 Juan B. Fontanet-Manzaneque
Juan B. Fontanet-Manzaneque Daniela M. Hernández
Daniela M. Hernández Andrea Giordano1,2
Andrea Giordano1,2 Ana I. Caño-Delgado
Ana I. Caño-Delgado- 1Department of Molecular Genetics, Centre for Research in Agricultural Genomics (CRAG) CSIC-IRTA-UAB-UB, Barcelona, Spain
- 2PLANeT Biotech, Parc de Recerca UAB, Barcelona, Spain
Climate change is intensifying drought events, posing a major threat to global food security. Sorghum bicolor (L.) Moench (Sorghum), a C4 monocot grass, is emerging as a valuable model for drought research due to its natural tolerance to water limitation and adaptability to semi-arid and arid environments. Sorghum cultivation requires significantly less water than major cereals such as rice, maize, and wheat, making it an attractive crop for sustaining agricultural productivity under water-limiting conditions. In fact, Sorghum uses up to 34% less water than rice in rainfed systems and up to 50% less under irrigation, with rice-to-Sorghum substitution potentially reducing water demand by 33%. Its lower water requirements, along with the compact growth of commonly used accessions such as TX430 and BTx623, make Sorghum a practical system for experimentation, particularly in genome editing studies. Maize, which shares close genetic similarity and also belongs to the Panicoideae subfamily, could particularly benefit from Sorghum-based insights. Sorghum also overcomes key limitations of model species such as Arabidopsis thaliana, offering greater relevance to monocot crops. Additionally, advances in metabolomics, transcriptomics, proteomics, phenomics, population genomics and pangenomics are expanding our understanding of the molecular and physiological mechanisms underlying Sorghum’s drought resilience. Despite these advantages, challenges remain in transformation efficiency and the availability of genomic tools. This review highlights Sorghum’s drought tolerance mechanisms, available omics and genetic tools, described drought-related genes and regulatory networks, and the limitations and progress in gene manipulation for climate-resilient crop development. Sorghum uniquely combines the advantages of a staple crop and a model organism, making it a powerful next-generation system for climate-resilient agriculture.
1 Introduction
Climate change has intensified the frequency of extreme events, such as heatwaves and droughts, since the 1950s, exposing millions to food and water insecurity, particularly in Africa, Asia, and Central and South America (IPCC, 2023). Heat and drought are the primary contributors to crop yield reduction under climate change (Rezaei et al., 2023). However, drought alone represents the greatest limitation to food production, causing more annual crop yield loss than all pathogens combined, with an estimated $30 billion in losses over the past decade (Gupta et al., 2020). Cereal crops are especially vulnerable: a 40% reduction in water availability leads to a 40% decrease in maize yield and a 20% decrease in wheat yield, while a 50% reduction in water results in a 60% decline in rice yield and a 30% decline in Sorghum yield (Daryanto et al., 2017). Moreover, even short periods of drought can significantly impact final yield outcomes, as the timing of drought within the plant cycle is critical, especially during germination, seedling establishment, floral induction, and grain development (Dietz et al., 2021; Barnabás et al., 2008).
In addition to climate change, demographic changes contribute to food security challenges. The global population is projected to increase from the current 8 billion to between 9.7 and 10.9 billion by the end of the century, raising global food demand by 35-62% by 2050 (United Nations, 2022; Van Dijk et al., 2021).
In conclusion, the combined pressures of climate change and a growing global population underscore the urgent need for crops better adapted to changing environments to meet the increasing food demand and ensure food security. However, many major cereal crops remain poorly equipped to withstand increasingly frequent and severe drought conditions. While Arabidopsis thaliana (Arabidopsis) has been instrumental in advancing our understanding of plant stress responses, its evolutionary distance from monocots limits its direct applicability to cereal crop improvement. To accelerate the development of climate-resilient cereals, new model systems are needed that combine experimental tractability with close genetic and physiological relevance to key crops. Sorghum bicolor (L.) Moench (Sorghum) emerges as a promising candidate.
Sorghum, a C4 monocot grass that diverged from maize approximately 15 million years ago, is the fifth most important cereal crop worldwide, with an annual production of around 57 million tons (Mullet et al., 2002; FAOSTAT, 2022). While serving as a staple food source for millions in Africa and Asia, Sorghum remains underutilized in more industrialized countries, where it is primarily grown for animal feed. Nonetheless, Sorghum holds considerable potential for human consumption, particularly as a gluten-free alternative. Sorghum-based food products, including bread, pasta, porridges, and parboiled rice-like products, present valuable dietary options, especially for people with celiac disease (Taylor et al., 2006).
In addition to its nutritional value, sweet Sorghum has emerged as a particularly promising bioenergy crop, particularly for bioethanol production, due to its high biomass yields on marginal lands not suitable for food or feed cultivation. Breeding efforts have enhanced traits such as sugar accumulation and secondary cell wall biosynthesis, further supporting its utility in biofuel applications (Guigou et al., 2011; Zhang et al., 2018).
The primary benefit of Sorghum cultivation is its drought tolerance mechanisms. Sorghum is predominantly grown in semi-arid and arid tropics of Africa and South Asia, with significant production also in China, Southeast Asia, and the Americas (Venkateswaran et al., 2019). Consequently, drought stress is considered the most common abiotic stressor that Sorghum encounters in its major production regions (Figure 1). Sorghum’s high protein content and minimal irrigation requirements make it an attractive alternative to water-intensive crops such as rice. For instance, replacing rice with sorghum could reduce water demand by 33% while increasing protein production by 1% (Davis et al., 2018). Moreover, sorghum demonstrates greater resilience to high temperatures compared to wheat, reinforcing its value under climate change scenarios (DeFries et al., 2023).
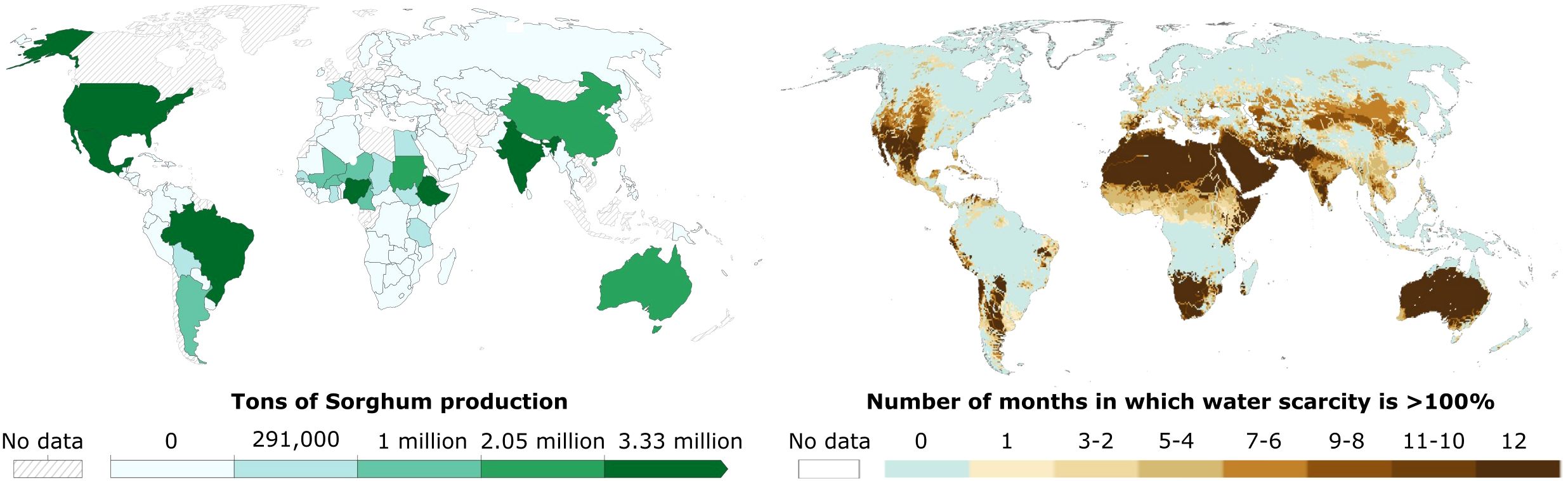
Figure 1. Drought is the sorghum’s most common abiotic stressor. Sorghum production is associated with water scarcity regions. In the left map there is the production of sorghum in 2023 (Food and Agriculture Organization of the United Nations (2025) – with major processing by Our World in Data) and in the right map the number of months/year with water scarcity (Adapted from: Figure Box 4.1.1 in Caretta, M.A., A. Mukherji, M. Arfanuzzaman, R.A. Betts, A. Gelfan, Y. Hirabayashi, T.K. Lissner, J. Liu, E. Lopez Gunn, R. Morgan, S. Mwanga, and S. Supratid, 2022: Water. In: Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [H.-O. Pörtner, D.C. Roberts, M. Tignor, E.S. Poloczanska, K. Mintenbeck, A. Alegría, M. Craig, S. Langsdorf, S. Löschke, V. Möller, A. Okem, B. Rama (eds.)]. Cambridge University Press, Cambridge, UK and New York, NY, USA, pp. 551-712, doi:10.1017/9781009325844.006).
Beyond its agronomic and ecological advantages, sorghum possesses several features that enhance its value as a model system. It has a relatively small diploid genome, in contrast to the tetraploid durum wheat, the allohexaploid bread wheat, or maize, which—though diploid—originated from a tetraploid ancestor, increasing its genetic redundancy (Swigonova et al., 2004; Maccaferri et al., 2019; Cavalet-Giorsa et al., 2024). Sorghum also develops a primary root that is amenable to physiological studies and confocal imaging in a manner comparable to the model dicot Arabidopsis (Blasco-Escámez et al., 2017; Fontanet-Manzaneque et al., 2024b; Rico-Medina et al., 2025). Furthermore, its life cycle is shorter than that of wheat and maize and can be further reduced through embryo rescue techniques (Rizal et al., 2014).
Together, these features, combined with the availability of omics resources, advancing genetic tools, recent progress in overcoming transformation recalcitrance, and an expanding body of knowledge on drought-responsive regulatory networks, position sorghum as a next-generation model for studying drought adaptation and for driving crop improvement strategies aimed at enhancing global food security.
2 Sorghum drought tolerance mechanisms
Sorghum is a key crop in water-limited environments. Its drought resilience is driven by a combination of morphological, physiological, and genetic adaptations that allow the plant to maintain productivity under water stress conditions (Figure 2). Physiological drought tolerance can be broadly defined as the plant’s ability to sustain photosynthetic carbon assimilation and regulate transpiration under water deficit conditions (Tardieu et al., 2018).
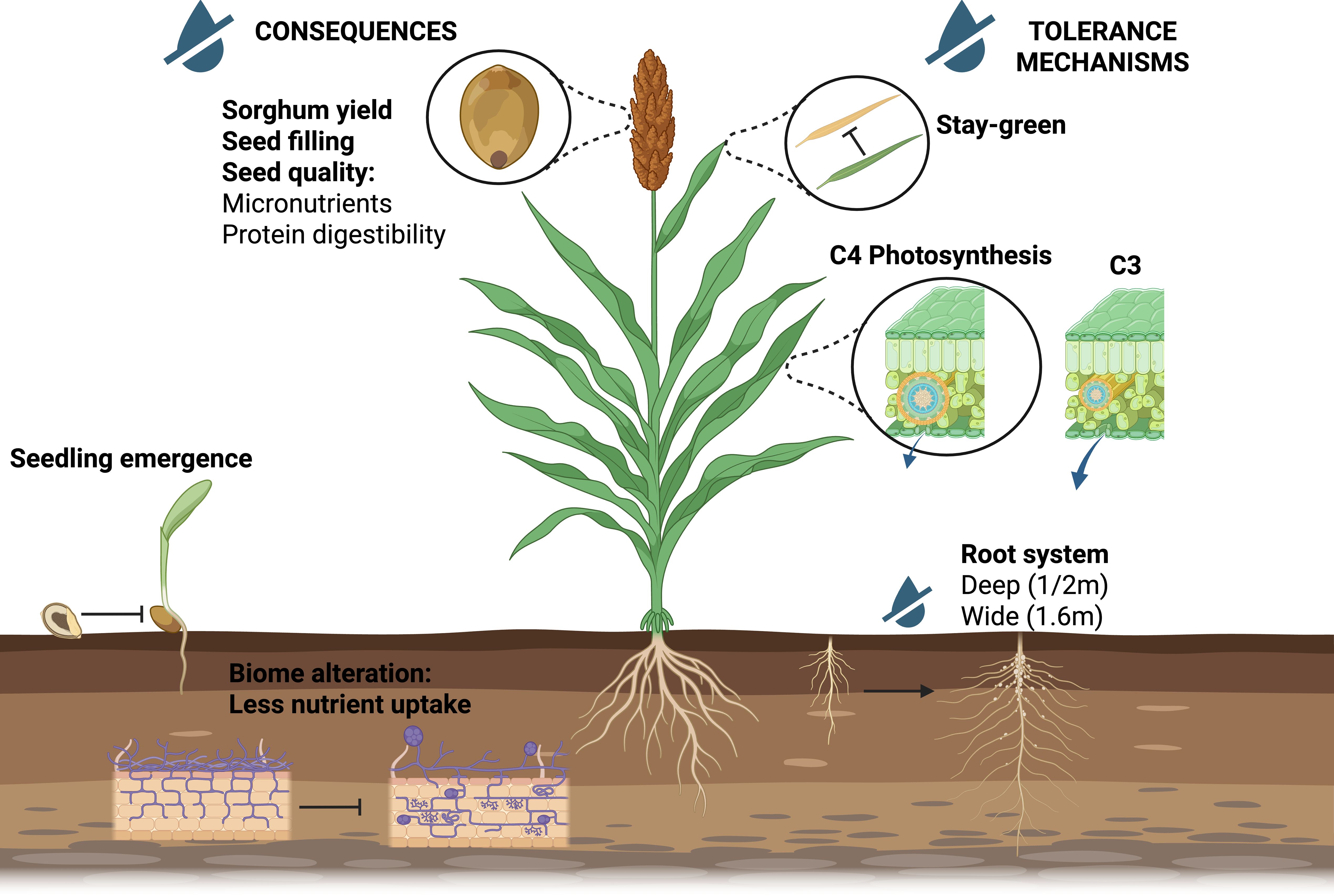
Figure 2. The impact of drought on Sorghum and its tolerance mechanisms. Drought stress affects Sorghum development at various stages, including seedling emergence, root establishment, and seed filling. However, Sorghum possesses several mechanisms to cope with this abiotic stress, such as deep and extensive root system, C4 photosynthesis to minimize water loss, and stay-green traits. Created in BioRender. Fontanet-Manzaneque, J. B. (2025) https://BioRender.com/91z4p60.
In the first physiological component, light capture and assimilation, Sorghum exhibits the retention of photosynthetically active leaves during periods of post-flowering drought, thereby extending the grain-filling period, a phenomenon known as the stay-green trait (Stg) (Borrell et al., 2000). This complex trait, involving different physiological processes, has been linked to four major quantitative trait loci (QTLs) known as Stg1–4 which are responsible for half of the phenotypic variance observed in stay-green genotypes (Xu et al., 2000; Sanchez et al., 2002). Subsequent studies have revealed that each Stg individually reduces post-flowering drought-induced leaf senescence, and Stg2 has been identified as the most prominent QTL, showing the greatest delay in leaf senescence, the highest green leaf area at maturity, and the lowest senescence rates among the individual QTLs (Harris et al., 2007). In experiments studying the interactions of these QTLs to determine the percentage of explained phenotypic variation, Stg2 alone accounted for 30% of the variation, and the Stg2+Stg3 combination explained nearly the 50% (49.8%), exceeding the sum of all individual effects (Subudhi et al., 2000; Xu et al., 2000).
In the second component, transpiration, the presence of each individual QTL (Stg1-4) in Sorghum cultivars is also characterized by a reduction in canopy and tillering at the anthesis stage, leading to a decrease in transpirational leaf area and better conservation of soil water prior to anthesis, for subsequent use during grain filling (Borrell et al., 2014). In this regard, it is noteworthy that the relationship between Stg traits and yield, although influenced by both environmental conditions and genetic background, exhibits a positive association in environments where yield is below 6 tons/ha. Consequently, the adoption of Sorghum hybrids with Stg could prove beneficial in enhancing yields across a wide range of environments, particularly since average yields worldwide are below 2.5 tons/ha (Jordan et al., 2012).
Additionally, Sorghum possesses the C4 photosynthetic pathway, which is crucial for its drought tolerance (Figure 2). Unlike many other plants that utilize the C3 photosynthesis pathway, in which CO2 is initially converted to a C3 compound, Sorghum rapidly produces C4 compounds using a specialized enzyme, phosphoenolpyruvate carboxylase, thereby reducing water loss through stomata (Pennisi, 2009). The CO2-concentrating mechanisms in C4 plants also avoids the oxygenase activity of RuBisCO, which is induced upon stomatal closure, thereby reducing photorespiration and improving carbon-use efficiency (Sage et al., 2012). This photosynthetic advantage, typical of plants adapted to harsh environments, relies on a specialized leaf anatomy known as Kranz anatomy. This anatomy, mainly defined as two distinct concentric layers of chlorenchyma cells, formed by a bundle sheath, surrounded by an outer layer of mesophyll cells, allows the segregation of the C4 synthesis and Calvin cycle in different cell types, having a CO2 concentration in the chlorenchyma cells (Lundgren et al., 2014).
Stomatal regulation is a central physiological mechanism enabling Sorghum to maintain productivity under drought conditions by balancing water conservation and carbon assimilation (Tari et al., 2013). Genotypic variability in stomatal behavior reflects distinct drought coping strategies: isohydric genotypes, such as ‘Gadambalia’, tend to close their stomata early to conserve water, though at the cost of reduced photosynthesis; whereas anisohydric types like ‘Tabat’ maintain stomatal opening and gas exchange even at lower water potentials (Tsuji et al., 2003).
Recent research has demonstrated that sorghum exhibits rapid stomatal kinetics, adjusting conductance within minutes of environmental changes. This dynamic response helps sustain photosynthesis and improves intrinsic water-use efficiency (WUE) under fluctuating conditions (Battle et al., 2024). Studies also associate faster stomatal closure, higher stomatal density, and narrower operational apertures with enhanced WUE, without compromising carbon assimilation (Al-Salman et al., 2023).
Considerable genetic variation among Sorghum genotypes has been reported in traits such as net carbon assimilation rate (A), transpiration rate (E), the A:E ratio, and WUE, under both well-watered and drought conditions. Increases in A:E and WUE have been linked to improved drought resilience, particularly during the pre-flowering stage (Balota et al., 2008). The genetic basis of stomatal conductance has also been explored, with associated QTLs identified (Lopez et al., 2017).
Beyond stomatal regulation, Sorghum also minimizes non-stomatal water loss through the deposition of epicuticular waxes on leaf surfaces. These hydrophobic layers form a barrier against cuticular transpiration, particularly under low relative humidity, to increase WUE by reducing the cuticular conductance to water vapor (Surwenshi et al., 2010).
Development and phenotypic analysis of wax-deficient mutants has facilitated the identification of key genetic components involved in cuticle and wax biosynthesis (Peterson et al., 1982; Jenks et al., 2000; Peters et al., 2009). These include QTL mapping, association studies, and map-based cloning (Burow et al., 2008, 2009; Awika et al., 2017; Punnuri et al., 2017; Uttam et al., 2017; Elango et al., 2020). More recently, spatial and developmental variation in wax composition across sorghum organs has been explored, alongside gene expression profiling of wax-related genes (Chemelewski et al., 2023).
In addition to its physiological advantages, the morphological feature of Sorghum that contributes to its drought tolerance is its root system (Figure 2). Sorghum roots can penetrate depths of 1 to 2 meters and efficiently extract water up to a lateral distance of 1.6 meters from the plant. This deep root system allows Sorghum plants to access moisture from deeper soil layers (Krupa et al., 2017). Experimental studies have demonstrated that Sorghum plants with deeper root systems exhibit higher yields and greater dry matter weight, which also influences the photosynthetic machinery (Chen et al., 2020). Root removal at 30 or 60 cm depths caused significant reductions in dry matter accumulation, photosynthesis and ultimately yield (Chen et al., 2020). In addition, several comparative studies with maize have further supported the direct correlation of deeper root systems with better drought adaptation. While maize exhibits higher above-ground dry weight in control conditions, Sweet sorghum showed a 27.2% increase in above-ground dry weight and a 200% increase in root dry weight under drought conditions in comparison with maize, highlighting the functional value of sorghum root biomass in drought tolerance (Schittenhelm and Schroetter, 2014).
Moreover, genetic studies have shown that QTLs for root dry weight (qRDW) and nodal root angle (qRA), which affects root density and distribution in the soil, are co-located with QTLs for stay-green traits and associated with better drought adaptation (Mace et al., 2012). This indicates that root angle and architecture play a role in influencing yield under drought stress conditions, even though there may not be a direct relationship with plant size (Singh et al., 2011).
QTL pyramiding has emerged as a strategic breeding approach in sorghum to enhance drought tolerance and productivity. This involves the combination of multiple beneficial QTLs into a single genotype using marker-assisted selection, though the process is labor-intensive and requires careful compatibility of gene activity (Kadam and Fakrudin, 2017; Nagaraja Reddy et al., 2013). When applied to traits like stay-green, pyramiding improves the potential expression of drought tolerance, as demonstrated by introgression lines containing Stg3 Stg1 and Stg2, which showed only a 10% yield reduction under water-stressed conditions compared to 18–23% of other Stg combinations, indicating enhanced tolerance (Kamal et al., 2018). Ultimately, only QTL combinations that demonstrate consistent expression across multi-environment trials are selected for varietal release, ensuring both yield stability and stress resilience (Gorthy et al., 2017; Kebede et al., 2001).
Summarizing the genetic linkage of the described traits, three main clusters can be identified: Stg1 and Stg2 QTLs on chromosome 3; Stg3 and the root dry weight QTL qRDW1_2 on chromosome 2; and Stg4, qRDW1_5 and the root angle QTL qRA1_5 on chromosome 5 (Xu et al., 2000; Mace et al., 2012; Deshpande et al., 2017).
3 Effects of drought stress on Sorghum
Despite being a drought-tolerant crop and thriving under low-input conditions, drought stress remains the most prevalent abiotic stressor faced by Sorghum in key production regions (Figure 1; Abreha et al., 2022). Drought stress impacts Sorghum development from the early stages, with seedling death during emergence and establishment being a common occurrence in drylands (Figure 2). Drought significantly hinders the germination rate in Sorghum (Smith et al., 1989). Subsequent studies have identified that the primary losses due to drought occur at various stages of early seedling development, including germination, emergence, seedling growth, and the lengths of coleoptiles, mesocotyls, radicles, and primary shoots (Bayu et al., 2005). Further investigations demonstrated the negative early effects of drought on germination rate and time, root and shoot length, dry matter, and seedling vigor (Queiroz et al., 2019).
Additionally, drought stress impacts Sorghum’s ability to uptake, mobilize and transport soil nutrients by altering the root-associated microbiome, specifically affecting symbiotic arbuscular mycorrhizal fungi (Symanczik et al., 2018; Figure 2).
Ultimately, drought is affecting its primary production trait, yield (Sarshad et al., 2021). However, the impact of water deficit on yield is highly dependent on growth stage and timing. Drought stress can reduce Sorghum yield by up to 36% when it occurs during the vegetative stage, and by as much as 55% during reproductive stages (Assefa et al., 2010). Furthermore, drought not only adversely affects Sorghum yield but also seed filling and grain quality (Sehgal et al., 2018; Figure 2). Total starch, amylose, and amylopectin content are reduced in grains exposed to drought during flowering (Bing et al., 2014). Water deficit also decreases protein digestibility and reduces micronutrient content, specifically zinc, manganese, and copper, in Sorghum grain nutrition (Impa et al., 2019).
Given the significant impact of drought stress on Sorghum, considerable attention must be directed towards developing tolerant cultivars and implementing efficient mitigation strategies in Sorghum production. Such strategies may include the use of early maturing varieties (Yahaya and Shimelis, 2022), optimized irrigation practices such as low irrigation at panicle initiation and termination at grain filling (Araya et al., 2018), marker-assisted selection and exploitation of stay-green traits (Mwamahonje et al., 2021), and genome editing approaches to develop improved drought-tolerant varieties.
4 Genetic and multi-omics resources available to improve abiotic stress resistance in sorghum
4.1 Sources of genetic variation for drought tolerance
While most cultivars and varieties of sorghum often comprise a very narrow genetic diversity, a valuable and underexploited source of drought tolerance is found in sorghum landraces and wild sorghum relatives unconstrained by domestication or stringent breeding (Ahmed et al., 2024, Sauer et al., 2024; Enyew et al., 2022; Ochieng et al., 2021; Nagesh Kumar et al., 2021). Wild and landrace sorghum germplasm represents a broad pool of new alleles for traits of drought resistance that can be applied to sorghum breeding.
Capturing and ensuring the conservation of such diversity has been the objective of many repositories such as the USDA Agriculture Research Service National Plant Germplasm System, currently holding over 7200 accessions of Ethiopian sorghum lines (Cuevas et al., 2017). Likewise, utilizing chemical mutagenesis techniques such as Ethyl Methane Sulfonate (EMS) mutagenesis and Targeting Induced Local Lesion In Genomes (TILLING) is a proven approach for producing new sorghum germplasm (Jiao et al., 2016; Blomstedt et al., 2012; Xin et al., 2009; Mason et al., 2024; Xin et al., 2008; Kalpande et al., 2022), which have already enabled the basic study of loci implicated in drought response and tolerance mechanisms in Sorghum (Fontanet‐Manzaneque et al., 2024b; Harris-Shultz et al., 2019).
4.2 Genomic resources and advancements in pangenomics
Sorghum molecular research was greatly boosted by the publication of the first reference genome (BTx623) in 2009 by Paterson et al., 2009 and further improved in McCormick et al. (2018), who already envisioned Sorghum, with its relatively small genome of around 800 Mb and 34,211 bona fide protein-coding genes, as a potential model species for C4 grasses. Molecular breeding approaches of Sorghum heavily rely on the BTx623, Tx430 (Deschamps et al., 2018) and Rio (Cooper et al., 2019) accession assemblies, with the BTx623 accession being the most used as a parent for grain hybrid generation.
Owing to the technical advancements, whole-genome sequencing in population and quantitative genomics studies has been useful for characterizing the variability of agronomic traits in sorghum (Faye et al., 2019; Morris et al., 2013). Resources like the West African sorghum association panel (WASAP), consisting of sorghum landraces and breeding lines from West Africa, allow the study of agronomical traits of interest (Faye et al., 2021a) with diverse accessions. WASAP has been applied in GWAS-based studies for identifying novel drought-response associated loci (Faye et al., 2021b; Maina et al., 2022), and pleiotropic loci were found to be associated with various drought tolerance traits in pre- and post-flowering drought scenarios. Furthermore, genome–environment association studies, which are relevant for identifying genotype-by-environment interactions and predicting phenotypic variation, have also been reported for drought and other climatic adaptation traits (Girma et al., 2020; Menamo et al., 2021; Lasky et al., 2015).
However, understanding drought adaptation needs the added perspective of studies tackling structural variation and gene presence/absence patterns in a broad number of genotypes beyond a single reference genome, an area where pangenomics is particularly valuable. Pangenomes are a powerful tool for genetic variability characterization, as has been evidenced for species such as wheat (Jiao et al., 2025), rice (Guo et al., 2025), and other agronomically important species.
The recently published sorghum pangenome (Tao et al., 2021) constructed with 13 cultivated genomes and 5 wild inter-fertile relatives genomes reveals a high number of dispensable genes (this is, genes unique between genome accessions), among which many Copy Number Variants and Presence/Absence Variants are enriched in abiotic and biotic stress Gene Ontology processes, highlighting again this underexploited source of genetic diversity. In the case of drought, Ruperao et al. (2021) mapped available transcriptomics data from contrasting resistant vs. susceptible genotypes to a sorghum pangenome and found 1,788 genes deregulated by drought, out of which 79 were newly identified with the pangenome assembly. A new study by Cole et al. (2025) employed a pangenome assembly of three sorghum accessions, BTx642 (a Stg genotype), Tx430, and BTx623 (senescent genotypes), to evaluate consistent drought-responsive gene expression across Tx430 and BTx623 over three consecutive years, thereby minimizing artifacts associated with using a single reference genome.
4.3 Omic profile of Sorghum’s drought adaptation
The development of integrated omics databases, such as those compiled by Liu et al. (2024), represents a valuable resource for the genetic improvement of Sorghum. Genomics, transcriptomics, proteomics, metabolomics and phenomics studies have evaluated Sorghum’s responses to drought. As a result, many of the specific genes and pathways involved in its drought adaptation mechanisms have been identified (Table 1).
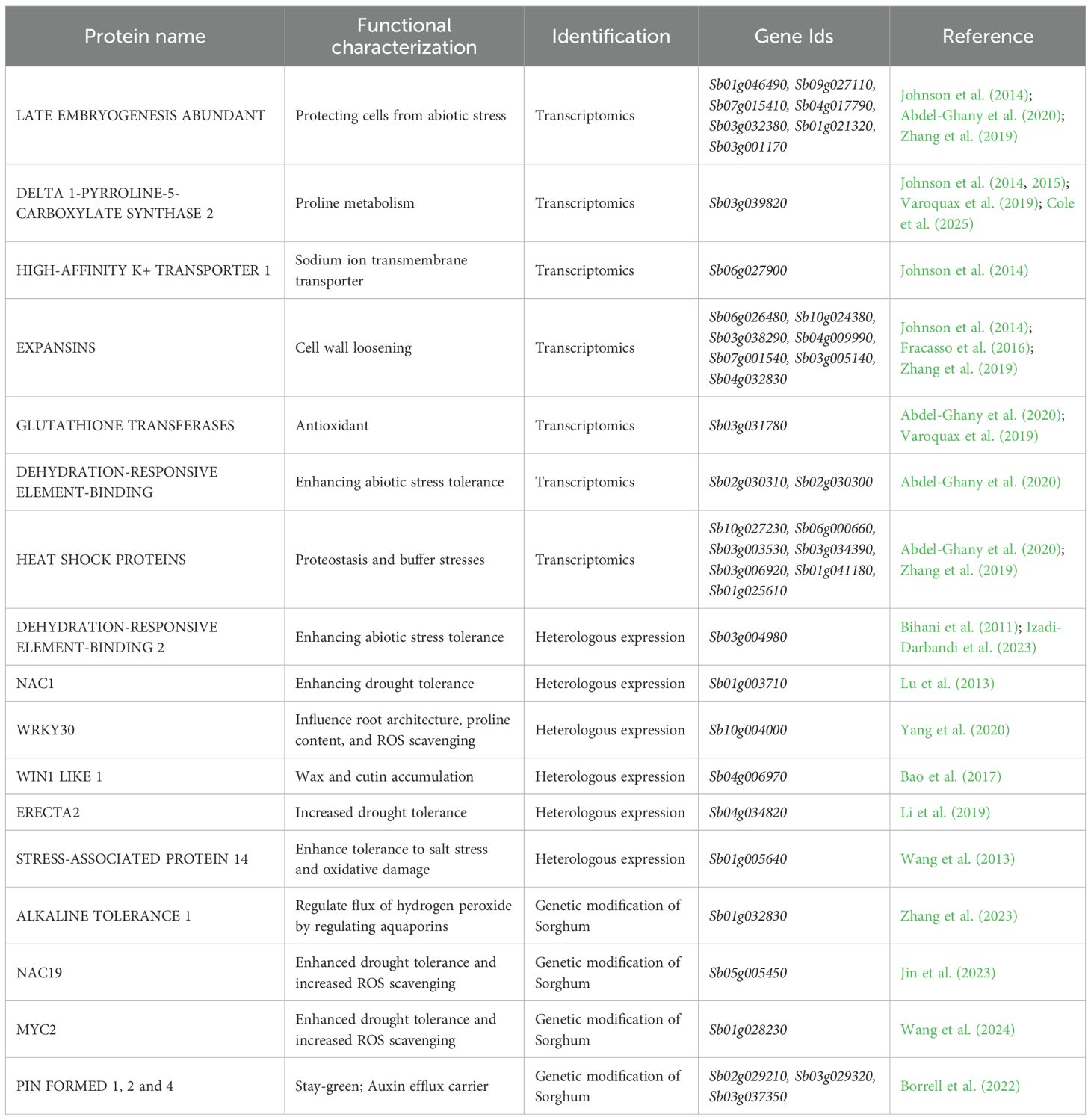
Table 1. Summary of currently available genetic tools in Sorghum.
4.4 Transcriptomics insights on drought responsive gene expression
Transcriptome profiling of Sorghum under drought stress has revealed key adaptive mechanisms. Among the most responsive genes, LATE EMBRYOGENESIS ABUNDANT (LEA) proteins, the sodium transporter HIGH-AFFINITY K+ TRANSPORTER 1 (HKT1), and the DELTA 1-PYRROLINE-5-CARBOXYLATE SYNTHASE 2 (P5CS2), involved in proline metabolism, were notably upregulated under drought conditions (Johnson et al., 2014; Abdel-Ghany et al, 2020; Zhang et al., 2019; Varoquaux et al., 2019). Later studies have consistently highlighted the importance of P5CS2 in drought adaptation. For example, P5CS2 is significantly upregulated in the Stg genotype B35 compared to the senescent genotype R16, and it maps within the Stg1 QTL (Johnson et al., 2015). However, a pangenome-based transcriptomic analysis expanding on the dataset of Varoquaux et al. (2019) identified P5CS2 as a core drought-responsive gene, being upregulated in both a Stg and a senescent genotype (Cole et al., 2025). This suggests that P5CS2-mediated proline accumulation may represent a general drought response mechanism rather than a stay-green–specific adaptation. Deregulation of glutathione transferases is also a common mechanism across Sorghum varieties in response to water deficit, suggesting a conserved drought response. Indeed, lower constitutive expression of glutathione transferase genes could explain the sensitivity upon water stress of drought-sensitive varieties (Abdel-Ghany et al., 2020; Fracasso et al., 2016).
Additionally, upregulation of expansins and HEAT SHOCK PROTEINS (HSPs) is a conserved mechanism of Sorghum to drought stress and in combination with heat stress (Johnson et al., 2014; Fracasso et al., 2016; Zhang et al., 2019). However, specific changes observed in drought-resistant varieties include upregulation of transcription factors such as DEHYDRATION-RESPONSIVE ELEMENT-BINDING PROTEINS (DREBs) and specific HSPs (Abdel-Ghany et al., 2020).
At transcript level, Sorghum possesses conserved responses involving deregulation of LEAs, HSPs, P5CS2, expansins and glutathione transferases (e.g., GST29), while stress-resilient varieties further exhibit elevated expression of DREBs and specific HSPs.
4.5 Proteomic signatures of drought adaptation in Sorghum
Compared to transcriptomics, proteomics offers a more accurate picture of the cellular state, as proteins are the actual effectors of biological activity. Proteomics data reveal post-transcriptional regulatory patterns that transcript data alone cannot provide (Naaz et al., 2024; Satrio et al., 2024).
In response to drought, Ngara et al., 2018 found that sorbitol-induced osmotic stress in Sorghum triggered increased abundance of proteins targeted to the secretory pathway, including expansins, redox proteins, proteases, and glycosyl hydrolases. In a comparative proteomic study, Goche et al., 2020 reinforced the relevance of fast stomatal closure as a key trait for drought tolerance. Other signature mechanisms seen in tolerant sorghum genotypes include the accumulation of the osmoprotectant glycine betaine and root architectural modifications that increase the root-to-shoot ratio, thereby improving water uptake efficiency.
Recently, a comprehensive quantitative proteomics dataset of two contrasting sorghum drought genotypes revealed distinct patterns of protein accumulation in leaves and roots under drought, heat, and combined stress conditions (Ali et al., 2025), providing a valuable resource to identify protein markers associated with drought tolerance and support Sorghum breeding programs.
4.6 Metabolomic profile in response drought in Sorghum
Metabolomics allows for direct profiling of the end products of drought stress response. Baker et al. (2023) investigated post-flowering drought responses of the reference genotypes BTx642 and RTx430, and observed distinct, genotype-specific accumulation of osmolytes. Metabolites such as galactinol, α-ketoglutarate, and aspartate positively correlate with stomatal conductance and decline as drought induces stomatal closure. In contrast, fumarate and maleate accumulate during drought and correlate inversely with stomatal opening, illustrating metabolic adaptation to water stress (Baker et al., 2023). Another untargeted metabolomics study explored how four genotypes (agriculturally low- or high-performing) modulate root metabolites to shape their rhizosphere microbiome under drought: the metabolome of low-performing genotypes was enriched in flavonoids, while high-performing genotypes showed greater modulation of other compounds, including pipecolinic acid, 13-(S)-hydroxyoctadecatrienoic acid (an oxylipin), and albiflorin (a terpenoid) (García et al., 2025). These findings suggest distinct metabolic strategies in root-microbiome interactions during drought. Further studies could aim to define consistent metabolic signatures linked to drought tolerance in sorghum.
4.7 Phenomics
Phenomics, which analyzes the set of observable traits in an organism, has become an essential tool for capturing genetic variation under field-relevant environmental conditions. As genomics and genotyping have become more accessible, phenotyping remains the main bottleneck in linking genetic variation to traits, due to the laborious nature and high costs of measuring traits at large scale and high throughput. Advances in imaging, sensors, and data analysis have made phenomics a valuable tool in sorghum, both for evaluating agronomic and drought tolerance traits (Bao and Tang, 2016; Salas Fernandez et al., 2017; Watanabe et al., 2017; Jadhav et al., 2024). Spindel et al. (2018) carried out a GWAS in sorghum by employing aerial drone imagery to capture phenotypic data. By analyzing patterns of historical recombination, the study identified specific genetic variants linked to key traits such as drought tolerance and biomass yield.
4.8 Integrative multi-omics for developing drought tolerant cultivars
The integration of omics approaches, while inherently challenging at the technical and analytical level, offers a more holistic understanding of complex biological processes, especially those as complex as drought response. Instead of isolated molecular markers or singular responses, multi-omics approaches identify regulatory networks and molecular interactions that collectively contribute to stress resilience in sorghum (Mukherjee et al., 2024; Seth et al., 2025; Ren et al., 2022). In this context, Yue et al. (2025) combined metabolomic and transcriptomic analyses to investigate drought responses and reaffirmed the involvement of the flavonoid biosynthetic pathway. Their results revealed that certain flavonoid biosynthesis genes correlated positively with metabolite levels, while other showed negative correlations, indicating a precise regulation under stress.
Further supporting the role of flavonoids in sorghum’s drought tolerance, Fontanet-Manzaneque et al. (2024b) employed transcriptomics, cistromics and metabolomics to demonstrate that SbBRI1 brassinosteroid receptor signaling is attenuated under drought, facilitating activation of the flavonoid pathway via BES1 transcription factor.
These integrative omics not only reveal mechanisms of stress adaptation but also directly contribute to and streamline the development of new cultivars, as they enable the identification of key genes, pathways, and observable traits associated with stress tolerance much more rapidly. Coupling this multilayered data with modern breeding techniques, such as genomic selection and marker-assisted selection (MAS), can significantly accelerate and improve the development of stress-tolerant cultivars (Hao et al., 2021; Liaqat et al., 2024).
5 Functional validation and genetic engineering of drought-responsive genes
More targeted approaches have addressed the functional validation of specific genes involved in drought tolerance mechanisms in Sorghum. However, due to the technical challenges associated with stable sorghum transformation, many genes have been validated using heterologous systems such as Arabidopsis thaliana, rice, or maize, species that are more amenable to genetic manipulation. While informative, these systems do not always replicate the full physiological context of Sorghum, and functional results may not fully translate across species.
Transcription factors such as SbDREB2, SbNAC1, and SbWRKY30 have been functionally validated in rice, maize, and Arabidopsis, where their overexpression consistently enhanced drought tolerance (Bihani et al., 2011; Lu et al., 2013; Yang et al., 2020; Izadi-Darbandi et al., 2023). Other engineering efforts include SbWINL1, a wax-inducing transcription factor that increased drought tolerance in Arabidopsis through the induction of wax and cutin biosynthesis (Bao et al., 2017); SbER2, a receptor-like kinase whose expression in maize and Arabidopsis improved water-use efficiency and photosynthetic performance under drought (Li et al., 2019); and SbSAP14, a STRESS-ASSOCIATED PROTEIN that mitigated salt-induced oxidative stress and delayed leaf senescence when expressed in rice (Wang et al., 2013).
Despite these advances in heterologous systems, only a few laboratories have successfully transformed sorghum to directly investigate gene function in planta. Notably, a Genome-Wide Association Study identified ALKALINE TOLERANCE 1 (AT1), which encodes an atypical G protein γ subunit. In Sorghum, overexpression of AT1 reduced alkaline tolerance, whereas knockout lines displayed enhanced stress resistance through increased regulation of H2O2 efflux via aquaporin phosphorylation (Zhang et al., 2023). This study highlights the mechanistic insights that can be obtained from direct genetic manipulation in Sorghum itself.
Additional examples of successful sorghum transformation include SbNAC9, whose overexpression conferred drought tolerance through improved detoxification of reactive oxygen species (ROS) (Jin et al., 2023), and SbMYC2, a bHLH transcription factor that reduced ROS accumulation under drought conditions. In contrast, silencing SbMYC2 compromised stress tolerance in sorghum seedlings (Wang et al., 2024).
Furthermore, three PIN-FORMED auxin transporters, SbPIN1, SbPIN2, and SbPIN4, linked to the Stg1–3 QTLs, were introduced into a non-stay-green sorghum cultivar. Their expression led to improved canopy development, enhanced root architecture, and increased panicle growth. Notably, these lines also exhibited reduced leaf area, which may contribute to lower transpiration rates under drought conditions (Borrell et al., 2022).
6 Sorghum transformation
Despite the identification of specific genes involved in drought-tolerant mechanisms, there is still limited progress in genetic engineering of sorghum varieties. Sorghum is still considered a crop recalcitrant to genetic transformation (Visarada and Kishore, 2015; Miller et al., 2023). Optimizing transformation efficiency is crucial for the development of Sorghum cultivars better adapted to the current climate change context.
6.1 Sorghum transformation limitations
Sorghum is considered the most recalcitrant crop among cereals for tissue culture due to the accumulation of phenolic compounds and a high degree of genotype dependence (Azhakanandam and Zhang, 2015).
Phenolic compound accumulation remains a primary bottleneck in Sorghum transformation. Several factors induce the accumulation of phenolic pigments in Sorghum in vitro cultures. Agrobacterium tumefaciens (Agrobacterium) infection and phosphinothricin selection trigger stress responses that lead Sorghum cells to release toxic levels of phenolics, compromising cell viability (Tadesse et al., 2003; Zhao et al., 2000).
Various strategies have been adopted to minimize phenolic compound accumulation such as performing short subculture intervals and adding 1% polyvinylpolypyrrolidone (Cai et al., 1987; Zhao et al., 2000); changing the selection system to mannose (Gao et al., 2005); changing phosphinothricin selection to geneticin (Tadesse et al., 2003); using activated charcoal to reduce black pigment production (Nguyen et al., 2007); and cold pre-treatment of immature seeds positively affecting both explant survival and callus formation while reducing phenolic compounds, likely due to the inhibition of key enzymes involved in phenolic compound synthesis (Dicko et al., 2006). Supplementing the media with L-asparagine (6.7 mM), L-proline (17.4 mM), and different concentrations of NO3-, NH4+, and PO4- resulted in increased induction and growth of Sorghum friable embryogenic callus without medium pigmentation (Elkonin and Pakhomova, 2000). Combining this supplemented media with geneticin selection further mitigated the compounding effect of phenolic secretion (Howe et al., 2006). These phenolic mitigation strategies are now widely integrated into modern protocols and are summarized in Table 2, alongside transformation efficiencies.
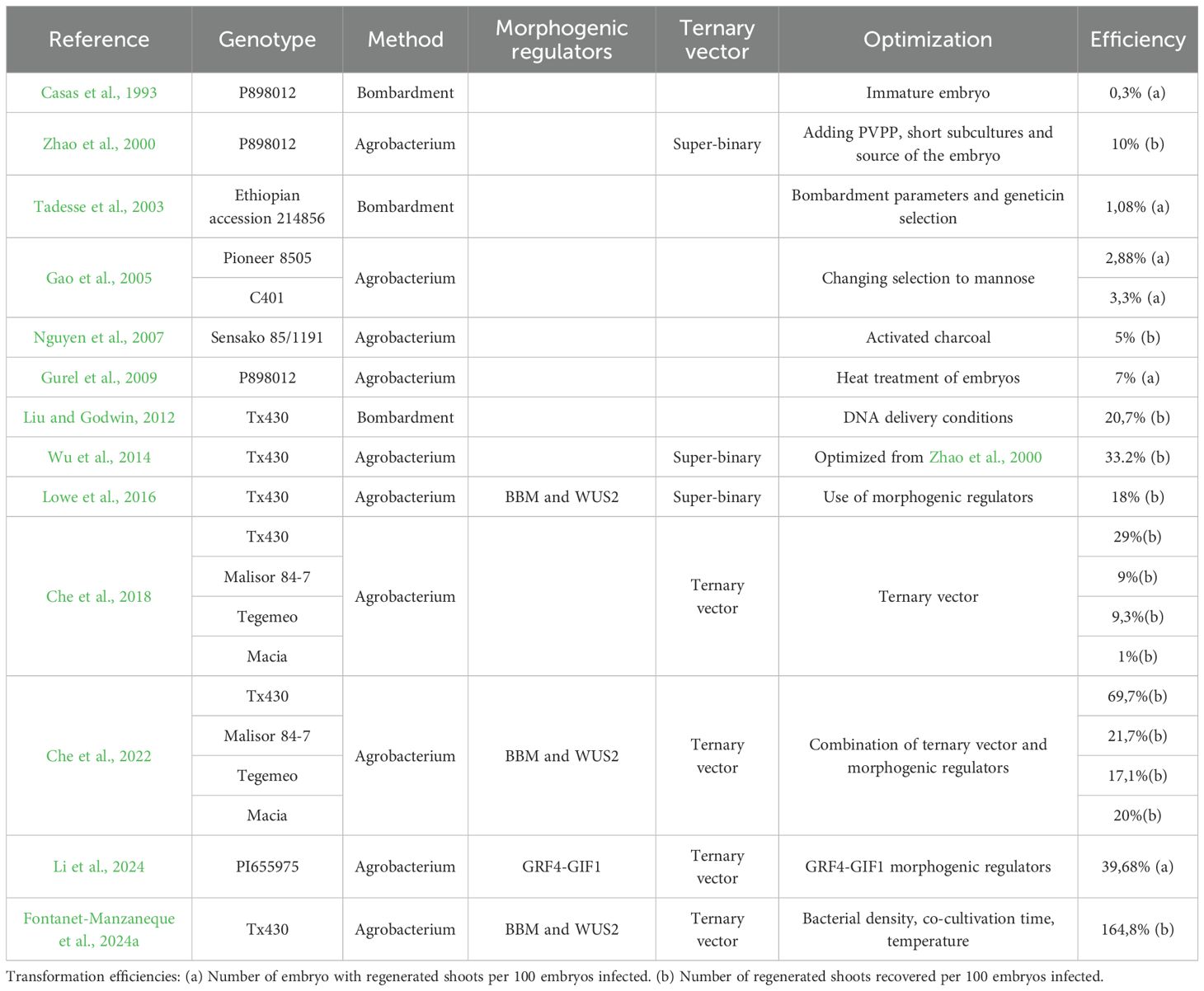
Table 2. Summary of the progress in Sorghum transformation.
Genotype dependence poses an equally critical limitation in Sorghum transformation. Sorghum callus induction and regeneration have also been quite limited by genotype. Even with established transformation methods, many elite and agronomically important varieties remain poorly responsive (Botella, 2019; Kaeppler and Pedersen, 1997; Hagio, 2002).
However, the implementation of new methodologies to ameliorate phenolic toxicity and increase regeneration frequencies has enabled the transformation of previously un-transformable elite varieties, such as Ramada, Malisor 84-7, Tegemeo and Macia (Raghuwanshi and Birch, 2010; Liu et al., 2015; Omer et al., 2018; Nelson-Vasilchik et al., 2018; Che et al., 2018; Flinn et al., 2020).
Overcoming genotype recalcitrance and phenolic accumulation is thus central to unlocking the full potential of Sorghum as a next-generation model system and as a resilient cereal crop for climate-adaptive agriculture.
6.2 Sorghum transformation progress
6.2.1 Microprojectile bombardment
The first successful genetic transformation of Sorghum was achieved using microprojectile bombardment using immature embryos as explants (Casas et al., 1993). Although various explants such as immature inflorescences, shoot tips, mature embryos, embryogenic calli, and leaf whorls have been used for Sorghum transformation, immature embryos remain the predominant target (Brettell et al., 1980; Tadesse et al., 2003; Silva et al., 2020).
The first report of sorghum transformation was achieved with microprojectile bombardment reaching 0.3% transformation efficiency (Casas et al., 1993) Optimization of bombardment parameters, including acceleration pressure, distance to the target, aperture of the helium inlet valve, gap width, and microprojectile travel distance, improved efficiency to 1.08% (Able et al., 2001; Tadesse et al., 2003). Further refinements in culture media ingredients, along with optimized DNA delivery conditions increased efficiency to 20.7% (Liu and Godwin, 2012).
6.2.2 Agrobacterium-mediated transformation
Simultaneously, Agrobacterium-mediated Sorghum transformation was optimized through adjustments in infection, co-cultivation, and selection conditions. This included using different selecting agents, applying various treatments like cold or heat pretreatment to promote callus induction, and reduce phenolic compounds (Gao et al., 2005; Nguyen et al., 2007; Gurel et al., 2009). Despite these efforts, Agrobacterium-mediated transformation efficiency remained lower than particle bombardment until Wu et al. (2014) achieved a 33% efficiency using super binary vectors. Super binary vectors, which enhance the virulence of Agrobacterium, were first utilized in plant transformation with Chenopodium quinoa (Komari, 1990) and are binary vectors (small T-DNA carrying plasmid) including additional virulence genes (virB, virC, and virG) (Komari et al., 2006).
Since then, the adoption of Agrobacterium has become the predominant transformation protocol [Anand et al. (2018); Che et al. (2018); Hoerster et al. (2020); Che et al. (2022); Johnson et al. (2023); Wang et al. (2023); Li et al. (2024), and Fontanet-Manzaneque et al. (2024a)].
6.2.3 Implementation of morphogenic regulators
The implementation of morphogenic regulators was a significant breakthrough in monocot transformation substantially increasing the transformation efficiencies of cereals, including Sorghum, maize, rice, and sugarcane (Lowe et al., 2016). This approach leveraged morphogenic transcription factors like maize BABY BOOM (BBM) and WUSCHEL2 (WUS), which promote the transition from vegetative to embryonic growth (Boutilier et al., 2002; Zuo et al., 2002), and increased Sorghum transformation frequencies from 1.9% to 18.3% with morphogenic genes present in the T-DNA of a super binary vector. However, to regenerate plants, it was necessary to remove BBM and WUS2 expression cassettes due to their pleiotropic effects. This was achieved using a drought-inducible maize promoter (pRAB17) to drive CRE recombinase gene expression, which excised the morphogenic regulators, flanked by loxP sites, upon desiccation (Lowe et al., 2016).
6.2.4 Implementation of ternary vectors
The development of a ternary vector system further improved transformation efficiencies. Traditional super binary vectors (e.g., pSB1) have limitations due to their large size and the need for a co-integration step. To address these limitations, new “pVIR” vectors were created, featuring small size, enhanced stability, improved bacterial selectable markers, and an optimal set of amended vir genes (operons virC, virD, and virE) (Anand et al., 2018; Lowe et al., 2018). This introduction of the pVIR plasmid in trans with T-DNA binary vector in the same Agrobacterium strain enabled the development of a rapid maize transformation system (Lowe et al., 2018).
Che et al. (2018) combined the ternary vector system with media optimization techniques described by Wu et al. (2014), achieving high transformation efficiencies in recalcitrant African varieties, including Macia, Malisor 84-7, and Tegemeo. The combination of the ternary vector system, media optimization, and morphogenic regulators (ZmWUS2 and ZmBBM) led to the highest reported Sorghum transformation efficiency of 69.7% (number of regenerated shoots recovered per 100 embryos infected) (Che et al., 2022). Additionally, the use of alternative morphogenic regulators like GRF4-GIF1 achieved transformation efficiencies of 39.68% (number of embryo with regenerated shoots per 100 embryos infected) (Li et al., 2024), demonstrating the versatility and effectiveness of these advanced transformation methodologies. Finally, Fontanet-Manzaneque et al. (2024a) combined the ternary vector pVS1-VIR2 with ZmBBM and ZmWUS2, and optimized the bacterial density, temperature and co-cultivation time leading to a 164.8% transformation efficiency (number of regenerated shoots recovered per 100 embryos infected), a 2.36-fold increase compared to Che et al., 2022.
The advancements in Sorghum genetic transformation have significantly progressed from the initial use of microprojectile bombardment to more sophisticated techniques involving Agrobacterium-mediated methods. The development and utilization of ternary vector systems, alongside media optimization and innovative morphogenic regulators, have culminated in unprecedented transformation efficiencies, even in recalcitrant Sorghum varieties. These methodological advancements not only enhance the feasibility of genetic transformation in Sorghum but also open new avenues for developing drought-resistant and climate-resilient Sorghum cultivars (Figure 3).
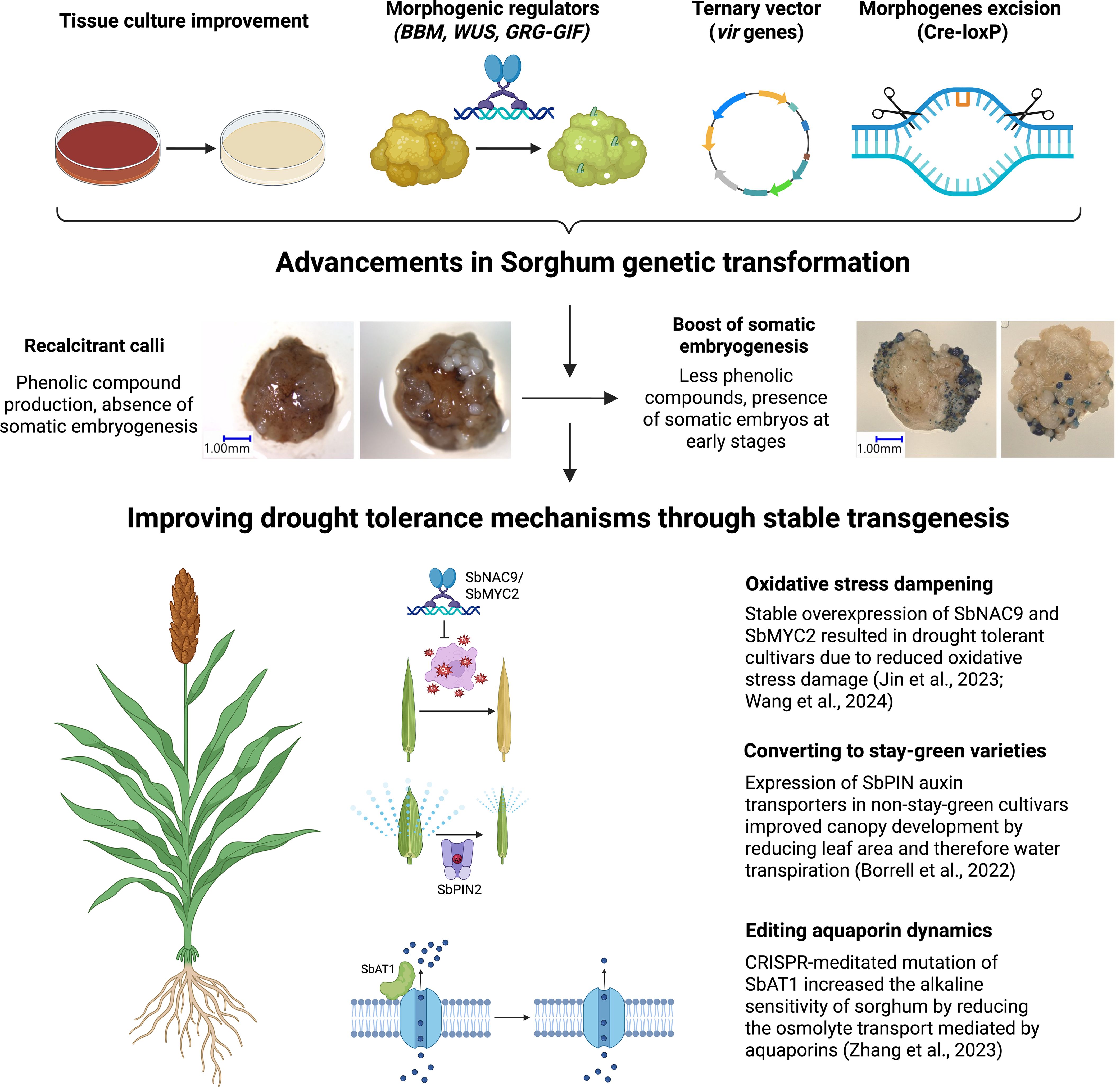
Figure 3. Advancements in Sorghum genetic transformation enable improvement of drought tolerance by stable transgenesis. The refinement of sorghum tissue culture and the development of new technologies including the use of morphogenic regulators, ternary vector system and morphogene excision have allowed the induction of somatic embryogenesis and overcome the sorghum calli recalcitrance. This paves the way to exploit the drought tolerance mechanisms of sorghum by stable transgenesis. Already existing examples reinforce the use of this technology to speed up the generation of crops better adapted to climate change Created in BioRender. Fontanet-Manzaneque, J. B. (2025) https://BioRender.com/xke8k9l.
7 Closing remarks and future prospects
Plant biology has long relied on model organisms such as Arabidopsis to dissect fundamental biological processes. These model species offer distinct experimental advantages, including short life cycles, compact and well-annotated genomes, high transformation and gene editing efficiencies, and simple cultivation requirements. However, their limited agronomic relevance necessitates substantial translational effort to transfer basic findings into field applications. In contrast, crop research targets agriculturally important species (such as wheat, rice, and maize) to generate immediate impact, but progress is often hampered by longer generation times, complex polyploid genomes, and recalcitrance to transformation. Ultimately, both research paradigms face a major bottleneck when translating knowledge from the laboratory to the field.
Sorghum combines several key features of model organisms—such as a small diploid genome, high genetic diversity, and growing omics resources—with the practical strengths of a staple crop: global cultivation, inherent drought tolerance, low input requirements, and multiple end-uses spanning food, feed, and bioenergy. These attributes make sorghum a promising system for both mechanistic studies of abiotic stress in monocots and direct crop improvement (Figure 4). However, important challenges remain before sorghum can be established as a model crop with the robustness and legacy of Arabidopsis.
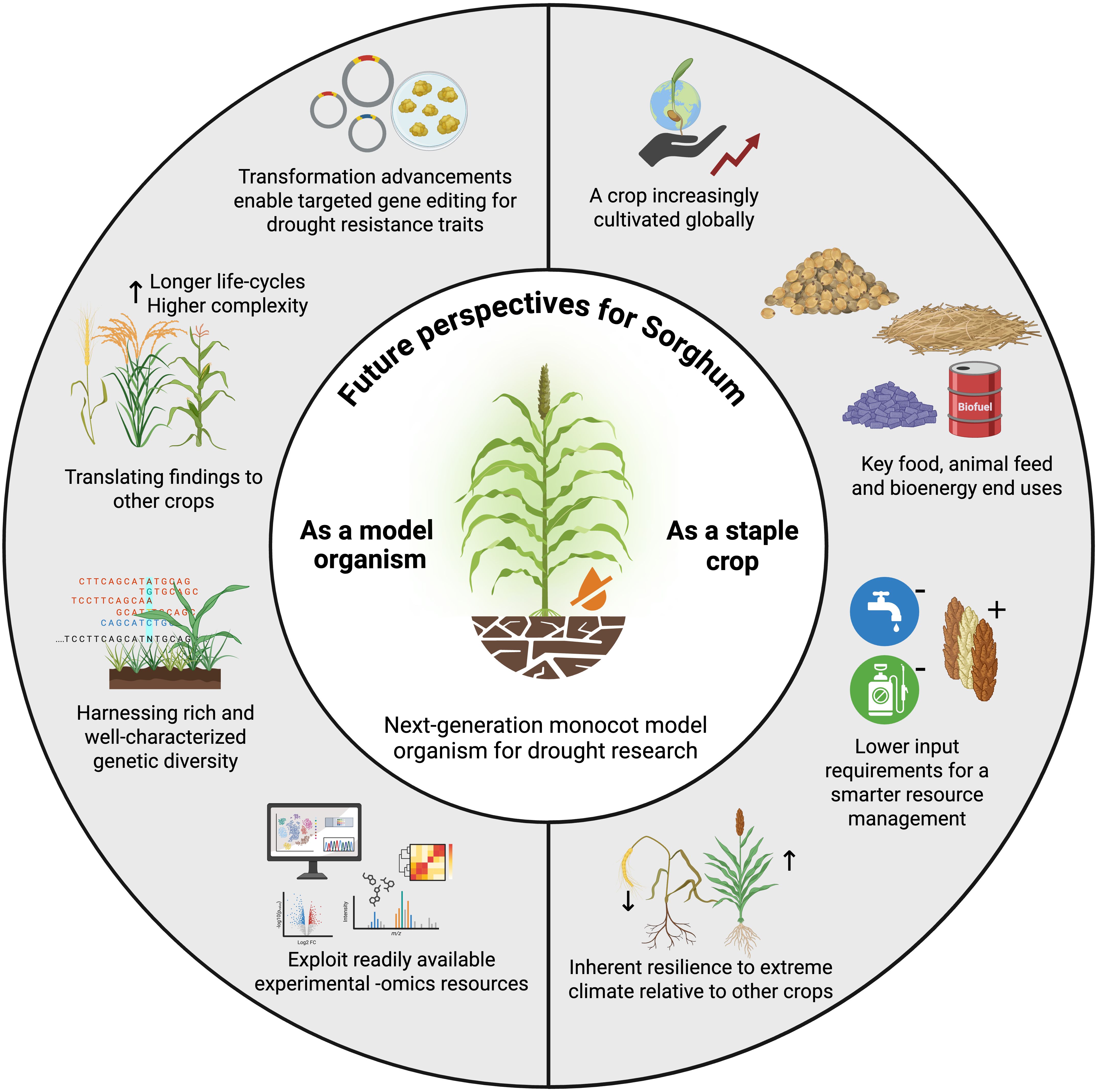
Figure 4. Future perspectives for Sorghum as model organism for the study of drought in monocots. Encompassing characteristics of a model plant and a staple crop, sorghum holds the potential of accelerating the development of climate-smart (particularly drought resistant) crops. Created in BioRender. Caño-Delgado, A. I. (2025) https://BioRender.com/496nmk2.
First, spatial resolution is lacking. While recent transcriptomic studies have attempted to address this by using Laser Capture Microdissection to profile tissue-type-specific expression in sorghum secondary cell wall development (Fu et al., 2024), the field still lacks tissue-specific markers and single-cell resolution tools. Techniques such as single-cell and single-nucleus RNA-seq, already well-established in Arabidopsis and maize (Tenorio-Berrío et al., 2022; Doll et al., 2025), remain largely underdeveloped in Sorghum.
In addition, although many transcriptomic studies have characterized gene expression responses to drought (Johnson et al., 2014; Abdel-Ghany et al., 2020; Zhang et al., 2019; Varoquaux et al., 2019), very few have explored the cistrome or epigenomic landscape, which are crucial to understanding where transcription factors bind across the genome and how chromatin modifications, such as DNA methylation or histone acetylation, regulate gene expression. Recent efforts, such as the study by Zhou et al. (2021), have begun to address this gap using transposase-accessible chromatin sequencing (ATAC-seq) and bisulfite sequencing to detect epigenetic marks. However, more work is needed to map chromatin accessibility and regulatory regions across different tissues and environmental conditions.
Finally, despite Sorghum’s reputation as a drought-tolerant crop, its water footprint remains suboptimal compared to major cereals. For instance, producing one ton of sorghum grain requires 5,695 m³ of water, whereas maize and wheat require only 2,522 m³/ton and 2,474 m³/ton, respectively (Mali et al., 2021). This highlights an urgent need to improve sorghum yield and resource use efficiency through advanced breeding and biotechnological tools, such as CRISPR-Cas9 and the recent breakthroughs in stable genetic transformation, which historically represented one of the biggest bottlenecks in Sorghum research.
In conclusion, while several technical and biological challenges remain, Sorghum stands out not only as a resilient cereal crop but also as a forward-looking model system to bridge basic and translational plant science. Its increasing amenability to genomic tools, transformation protocols, and integrative multi-omics approaches positions it as a strategic platform for accelerating drought tolerance research and driving innovation in cereal crop improvement under climate stress.
Author contributions
JBF-M: Writing – original draft, Writing – review & editing, Visualization, Supervision, Conceptualization. DMH: Writing – review & editing, Visualization. AG: Writing – review & editing, Funding acquisition. AIC-D: Writing – original draft, Writing – review & editing, Funding acquisition, Supervision, Conceptualization.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 Research and Innovation Programme (grant agreement No 683163) and ERC-2022-POC1 (GA#101069461- PLANeT) Juan B. Fontanet-Manzaneque has been funded by European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (grant agreement No 683163) awarded to Ana I. Caño-Delgado and by Grant PID2020-118218RB-I00 funded by MCIU/AEI/10.13039/501100011033. Juan B. Fontanet-Manzaneque was additionally supported by EMBO Scientific Exchange Grant (number 9889). Daniela M. Hernández has been funded by the AGAUR-FI ajuts predoctoral program Joan Oró (2025 FI-1-01284) by the Secretariat of Universities and Research of the Department of Research and Universities of the Generalitat of Catalonia and the European Social Fund Plus and by the ERC-2022-POC1-GA#101069461- PLANeT Funded by the European Union. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency (ERCEA). Neither the European Union nor the granting authority can be held responsible for them. PLANeT Biotech received funding from ERC-2022-POC1-GA#101069461- PLANeT, Startup Capital, ACCIO (grant number ACE117/24/000088). Andrea Giordano was funded by grant TQ2022-012643 funded by MICIU/AEI /10.13039/501100011033 and by European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme (IDRICA grant agreement No 683163).
Conflict of interest
Authors AG and AC-D were employed by the company PLANeT Biotech.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abdel-Ghany, S. E., Ullah, F., Ben-Hur, A., and Reddy, A. S. N. (2020). Transcriptome analysis of drought-resistant and drought-sensitive sorghum (Sorghum bicolor) genotypes in response to PEG-induced drought stress. Int. J. Mol. Sci. 21, 772. doi: 10.3390/ijms21030772
Able, J. A., Rathus, C., and Godwin, I. D. (2001). The investigation of optimal bombardment parameters for transient and stable transgene expression in Sorghum. In Vitro Cell. Dev. Biology-Plant 37, 341–348. doi: 10.1007/s11627-001-0061-7
Abreha, K. B., Enyew, M., Carlsson, A. S., Vetukuri, R. R., Feyissa, T., Motlhaodi, T., et al. (2022). Sorghum in dryland: morphological, physiological, and molecular responses of Sorghum under drought stress. Planta 255, 1–23. doi: 10.1007/s00425-021-03799-7
Ahmed, A., Abdalla, A. A. A., Elsafy, M., Ezzeldin, A., Rahmatov, M., and Abdelhalim, T. (2025). Harnessing genetic diversity in Sudanese sorghum wild relatives for stay-green drought tolerance via microsatellite (SSR) marker assessment. Genetic Resources and Crop Evolution 72, 4699–4711.
Ali, A. E. E., Sharp, R. E., Greeley, L., Peck, S. C., Tabb, D. L., and Ludidi, N. (2025). Proteomic dataset of sorghum leaf and root responses to single and combined drought and heat stress. Sci. Data 12, 97. doi: 10.1038/s41597-025-04369-2
Al-Salman, Y., Ghannoum, O., and Cano, F. J. (2023). Midday water use efficiency in sorghum is linked to faster stomatal closure rate, lower stomatal aperture and higher stomatal density. Plant J. 115, 1661–1676. doi: 10.1111/tpj.16346
Anand, A., Bass, S. H., Wu, E., Wang, N., McBride, K. E., Annaluru, N., et al. (2018). An improved ternary vector system for Agrobacterium-mediated rapid maize transformation. Plant Mol. Biol. 97, 187–200. doi: 10.1007/s11103-018-0732-y
Araya, A., Kisekka, I., Gowda, P. H., and Prasad, P. V. V. (2018). Grain sorghum production functions under different irrigation capacities. Agric. Water Manage. 203, 261–271. doi: 10.1016/j.agwat.2018.03.010
Assefa, Y., Staggenborg, S. A., and Prasad, V. P. (2010). Grain sorghum water requirement and responses to drought stress: A review. Crop Manage. 9, 1–11. doi: 10.1094/CM-2010-1109-01-RV
Awika, H. O., Hays, D. B., Mullet, J. E., Rooney, W. L., and Weers, B. D. (2017). QTL mapping and loci dissection for leaf epicuticular wax load and canopy temperature depression and their association with QTL for staygreen in Sorghum bicolor under stress. Euphytica 213, 207. doi: 10.1007/s10681-017-1990-5
Azhakanandam, K. and Zhang, Z. J. (2015). “Sorghum transformation: achievements, challenges, and perspectives,” in Recent advancements in gene expression and enabling technologies in crop plants, 291–312.
Baker, C. R., Patel-Tupper, D., Cole, B. J., Ching, L. G., Dautermann, O., Kelikian, A. C., et al. (2023). Metabolomic, photoprotective, and photosynthetic acclimatory responses to post-flowering drought in sorghum. Plant Direct 7, e545. doi: 10.1002/pld3.545
Balota, M., Payne, W. A., Rooney, W., and Rosenow, D. (2008). Gas exchange and transpiration ratio in sorghum. Crop Sci. 48, 2361–2371. doi: 10.2135/cropsci2008.01.0051
Bao, S. G., Shi, J. X., Luo, F., Ding, B., Hao, J. Y., Xie, X. D., et al. (2017). Overexpression of Sorghum WINL1 gene confers drought tolerance in Arabidopsis thaliana through the regulation of cuticular biosynthesis. Plant Cell Tissue Organ Culture (PCTOC) 128, 347–356. doi: 10.1007/s11240-016-1114-2
Bao, Y. and Tang, L. (2016). Field-based robotic phenotyping for sorghum biomass yield component traits characterization using stereo vision. IFAC-PapersOnLine 49, 265–270. doi: 10.1016/j.ifacol.2016.10.049
Barnabás, B., Jäger, K., and Fehér, A. (2008). The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 31, 11–38. doi: 10.1111/j.1365-3040.2007.01727.x
Battle, M. W., Vialet-Chabrand, S., Kasznicki, P., Simkin, A. J., and Lawson, T. (2024). Fast stomatal kinetics in sorghum enable tight coordination with photosynthetic responses to dynamic light intensity and safeguard high water use efficiency. J. Exp. Bot. 75, 6796–6809. doi: 10.1093/jxb/erae389
Bayu, W., Rethman, N. F. G., Hammes, P. S., Pieterse, P. A., Grimbeek, J., and Van Der Linde, M. (2005). Water stress affects the germination, emergence, and growth of different Sorghum cultivars. SINET: Ethiopian J. Sci. 28, 119–128. doi: 10.4314/sinet.v28i2.18248
Bihani, P., Char, B., and Bhargava, S. (2011). Transgenic expression of Sorghum DREB2 in rice improves tolerance and yield under water limitation. J. Agric. Sci. 149, 95–101. doi: 10.1017/S0021859610000742
Bing, Y. I., Zhang, Z., Yi, H. A. N., Wenjuan, X. U., and Huang, R. D. (2014). Effect of drought stress during flowering stage on starch accumulation and starch synthesis enzymes in sorghum grains. J. Integr. Agric. 13, 2399–2406. doi: 10.1016/S2095-3119(13)60694-2
Blasco-Escámez, D., Lozano-Elena, F., Fàbregas, N., and Caño-Delgado, A. I. (2017). “The primary root of Sorghum bicolor (L. Moench) as a model system to study brassinosteroid signaling in crop,” in Brassinosteroids: Methods and Protocols (Springer New York, New York, NY), 181–192.
Blomstedt, C. K., Gleadow, R. M., O’Donnell, N., Naur, P., Jensen, K., Laursen, T., et al. (2012). A combined biochemical screen and TILLING approach identifies mutations in Sorghum bicolor L. Moench resulting in acyanogenic forage production. Plant Biotechnol. J. 10, 54–66. doi: 10.1111/j.1467-7652.2011.00646.x
Borrell, A. K., Hammer, G. L., and Henzell, R. G. (2000). Does maintaining green leaf area in Sorghum improve yield under drought? II. Dry matter production and yield. Crop Sci. 40, 1037–1048. doi: 10.2135/cropsci2000.4041037x
Borrell, A. K., van Oosterom, E. J., Mullet, J. E., George-Jaeggli, B., Jordan, D. R., Klein, P. E., et al. (2014). Stay-green alleles individually enhance grain yield in Sorghum under drought by modifying canopy development and water uptake patterns. New Phytol. 203, 817–830. doi: 10.1111/nph.12869
Borrell, A. K., Wong, A. C., George-Jaeggli, B., van Oosterom, E. J., Mace, E. S., Godwin, I. D., et al. (2022). Genetic modification of PIN genes induces causal mechanisms of stay-green drought adaptation phenotype. J. Exp. Bot. 73, 6711–6726. doi: 10.1093/jxb/erac336
Botella, J. R. (2019). Now for the hard ones: is there a limit on CRISPR genome editing in crops? J. Exp. Bot. 70, 734–737. doi: 10.1093/jxb/erz007
Boutilier, K., Offringa, R., Sharma, V. K., Kieft, H., Ouellet, T., Zhang, L., et al. (2002). Ectopic expression of BABY BOOM triggers a conversion from vegetative to embryonic growth. Plant Cell 14, 1737–1749. doi: 10.1105/tpc.001941
Brettell, R. I. S., Wernicke, W., and Thomas, E. (1980). Embryogenesis from cultured immature inflorescences of Sorghum bicolor. Protoplasma 104, 141–148. doi: 10.1007/BF01279376
Burow, G. B., Franks, C. D., Acosta-Martinez, V., and Xin, Z. (2009). Molecular mapping and characterization of BLMC, a locus for profuse wax (bloom) and enhanced cuticular features of Sorghum (Sorghum bicolor (L.) Moench.). Theor. Appl. Genet. 118, 423–431. doi: 10.1007/s00122-008-0908-y
Burow, G. B., Franks, C. D., and Xin, Z. (2008). Genetic and physiological analysis of an irradiated bloomless mutant (epicuticular wax mutant) of sorghum. Crop Sci. 48, 41–48. doi: 10.2135/cropsci2007.02.0119
Cai, T., Daly, B., and Butler, L. (1987). Callus induction and plant regeneration from shoot portions of mature embryos of high tannin Sorghums. Plant cell Tissue Organ culture 9, 245–252. doi: 10.1007/BF00040810
Casas, A. M., Kononowicz, A. K., Zehr, U. B., Tomes, D. T., Axtell, J. D., Butler, L. G., et al. (1993). Transgenic Sorghum plants via microprojectile bombardment. Proc. Natl. Acad. Sci. 90, 11212–11216. doi: 10.1073/pnas.90.23.11212
Cavalet-Giorsa, E., González-Muñoz, A., Athiyannan, N., Holden, S., Salhi, A., Gardener, C., et al. (2024). Origin and evolution of the bread wheat D genome. Nature 633, 848–855. doi: 10.1038/s41586-024-07808-z
Che, P., Anand, A., Wu, E., Sander, J. D., Simon, M. K., Zhu, W., et al. (2018). Developing a flexible, high-efficiency Agrobacterium-mediated Sorghum transformation system with broad application. Plant Biotechnol. J. 16, 1388–1395. doi: 10.1111/pbi.12879
Che, P., Wu, E., Simon, M. K., Anand, A., Lowe, K., Gao, H., et al. (2022). Wuschel2 enables highly efficient CRISPR/Cas-targeted genome editing during rapid de novo shoot regeneration in Sorghum. Commun. Biol. 5, 344. doi: 10.1038/s42003-022-03308-w
Chemelewski, R., McKinley, B. A., Finlayson, S., and Mullet, J. E. (2023). Epicuticular wax accumulation and regulation of wax pathway gene expression during bioenergy Sorghum stem development. Front. Plant Sci. 14, 1227859. doi: 10.3389/fpls.2023.1227859
Chen, X., Wu, Q., Gao, Y., Zhang, J., Wang, Y., Zhang, R., et al. (2020). The role of deep roots in Sorghum yield production under drought conditions. Agronomy 10, 611. doi: 10.3390/agronomy10040611
Cole, B., Zhang, W., Shi, J., Wang, H., Baker, C., Varoquaux, N., et al. (2025). Multi-season analysis reveals hundreds of drought-responsive genes in sorghum. bioRxiv, 2025–2006. doi: 10.1101/2025.06.27.662006
Cooper, E. A., Brenton, Z. W., Flinn, B. S., Jenkins, J., Shu, S., Flowers, D., et al. (2019). A new reference genome for Sorghum bicolor reveals high levels of sequence similarity between sweet and grain genotypes: implications for the genetics of sugar metabolism. BMC Genomics 20, 1–13. doi: 10.1186/s12864-019-5734-x
Cuevas, H. E., Rosa-Valentin, G., Hayes, C. M., Rooney, W. L., and Hoffmann, L. (2017). Genomic characterization of a core set of the USDA-NPGS Ethiopian sorghum germplasm collection: implications for germplasm conservation, evaluation, and utilization in crop improvement. BMC Genomics 18, 1–17. doi: 10.1186/s12864-016-3475-7
Daryanto, S., Wang, L., and Jacinthe, P. A. (2017). Global synthesis of drought effects on cereal, legume, tuber and root crops production: A review. Agric. Water Manage. 179, 18–33. doi: 10.1016/j.agwat.2016.04.022
Davis, K. F., Chiarelli, D. D., Rulli, M. C., Chhatre, A., Richter, B., Singh, D., et al. (2018). Alternative cereals can improve water use and nutrient supply in India. Sci. Adv. 4, eaao1108. doi: 10.1126/sciadv.aao1108
DeFries, R., Liang, S., Chhatre, A., Davis, K. F., Ghosh, S., Rao, N. D., et al. (2023). Climate resilience of dry season cereals in India. Sci. Rep. 13, 9960. doi: 10.1038/s41598-023-37109-w
Deschamps, S., Zhang, Y., Llaca, V., Ye, L., Sanyal, A., King, M., et al. (2018). A chromosome-scale assembly of the sorghum genome using nanopore sequencing and optical mapping. Nat. Commun. 9, 4844. doi: 10.1038/s41467-018-07271-1
Deshpande, S., Rakshit, S., Manasa, K. G., Pandey, S., and Gupta, R. (2017). “Genomic approaches for abiotic stress tolerance in Sorghum,” in The Sorghum Genome (Springer International Publishing, Cham), 169–187.
Dicko, M. H., Gruppen, H., Traoré, A. S., Voragen, A. G., and van Berkel, W. J. (2006). Phenolic compounds and related enzymes as determinants of Sorghum for food use. Biotechnol. Mol. Biol. Rev. 1, 20–37.
Dietz, K. J., Zörb, C., and Geilfus, C. M. (2021). Drought and crop yield. Plant Biol. 23, 881–893. doi: 10.1111/plb.13304
Doll, N. M., Fierlej, Y., Eekhout, T., Elias, L., Bellot, C., Sun, G., et al. (2025). KIL transcription factors facilitate embryo growth in maize by promoting endosperm elimination via lytic cell death. Plant Cell, koaf162. doi: 10.1093/plcell/koaf162
Elango, D., Xue, W., and Chopra, S. (2020). Genome wide association mapping of epi-cuticular wax genes in Sorghum bicolor. Physiol. Mol. Biol. Plants 26, 1727–1737. doi: 10.1007/s12298-020-00848-5
Elkonin, L. A. and Pakhomova, N. V. (2000). Influence of nitrogen and phosphorus on induction embryogenic callus of Sorghum. Plant cell Tissue Organ culture 61, 115–123. doi: 10.1023/A:1006472418218
Enyew, M., Carlsson, A. S., Geleta, M., Tesfaye, K., Hammenhag, C., Seyoum, A., et al. (2022). Novel sources of drought tolerance in sorghum landraces revealed via the analyses of genotype-by-environment interactions. Front. Plant Sci. 13, 1062984. doi: 10.3389/fpls.2022.1062984
Faye, J. M., Maina, F., Hu, Z., Fonceka, D., Cisse, N., and Morris, G. P. (2019). Genomic signatures of adaptation to Sahelian and Soudanian climates in sorghum landraces of Senegal. Ecology and evolution 9, 6038–6051.
Faye, J. M., Akata, E. A., Sine, B., Diatta, C., Cisse, N., Fonceka, D., et al. (2021b). Quantitative and population genomics suggest a broad role of stay-green loci in the drought adaptation of sorghum. Plant Genome 15, e20176. doi: 10.1002/tpg2.20176
Faye, J. M., Maina, F., Akata, E. A., Sine, B., Diatta, C., Mamadou, A., et al. (2021a). A genomics resource for genetics, physiology, and breeding of West African sorghum. Plant Genome 14, e20075. doi: 10.1002/tpg2.20075
Flinn, B., Dale, S., Disharoon, A., and Kresovich, S. (2020). Comparative analysis of in vitro responses and regeneration between diverse bioenergy Sorghum genotypes. Plants 9, 248. doi: 10.3390/plants9020248
Fontanet-Manzaneque, J. B., Haeghebaert, J., Aesaert, S., Coussens, G., Pauwels, L., and Caño-Delgado, A. I. (2024a). Efficient sorghum and maize transformation using a ternary vector system combined with morphogenic regulators. Plant J. 120, 2076–2088. doi: 10.1111/tpj.17101
Fontanet-Manzaneque, J. B., Laibach, N., Herrero-García, I., Coleto-Alcudia, V., Blasco-Escámez, D., Zhang, C., et al. (2024b). Untargeted mutagenesis of brassinosteroid receptor SbBRI1 confers drought tolerance by altering phenylpropanoid metabolism in Sorghum bicolor. Plant Biotechnol. J. 22, 3406–3423. doi: 10.1111/pbi.14461
Fracasso, A., Trindade, L. M., and Amaducci, S. (2016). Drought stress tolerance strategies revealed by RNA-Seq in two Sorghum genotypes with contrasting WUE. BMC Plant Biol. 16, 1–18. doi: 10.1186/s12870-016-0800-x
Fu, J., McKinley, B., James, B., Chrisler, W., Markillie, L. M., Gaffrey, M. J., et al. (2024). Cell-type-specific transcriptomics uncovers spatial regulatory networks in bioenergy sorghum stems. Plant J. 118, 1668–1688. doi: 10.1111/tpj.16690
Gao, Z., Xie, X., Ling, Y., Muthukrishnan, S., and Liang, G. H. (2005). Agrobacterium tumefaciens-mediated Sorghum transformation using a mannose selection system. Plant Biotechnol. J. 3, 591–599. doi: 10.1111/j.1467-7652.2005.00150.x
Garcia, C., Pauli, D., and Arnold, A. E. (2025). Untargeted metabolomics unveils an accumulation of flavonoids associated with the microbiome of low-performing sorghum. Discover Plants 2, 197. doi: 10.1007/s44372-025-00273-6
Girma, G., Nida, H., Tirfessa, A., Lule, D., Bejiga, T., Seyoum, A., et al. (2020). A comprehensive phenotypic and genomic characterization of Ethiopian sorghum germplasm defines core collection and reveals rich genetic potential in adaptive traits. Plant Genome 13, e20055. doi: 10.1002/tpg2.20055
Goche, T., Shargie, N. G., Cummins, I., Brown, A. P., Chivasa, S., and Ngara, R. (2020). Comparative physiological and root proteome analyses of two sorghum varieties responding to water limitation. Sci. Rep. 10, 11835. doi: 10.1038/s41598-020-68735-3
Gorthy, S., Narasu, L., Gaddameedi, A., Sharma, H. C., Kotla, A., Deshpande, S. P., et al. (2017). Introgression of shoot fly (Atherigona soccata L. Moench) resistance QTLs into elite post-rainy season sorghum varieties using marker assisted backcrossing (MABC). Front. Plant Sci. 8, 1494. doi: 10.3389/fpls.2017.01494
Guigou, M., Lareo, C., Pérez, L. V., Lluberas, M. E., Vázquez, D., and Ferrari, M. D. (2011). Bioethanol production from sweet Sorghum: Evaluation of post-harvest treatments on sugar extraction and fermentation. Biomass Bioenergy 35, 3058–3062. doi: 10.1016/j.biombioe.2011.04.028
Guo, D., Li, Y., Lu, H., Zhao, Y., Kurata, N., Wei, X., et al. (2025). A pangenome reference of wild and cultivated rice. Nature, 1–10. doi: 10.1038/s41586-025-08883-6
Gupta, A., Rico-Medina, A., and Caño-Delgado, A. I. (2020). The physiology of plant responses to drought. Science 368, 266–269. doi: 10.1126/science.aaz7614
Gurel, S., Gurel, E., Kaur, R., Wong, J., Meng, L., Tan, H. Q., et al. (2009). Efficient, reproducible Agrobacterium-mediated transformation of sorghum using heat treatment of immature embryos. Plant Cell Rep. 28, 429–444. doi: 10.1007/s00299-008-0655-1
Hagio, T. (2002). Adventitious shoot regeneration from immature embryos of Sorghum. Plant cell Tissue Organ culture 68, 65–72. doi: 10.1023/A:1012918316140
Hao, H., Li, Z., Leng, C., Lu, C., Luo, H., Liu, Y., et al. (2021). Sorghum breeding in the genomic era: opportunities and challenges. Theor. Appl. Genet. 134, 1899–1924. doi: 10.1007/s00122-021-03789-z
Harris, K., Subudhi, P. K., Borrell, A., Jordan, D., Rosenow, D., Nguyen, H., et al. (2007). Sorghum stay-green QTL individually reduce post-flowering drought-induced leaf senescence. J. Exp. Bot. 58, 327–338. doi: 10.1093/jxb/erl225
Harris-Shultz, K. R., Hayes, C. M., and Knoll, J. E. (2019). Mapping QTLs and identification of genes associated with drought resistance in sorghum. Sorghum: Methods Protoc., 11–40. doi: 10.1007/978-1-4939-9039-9_2
Hoerster, G., Wang, N., Ryan, L., Wu, E., Anand, A., McBride, K., et al. (2020). Use of non-integrating Zm-Wus2 vectors to enhance maize transformation. In Vitro Cell. Dev. Biol. Plant 56, 265–279. doi: 10.1007/s11627-019-10042-2
Howe, A., Sato, S., Dweikat, I., Fromm, M., and Clemente, T. (2006). Rapid and reproducible Agrobacterium-mediated transformation of Sorghum. Plant Cell Rep. 25, 784–791. doi: 10.1007/s00299-005-0081-6
Impa, S. M., Perumal, R., Bean, S. R., Sunoj, V. J., and Jagadish, S. K. (2019). Water deficit and heat stress induced alterations in grain physico-chemical characteristics and micronutrient composition in field grown grain Sorghum. J. Cereal Sci. 86, 124–131. doi: 10.1016/j.jcs.2019.01.013
Intergovernmental Panel on Climate Change (IPCC) (2023). Summary for Policymakers. In: Climate Change 2023: Synthesis Report. A Report of the Intergovernmental Panel on Climate Change. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change. II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change.
Izadi-Darbandi, A., Alameldin, H., Namjoo, N., and Ahmad, K. (2023). Introducing Sorghum DREB2 gene in maize (Zea mays L.) to improve drought and salinity tolerance. Biotechnol. Appl. Biochem. 70, 1480–1488. doi: 10.1002/bab.2458
Jadhav, Y., Thakur, N. R., Ingle, K. P., and Ceasar, S. A. (2024). The role of phenomics and genomics in delineating the genetic basis of complex traits in millets. Physiologia Plantarum 176, e14349. doi: 10.1111/ppl.14349
Jenks, M. A., Rich, P. J., Rhodes, D., Ashworth, E. N., Axtell, J. D., and Ding, C. K. (2000). Leaf sheath cuticular waxes on bloomless and sparse-bloom mutants of Sorghum bicolor. Phytochemistry 54, 577–584. doi: 10.1016/S0031-9422(00)00153-9
Jiao, Y., Burke, J., Chopra, R., Burow, G., Chen, J., Wang, B., et al. (2016). A Sorghum Mutant Resource as an Efficient Platform for Gene Discovery in Grasses. Plant Cell 28, 1551–1562. doi: 10.1105/tpc.16.00373
Jiao, C., Xie, X., Hao, C., Chen, L., Xie, Y., Garg, V., et al. (2025). Pan-genome bridges wheat structural variations with habitat and breeding. Nature 637, 384–393. doi: 10.1038/s41586-024-08277-0
Jin, X., Zheng, Y., Wang, J., Chen, W., Yang, Z., Chen, Y., et al. (2023). SbNAC9 improves drought tolerance by enhancing scavenging ability of reactive oxygen species and activating stress-responsive genes of Sorghum. Int. J. Mol. Sci. 24, 2401. doi: 10.3390/ijms24032401
Johnson, K., Cao Chu, U., Anthony, G., Wu, E., Che, P., and Jones, T. J. (2023). Rapid and highly efficient morphogenic gene-mediated hexaploid wheat transformation. Front. Plant Sci. 14, 1151762. doi: 10.3389/fpls.2023.1151762
Johnson, S. M., Cummins, I., Lim, F. L., Slabas, A. R., and Knight, M. R. (2015). Transcriptomic analysis comparing stay-green and senescent Sorghum bicolor lines identifies a role for proline biosynthesis in the stay-green trait. J. Exp. Bot. 66, 7061–7073. doi: 10.1093/jxb/erv405
Johnson, S. M., Lim, F. L., Finkler, A., Fromm, H., Slabas, A. R., and Knight, M. R. (2014). Transcriptomic analysis of Sorghum bicolor responding to combined heat and drought stress. BMC Genomics 15, 1–19. doi: 10.1186/1471-2164-15-456
Jordan, D. R., Hunt, C. H., Cruickshank, A. W., Borrell, A. K., and Henzell, R. G. (2012). The relationship between the stay-green trait and grain yield in elite Sorghum hybrids grown in a range of environments. Crop Sci. 52, 1153–1161. doi: 10.2135/cropsci2011.06.0326
Kadam, S. R. and Fakrudin, B. (2017). Marker assisted pyramiding of root volume QTLs to improve drought tolerance in rabi sorghum. Res. Crops 18, 683–692. doi: 10.5958/2348-7542.2017.00114.0
Kaeppler, H. F. and Pedersen, J. F. (1997). Evaluation of 41 elite and exotic inbred Sorghum genotypes for high quality callus production. Plant cell Tissue Organ culture 48, 71–75. doi: 10.1023/A:1005765009568
Kalpande, H., Surashe, S., Badigannavar, A., More, A., and Ganapathi, T. (2022). Induced variability and assessment of mutagenic effectiveness and efficiency in sorghum genotypes [Sorghum bicolor (L.) Moench. Int. J. Radiat. Biol. 98, 230–243. doi: 10.1080/09553002.2022.2003466
Kamal, N. M., Gorafi, Y. S. A., Tsujimoto, H., and Ghanim, A. M. A. (2018). Stay-Green QTLs Response in Adaptation to Post-Flowering Drought Depends on the Drought Severity. Biomed. Res. Int. 7082095. doi: 10.1155/2018/7082095
Kebede, H., Subudhi, P. K., Rosenow, D. T., and Nguyen, H. T. (2001). Quantitative trait loci influencing drought tolerance in grain sorghum (Sorghum bicolor L. Moench). Theor. Appl. Genet. 103, 266–276. doi: 10.1007/s001220100541
Komari, T. (1990). Transformation of cultured cells of Chenopodium quinoa by binary vectors that carry a fragment of DNA from the virulence region of pTiBo542. Plant Cell Rep. 9, 303–306. doi: 10.1007/BF00232856
Komari, T., Takakura, Y., Ueki, J., Kato, N., Ishida, Y., and Hiei, Y. (2006). Binary vectors and super-binary vectors. Agrobacterium Protoc., 15–42. doi: 10.1385/1-59745-130-4:15
Krupa, K. N., Dalawai, N., Shashidhar, H. E., Harinikumar, K. M., Manojkumar, H. B., Bharani, S., et al. (2017). Mechanisms of drought tolerance in Sorghum: a review. Int. J. Pure Appl. Biosci. 5, 221–237. doi: 10.18782/2320-7051.2845
Lasky, J. R., Upadhyaya, H. D., Ramu, P., Deshpande, S., Hash, C. T., Bonnette, J., et al. (2015). Genome-environment associations in sorghum landraces predict adaptive traits. Sci. Adv. 1, e1400218. doi: 10.1126/sciadv.1400218
Li, H., Han, X., Liu, X., Zhou, M., Ren, W., Zhao, B., et al. (2019). A leucine-rich repeat-receptor-like kinase gene SbER2–1 from Sorghum (Sorghum bicolor L.) confers drought tolerance in maize. BMC Genomics 20, 1–15. doi: 10.1186/s12864-019-6143-x
Li, J., Pan, W., Zhang, S., Ma, G., Li, A., Zhang, H., et al. (2024). A rapid and highly efficient Sorghum transformation strategy using GRF4-GIF1/ternary vector system. Plant J. 117, 1604–1613. doi: 10.1111/tpj.16575
Liaqat, W., Altaf, M. T., Barutçular, C., Mohamed, H. I., Ali, Z., and Khan, M. O. (2024). Drought stress in sorghum: physiological tools, breeding technology, Omics approaches and Genomic-assisted breeding-A review. J. Soil Sci. Plant Nutr. 24, 1665–1691. doi: 10.1007/s42729-024-01702-3
Liu, G., Gilding, E. K., and Godwin, I. D. (2015). A robust tissue culture system for Sorghum [Sorghum bicolor (L.) Moench. South Afr. J. Bot. 98, 157–160. doi: 10.1016/j.sajb.2015.03.179
Liu, G. and Godwin, I. D. (2012). Highly efficient Sorghum transformation. Plant Cell Rep. 31, 999–1007. doi: 10.1007/s00299-011-1218-4
Liu, F., Wodajo, B., Zhao, K., Tang, S., Xie, Q., and Xie, P.. (2025). Unravelling sorghum functional genomics and molecular breeding: past achievements and future prospects. J. Genet. Genom. 52, 719–732. doi: 10.1016/j.jgg.2024.07.016
Lopez, J. R., Erickson, J. E., Munoz, P., Saballos, A., Felderhoff, T. J., and Vermerris, W. (2017). QTLs associated with crown root angle, stomatal conductance, and maturity in sorghum. Plant Genome 10, plantgenome2016–04. doi: 10.3835/plantgenome2016.04.0038
Lowe, K., La Rota, M., Hoerster, G., Hastings, C., Wang, N., Chamberlin, M., et al. (2018). Rapid genotype “independent” Zea mays L.(maize) transformation via direct somatic embryogenesis. In Vitro Cell. Dev. Biology-Plant 54, 240–252. doi: 10.1007/s11627-018-9905-2
Lowe, K., Wu, E., Wang, N., Hoerster, G., Hastings, C., Cho, M. J., et al. (2016). Morphogenic regulators Baby boom and Wuschel improve monocot transformation. Plant Cell 28, 1998–2015. doi: 10.1105/tpc.16.00124
Lu, M., Zhang, D. F., Shi, Y. S., Song, Y. C., Wang, T. Y., and Li, Y. (2013). Expression of SbSNAC1, a NAC transcription factor from Sorghum, confers drought tolerance to transgenic Arabidopsis. Plant Cell Tissue Organ Culture (PCTOC) 115, 443–455. doi: 10.1007/s11240-013-0375-2
Lundgren, M. R., Osborne, C. P., and Christin, P. A. (2014). Deconstructing Kranz anatomy to understand C4 evolution. J. Exp. Bot. 65, 3357–3369. doi: 10.1093/jxb/eru186
Maccaferri, M., Harris, N. S., Twardziok, S. O., Pasam, R. K., Gundlach, H., Spannagl, M., et al. (2019). Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 51, 885–895. doi: 10.1038/s41588-019-0381-3
Mace, E. S., Singh, V., Van Oosterom, E. J., Hammer, G. L., Hunt, C. H., and Jordan, D. R. (2012). QTL for nodal root angle in Sorghum (Sorghum bicolor L. Moench) co-locate with QTL for traits associated with drought adaptation. Theor. Appl. Genet. 124, 97–109. doi: 10.1007/s00122-011-1690-9
Maina, F., Harou, A., Hamidou, F., and Morris, G. P. (2022). Genome-wide association studies identify putative pleiotropic locus mediating drought tolerance in sorghum. Plant direct 6, e413. doi: 10.1002/pld3.413
Mali, S. S., Shirsath, P. B., and Islam, A. (2021). A high-resolution assessment of climate change impact on water footprints of cereal production in India. Sci. Rep. 11, 8715. doi: 10.1038/s41598-021-88223-6
Mason, P. J., Blaakmeer, A., Furtado, A., Stuart, P. N., Nomula, R., Bjarnholt, N., et al. (2024). Harnessing the power of an extensive EMS-induced sorghum population for rapid crop improvement. Physiologia Plantarum 176, e14449. doi: 10.1111/ppl.14449
McCormick, R. F., Truong, S. K., Sreedasyam, A., Jenkins, J., Shu, S., Sims, D., et al. (2018). The Sorghum bicolor reference genome: improved assembly, gene annotations, a transcriptome atlas, and signatures of genome organization. Plant J. 93, 338–354. doi: 10.1111/tpj.13781
Menamo, T., Kassahun, B., Borrell, A. K., Jordan, D. R., Tao, Y., Hunt, C., et al. (2021). Genetic diversity of Ethiopian sorghum reveals signatures of climatic adaptation. Theor. Appl. Genet. 134, 731–742. doi: 10.1007/s00122-020-03727-5
Miller, S., Rønager, A., Holm, R., Fontanet-Manzaneque, J. B., Caño-Delgado, A. I., and Bjarnholt, N. (2023). New methods for Sorghum transformation in temperate climates. AoB Plants 15, plad030. doi: 10.1093/aobpla/plad030
Morris, G. P., Ramu, P., Deshpande, S. P., Hash, C. T., Shah, T., Upadhyaya, H. D., et al. (2013). Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. 110, 453–458. doi: 10.1073/pnas.1215985110
Mukherjee, A., Maheshwari, U., Sharma, V., Sharma, A., and Kumar, S. (2024). Functional insight into multi-omics-based interventions for climatic resilience in sorghum (Sorghum bicolor): a nutritionally rich cereal crop. Planta 259, 91. doi: 10.1007/s00425-024-04365-7
Mullet, J. E., Klein, R. R., and Klein, P. E. (2002). Sorghum bicolor—an important species for comparative grass genomics and a source of beneficial genes for agriculture. Curr. Opin. Plant Biol. 5, 118–121. doi: 10.1016/S1369-5266(02)00232-7
Mwamahonje, A., Eleblu, J. S. Y., Ofori, K., Deshpande, S., Feyissa, T., and Tongoona, P. (2021). Drought tolerance and application of marker-assisted selection in sorghum. Biology 10, 1249. doi: 10.3390/biology10121249
Naaz, N., Choudhary, S., and Hasan, N. (2024). “Proteomics Application in Plant Breeding,” in Plant Molecular Breeding in Genomics Era: Concepts and Tools (Springer Nature Switzerland, Cham), 243–282.
Nagaraja Reddy, R., Madhusudhana, R., Murali Mohan, S., Chakravarthi, D. V. N., Mehtre, S. P., Seetharama, N., et al. (2013). Mapping QTL for grain yield and other agronomic traits in post-rainy sorghum [Sorghum bicolor (L.) Moench. Theor. Appl. Genet. 126, 1921–1939. doi: 10.1007/s00122-013-2107-8
Nagesh Kumar, M. V., Ramya, V., Govindaraj, M., Sameer Kumar, C. V., Maheshwaramma, S., Gokenpally, S., et al. (2021). Harnessing sorghum landraces to breed high-yielding, grain mold-tolerant cultivars with high protein for drought-prone environments. Front. Plant Sci. 12, 659874. doi: 10.3389/fpls.2021.659874
Nelson-Vasilchik, K., Hague, J., Mookkan, M., Zhang, Z. J., and Kausch, A. (2018). Transformation of recalcitrant Sorghum varieties facilitated by baby boom and Wuschel2. Curr. Protoc. Plant Biol. 3, e20076. doi: 10.1002/cppb.20076
Ngara, R., Ramulifho, E., Movahedi, M., Shargie, N. G., Brown, A. P., and Chivasa, S. (2018). Identifying differentially expressed proteins in sorghum cell cultures exposed to osmotic stress. Sci. Rep. 8, 8671. doi: 10.1038/s41598-018-27003-1
Nguyen, T. V., Thanh Thu, T., Claeys, M., and Angenon, G. (2007). Agrobacterium-mediated transformation of Sorghum (Sorghum bicolor (L.) Moench) using an improved in vitro regeneration system. Plant cell Tissue Organ culture 91, 155–164. doi: 10.1007/s11240-007-9228-1
Ochieng, G., Ngugi, K., Wamalwa, L. N., Manyasa, E., Muchira, N., Nyamongo, D., et al. (2021). Novel sources of drought tolerance from landraces and wild sorghum relatives. Crop Sci. 61, 104–118. doi: 10.1002/csc2.20300
Omer, R. A., Asami, P., and Birungi, J.. (2018). Callus induction and plant regeneration from immature embryos of sweet sorghum (Sorghum bicolor Moench). Biotechnology (Faisalabad, Pakistan) 17 (1), 10–3923. doi: 10.3923/biotech.2018.12.18
Paterson, A. H., Bowers, J. E., Bruggmann, R., Dubchak, I., Grimwood, J., Gundlach, H., et al. (2009). The Sorghum bicolor genome and the diversifcation of grasses. Nature 457 (7229), 551–556. doi: 10.1038/nature07723
Pennisi, E. (2009). How Sorghum withstands heat and drought. Science 323, 573–573. doi: 10.1126/science.323.5914.573
Peters, P. J., Jenks, M. A., Rich, P. J., Axtell, J. D., and Ejeta, G. (2009). Mutagenesis, selection, and allelic analysis of epicuticular wax mutants in sorghum. Crop Sci. 49, 1250–1258. doi: 10.2135/cropsci2008.08.0461
Peterson, G. C., Suksayretrup, K., and Weibel, D. E. (1982). Inheritance of some bloomless and sparse-bloom mutants in sorghum 1. Crop Sci. 22, 63–67. doi: 10.2135/cropsci1982.0011183X002200010014x
Punnuri, S., Harris-Shultz, K., Knoll, J., Ni, X., and Wang, H. (2017). The genes bm2 and blmc that affect epicuticular wax deposition in sorghum are allelic. Crop Sci. 57, 1552–1556. doi: 10.2135/cropsci2016.11.0937
Queiroz, M. S., Oliveira, C. E., Steiner, F., Zuffo, A. M., Zoz, T., Vendruscolo, E. P., et al. (2019). Drought stresses on seed germination and early growth of maize and Sorghum. J. Agric. Sci. 11, 310. doi: 10.5539/jas.v11n2p310
Raghuwanshi, A. and Birch, R. G. (2010). Genetic transformation of sweet Sorghum. Plant Cell Rep. 29, 997–1005. doi: 10.1007/s00299-010-0885-x
Ren, G., Yang, P., Cui, J., Gao, Y., Yin, C., Bai, Y., et al. (2022). Multiomics analyses of two sorghum cultivars reveal the molecular mechanism of salt tolerance. Front. Plant Sci. 13, 886805. doi: 10.3389/fpls.2022.886805
Rezaei, E. E., Webber, H., Asseng, S., Boote, K., Durand, J. L., Ewert, F., et al. (2023). Climate change impacts on crop yields. Nat. Rev. Earth Environ. 4, 831–846. doi: 10.1038/s43017-023-00491-0
Rico-Medina, A., Laibach, N., Fontanet-Manzaneque, J. B., Blasco-Escámez, D., Lozano-Elena, F., Martignago, D., et al. (2025). Molecular and physiological characterization of brassinosteroid receptor BRI1 mutants in Sorghum bicolor. New Phytol. 246, 1113–1127. doi: 10.1111/nph.20443
Rizal, G., Karki, S., Alcasid, M., Montecillo, F., Acebron, K., Larazo, N., et al. (2014). Shortening the breeding cycle of sorghum, a model crop for research. Crop Sci. 54, 520–529. doi: 10.2135/cropsci2013.07.0471
Ruperao, P., Thirunavukkarasu, N., Gandham, P., Selvanayagam, S., Govindaraj, M., Nebie, B., et al. (2021). Sorghum pan-genome explores the functional utility for genomic-assisted breeding to accelerate the genetic gain. Front. Plant Sci. 12, 666342. doi: 10.3389/fpls.2021.666342
Sage, R. F., Sage, T. L., and Kocacinar, F. (2012). Photorespiration and the evolution of C4 photosynthesis. Annu. Rev. Plant Biol. 63, 19–47. doi: 10.1146/annurev-arplant-042811-105511
Salas Fernandez, M. G., Bao, Y., Tang, L., and Schnable, P. S. (2017). A high-throughput, field-based phenotyping technology for tall biomass crops. Plant Physiol. 174, 2008–2022. doi: 10.1104/pp.17.00707
Sanchez, A. C., Subudhi, P. K., Rosenow, D. T., and Nguyen, H. T. (2002). Mapping QTLs associated with drought resistance in Sorghum (Sorghum bicolor L. Moench). Plant Mol. Biol. 48, 713–726. doi: 10.1023/A:1014894130270
Sarshad, A., Talei, D., Torabi, M., Rafiei, F., and Nejatkhah, P. (2021). Morphological and biochemical responses of Sorghum bicolor (L.) Moench under drought stress. SN Appl. Sci. 3, 81. doi: 10.1007/s42452-020-03977-4
Satrio, R. D., Fendiyanto, M. H., and Miftahudin, M. (2024). “Tools and techniques used at global scale through genomics, transcriptomics, proteomics, and metabolomics to investigate plant stress responses at the molecular level,” in Molecular dynamics of plant stress and its management (Springer Nature Singapore, Singapore), 555–607.
Sauer, A. M., Loftus, S., Schneider, E. M., Sudhabindu, K., Hajjarpoor, A., Sivasakthi, K., et al. (2024). Sorghum landraces perform better than a commonly used cultivar under terminal drought, especially on sandy soil. Plant Stress 13, 100549. doi: 10.1016/j.stress.2024.100549
Schittenhelm, S. and Schroetter, S. (2014). Comparison of drought tolerance of maize, sweet sorghum and sorghum-Sudangrass hybrids. J. Agron. Crop Sci. 200, 46–53. doi: 10.1111/jac.12039
Sehgal, A., Sita, K., Siddique, K. H., Kumar, R., Bhogireddy, S., Varshney, R. K., et al. (2018). Drought or/and heat-stress effects on seed filling in food crops: impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 9, 1705. doi: 10.3389/fpls.2018.01705
Seth, D., Kumari, P., Pramanik, S., Das, A., and Madhusudhana, R. (2025). Role of omics for enhancing abiotic stress resilience in sorghum (Sorghum bicolor L. Moench). J. Plant Biochem. Biotechnol., 1–18. doi: 10.1007/s13562-025-00994-5
Silva, T. N., Kelly, M. E., and Vermerris, W. (2020). Use of Sorghum bicolor leaf whorl explants to expedite regeneration and increase transformation throughput. Plant Cell Tissue Organ Culture (PCTOC) 141, 243–255. doi: 10.1007/s11240-020-01783-9
Singh, V., van Oosterom, E. J., Jordan, D. R., Hunt, C. H., and Hammer, G. L. (2011). Genetic variability and control of nodal root angle in Sorghum. Crop Sci. 51, 2011–2020. doi: 10.2135/cropsci2011.01.0038
Smith, R. L., Hoveland, C. S., and Hanna, W. W. (1989). Water stress and temperature in relation to seed germination of pearl millet and Sorghum. Agron. J. 81, 303–305. doi: 10.2134/agronj1989.00021962008100020032x
Spindel, J. E., Dahlberg, J., Colgan, M., Hollingsworth, J., Sievert, J., Staggenborg, S. H., et al. (2018). Association mapping by aerial drone reveals 213 genetic associations for Sorghum bicolor biomass traits under drought. BMC Genomics 19, 679. doi: 10.1186/s12864-018-5055-5
Subudhi, P. K., Rosenow, D. T., and Nguyen, H. T. (2000). Quantitative trait loci for the stay green trait in sorghum (Sorghum bicolor L. Moench): consistency across genetic backgrounds and environments. Theor. Appl. Genet. 101, 733–741. doi: 10.1007/s001220051538
Surwenshi, A., Chimmad, V. P., Jalageri, B. R., Kumar, V., