matter translocation and accumulation
Year and K application highly significantly affected post-heading dry matter translocation (Table 3). N application rate and the K × N interaction significantly or highly significantly influenced total dry matter accumulation and post-heading translocation. The three-way interaction (year × K × N) significantly affected the contribution rate of post-heading translocation.

Table 3. Effects of N-K management strategies on dry matter accumulation and translocation in machine-transplanted rice.
Across K treatments, total dry matter accumulation at various growth stages mostly showed nonsignificant differences. Post-heading translocation was higher in K2 than in K1, with mostly significant differences among treatments. On average, K2 increased translocation proportion and contribution rate by 5.29% and 7.64%, respectively, compared with K1 over two years. Under K2, total dry matter accumulation at all growth stages peaked in N3, with significant differences observed. At maturity, N3 increased accumulation by 19.02% compared with N1 and by 9.59% compared with N2. Post-heading translocation also peaked in N3, exceeding N1 in translocation amount, proportion, and contribution rate by 25.54%, 5.37%, and 7.42%, and N2 by 12.68%, 2.76%, and 2.57%, respectively.
3.3 Assimilate translocation from vegetative organs
Year, N application rate, and the interactions of year with K or N application significantly or highly significantly affected stem–sheath dry matter output rate, stem–sheath dry matter conversion rate, leaf dry matter output rate, and leaf dry matter conversion rate (Table 4). K application highly significantly influenced stem–sheath dry matter output rate and stem–sheath dry matter conversion rate. The three-way interaction (year × K × N) highly significantly affected stem–sheath dry matter output rate and stem–sheath dry matter conversion rate, and significantly affected leaf dry matter output rate.
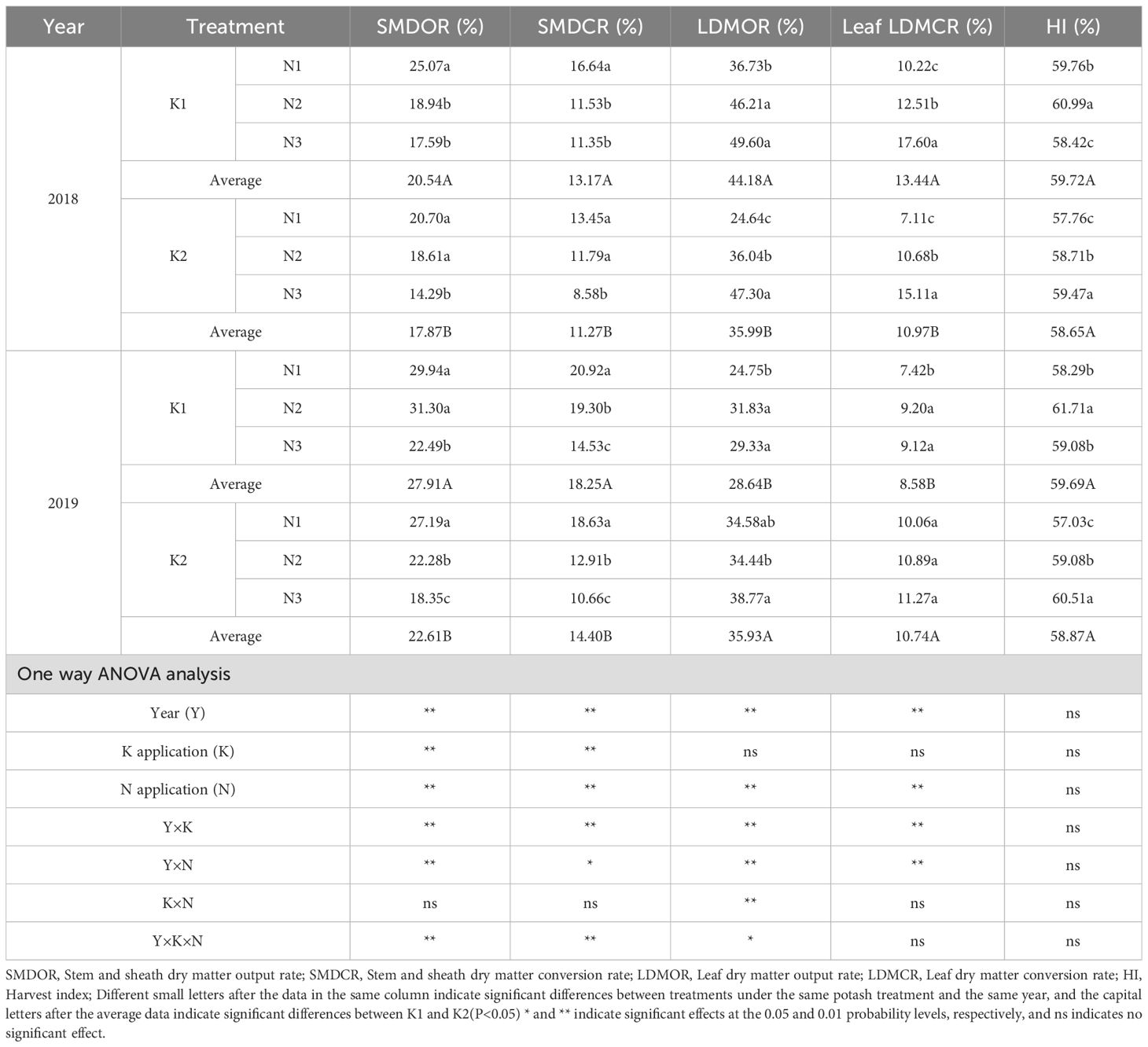
Table 4. Effects of N-K management strategies on assimilating translocation from vegetative organs in machine-transplanted rice.
Across K treatments, stem–sheath dry matter output rate, stem–sheath dry matter conversion rate, and harvest index were higher in K1 than in K2. On average, K1 increased these indices by 19.69%, 22.40%, and 1.61%, respectively, compared with K2 across two years. Under K2, stem–sheath dry matter output rate and conversion rate were highest in N1 (N1 > N2 > N3). Conversely, leaf dry matter output rate, leaf dry matter conversion rate, and harvest index peaked in N3, with mostly significant differences. N3 increased leaf dry matter output rate, leaf dry matter conversion rate, and harvest index by 45.34%, 53.64%, and 4.52% compared with N1, and by 22.12%, 22.30%, and 1.86% compared with N2.
3.4 Nitrogen accumulation in organs
Year highly significantly affected N accumulation at the jointing and heading stages (Table 5). K application significantly influenced N accumulation at peak tillering, heading, and maturity. N application rate highly significantly affected all indices except N accumulation at peak tillering. The K × N interaction significantly or highly significantly impacted N accumulation at all stages and the N harvest index. The year × N interaction and the three-way interaction (year × K × N) highly significantly affected N accumulation at jointing.
