 Ekaterina D. Badaeva1,2*†
Ekaterina D. Badaeva1,2*† Maida Jazmin González Franco1†
Maida Jazmin González Franco1† Olga Razumova1†
Olga Razumova1† Nadezhda A. Tereshchenko2†
Nadezhda A. Tereshchenko2† Mikhail Divashuk1†
Mikhail Divashuk1†- 1Laboratory of Applied Genomics and Crop Breeding, All-Russian Research Institute of Agricultural Biotechnology, Russian Academy of Agricultural Sciences, Moscow, Russia
- 2Laboratory of Genetic Basis of Plant Identification, Vavilov Institute of General Genetics, Russian Academy of Sciences, Moscow, Russia
The genus Aegilops L. is the closest wild relative of wheat (Triticum L.), which contributed two of the three genomes to cultivated wheat. The genus Aegilops comprises 23 species differing in ploidy level and genome composition; diploid species possess the C, D, M, N, S, and U genome types, whereas various genome combinations are identified in tetraploid and hexaploid forms. The U genome is present in diploid Ae. umbellulata and eight polyploid species [Ae. triuncialis, Ae. biuncialis, Ae. geniculata, Ae. peregrina, Ae. kotschyi, Ae. columnaris, Ae. neglecta (4× and 6×), and Ae. juvenalis]. Some of these species have a wide distribution range, resulting in high adaptive capacity to various environmental conditions, and can serve as a valuable source of genetic diversity and useful genes for wheat breeding. The U genome is substantially rearranged relative to the genomes of common wheat, which hampers the direct transfer of useful traits from Aegilops to wheat. However, many genes conferring resistance to leaf rust (Lr9, Lr76, Lr57, Lr54, Lr59, Lr58), stripe rust (Yr70, Yr40, Yr37, Yr42), stem rust (Sr53), nematodes (CreX, CreY, Cre7), and various abiotic stresses have been successfully introgressed from Aegilops into the wheat genome. In this review, we describe the status of the contribution of Aegilops species carrying the U genome to wheat improvement, the methods used by different scientific teams to transfer genetic material, and the future prospective of exploitation of their useful traits in practical breeding.
Introduction
The genus Aegilops L. (family Poaceae) comprises 23 annual herbaceous species (Van Slageren, 1994; Kilian et al., 2011). Most of them are native to the Middle East and the Mediterranean, but several species also grow in other regions, such as northwestern Europe, Ukraine, Crimea, the Caucasus, north-Saharan Africa, Central Asia, and western China (Zhukovsky, 1928; Eig, 1929; Hammer, 1980; Boguslavsky and Golik, 2003; Kilian et al., 2011; Feldman and Levy, 2023). Several allotetraploid species have been introduced into the United States, of which Ae. cylindrica is now becoming widespread (Gandhi et al., 2009). The greatest diversity of Aegilops species is concentrated in the central part of the genus’ range, in the Fertile Crescent (Zhukovsky, 1928; Hammer, 1980; Van Slageren, 1994; Kilian et al., 2011). In such countries as Iran, Iraq, Syria, and Turkey, up to 13–17 of the 23 Aegilops species occur, while at the periphery of the range, e.g., in Hungary and Ukraine (eastern Europe), Kazakhstan (Middle Asia), or China and India (South-Eastern Asia), only 2–3 species are recorded (Zhukovsky, 1928).
Aegilops are classified into diploid (2n=2x=14), tetraploid (2n=2x=28), and hexaploid (2n=2x=42) species; at all ploidy levels, the species differ in genomic composition (Kihara, 1954; Hammer, 1980; Badaeva et al., 1996; Dvořák, 1998; Badaeva et al., 2002; Badaeva et al., 2021). Six main types of diploid nuclear genomes are currently distinguished among Aegilops species, in particular, S (the Sitopsis section), D (Ae. tauschii), M (Ae. comosa), N (Ae. uniaristata), U (Ae. umbellulata), and C (Ae. caudata) (Kihara, 1954; Kimber and Tsunewaki, 1988; Badaeva et al., 1996; Dvořák, 1998; Kilian et al., 2011; Sharma et al., 2021). The genome of Amblyopyrum muticum, which some taxonomists also classify as a member of the genus Aegilops (Zhukovsky, 1928; Ohta and Saruhashi, 1999), is designated by the symbol T (Hammer, 1980; Dvořák, 1998; Edet et al., 2018) (Table 1). Recent studies showed that the S genome of Ae. speltoides differs significantly from the S* genomes of other Sitopsis species (Dvořák, 1998; Ruban and Badaeva, 2018; Avni et al., 2022; Li et al., 2022) and is phylogenetically close to the T genome of A. mutica (Ohta and Saruhashi, 1999; Edet et al., 2018; Parisoid and Badaeva, 2020; Avni et al., 2022; Adhikari et al., 2023). Some diploid genomes, like D, S, and T, are not modified relative to the ancestral, and some modifications are found in the M genome, whereas U, C, and N genomes underwent significant structural rearrangements due to species-specific translocations and inversions (Iqbal et al., 2000; Danilova et al., 2017; Parisoid and Badaeva, 2020; Li et al., 2021; Said et al., 2021; Abrouk et al., 2023; Li et al., 2024; Singh et al., 2024).
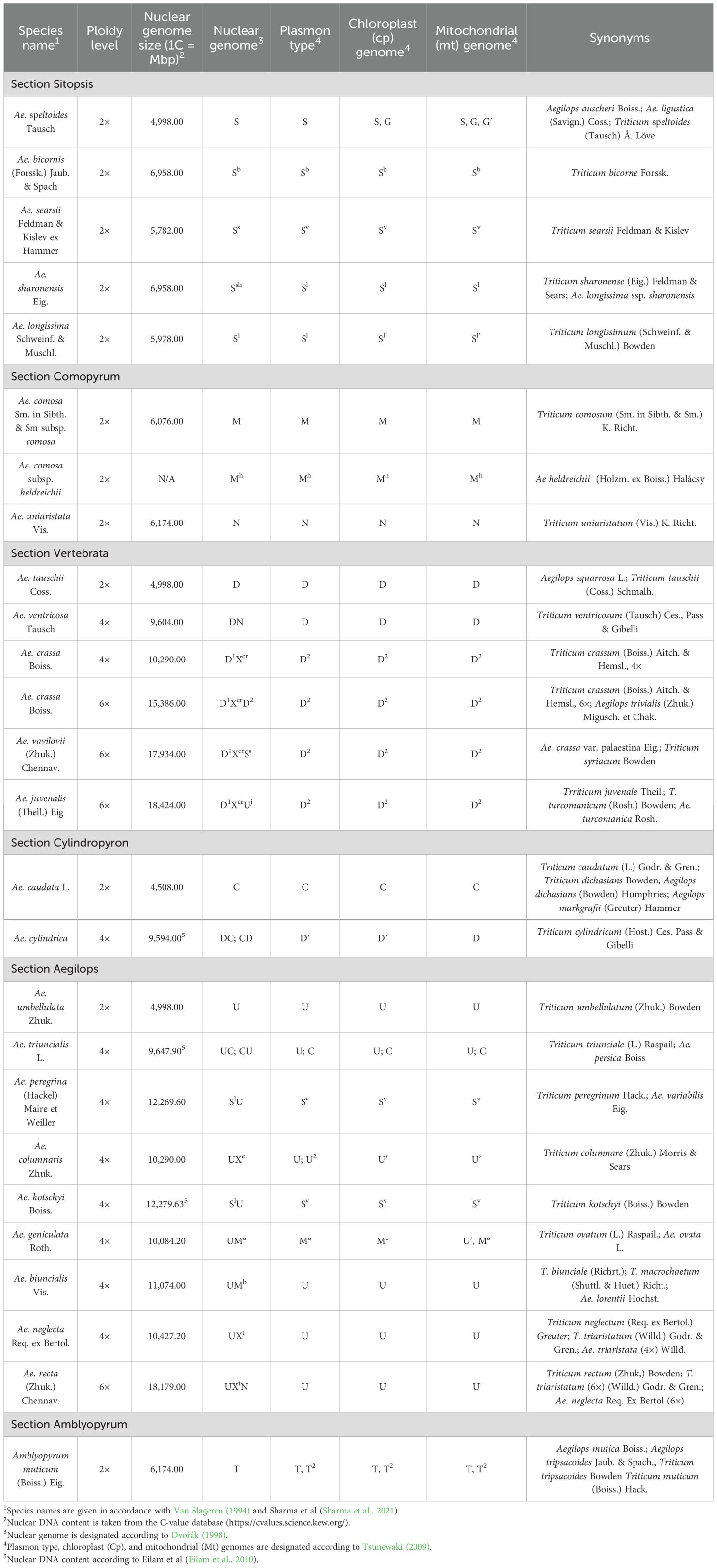
Table 1. Characteristics of genomes of Aegilops species.
The U genome of Ae. umbellulata is one of the best characterized genomes. Nearly complete sets of wheat-Ae. umbellulata chromosome addition, substitution, and translocation lines have been produced (Schneider et al., 2008). The homoeologous relationships of Ae. umbellulata chromosomes were established based on meiotic chromosome pairing behavior and compensation ability in substitution lines (Chapman and Riley, 1970; Athwal and Kimber, 1972; Riley et al., 1973; Chapman et al., 1975; Koebner and Shepherd, 1986; Reader and Miller, 1987), analysis of storage protein (Shepherd, 1973; Brown et al., 1979; Lawrence and Shepherd, 1981) and isozymes (Benito et al., 1987), and chromosome painting with bulked group-specific oligo-probes (Li et al., 2021). The species karyotype was constructed, and the intraspecific chromosome diversity of Ae. umbellulata was assessed by C-banding (Friebe et al., 1995) and FISH with various DNA probes (Badaeva et al., 1996; Molnár et al., 2011; Song et al., 2020). The U genome chromosomes were found to be highly asymmetric due to species-specific translocations (Figure 1A). Thus, according to RFLP analysis of wheat-Ae. umbellulata addition lines, the U genome diverged as a result of 11 translocations and inversions involving all seven chromosomes (Zhang et al., 1998). Results of physical mapping of cDNA probes and comparative single-gene FISH analysis confirmed these translocations and showed that 4U and 6U are the most rearranged chromosomes of Ae. umbellulata (Figure 1B). According to the authors’ data, four out of the seven group 4-specific cDNA probes were mapped on 6U and all the six group 6-specific probes—on 4U pointing to multiple reciprocal translocations between 4U and 6U. Based on these data, Zhang et al. (1998) and Said et al. (2021) suggested to correct the genetic classification of Ae. umbellulata chromosome by re-placing 4U and 6U. Another species-specific translocation involved chromosomes 3U and 7U (Said et al., 2021).
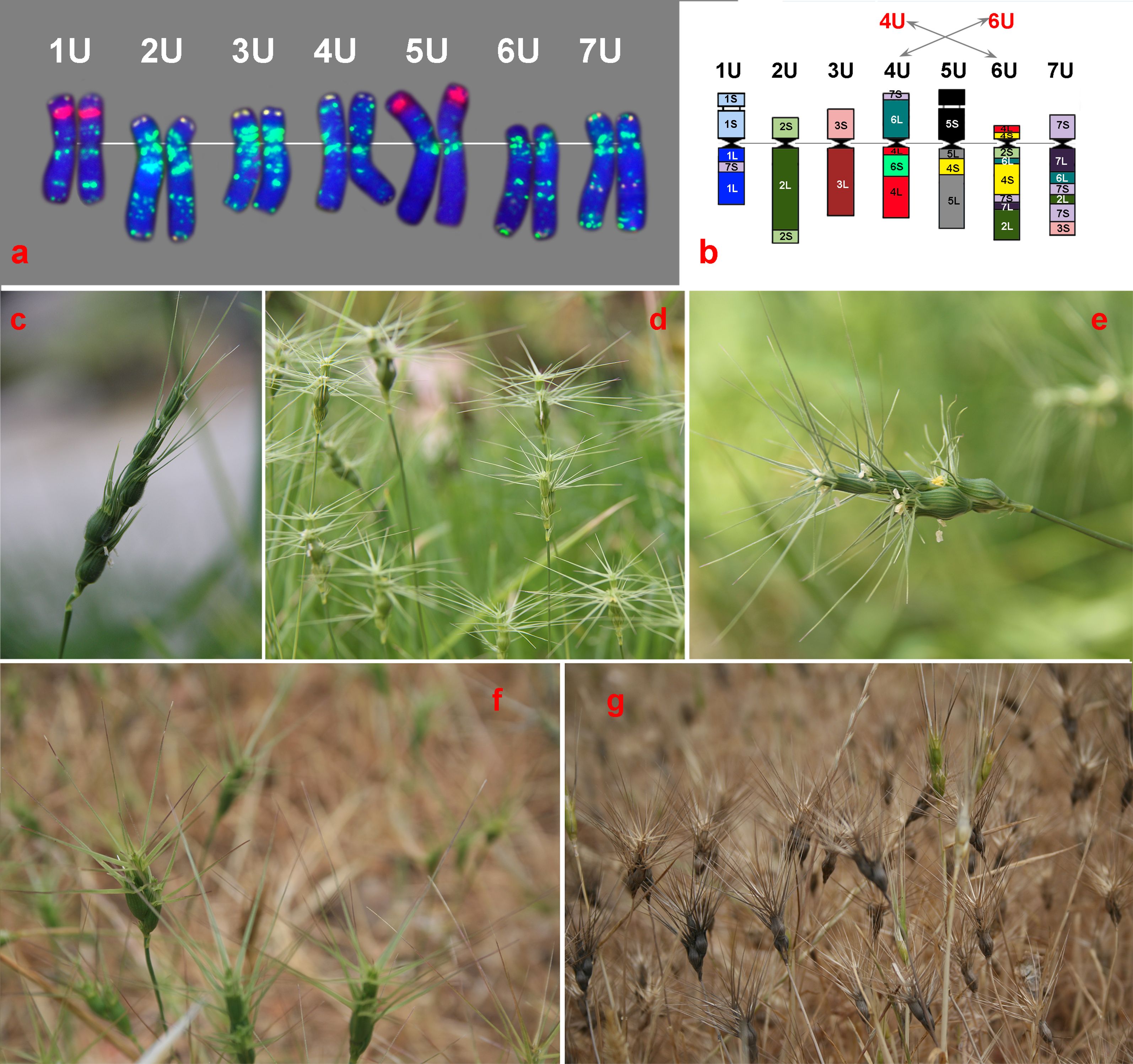
Figure 1. Chromosome organization of the U genome (a, b) and morphological variation of diploid Aegilops umbellulata (d) and some polyploid species of the U genome group: (c) Ae. juvenalis, (e) Ae. columnaris, (f) Ae. peregrina, and (g) Ae. geniculata. (a) Distribution of GAAn (green), pTa71 (red), and pSc119.2 (yellow) probes on Ae. umbellulata (AE 822) visualized by FISH; (b) the structural rearrangements of the U genome chromosomes [modified after Said et al (Said et al., 2021)].
Sequencing and chromosome-scale assembly of Ae. umbellulata, acc. TA1851 (Abrouk et al., 2023), as well as high-quality near telomere-to-telomere genome assembly (acc. PI 554389 (Singh et al., 2024), confirmed significant structural reorganization of the U genome. As in other Aegilops species, the major part of the U genomes was found to be composed of mobile elements, particularly retrotransposons. Genome assembly of PI 554389 allowed annotation of 78,076 gene models, 48,366 of which were high-confidence (HC) genes. A total of 2,162 among the HC genes were resistance gene analogs (RGAs) (Singh et al., 2024). This finding implies the great potential of Ae. umbellulata as a source of resistance genes for wheat improvement.
Polyploid Aegilops species have different genome compositions (Kihara, 1954; Hammer, 1980; Kimber and Tsunewaki, 1988; Dvořák, 1998; Badaeva et al., 2002; Kilian et al., 2011; Badaeva et al., 2021). Based on the presence of the “pivotal” genome, Kimber and Feldman (1987) divided them into two main clusters: the U genome cluster and the D genome cluster. The U genome cluster includes eight species: tetraploid Ae. triuncialis, Ae. kotschyi, Ae. peregrina, Ae. biuncialis, Ae. geniculata, Ae. columnaris, Ae. neglecta, 4x, and hexaploid Ae. neglecta subsp. recta (thereafter Ae. recta) (Table 1). In addition, the U genome is found in hexaploid Ae. juvenalis, belonging to the D genome cluster (Kihara et al., 1959; Kimber and Feldman, 1987; Dubkovsky and Dvořák, 1995). Analysis of Ae. juvenalis using C-banding and FISH revealed significant rearrangements of the Uj genome chromosomes relative to Ae. umbellulata (Badaeva et al., 2021). Most species from the U genome cluster grow in the Mediterranean and the Middle East, while Ae. juvenalis occurs mainly in the Middle and Central Asia (Figure 2).
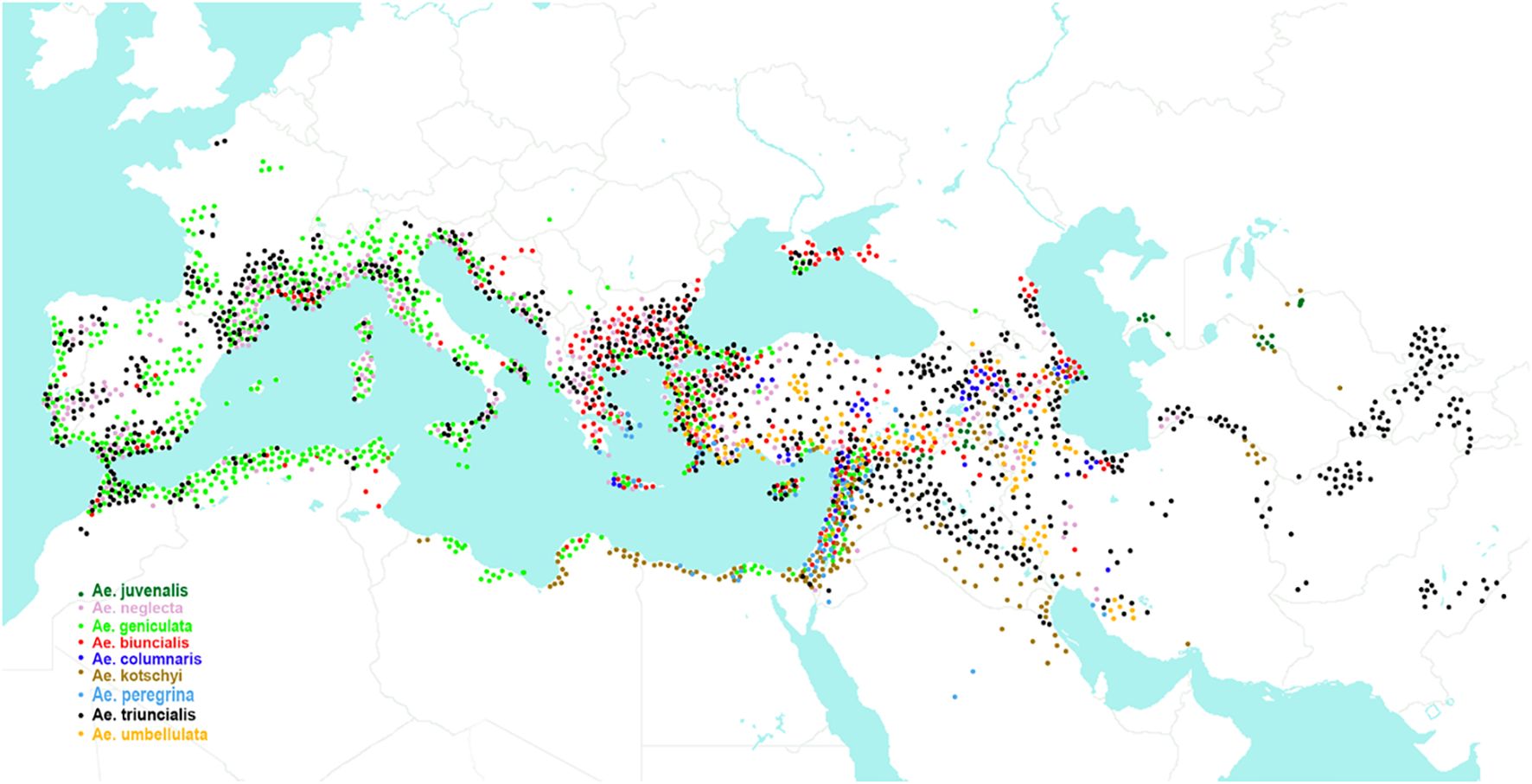
Figure 2. Distribution area of Aegilops species containing the U genome.
The cytoplasmic genome (plasmon) of five species of this group is identical or slightly modified relative to the cytoplasmic genome of Ae. umbellulata, indicating that it was their maternal parent and cytoplasm donor (Table 1). The plasmon of Ae. kotschyi and Ae. peregrina corresponds to the cytoplasmic genome of Ae. searsii, while the plasmon of Ae. juvenalis is identical to that of Ae. crassa (Tsunewaki, 2009). The origin of Ae. geniculata plasmon is not clearly understood, since it differs significantly from other Aegilops species. A proposed hypothesis suggests its inheritance from Ae. mutica (Maan, 1977; Tsunewaki, 2009).
Phylogenetically, the genus Aegilops is closely related to the genus Triticum and played a major role in the evolution of wheat (Levy and Feldman, 2022). Hexaploid bread wheat Triticum aestivum and tetraploid durum wheat T. durum share two common genomes, A and B, which are supplemented by the D genome in hexaploid wheat (Kihara, 1944; McFadden and Sears, 1946; Lilienfeld, 1951; Dvořák et al., 1998; El Baidouri et al., 2017; Levy and Feldman, 2022). The A genome of T. durum and T. aestivum is most similar to the Au genome of diploid wild wheat T. urartu (Dvořák et al., 1988; Dvořák, 1998; Badaeva et al., 2015). The immediate donor of the B genome is not exactly determined, but it is thought to derive from currently extinct species related to Ae. speltoides (Badaeva et al., 1996; Petersen et al., 2006; Salse et al., 2008; Marcussen et al., 2014; Avni et al., 2022; Levy and Feldman, 2022; Adhikari et al., 2023). The donor of the D genome of hexaploid wheat was Ae. tauschii (Kihara, 1944; McFadden and Sears, 1946; Lelley et al., 2000; Mizuno et al., 2010; Luo et al., 2017; Singh et al., 2019). Thus, the species from the genus Aegilops contributed two of the three genomes to bread wheat (Kihara, 1954; Hammer, 1980; Marcussen et al., 2014; Sharma et al., 2021).
