 Xing Fan1,2
Xing Fan1,2 Yuanpeng Zhu3Yan Jia1Peishen Du1Weini Wang4Junmei Liu4Zhijun Lv5
Yuanpeng Zhu3Yan Jia1Peishen Du1Weini Wang4Junmei Liu4Zhijun Lv5 Ronghao Liu1,2*Xiaobin Li6*
Ronghao Liu1,2*Xiaobin Li6*- 1College of Water Resources Science and Engineering, Taiyuan University of Technology, Taiyuan, China
- 2Shanxi Key Laboratory of Collaborative Utilization of River Basin Water Resources, Taiyuan, China
- 3Anhui Provincial Academy of Eco-Environmental Science Research, Hefei, China
- 4Ordos Agriculture and Animal Husbandry Ecology and Resource Protection Center, Ordos, Inner Mengulia, China
- 5Agricultural and Animal Husbandry Technology Extension Center, Dalat Banner, Ordos, Inner Mengulia, China
- 6Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing, China
The availability of soil potassium plays a critical role in yield increases. Potassium-solubilizing microorganisms (KSM) offer a promising biological solution to improve potassium availability, but their efficacy across diverse global environments remains uncertain. Through a global meta-analysis of 102 studies (846 paired observations), we systematically evaluated the effects of KSM application on crop yields across five key dimensions: microorganism types, soil factors, crop classifications, field management, and stress types. KSM inoculation significantly increased soil available potassium (+28.9%), crop yield (+23.4%), and key growth indices, such as root length (+29.50%) and leaf area (+44.7%). This study identified Aspergillus spp. as the most suitable microorganism, and revealed that KSM efficacy is highly dependent on context: yield responses were greatest in clay loam soils, vegetable crops, and greenhouse conditions. Structural equation modeling indicated that microbial abundance, climate, soil available potassium, and plant growth (root length and leaf area) are key direct and indirect drivers of yield enhancement. The results indicate that the application of KSM is an effective strategy to increase crop yields in various environments. By identifying the optimal conditions for KSM application, the identification of optimal application parameters, derived from cross-study analysis, provides a robust strategy for leveraging microbial communities to boost soil potassium availability and nutrient efficiency, thereby contributing to the transition toward more sustainable and climate-resilient agriculture.
1 Introduction
Potassium (K) is an essential plant nutrient and the most abundant cation in plant cells, playing a critical role in a myriad of physiological and metabolic processes, including enzyme activation, osmotic regulation, and photosynthesis (Sheng, 2005; Sattar et al., 2019; Li et al., 2023b). Consequently, potassium availability directly governs photosynthetic efficiency, crop stress resistance, growth, and ultimately yield and quality (Yang et al., 2023; Wang et al., 2024; Zhao et al., 2024). However, the natural soil potassium pool often lacks sufficient capacity to convert insoluble K into plant-available forms, frequently failing to meet crop demand (Xu et al., 2025). This limitation represents a fundamental constraint on agricultural productivity and sustainability, contributing to inefficient fertilizer use and environmental imbalances (Zhao et al., 2024; Liu et al., 2025).
Addressing this challenge requires a shift from purely chemical solutions towards biological strategies that harness soil ecosystem processes. Potassium-solubilizing microorganisms (KSM) are a key functional group within the soil microbiome that can enhance mineral weathering and mobilize fixed potassium pools through biological mineralization (Sattar et al., 2019). Their mechanisms (including acid production, phytohormone secretion, and extracellular enzyme release) not only improve potassium availability but also enhance soil health and plant resilience (Meena et al., 2014; Sun et al., 2019; Chen et al., 2022). This positions KSM application at the intersection of microbial ecology and plant nutrition, offering a pathway to more sustainable agricultural intensification by working with, rather than against, soil biological processes (Shen et al., 2016).
Numerous targeted studies have demonstrated the efficacy of specific KSM strains (e.g., Bacillus, Pseudomonas) in increasing potassium solubility and crop yields under controlled or local conditions (Khanghahi et al., 2018; Sarikhani et al., 2018; Nawaz et al., 2023). However, the transition from promising microbe-level effects to reliable field-scale outcomes remains a major hurdle in microbial ecology applied to agriculture. A critical gap exists between demonstrating efficacy in discrete settings and predicting effectiveness across the heterogeneous landscapes of global agriculture. The intricate relationships between KSM functionality, environmental context (e.g., soil type, stress type), and field management are poorly quantified. Consequently, it is unclear whether the benefits of KSM are generalizable or contingent on specific, and often unknown, ecological interactions and agronomic conditions. This uncertainty hinders the development of predictive models and evidence-based recommendations for integrating KSM into sustainable farming systems.
To bridge this gap between microbial ecology and agricultural practice, we conducted a comprehensive global meta-analysis of studies from 2015-2024. By synthesizing evidence across five key dimensions: microbial species, soil types, crop types, field management, and stress types (Table 1). We aim to: (i) quantify the overarching impact of KSM on soil potassium dynamics and plant performance; (ii) identify the contextual factors that most strongly regulate KSM efficacy; and (iii) elucidate the complex pathways through which KSM influence crop yield, integrating microbial, plant, and environmental variables. Our work provides a mechanistic, evidence-based framework for understanding KSM ecology in agricultural systems, ultimately guiding the development of targeted microbial strategies for enhancing potassium use efficiency and advancing sustainable crop production.