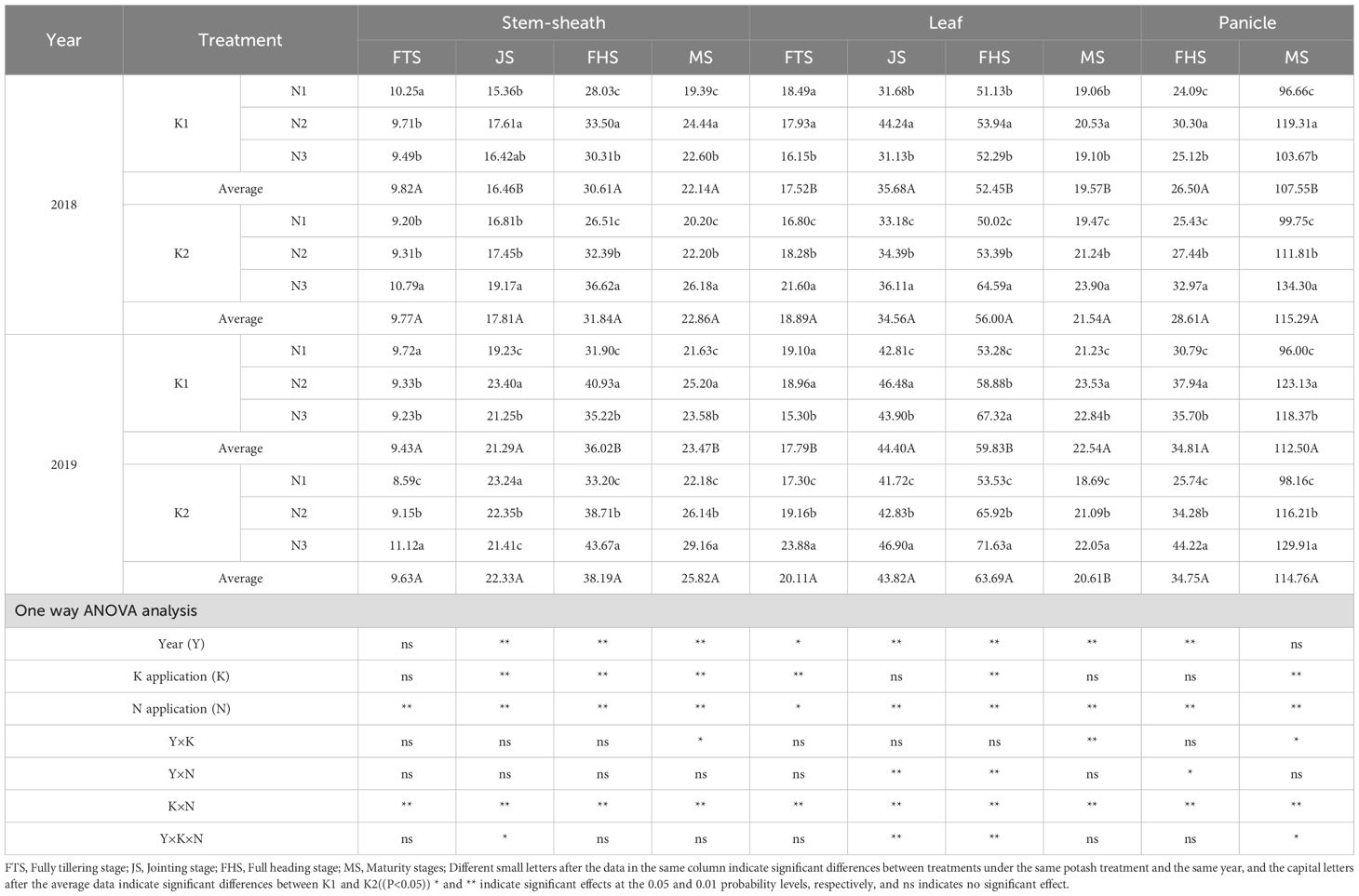
Table 5. Effects of different N-K management strategies on nitrogen accumulation in organs of machine-transplanted rice.
Across K treatments, plant N accumulation during growth stages was higher in K2 than in K1, although differences were nonsignificant. At maturity, K2 averaged a 4.11% increase compared with K1. N productivity (dry matter basis) was higher in K1, while the N harvest index was higher in K2, though both differences were nonsignificant. Under K2, N accumulation increased with N application (N3 > N2 > N1), showing significant differences: at heading and maturity, N3 increased accumulation by 36.59% and 31.27% compared with N1, and by 16.16% and 14.69% compared with N2. Grain N productivity decreased with increasing N application (N1 > N2 > N3), with significant differences. The N harvest index peaked in N3, significantly exceeding N1 by 1.70% and N2 by 1.01%.
3.5 Nitrogen translocation in vegetative organs
Year highly significantly affected all N export parameters in machine-transplanted rice (Table 6). K application, N application rate, and their interaction highly significantly influenced N export amount and increased grain N. The year × K and year × N interactions significantly or highly significantly affected N export amount and the export contribution rate. The three-way interaction (year × K × N) significantly or highly significantly impacted export contribution rate and increased grain N.
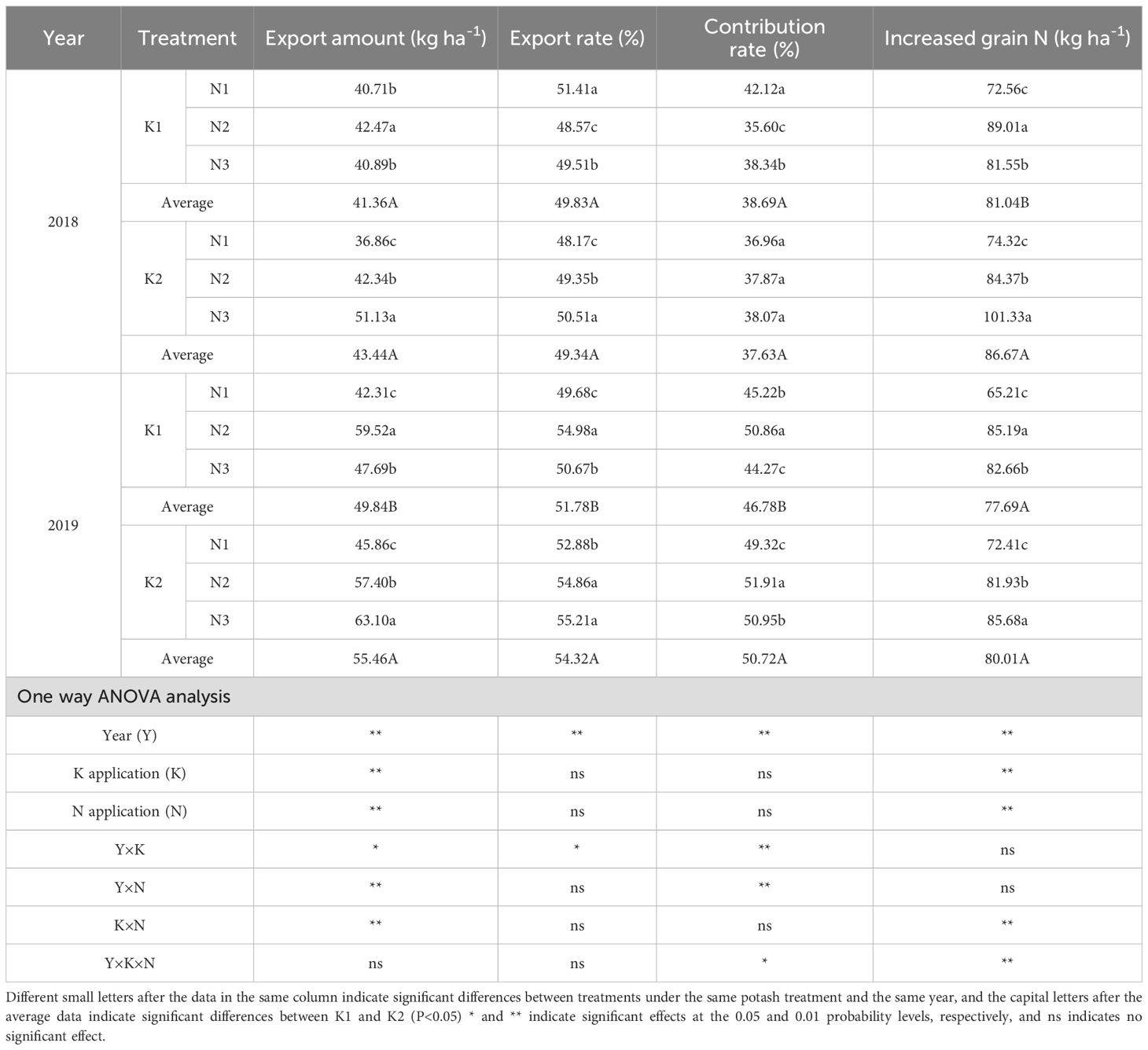
Table 6. Effects of N-K management strategies on nitrogen translocation in vegetative organs of machine-transplanted rice.
Across K treatments, most N export parameters were higher in K2 than in K1. On average, N export amount and increased grain N rose by 8.44% and 5.01%, respectively, compared with K1 over two years. Under K2, N export amount, export rate, and increased grain N increased with higher panicle N application, peaking in N3 (N3 > N2 > N1). N3 increased these parameters by 38.09%, 4.62%, and 27.45% compared with N1, and by 14.53%, 1.45%, and 12.45% compared with N2. The N export contribution rate showed inconsistent trends over the years.
3.6 Potassium uptake and utilization
Year highly significantly affected total K accumulation, K dry matter production efficiency, and K harvest index at late growth stages (Table 7). K application highly significantly influenced total K accumulation at maturity and K harvest index. N application significantly or highly significantly impacted total K accumulation at all stages and K dry matter production efficiency. The year × K and year × N interactions significantly or highly significantly affected the K harvest index. The K × N interaction highly significantly influenced total K accumulation at all stages. The three-way interaction (year × K × N) highly significantly affected K grain production efficiency.

Table 7. Effects of N-K management strategies on potassium uptake and utilization in machine-transplanted rice.
Across K treatments, total K accumulation during late growth stages was higher in K2 than in K1, averaging a 3.46% increase at maturity. K dry matter production efficiency was highest in K1, though differences among treatments were nonsignificant. The K harvest index peaked in K2, averaging an 8.49% increase compared with K1. Under K2, total K accumulation peaked in N3, differing significantly from N1 and N2: at heading and maturity, N3 increased accumulation by 21.82% and 26.94% compared with N1, and by 8.83% and 11.90% compared with N2. K dry matter production efficiency peaked in N1 with significant differences, while the K harvest index showed inconsistent trends between years.
3.7 Potassium translocation
Year and K application highly significantly affected K export amount, export rate, and increased grain K in machine-transplanted rice (Table 8). N application rate, the year × K interaction, the year × N interaction, and the K × N interaction significantly or highly significantly influenced all K export parameters.
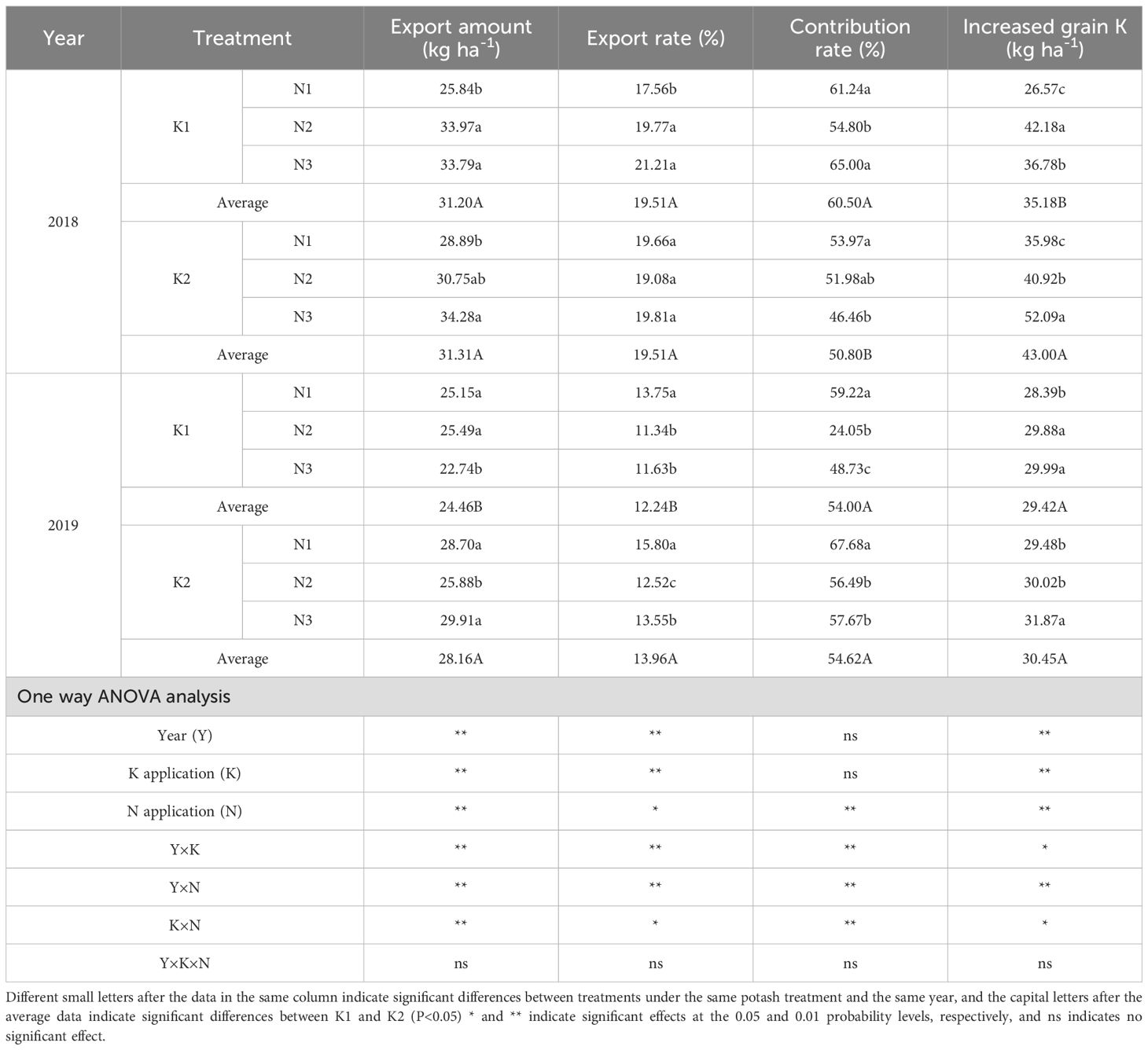
Table 8. Effects of N-K management strategies on potassium translocation in machine-transplanted rice.
Across K treatments, K export amount and increased grain K were higher in K2 than in K1, averaging increases of 6.85% and 13.70%, respectively, compared with K1 across two years. Under K2, K export amount and increased grain K increased with higher panicle N application, peaking in N3. N3 increased these parameters by 11.46% and 28.26% compared with N1, and by 13.35% and 18.35% compared with N2. K export rate and contribution rate showed inconsistent trends between years.
3.8 Culm physical traits
Year significantly or highly significantly affected internode length, wall thickness, and plant height in machine-transplanted rice (Table 9). K application highly significantly influenced the wall thickness of S2 and S3 internodes. N application and the year × K interaction significantly or highly significantly affected wall thickness. The K × N interaction significantly or highly significantly impacted internode length and wall thickness. The three-way interaction (year × K × N) highly significantly affected S4 internode length and the wall thickness of S2 and S4 internodes.
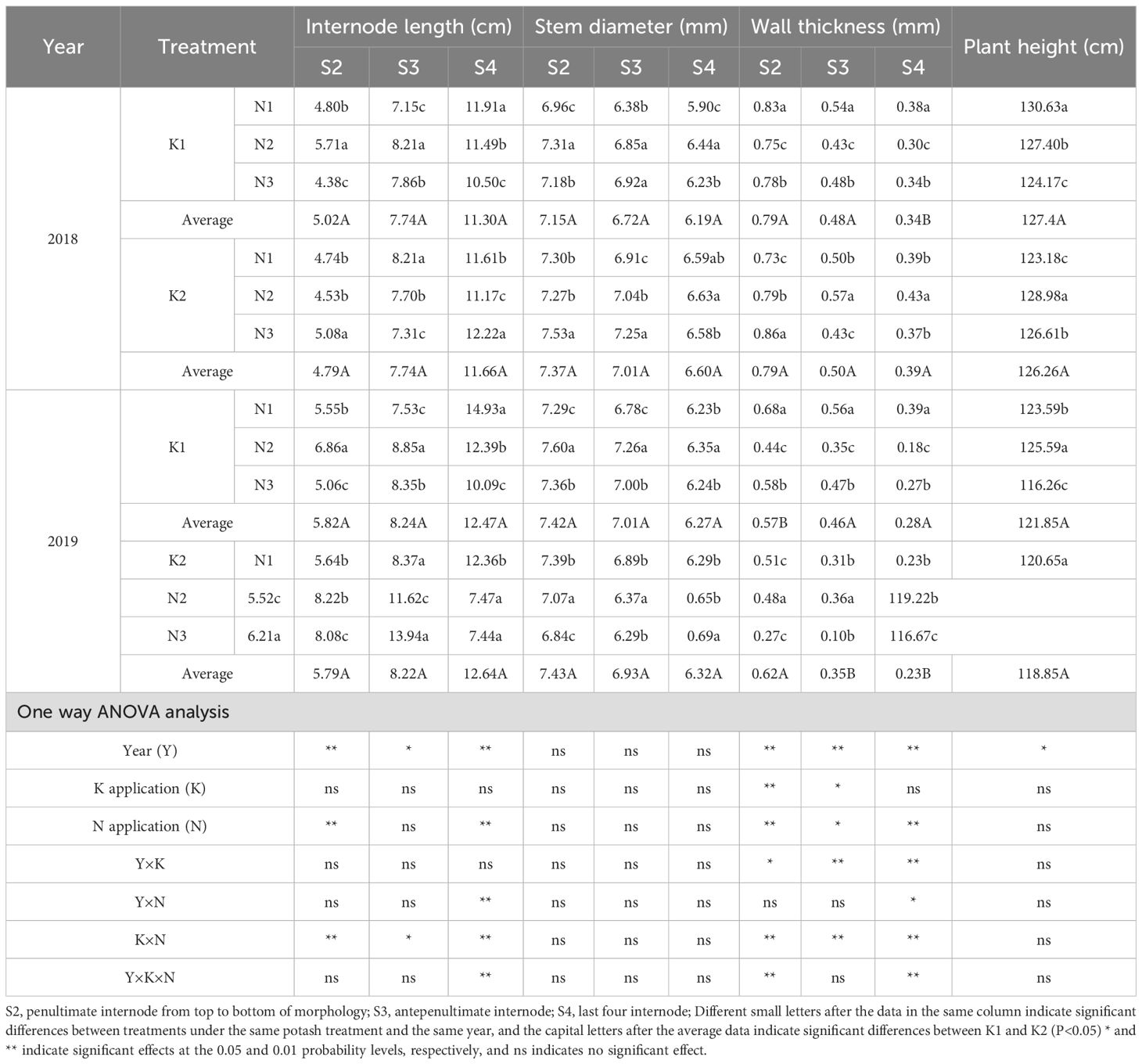
Table 9. Effects of N-K management strategies on culm physical traits in machine-transplanted rice.
Across K treatments, S2 internode length and S4 stem diameter were higher in K2 than in K1, although differences were nonsignificant; plant height was higher in K1 than in K2, also with nonsignificant differences. Under K2, S2 and S4 internode lengths peaked in N3, while S3 internode length peaked in N1. S2 stem diameter peaked in N3. S2 wall thickness peaked in N3, while S3 and S4 wall thickness peaked in N2, all with significant differences.
3.9 Lodging index
Year and K application highly significantly affected internode resistance and breaking moment of all internodes (Table 10). N application significantly or highly significantly influenced all parameters except the S4 bending moment. The year × K interaction significantly or highly significantly impacted the S2 and S3 bending moments and the breaking moment. The K × N interaction significantly or highly significantly affected the S2 and S3 bending moments and the S4 breaking moment.
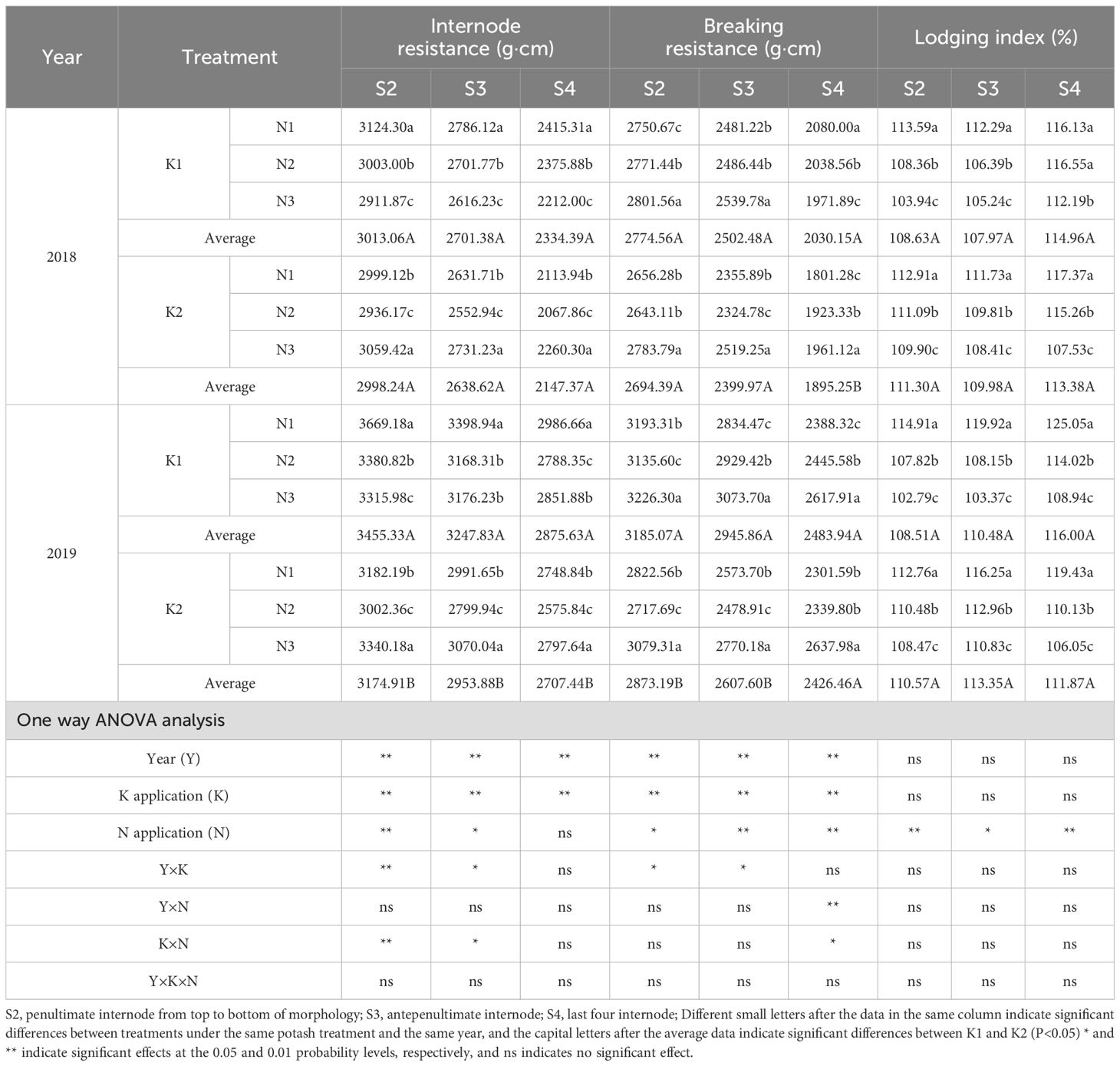
Table 10. Effects of N-K management strategies on lodging index in machine-transplanted rice.
Across K treatments, internode resistance and breaking moment were higher in K1 than in K2. The lodging index of S2 and S3 was higher in K2 than in K1, while that of S4 was higher in K1 than in K2; all differences were nonsignificant. Under K2, internode resistance followed the trend N3 > N1 > N2, with significant differences: N3 increased resistance by 3.53% (S2), 3.16% (S3), and 4.01% (S4) compared with N1, and by 7.76%, 8.38%, and 8.92% compared with N2. The breaking moment of all internodes peaked in N3, with significant differences. The lodging index showed the trend N1 > N2 > N3, with significant differences.
3.10 Correlation analysis
Yield showed significant positive correlations with total dry matter accumulation, N accumulation, the N harvest index (NHI), and K accumulation, but significant negative correlations with rice grain N productivity and rice grain K productivity. Both dry matter N productivity and rice grain N productivity, as well as dry matter K productivity and rice grain K productivity, were significantly positively correlated with the S4 lodging index. Additionally, the S2 lodging index was significantly positively correlated with the S3 lodging index.
4 Discussion
4.1 Effects of N and K ratios on yield and yield components in mechanical transplanting rice
Nitrogen and potassium, as essential macronutrients for rice growth, critically influence yield and its components. Our results demonstrate that under one-time basal potassium application (K1), moderate panicle nitrogen (N2) significantly increased the number of productive panicles and total spikelets by coordinating nutrient allocation between tillering and panicle initiation stages. This effect is attributed to optimized dry matter accumulation from peak tillering to heading under N2, which enhanced photosynthate translocation to panicles (Li et al., 2025). However, split potassium application (K2) altered nitrogen response mechanisms. Under post-anthesis potassium postponement, high panicle nitrogen application (N3) synergistically enhanced yield through three integrated physiological mechanisms: (i) delayed potassium application precisely matched the peak potassium demand during panicle differentiation, activating nitrate reductase and pyruvate kinase activity to promote photoassimilate translocation to panicles, resulting in a 25.54% increase in post-heading dry matter remobilization (Shova et al., 2023); (ii) increased panicle N application delayed root senescence during grain filling and improved N assimilation efficiency, elevating spikelets per panicle by 11.18%; and (iii) split potassium application regulated cell turgor pressure to optimize vascular bundle development, effectively mitigating canopy overcrowding induced by high nitrogen inputs (Islam and Muttaleb, 2016). Insufficient spikelet number constitutes a primary yield-limiting factor in machine-transplanted rice. Potassium supplementation effectively optimizes nutrient supply and physiological conditions during this critical phase, promoting spikelet differentiation while reducing degeneration, thereby significantly increasing grains per panicle (Zhang et al., 2010). Concurrently, enhanced panicle nitrogen synergized with delayed potassium application to boost dry matter accumulation and translocation efficiency during grain filling, ultimately enlarging sink capacity and strengthening source flow for substantial yield improvement.
Regarding yield components under different N–K management strategies, the K2N3 regime exhibited superior performance in productive panicles, spikelets per panicle, and total spikelets. This superiority likely stemmed from improved root uptake of water and nutrients by panicle-stage N–K application, which enhanced tillering capacity and plant vigor. Furthermore, nitrogen application prolonged the active grain-filling phase (Jiang et al., 2022), while potassium regulated filling rate to synergistically increase grain weight (Liu et al., 2024), explaining the yield advantage of K2N3 observed herein. Nevertheless, this combination reduced the seed-setting rate, potentially due to excessive vegetative growth from increased nitrogen proportion at the booting stage, which disrupted nutrient allocation during anthesis and impaired pollination. This observation corroborates the conclusion that N–K imbalance may restrict grain-filling completion through sink–source limitations (Chen et al., 2024). Simultaneously, our analysis demonstrates that optimized N–K stoichiometry significantly enhances economic returns through yield improvement, particularly in the K2N3 treatment, which could effectively increase farmers’ income in local agricultural systems.
4.2 Effects of N–K ratios on nutrient uptake and translocation in mechanical transplanting rice
Nitrogen uptake and translocation efficiency constitute the core physiological basis for rice yield formation. Studies indicate that potassium activates nitrate reductase activity, enhancing nitrogen assimilation and partitioning to panicles (Gao et al., 2019) while reducing nitrogen retention in stems and sheaths. Topdressing 50 kg ha-1 K at the jointing stage significantly increased plant nitrogen accumulation at flowering, grain nitrogen accumulation at maturity, and the nitrogen translocation contribution rate (Guo et al., 2016). The split application of potassium fertilizer further enhanced nitrogen translocation efficiency by 38.09% through activation of H+-ATPase pumps that drive nitrate transporters. The underlying physiological mechanism involves potassium serving as an activator of nitrate reductase, thereby promoting the allocation of nitrogen assimilation products to panicles while reducing nitrogen retention in stems and leaf sheaths (Vijayakumar et al., 2022). When 50 kg ha-¹ of potassium was top-dressed at the jointing stage, the treatment significantly increased plant nitrogen accumulation at flowering, grain nitrogen accumulation at maturity, and nitrogen use efficiency (Wang et al., 2025). Concurrently, machine-transplanted rice exhibits significantly elevated nitrogen uptake from heading to maturity. Appropriately increasing the proportion of panicle nitrogen promotes root vitality, facilitating more efficient soil nitrogen absorption and accumulation (Hu et al., 2018; Zheng et al., 2023). Panicle nitrogen application precisely matches the critical period from panicle initiation to grain filling, enhancing shoot nitrogen accumulation and translocation capacity from stems and sheaths to leaves, while increasing nitrogen recovery efficiency by 7.27%–26.06% (Zhang et al., 2022). Thus, split potassium application potentiates nitrogen metabolism, fully unlocking the yield potential of panicle nitrogen to synergistically achieve efficient nitrogen supply for grain sink filling.
The panicle initiation to heading phase represents the peak potassium uptake window, where potassium supply efficacy directly governs subsequent K translocation to grains (Yu et al., 2012). Under the K2N3 regime, potassium accumulation in mature plants increased by 18%–25%. The molecular mechanism involves panicle nitrogen application activating the expression of potassium channel proteins and transporters, which enhances root potassium uptake capacity while optimizing vascular bundle potassium translocation efficiency (Vijayakumar et al., 2024). This finding is corroborated by studies in Iranian rice systems, which demonstrated that in water-scarce regions, application of 120 kg ha-1 nitrogen combined with 80 kg ha-1 potassium comprehensively improved rice osmoregulation capacity, protected photosynthetic organs, and ultimately enhanced drought resistance while maintaining grain yield (Rabiei et al., 2021). However, excessive potassium application can induce potassium retention in vegetative tissues, inhibiting redistribution to grains. Optimal K application rates have been reported at 108–120 kg ha-1 (Hou et al., 2018). Notably, panicle nitrogen systematically enhances fertilizer use efficiency and significantly reduces agricultural greenhouse gas emissions (Poudel et al., 2025) by activating root uptake capacity, strengthening vascular bundle transport efficiency, and optimizing inter-organ allocation strategies. Therefore, by synchronizing nutrient supply with uptake peaks and improving redistribution, K2 synergistically interacts with N3 to systematically enhance productivity, thereby establishing the physiological foundation for efficient potassium accumulation in grains and yield improvement.
4.3 Effects of N–K ratios on lodging resistance in mechanical transplanting rice
Machine-transplanted rice exhibits significantly weaker lodging resistance than manually transplanted rice due to inferior stem solidity and root anchorage, necessitating precise N–K co-regulation to synergistically enhance morphological and mechanical traits (Sun et al., 2017). This study is the first to elucidate the mechanism at the level of cell wall construction: under split potassium application (K2), the wall thickness and bending resistance of the S2 internode significantly increased. This improvement is attributed to potassium ions activating the phenylpropanoid pathway, which promotes lignin monomer synthesis and cell wall cross-linking (Ding et al., 2021). Under the K2N3 treatment, the wall thickness and stem diameter of both S2 and S4 internodes increased while shorter basal internodes, directly improving stem bending resistance. At the mechanical level, this combination significantly enhanced bending moment resistance by increasing cellulose deposition and vascular bundle development, thereby minimizing the lodging index. This finding is consistent with Zhang et al. (2021), who reported that “increased potassium application significantly reduces the lodging index by increasing lignin content or cell wall thickness in rice stems.” Moreover, split nitrogen application optimized the enrichment of photosynthetic products and nitrogen in the basal stems, supporting early-stage population establishment while avoiding stem weakness caused by late-stage nitrogen excess, thereby preventing yield losses due to lodging stress (Lu et al., 2025). However, excessive basal nitrogen application (e.g., K1N1) induced gibberellin synthesis, which promoted internode elongation and diluted cell wall material concentration, ultimately reducing mechanical strength.
As essential macronutrients for plants, adequate nitrogen and potassium directly promote the accumulation of photosynthetic products and assimilate translocation, laying the material foundation for high yields. However, under high-yield conditions, grain output efficiency per unit nutrient decreases, which may stem from the “yield–efficiency trade-off” effect, whereby nutrient use efficiency tends to decline as yields approach higher levels. Regarding stem lodging resistance characteristics, the lodging indices of S2 and S3 internodes showed a significant positive correlation (Figure 2), possibly because vascular bundle development in basal internodes is continuous. When the cell wall thickness of the S2 internode decreases, the resulting reduction in bending resistance increases the mechanical load on upper internodes (Yuan et al., 2025). Notably, the dry matter productivity of nitrogen and potassium showed a significant positive correlation with the lodging index of the S4 internode. This result is the first to reveal the intrinsic relationship between nutrient use efficiency and stem mechanical properties. We speculate that the mechanism may involve plants preferentially allocating more resources to grains rather than to the synthesis of stem structural components (e.g., cellulose and lignin) when nitrogen and potassium use efficiency is excessively high, thereby reducing stem mechanical strength. These findings provide new evidence supporting the “yield–lodging resistance synergy” theory (Zhang et al., 2025).
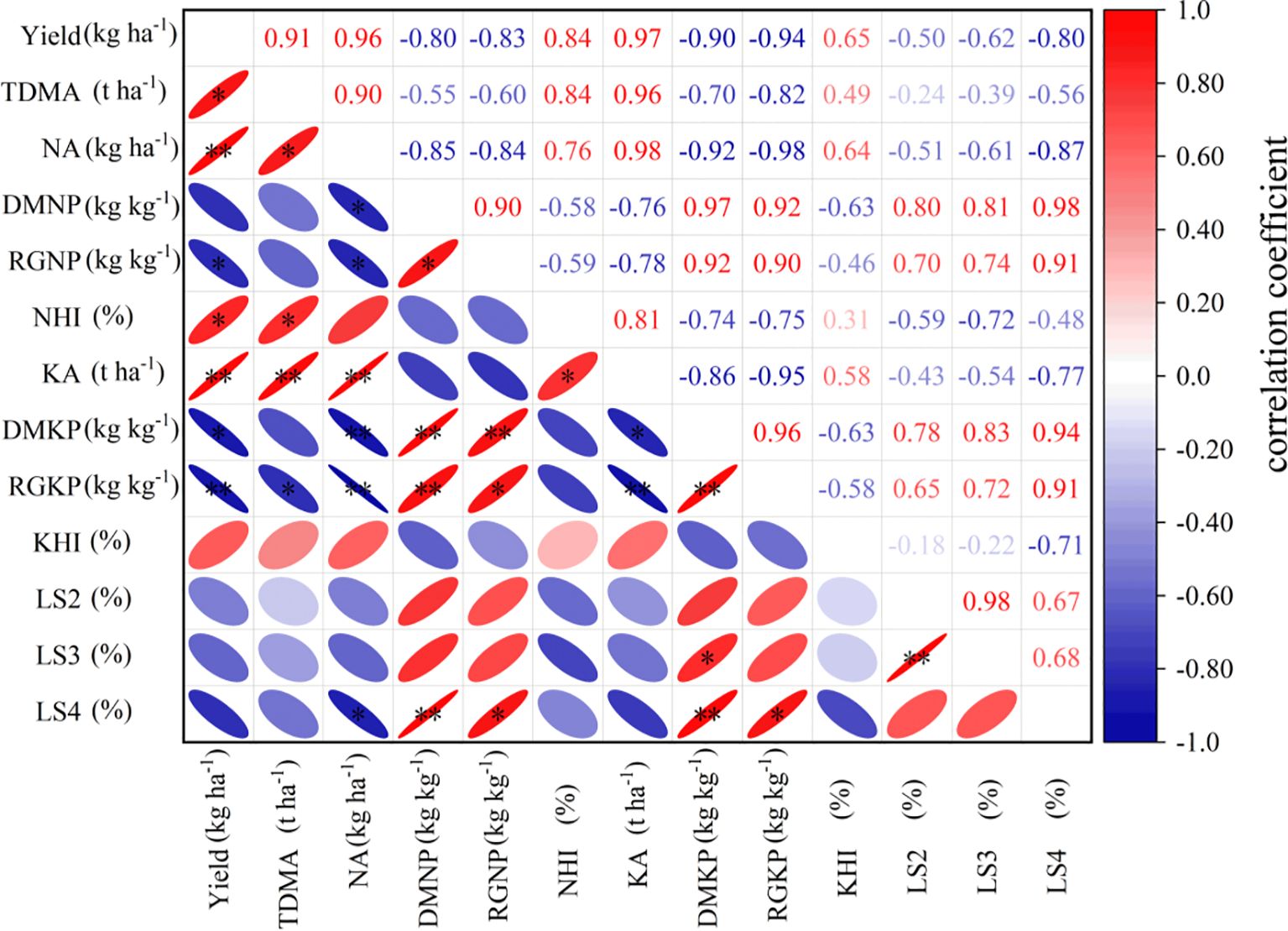
Figure 2. Correlation analysis FHS - Full heading stage, MS - Maturity stages; S2 - penultimate internode from top to bottom of morphology; S3 - antepenultimate internode; S4 – last four internode; Different small letters after the data in the same column indicate significant differences between treatments under the same potash treatment and the same year, and the capital letters after the average data indicate significant differences between K1 and K2 (P<0.05) * and ** indicate significant effects at the 0.05 and 0.01 probability levels, respectively, and ns indicates no significant effect.
Although this study systematically analyzed the regulatory mechanisms of nitrogen–potassium coordination on yield formation and stress resistance in machine-transplanted rice, some limitations remain. First, the conclusions are based on short-term observations of specific indica rice varieties, and their applicability to japonica ecotypes and long-term cultivation requires further validation. Second, although the improvement in stem lodging resistance is associated with potassium management, key evidence on cell wall component reconstruction (e.g., spatial deposition patterns of lignin) remains insufficient. Third, the environmental risks under high panicle fertilizer application were not assessed. Future research should combine long-term, multi-ecological field trials to deepen the synergistic analysis of post-anthesis nutrient translocation barriers and stem microstructure development, and establish an environmental footprint accounting system to improve the sustainable application of nitrogen–potassium management in machine-transplanted rice.
5 Conclusions
Compared with one-time basal potassium application (K1), split potassium application (K2) increased rice yield by optimizing panicle traits, enhancing post-heading dry matter translocation efficiency, and strengthening lodging resistance. When K2 was combined with high panicle nitrogen (N3), the strategy demonstrated amplified effects: it significantly promoted post-heading dry matter accumulation and translocation, improved targeted nitrogen–potassium partitioning to panicles, and elevated grain yield by 4.77%–12.17% compared with conventional nitrogen regimes. Concurrently, this combination reduced lodging risk through increased internode diameter and wall thickness. Collectively, the cultural strategy that integrates delayed potassium with high panicle nitrogen provides an effective agronomic solution for lodging-resistant yield enhancement in machine-transplanted rice by coordinating post-flowering assimilate allocation, strengthening panicle nutrient accumulation, and reinforcing stem mechanical strength.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
ZL: Data curation, Writing – original draft. MH: Conceptualization, Writing – original draft. TL: Investigation, Writing – original draft. YT: Writing – original draft, Investigation. RL: Investigation, Writing – original draft. ZBP: Investigation, Writing – original draft. CW: Investigation, Writing – original draft. ZLP: Writing – original draft, Investigation. ZW: Writing – review & editing, Conceptualization. ZC: Writing – review & editing, Investigation. ZY: Investigation, Writing – review & editing. YS: Investigation, Writing – review & editing. JM: Conceptualization, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the National Key Research and Development Program Project (Grant No. 2017YFD0301700).
Acknowledgments
We would like to thank our teacher for carefully reading and correcting our manuscript and providing technical assistance and financial support for the study, as well as our scientific research team for their contribution to this paper.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Begum, S., Islam, M. S., Rashid, M. M., Manir, M. R., Rahman, M. H., and Hossen, M. A. (2022). Efficacy of mechanical seedling transplanter and deep placement of mixed fertilizer on rice yield. Asian J. Adv. Agric. Res. 20, 45–51. doi: 10.9734/AJAAR/2022/V20I2395
Bhatt, P. S., Reddy, P. R. R., Varma, N. R. G., Babu, T. K., and Kumar, P. (2023). Studies of farmer participatory on machine transplanting of rice. Int. J. Environ. Climate Change 13, 752–755. doi: 10.9734/IJECC/2023/V13I123737
Chen, G., Duan, Q., Wu, C., He, X., Hu, M., Li, C., et al. (2024). Optimizing rice yield, quality and nutrient use efficiency through combined application of nitrogen and potassium. Front. Plant Sci. 15. doi: 10.3389/FPLS.2024.1335744, PMID: 38529059
Chen, S., Liu, S., Zheng, X., Yin, M., Chu, G., Xu, C., et al. (2018). Effect of various crop rotations on rice yield and nitrogen use efficiency in paddy–upland systems in southeastern China. Crop J. 6, 576–588. doi: 10.1016/j.cj.2018.07.007
Deng, J., Liu, K., Xiong, X., Hussain, T., Huang, L., de Voil, P., et al. (2024). Achieving sustainable rice production through nitrogen-potassium harmony for enhanced economic and environmental gains. Agric. Water Manage. 301, 108949. doi: 10.1016/J.AGWAT.2024.108949
Ding, C., Luo, X., Wu, Q., Lu, B., Ding, Y., Wang, S., et al. (2021). Compact plant type rice has higher lodging and N resistance under machine transplanting. J. Integr. Agric. 20, 65–77. doi: 10.1016/S2095-3119(20)63229-4
Gao, Z. Y., Wang, Y. F., Chen, G., Zhang, A. P., and Yang, S. L. (2019). The indica nitrate reductase gene OsNR2 allele enhances rice yield potential and nitrogen use efficiency. Nat. Commun. 10, 5207. doi: 10.1038/s41467-019-13110-8, PMID: 31729387
Gao, Z., Ye, T., Cui, X., Lu, J., Ren, T., Cong, R., et al. (2025). Dynamics of potassium concentration in paddy field water, soil and plant affected by potassium fertilizer levels. Nutrient Cycling Agroecosystems 130, 1–14. doi: 10.1007/S10705-025-10396-7
Guo, M. M., Zhao, G. C., Guo, W. S., Chang, X. H., Wang, D. M., Yang, Y. S., et al. (2016). Regulation of nitrogen topdressing stage and potassium fertilizer rate on absorption and translocation of nitrogen by wheat. J. Plant Nutr. Fertilizer Sci. 22, 590–597. doi: 10.11674/zwyf.15089
Hou, Y. P., Yang, J., Kong, L. L., Li, Q., and Wang, L. C. (2018). Response of nutrient absorption, translocation, and yield of rice to potassium fertilizer application. J. Jilin Agric. Univ. 40, 17–24. doi: 10.13327/j.jjlau.2017.3542
Hu, Y. J., Wu, P., Zhu, M., Xing, Z. P., Dai, Q. G., Huo, Z. Y., et al. (2018). Characteristics of nitrogen uptake and utilization of mechanically-transplanted pot-tray-nursed rice seedlings. Chin. J. Rice Sci. 32, 257–264. doi: 10.16819/j.1001-7216.2018.7086
Islam, M. and Muttaleb (2016). Effect of potassium fertilization on yield and potassium nutrition of Boro rice in a wetland ecosystem of Bangladesh. Arch. Agron. Soil Sci. 62, 1530–1540. doi: 10.1080/03650340.2016.1157259
Jiang, M. J., Xu, W. B., Li, L. J., Zhang, J. F., Wang, R. J., Ji, G. M., et al. (2024). Balanced nitrogen reduction for improved grain yield and eating quality in mechanically transplanted hybrid indica rice. Agriculture 14, 1313–1313. doi: 10.3390/AGRICULTURE14081313
Jiang, P., Zhou, X., Zhang, L., Liu, M., Xiong, H., Guo, X., et al. (2022). Delaying application and reducing the N rate enhances grain yield and N use efficiency in no-tillage, direct-seeded hybrid rice. Agronomy 12, 2092. doi: 10.3390/AGRONOMY12092092
Jinjala, V. R., Gudadhe, N. N., Tripathi, S., Chudasama, S. D., and Ninama, A. R. (2024). Influence of iron and zinc biofortification on growth and yield of drilled rice (Oryza sativa L.). Asian J. Soil Sci. Plant Nutr. 10, 141–156. doi: 10.9734/AJSSPN/2024/V10I4390
Lei, X. L., Liu, L., Gou, W., and Ma, C. R. (2013). Effects of planting methods on the lodging resistance of hybrid indica rice plants. Acta Agronomica Sin. 39, 1814–1825. doi: 10.3724/SP.J.1006.2013.01814
Lei, X., Wu, Y., Wang, J., Tao, J., Wan, C., Wang, M., et al. (2025). Effects of planting density and NPK-fertilization level on lodging resistance and grain yield of common buckwheat. Field Crops Res. 322, 109738. doi: 10.1016/J.FCR.2025.109738
Li, Z. X., Hu, M. M., Tang, Y., Wang, C., Wang, Z. L., Liu, T., et al. (2025). Suitable partition ratios of nitrogen and potassium fertilizers increase the accumulation and translocation of dry matter and nutrients and the yield of mechanically direct-seeded rice. J. Plant Nutr. Fertilizers 31, 267–281. doi: 10.11674/zwyf.2024355
Liu, A., Ye, T., Zhang, Y., Liao, S., and Li, X. (2024). Effects of potassium fertilization on grain yield, taste quality, and starch characteristics of rice (Oryza sativa L.) grain. J. Sci. Food Agric. 105, 3036–3044. doi: 10.1002/JSFA.14096, PMID: 39723469
Lu, Y., Cui, J., Bao, S., Liu, W., Geng, Y., Liang, X., et al. (2025). Effects of nitrogen fertilizer application rate on lodging resistance for rice (Oryza sativa L.) stem. Sci. Rep. 15, 2149. doi: 10.1038/S41598-025-85641-8, PMID: 39820090
Mohapatra, S., Tripathy, S. K., Mohanty, A. K., Barison, J., Dixit, A., Khurana, J., et al. (2022). Evaluation of fertilizer doses under different planting methods of rice (Oryza sativa) in west central table land zone of Odisha. Indian J. Agron. 67, 123–128. doi: 10.59797/ija.v67i2.107
Poudel, A., Saini, M. R., Phogat, S., Roy, J., Chauhan, H., Madhavan, J., et al. (2025). Growing Indica and Aus type rice till maturity in hydroponics under N-optimum and N-stressed conditions: A detailed account. J. Plant Growth Regul., 1–15. doi: 10.1007/S00344-025-11794-5
Rabiei, Z., Roshan, N. M., Sadeghi, S. M., Amiri, E., and Doroudian, H. R. (2021). Effect of drought stress and different levels of nitrogen and potassium fertilizers on the accumulation of osmolytes and chlorophyll in rice (Oryza sativa L.). Gesunde Pflanzen 73, 1–10. doi: 10.1007/S10343-021-00550-W
Shova, A., Md, K., Zulfikar, M. K., and Sadiqul, M. A. (2023). Enhanced potassium fertilization improved rice (Oryza sativa) yield and nutrient uptake in coastal saline soil of Bangladesh. J. Soil Sci. Plant Nutr. 23, 1884–1895. doi: 10.1007/S42729-023-01144-3
Song, Y., Dong, M., Jin, M., Gu, J., Chen, F., Chen, P., et al. (2025). Nitrogen management in rice under crop rotation and nitrogen level adjustment: Comprehensive responses of soil, roots, and plant growth. Rhizosphere 33, 101038–101038. doi: 10.1016/J.RHISPH.2025.101038
Sun, J. W., Li, N., Wang, C. Y., Zhao, J. H., Zhang, S. W., Jiang, J. M., et al. (2017). Effects of transplanting methods and potassium rates on lodging resistance of hybrid rice. J. Nucl. Agric. Sci. 31, 2408–2417. doi: 10.11869/j.issn.100-8551.2017.12.2408
Tian, K., Zhong, X. H., Huang, N. G., and Pan, J. F. (2014). Fertilizer rates on yield formation and lodging of rice. China Rice 3), 54–57. doi: 10.3969/j.issn.1006-8082.2014.03.012
Vijayakumar, S., Dinesh, K., Kulasekaran, R., and Rajpoot, S. K. (2022). Effect of potassium fertilization on water productivity, irrigation water use efficiency, and grain quality under direct seeded rice-wheat cropping system. J. Plant Nutr. 45, 2023–2038. doi: 10.1080/01904167.2022.2046071
Vijayakumar, S., Kumar, D., Kulasekaran, R., Bhargavi, B., Kaje, V. V., and Shivay, Y. S. (2024). Effect of split application of potassium on nutrient recovery efficiency, soil nutrient balance, and system productivity under rice-wheat cropping system (RWCS). J. Plant Nutr. 47, 1546–1563. doi: 10.1080/01904167.2024.2315974
Wang, Y., Yin, X., Wang, X., Ali, M. F., Lin, X., Gu, S., et al. (2025). Split potassium application delays senescence and increases grain yield in winter wheat grown on sandy and silt loam soils. Front. Plant Sci. 16. doi: 10.3389/FPLS.2025.1599296, PMID: 40475902
Wu, J., Liao, Q., Shah, F., Li, Z., Tao, Y., Wang, P., et al. (2025). The potential role of nitrogen management in enhancing grain yield and lodging resistance of Shanlan upland rice (Oryza sativa L.). Agronomy 15, 614. doi: 10.3390/AGRONOMY15030614
Wu, W. G., Zhang, H. C., Chen, Y., Li, J., Qian, Y. F., Wu, G. C., et al. (2008). Dry-matter accumulation and nitrogen absorption and utilization in middle-season indica super hybrid rice. Acta Agronomica Sin. 6), 1060–1068. doi: 10.3724/SP.J.1006.2008.01060
Yang, Z. Y., Sun, Y. J., Xu, H., Qing, J., and Jia, X. W. (2013). Effects of different cultivation methods on accumulation and transport of assimilation products and lodging resistance of stem-sheaths of no-tillage rice. Chin. J. Rice Sci. 27, 511–519. doi: 10.3969/j.issn.1001-7216.2013.05.008
Yao, B., He, H.-B., Xu, H.-C., Zhu, T.-Z., Liu, T., Ke, J., et al. (2021). Determining nitrogen status and quantifying nitrogen fertilizer requirement using a critical nitrogen dilution curve for hybrid indica rice under mechanical pot-seedling transplanting pattern. J. Integr. Agric. 20, 1474–1486. doi: 10.1016/S2095-3119(21)63622-5
Yu, L. H., Li, G. H., Xu, J. J., Yang, J. J., Wang, S. H., Liu, Z. H., et al. (2012). Characteristics of uptake of nitrogen, phosphorus, and potassium and partitioning in mechanical transplanting japonica rice. Acta Agronomica Sin. 38, 707–716. doi: 10.3724/SP.J.1006.2012.00707
Yuan, S., Su, Y., Chen, P., Tu, N., Zhou, W., and Yi, Z. (2025). Effect of split application of nitrogen combined with chitooligosaccharide on lodging in rice. Plant Soil, 1–17. doi: 10.1007/S11104-025-07508-1
Zhang, T., He, X., and Chen, B. (2021). Effects of different potassium (K) fertilizer rates on yield formation and lodging of rice. Phyton (Buenos Aires) 90, 815–826. doi: 10.32604/PHYTON.2021.014168
Zhang, L., Ma, Z., He, N., Tang, Z., Wang, C., Zheng, W., et al. (2025). Lodging resistance of Japonica hybrid rice plants studied in relation to mechanical and physicochemical characteristics. Agronomy 15, 699–699. doi: 10.3390/AGRONOMY15030699
Zhang, Y. J., Wang, Z. Q., Ma, P., Yang, Z. Y., and Sun, Y. J. (2022). Effects of water-nitrogen coupling on nitrogen uptake, utilization, and yield of rice under wheat straw returning. Chin. J. Rice Sci. 36, 388–398. doi: 10.16819/j.1001-7216.2022.210803
Zhang, H., Xu, M., Shi, X., Li, Z., Huang, Q., and Wang, X. (2010). Rice yield, potassium uptake and apparent balance under long-term fertilization in rice-based cropping systems in southern China. Nutrient Cycling Agroecosystems 88, 341–349. doi: 10.1007/s10705-010-9359-3
Keywords: rice, mechanical transplanted, N fertilizer, K fertilizer, nutrient utilization, yield
Citation: Li Z, Hu M, Liu T, Tang Y, Liu R, Peng Z, Wang C, Peng Z, Wang Z, Chen Z, Yang Z, Sun Y and Ma J (2025) Effects of N-K management strategies on nutrient uptake efficiency, lodging resistance, and yield in machine-transplanted rice. Front. Plant Sci. 16:1658245. doi: 10.3389/fpls.2025.1658245
Received: 02 July 2025; Accepted: 19 August 2025;
Published: 08 September 2025.
Edited by:
Sangeeta Lenka, Indian Institute of Soil Science (ICAR), IndiaReviewed by:
Bharat Prakash Meena, Indian Institute of Soil Science (ICAR), IndiaAk Biswas, Indian Council of Agricultural Research (ICAR), India
Copyright © 2025 Li, Hu, Liu, Tang, Liu, Peng, Wang, Peng, Wang, Chen, Yang, Sun and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Ma, bWFqdW5wMjAwMkAxNjMuY29t
†These authors have contributed equally to this work and share first authorship