Methods for transferring genetic material from Aegilops species to bread wheat
Aegilops species are a valuable genetic resource for expanding the genetic diversity of bread and durum wheat (Feldman and Sears, 1981; Gill et al., 1985; Boguslavsky and Golik, 2003; Colmer et al., 2006; Mujeeb-Kazi et al., 2009; Imadi and Gul, 2019; Kishii, 2019; Kumar et al., 2019; Sharma et al., 2021). Many species of this genus, including Ae. umbellulata and the U genome polyploids, exhibit broad morphological (Figures 1C–G) and genetic diversity (Dhaliwal et al., 1991; Damania et al., 1992; Monneveux et al., 2000; Zaharieva et al., 2001; Boguslavsky and Golik, 2003; Zaharieva et al., 2003; Molnár et al., 2004; Smith et al., 2004; Ahmadpoor et al., 2014; Ivanizs et al., 2019; Singh et al., 2024), driven by adaptation to diverse eco-geographical conditions (Van Slageren, 1994; Zaharieva et al., 2001; Kilian et al., 2011). Consequently, individual species may contain accessions with contrasting expression of traits, such as drought and heat tolerance (Molnár et al., 2004) and rust resistance (Gill et al., 1985; Valkoun et al., 1985).
Many Aegilops species, including the U genome group, do not share common genomes with bread wheat and, according to Harlan and de Wet (1971), belong to the secondary or even tertiary gene pool. Direct transfer of genetic material from such forms to bread wheat is challenging due to the non-crossability of the species and hybrid lethality. Significant modification of the U genome relative to wheat genomes (Zhang et al., 1998; Said et al., 2021; Abrouk et al., 2023; Singh et al., 2024) is an additional constraint restricting its direct utilization in wheat breeding, because structural rearrangements of homoeologous chromosomes decrease their compensation ability and pairing capacity (Zhang et al., 2015; Laugerotte et al., 2022).
A standard scheme for transferring genetic material from species of the secondary and tertiary gene pool, including Aegilops, involves the following steps. First, identification of potential donors of target traits by screening accessions from diverse geographic regions. The selected Aegilops accession is then crossed to bread wheat to produce F1 hybrids. The embryos of such hybrids are mainly weak and underdeveloped and often lack the endosperm (Bai et al., 1994). To overcome these problems and recover viable plants, the embryo rescue technique has been developed (Laibach, 1925; Sharma et al., 1996; Rogo et al., 2023). Chromosome doubling in F1 plants is usually induced by colchicine treatment (Blakeslee and Avery, 1937), though spontaneous doubling may also occur through fusion of unreduced gametes or autonomous genome duplication in an allohaploid hybrid (Pyralov; Matsuoka et al., 2014; Fakhri et al., 2016). Alien addition and substitution lines are produced by backcrossing the resulting amphidiploid by bread wheat (Feldman and Sears, 1981). Due to the tendency toward elimination of alien chromosomes over successive generations of interspecific hybrids (Blüthner et al., 1988; Bai et al., 1994; Zhang et al., 2015; Badaeva et al., 2024), it is necessary to monitor the inheritance of Aegilops chromosomes using morphological, molecular, or cytological markers.
Since alien chromosomes, in addition to target genes, contain linked loci coding undesirable traits (“linkage drag”), geneticists and breeders are aiming to reduce the size of the introgressed chromosome fragment by inducing wheat–alien translocations (Knott and Dvořák, 1976; Qi et al., 2007). Normally, homoeologous wheat and alien chromosomes do not pair in meiosis (Riley et al., 1959; Naranjo and Benavente, 2015; Perničková et al., 2019; Laugerotte et al., 2022); thus, methods such as irradiation, gametocidal (Gc) factors, genes-promoters of homoeologous pairing, or Ph1 (Pairing homoeologous 1—the key gene regulating meiotic pairing in wheat) suppressors are employed to induce the translocations (Qi et al., 2007; Zhang et al., 2015).
Irradiation and gametocidal factors cause random chromosome breaks, and fusion of the resulting fragments leads to the formation of translocations (Tsujimoto and Tsunewaki, 1985; Chen et al., 2009; Wang et al., 2012; Niranjana, 2017; Said et al., 2024). Most translocations will be non-compensatory, because they occur between non-homoeologous chromosomes (Knott and Dvořák, 1976; Qi et al., 2007; Rather et al., 2017). Rearrangements induced by X-rays cause a reduction of fertility and genomic stability in consecutive generations of the amphiploid and introgressive lines (Molnár et al., 2009; Wang et al., 2012; Wang et al., 2019). On the other hand, irradiation is effective in inducing translocations between chromosomes that do not pair in meiosis, in particular, wheat–alien translocations, since the presence of even a small translocated fragment at the end of a chromosome blocks its recombination with the normal homologous chromosome (Sears, 1976; Jiang et al., 1993; Laugerotte et al., 2022). Irradiation and a gametocidal system allow breaking the linkage between tightly linked loci and thus promote translocation of very small alien chromosome fragments (Masoudi-Nejad et al., 2002); irradiation is also effective in breaking the linkage between target and gametocidal genes (Ragini et al., 2024). Due to a random occurrence of irradiation-induced chromosomal breaks and rejoining, only a small fraction of translocations obtained by these methods found application in breeding (Friebe et al., 1996a; Qi et al., 2007; Kishii, 2019).
Several systems have been developed to produce wheat–alien translocations by inducing pairing and recombination of homoeologous chromosomes. These rely on either suppressing the Ph1, which regulates meiotic chromosome pairing in wheat, or introducing genes—promoters of homoeologous pairing from other species (Qi et al., 2007).
The system employing the recessive mutant of common wheat Chinese Spring (thereafter CS), designated CS ph1b, is most widely used for inducing wheat–alien translocations via recombination of homoeologous chromosomes. This mutant was obtained by E.R. Sears by irradiating immature pollen (before the onset of meiosis) of Chinese Spring wheat with subsequent pollination of the genetically marked CS line monosomic for 5B chromosome (Sears, 1977). Cytological (Gill and Gill, 1991) and molecular marker analyses (Gyawali et al., 2019) revealed that the ph1b mutation is caused by the deletion of a fragment of chromosome 5BL carrying the Ph1 gene. Another suppressor gene, Ph2, with a less pronounced effect on chromosome pairing, was mapped on 3DS (Mello-Sampayo, 1971; Sears, 1976). Recombination between wheat and Aegilops chromosomes can also be induced by using CS lines nullisomic or monosomic for chromosome 5B (Sears and Okamato, 1958; Riley and Kempanna, 1963; Koebner and Shepherd, 1986), as well as by the replacement of chromosome 5B by 5RL of rye in substitution or translocation lines (Bielig and Driscoll, 1970; Merker, 1992); however, none of these methods has been used as widely as CS ph1b. A temperature-sensitive mutation, ph1c, was later produced in durum wheat cultivar Cappelli using irradiation (Dvořák et al., 1984).
The frequency of recombination between wheat and Aegilops chromosomes can be increased by gene promoters of homoeologous pairing introduced into the wheat genome from other species. Such genes were found in Ae. speltoides (Dvořák et al., 2006; Li et al., 2017), Ae. peregrina (Farooq et al., 1990), Ae. umbellulata (Riley et al., 1973), Ae. geniculata (Koo et al., 2020), and Thinopyrum elongatum (Dvořák, 1987). Two dominant genes suppressing Ph1—the Su1-Ph1 and Su2-Ph1—were identified in Ae. speltoides and mapped on chromosomes 3S and 7S, respectively (Dvořák et al., 2006; Li et al., 2017). Chen et al. (1994) transferred the Su-Ph1 to the long arm of chromosome 4D of CS; the translocated chromosome was able to suppress Ph1 even in the monosomic state. The resulting line, designated CS Ph1, was actively used to induce heterologous translocations in wheat–Aegilops hybrids (Aghaee-Sarbarzeh et al., 2000; Kuraparthy et al., 2007b; Chhuneja et al., 2008). Another effective gene-promoter of homoeologous chromosome pairing, the Hpp-5Mg, was identified in the substitution Ae. geniculata–T. aestivum line DS 5Mg#1(5D) and mapped on chromosome 5MgS (Koo et al., 2020). A many-fold increase in the frequency of recombinant chromosomes was recorded in a double mutant ph1b ph1b/Hpp-5Mg Hpp-5Mg (Koo et al., 2017; Koo et al., 2020); the authors also noticed frequent occurrence of translocations with proximal breakpoints or interstitial translocations emerging due to the double recombination event.
Aegilops species as a source of economically valuable traits for improving bread wheat
Disease and pest resistance is among the most important traits transferred into wheat from the related species (Knott and Dvořák, 1976; Valkoun et al., 1985; Marais et al., 2003; Volkova et al., 2008; Olivera et al., 2018; Imadi and Gul, 2019; Sadeghabad et al., 2020; Kumar et al., 2022). According to the latest edition of the Wheat Gene Symbol Catalog (Wheat Gene Catalogue, 2024), over 60 genes conferring resistance to foliar diseases have been introduced from Aegilops into bread wheat including 26 genes of leaf rust resistance (caused by Puccinia triticina Eriks): Lr9, 21, 22a, 28, 32, 35, 36, 37, 39, 40, 41, 42, 43, 50, 51, 53, 54, 56, 57, 58, 59, 62, 66, 76, 85, and LrAc; 8 of yellow rust resistance (caused by P. striiformis Westend. f. sp. tritici): Yr8, 17, 28, 37, 38, 40, 42, and 70; 12 of stem rust resistance (caused by P. graminis Pers. f. sp. tritici): Sr32, 33, 34, 38, 39, 45, 46, 47, 51, 53, 62, and 66; and 16 of powdery mildew (caused by Blumeria graminis f. sp. tritici) resistance genes (Pm2a, 2mb, 12, 13, 19, 29, 32, 34, 35, 53, 57, 58, 66, 6Sl, Y39, and MlUM15). Aside from them, Aegilops also served as a source of resistance to a number of other diseases and pests: cereal aphid (Schizaphis graminum Rond.)—Gb3, 5, 7, and 9; Hessian fly (Mayetiola destructor (Say)—H13, 23, 26, 27, 30, and 34; cereal cyst nematode (Heterodera avenae Woll.)—Cre2, 4, 5, 6, 7, CreX, and CreY; root rot nematodes (Meloidogyne spp.)—Rkn1, 2, and 3; aphid (Diuraphis noxia (Mordvilko)—Dn3; Septoria tritici blotch (Mycosphaerella graminicola (Fuckel) Schroeter)—Stb5; Septoria nodorum blotch (Phaeosphaeria nodorum (E. Muller) Hedjaroude)—Sn3; mosaic virus—Wsm3; and others. There is no doubt that the genetic potential of Aegilops is far from being exhausted, and new promising sources of economically useful traits for breeding resistant wheat varieties can be found among them (Kishii, 2019; King et al., 2024).
Although Aegilops species from the U genome group do not share common genomes with wheat, they have already made a significant contribution to wheat improvement, especially in breeding for pest resistance. One of the most widespread and destructive diseases of bread wheat is rust, which occurs in nearly all regions of wheat cultivation (Singh et al., 2015; Schwessinger, 2017; Lewis et al., 2018; Prasad et al., 2020; Kumar et al., 2022; Patpour et al., 2022). Development of wheat varieties with genetically determined resistance remains the most effective and environmentally sustainable strategy to combat rust (Knott and Dvořák, 1976; Singh et al., 2015; Schwessinger, 2017; Kumar et al., 2022). Notably, several Aegilops species from the U genome group have already been donors of resistance genes against leaf, yellow, and stem rust (Table 2).
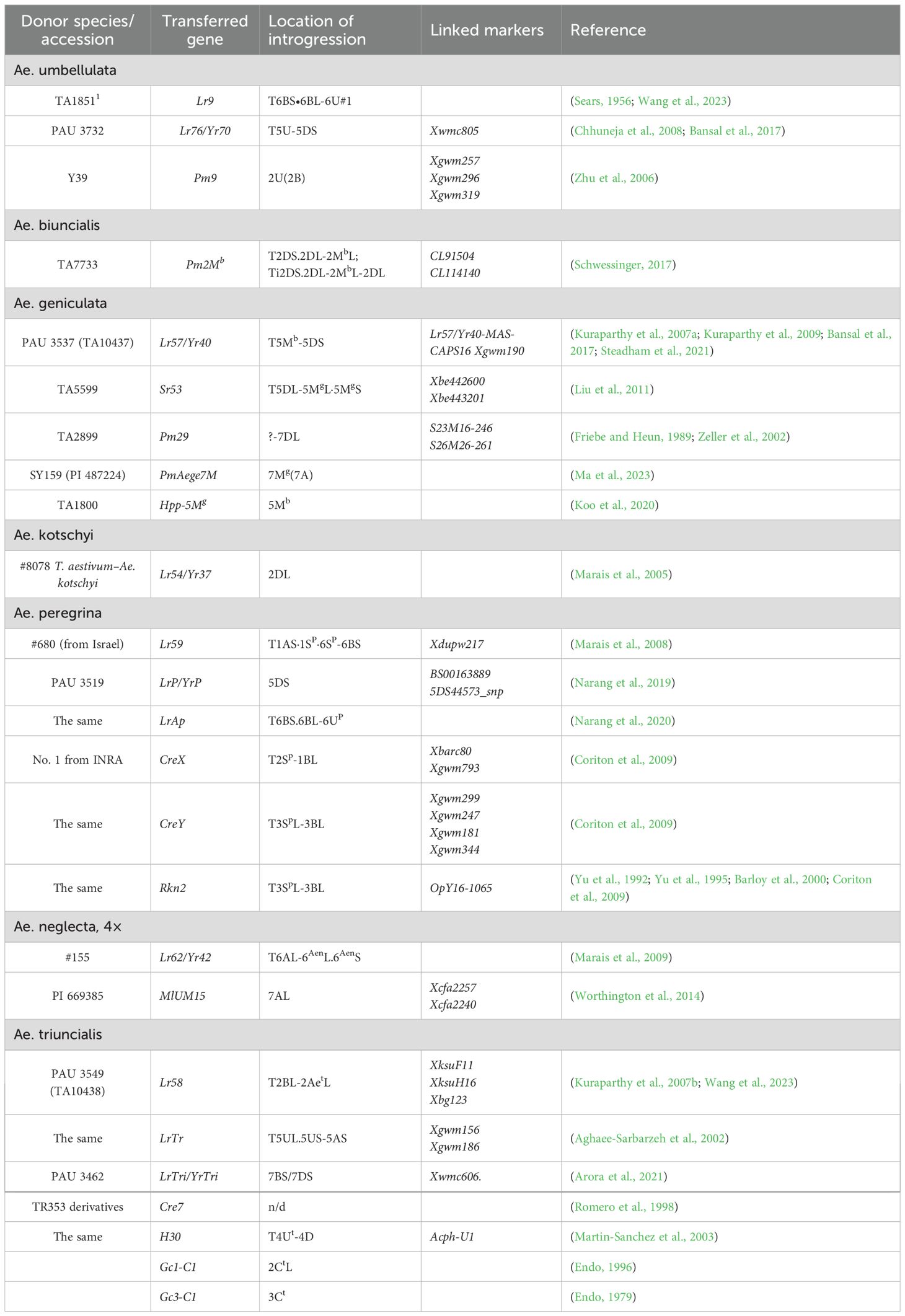
Table 2. Genes for economically valuable traits transferred into the wheat genome from the U genome-containing Aegilops species.
Aegilops umbellulata (Figure 1D): The first successful transfer of the leaf rust resistance gene from Ae. umbellulata to common wheat was performed by Dr. E.R. Sears (1956). He identified Ae. umbellulata accession with high resistance to leaf rust and crossed it with T. dicoccoides to produce an amphiploid, which, in turn, was crossed with CS. Following two backcrosses with CS and subsequent selection for resistance, Sears obtained a leaf rust-resistant wheat plant with Ae. umbellulata chromosome addition, which carried many disadvantageous phenotype modifications. Based on this line, he obtained an alien isochromosome addition composed of the resistance-determining arms. It was irradiated with X-rays to pollinate normal CS. In the progeny, Sears selected only resistant plants. Over 40 offspring carried one of at least 17 different wheat–Aegilops translocations; however, only one cytologically undetectable translocation showed normal transmission through the gametes. The line carrying this translocation was named “Transfer,” while the gene conferring its resistance was designated Lr9. C-banding and genomic in situ hybridization (GISH) analyses of the “Transfer” showed that the translocation designated T6BS•6BL-6U#1 occurred between homoeologous chromosomes and that the alien fragment is very small and located terminally in the long arm of wheat chromosome 6B (Friebe et al., 1996a). Although this introgression causes some yield reduction (Knott and Dvořák, 1976; Ortelli et al., 1996a; Ortelli et al., 1996b), it was widely adopted in modern wheat breeding. According to GRIS (http://wheatpedigree.net), the Lr9 is present in 79 common wheat cultivars, including Abe, Arthur-71, Agra, Agripro-Mason, Agripro-Natchez, Chiarano, Clemson 201, Coker-762, Coker-833, Coker-9024, Fundulea-29, McNair-701, Oasis, Pennmore, Riley-67, Sullivan, Tribute, etc., and in 49, it was proven to derive from Ae. umbellulata. A number of Lr9-carrying breeding lines have been developed in the Ural region of Russia and in Kazakhstan (Gultyaeva et al., 2024). Most Lr9 cultivars were produced and grown in North America, and their ratio is rising with time (Martynov et al., 2015). Lr9 is still effective in many wheat-growing areas, despite virulent Pt isolates emerging soon after the release of the first Lr9-carrying cultivars (Huerta-Espino et al., 2008).
Recently, Wang et al. (2023) cloned the Lr9 gene through mutagenesis and transcriptome sequencing and demonstrated that it encodes an unusual tandem kinase fusion protein. Long-read sequencing of the “Transfer” and Ae. umbellulata accession TA1851, which was identified as a putative donor of the Lr9 gene, enabled reconstruction of the introgressed ~28.4-Mb Lr9 region and precise localization of translocation breakpoints. The loss of a 5.58-Mb fragment from chromosome 6B, containing 87 high-confidence predicted genes, was identified. This deletion, combined with linkage drag from undesirable Ae. umbellulata genes co-introgressed with Lr9, may cause yield reduction in translocation-carrying lines (Huerta-Espino et al., 2008). A telomere-to-telomere genome assembly of Ae. umbellulata PI 554389 and resequencing 20 other Ae. umbellulata genomes identified six new haplotypes of Lr9 (Singh et al., 2024). Several T. aestivum–Ae. umbellulata introgressive lines resistant to leaf rust (mediated by the Lr9 gene) have been recently developed by Tyryshkin and Kolesova (2020).
New genes conferring resistance to leaf and stripe rust were identified in other accessions of Ae. umbellulata and used to develop introgressed disease-resistant wheat lines. For instance, Chhuneja et al. (2008) transferred two novel leaf rust resistance genes and a stripe rust resistance gene from Ae. umbellulata accession #3732 to bread wheat (Table 2). They crossed an amphidiploid WH890 (Ae. umbellulata × T. durum), line WH890, with CS PhI to induce homoeologous pairing between wheat and Ae. umbellulata chromosomes. Their F1 hybrid was crossed with the susceptible bread wheat cultivar WL711, and resistant plants were selected in the backcross progeny. According to rust reaction and allelism tests, the resistant lines split into two groups. The first group (exhibiting leaf rust resistance at the seedling stage) carried the Lr9 gene, while the second group harbored at least two novel leaf rust resistance genes (temporarily designated LrU1 and LrU2) and one stripe rust resistance gene (YrU1). Targeted GISH analysis revealed that the first lines possessed a complete Ae. umbellulata chromosome. Two lines with no apparent linkage drag carried translocations of Ae. umbellulata genetic material to the short arm of chromosome 5D. These lines can serve as promising donors of resistance genes for breeding programs (Özgen et al., 2004; Chhuneja et al., 2008).
Genetic mapping revealed that two of the newly identified genes, Lr76 (LrUmb) and Yr70 (=YrU1), are closely linked and located at the distal end of chromosome 5DS, being 7.6 cM distal to the marker Xgwm190-5D (Bansal et al., 2017). The introgressed region spanned approximately 9.47 Mb and was predicted to contain six NB-LRR (nucleotide-binding site leucine-rich repeat) (Bansal et al., 2020).
The powdery mildew resistance gene Pm9 was introduced by Zhu et al. (2006) from Ae. umbellulata accession Y39 into bread wheat using a complex combination of crosses. First, they produced a mildew-resistant amphidiploid Am9 by crossing Ae. umbellulata × Triticum carthlicum and crossed it with the susceptible bread wheat cultivar Laizhou953, followed by three backcrosses with Laizhou953 to obtain a resistant cultivar. Microsatellite analysis revealed a complete 2B(2U) substitution; among others, SSR markers Xgwm257/185, Xgwm296/130, and Xgwm319/180 co-segregated with powdery mildew reaction, and Xgwm296 was “resistance-dominant” (Zhu et al., 2006). All these group 2-specific wheat markers were amplified only in Ae. umbellulata Y39 and in the resistant plants, indicating that the resistance was controlled by the Ae. umbellulata chromosome, presumably 2U.
Aegilops biuncialis possesses a complex of valuable traits that can be used in wheat breeding. It is characterized by high adaptability, in particular salt tolerance, drought tolerance, and heat resistance (Table 3). Aegilops biuncialis is more drought-resistant than wheat (Molnár et al., 2004). To transfer drought tolerance from Ae. biuncialis to bread wheat, wheat cultivar Mv9kr1 was crossed with Ae. biuncialis accessions MvGB470 and MvGB1112 (Logojan and Molnár-Láng, 2000). The resulting amphiploids were more tolerant to drought and were able to maintain photosynthetic processes under moderate reduction of water content during osmotic stress (Dulai et al., 2014). The highest drought tolerance was recorded in T. aestivum–Ae. biuncialis lines Ae9041 and Ae9061 with 6Ub chromosome addition (Zhao et al., 2013).