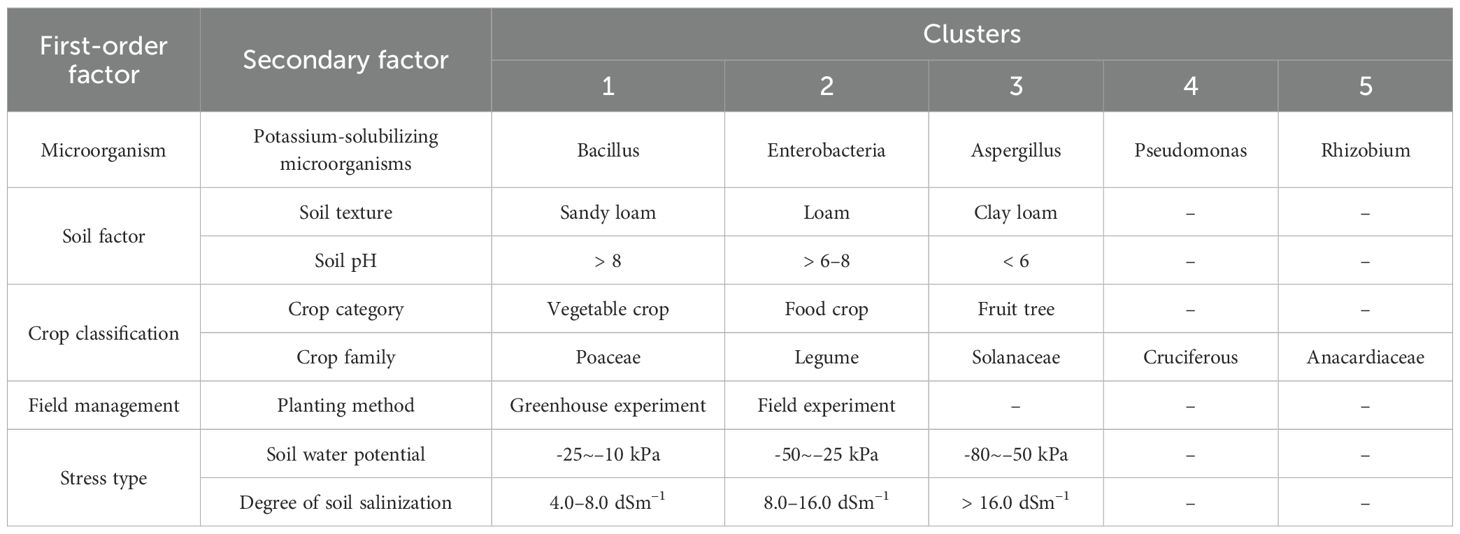
Table 1. Subgroup classification table for the forest plots.
2 Materials and methods
2.1 Data retrieval strategies and filtering criteria
The data for the meta-analysis were retrieved from Web of Science (WOS) and the China National Knowledge Infrastructure (CNKI), and the keywords searched included “potassium-solubilizing bacteria”, “yield”, “rhizosphere potassium-solubilizing bacteria”, “microorganisms”, “enzyme activity” and “potassium solubilization”. For the meta-analysis, 102 relevant published studies were collated, including 95 English and 7 Chinese studies (Figure 1), containing 846 pairs of paired observations(Among them, 806 pairs of data are from English databases, and 40 pairs of data are from Chinese databases). The selected studies had to satisfy the following requirements: (1) The retrieved variables included the mean, standard deviation (or standard error), and repeat group. (2) All the control and treatment groups belonged to the same ecosystem and experienced the same environmental and growth conditions. (3) Data from only the control and treatment groups inoculated with potassium-solubilizing microorganisms were analyzed in multiple controlled studies with experimental inoculation treatments. (4) The extracted literature was limited to articles published from 2015 to 2025.
Figure 1. Map of study sites included in the meta-analysis.
2.2 Data collection and classification
We also recorded climatic characteristics such as the mean annual temperature (MAT) and mean annual precipitation (MAP), collecting climatic data from the NASA Surface Meteorology and Solar Energy Location site (https://power.larc.nasa.gov/) on the basis of the latitude and longitude of the study site, if not reported in the original study.
Data from all published literature tables were extracted manually, whereas data from figures were extracted via the Get Data (https://getdata.sourceforge.net/download.html) tool.
2.3 Meta-data analysis
We used Meta-Win 2.0 software (https://www.metawinsoft.com/) to analyze the data for random effects. The effect value was calculated via the natural logarithm of the response ratio (lnRR) in Equation (2) (Luo et al., 2018), which was then converted to upper and lower 95% confidence intervals (CI) in Equation (4) to evaluate the impact of the KSM on crop yield, soil physicochemical parameters, and crop growth indicators. In several studies, the standard error (SE) in Equation (1) had to be translated to the standard deviation (SD) in Equation (1), which was calculated as (Jian et al., 2016):
The variance of the effect value is as follows:
where is the mean of the variable in the treatment group and where is the mean of the variable in the control group in Equation (3). St is the standard deviation of the variable in the treatment group; Sc is the standard deviation of the variable in the control group; n is the sample size; and nt and nc are the sample sizes of the treatment and control groups, respectively in Equation (3). We converted lnRR and its corresponding confidence intervals to the corresponding percentage change.
2.4 Heterogeneity test and publication bias test
The I2 statistic was used to assess heterogeneity, and the test value Q was calculated as follows in Equation (5) and (6) (Huo et al., 2017):
where wi is the weight of group i of data, n is the number of effect sizes, Ei is the effect size of the ith group, and E is the average of all data effect sizes in Equation (5). Calculations were made using the random effect model when the heterogeneity test result I2 was greater than 50% and the fixed effect model when it was less than 50%. The Rosenthal loss of safety coefficient method was used for the publication bias test. N>5 m+10 (N is the loss of safety coefficient, and m is the sample size) indicates no publication bias. We used AMOS software (IBM SPSS AMOS 20.0.0) to untangle the indirect and direct effects of climate (MAT and MAP), microbial count, soil available potassium content, crop growth indicators, and soil enzyme activities on crop yields (Eisenhauer et al., 2015) via structural equation modeling (SEM). Before modeling, all the data were normalized (ensuring that the data followed a standard normal distribution with a mean value of 0 and standard deviation of 1), and an a priori model was designed based on the known effects and relationships among the drivers that had a significant impact on crop yields in our previous analyses. SEM was used instead of multiple regressions since directions can be assigned to several relationships, resulting in multiple explanatory and response variables in one model. Furthermore, the structure of such a model can reveal whether a significant bivariate relationship results from a significant relationship between two given variables and a third variable. Despite using fail-safe N to assess publication bias, our results may still be affected by the underreporting of studies with non-significant or negative findings. While subgroup analysis explored substantial heterogeneity (indicated by I²), residual heterogeneity could remain due to unmeasured factors like experimental duration, KSM strain potency, and unrecorded soil microbial communities, which may limit the generalizability of pooled effects (Jin et al., 2015; Mathur and VanderWeele, 2022).
3 Results
3.1 Effect of KSM inoculation on crop yield
The calculation of the effect size on yield revealed that KSM inoculation improved overall crop yield by 23.43% compared with the control, confirming its potential as a yield-enhancing agricultural practice (Figure 2). Note: The four KSM genera analyzed (Aspergillus, Pseudomonas, Bacillus, Enterobacter) were selected based on two criteria: (1) their high frequency of reporting in existing KSM-related studies (accounting for >70% of eligible literature included in this meta-analysis); (2) their well-documented potassium-solubilizing mechanisms and agricultural application records, ensuring data robustness and practical relevance. Among these genera, Aspergillus (+36.27%) and Pseudomonas (+32.86%) were consistently demonstrated to outperform Bacillus and Enterobacter in boosting crop yields (P<0.05).

Figure 2. Effect value of KSM inoculation over uninoculated KSM on crop yield (the significant signal means *P<0.05, **P<0.01, ***P<0.001).
Soil texture emerged as a key modulating factor: clay loam soil (+38.82%) showed a significantly stronger yield response to KSM than sandy loam and loam (P<0.05), while soil pH had no substantial impact. Crop type and family also drove marked differences: vegetable crops (+34.25%) exhibited the greatest yield increase relative to food crops and fruit trees (P<0.05), with cruciferous (+42.74%) and leguminous (+40.08%) crops outperforming Poaceae and Anacardiaceae (P<0.05). Additionally, greenhouse experiments (+34.17%) yielded better results than field trials (P<0.05), whereas soil water potential and EC values did not induce significant variations in yield response.
3.2 Effect of KSM inoculation on soil physicochemical properties
3.2.1 Effect of KSM inoculation on soil nutrients and pH
Compared with the control, KSM inoculation significantly modified soil potassium status and pH, with soil total potassium and available potassium increasing by 18.28% and 28.91%, respectively, and soil pH decreasing by 18.10% (Supplementary Figures S2A, B).
Notably, the efficacy of KSM in boosting soil available potassium varied markedly by biological and environmental factors: Enterobacter (+39.22%) showed a significantly stronger capacity to mobilize available potassium than Bacillus and Pseudomonas (P<0.05), while loam soil (+32.25%) outperformed clay loam and sandy loam in this regard (P<0.05). Crop-related factors also drove substantial differences: vegetable crops (+50.4%) and Poaceae (+32.71%) exerted the greatest positive effects on soil potassium availability among crop types and families, respectively (P<0.05). In contrast, soil pH had no substantial impact on KSM-mediated available potassium augmentation, indicating that KSM can stably improve potassium availability across diverse soil acid-base conditions.
For soil pH regulation, all tested KSM genera induced negative effects, though with no significant differences between them. Soil texture and initial pH modulated this effect: loam (–18.32%) and clay loam (–25.06%) showed stronger pH reduction than sandy loam, and KSM inoculation at pH>8 (–25.38%) had a significantly greater acidifying effect than at pH 6–8 (P<0.05), suggesting that KSM are more effective at lowering pH in alkaline soils.
3.2.2 Effect of KSM inoculation on soil enzyme activity
KSM inoculation significantly enhanced the activities of both antioxidant-related and metabolism-related enzymes, with peroxidase (+41.0%) and cellulase (+147.3%) showing the highest enhancement rates, respectively (Supplementary Figure S2A, C).
For antioxidant-related enzymes, superoxide dismutase activity trended positive in sandy loam and loam but decreased in clay loam, while peroxidase was beneficial only in loam soil. In contrast, catalase activity showed clear strain and texture specificity: Pseudomonas (+50.92%) was significantly more effective than Bacillus (P<0.05), and clay loam/loam supported positive responses whereas sandy loam showed a negative effect. Consistently across all antioxidant enzymes, KSM promoted activity at soil pH<6 but suppressed it at pH>8 relative to the control, highlighting soil pH as a key regulator of KSM-induced antioxidant defense.
Metabolism-related enzymes showed universal enhancement, with protease (+58.9%), chitinase (+62.7%), and cellulase (+147.3%) exhibiting the strongest activity increases, indicating that KSM play a critical role in accelerating soil organic matter decomposition and nutrient cycling. Sucrase showed the weakest response (+13.6%), suggesting that KSM have enzyme-specific regulatory effects on soil metabolic processes.
3.3 Effect of KSM inoculation on crop growth
3.3.1 Effect of KSM inoculation on crop potassium content
KSM inoculation significantly enhanced potassium uptake in crops, with root K concentration and shoot K concentration both showing marked increases relative to the control (Figures 3A, B).
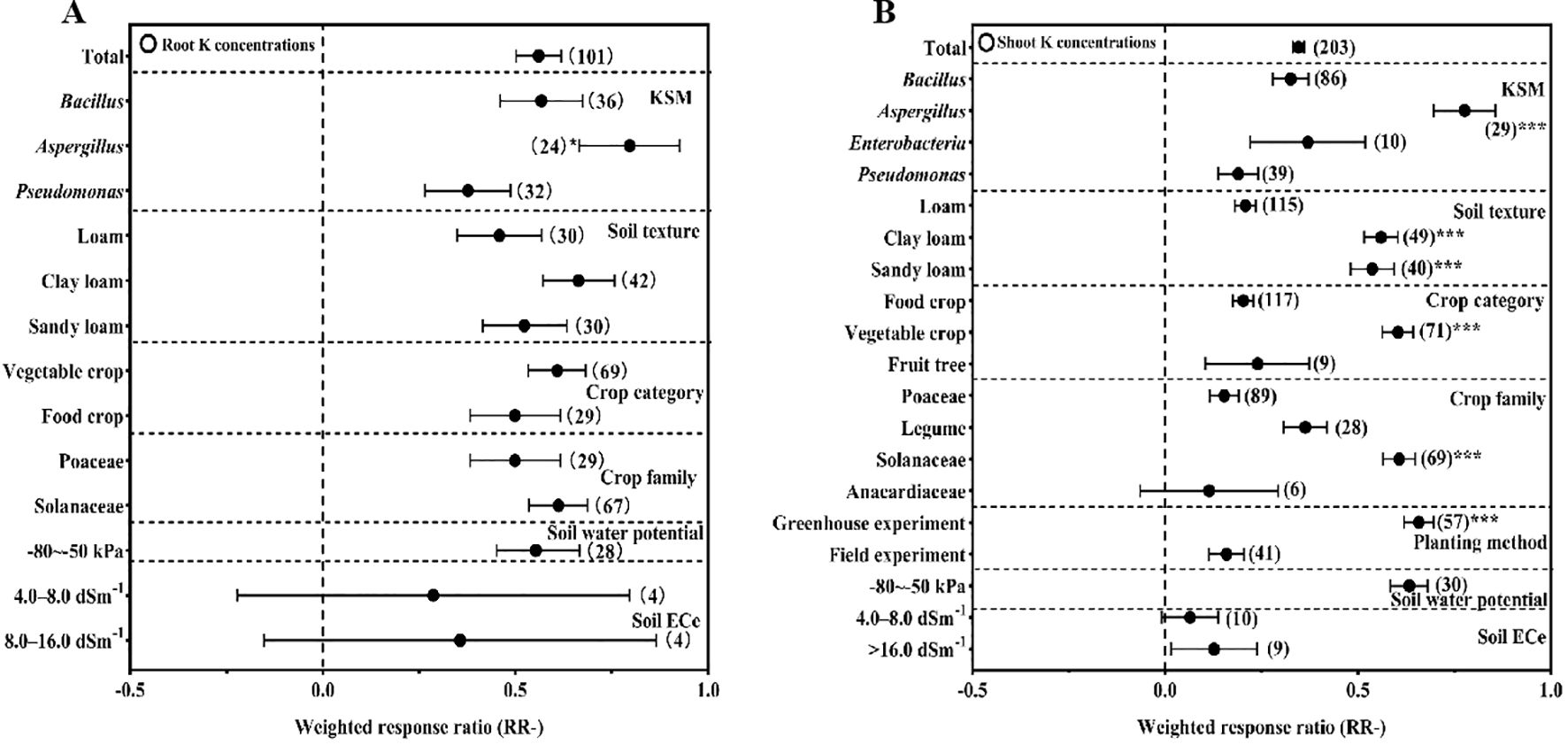
Figure 3. Effect of different conditions on K concentrations in roots (A) and in shoots (B) under inoculation with KSM (the significant signal means *P<0.05, **P<0.01, ***P<0.001).
For root K concentration, Aspergillus (+79.64%) was the most effective genus, outperforming Bacillus and Pseudomonas (P<0.05). Notably, the effect values for root K concentration were consistently positive across soil textures, crop types, crop families, and soil ECe conditions, with no significant variations between subgroups—indicating that KSM promotes root potassium accumulation stably across diverse contexts.
Shoot K concentration was increased by 31.11% overall, with Aspergillus (+77.65%) again outperforming Enterobacter, Bacillus, and Pseudomonas (P<0.05). Soil texture significantly modulated this effect: clay loam (+55.99%) and sandy loam (+53.71%) were more effective than loam (P<0.05). By crop category, vegetable crops (+60.38%) showed a stronger response than food crops and fruit trees (P<0.05), and Solanaceae (+60.67%) outperformed other crop families (P<0.05). No significant differences in shoot K concentration were detected under various soil ECe conditions.
3.3.2 Effect of KSM inoculation on growth indices
Compared with the control, KSM significantly increased all measured crop growth indices, with dry root weight (+59.21%) and total chlorophyll content (+56.06%) showing the most dramatic improvements, followed by leaf area (+44.67%) and root fresh weight (+40.79%) (Supplementary Figure S3A).
For root length (Figure 4A), KSM exerted positive effects across all subgroups, with clay loam soil (+52.71%) and field experiments (+57.20%) standing out as the most favorable conditions (P<0.05). Notably, plant height responded strongly to Aspergillus inoculation (+54.33%), which was significantly more effective than other genera (P<0.05); sandy loam soil (+36.71%) and vegetable crops (+43.30%), particularly solanaceae (+47.11%), also showed enhanced plant height under KSM treatment (P<0.05). In contrast, soil pH had no significant influence on plant height. Greenhouse experiments (+49.86%) outperformed field trials in promoting plant height (P<0.05), and the optimal soil water potential range was –80 to –50 kPa (+55.64%, P<0.05).

Figure 4. Effect of different conditions on root length (A), leaf area (B) under inoculation with KSM (the significant signal means *P<0.05, **P<0.01, ***P<0.001).
Leaf area was most notably increased by Aspergillus (+76.00%, P<0.05), with vegetable crops (+62.89%) and solanaceae (+63.37%) again showing stronger responses than other crop types/families (P<0.05, Figure 4B). Greenhouse conditions (+71.24%) also facilitated a greater increase in leaf area compared to field settings (P<0.05), while soil texture and pH had no significant differential effects.
3.4 Effect of KSM inoculation on the K content in soil and crops in relation to environmental factors
A key chain emerged from the correlation analysis (Figure 5): soil available potassium was significantly and positively correlated with both crop shoot K concentrations (P<0.05) and root K concentrations (P<0.05), while root K concentrations further positively linked to shoot K concentrations (P<0.05). Critically, both soil available potassium and crop shoot K concentrations showed direct positive correlations with crop yield (P<0.05), highlighting that KSM’s yield-enhancing effect is likely mediated through improved soil K availability and subsequent crop K uptake.

Figure 5. Distribution of the variable importance for (A) Yield, (B) Root length, (C) Plant height, (D) Leaf area, (E) Root K concentrations, and (F) Shoot K concentrations random forest models including the different Microbial count, climatic conditions, soil properties and Growth indicators as variable, the significant signal means *P<0.05, **P<0.01.
3.5 Relationships between effect sizes of inoculation with KSM on yield and other growth indicators
The results of the random forest analysis are presented in Figure 6. We assigned significance to factors influencing yield and other growth indicator changes by considering those with a significance level greater than 0.1. For yield, the key control factor is microbial count and soil available K. For root length, the key control factor is soil available K. For plant height, the key control factors are root length, soil available K, MAT, microbial count, soil pH, MAP, shoot K concentrations, and soil water potential. For leaf area, significant control factors included soil pH, microbial count, and plant height. For root K concentrations, significant control factors included microbial count, MAT, and soil available K. For shoot K concentrations, the significant control factors included soil available K and root K concentrations.

Figure 6. Relationship between the effect size of soil available K and (A) yield, B) root K concentrations, (C) shoot K concentrations, (D) root length and (E) plant height; root K concentrations and (F) shoot K concentrations, (G) plant height and (H) leaf area; shoot K concentrations and (I) plant height, (G) leaf area; plant height and (K) yield. Reported statistical results from two-sided linear regressions. Gray areas represent 95%Cl.
3.6 Impact of microbial count, MAT, and MAP on crop yield following KSM inoculation
Structural equation modeling (SEM) was used to investigate how the various explanatory variables and their interactions influenced crop yield under KSM inoculation (Figure 7). The final structural equation model explained 19% of the overall variation in crop yield (χ2 = 5.09; P = 0.955, AIC = 113.09). The microbial count, MAP, MAT, soil available potassium, root K concentration, root length, leaf area, and oxidase were the most crucial factors affecting crop yield, both directly and indirectly. Overall, the microbial count indirectly affects crop yield by altering root length (path coefficient = 0.76), root K concentration (path coefficient = 0.93), and leaf area (path coefficient = 1.49). MAP indirectly affects crop yield by influencing root length (path coefficient = 0.81), root K concentration (path coefficient = 0.92), and leaf area (path coefficient = 1.69). The MAT had an indirect effect (path coefficient = 0.81), and the root K concentration (path coefficient = 0.92) and leaf area (path coefficient = 1.69) were affected. MAT also indirectly improved crop yield by influencing root K concentrations (path coefficient =–0.21) and leaf area (path coefficient =–0.15). Soil available potassium indirectly affected crop yield by influencing root length (path coefficient = 0.31) and root K concentrations (path coefficient = 0.29). The root K concentrations also indirectly influenced crop yield because of their impact on root length (path coefficient =–0.32). Root length had a direct positive effect on crop yield (path coefficient = 0.21), and leaf area had a direct positive effect on crop yield (path coefficient = 0.24). Oxidase had a direct positive effect on crop yield (path coefficient = 0.14). These results confirm the potential of KSM to increase crop production through inoculation.

Figure 7. Structural equation modeling (SEM) describing the direct and indirect effects of predictor variables on effect size of field.
4 Discussion
4.1 Effects of KSM on different microbial genera and soil factors
Our results showed that KSM significantly increased crop yield, soil available K, and crop potassium uptake and growth, confirming their role in bioweathering potassium-containing minerals and mediating soil potassium transformation (Chen et al., 2022). Elevated root and shoot K concentrations further promote crop growth and yield, which aligns with Olaniyan who emphasized that enhanced potassium nutrition improves root vigor and photosynthetic efficiency (Olaniyan et al., 2022).
Crop yield and development varied with KSM genus, soil texture, and other contextual factors. Among tested microorganisms, Aspergillus showed the strongest overall effect on crop K content and yield, outperforming Bacillus and Pseudomonas. This superiority stems from its synergistic mechanisms: it secretes multiple extracellular organic acids (e.g., oxalic acid) to disrupt potassium-bearing mineral lattices and releases phosphatases/glucosidases (Ashrafi-Saiedlou et al., 2024), while producing phytohormones and iron transporters to optimize root physiological functions (Kumawat et al., 2024). Unlike Bacillus (which primarily relies on acidolysis), Aspergillus also improves rhizosphere microecology and soil aggregation (Imran et al., 2021), making it a key candidate for microbial preparation development and potash fertilizer efficiency improvement.
With respect to soil texture, clay loam showed the strongest yield response to KSM, which is attributed to the unique improvement effect of KSM on this soil type. Clay loam typically suffers from poor aeration that hinders root nutrient uptake (Hsiao et al., 2018), but KSM inoculation maintains stable available potassium levels during the crop reproductive period (Singh et al., 2022) and produces extracellular polysaccharides to promote soil aggregation (Wang et al., 2022). This structural improvement increases aeration and nutrient diffusion, enhancing root K uptake and root length (a trend consistent with the significant increase in root K concentration and length observed in clay loam). These findings indicate that KSM can targeted alleviate the limitations of clay loam soils, providing a solution for yield improvement in such agricultural systems.
4.2 Effects of KSM under different crop classifications, field management practices, and stress types
With respect to crop classification, our synthesis indicated that vegetable crops responded more positively to KSM than other crop types, which can be explained by their biological traits: most vegetables have short growth cycles, high potassium demand, and quality formation closely linked to potassium nutrition (Zoerb et al., 2014). Compared with cereals, vegetables such as sugar beet exhibit higher K mobilization efficiency per unit root length (El Dessougi et al., 2002; Wang et al., 2011), making them more sensitive to KSM-mediated potassium supplementation. This finding provides a basis for targeted KSM application—prioritizing vegetable crops in agricultural production to maximize the return on microbial inoculation.
With respect to field management, greenhouse conditions significantly enhanced KSM efficacy relative to field trials, as the controlled temperature, humidity, and light optimize both crop growth and microbial survival (Shamshiri et al., 2018; Farvardin et al., 2024). This suggests that KSM application should be paired with appropriate environmental regulation to fully exploit its potential.
Under stress types, KSM consistently improved crop performance across drought and salinity gradients, which is achieved through dual mechanisms: supplementing potassium to maintain physiological functions (e.g., photosynthesis, enzyme activity) (Munsif et al., 2022) and enhancing stress defense systems (e.g., increasing antioxidant enzyme activity) (Feng et al., 2019). For salinity stress, KSM also helps maintain K+/Na+ balance (Romero-Munar and Aroca, 2023), further confirming its role in improving crop stress resilience.
4.3 Key influences of the KSM on improving crop yields
This study demonstrated a significant linear correlation among soil available potassium, root/shoot potassium concentrations, and crop yield following KSM inoculation, consistent with previous findings (Shen et al., 2016; Ai et al., 2022). The increase in yield can be attributed to a well-defined physiological pathway initiated by KSM-mediated potassium solubilization.
KSM enhances the availability of soil potassium, leading to elevated potassium uptake by roots and subsequent translocation to shoots (Chen et al., 2021). The increased potassium levels stimulate root growth, including surface area, vigor, and biomass, which further expands nutrient absorption capacity and improves water retention (Sustr et al., 2019). Simultaneously, potassium promotes shoot development by facilitating cell expansion, leaf area enlargement, and photosynthetic efficiency (Hu et al., 2022; Ding et al., 2024).
Our SEM analysis revealed a multi-level regulatory network of KSM on yield, with soil available potassium and microbial count as core indirect drivers, and root length/leaf area as direct effectors. Soil available potassium promotes root tip cell division and elongation by regulating growth hormone transport (Sheng, 2005), while microbial count modulates root traits and nutrient availability (Sun et al., 2023), thereby establishing a coherent pathway from potassium activation to root growth and, ultimately, yield increase. Climate factors also play a role: MAP improves soil moisture to enhance root uptake (Li et al., 2023a), while MAT shows dual effects—moderate temperatures promote metabolism, but extreme temperatures inhibit root K uptake (Jacobson et al., 1957), highlighting the need for temperature adaptation in KSM application.
These findings have important implications for sustainable agriculture: KSM can reduce reliance on mineral potash fertilizers by improving soil potassium use efficiency, particularly in clay loam and vegetable cropping systems. However, practical application must consider contextual limitations, for example, KSM efficacy may decline in highly saline soils due to microbial survival constraints. Future research should focus on optimizing inoculation strategies and developing climate-adapted KSM formulations to enhance applicability across diverse agricultural environments.
5 Conclusions
This meta-analysis clarified the efficacy and regulatory mechanisms of KSM across diverse environments, filling the gap between local trials and global application guidelines. Key findings: (1) KSM inoculation significantly improved multiple agronomic parameters via a sequential process initiated by boosting soil available potassium, followed by enhanced plant potassium assimilation, and culminating in yield increase.; (2) Structural equation modeling identified microbial count and soil available K as core indirect drivers, with root length/leaf area as direct effectors; (3) Aspergillus was the most effective genus, with optimal responses in clay loam, vegetable crops, and greenhouse conditions. These findings provide a data-driven basis for reducing mineral potash fertilizer reliance—practically, we recommend targeted inoculation of Aspergillus in clay loam vegetable fields, combined with greenhouse environmental regulation. In summary, KSM is a potent biological strategy for sustainable agriculture, and our work offers actionable pathways for its optimized application.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author contributions
XF: Writing – original draft, Writing – review & editing. YZ: Writing – original draft. YJ: Writing – review & editing. PD: Writing – review & editing. WW: Funding acquisition, Writing – original draft. JL: Funding acquisition, Writing – original draft. ZL: Funding acquisition, Writing – original draft. RL: Writing – original draft, Writing – review & editing. XL: Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by the Key Research and Development Program Young Scientist Project (2023YFD1901900), the Natural Science Foundation of Shanxi province (202203021211139), the Inner Mongolia Ordos City Science and Technology Program, and Research and Promotion Project of Water Conservancy Science and Technology in Shanxi Province (2025GM15).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2025.1659478/full#supplementary-material
References
Ai, W., Guo, T., Lay, K.D., Ou, K., Cai, Y.Ding K., et al. (2022). Isolation of soybean-specific plant growth-promoting rhizobacteria using soybean agglutin and evaluation of their effects to improve soybean growth, yield, and soil nutritional status. Microbiol. Res. 261, 127076. doi: 10.1016/j.micres.2022.127076
Ashrafi-Saiedlou, S., Rasouli-Sadaghiani, M., Samadi, A., Barin, M., Sepehr, E., et al. (2024). Aspergillus Niger as an eco-friendly agent for potassium release from K- bearing minerals: Isolation, screening and culture medium optimization using Plackett-Burman design and response surface methodology. Heliyon. 10 (7). doi: 10.10.1016/j.heliyon.2024.e29117
Chen, H., Zhang, Q., Wang, X., Zhang, J., Ismail, A. M., Zhang, Z., et al. (2021). Nitrogen form-mediated ethylene signal regulates root-to-shoot K+ translocation via NRT1.5. Plant Cell Environ. 44, 3576–3588. doi: 10.1111/pce.14182
Chen, Y., Yang, H., Shen, Z., and Ye, J.. (2022). Whole-genome sequencing and potassium-solubilizing mechanism of bacillus aryabhattai SK1-7. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.722379
Ding, X., Wu, Y., Zheng, B., Zhong, G., Song, X., Geng, Y., et al. (2024). Assessment of nitrogen, phosphorus and potassium flows in China’s food system during 2010-2019: Implications for integrated management. Resour. Conserv. Recycl. 207, 107652. doi: 10.1016/j.resconrec.2024.107652
Eisenhauer, N., Bowker, M. A., Grace, J. B., and Powell, J. R.. (2015). From patterns to causal understanding: Structural equation modeling (SEM) in soil ecology. Pedobiologia 58, 65–72. doi: 10.1016/j.pedobi.2015.03.002
El Dessougi, H., Claassen, N., and Steingrobe, B. (2002). Potassium efficiency mechanisms of wheat, barley, and sugar beet grown on a K fixing soil under controlled conditions. J. Plant Nutr. Soil Sci. 165, 732–737. doi: 10.1002/jpln.200290011
Farvardin, M., Taki, M., Gorjian, S., Shabani, E., Sosa-Savedra, J. C., et al. (2024). Assessing the physical and environmental aspects of greenhouse cultivation: A comprehensive review of conventional and hydroponic methods. Sustainability 16, 1273. doi: 10.3390/su16031273
Feng, K., Cai, Z., Ding, T., Yan, H., Liu, X., Zhang, Z., et al. (2019). Effects of potassium-solubilizing and photosynthetic bacteria on tolerance to salt stress in maize. J. Appl. Microbiol. 126, 1530–1540. doi: 10.1111/jam.14220
Hsiao, C.-J., Sassenrath, G. F., Zeglin, L. H., Hettiarachchi, G. M., Rice, C. W., et al. (2018). Vertical changes of soil microbial properties in claypan soils. Soil Biol. Biochem. 121, 154–164. doi: 10.1016/j.soilbio.2018.03.012
Hu, W., Lu, Z., Gu, H., Ye, X., Li, X., Cong, R., et al. (2022). Potassium availability influences the mesophyll structure to coordinate the conductance of CO2 and H2O during leaf expansion. Plant Cell Environ. 45, 2987–3000. doi: 10.1111/pce.14405
Huo, C., Luo, Y., and Cheng, W. (2017). Rhizosphere priming effect: A meta-analysis. Soil Biol. Biochem. 111, 78–84. doi: 10.1016/j.soilbio.2017.04.003
Imran, M., Abulreesh, H. H., Monjed, M. K., Elbanna, K., Samreen, Ahmad, I., et al. (2021). Multifarious functional traits of free-living rhizospheric fungi, with special reference to Aspergillus spp. isolated from North Indian soil, and their inoculation effect on plant growth. Ann. Microbiol. 71, 31. doi: 10.1186/s13213-021-01643-7
Jacobson, L., Overstreet, R., Carlson, R. M., and Chastain, J. A. (1957). The effect of pH and temperature on the absorption of potassium and bromide by barley roots. Plant Physiol. 32, 658–662. doi: 10.1104/pp.32.6.658
Jian, S., Li, J., Chen, J., Wang, G, Mayes, M. A., Dzantor, K. E., et al. (2016). Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 101, 32–43. doi: 10.1016/j.soilbio.2016.07.003
Jin, Z.-C., Zhou, X.-H., and He, J. (2015). Statistical methods for dealing with publication bias in meta-analysis. Stat Med. 34, 343–360. doi: 10.1002/sim.6342
Khanghahi, M. Y., Pirdashti, H., Rahimian, H., Nematzadeh, G., Sepanlou, M. G., et al. (2018). Potassium solubilizing bacteria (KSB) isolated from rice paddy soil: from isolation, identification to K use efficiency. Symbiosis 76, 13–23. doi: 10.1007/s13199-017-0533-0
Kumawat, A., Kumar, D., Shivay, Y. S., Yadav, D., Sadhukhan, R., Gawdiya, S., et al. (2024). Sustainable basmati rice yield and quality enhancement through long-term organic nutrient management in the Indo-Gangetic Plains. Field Crops Res. 310, 109356. doi: 10.1016/j.fcr.2024.109356
Li, X., Li, M., Cen, X., Xu, L., and He, N.. (2023a). High precipitation rates increase potassium density in plant communities in the Tibetan Plateau. Commun. Earth Environ. 4, 368. doi: 10.1038/s43247-023-01033-8
Li, Z., Qiu, L., Zhang, T., Gaoyang, E., Zhang, L., Wang, L., et al. (2023b). Long-term application of controlled-release potassium chloride increases maize yield by affecting soil bacterial ecology, enzymatic activity and nutrient supply. Field Crops Res. 297, 108946. doi: 10.1016/j.fcr.2023.108946
Liu, Z., Yuan, X., Zhang, Z., Yang, X., Ai, C., Wang, Z., et al. (2025). Revisiting potassium-induced impacts on crop production and soil fertility based on thirty-three Chinese long-term experiments. Field Crops Res. 322, 109732. doi: 10.1016/j.fcr.2024.109732
Luo, G., Li, L., Friman, V. P., Guo, J., Guo, S., Shen, Q., et al. (2018). Organic amendments increase crop yields by improving microbe-mediated soil functioning of agroecosystems: A meta-analysis. Soil Biol. Biochem. 124, 105–115. doi: 10.1016/j.soilbio.2018.06.002
Mathur, M. B. and VanderWeele, T. J. (2022). Methods to address confounding and other biases in meta-analyses: review and recommendations. Annu. Rev. Public Health 43, 19–35. doi: 10.1146/annurev-publhealth-051920-114020
Meena, V. S., Maurya, B. R., and Verma, J. P. (2014). Does a rhizospheric microorganism enhance K+ availability in agricultural soils? Microbiol. Res. 169, 337–347. doi: 10.1016/j.micres.2013.09.003
Munsif, F., Shah, T., Arif, M., Jehangir, M., Afridi, M. Z., Ahmad, I., et al. (2022). Combined effect of salicylic acid and potassium mitigates drought stress through the modulation of physio-biochemical attributes and key antioxidants in wheat. Saudi. J. Biol. Sci. 29, 103294. doi: 10.1016/j.sjbs.2022.103294
Nawaz, A., Ul Qamar, Z., Marghoob, M. U., Imtiaz, M., Imran, A., Mubeen, F., et al. (2023). Contribution of potassium solubilizing bacteria in improved potassium assimilation and cytosolic K/Na ratio in rice (Oryza sativa L.) under saline-sodic conditions. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1196024
Olaniyan, F. T., Alori, E. T., Adekiya, A. O., Ayorinde, B. B., Daramola, F. Y., Osemwegie, O. O., et al. (2022). The use of soil microbial potassium solubilizers in potassium nutrient availability in soil and its dynamics. Ann. Microbiol. 72, 45. doi: 10.1186/s13213-022-01701-8
Romero-Munar, A. and Aroca, R. (2023). A non-K+-solubilizing PGPB (Bacillus megaterium) increased K+ deprivation tolerance in Oryza sativa seedlings by up-regulating root K+ transporters. Plant Physiol. Biochem. 196, 774–782. doi: 10.1016/j.plaphy.2023.02.027
Sarikhani, M. R., Oustan, S., Ebrahimi, M., and Aliasgharzad, N.. (2018). Isolation and identification of potassium-releasing bacteria in soil and assessment of their ability to release potassium for plants. Eur. J. Soil Sci. 69, 1078–1086. doi: 10.1111/ejss.12708
Sattar, A., Naveed, M., Ali, M., Zahir, Z. A., Nadeem, S. M., Yaseen, M., et al. (2019). Perspectives of potassium solubilizing microbes in sustainable food production system: A review. Appl. Soil Ecol. 133, 146–159. doi: 10.1016/j.apsoil.2018.09.012
Shamshiri, R. R., Jones, J. W., Thorp, K. R., Ahmad, D., Man, H. C., Taheri, S., et al. (2018). Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: a review. Int. Agrophys. 32, 287–302. doi: 10.1515/intag-2017-0005
Shen, H., He, X., Liu, Y., Chen, Y., Tang, J., Guo, T., et al. (2016). A complex inoculant of N2-fixing, P- and K-solubilizing bacteria from a purple soil improves the growth of kiwifruit (Actinidia chinensis) plantlets. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00841
Sheng, X. F. (2005). Growth promotion and increased potassium uptake of cotton and rape by a potassium releasing strain of Bacillus edaphicus. Soil Biol. Biochem. 37, 1918–1922. doi: 10.1016/j.soilbio.2005.02.026
Singh, S. K., Wu, X., Shao, C., and Zhang, H. (2022). Microbial enhancement of plant nutrient acquisition. Stress Biol. 2, 3–3. doi: 10.1007/s44154-021-00027-w
Sun, F., Ou, Q., Yu, H., Li, N., Peng, C., et al. (2019). The invasive plant Mikania micrantha affects the soil foodweb and plant-soil nutrient contents in orchards. Soil Biol. Biochem. 139, 107630. doi: 10.1016/j.soilbio.2019.107630
Sun, L., Wang, S., Rao, M. P. N., Shi, Y., Lian, Z. H., Jin, P. J., et al. (2023). The shift of soil microbial community induced by cropping sequence affect soil properties and crop yield. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1095688
Sustr, M., Soukup, A., and Tylova, E. (2019). Potassium in root growth and development. Plants (Basel). 8, 435. doi: 10.3390/plants8100435
Wang, H. Y., Shen, Q. H., Zhou, J. M., Wang, J., Du, C. W., Chen, X. Q., et al. (2011). Plants use alternative strategies to utilize nonexchangeable potassium in minerals. Plant Soil 343, 209–220. doi: 10.1007/s11104-011-0726-x
Wang, D., Lin, J. Y., Sayre, J. M., Schmidt, R., Fonte, S.J., Rodrigues, J. L. M., et al. (2022). Compost amendment maintains soil structure and carbon storage by increasing available carbon and microbial biomass in agricultural soil-A six-year field study. Geoderma 427, 116117. doi: 10.1016/j.geoderma.2022.116117
Wang, C., Xie, Y., and Tan, Z.. (2024). Soil potassium depletion in global cereal croplands and its implications. Sci. Total. Environ. 907, 167875. doi: 10.1016/j.scitotenv.2023.167875
Xu, Z., Lai, T., Li, S., Si, D., Zhang, C., et al. (2025). Effective potassium management for sustainable crop production based on soil potassium availability. Field Crops Res. 326, 109865. doi: 10.1016/j.fcr.2025.109865
Yang, C., Zhang, W., Gu, H., Liu, A., Guo, Q., Chen, Y., et al. (2023). Field, plant, to leaf: A meta-analysis on crop water use efficiency response to potassium fertilization. J. Hydrol. 621, 129578. doi: 10.1016/j.jhydrol.2023.129578
Zhao, Y., Liang, H., Zhang, J., Chen, Y., Dhital, Y. P., Zhao, T., et al. (2024). Isolation and characterization of potassium-solubilizing rhizobacteria (KSR) promoting cotton growth in saline-sodic regions. Microorganisms 12, 1474. doi: 10.3390/microorganisms12071474
Keywords: potassium-solubilizing microorganisms, soil available potassium, crop yield, meta-analysis, sustainable agriculture
Citation: Fan X, Zhu Y, Jia Y, Du P, Wang W, Liu J, Lv Z, Liu R and Li X (2025) Multi-environment meta-analysis reveals the mechanism of action of potassium-solubilizing microorganisms on crop yields. Front. Plant Sci. 16:1659478. doi: 10.3389/fpls.2025.1659478
Received: 04 July 2025; Accepted: 20 October 2025;
Published: 03 November 2025.
Edited by:
M. J. I. Shohag, University of Florida, United StatesReviewed by:
Shafaqat Ali, Government College University, PakistanAbdoulaye Soumare, Ziguinchor University, Senegal
Copyright © 2025 Fan, Zhu, Jia, Du, Wang, Liu, Lv, Liu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ronghao Liu, bGl1cm9uZ2hhb0B0eXV0LmVkdS5jbg==; Xiaobin Li, bGl4aWFvYmluMDFAY2Fhcy5jbg==