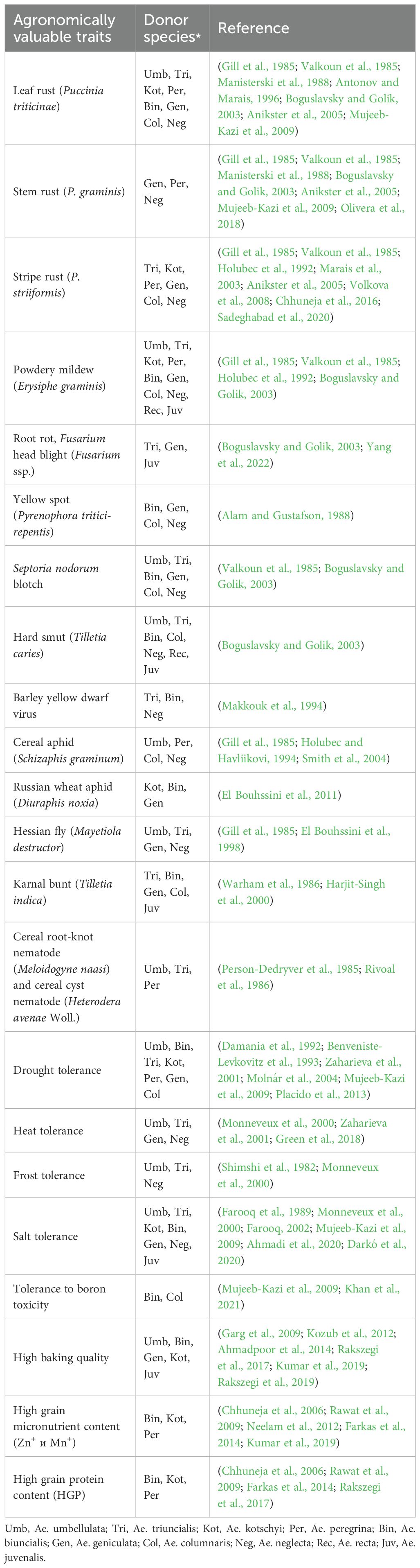
Table 3. Agronomically valuable traits of Aegilops species containing the U genome potentially useful in wheat breeding.
Aegilops biuncialis is a useful source of genes for salt tolerance. This complex trait occurs via different mechanisms controlled by many genes. According to current concepts, plant tolerance to elevated Na+ content can be developed through a limited Na+ uptake and transport between roots and shoots, Na+ excretion through the cytosol, and plant protection from salt-induced osmotic stress through the accumulation of osmoprotectants (Liang et al., 2018). The latter are water-soluble sugars, proline, and glycine betaine, the main organic substances that accumulate in plant tissues in response to biotic stresses (Ashraf and Foolad, 2007). Testing of parental forms and a series of introgressed Ae. biuncialis × T. aestivum lines for salt tolerance revealed that the wild type, as well as introgression lines with the addition of chromosomes 2Mb and 3Mb and translocation of 3Mb.4BS, showed higher germination potential, shoot and root growth, better CO2 assimilation capacity, and lesser chlorophyll degradation upon salt stress compared to the parental wheat form. The paternal Aegilops form accumulated less Na+ in the roots due to the upregulation of the SOS1, SOS2, and HVP1 genes, leading to an increase in the content of proline and a number of sugars (Darkó et al., 2020). Low Na+ levels in the leaves were accompanied by an increase in proline content due to enhanced expression of the NHX2 gene. Plants with chromosome additions 3Mb and 3Mb.4BS were characterized by accelerated accumulation of sugars and proline in the roots, whereas in the chromosome 2Mb addition line, an excess of Na+ in the leaves was excreted into vacuoles due to overexpression of HVP1 in the roots and NHX2 in the leaves. These observations pointed to different mechanisms of salt tolerance controlled by these chromosomes (Darkó et al., 2020). In addition to Na+ tolerance, some Ae. biunicalis accessions were also tolerant to boron toxicity and, thus, can serve as valuable donors of this trait in wheat breeding (Khan et al., 2021).
Triticum aestivum × Ae. biuncialis introgressive lines with 3Mb and 3Mb.4BS chromosome additions also showed an increased content of microelements in grain (Farkas et al., 2014), while wheat–Ae. biuncialis 1Ub addition line carried alleles of high-molecular weight glutenin subunits (HMG-GS), associated with high baking quality (Zhou et al., 2014). The addition of Ae. biuncialis group 5 and 7 chromosomes to wheat resulted in an increase in the protein and dietary fiber content in grain, thereby improving the nutritional value of flour (Rakszegi et al., 2017; Rakszegi et al., 2019).
Aegilops biuncialis is characterized by high resistance to powdery mildew and moderate resistance to leaf rust, but is affected by Hessian fly and cereal aphid (Gill et al., 1985). Resistance to stem rust in this species turned out to be race-specific (Olivera et al., 2018).
To transfer powdery mildew resistance from Ae. biuncialis to bread wheat, Men et al. (2022) crossed disomic 2Mb-addition CS line TA7733 with CS ph1b to induce homoeologous recombination between 2Mb and bread wheat chromosomes 2A/2D. This resulted in 65 recombinants, classified into 12 variants using 2Mb-specific molecular markers. The authors determined the size of the introgressed fragment using GISH and tested recombinant lines for powdery mildew resistance. As a result, a novel resistance gene, temporarily designated Pm2Mb, was identified and localized on the long arm of chromosome 2Mb within the interval of fraction length (FL) 0.49 to 0.66 (Men et al., 2022). Molecular markers were developed for this gene to facilitate tracing of this introgression in breeding material, while two recombinants with the smallest introgression size could be of greatest interest for the selection of mildew-resistant wheat cultivars.
Testing of Ae. biuncialis for resistance to barley yellow dwarf mosaic virus resulted in the identification of several resistant accessions that originated from Bulgaria; i.e., this species can potentially serve as a donor of resistance genes to this disease (Makkouk et al., 1994).
Aegilops geniculata (Figure 1G) is one of the most promising donors of agronomically valuable traits including disease and pest resistance (Table 3). Currently, at least four Ae. geniculata-derived disease resistance genes have been introduced into common wheat.
Leaf rust resistance gene Lr57 was identified by Aghaee-Sarbarzeh et al. (2002) in the disomic substitution line DS 5Mg(5D) obtained from a cross between leaf and stripe rust-susceptible wheat cultivar WL711 and the resistant Ae. geniculata accession #3537 (=TA10437). To reduce the size of the introgressed fragment, homoeologous recombination between 5D of wheat and Ae. geniculata 5Mg chromosomes was induced using the PhI-mediated system (Aghaee-Sarbarzeh et al., 2002). Putative recombinants were tested for rust resistance and analyzed by GISH and with SSR markers. GISH identified three types of introgressions: one was complete chromosome substitution, and the second was a translocated chromosome composed of the whole short arm and most part of the long arm of 5Mg with a small fragment of the wheat chromosome. The third introgression (recombinant line 2K-11-1) was cytologically undetectable (“cryptic” translocation). The recombinant chromosome consisted of the whole short arm and the major part of the long arm of 5D and a small part of 5MgL (nearly 15% of the 5MgL arm length). The cryptic introgression, which harbors the novel leaf and stripe rust resistance genes Lr57 and Yr40, comprised less than 5% of the 5DS arm (Aghaee-Sarbarzeh et al., 2002; Dhaliwal et al., 2002; Kuraparthy et al., 2007a; Kuraparthy et al., 2009). To map novel genes, the line TA5602 carrying cryptic introgression was crossed with susceptible wheat–Ae. geniculata 5Mg(5D) addition line to develop a mapping panel for identifying target genes. The 5Mg-specific markers were generated based on data of next-generation sequencing of flow-sorted 5Mg chromosome, which allowed localizing Lr57 and Yr40 within a physical interval of ~1.5 Mb (Steadham et al., 2021).
Another gene, Sr53, conferring resistance to stem rust Ug99 races RKQQC and TTKSK, was transferred from the T5DL-5MgL.5MgS chromosome to the long arm of bread wheat chromosome 5D by inducing heterologous recombination in a hybrid of translocation line TA5599 × CS ph1b (Liu et al., 2011; Kumar et al., 2022). Nine spontaneous recombinants with the reduced introgressed fragment of 5Mg were identified. Three recombinants carried interstitial translocations (T5DS.5DL-5MgL-5DL), which retained the proximal part (~20%–30%) of the introgressed 5MgL segment. Six lines possessed T5DL-5MgL.5MgS with a shortened 5MgL fragment (32%–45% of the total arm length). Based on analysis of the recombinant lines using GISH and 5D- and 5Mg-specific SSR/STS-PCR markers, all lines were considered as genetically compensated, and three Sr53-carrying lines were recommended as new sources of rust resistance (Liu et al., 2011).
Powdery mildew resistance gene Pm29 was transferred from Ae. geniculata to the bread wheat line Pova from the addition line VI T. aestivum cv. Poros–Ae. geniculata (accession TA2899) (Friebe and Heun, 1989). Using monosomic analysis and AFLP mapping, Pm29 was localized on the long arm of wheat chromosome 7D (Zeller et al., 2002).
Another novel powdery mildew resistance gene, tentatively designated PmAege7M, was identified in the 7Mg(7A) disomic substitution line W16998, obtained from a cross between CS and mildew-resistant Ae. geniculata accession SY159=PI 487224 (Wang et al., 2022; Ma et al., 2023). In addition to resistance, the W16998 exhibited a number of agronomically valuable traits, such as high tillering, high grain number, and 1,000-grain weight, making it a promising breeding material. The line W19513, isolated from the same cross combination and identified as 3Mg disomic addition, showed a moderate level of resistance to powdery mildew and high resistance to stripe rust (Wang et al., 2021).
Two CS × Ae. geniculata substitution lines (W623 and W637), which were derived from the 7Mg disomic addition line W166, showed high resistance to powdery mildew, yellow rust, and Fusarium head blight (FHB); resistance to FHB in wheat–Ae. geniculata lines was found for the first time (Yang et al., 2022). The alien chromosome was identified as 7Mg using GISH, FISH, EST-STS, and PLUG markers for different homoeologous groups. As a result, a disomic substitution 7Mg(7A) was identified in W623 as well as a disomic substitution 7Mg(7D) in W637. With the help of functional or linkage markers to FHB resistance genes or QTL, Yang et al. (2022) proved that W623 and W637 possess novel gene(s) controlling resistance to FHB, which are located on chromosome 7Mg of Ae. geniculata. Both 7Mg substitution lines and their parental form W166 were totally immune to powdery mildew race E09 at the seedling and adult stages and nearly immune to stripe rust. Thus, these lines could serve as promising material for developing wheat varieties with improved complex disease resistance.
Aegilops geniculatа can also act as a source of tolerance to abiotic stressors. In particular, it can be a potential donor of salt (Gorham, 1990; Farooq, 2002; Colmer et al., 2006) and drought tolerance (Zaharieva et al., 2001; Zaharieva et al., 2003; Pradhan et al., 2012a; Pradhan et al., 2012b), and the Ae. geniculatа–T. aestivum 3Mg introgressive line was characterized by resistance to copper ions (Landjeva et al., 1998). Chromosomes 1Ug and 1Mg have a positive effect on the baking quality of wheat flour (Garg et al., 2009; Garg et al., 2016; Rakszegi et al., 2017; Kumar et al., 2019; Rakszegi et al., 2019), while 1Ug chromosome addition improved rheological properties of dough by changing the composition and microstructure of gluten (Guo et al., 2021). Aegilops geniculata showed high micronutrient content in the leaves and grain (Rawat et al., 2009; Kumar et al., 2019) and can serve as a useful donor of these traits in wheat biofortification.
Aegilops geniculata can be used in wheat breeding as a carrier of gametocidal (Gc) genes. A new Gc gene causing chromosomal breaks in gametophytes, which lost the Gc factor, was identified on chromosome 4Mg of Ae. geniculata (Kynast et al., 2000). Kwiatek et al. (2017) induced androgenesis at post-meiotic pollen divisions in monosomic plants of hexaploid triticale (AABBRR) with 4Mg addition, followed by production of dihaploids (DHs), to maintain the chromosome aberrations caused by the gametocidal action. As a result, they obtained 41 DH lines, 17 of which carried different types of chromosomal aberrations in a homozygous state. Aegilops geniculata also possess genes affecting chromosome pairing. Thus, Koo et al. (2017) identified a new homoeologous pairing promoter gene(s) Hpp-5Mg on the chromosome 5Mg in the CS DS5Mg#2(5D) double disomic substitution line. The Hpp-5Mg gene significantly increased chromosome pairing and recombination frequency between wheat and homoeologous alien chromosomes, especially when combined with ph1b, permitting to induce of crossing over not only in the distal but also in the proximal regions of chromosomes, resulting in proximal and interstitial translocations (Koo et al., 2020).
Aegilops kotschyi possess a number of valuable traits that are in demand in breeding (Table 3). Antonov and Marais (1996) discovered Ae. kotschyi accession from Israel resistant to brown and yellow rust pathotypes common in South Africa (Antonov and Marais, 1996; Marais et al., 2003). They crossed it with CS and tested their offspring for rust resistance. The resistant form was backcrossed by CS, and the line 8078 containing a pair of Ae. kotschyi addition chromosomes was selected. The line had tenacious glumes inherited from the wild parent. In wheat, this trait is controlled by the dominant Tg gene localized on chromosome 2DS introgressed from Ae. tauschii (Wheat Gene Catalogue, 2024); thus, the alien chromosome could belong to homoeologous group 2. The line was crossed with three monosomic lines (CS M2A, CS N2B, CS M2D), and the resistant F1 plants with 2n=42, probably with group 2 wheat/group 2 Aegilops monosomic chromosome substitution, were selected from each cross and backcrossed as male parent to CS or another susceptible variety. The TF1 progeny was tested and the resistant plants were raised to TF2 families, which were again tested for leaf rust resistance. The alien chromosomes in monosomic addition lines (2n=42w+1Ae) always showed a low transmission rate, which was confirmed by screening TF1 and TF2 progenies for rust resistance.
Based on this knowledge, Marais et al. (2005) proposed that a higher transmission rate could be an indicator of compensating translocation with alien chromosome carrying resistance genes. Indeed, one population, S14, obtained in a test cross of the double monosomic 2D/Ae. kotschyi group2//CS contained 96% of the resistant plants and apparently carried a translocated chromosome (Marais et al., 2005). The authors selected resistant plants with 2n=42, which displayed normal threshability, shorter stature, and a much shorter growth period compared to the addition line. These plants presumably carried Robertsonian translocation between T2DS and the long arm of an undefined group 2 chromosome of Ae. kotschyi (Marais et al., 2005; Heyns et al., 2011). The translocated chromosome showed normal transmission through female gametes, but strong preferential transfer of male gametes carrying the resistance genes (Marais et al., 2005). The authors suggested that Ae. kotschyi chromosome present in the addition line 8078 possessed two linked genes conferring resistance to leaf rust and yellow rust designated Lr54/Yr37. These genes were localized on the long arm of Ae. kotschyi chromosome, which was translocated on 2DS in line S14 (Kuraparthy et al., 2007a). To reduce the size of the introgressed chromatin, Heyns et al. (2011) induced recombination between T2DS-2AekL and 2DL by using the CS ph1b system and obtained 10 stable recombinant lines with shortened alien fragment. Recombinant translocations presumably occurred due to single crossovers, leading to the replacement of a distal part of the alien chromosome fragment with wheat chromatin. Translocation recombinants were characterized using microsatellite, SCAR, and AFLP markers for 2DL. The resistant recombinants were classified into three categories according to the length of alien chromatin retained in the recombinant chromosomes, and the shortest alien fragment occupying the proximal half of the chromosome was detected in recombinant #74 (Heyns et al., 2011). To assess the perspectives of Lr54/Yr57 in wheat breeding, the authors developed near-isogenic lines of rec. #74 in adapted varieties and developed a dominant SCAR marker for target genes and several microsatellite markers linked to Lr54/Yr57 to trace them in breeding material (Heyns et al., 2011).
A similar translocation, but between the rye chromosome 2RS and the 2SkL chromosome of Ae. kotschyi, has later been produced by Ulaszewski et al. (2019) by crossing the ditelosomic (40T + D2RS + D2RL) line of triticale cv. Sekundo × monosomic substitution line of triticale carrying a single copy of 2Sk chromosome. GISH confirmed the presence of Robertsonian translocation between rye and Aegilops chromosomes, whereas FISH identified Ae. kotschyi chromosome as 2Sk (Ulaszewski et al., 2019). The application of molecular markers developed earlier for Lr54/Yr57 genes (Heyns et al., 2011) to recombinant triticale lines revealed an amplicon of the expected size, which indicated that generated 2Sk.2R RobTs translocation carried the same resistance genes and the recombinant chromosome described by Marais et al. (2005) composed of 2DS and Ae. kotschyi chromosome 2SkL.
Aegilops kotschyi is characterized by high grain microelement content (HGMC) compared to cultivated wheat and can be used as a valuable donor in wheat breeding (Chhuneja et al., 2006; Rawat et al., 2011; Verma et al., 2016a; Verma et al., 2016b; Kumar et al., 2019). To transfer GMC, Rawat et al. (2009) crossed the CS PhI line and wheat cultivars lacking the PhI gene with Ae. kotschyi, which was chosen from 90 Triticum and Aegilops accessions based on the three- to fourfold higher content of iron and zinc and ~33% higher protein content in grains. Most F1 hybrids produced in these crosses were sterile because of meiotic disturbances (Rawat et al., 2009). The sterile F1 hybrids were backcrossed as female parent with wheat, and some of the obtained BC1 hybrids and their BC2 progeny showed nearly 60% higher grain ash iron and zinc content than the recurrent wheat parent, suggesting that the useful variability of Ae. kotschyi for higher ash Fe+ and Zn+ concentration has been transferred to wheat.
Thirteen BC2F2 and BC1F3 lines obtained from the cross CS (PhI) × Ae. kotschyi (acc. 396), which were morphologically similar to the recurrent wheat parent and showed high microelement and protein content in grain, were selected for the assessment of introgressions by using morphological traits, HMW-GS profiles, wheat microsatellite markers, meiotic analysis, and GISH (Tiwari et al., 2010; Rawat et al., 2011). The authors found that HGMC correlates with the presence of addition and substitution of group 2 and 7 chromosomes of Ae. kotschyi. The 2Sk addition was found in 10 out of 13 derivatives with the enhanced grain protein and Zn+ concentration, and the group 7 chromosome introgression was identified in 10 lines (Rawat et al., 2011). Molecular markers for group 2 and 7 chromosomes revealed that the introgressions (additions and substitutions) were represented by whole chromosomes. Besides 2Sk and 7Uk/7Sk, hybrid lines may contain other Ae. kotschyi chromosomes, but they have no effect on the target traits. The plants with exceptionally high grain iron and zinc contents possessed additional HMW-glutenin subunits inherited from Ae. kotschyi #396, showing that 1Uk/1Sk chromosome(s) introgressions conferred very high grain iron and zinc content in the derivatives. These chromosomes also had the greatest effect on increasing grain micronutrient content.
One line containing the whole chromosome 7Sk and a translocated 2Sk chromosome with a fragment of wheat chromosome on the tip of the short arm did not show any increase in grain protein content (GPC); it had waxy leaves and spikes. The low protein content in this derivative suggests that the regulatory gene for high GPC in Ae. kotschyi #396 may be on the telomeric end of chromosome 2SkS (Rawat et al., 2011).
To transfer the HGMC from Ae. kotschyi to wheat, Verma et al. (2016b) employed the system of induced homoeologous chromosome pairing by crossing Ae. kotschyi (acc. 3790) with Pavon-mono-5B variety. They obtained F1 plants with 35 and 34 chromosomes (monosomic for 5B), which were completely sterile. The F1 plants with 34 chromosomes (lacking 5B) showed high homoeologous pairing. They were crossed with the wheat variety PBW343 (Lt24 + Yr36), and the resulting partially fertile BC1F1 plants were backcrossed by PBW343 and their BC2F1 progenies were screened for pollen viability, chromosome number, and meiotic chromosome pairing. The somatic chromosome number in the offspring varied from 43 to 60, and different lines possessed introgressions such as whole Uk/Sk chromosome(s) as well as one to several terminal or interstitial translocations of variable size involving both wheat and Aegilops chromosomes. Analysis of HMW-GS profiles of the BC2F1 plants allowed the identification of three lines with 1Uk/1Sk introgression, all showing higher grain mineral micronutrient content. Some BC2F2 plants exhibited a 125% increase in Fe+ content (MB-27, 2n=42, 19II + 4I) and a 158% increase in Zn+ content (BM-35, 2n=57, 23II + 11I) relative to the PBW343 variety and can be used for wheat improvement.
Aegilops kotschyi can also be used as a source of resistance to unfavorable abiotic factors: high temperatures, salinity and drought (Benveniste-Levkovitz et al., 1993; Bocianowski and Prażak, 2022), and water stress (Shimshi et al., 1982). The cytoplasm of some Ae. kotschyi accessions is capable of inducing cytoplasmic male sterility (thermo-sensitive male sterility based on cytoplasm, K-TCMS), which is of considerable interest for hybrid wheat breeding (Mukai and Tsunewaki, 1979; Gupta et al., 2019; Li et al., 2019; Fan et al., 2024). Alloplasmic lines of bread and durum wheat with Ae. kotschyi cytoplasm can also be used as haploproducers (Tsunewaki et al., 1974; Mukai and Tsunewaki, 1979; Fan et al., 2024).
Aegilops peregrina (Figure 1F) is characterized by high resistance to a number of biotic and abiotic stressors. To date, genes controlling resistance to leaf rust (Lr59, LrP, and LrAp), stripe rust (YrP) (Marais et al., 2008; Marais et al., 2010a; Pirseyedi et al., 2015; Narang et al., 2018; Narang et al., 2019; Narang et al., 2020) and still uncharacterized dominant gene of stripe rust resistance (Liu et al., 2010), cereal cyst nematode (CreX and CreY) (Coriton et al., 2009), root-knot nematode (Rkn2) (Yu et al., 1990; Yu et al., 1995), and uncharacterized powdery mildew resistance gene (Spetsov et al., 1997) have been transferred from this species to the wheat genome.
The leaf rust resistance gene Lr59 was introgressed from Ae. peregrina to wheat chromosome 1AL. Resistant Ae. peregrina accession 680 from Israel was used to pollinate CS followed by chromosome doubling in hybrids by colchicine treatment. The resulting C1 plants were backcrossed by CS and by the wheat breeding line W84-17; each generation was tested for leaf rust resistance against five South African pathotypes (Marais et al., 2008). The Ae. peregrina-derived resistance was found to be controlled by monosomic addition chromosome (in CS background) and by a translocation chromosome (in W84–17 background). The translocation occurred spontaneously in the line selected from the 4th backcross with W84-17. The addition chromosome was assigned to homoeologous group 1, which agreed with the cytogenetic analysis—the added chromosome showed C-banding pattern similar to the 1Sp of Ae. peregrina (Friebe et al., 1996b). The translocation chromosome had normal female transmission, while male transmission favored resistant gametes. Meiotic, monosomic, and microsatellite analyses showed that the fragment of Lr59-carrying Ae. peregrina chromosome constituted almost the entire long arm of 1A; i.e., this translocation is most likely Robertsonian (Marais et al., 2008). Later, GISH revealed that the terminal region of the long arm of the full-length Lr59 translocation belonged to wheat chromosome; i.e., this was an interstitial translocation. Further study confirmed that the distal part of the original translocation is homoeologous to subtelomeric region of 6BS, so the structure of the rearranged chromosome can be described as 1AS-1LP-6SP-6BS (Pirseyedi et al., 2015).
To reduce the size of the alien segment, plants heterozygous for the full-length Lr59 translocation and homozygous for CS ph1b mutation were pollinated with either nulli-1A/tetra-1B or nulli-1A/tetra-1D CS plants. Eight potentially useful recombinant resistant lines have been found among selfed F2 and F3 progenies (Marais et al., 2010a). The obtained recombinants and resistant test-cross F1 progeny monosomic for 1A were analyzed using microsatellite markers to localize Lr59 on genetic and physical maps. Eight of the resulting recombinants, which inherited rust resistance, differed in size and localization of the introgressed region. Two resistant plants were nullisomic for 1A; thus, the Lr59 fragment was retained on a different chromosome. Probably, this gene was transferred to other wheat chromosome due to allosyndetic homoeologous recombination (Marais et al., 2010a). To identify the non-1AL chromosome(s) involved in the allosyndetic recombination events, the authors built a genetic map of one of the seven recombinants, which did not involve 1AL. The Lr59 was assigned to the linkage group 6B; the respective locus was mapped 0.5 cM distally from co-segregating SNPs IWA1495, IWA6704, IWA2098, and IWA969. Eleven additional SNPs were mapped within an interval 0.5–6.5 cM proximally of Lr59 (Pirseyedi et al., 2015). The authors showed that a homoeoallele of Xdupw217 was associated with the Lr59 alien insert and therefore can be used to trace the resistance gene in breeding populations. By using GISH, the authors found that all—the original and seven resistant recombined Lr59 translocations—contained a small, similar-sized subterminal wheat chromosome segment and recommended recombinant line Lr59–151 with the smallest introgressed segment for further use in wheat breeding.
Two novel linked genes conferring leaf and stripe rust resistance temporarily designated LrP and YrP have been introgressed to common wheat from another Ae. peregrina accession (pau3519). Narang et al. (2018) crossed it with CS PhI to induce homoeologous recombination and backcrossed the F1 hybrid by the rust-susceptible wheat cultivar WL711. Their hybrids were tested for rust resistance, and the resistant plants were backcrossed again by WL711. The resistant BC1F1 plants with good agronomic performance have been selfed over five generations to develop stable homozygous BC2F6 introgression lines (IL). Two lines, IL pau16058 and IL pau16061, with chromosome number 2n=42 were selected for further analyses. They were screened for resistance against six Puccinia triticina and two P. striiformis f. sp. tritici pathotypes at the juvenile stage and against a mixture of pathotypes of both pathogens prevailing in India at the adult plant stage. IL pau16061 was resistant only to the leaf rust, while IL pau16058 showed resistance to both diseases at all stages of development (Narang et al., 2018). Microsatellite analysis identified a number of alien introgressions of different size in both lines: in IL pau16061, the largest introgression was detected on homoeologous group 6, followed by group 2, whereas in IL pau16058, the largest introgression was observed on homoeologous group 2, followed by group 5. According to SSR analysis, the alien fragment, controlling resistance in IL16058, was translocated to group 5 chromosome of wheat. Later, based on nulli-tetrasomic analysis, the location of the resistance genes was specified to chromosome arm 5DS (Narang et al., 2019). Introgression showed no deleterious phenotypic effects; thus, Narang et al. (2018) suggested that it involved a small alien segment with minimum linkage drag and might be compensatory.
The IL pau16058 was then crossed with the susceptible wheat variety WL711 to develop F2:3 mapping population (Narang et al., 2019). Analysis of this population revealed that leaf and stripe rust resistance were inherited as dominant co-segregating traits and therefore were controlled by two independent genes, temporarily designated LrP and YrP. Genomic origin of the introgressed Ae. peregrina fragment was defined using COS and KASP markers, designed for the U genome chromosomes. The results showed that the introgression carrying leaf rust and stripe rust resistance genes in IL pau16058 probably originated from the short arm of 5Up chromosome of Ae. peregrina (Narang et al., 2019).
Using the Illumina Infinium iSelect 90K wheat array and resistance gene enrichment sequencing (RenSeq) markers, the LrP and YrP genes were mapped within the 4.19-cM stretch in the distal part of 5DS containing eight SNPs and one microsatellite marker. LrP and YrP co-segregated with markers BS00163889 and 5DS44573_snp and were flanked by SNP markers BS00129707 and proximally 5DS149010 (Narang et al., 2019).
Another dominant gene conditioning leaf rust resistance at all stages of plant development, temporarily designated LrAp (Narang et al., 2020), was found in the second line, IL pau16061, isolated earlier by Narang et al (Narang et al., 2018). Chromosome localization of the introgressed fragments was determined using GISH with genomic DNA of Ae. umbellulata (Up) and Ae. speltoides (S) as probes (Narang et al., 2020). Two introgressions have been identified: one from the Sp and the other from the Up chromosomes. Rehybridization of the same metaphase cells with chromosome-specific probes showed that the Up genome segment was introgressed to the end of the long arm of wheat chromosome 6B and the translocation can be described as 6UpL-6BL. This was a compensating translocation and the novel leaf rust resistance gene temporarily designated LrAp was localized on the genetic map of 6BL using KASP markers. Alien segment spans along the 2.91-Mb region at the telomeric end of wheat chromosome 6BL (Narang et al., 2020). The S genome introgression was observed in both resistant and susceptible plants, and therefore, it was not associated with resistance.
Spetsov et al. (1997) transferred resistance to powdery mildew from unspecified Ae. peregrina accession of the IWS-General Toshevo collection to the wheat cultivar Rusalka. According to cytological and electrophoretic analyses of HMW glutenins, resistance was controlled by chromosome 1Uр, introgressed either as a 1Up chromosome addition or 1Up(1B) substitution (Spetsov et al., 1997).
Besides the resistance to foliar disease, some Ae. peregrina accessions showed resistance to cereal cyst nematode (CCN) caused by Heterodera avenae and root-knot nematode (RKN) caused by Meloidogyne naasi (Person-Dedryver et al., 1985; Yu et al., 1990; Yu and Jahier, 1992; Yu et al., 1992; Yu et al., 1995; Barloy et al., 2000; Coriton et al., 2009). Person-Dedryver at al (Person-Dedryver et al., 1985). evaluated the collection of wild wheats for nematode resistance and identified several resistant accessions of Ae. umbellulata and Ae. peregrina. Resistance of Ae. peregrina was due to the ability to suppress the development of larvae into females and nematode reproduction (Person-Dedryver et al., 1985; Yu et al., 1990). CS was pollinated with resistant Ae. peregrina accession No. 1, and the F1 hybrid was backcrossed twice by wheat cultivars Lutin and Rescler. The BC2 plants were then selfed up to the BC2F5 generation, and at each stage, all plants were evaluated for nematode resistance and chromosome number. Two disomic addition lines have been isolated in the progeny from this cross: the BC2F2 line X35 was resistant to both nematodes (CreY and Rkn2, previously designated Rkn-mn1), and line N was resistant to CCN (CreX gene) (Jahier et al., 1998; Coriton et al., 2009). The addition line LX was obtained via three backcrosses of the resistant addition line X35 with the susceptible wheat cultivar Lutin. The stable resistant translocation line X8 was also derived from X35; all X8 plants had 42 chromosomes and showed regular chromosome pairing in meiosis. Yu et al. (1990) suggested that the resistance to root-knot nematode in X8 and X35 is controlled by a single dominant gene Rkn2 (previously Rkn-mn1). As a similar resistance gene was earlier revealed in some accessions of Ae. longissima, the donor of Ae. peregrina Sp genome, the authors proposed that Rkn-2 could be located on the Sp genome chromosome. Monosomic analysis and telocentric mapping of the translocation line X8 and the addition line LX showed that the Ae. peregrina segment carrying the Rkn2 and CreY genes was transferred on wheat chromosome 3B (Yu et al., 1995; Barloy et al., 2000). The alien segment was located distally on 3BL, and Rkn2 and CreY were independent of the centromere (Yu et al., 1995). GISH and SSR mapping (Coriton et al., 2009) were fully consistent with this suggestion. In wheat–Ae. peregrina addition line LX, four 3BL-specific SSR markers and three SSRs of 7BL, 2BS, and 4BS were detected. In the translocation line X8, all SSRs developed for 3BL amplified products specific for Ae. peregrina, while all wheat homoeoloci were absent. Based on these data, Coriton et al. (2009) proposed that a recombination event between the additional chromosome of LX line and wheat took place within the distal part of 3BL.
Jahier et al. (1998) suggested that a segment of Ae. peregrina chromosome carrying the CCN resistance gene (CreX) in a second line—D, a derivative of line N, was introgressed into unknown wheat chromosome. FISH identified the additional Ae. peregrina CreX-chromosome as 6Sp; however, SSR markers amplified on this chromosome were specific for groups 1, 2, 4, and 6 (Coriton et al., 2009). The SSRs of group 2 were assigned to deletion bins of distal, terminal, and centromeric regions of 2AS or 2DS; thus, the authors proposed that the addition chromosome should be classified as 2Sp. A CreX-homozygous line D3 with a much shorter Ae. peregrina segment was obtained by backcrossing translocation line D by Lutin (Barloy et al., 2000). Based on microsatellite analysis, the CreX gene in D3 line was translocated on 1BL being in the vicinity to the Xgwm818 SSR marker (Coriton et al., 2009).
In addition to resistance genes, Ae. peregrinа has a number of valuable traits that may be utilized in breeding (Table 3). According to Damania et al (Damania et al., 1992), Ae. peregrinа is one of the most drought-tolerant species in the genus Aegilops. Liu et al. (2015) tested over 80 wheat–Aegilops introgressive lines developed on a CS background and found that wheat–Ae. peregrina 4Sp and 3Up addition line showed excellent performance under drought stress and should therefore be deeply examined and included in breeding programs for drought resistance. According to Farooq et al (Farooq et al., 1989), Ae. peregrinа also had a high potential for enhancing the salt tolerance of wheat. Introgression of group 4 and 7 chromosomes doubled the grain microelement content in the respective addition lines compared to the recipient variety (Neelam et al., 2012). In addition, the ability of Ae. peregrinа cytoplasm to induce cytoplasmic male sterility (Mukai and Tsunewaki, 1979) can be utilized in hybrid wheat breeding.
Aegilops neglecta: Unlike most species from the U genome group, Ae. neglecta and Ae. recta have been little used for wheat improvement. Only three successful transfers of the resistance genes, particularly leaf and stripe rust resistance Lr62/Yr42 (Marais et al., 2009), other leaf and stripe rust resistance genes not characterized yet (Bai et al., 1994), and powdery mildew resistance gene MlUM15 (Marais et al., 2005), from these species to bread wheat have been reported.
A dominant powdery mildew resistance gene, temporarily designated MlUM15, was introgressed from tetraploid Ae. neglecta accession TTCC 223 (collected by D. Marshall, USDA-ARS in 1992 on a rocky hillside near the town of Karamusa in north-central Turkey) to common wheat (Marais et al., 2005). The winter bread wheat cultivar “Saluda” was crossed with Ae. neglecta followed by two backcrosses by the recurrent parent and subsequent selfing. The derived BC2F8 line NC-UM15 was resistant to powdery mildew, which is supposed to be controlled by a novel resistance gene. To map this gene, Worthington et al. (Worthington et al., 2014) developed a mapping population by crossing NC-UM15 × Saluda with subsequent selfing of the F1 plants to produce F2 seeds, whose 198 F2:3 families were tested for disease resistance. DNA was isolated from all F2 plants, which were used for developing F2:3 families. Depending on the testing results, the F2:3 families were classified as homozygous resistant, homozygous susceptible, or segregating. Worthington et al. (Worthington et al., 2014) pooled DNA isolated from 1) 10 F2:3 families with consistently resistant phenotypes, 2) 10 families with consistently susceptible phenotypes, 3) resistant parents, and 4) susceptible parents. These groups were screened using more than 300 SSR markers covering the whole genome of wheat. Markers showing polymorphism between the resistant parent + bulk, on one hand, and the susceptible parent + bulk, on the other, were preliminarily considered as linked to the resistance gene and were taken to screen the entire F2 population. Three dominant SNP markers, IWA2929, IWA4434, and IWA8057, were linked in coupling, and Xcfa2257 was linked in repulsion with the resistance allele. Eleven SSR, STS, and SNP markers provided a basic map for calculating recombination frequencies, and the resistance gene MlUM15 was mapped to a 1.2-cM interval between Xcfa2257 and Xcfa2240. The physical location on the tip of 7AL was confirmed by analysis of CS aneuploid lines (Marais et al., 2005).
Two linked genes conferring resistance to leaf and yellow rust have been introgressed to common wheat from Ae. neglecta #155 (Marais et al., 2009), which was selected among 877 Aegilops accessions belonging to 27 species as highly resistant to leaf rust (Antonov and Marais, 1996). This accession was crossed with CS, and their resistant F1 hybrid was backcrossed to CS four times to develop a nearly isogenic line with a shortened culm (CS-S). Following five backcrosses by wheat, a monosomic addition plant (8048) was obtained, and its progeny was selfed and tested for disease resistance (Marais et al., 2009). A broad range of infection types has been observed, probably because the gene designated Lr62 was localized on the addition chromosome assigned to group 3 based on RFLP analysis (Marais et al., 2003). The disomic addition line, 8048-44, was selected from 8048. The Ae. neglecta addition chromosome showed mixed homoeology to wheat chromosomes. Its short arm and a proximal part of the long arm were homoeologous to wheat group 6, but a distal region on the long arm was derived from group 3 Ae. neglecta chromosome (Marais et al., 2010b). The line 8048 had an extremely poor agrotype and expressed strong hybrid necrosis, but was highly resistant to leaf rust.
On the next step, Marais et al. (2009) aimed to transfer the resistance gene(s) to the homoeologous wheat chromosome and characterize the introgression using genetic markers. Heterologous chromosome pairing was induced by crossing a monosomic addition plant (derived from a backcross of 8048 × CS-S) with a CS double monosomic 3B/5B plant followed by selection of the resistant double monosomic F1 progeny with 19″ + 3′. Hybrids were crossed with CS ph1b and with the susceptible tester line. The obtained progeny was scored for disease resistance, the resistant plants were selfed, and their F2 progeny was scored again for resistance, fertility, and the absence of hybrid necrosis (Marais et al., 2009). Segregation ratio (3:1) obtained in the F3 progeny implied that the resistance was controlled by translocation rather than by the addition chromosome. Test-crosses with three CS lines monosomic for 3A, 3B, 3D revealed that the translocation involved wheat chromosome from a different genetic group. The respective wheat chromosome was first searched using monosomic and nulli-tetrasomic analysis, and after that, wheat chromosomes, selected as possible candidates, were tested with arm-specific microsatellite markers. Based on this analysis, wheat chromosome was identified as 6A (Marais et al., 2009). The spontaneous exchange between the addition chromosome in 8048–44 and 6AL of wheat that gave rise to the full-length Lr62 translocation probably occurred within group 6 homoeologous regions, close to and proximally from the chromosome 3 translocation breakpoint (Somo et al., 2017). As a consequence of translocation, all group 3-derived chromatin on Ae. neglecta addition chromosome was replaced with wheat 6AL chromatin.
The addition line 8048–44 also carried a promising yellow rust resistance gene (Yr42) effective against South African pathotypes of P. striiformis at the seedling stage (Marais et al., 2009). Resistance was not co-transferred in the translocation event and thus may occur within the group 3 homoeologous region. Therefore, it was possible to transfer this gene by inducing homoeologous chromosome pairing.
The Lr62 gene was efficient against a broad range of South African and Canadian pathotypes of P. triticina; however, its carrier had poor agronomical performance. To increase the commercial attractiveness of the obtained translocations, Marais et al. (2010b) attempted to reduce their size by inducing heterologous chromosome pairing. Plants heterozygous for Lr62/Yr42 translocation and lacking the Ph1 locus were crossed with CS nulli-6A/tetra-6B or nulli-6A/tetra-6D plants. Resistant (Lr62) test-cross F1 progeny was evaluated for the presence of three specific SSR markers, resulting in the identification of 41 recombinants. The Lr62/Yr42 was localized toward the distal end of 6AS, which was homoeologous to the translocated chromosome. Major structural differences found between the CS ph1b mutant and the translocated chromosome 6A were caused by a duplication that was probably inherent in CS ph1b. Four promising recombinants, which retained both Lr62 and Yr42 within a comparatively small introgressed region at the 6AS terminus, were isolated. Subsequent analysis of the recombinants and the original Lr62 line using 6A-specific microsatellite markers and GISH showed that in three lines, the exchanges occurred near the telomeric region of 6AS (Somo et al., 2017). As recombinant lines retained the Gli-A2 locus, their translocations should not adversely affect baking quality and can be recommended for practical use (Somo et al., 2017).
There was only one report on the introgression of agronomically valuable traits from hexaploid Ae. recta to wheat. Thus, Bai et al. (1994) crossed six Ae. recta accessions with good resistance to leaf and stem rust with two susceptible durum and four susceptible bread wheat cultivars. All F1 hybrids expressed Ae. recta-derived resistance to both diseases. The F1 hybrids were backcrossed to their wheat parents to produce BC1F1 plants, which had a much lower seed set (0%–7.14%) compared to F1 (seed set 12.50%–78.33%); both characters varied depending on genotypes of parental species. Meiotic analysis showed that the low female fertility of the F1 hybrids was caused by poor chromosome pairing, and only gametes with complete or nearly complete genomes were viable. The BC1 plants, obtained only from backcrosses of Ae. recta to common wheat, were backcrossed to the susceptible wheat parent one or more times until the fertility was recovered. The selfed progeny was tested for disease resistance, and three families containing resistant plants have been selected.
Nearly half of the BC1F1 plants (17 vs. 39) had chromosome numbers close to the expected (2n=53–56), whereas chromosome numbers in the remaining 22 lines ranged from 44 to 52. The BC2F2–BC2F4 plants were segregated for disease resistance. Most susceptible lines had 2n=42 (ranged from 40 to 44) and formed 21 bivalents in meiosis. Chromosome number of the resistant lines varied depending on cross combination from 2n=43 or 44 with 21″ + 1′ or 21″ + 1″ in one combination to 43–50 in two others (Bai et al., 1994). These data indicated that the resistance genes must be located in alien chromosomes added to wheat. Aegilops recta chromosome associated with disease resistance was identified using 14 RFLP markers specific for seven homoeologous groups of wheat (Bai et al., 1994). The addition chromosomes carrying leaf resistance genes in lines 1 and 2 were assigned to homoeologous groups 5 and 2, and the stem rust resistance gene was localized on the group 7 chromosome of Ae. recta (Bai et al., 1994).
Genetic potential of Ae. neglecta, however, is not fully exploited (Table 3). More than 80% of Ae. neglecta accessions were resistant to Ug99 races TTKSK, TTTTF, and TRTTF (Olivera et al., 2018). Some accessions showed multiple aphid resistance (El Bouhssini et al., 2011) and can be useful donors of this trait.
Aegilops triuncialis is geographically one of the most widespread Aegilops species, which is adapted to a broad range of biotic and abiotic stressors (Table 3). Many accessions of Ae. triuncialis showed high resistance to diseases (Gill et al., 1985; Harjit-Singh et al., 2000) and pests (Romero et al., 1998) and served as donors of these traits in wheat breeding (Table 2). Thus, Harjit-Singh et al. (2000) crossed susceptible wheat cultivars WL711 with Ae. triuncialis acc. 3549 (=TA10438) with complex resistance to leaf rust, Karnal bunt, powdery mildew, and cereal cyst nematode (CCN) and backcrossed the sterile F1 hybrid by the recurrent wheat parent. The resistant BC2/BC3 plants gave rise to two groups of derivatives. One group with 2n=42 was resistant to CCN and powdery mildew and moderately resistant to leaf rust, while the second group with 2n=44 possessed leaf rust, Karnal bunt, and powdery mildew resistance. C-banding analysis of the first group identified a complete 5A(5Ut) chromosome substitution, which was confirmed by GISH. GISH also revealed an additional, segregating wheat–Ae. triuncialis recombinant chromosome, possessing a small terminal translocation from the Ut chromosome to the short arm of wheat chromosome (Aghaee-Sarbarzeh et al., 2002). GISH on the second group of introgressive lines (2n=44) detected disomic addition of acrocentric Ae. triuncialis chromosome and Robertsonian translocation between the unknown wheat (long arm) and Ae. triuncialis (short arm) chromosomes (Harjit-Singh et al., 2000). The authors suggested that gene(s) conferring resistance to leaf rust, Karnal bunt, and powdery mildew could be linked and located either on wheat–alien translocated chromosome or on the alien acrocentric chromosome. Alternatively, gene(s) for resistance to one or two diseases could be split between different introgressions (Harjit-Singh et al., 2000).
Kuraparthy et al. (2007b) transferred the leaf rust resistance gene, designated Lr58, from Ae. triuncialis, #3549 to wheat. The introgression line was developed by crossing the susceptible cultivar WL711 with the resistant Ae. triuncialis accession #3549 and backcrossing the resistant F1 plants by WL711 (Harjit-Singh et al., 2000; Aghaee-Sarbarzeh et al., 2002). Resistant BC1F1 plants were selected, backcrossed by WL711, and selfed to develop BC3F11 lines (starting from BC2F1 and BC3F1, only resistant plants with a complete set of wheat chromosome were selected for subsequent selfing). The resistant BC3F11 line TA5605 had normal plant growth and development. This line, its parental forms Ae. triuncialis #3549, WL711, and CS, and wheat cultivar Jagger were subjected to molecular-genetic and cytogenetic analyses. The segregation ratio 3:1 (resistant:susceptible), obtained in the F2 population from the Jagger × TA5605 cross, suggested a monogenic dominant inheritance (Kuraparthy et al., 2007b). The authors failed to determine the chromosome location of the introgression using 23 5A-specific SSR markers. No signal from the alien segment was revealed by GISH, indicating that the introgressed segment is very small and cytologically undetectable. Polymorphic alleles between the resistant and susceptible bulks were revealed by the RFLP probe KSUF11, specific for group 2 of wheat. Eighteen additional 2BL-specific probes were further selected for characterizing the translocation in TA5605, and two of them, KSUH16 and BG123, showed Ae. triuncialis-specific diagnostic polymorphism between resistant and susceptible bulks. Three probes co-segregated with leaf rust resistance in the F2 mapping population. The authors concluded that the translocation in TA5605 involved the group 2 chromosome of Ae. triuncialis and the 2L arm of wheat chromosome and occurred through homoeologous recombination (Kuraparthy et al., 2007b). The physical location of the translocation breakpoint was deduced considering the position of resistance-associated/not-associated RFLP markers on the genetic map. Translocation breakpoint in TA5605 was localized in the deletion bin 2L-0.89–1.00 of the consensus physical map, and the size of the introgressed segment was <10% of the long arm of wheat chromosome 2BL (Kuraparthy et al., 2007b). Genomic origin of Ae. triuncialis chromosome (2Ut or 2Ct) involved in this cryptic translocation designated T2BS·2BL-2tL(0.95) remained unknown.
The leaf rust-resistant introgression line 2K-69–4 was produced by backcrossing CS5A(5Ut) substitution line with CS phI to induce heterologous chromosome pairing (Aghaee-Sarbarzeh et al., 2002). GISH revealed a homozygous translocation; the major part of the recombinant chromosome was from chromosome 5Ut, and only the distal part of the short arm corresponded to 5AS. Aegilops triuncialis accession #3549, DS5Ut(5A), and the translocation line 2K-69–4 were all resistant to the most virulent in Indian leaf rust pathotypes 77A-1 and 77-2. Moderate susceptibility of DS5Ut(5A) to leaf rust under field conditions (compared to high susceptibility of the recurrent wheat parent) suggested that chromosome 5Ut carried a gene conditioning slow rusting against the prevalent leaf rust races. Because the recombinant chromosome of 2K-69–4 consisted mainly of 5Ut, which may bring many agronomically undesirable genes, this translocation cannot be directly used for wheat improvement.
To transfer the mildew resistance gene located on 5Ut (Harjit-Singh et al., 2000) and to reduce the size of the introgressed fragment, the DS 5Ut(5A) line was crossed and backcrossed with Pavon ph1bph1b to obtain a hybrid homozygous for ph1b (Kamboj et al., 2020). The plants homozygous for ph1b and monosomic for 5Ut/5A were selected using molecular markers and crossed with WL711 to induce 5Ut-5A recombinants by homoeologous pairing. Homozygous recombinants were obtained after five rounds of selfing, and in each generation, only plants with square heads were propagated (“spelta” heads served as a marker for the absence of recombination, and plants were abounded). A total of 367 F4 substitution recombinant lines were screened for mildew resistance in the lab and under field conditions, and 68 F5 lines with varying incidence of powdery mildew (0–9 score) were genotyped using 29 wheat 5A-specific SSR markers. Of the 367 F5 lines, only eight were resistant to powdery mildew (two lines with spelt head and six with square head), while the remaining 259 were affected by the disease to varying degrees. The segregation ratio of the disease resistance presumed that it is governed by several genes. Analysis of 68 selected lines with different responses to infection using 18 transferable and polymorphic 5A-specific SSR markers allowed to identify three main regions of 5Ut—one on 5UtS/5AS and the other two on 5UtL/5AL, associated with powdery mildew resistance (Kamboj et al., 2020). The major resistance gene was apparently localized in the short 5Ut/5A arm. Meiotic analysis of four randomly selected mildew-resistant lines showed normal chromosome pairing (21″), indicating that the translocation of 5Ut fragments conferring mildew resistance into 5A was compensating (Kamboj et al., 2020). Noteworthy, the resistance gene(s), which was introgressed from Ae. triuncialis to wheat more than 25 years ago (Harjit-Singh et al., 2000), continued to maintain resistance against the diverse prevalence in Indian pathotypes of Blumeria graminis for over 18 years (Kamboj et al., 2020).
Introgressive DS 5Ut(5A) line (BTC17) also served as donor of the extra-soft grain texture trait controlled by puroindolines (Sharma et al., 2020). A total of 367 5U-5A substitution recombinant lines developed by Kamboj et al. (2020) were screened for the presence of Ae. triuncialis-derived Pina and Pinb, but only 23 of them were found to contain both alleles (Sharma et al., 2020). Grain hardness index of recombinants with functional puroindolines was reduced from 6% to 67%. Using the 5A-specific SSR markers, the authors established that the 5Ut fragment containing puroindoline loci was translocated to the telomeric end of wheat chromosome 5AS, which was confirmed by sequencing of Pina and Pinb genes (Sharma et al., 2020). The obtained lines could be in demand in breeding soft wheat varieties for the cookie and biscuit industries. As puroindoline proteins extracted from soft-grain lines exhibited antimicrobial activity against gram-positive and gram-negative bacteria, they can be utilized in the pharmaceutical industry.
Two novel genes controlling leaf and stripe rust resistance were introduced from Ae. triuncialis accession # pau 3462 to the susceptible wheat cultivar WL711 (Arora et al., 2021). Aegilops triuncialis was crossed and backcrossed by CS ph1b to induce recombination between Aegilops and wheat chromosomes. BC2 progeny with the recurrent plant type and resistant to both pathogens was selected in subsequent generations to develop the stable, resistant BC2F7 plants phenotypically similar to WL711. Inheritance and chromosome location of resistance genes were studied in the F2 and F2:3 mapping populations obtained by crossing the stable introgression line IL tri with the wheat cultivar WL711. According to the segregation ratio of resistant/susceptible plants, both diseases are controlled by single dominant genes, temporarily designated LrTri and YrTri, which were transferred to wheat as independent fragments (Arora et al., 2021). Based on molecular-genetic analysis with 614 SSR markers to all wheat chromosomes, the LrTri was mapped on chromosome 7BS/7DS at a distance of 11.2 cM from the SSR marker Xwmc606 (Arora et al., 2021), although the location of the yellow rust resistance gene remained unknown.
Aegilops triuncialis is highly resistant to cereal cyst nematode caused by H. avenae (Person-Dedryver et al., 1985; Benveniste-Levkovitz et al., 1993; Narang et al., 2019). To transfer this trait to wheat, Romero et al. (1998) obtained two vigorous male-sterile amphihaploids T. turgidum (H-1-1) × Ae. triuncialis (A-1), which were crossed with the bread wheat cultivar Almatense (H-10-15). The resulting hybrids were highly sterile, but produced eight seeds. The plants obtained from these seeds contained 28–41 chromosomes and were highly sterile. After eight rounds of selfing, 49 translocation lines (TR) with chromosome number 40 (95%) or 41 (5%) have been developed. The fertility of hybrids was still very low, but the surviving plants were vigorous. Low fertility was probably caused by the Ph-suppressing effect of the Ct genome (Romero et al., 1998) or by the action of Gc factors carried by Ae. triuncialis chromosomes 2Ct and 3Ct (Endo, 2007). Gc genes could also induce heterologous chromosome translocations in hybrids.
According to field tests, 9 of 49 TR lines were not infested with CCN pathotype Ha71, and one of these lines—TR-535—was chosen for further studies because of its good performance in all resistance tests. Segregation analysis of the BC5–6 families obtained from the TR-535 × T. aestivum cross showed that CCN resistance transferred from Ae. triuncialis is controlled by a single dominant gene tentatively designated CreAet (Romero et al., 1998), currently Cre7 (Wheat Gene Catalogue, 2024). The authors suggested that CCN resistance in TR-535 was derived from the Ct genome (Romero et al., 1998); however, the chromosome location of the Cre7 was not determined, and its genome origin still needs to be confirmed.
The same translocation line TR-353 (2n=41) and its euploid derivative TR-3531 (2n=42) also served as donors of Hessian fly resistance to wheat. Martin-Sanchez and co-authors (Martin-Sanchez et al., 2003) found that Ae. triuncialis-derived resistance is inherited as a single Mendelian factor, and the newly discovered gene designated H30 is non-allelic to already known genes. The resistance was associated with the Acph-U1 marker, which was also identified in Ae. triuncialis, Ae. umbellulata, and amphiploid CS-Ae. umbellulata, but was absent in wheat–Ae. umbellulata addition lines and in Ae. caudata. Based on these data, Martin-Sanchez et al. (2003) proposed that Hessian fly resistance was transferred from the Ut genome, and the H30 gene is located on a segment of chromosome 4Ut translocated to chromosome 4D of wheat. Importantly, the authors were able to introduce resistance to the Hessian fly (less often, together with the oat nematode) to 16 elite breeding lines; i.e., their material became a valuable source of these traits for wheat improvement.
Gametocidal chromosomes of Ae. triuncialis were first discovered in 1975 by Endo and Twunewaki (Endo and Tsunewaki, 1975). They noticed that all wheat–Ae. triuncialis alloplasmic lines were characterized by extreme female sterility. All wheat lines with substituted cytoplasm from Ae. triuncialis, without any exceptions, carried additional acrocentric chromosome from their cytoplasm donor. The author supposed that sterility was caused by the unviability of gametes lacking the alien chromosome, which were named gametocidal or “gamete killing” (Endo, 2015; Said et al., 2024). Two gametocidal chromosomes, 2Ct and 3Ct, were identified in Ae. triuncialis, which carried the Gc1-C1 and Gc3-C1 genes, respectively (Endo, 1979; Endo, 1996). Gc-chromosome 2Ct expressed mild, semilethal effects in all wheat varieties (Endo, 1982; Endo, 2007). Chromosomes 3Ct and its derivative 3CSAT expressed different actions in CS background: that of 3Ct was lethal but that of 3CSAT was semilethal (Endo TR, 1996). Structurally rearranged chromosomes of CS that emerged under the action of Gc factors of 2Ct and 3CSAT were cytologically identified and isolated, giving rise to a series of wheat deletion stocks (Endo TR, 1996), as well as to deletion stocks of rye (Masoudi-Nejad et al., 2002; Li et al., 2013; Endo, 2021) and barley (H. vulgare and H. chilense) chromosomes added to CS (Serizawa et al., 2001; Nasuda et al., 2005; Said and Cabrera, 2009; Sakai et al., 2009). Due to their ability to induce spontaneous chromosome breaks, Gc chromosomes were employed for reducing the size of alien introgressions. The ability of Ae. triuncialis cytoplasm to induce cytoplasmic male sterility could find practical application in hybrid wheat breeding (Tsunewaki et al., 1974).
Currently, no reports on the practical utilization of the genetic material of Ae. columnaris (Figure 1E) have been reported, although this species possesses a number of valuable traits, including resistance to diseases and different abiotic factors (Table 3). Three primary amphidiploids were produced in the All-Russian Research Institute of South-East, Saratov (ARISER) by crossing three common wheat cultivars, Dobrynya, Saratovskaya-68, and L-503 with Ae. columnaris (acc. k-1193). Hybrid seeds were well-developed and viable, but the F1 plants were sterile and were backcrossed to the recurrent wheat cultivars. The BC1 and BC2 progenies were either backcrossed again to wheat or self-pollinated. As a result, 57 introgressive lines with different substitutions and additions have been developed (Badaeva et al., 2018). According to preliminary observations, introgression of chromosome 5Xc was associated with leaf rust resistance in hot and dry conditions of the middle Volga region (Badaeva et al., 2018). Some substitutions were associated with enhanced salt tolerance (Dashtoian et al., 2023). The cytoplasm of this species induces cytoplasmic male sterility in wheat (Tsunewaki et al., 1974).
We failed to find any publications on the transfer of useful traits from hexaploid Ae. juvenalis (Figure 1C), although all accessions of this species are immune to powdery mildew (Valkoun et al., 1985; Boguslavsky and Golik, 2003) and highly resistant to Fusarium root rot (Boguslavsky and Golik, 2003). A relatively recent article reported the production of a decaploid hybrid Ae. juvenalis × durum wheat (Takumi et al., 2020), and hybrid and several introgressive lines of Ae. juvenalis with triticale have been developed, although not characterized further (Gradzielewska et al., 2012).
Conclusion
Summarizing the above information, Aegilops species containing the U genome (Table 1) have already contributed many valuable genes, especially disease resistance genes, some of which have found a broad application in common wheat breeding (Table 2). Thus, over 79 commercial wheat cultivars, predominantly from the US, possess the Ae. umbellulata-derived gene Lr9. Many other genes conferring leaf rust (Lr54, Lr76), stem rust (Sr53), yellow rust (Yr37, Yr40), powdery mildew (Pm9, Pm29), and nematodes (CreX, CreY, Cre7) have also been introgressed from species of the U genome group into wheat, although they have not been used as extensively as Lr9. All relevant publications demonstrate that Aegilops species carrying the U genome constitute a unique and highly promising genetic resource for improving cultivated wheat, not only with regard to disease resistance but also to other agronomically important traits. They can serve and have already been used as donors of draught, heat and salt tolerance, high micronutrient content, and high baking quality. The mechanisms underlying these traits, however, remain not fully understood and need further investigation. Detailed examination of the gene pool of Aegilops species, including the U genome group, holds a great promise for developing wheat varieties of the next generation that combine high productivity with multifaceted resistance to both biotic and abiotic stresses. There is no doubt that the genetic potential of the U genome group of Aegilops is far from exhausted and they can become donors of many new important traits (Table 3) and expand the genetic diversity of durum and common wheat. In summary, the U genome-bearing Aegilops species represent a unique genetic reservoir whose significance will only grow with the development of modern breeding technologies.
Author contributions
EB: Methodology, Supervision, Data curation, Formal Analysis, Conceptualization, Funding acquisition, Writing – review & editing, Writing – original draft. MG: Writing – review & editing, Data curation, Writing – original draft, Methodology, Formal Analysis. OR: Writing – review & editing, Methodology, Validation, Formal Analysis, Data curation. NT: Formal Analysis, Data curation, Writing – review & editing, Methodology. MD: Funding acquisition, Resources, Writing – review & editing, Project administration, Supervision.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The work was prepared with the financial support of the Russian Science Foundation grant no. 24-16-00145, https://ias.rscf.ru/user/doc/a.w.p.2024.92.legacy/199702.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
CS, Chinese Spring (wheat cultivar); IL, introgression line; CCN, cereal cyst nematode; RKN, root-knot nematode; FHB, Fusarium head blight; HMG-GS, high-molecular weight glutenin subunits; HGMC, high grain microelement content; GPC, grain protein content; GISH, genomic in situ hybridization; FISH, fluorescence in situ hybridization.
References
Abrouk, M., Wang, Y., Cavalet-Giorsa, E., Troukhan, M., Kravchuk, M., and Krattinger, S. G. (2023). Chromosome-scale assembly of the wild wheat relative Aegilops umbellulata. Sci. Data 10, 739. doi: 10.1038/s41597-023-02658-2
Adhikari, L., Raupp, J., Wu, S., Koo, D.-H., Friebe, B., and Poland, J. (2023). Genomic characterization and gene bank curation of Aegilops: the wild relatives of wheat. Front. Plant Sci. 14. doi: 10.3389/fpls.2023.1268370
Aghaee-Sarbarzeh, M., Ferrahi, M., Singh, S., Singh, H., Friebe, B., Gill, B., et al. (2002). Ph1-induced transfer of leaf and stripe rust-resistance genes from Aegilops triuncialis and Ae. geniculata to bread wheat. Euphytica 127, 377–3 82. doi: 10.1023/A:1020334821122
Aghaee-Sarbarzeh, M., Harjit-Singh, and Dhaliwal, H. (2000). Brief communication. Ph′ gene derived from Aegilops speltoides induces homoeologous chromosome pairing in wide crosses of Triticum aestivum. J. Heredity. 91, 417–421. doi: 10.1093/jhered/91.5.417
Ahmadi, J., Pour-Aboughadareh, A., Fabriki Ourang, S., Khalili, P., and Poczai, P. (2020). Unraveling salinity stress responses in ancestral and neglected wheat species at early growth stage: A baseline for utilization in future wheat improvement programs. Physiol. Mol. Biol. Plants. 26, 537–549. doi: 10.1007/s12298-020-00768-4
Ahmadpoor, F., Rasool, A.-Z., Behnam, F., and Shahbazi, H. (2014). Investigation of diversity in Aegilops biuncialis and Aegilops umbellulata by A-PAGE. Natural Product Res. 28, 1626–1636. doi: 10.1080/14786419.2014.931392
Alam, K. B. and Gustafson, J. P. (1988). Tan-spot resistance screening of Aegilops species. Plant Breeding. 100, 112–118. doi: 10.1111/j.1439-0523.1988.tb00225.x
Anikster, Y., Manisterski, J., Long, D. L., and Leonard, K. J. (2005). Resistance to leaf rust, stripe rust, and stem rust in Aegilops spp. in Israel. Plant Disease. 89, 303–308. doi: 10.1094/PD-89-0303
Antonov, A. I. and Marais, G. F. (1996). Identification of leaf rust resistance genes in Triticum species for transfer to common wheat. South Afr. J. Plant Soil. 13, 55–60. doi: 10.1080/02571862.1996.10634376
Arora, S., Kaur, S., Dhillon, G. S., Singh, R., Kaur, J., Sharma, A., et al. (2021). Introgression and genetic mapping of leaf rust and stripe rust resistance in Aegilops triuncialis. J. Genet. 100, 6. doi: 10.1007/s12041-020-01253-3
Ashraf, M. and Foolad, M. R. (2007). Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Botany. 59, 206–216. Available online at: https://www.sciencedirect.com/science/article/pii/S0098847206000268 (Accessed March 24, 2025).
Athwal, R. S. and Kimber, G. (1972). The pairing of an alien chromosome with homoeologous chromosome of wheatT. Can. J. Genet. Cytology. 14, 325–333. doi: 10.1139/g72-040
Avni, R., Lux, T., Minz-Dub, A., Millet, E., Sela, H., Distelfeld, A., et al. (2022). Genome sequences of three Aegilops species of the section Sitopsis reveal phylogenetic relationships and provide resources for wheat improvement. Plant J. 110, 179–192. doi: 10.1111/tpj.15664
Badaeva, E. D., Amosova, A. V., Goncharov, N. P., Macas, J., Ruban, A. S., Grechishnikova, I. V., et al. (2015). A set of cytogenetic markers allows the precise identification of all A-genome chromosomes in diploid and polyploid wheat. Cytogenetic Genome Res. 146, 71–79. doi: 10.1159/000433458
Badaeva, E. D., Amosova, A. V., Muravenko, O. V., Samatadze, T. E., Chikida, N. N., Zelenin, A. V., et al. (2002). “Genome differentiation in Aegilops. 3. Evolution of the D-genome cluster,” in Plant Systematics and Evolution, vol. 231. (Austria, Vienna: Springer-Verlag), 163–190. doi: 10.1007/s006060200018
Badaeva, E. D., Chikida, N. N., Belousova, M. K., Ruban, A. S., Surzhikov, S. A., and Zoshchuk, S. A. (2021). A new insight on the evolution of polyploid Aegilops species from the complex Crassa: molecular-cytogenetic analysis. Plant Systematics Evolution. 307, 3. doi: 10.1007/s00606-020-01731-2
Badaeva, E. D., Davoyan, R. O., Tereshchenko, N. A., Lyalina, E. V., Zoshchuk, S. A., and Goncharov, N. P. (2024). Cytogenetic peculiarities in interspecific amphiploid and genome-substituted forms of wheat. Vavilov J. Genet. Breeding. 28, 716–730. doi: 10.18699/vjgb-24-80
Badaeva, E. D., Friebe, B., and Gill, B. S. (1996). Genome differentiation in Aegilops. 1. Distribution of highly repetitive DNA sequences on chromosomes of diploid species. Genome 39, 293–306. doi: 10.1139/g96-040
Badaeva, E. D., Ruban, A. S., Shishkina, A. A., Sibikeev, S. N., Druzhin, A. E., Surzhikov, S. A., et al. (2018). Genetic classification of Aegilops columnaris Zhuk. (2n=4x=28, UcUcXcXc) chromosomes based on FISH analysis and substitution patterns in common wheat × Ae. columnaris introgressive lines. Genome 61, 131–143. doi: 10.1139/gen-2017-0186
Bai, D., Scoles, G. J., and Knott, D. R. (1994). Transfer of leaf rust and stem rust resistance genes from Triticum triaristatum to durum and bread wheats and their molecular cytogenetic localization. Genome 37, 410–418. doi: 10.1139/g94-058
Bansal, M., Adamski, N. M., Toor, P. I., Kaur, S., Molnár, I., Holušová, K., et al. (2020). Aegilops umbellulata introgression carrying leaf rust and stripe rust resistance genes Lr76 and Yr70 located to 9.47-Mb region on 5DS telomeric end through a combination of chromosome sorting and sequencing. Theor. Appl. Genet. 133, 903–915. doi: 10.1007/s00122-019-03514-x
Bansal, M., Kaur, S., Dhaliwal, H. S., Bains, N. S., Bariana, H. S., Chhuneja, P., et al. (2017). Mapping of Aegilops umbellulata-derived leaf rust and stripe rust resistance loci in wheat. Plant Pathology. 66, 38–44. doi: 10.1111/ppa.12549
Barloy, D., Lemoine, J., Dredryver, F., Jahier, J., and McIntosh, R. A. (2000). Molecular markers linked to the Aegilops variabilis-derived root-knot nematode resistance gene Rkn-mn1 in wheat. Plant Breeding. 119, 169–172. doi: 10.1046/j.1439-0523.2000.00451.x
Benito, C., Figuerias, A. M., and Gonzales-Jaen, M. T. (1987). Location of genes coding isozyme markers on Aegilops umbellulata chromosomes adds data on homoeology. Theor. Appl. Genet. 73, 581–588. doi: 10.1007/BF00289198
Benveniste-Levkovitz, P., Canaani, O., Gromet-Elhanan, Z., and Atsmon, D. (1993). Characterization of drought resistance in a wild relative of wheat, Triticum kotschyi. Photosynthesis Res. 35, 149–158. doi: 10.1007/BF00014745
Bielig, L. M. and Driscoll, C. J. (1970). Substitution of rye chromosome 5RL for chromosome 5B of wheat and its effect on chromosome pairing. Genetics 65, 241–247. doi: 10.1093/genetics/65.2.241
Blakeslee, A. F. and Avery, A. G. (1937). Methods of inducing doubling of chromosomes in plants by treatment with colchicine. J. Heredity. 28, 393–411. doi: 10.1093/oxfordjournals.jhered.a104294
Blüthner, W.-D., Schubert, V., and Mettin, D. (1988). “Instability in amphiploids and backcross derivatives of a Triticum aestivum x Ae. caudata cross,” in Proceedings of the 7th International Wheat Genetics Symposium(Cambridge: Bath Press, Avon).
Bocianowski, J. and Prażak, R. (2022). Genotype by year interaction for selected quantitative traits in hybrid lines of Triticum aestivum L. with Aegilops kotschyi Boiss. and Ae. variabilis Eig. using the additive main effects and multiplicative interaction model. Euphytica 218, 11. doi: 10.1007/s10681-022-02967-4
Boguslavsky, R. L. and Golik, O. V. (2003). The genus Aegilops L. as the genetic resourse for breeding Kharkov (Ukraine: Yuriev Institute of Plant Industry, NAAN), 129.
Brown, J. W. S., Kemble, R. J., Law, C. N., and Flavell, R. B. (1979). Control of endosperm proteins in Triticum aestivum (var. Chinese Spring) and Aegilops umbellulata by homoeologous group 1 chromosomes. Genetics 93, 189–200. doi: 10.1093/genetics/93.1.189
Chapman, V. and Riley, R. (1970). Homoeologous meiotic chromosome pairing in Triticum aestivum in which chromosome 5B is replaced by an alien homoeologue. Nature 226, 376–377. doi: 10.1038/226376a0
Chapman, V., Riley, R., and Miller, T. (1975). Alien chromosome addition and substitution lines (Cambridge: Plant Breeding Institute).
Chen, P. D., Bie, T. D., Cao, Y. P., Chen, S. W., and Wang, X. E. (2009). Irradiation-induced wheat-alien translocation lines and their application in wheat breeding (Rome, Italy: Food and Agriculture Organization of the United Nations (FAO).
Chen, P. D., Tsujimoto, H., and Gill, B. S. (1994). Transfer of Ph(I) genes promoting homoeologous pairing from Triticum speltoides to common wheat. Theor. Appl. Genet. 88, 97–101. doi: 10.1007/BF00222400
Chhuneja, P., Dhaliwal, H. S., Bains, N. S., and Singh, K. (2006). Aegilops kotschyi and Aegilops tauschii as sources for higher levels of grain Iron and Zinc. Plant Breeding. 125, 529–531. doi: 10.1111/j.1439%2D0523.2006.01223.x
Chhuneja, P., Kaur, S., and Dhaliwal, H. S. (2016). “Introgression and exploitation of biotic stress tolerance from related wild species in wheat cultivars,” in Molecular Breeding for Sustainable Crop Improvement, vol. 2 . Eds. Rajpal, V. R., Rao, S. R., and Raina, S. N. (Springer International Publishing, Cham), 269–324.
Chhuneja, P., Kaur, S., Goel, R. K., Aghaee-Sarbarzeh, M., Prashar, M., and Dhaliwal, H. S. (2008). Transfer of leaf rust and stripe rust resistance from Aegilops umbellulata Zhuk. to bread wheat (Triticum aestivum L.). Genet. Resour. Crop Evolution. 55, 849–859. doi: 10.1007/s10722-007-9289-3
Colmer, T. D., Flowers, T. J., and Munns, R. (2006). Use of wild relatives to improve salt tolerance in wheat. J. Exp. Botany. 57, 1059–1078. doi: 10.1093/jxb/erj124
Coriton, O., Barloy, D., Huteau, V., Lemoine, J., Tanguy, A.-M., and Jahier, J. (2009). Assignment of Aegilops variabilis Eig chromosomes and translocations carrying resistance to nematodes in wheat. Genome 52, 338–346. doi: 10.1139/g09-011
Damania, A. B., Altunji, H., and Dhaliwal, H. S. (1992). Evaluation of Aegilops spp. for drought and frost tolerance.
Danilova, T. V., Akhunova, A. R., Akhunov, E. D., Friebe, B., and Gill, B. S. (2017). Major structural genomic alterations can be associated with hybrid speciation in Aegilops markgrafii (Triticeae). Plant J. 92, 317–330. doi: 10.1111/tpj.13657
Darkó, É., Khalil, R., Dobi, Z., Kovács, V., Szalai, G., Janda, T., et al. (2020). Addition of Aegilops biuncialis chromosomes 2M or 3M improves the salt tolerance of wheat in different way. Sci. Rep. 10. doi: 10.1038/s41598-020-79372-1
Dashtoian, J. V., Kalinina, A. V., and Korobko, V. V. (2023). Effect of salinity on the morphogenesis of seedlings of introgressive lines of spring bread wheat with genetic material Aegilops columnaris. Saratov Univ New Ser. Ser. Chem. Biol. Ecol. 23, 308–317. doi: 10.18500/1816-9775-2023-23-3-308-317
Dhaliwal, H. S., Harjit-Singh, Gupta, S., Bagga, P. S., and Gill, K. S. (1991). Evaluation of Aegilops and wild Triticum species for resistance to leaf rust (Puccinia recondita f.sp. tritici) of wheat. Int. J. Trop. Agric. 9, 118–121.
Dhaliwal, H. S., Harjit, S., and William, M. (2002). Transfer of rust resistance from Aegilops ovata into bread wheat (Triticum aestivum L.) and molecular characterisation of resistant derivatives. Euphytica 126, 153–159. doi: 10.1023/A:1016312723040
Dubkovsky, J. and Dvořák, J. (1995). Genome identification of the Triticum crassum complex (Poaceae) with the restriction patterns of repeated nucleotide sequences. Am. J. Botany. 182, 131–140. doi: 10.1002/j.1537-2197.1995.tb15657.x
Dulai, S., Molnár, I., Szopkó, D., Darkó, É., Vojtkó, A., Sass-Gyarmati, A., et al. (2014). Wheat-Aegilops biuncialis amphiploids have efficient photosynthesis and biomass production during osmotic stress. J. Plant Physiol. 171, 509–517. Available online at: https://www.sciencedirect.com/science/article/pii/S0176161713004641 (Accessed June 13, 2025).
Dvořák, J. (1987). Chromosomal distribution of genes in diploid Elytrigia elongata that promote or suppress pairing of wheat homoeologous chromosomes. Genome 29, 34–40. doi: 10.1139/g87-006
Dvořák, J. (Ed.) (1998). “Genome analysis in the Triticum-Aegilops alliance,” in Proceedings of the 9th International Wheat Genetics Symposium (Printcrafters Inc, Saskatoon, Saskatchewan). 2–7 August 1998.
Dvořák, J., Chen, K.-C., and Giorgi, B. (1984). The C-band pattern of a Ph– mutant of durum wheat. Can. J. Genet. Cytology. 26, 360–363. doi: 10.1139/g84-056
Dvořák, J., Deal, K. R., and Luo, M. C. (2006). Discovery and mapping of wheat Ph1 suppressors. Genetics 174, 17–27. Available online at: http://www.genetics.org/cgi/content/abstract/174/1/17 (Accessed May 25, 2013).
Dvořák, J., Luo, M. C., Yang, Z. L., and Zhang, H. B. (1998). The structure of the Aegilops tauschii genepool and the evolution of hexaploid wheat. Theor. Appl. Genet. 97, 657–670. doi: 10.1007/s001220050942
Dvořák, J., Mcguire, P. E., and Cassidy, B. (1988). Apparent sources of the А genomes of wheats inferred from polymorphism in abundance and restriction fragment length of repeated nucleotide sequences. Genome 30, 680–689. doi: 10.1139/g88-115
Edet, O. U., Gorafi, Y. S. A., Nasuda, S., and Tsujimoto, H. (2018). DArTseq-based analysis of genomic relationships among species of tribe Triticeae. Sci. Rep. 8, 16397. doi: 10.1038/s41598-018-34811-y
Eilam, T., Anikster, Y., Millet, E., Manisterski, J., and Feldman, M. (2010). Genome size in diploids, allopolyploids, and autopolyploids of Mediterranean Triticeae. J. Bot. 2010, 1–12 pp. doi: 10.1155/2010/341380
El Baidouri, M., Murat, F., Veyssiere, M., Molinier, M., Flores, R., Burlot, L., et al. (2017). Reconciling the evolutionary origin of bread wheat (Triticum aestivum). New Phytologist. 213, 1477–1486. doi: 10.1111/nph.14113
El Bouhssini, M., Benlhabib, O., Nachit, M. M., Houari, A., Bentika, A., Nsarellah, N., et al. (1998). Identification in Aegilops species of resistant sources to Hessian fly (Diptera: Cecidomyiidae) in Morocco. Genet. Resour. Crop Evolution. 45, 343–345. doi: 10.1023/A:1008675029389
El Bouhssini, M., Street, K., Amri, A., Mackay, M., Ogbonnaya, F. C., Omran, A., et al. (2011). Sources of resistance in bread wheat to Russian wheat aphid (Diuraphis noxia) in Syria identified using the Focused Identification of Germplasm Strategy (FIGS). Plant Breeding. 130, 96–97. doi: 10.1111/j.1439-0523.2010.01814.x
Endo, T. R. (1979). “On the Aegilops chromosome having gametocidal action on common wheat,” in Proceedings of the 5th International Wheat Genetics Symposium. Ed. Ramanujam, S.(New Delhi, India: Indian Society of Genetics and Plant Breeding), 306–314.
Endo, T. R. (1982). Gametocidal chromosomes of three Aegilops species in common wheat. Can. J. Genet. Cytology. 24, 201–296. doi: 10.1266/jjg.65.135
Endo, T. (1996). Allocation of a gametocidal chromosome of Aegilops cylindrica to wheat homoeologous group 2. Genes Genet. Systems. 71, 243–24 6. doi: 10.1266/ggs.71.243
Endo, T. R. (2007). The gametocidal chromosome as a tool for chromosome manipulation in wheat. Chromosome Res. 15, 67–75. doi: 10.1007/s10577-006-1100-3
Endo, T. R. (2015). “Gametocidal genes,” in Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics. Eds. Molnár-Laáng, M., Ceoloni, C., and Doležel, J. (IG Switzerland: Springer International Publishing Cham), 121–131.
Endo, T. R. (2021). “Dissection of the rye genome by the gametocidal system,” in The Rye Genome. Eds. Rabanus-Wallace, M. T. and Stein, N. (Springer International Publishing, Cham), 77–84.
Endo, T. R. and Tsunewaki, K. (1975). Sterility of common wheat with Aegilops triuncialis cytoplasm. Heredity 66, 13–18. doi: 10.1093/oxfordjournals.jhered.a108562
Endo TR, B. S. G. (1996). The deletion stocks of common wheat. J. Heredity. 87, 295–307. doi: 10.1093/oxfordjournals.jhered.a023003
Fakhri, Z., Mirzaghaderi, G., Ahmadian, S., and Mason, A. S. (2016). Unreduced gamete formation in wheat × Aegilops spp. hybrids is genotype specific and prevented by shared homologous subgenomes. Plant Cell Rep. 35, 1143–1154. doi: 10.1007/s00299-016-1951-9
Fan, C., Melonek, J., and Lukaszewski, A. J. (2024). New observations of the effects of the cytoplasm of Aegilops kotschyi Boiss. in bread wheat Triticum aestivum L. Genes 15, 855. Available online at: https://www.mdpi.com/2073-4425/15/7/855 (Accessed March 30, 2025).
Farkas, A., Molnár, I., Dulai, S., Rapi, S., Oldal, V., Cseh, A., et al. (2014). Increased micronutrient content (Zn, Mn) in the 3Mb(4B) wheat – Aegilops biuncialis substitution and 3Mb.4BS translocation identified by GISH and FISH. Genome 57, 61–67. doi: 10.1139/gen-2013-0204
Farooq, S. (2002). “Aegilops ovata: a potential gene source for improvement of salt tolerance of wheat,” in Prospects for saline agriculture, vol. 37 . Eds. Ahmad, R. and Malik, K. ,. H. (Kluwer Academic Publishers, Dordrecht), 123–130.
Farooq, S., Iqbal, N., and Shah, T. M. (1990). Promotion of homoeologous chromosome pairing in hybrids of Triticum aestivum x Aegilops variabilis. Genome 33, 825–828. doi: 10.1139/g90-124
Farooq, S., Niazi, M. L. K., Iqbal, N., and Shah, T. M. (1989). Salt tolerance potential of wild resources of the tribe Triticeae. Plant Soil. 119, 255–260. doi: 10.1007/BF02370417
Feldman, M. and Levy, A. A. (2023). Wheat evolution and domestication (Cham, Switzerland: Springer Cham).
Feldman, M. and Sears, E. R. (1981). The wild gene resources of wheat. Sci. Am. 244, 102–112. doi: 10.1038/scientificamerican0181-102
Friebe, B. and Heun, M. (1989). C-banding pattern and powdery mildew resistance of Triticum ovatum and four T. aestivum-T. ovatum chromosome addition lines. Theor. Appl. Genet. 78, 417–424. doi: 10.1007/BF00265306
Friebe, B., Jiang, J., Raupp, W. J., McIntosh, R. A., and Gill, B. S. (1996a). Characterization of wheat-alien translocations conferring resistance to diseases and pests: current status. Euphytica 91, 59–87. doi: 10.1007/BF00035277
Friebe, B., Jiang, J., Tuleen, N., and Gill, B. S. (1995). Standard karyotype of Triticum umbellulatum and the characterization of derived chromosome addition and translocation lines in common wheat. Theor. Appl. Genet. 90, 150–156. doi: 10.1007/BF00221010
Friebe, B., Tuleen, N. A., Badaeva, E. D., and Gill, B. S. (1996b). Cytogenetic identification of Triticum peregrinum chromosomes added to common wheat. Genome 39, 272–276. doi: 10.1139/g96-037
Gandhi, H. T., Vales, M. I., Mallory-Smith, C., and Riera-Lizarazu, O. (2009). Genetic structure of Aegilops cylindrica Host in its native range and in the United States of America. Theor. Appl. Genet. 119, 1013–1025. doi: 10.1007/s00122-009-1105-3
Garg, M., Tanaka, H., and Tsujimoto, H. (2009). Exploration of Triticeae seed storage proteins for improvement of wheat end-product quality. Breed. Science. 59, 519–528. doi: 10.1270/jsbbs.59.519
Garg, M., Tsujimoto, H., Gupta, R. K., Kumar, A., Kaur, N., Kumar, R., et al. (2016). Chromosome specific substitution lines of Aegilops geniculata alter parameters of bread making quality of wheat. PloS One 11, e0162350. doi: 10.1371/journal.pone.0162350
Gill, K. S. and Gill, B. S. (1991). A DNA fragment mapped within the submicroscopic deletion of Ph1, a chromosome pairing regulator gene in polyploid wheat. Genetics 129, 257–259. Available online at: http://www.genetics.org/content/129/1/257.abstract (Accessed April 16, 2015).
Gill, B. S., Sharma, C., Raupp, W. J., Browder, L. E., Heachett, J. H., Harvey, T. L., et al. (1985). Evaluation of Aegilops species for resistance to wheat powdery mildew, wheat leaf rust, Hessian fly, and greenbug. Plant Dis. 69, 314–316.
Gorham, J. (1990). Salt tolerance in the Triticeae: K/Na discrimination in Aegilops species. J. Exp. Botany. 41, 615–621. doi: 10.1093/jxb/41.5.615
Gradzielewska, A., Gruszecka, D., Leśniowska-Nowak, J., and Paczos-Grzęda, E. (2012). Identification of hybrids between triticale and Aegilops juvenalis (Thell.) Eig and determination of genetic similarity with ISSRs. Genet. Mol. research: GMR 11, 2147–2155. doi: 10.4238/2012.July.10.1
Green, A., Friebe, B., Prasad, P. V. V., and Fritz, A. (2018). Evaluating heat tolerance of a complete set of wheat-Aegilops geniculata chromosome addition lines. J. Agron. Crop Science. 204, 588–593. doi: 10.1111/jac.12282
Gultyaeva, E., Shaydayuk, E., Shreyder, E., Kushnirenko, I., and Shamanin, V. (2024). Genetic diversity of promising spring wheat accessions from Russia and Kazakhstan for rust resistance. Plants 13, 2469. Available online at: https://www.mdpi.com/2223-7747/13/17/2469 (Accessed August 10, 2025).
Guo, L., Yu, L., Tong, J., Zhao, Y., Yang, Y., Ma, Y., et al. (2021). Addition of Aegilops geniculata 1Ug chromosome improves the dough rheological properties by changing the composition and micro-structure of gluten. Food Chem. 358, 129850. Available online at: https://www.sciencedirect.com/science/article/pii/S0308814621008566 (Accessed July 01, 2025).
Gupta, P. K., Balyan, H. S., Gahlaut, V., Saripalli, G., Pal, B., Basnet, B. R., et al. (2019). Hybrid wheat: past, present and future. Theor. Appl. Genet. 132, 2463–2483. doi: 10.1007/s00122-019-03397-y
Gyawali, Y., Zhang, W., Chao, S., Xu, S., and Cai, X. (2019). Delimitation of wheat ph1b deletion and development of ph1b-specific DNA markers. Theor. Appl. Genet. 132, 195–204. doi: 10.1007/s00122-018-3207-2
Hammer, K. (1980). Vorarbeiten zur monographischen Darstellung von Wildpflanzensortimenten: Aegilops L. Kulturpflanze 28, 33–180. doi: 10.1007/BF02014641
Harjit-Singh, Tsujimoto, H., Sakhuja, P. K., Sigh, T., and Dhaliwal, H. S. (2000). Transfer of resistance to wheat pathogens from Aegilops triuncialis into bread wheat. Wheat Inf. Service 90, 21–30.
Harlan, J. R. and de Wet, J. M. J. (1971). Toward a rational classification of cultivated plants. Taxon 20, 509–517. Available online at: http://www.jstor.org/stable/1218252 (Accessed September 07, 2021).
Heyns, I., Pretorius, Z., and Francois, M. (2011). Derivation and characterization of recombinants of the Lr54/Yr37 translocation in common wheat. Open Plant Sci. J. 5, 1–8 pp. doi: 10.2174/1874294701105010001
Holubec, V., Hanušova, R., and Kostkanovǎ, E. (1992). The Aegilops collection in the Praha-Ruzyné (CzechoSlovakia) Gene Bank: collecting, evaluation and documentation. Hereditas 116, 271–276. doi: 10.1111/j.1601-5223.1992.tb00835.x
Holubec, V. and Havliikovi, H. (1994). Interspecific differences in cereal aphid infestation of 20 Aegilops species. Genet. Slecht. 30, 81–87.
Huerta-Espino, J., Singh, R. P., and Reyna-Martinez, J. (2008). First detection of virulence to genes Lr9 and Lr25 conferring resistance to leaf rust of wheat caused by Puccinia triticina in Mexico. Plant Disease. 92, 311. doi: 10.1094/PDIS-92-2-0311A
Imadi, S. R. and Gul, A. (2019). “Wild relatives and their role in building resistance in bread wheat: at a glance,” in Crop Production Technologies for Sustainable Use and Conservation/Physiological and Molecular Advances. Eds. Ozturk, M., Hakeem, K. R., Ashraf, M., and Ahmad, M. S. A. (Apple Academic press Inc, Oakville, Canada), 395–416.
Iqbal, N., Reader, S. M., Caligari, P. D. S., and Miller, T. E. (2000). Characterization of Aegilops uniaristata chromosomes by comparative DNA marker analysis and repetitive DNA sequence in situ hybridization. Theor. Appl. Genet. 101, 1173–1179. doi: 10.1007/s001220051594
Ivanizs, L., Monostori, I., Farkas, A., Megyeri, M., Mikó, P., Türkösi, E., et al. (2019). Unlocking the genetic diversity and population structure of a wild gene source of wheat, Aegilops biuncialis Vis., and its relationship with the heading time. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.01531
Jahier, J., Rivoal, R., Yu, M. Q., Abelard, P., Tanguy, A. M., and Barloy, D. (1998). Transfer of genes for resistance to cereal cyst nematode from Aegilops variabilis Eig to wheat. J. Genet. Breeding. 52, 253–257. Available online at: https://hal.inrae.fr/hal-02693092 (Accessed June 30, 2025).
Jiang, J., Friebe, B., and Gill, B. S. (1993). Recent advances in alien gene transfer in wheat. Euphytica 73, 199–212. doi: 10.1007/BF00036700
Kamboj, R., Sharma, S., Kumar, R., Sharma, P., Ravat, V. K., Chhuneja, P., et al. (2020). Introgression of powdery mildew resistance from Aegilops triuncialis into wheat through induced homeologous pairing. J. Plant Biochem. Biotechnol. 29, 418–426. doi: 10.1007/s13562-020-00548-x
Khan, M. K., Pandey, A., Hamurcu, M., Avsaroglu, Z. Z., Ozbek, M., Omay, A. H., et al. (2021). Variability in physiological traits reveals boron toxicity tolerance in Aegilops species. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.736614
Kihara, H. (1954). Considerations on the evolution and distribution of Aegilops species based on the analyser-method. Cytologia 19, 336–357. doi: 10.1508/cytologia.19.336
Kihara, H., Yamashita, K., and Tanaka, M. (1959). Genomes of 6x species of Aegilops. Wheat Inf. Service 8, 3–5.
Kilian, B., Mammen, K., Millet, E., Sharma, R., Graner, A., Salamini, F., et al. (2011). “Aegilops,” in Wild crop relatives: Genomics and breeding resources Cereals. Ed. Kole, C. (Springer, Berlin Heidelberg), 1–76.
Kimber, G. and Feldman, M. (1987). Wild wheat, an Introduction (Columbia, Mo, U.S.A: College of Agriculture University of Missouri), 142 p.
Kimber, G. and Tsunewaki, K. (Eds.) (1988). “Genome symbols and plasma types in the wheat group,” in Proceedings of the 7th International Wheat Genetics Symposium (Bath Press, Avon, Cambridge, England). 13–19 July 1988.
King, J., Dreisigacker, S., Reynolds, M., Bandyopadhyay, A., Braun, H. J., Crespo-Herrera, L., et al. (2024). Wheat genetic resources have avoided disease pandemics, improved food security, and reduced environmental footprints: A review of historical impacts and future opportunities. Global Change Biol. 30, e17440. doi: 10.1111/gcb.17440
Kishii, M. (2019). An update of recent use of Aegilops species in wheat breeding. Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00585
Knott, D. R. and Dvořák, J. (1976). Alien germ plasm as a source of resistance to disease. Annu. Rev. Phytopatology. 14, 211–235. doi: 10.1146/annurev.py.14.090176.001235
Koebner, R. M. D. and Shepherd, K. W. (1986). Controlled introgression to wheat of genes from rye chromosome arm 1RS by induction of allosyndesis. Theor. Appl. Genet. 73, 197–208. doi: 10.1007/BF00289275
Koo, D.-H., Friebe, B., and Gill, B. S. (2020). Homoeologous recombination: A novel and efficient system for broadening the genetic variability in wheat. Agronomy 10, 1059. Available online at: https://www.mdpi.com/2073-4395/10/8/1059 (Accessed March 18, 2025).
Koo, D.-H., Liu, W., Friebe, B., and Gill, B. S. (2017). Homoeologous recombination in the presence of Ph1 gene in wheat. Chromosoma 126, 531–540. doi: 10.1007/s00412-016-0622-5
Kozub, N. A., Sozinov, I. A., and Sozinov, A. A. (2012). Identification of alleles at the gliadin loci Gli-U1 and Gli-Mb1 in Aegilops biuncialis Vis. Russian J. Genet. 48, 390–395. doi: 10.1134/S1022795412030052
Kumar, A., Choudhary, A., Kaur, H., and Mehta, S. (2022). A walk towards Wild grasses to unlock the clandestine of gene pools for wheat improvement: A review. Plant Stress. 3, 100048. Available online at: https://www.sciencedirect.com/science/article/pii/S2667064X21000476 (Accessed March 17, 2025).
Kumar, K., Jan, I., Saripalli, G., Sharma, P. K., Mir, R. R., Balyan, H. S., et al. (2022). An update on resistance genes and their use in the development of leaf rust resistant cultivars in wheat. Front. Genet. 13. doi: 10.3389/fgene.2022.816057
Kumar, A., Kapoor, P., Chunduri, V., Sharma, S., and Garg, M. (2019). Potential of Aegilops sp. for improvement of grain processing and nutritional quality in wheat (Triticum aestivum). Front. Plant Sci. 10. doi: 10.3389/fpls.2019.00308
Kuraparthy, V., Chhuneja, P., Dhaliwal, H. S., Kaur, S., Bowden, R. L., and Gill, B. S. (2007a). Characterization and mapping of cryptic alien introgression from Aegilops geniculata with new leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat. Theor. Appl. Genet. 114, 1379–1389. doi: 10.1007/s00122-007-0524-2
Kuraparthy, V., Sood, S., Chhuneja, P., Dhaliwal, H. S., Kaur, S., Bowden, R. L., et al. (2007b). A cryptic wheat-Aegilops triuncialis translocation with leaf rust resistance gene Lr58. Crop Sci. 47, 1995–2003. doi: 10.2135/cropsci2007.01.0038
Kuraparthy, V., Sood, S., and Gill, B. S. (2009). Molecular genetic description of the cryptic wheat–Aegilops geniculata introgression carrying rust resistance genes Lr57 and Yr40 using wheat ESTs and synteny with rice. Genome 52, 1025–1036. doi: 10.1139/g09-076
Kwiatek, M. T., Wiśniewska, H., Ślusarkiewicz-Jarzina, A., Majka, J., Majka, M., Belter, J., et al. (2017). Gametocidal factor transferred from Aegilops geniculata Roth can be adapted for large-scale chromosome manipulations in cereals. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.00409
Kynast, R. G., Friebe, B., and Gill, B. S. (2000). Fate of multicentric and ring chromosomes induced by a new gametocidal factor located on chromosome 4Mg of Aegilops geniculata. Chromosome Res. 8, 133–139. doi: 10.1023/A:1009294519798
Laibach, F. (1925). Das taubwerden von bastardsamen und die künstliche Aufzucht früh absterbender bastardembryonen. Z Bot. 17, 417–459.
Landjeva, S., Merakchijska, M., and Ganeva, G. (1998). Seedling copper tolerance and cytogenetic characterization of wheat-Aegilops ovata hybrid lines. Wheat Inf. Service. 87, 27–30.
Laugerotte, J., Baumann, U., and Sourdille, P. (2022). Genetic control of compatibility in crosses between wheat and its wild or cultivated relatives. Plant Biotechnol. J. 20, 812–832. doi: 10.1111/pbi.13784
Lawrence, G. J. and Shepherd, K. W. (1981). Chromosomal location of genes controlling seed proteins in species related to wheat. Theor. Appl. Genet. 59, 25–31. doi: 10.1007/BF00275771
Lelley, T., Stachel, M., Grausgruber, H., and Vollmann, J. (2000). Analysis of relationships between Aegilops tauschii and the D genome of wheat utilizing microsatellites. Genome 43, 661–668. doi: 10.1139/gen-43-4-661
Levy, A. A. and Feldman, M. (2022). Evolution and origin of bread wheat. Plant Cell. 34, 2549–2567. doi: 10.1093/plcell/koac130
Lewis, C. M., Persoons, A., Bebber, D. P., Kigathi, R. N., Maintz, J., Findlay, K., et al. (2018). Potential for re-emergence of wheat stem rust in the United Kingdom. Commun. Biol. 1, 13. doi: 10.1038/s42003-018-0013-y
Li, H., Deal, K. R., Luo, M.-C., Ji, W., Distelfeld, A., and Dvorak, J. (2017). Introgression of the Aegilops speltoides Su1-Ph1 suppressor into wheat. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.02163
Li, S., Liu, Z., Jia, Y., Ye, J., Yang, X., Zhang, L., et al. (2019). Analysis of metabolic pathways related to fertility restoration and identification of fertility candidate genes associated with Aegilops kotschyi cytoplasm in wheat (Triticum aestivum L.). BMC Plant Biol. 19, 252. doi: 10.1186/s12870-019-1824-9
Li, J., Nasuda, S., and Endo, T. (2013). Dissection of rye chromosomes by the gametocidal system. Genes Genet. Systems. 88, 321–327. doi: 10.1266/ggs.88.321
Li, H., Rehman, S., Song, R., Qiao, L., Hao, X., Zhang, J., et al. (2024). Chromosome-scale assembly and annotation of the wild wheat relative Aegilops comosa. Sci. Data 11, 1454. doi: 10.1038/s41597-024-04346-1
Li, L.-F., Zhang, Z.-B., Wang, Z.-H., Li, N., Sha, Y., Wang, X.-F., et al. (2022). Genome sequences of five Sitopsis species of Aegilops and the origin of polyploid wheat B subgenome. Mol. Plant 15, 488–503. doi: 10.1016/j.molp.2021.12.019
Li, G., Zhang, T., Yu, Z., Wang, H., Yang, E., and Yang, Z. (2021). An efficient Oligo-FISH painting system for revealing chromosome rearrangements and polyploidization in Triticeae. Plant J. 105, 978–993. doi: 10.1111/tpj.15081
Liang, W., Ma, X., Wan, P., and Liu, L. (2018). Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 495, 286–291. Available online at: https://www.sciencedirect.com/science/article/pii/S0006291X17322209 (Accessed March 27, 2025).
Lilienfeld, F. A. H. (1951). Kihara: genome-analysis in Triticum and Aegilops. X. Concluding review. Cytologia 16, 101–123. doi: 10.1508/cytologia.16.101
Liu, W., Rouse, M., Friebe, B., Jin, Y., Gill, B., and Pumphrey, M. (2011). Discovery and molecular mapping of a new gene conferring resistance to stem rust, Sr53, derived from Aegilops geniculata and characterization of spontaneous translocation stocks with reduced alien chromatin. Chromosome Res. 19, 669–682. doi: 10.1007/s10577-011-9226-3
Liu, D., Xiang, Z., Zhang, L., Zheng, Y., Yang, W., Chen, G., et al. (2010). Transfer of stripe rust resistance from Aegilops variabilis to bread wheat. Afr. J. Biotechnol. 10, 136–139. doi: 10.5897/AJB09.1719
Liu, C., Yang, Z., and Hu, Y.-G. (2015). Drought resistance of wheat alien chromosome addition lines evaluated by membership function value based on multiple traits and drought resistance index of grain yield. Field Crops Res. 179, 103–112. Available online at: https://www.sciencedirect.com/science/article/pii/S0378429015001409 (Accessed May 17, 2016).
Logojan, A. A. and Molnár-Láng, M. (2000). Production of Triticum aestivum — Aegilops biuncialis chromosome additions. Cereal Res. Commun. 28, 221–228. doi: 10.1007/BF03543597
Luo, M.-C., Gu, Y. Q., Puiu, D., Wang, H., Twardziok, S. O., Deal, K. R., et al. (2017). Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 551, 498. doi: 10.1038/nature24486
Ma, C., Qian, J., Feng, Y., Sehgal, S. K., Zhao, Y., Chen, Q., et al. (2023). Genetic mapping of a novel gene PmAege7M from Aegilops geniculata conferring resistance to wheat powdery mildew. Plant Disease. 107, 3608–3615. doi: 10.1094/PDIS-04-23-0764-RE
Maan, S. S. (1977). Cytoplasmic homology between Aegilops mutica Boiss. and Ae. ovata L. Euphytica 26, 601–613. doi: 10.1007/BF00021685
Makkouk, K. M., Ghulam, W., and Comeau, A. (1994). Resistance to barley yellow dwarf luteovirus in Aegilops species. Can. J. Plant Science. 74, 631–634. doi: 10.4141/cjps94-113
Manisterski, J., Segal, A., Levy, A., and Feldman, M. (1988). Evaluation of Israeli Aegilops and Agropyron species for resistance to wheat leaf rust. Plant Disease. 72, 941–944. doi: 10.1094/PD-72-0941
Marais, G. F., Kotze, L., and Eksteen, A. (2010a). Allosyndetic recombinants of the Aegilops peregrina-derived Lr59 translocation in common wheat. Plant Breeding. 129, 356–361. doi: 10.1111/j.1439-0523.2009.01713.x
Marais, G. F., Marais, A. S., Eksteen, A., and Pretorius, Z. A. (2010b). Modification of the Aegilops neglecta - common wheat Lr62/Yr42 translocation through allosyndetic pairing induction. South Afr. J. Plant Soil. 27, 142–151. doi: 10.10520/EJC87975
Marais, F., Marais, A., McCallum, B., and Pretorius, Z. (2009). Transfer of leaf rust and stripe rust resistance genes Lr62 and Yr42 from Aegilops neglecta Req. ex Bertol. to common wheat. Crop Science. 49, 871–879. doi: 10.2135/cropsci2008.06.0317
Marais, G. F., McCallum, B., and Marais, A. S. (2008). Wheat leaf rust resistance gene Lr59 derived from Aegilops peregrina. Plant Breed. 127, 340–345. doi: 10.1111/j.1439-0523.2008.01513.x
Marais, G. F., McCallum, B., Snyman, J. E., Pretorius, Z. A., and Marais, A. S. (2005). Leaf rust and stripe rust resistance genes Lr54 and Yr37 transferred to wheat from Aegilops kotschyi. Plant Breed. 124, 538–541. doi: 10.1111/j.1439-0523.2005.01116.x
Marais, G. F., Pretorius, Z. A., Marias, A. S., and Wellings, C. R. (2003). Transfer of rust resistance genes from Triticum species to common wheat. South Afr. J. Plant Soil. 20, 193–198. doi: 10.1080/02571862.2003.10634934
Marcussen, T., Sandve, S. R., Heier, L., Spannagl, M., Pfeifer, M., Jakobsen, K. S., et al. (2014). Ancient hybridizations among the ancestral genomes of bread wheat. Science 345. Available online at: http://science.sciencemag.org/content/345/6194/1250092.abstract (Accessed March 19, 2019).
Martin-Sanchez, J., Gomez-Colmenarejo, M., Del Moral, J., Sin, E., Montes, M., Gonzalez-Belinchon, C., et al. (2003). A new Hessian fly resistance gene (H30) transferred from the wild grass Aegilops triuncialis to hexaploid wheat. Theor. Appl. Genet. 106, 1248–12 55. doi: 10.1007/s00122-002-1182-z
Martynov, S. P., Dobrotvorskaya, T. V., and Mitrofanova, O. P. (2015). Genealogical analysis of the use of Aegilops (Aegilops L.) genetic material in wheat (Triticum aestivum L.). Russian J. Genet. 51, 855–862. doi: 10.1134/S1022795415090070
Masoudi-Nejad, A., Nasuda, S., McIntosh, R. A., Endo, T. R., and Endo, T. R. (2002). Transfer of rye chromosome segments to wheat by a gametocidal system. Chromosome Res. 10, 349–357. doi: 10.1023/A:1016845200960
Matsuoka, Y., Takumi, S., and Nasuda, S. (2014). “Chapter Four - Genetic mechanisms of allopolyploid speciation through hybrid genome doubling: Novel insights from wheat (Triticum and Aegilops) studies,” in International Review of Cell and Molecular Biology, vol. 309 . Ed. Jeon, K. W. (U.S.A.: Academic Press), 199–258.
McFadden, E. S. and Sears, E. R. (1946). The origin of Triticum spelta and its free-threshing hexaploid relatives. J. Heredity 37, 81–107. doi: 10.1093/oxfordjournals.jhered.a105590
Mello-Sampayo, T. (1971). Genetic regulation of meiotic chromosome pairing by chromosome 3D of Triticum aestivum. Nat. New Biol. 230, 22. doi: 10.1038/newbio230022a0
Men, W., Fan, Z., Ma, C., Zhao, Y., Wang, C., Tian, X., et al. (2022). Mapping of the novel powdery mildew resistance gene Pm2Mb from Aegilops biuncialis based on ph1b-induced homoeologous recombination. Theor. Appl. Genet. 135, 2993–3003. doi: 10.1007/s00122-022-04162-4
Merker, A. (1992). The Triticeae in cereal breeding. Hereditas 116, 277–280. doi: 10.1111/j.1601-5223.1992.tb00154.x
Mizuno, N., Yamasaki, M., Matsuoka, Y., Kawahara, T., and Takumi, S. (2010). Population structure of wild wheat D-genome progenitor Aegilops tauschii Coss.: implications for intraspecific lineage diversification and evolution of common wheat. Mol. Ecol. 19, 999–1013. doi: 10.1111/j.1365-294X.2010.04537.x
Molnár, I., Benavente, E., and Molnár-Láng, M. (2009). Detection of intergenomic chromosome rearrangements in irradiated Triticum aestivum–Aegilops biuncialis amphiploids by multicolour genomic in situ hybridization. Genome 52, 156–165. doi: 10.1139/g08-114
Molnár, I., Gáspár, L., Sárvári, É, Dulai, S., Hoffmann, B., Molnár-Láng, M., et al. (2004). Physiological and morphological responses to water stress in Aegilops biuncialis and Triticum aestivum genotypes with differing tolerance to drought. Funct. Plant Biol. 31, 1149–1159. Available online at: https://www.publish.csiro.au/paper/FP03143 (Accessed March 25, 2025).
Molnár, I., Kubaláková, M., Šimková, H., Cseh, A., Molnár-Láng, M., and Doležel, J. (2011). Chromosome isolation by flow sorting in Aegilops umbellulata and Ae. comosa and their allotetraploid hybrids Ae. biuncialis and Ae. geniculata. PLoS One 6, e27708. doi: 10.1371/journal.pone.0027708
Monneveux, P., Zaharieva, M., and Rekika, D. (2000). “The utilisation of Triticum and Aegilops species for the improvement of durum wheat,” in Durum wheat improvement in the Mediterranean region: New challenges, vol. 40 . Eds. Royo, C., Nachit, M., Di Fonzo, N., and Araus, J. L. (CIHEAM, Zaragoza), 71–781.
Mujeeb-Kazi, A., Gul, A., Ahmad, I., Farooq, M., Rauf, Y., -ur Rahman, A., et al. (2009). “Genetic resources for some wheat abiotic stress tolerances,” in Salinity and Water Stress: Improving Crop Efficiency. Eds. Ashraf, M., Ozturk, M., and Athar, H. R. (Springer Netherlands, Dordrecht), 149–163.
Mukai, Y. and Tsunewaki, K. (1979). Basic studies on hybrid wheat breeding. Theor. Appl. Genet. 54, 153–160. doi: 10.1007/BF00263045
Narang, D., Kaur, S., Steuernagel, B., Ghosh, S., Bansal, U., Li, J., et al. (2020). Discovery and characterisation of a new leaf rust resistance gene introgressed in wheat from wild wheat Aegilops peregrina. Sci. Rep. 10, 7573. doi: 10.1038/s41598-020-64166-2
Narang, D., Kaur, S., Steuernagel, B., Ghosh, S., Dhillon, R., Bansal, M., et al. (2019). Fine mapping of Aegilops peregrina co-segregating leaf and stripe rust resistance genes to distal-most end of 5DS. Theor. Appl. Genet. 132, 1473–1485. doi: 10.1007/s00122-019-03293-5
Narang, D., Satinder, K., Jyoti, S., and Chhuneja, P. (2018). Development and molecular characterization of wheat-Aegilops peregrina introgression lines with resistance to leaf rust and stripe rust. J. Crop Improvement. 32, 59–70. doi: 10.1080/15427528.2017.1398117
Naranjo, T. and Benavente, E. (2015). “The mode and regulation of chromosome pairing in wheat–alien hybrids (Ph genes, an updated view),” in Alien Introgression in Wheat: Cytogenetics, Molecular Biology, and Genomics. Eds. Molnár-Láng, M., Ceoloni, C., and Doležel, J. (Springer International Publishing, Cham), 133–162.
Nasuda, S., Kikkawa, Y., Ashida, T., Islam, A. K. M. R., Sato, K., and Endo, T. R. (2005). Chromosomal assignment and deletion mapping of barley EST markers. Genes Genet. Systems. 80, 357–366. doi: 10.1266/ggs.80.357
Neelam, K., Rawat, N., Tiwari, V., Prasad, R., Tripathi, S. K., Randhawa, G. S., et al. (2012). Evaluation and identification of wheat-Aegilops addition lines controlling high grain Iron and Zinc concentration and mugineic acid production. Cereal Res. Commun. 40, 53–61. doi: 10.1556/CRC.40.2012.1.7
Niranjana, M. (2017). Gametocidal genes of Aegilops: segregation distorters in wheat–Aegilops wide hybridization. Genome 60, 639–647. doi: 10.1139/gen-2017-0023
Ohta, S. and Saruhashi, Y. (1999). Geographical distribution of B chromosomes in Aegilops mutica Boiss., a wild relative of wheat. Hereditas 130, 177–183. doi: 10.1111/j.1601-5223.1999.00177.x
Olivera, P. D., Rouse, M. N., and Jin, Y. (2018). Identification of new sources of resistance to wheat stem rust in Aegilops spp. in the tertiary genepool of wheat. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01719
Ortelli, S., Winzeler, H., Winzeler, M., Fried, P. M., and Nösberger, J. (1996a). Leaf rust resistance gene Lr9 and winter wheat yield reduction: I. Yield and yield components. Crop Sci. 36, 1590–1595. doi: 10.2135/cropsci1996.0011183X003600060030x
Ortelli, S., Winzeler, M., Winzeler, H., and Nösberger, J. (1996b). Leaf rust resistance gene Lr9 and winter wheat yield reduction: II. Leaf gas exchange and root activity. Crop Sci. 36, 1595–1601. doi: 10.2135/cropsci1996.0011183X003600060031x
Özgen, M., Yildiz, M., Ulukan, H., and Koyuncu, N. (2004). Association of gliadin protein pattern and rust resistance derived from Aegilops umbellulata Zhuk. in winter Triticum durum Desf. Breed. Science. 54, 287–290. doi: 10.1270/jsbbs.54.287
Parisoid, C. and Badaeva, E. D. (2020). Chromosome restructuring among hybridizing wild wheats. New Phytologist. 226, 1263–1273. doi: 10.1111/NPH.16415
Patpour, M., Hovmøller, M. S., Rodriguez-Algaba, J., Randazzo, B., Villegas, D., Shamanin, V. P., et al. (2022). Wheat stem rust back in Europe: Diversity, prevalence and impact on host resistance. Front. Plant Sci. 13. doi: 10.3389/fpls.2022.882440
Perničková, K., Koláčková, V., Lukaszewski, A. J., Fan, C., Vrána, J., Duchoslav, M., et al. (2019). Instability of alien chromosome introgressions in wheat associated with improper positioning in the nucleus. Int. J. Mol. Science. 20, 1448. doi: 10.3390/ijms20061448
Person-Dedryver, F., Jahier, J., and Danielle, P. (1985). Les céréales à paille hôtes de Meloidogyne naasi Franklin. III. - Recherches de sources de résistance parmi les espèces voisines du blé tendre. Agronomie 5, 573–57 8. doi: 10.1051/agro:19850702
Petersen, G., Seberg, O., Yde, M., and Berthelsen, K. (2006). Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum). Mol. Phylogenet. Evolution. 39, 70–82. doi: 10.1016/j.ympev.2006.01.023
Pirseyedi, S.-M., Somo, M., Poudel, R. S., Cai, X., McCallum, B., Saville, B., et al. (2015). Characterization of recombinants of the Aegilops peregrina-derived Lr59 translocation of common wheat. Theor. Appl. Genet. 128, 2403–2414. doi: 10.1007/s00122-015-2594-x
Placido, D. F., Campbell, M. T., Folsom, J. J., Cui, X., Kruger, G. R., Baenziger, P. S., et al. (2013). Introgression of novel traits from a wild wheat relative improves drought adaptation in wheat. Plant Physiol. 161, 1806–1819. doi: 10.1104/pp.113.214262
Pradhan, G. P., Prasad, P. V. V., Fritz, A. K., Kirkham, M. B., and Gill, B. S. (2012a). Response of Aegilops species to drought stress during reproductive stages of development. Funct. Plant Biol. 39, 51–59. Available online at: https://www.publish.csiro.au/paper/FP11171 (Accessed March 31, 2025).
Pradhan, G. P., Prasad, P. V. V., Fritz, A. K., Kirkham, M. B., and Gill, B. S. (2012b). High temperature tolerance in Aegilops species and Its potential transfer to wheat. Crop Science. 52, 292–304. doi: 10.2135/cropsci2011.04.0186
Prasad, P., Savadi, S., Bhardwaj, S. C., and Gupta, P. K. (2020). The progress of leaf rust research in wheat. Fungal Biol. 124, 537–550. Available online at: https://www.sciencedirect.com/science/article/pii/S1878614620300337 (Accessed March 17, 2025).
Pyralov, G. R. (1976). “Results of hybridization of wheat with Aegilops, rye, Haynaldia and Agropyron,” in Genetics and Breeding in Azerbaijan. IV. Baku, Azerbaijan: Elm 136–137.
Qi, L., Friebe, B., Zhang, P., and Gill, B. S. (2007). Homoeologous recombination, chromosome engineering and crop improvement. Chrom Res. 15, 3–19. doi: 10.1007/s10577-006-1108-8
Ragini, R., Murukan, N., Sekhon, N. K., Chugh, C., Agarwal, P., Yadav, P., et al. (2024). Breaking the association between gametocidal gene(s) and leaf rust resistance gene (LrS2427) in Triticum aestivum-Aegilops speltoides derivative by gamma irradiation. Mol. Breeding. 44, 54. doi: 10.1007/s11032-024-01491-8
Rakszegi, M., Darkó, É., Lovegrove, A., Molnár, I., Láng, L., Bedő, Z., et al. (2019). Drought stress affects the protein and dietary fiber content of wholemeal wheat flour in wheat/Aegilops addition lines. PLoS One 14, e0211892. doi: 10.1371/journal.pone.0211892
Rakszegi, M., Molnár, I., Lovegrove, A., Darkó, É., Farkas, A., Láng, L., et al. (2017). Addition of Aegilops U and M chromosomes affects protein and dietary fiber content of wholemeal wheat flour. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01529
Rather, S., Sharma, D., Indra Deo, P., and Joshi, N. (2017). “Alien gene introgression in wheat,” in Wheat: A premier Food Crop. Eds. Birendra, P., Kumar, A., and Kumar, A. (Kalyani Publishers, New Delhi, India), 101–120.
Rawat, N., Neelam, K., Tiwari, V. K., Randhawa, G. S., Friebe, B., Gill, B. S., et al. (2011). Development and molecular characterization of wheat-Aegilops kotschyi addition and substitution lines with high grain protein, iron, and zinc. Genome 54, 943–953. doi: 10.1139/g11-059
Rawat, N., Tiwari, V. K., Singh, N., Randhawa, G. S., Singh, K., Chhuneja, P., et al. (2009). Evaluation and utilization of Aegilops and wild Triticum species for enhancing iron and zinc content in wheat. Genet. Resour. Crop Evolution. 56, 53–64. doi: 10.1007/s10722-008-9344-8
Reader, S. M. and Miller, T. E. (1987). The simultaneous substitution of two pairs of chromosomes from two alien species into Triticum aestivum cv. Chinese Spring. Cereal Res. Commun. 15, 39–42. Available online at: http://www.jstor.org/stable/23782827 (Accessed August 10, 2025).
Riley, R., Chapman, V., and Kimber, G. (1959). Genetic control of chromosome pairing in intergeneric hybrids with wheat. Nature 183, 1244–1246. doi: 10.1038/1831244a0
Riley, R., Chapman, V., and Miller, T. (1973). “The determination of meiotic chromosome pairing,” in Proceedings of the 4th International Wheat Genetics Symposium. Eds. Sears, E. R. and Sears, L. M. S. (Mo. Agr. Exp. Stn, Columbia, Missouri), 731–738.
Riley, R. and Kempanna, C. (1963). The homoeologous nature of the non-homologous meiotic pairing in Triticum aestivum deficient for chromosome V (5B). Heredity 18, 287–306. doi: 10.1038/hdy.1963.31
Rivoal, R., Doussinault, G., and Jahier, J. (1986). Resistance au développment d’Heterodera avenae Woll. chez diffréntes espèces de Triticum. Agronomie 6, 759–765. doi: 10.1051/AGRO:19860809
Rogo, U., Fambrini, M., and Pugliesi, C. (2023). Embryo rescue in plant breeding. Plants 12, 3106. Available online at: https://www.mdpi.com/2223-7747/12/17/3106 (Accessed July 01, 2025).
Romero, M., Montes, M., Sin, E., Lopez-Brana, I., Duce, A., Martin-Sanchez, J., et al. (1998). A cereal cyst nematode (Heterodera avenae Woll.) resistance gene transferred from Aegilops triuncialis to hexaploid wheat. Theor. Appl. Genet. 96, 1135–11 40. doi: 10.1007/s001220050849
Ruban, A. S. and Badaeva, E. D. (2018). Evolution of the S-genomes in Triticum-Aegilops alliance: Evidences from chromosome analysis. Front. Plant Sci. 9. doi: 10.3389/fpls.2018.01756
Sadeghabad, A. A., Dadkhodaie, A., and Heidari, B. (2020). Phenotypic and genetic diversity of leaf rust resistance in wheat wild relatives. J. Phytopathol. 168, 428–438. doi: 10.1111/jph.12907
Said, M. and Cabrera, A. (2009). A physical map of chromosome 4Hch from H. Chilense containing SSR, STS and EST-SSR molecular markers. Euphytica 167, 253–259. doi: 10.1007/s10681-009-9895-6
Said, M., Gaál, E., Farkas, A., Molnár, I., Bartoš, J., Doležel, J., et al. (2024). Gametocidal genes: from a discovery to the application in wheat breeding. Front. Plant Sci. 15. doi: 10.3389/fpls.2024.1396553
Said, M., Holušová, K., Farkas, A., Ivanizs, L., Gaál, E., Cápal, P., et al. (2021). Development of DNA markers from physically mapped loci in Aegilops comosa and Aegilops umbellulata using single-gene FISH and chromosome sequences. Front. Plant Sci. 12. doi: 10.3389/fpls.2021.689031
Sakai, K., Nasuda, S., Sato, K., and Endo, T. R. (2009). Dissection of barley chromosome 3H in common wheat and a comparison of 3H physical and genetic maps. Genes Genet. Systems. 84, 25–34. doi: 10.1266/ggs.84.25
Salse, J., Chagué, V., Bolot, S., Magdelenat, G., Huneau, C., Pont, C., et al. (2008). New insights into the origin of the B genome of hexaploid wheat: Evolutionary relationships at the SPA genomic region with the S genome of the diploid relative Aegilops speltoides. BMC Genomics 9, 1–12. doi: 10.1186/1471-2164-9-555
Schneider, A., Molnár, I., and Molnár-Láng, M. (2008). Utilisation of Aegilops (goatgrass) species to widen the genetic diversity of cultivated wheat. Euphytica 163, 1–19. doi: 10.1007/s10681-007-9624-y
Schwessinger, B. (2017). Fundamental wheat stripe rust research in the 21st century. New Phytologist. 213, 1625–1631. doi: 10.1111/nph.14159
Sears, E. R. (1956). The transfer of leaf rust resistance from Aegilops umbellulata into wheat. Brookhaven Symp. Biol. 9, 1–21.
Sears, E. R. (1976). Genetic control of chromosome pairing in wheat. Ann. Rev. Genet. 30, 31–51. doi: 10.1146/annurev.ge.10.120176.000335
Sears, E. R. (1977). An induced mutant with homoeologous pairing in common wheat. Can. J. Genet. Cytology. 19, 585–593. doi: 10.1139/g77-063
Sears, E. R. and Okamato, M. (Eds.) (1958). “Intergenomic chromosome relationship in hexaploid wheat,” in Proceedings of the 10th International Congress of Genetics (University of Toronto Press, Montreal, Canada). 20–27 August 1958.
Serizawa, N., Nasuda, S., Shi, F., Endo, T. R., Prodanovic, S., Schubert, I., et al. (2001). Deletion-based physical mapping of barley chromosome 7H. Theor. Appl. Genet. 103, 827–834. doi: 10.1007/s001220100703
Sharma, D. R., Kaur, R., and Kumar, K. (1996). Embryo rescue in plants - a review. Euphytica 89, 325–337. doi: 10.1007/BF00022289
Sharma, S., Schulthess, A. W., Bassi, F. M., Badaeva, E. D., Neumann, K., Graner, A., et al. (2021). Introducing beneficial alleles from plant genetic resources into the wheat germplasm. Biology 10, 982. Available online at: https://www.mdpi.com/2079-7737/10/10/982 (Accessed November 03, 2024).
Sharma, Y., Sheikh, I., Sharma, A., Yadav, A. N., Kumar, K., Chhuneja, P., et al. (2020). Transfer of grain softness from 5U-5A wheat-Aegilops triuncialis substitution line to bread wheat through induced homeologous pairing. J. Plant Biochem. Biotechnol. 29, 407–417. doi: 10.1007/s13562-020-00554-z
Shepherd, K. (1973). “Homoeology of wheat and alien chromosomes controlling endosperm protein phenotypes,” in Proc 4th Int Wheat Genet Symp. Eds. Sears, E. and Sears, L. (University of Missouri, Columbia, Mo), 745–760.
Shimshi, D., Mayoral, M. L., and Atsmon, D. (1982). Responses to water stress in wheat and related wild species. Crop Sci. 22, 123–128. doi: 10.2135/cropsci1982.0011183X002200010028x
Singh, J., Gudi, S., Maughan, P. J., Liu, Z., Kolmer, J., Wang, M., et al. (2024). Genomes of Aegilops umbellulata provide new insights into unique structural variations and genetic diversity in the U-genome for wheat improvement. Plant Biotechnol. J. 22, 3505–3519. doi: 10.1111/pbi.14470
Singh, R., Hodson, D., Jin, Y., Lagudah, E., Ayliffe, M., Bhavani, S., et al. (2015). Emergence and spread of new races of wheat stem rust fungus: Continued threat to food security and prospects of genetic control. Phytopathology 105, PHYTO01150030FI. doi: 10.1094/PHYTO-01-15-0030-FI
Singh, N., Wu, S., Tiwari, V., Sehgal, S., Raupp, J., Wilson, D., et al. (2019). Genomic analysis confirms population structure and identifies inter-lineage hybrids in Aegilops tauschii. Front. Plant Science. 10. doi: 10.3389/fpls.2019.00009
Smith, C. M., Havlícková, H., Starkey, S., Gill, B. S., and Holubec, V. (2004). Identification of Aegilops germplasm with multiple aphid resistance. Euphytica 135, 265–273. doi: 10.1023/B:EUPH.0000013306.15133.33
Somo, M., Pirseyedi, S. M., Cai, X., and Marais, F. (2017). Engineered versions of the wheat Lr62 translocation. Crop Science. 57, 1898–1905. doi: 10.2135/cropsci2016.08.0683
Song, Z., Dai, S., Bao, T., Zuo, Y., Xiang, Q., Li, J., et al. (2020). Analysis of structural genomic diversity in Aegilops umbellulata, Ae. markgrafii, Ae. comosa, and Ae. uniaristata by Fluorescence in situ hybridization karyotyping. Front. Plant Science. 11, 710–. doi: 10.3389/fpls.2020.00710
Spetsov, P., Mingeot, D., Jacquemin, J. M., Samardjieva, K., and Marinova, E. (1997). Transfer of powdery mildew resistance from Aegilops variabilis into bread wheat. Euphytica 93, 49–54. doi: 10.1023/A:1002904123885
Steadham, J., Schulden, T., Kalia, B., Koo, D.-H., Gill, B., Bowden, R., et al. (2021). An approach for high-resolution genetic mapping of distant wild relatives of bread wheat: example of fine mapping of Lr57 and Yr40 genes. Theor. Appl. Genet. 134, 1–16. doi: 10.1007/s00122-021-03851-w
Takumi, S., Tanaka, S., Yoshida, K., and Ikeda, T. M. (2020). Production and phenotypic characterization of nascent synthetic decaploids derived from interspecific crosses between a durum wheat cultivar and hexaploid Aegilops species. Genet. Resour. Crop Evolution. 67, 1905–1917. doi: 10.1007/s10722-020-00949-w
Tiwari, V., Rawat, N., Neelam, K., Kumar, S., Randhawa, G., and Dhaliwal, H. (2010). Substitutions of 2S and 7U chromosomes of Aegilops kotschyi in wheat enhance grain iron and zinc concentration. Theor. Appl. Genet. 121, 259–269. doi: 10.1007/s00122-010-1307-8
Tsujimoto, H. and Tsunewaki, K. (1985). Hybrid dysgenesis in common wheat caused by gametocidal genes. Japanese J. Genet. 60, 565–578. doi: 10.1266/jjg.60.565
Tsunewaki, K. (2009). Plasmon analysis in the Triticum-Aegilops complex. Breed. Science. 59, 455–470. doi: 10.1270/jsbbs.59.455
Tsunewaki, K., Endo, T. R., and Mukai, Y. (1974). Further discovery of alien cytoplasms inducing haploids and twins in common wheat. Theor. Appl. Genet. 45, 104–109. doi: 10.1007/BF00291138
Tyryshkin, L. G. and Kolesova, M. A. (2020). The use of molecular-genetic and phytopathological methods to identify genes for effective leaf rust resistance in Aegilops accessions. Proc. Appl. Botany Genet. Breeding. 181, 87–95. doi: 10.30901/2227-8834-2020-2-87-95
Ulaszewski, W., Belter, J., Wiśniewska, H., Szymczak, J., Bobrowska, R., Phillips, D., et al. (2019). Recovery of 2R.2S Triticale-Aegilops kotschyi Robertsonian chromosome translocations. Agronomy 9, 646. doi: 10.3390/agronomy9100646
Valkoun, J., Hammer, K., Kučerová, D., and Bartoš, P. (1985). Disease resistance in the genus Aegilops L. — stem rust, leaf rust, stripe rust, and powdery mildew. Die Kulturpflanze. 33, 133–153. doi: 10.1007/BF01997267
Van Slageren, M. W. (1994). Wild Wheats: a monograph of Aegilops L. and Amblyopyrum (Jaub. et Spach) Eig (Poaceae) (Wageningen: Wageningen Agricultural University, Wageningen and ICARDA, Aleppo, Syria), 514 p.
Verma, S. K., Kumar, S., Sheikh, I., Malik, S., Mathpal, P., Chugh, V., et al. (2016a). Transfer of useful variability of high grain iron and zinc from Aegilops kotschyi into wheat through seed irradiation approach. Int. J. Radiat. Biol. 92, 132–139. doi: 10.3109/09553002.2016.1135263
Verma, S. K., Kumar, S., Sheikh, I., Sharma, P., Mathpal, P., Malik, S., et al. (2016b). Induced homoeologous pairing for transfer of useful variability for high grain Fe and Zn from Aegilops kotschyi into wheat. Plant Mol. Biol. Reporter. 34, 1083–1094. doi: 10.1007/s11105-016-0989-8
Volkova, G. V., Anpilogova, L. K., Andronova, A. E., Kremneva, O. Y., and Kovalenko, L. C. (2008). Sources of wheat and goat grass resistance to leaf rust, stripe rust, tan spot, and Septoria leaf blotch for development cultivars and hybrids with multiple resistance to hazardous organisms. Catalog of VIR collection 768, 25.
Wang, Y., Abrouk, M., Gourdoupis, S., Koo, D.-H., Karafiátová, M., Molnár, I., et al. (2023). An unusual tandem kinase fusion protein confers leaf rust resistance in wheat. Nat. Genet. 55, 914–920. doi: 10.1038/s41588-023-01401-2
Wang, Y., Cheng, X., Yang, X., Wang, C., Zhang, H., Deng, P., et al. (2021). Molecular cytogenetics for a wheat–Aegilops geniculata 3Mg alien addition line with resistance to stripe rust and powdery mildew. BMC Plant Biol. 21, 575. doi: 10.1186/s12870-021-03360-4
Wang, Y., Fan, J., Xiao, Y., Feng, X., Zhang, H., Chen, C., et al. (2022). Genetic analysis of resistance to powdery mildew on 7Mg chromosome of wheat–Aegilops geniculata, development and utilization of specific molecular markers. BMC Plant Biol. 22, 564. doi: 10.1186/s12870-022-03934-w
Wang, H. Y., Liu, Z. H., Chen, P. D., and Wang, X. E. (2012). Irradiation - facilitated chromosomal translocation: wheat as an example (Vienna, Austria: CABI), 223–239. doi: 10.1079/9781780640853.0223
Wang, H., Yu, Z., Li, G., and Yang, Z. (2019). Diversified chromosome rearrangements detected in a wheat–Dasypyrum breviaristatum substitution line induced by gamma-ray irradiation. Plants (Basel). 8, 1–14. doi: 10.3390/plants8060175
Warham, E. J., Abdul, M.-K., and Rosas, V. (1986). Karnal bunt (Tilletia indica) resistance screening of Aegilops species and their practical utilization for Triticum aestivum improvement. Can. J. Plant Pathology. 8, 65–70. doi: 10.1080/07060668609501844
Wheat Gene Catalogue (2024). Komugi - Wheat Genetic Resource Database. Available online at: http://shigen.nig.ac.jp/wheat/komugi/genes/symbolClassList.jsp (Accessed March 31, 2025).
Worthington, M., Lyerly, J., Petersen, S., Brown-Guedira, G., Marshall, D., Cowger, C., et al. (2014). MlUM15: an Aegilops neglecta-derived powdery mildew resistance gene in common wheat. Crop Science. 54, 1397–1406. doi: 10.2135/cropsci2013.09.0634
Yang, X., Xu, M., Wang, Y., Cheng, X., Huang, C., Zhang, H., et al. (2022). Development and molecular cytogenetic identification of two wheat-Aegilops geniculata Roth 7Mg chromosome substitution lines with resistance to Fusarium head blight, powdery mildew and stripe rust. Int. J. Mol. Sci. 23, 7056. Available online at: https://www.mdpi.com/1422-0067/23/13/7056 (Accessed June 13, 2025).
Yu, M. Q. and Jahier, J. (1992). Origin of Sv genome of Aegilops variabilis and utilization of the Sv as analyser of the S genome of the Aegilops species in the Sitopsis section. Plant Breeding. 108, 290–295. doi: 10.1111/j.1439-0523.1992.tb00133.x
Yu, M. Q., Jahier, J., and Person-Dedryver, F. (1992). Genetics of two mechanisms of resistance to Meloidogyne naasi (FRANKLIN) in an Aegilops variabilis Eig. accession. Euphytica 58, 267–273. doi: 10.1007/BF00025259
Yu, M., Jahier, J., and Person-Dedryver, F. (1995). Chromosomal location of a gene (Rkn-mnl) for resistance to the root-knot nematode transferred into wheat from Aegilops variabilis. Plant Breed. 1995, 358–360. doi: 10.1111/j.1439-0523.1995.tb01250.x
Yu, M. Q., Person-Dedryver, J., and Jahier, J. (1990). Resistance to root knot nematode, Meloidogyne naasi Franklin transferred from Aegilops variabilis Eig. to bread wheat. Agronomie 10, 451–456. doi: 10.1051/agro:19900603
Zaharieva, M., Dimov, A., Stankova, P., David, J., and Monneveux, P. (2003). Morphological diversity and potential interest for wheat improvement of three Aegilops L. species from Bulgaria. Genet. Resour. Crop Evolution. 50, 507–517. doi: 10.1023/A:1023938621936
Zaharieva, M., Gaulin, E., Havaux, M., Acevedo, E., and Monneveux, P. (2001). Drought and heat responses in the wild wheat relative Aegilops geniculata Roth: Potential interest for wheat improvement. Crop Science. 41, 1321–1329. doi: 10.2135/cropsci2001.4141321x
Zeller, F. J., Kong, L., Hartl, L., Mohler, V., and Hsam, S. L. K. (2002). Chromosomal location of genes for resistance to powdery mildew in common wheat (Triticum aestivum L. em Thell.) 7. Gene Pm29 in line Pova. Euphytica 123, 187–194. doi: 10.1023/A:1014944619304
Zhang, P., Dundas, I. S., McIntosh, R. A., Xu, S. S., Park, R. F., Gill, B. S., et al. (2015). “Wheat– Aegilops introgressions,” in Alien Introgression in Wheat Cytogenetics, Molecular Biology, and Genomics. Eds. Molnár-Láng, M., Ceoloni, C., and Doležel, J. (Springer International Publishing, Switzerland), 221–244.
Zhang, H. B., Jia, J., Gale, M. D., and Devos, K. M. (1998). Relationships between the chromosomes of Aegilops umbellulata and wheat. Theor. Appl. Genet. 96, 69–75. doi: 10.1007/s001220050710
Zhao, H., Zhang, W., Wang, J., Li, F., Cui, F., Ji, J., et al. (2013). Comparative study on drought tolerance of wheat and wheat-Aegilops biuncialis 6Ub addition lines. J. Food Agric. Environment. 11, 1046–1052.
Zhou, J. P., Yao, C. H., Yang, E. N., Yin, M. Q., Liu, C., and Ren, Z. L. (2014). Characterization of a new wheat-Aegilops biuncialis addition line conferring quality-associated HMW glutenin subunits. Genet. Mol. Res. 13, 660–669. doi: 10.4238/2014.January.28.11
Zhu, Z., Zhou, R., Kong, X., Dong, Y., and Jia, J. (2006). Microsatellite marker identification of a Triticum aestivum -Aegilops umbellulata substitution line with powdery mildew resistance. Euphytica 150, 149–153. doi: 10.1007/s10681-006-9103-x
Keywords: wheat, Aegilops, u-genome, interspecific hybridization, alien introgression, agriculturally valuable traits
Citation: Badaeva ED, González Franco MJ, Razumova O, Tereshchenko NA and Divashuk M (2025) Perspectives on the utilization of Aegilops species containing the U genome in wheat breeding: a review. Front. Plant Sci. 16:1661257. doi: 10.3389/fpls.2025.1661257
Received: 07 July 2025; Accepted: 09 October 2025;
Published: 27 October 2025.
Edited by:
Yi Tian, Agricultural University of Hebei, ChinaReviewed by:
Panagiotis Madesis, University of Thessaly, GreeceGuanglong Hu, Beijing Academy of Agricultural and Forestry Sciences, China
Copyright © 2025 Badaeva, González Franco, Razumova, Tereshchenko and Divashuk. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ekaterina D. Badaeva, a2F0ZXJpbmFiYWRhZXZhQGdtYWlsLmNvbQ==
†ORCID: Ekaterina D. Badaeva, orcid.org/0000-0001-7101-9639
Maida Jazmin González Franco, orcid.org/0009-0004-0283-898X
Olga Razumova, orcid.org/0000-0001-5157-0087
Nadezhda A. Tereshchenko, orcid.org/0000-0001-9766-3000
Mikhail Divashuk, orcid.org/0000-0001-6221-3659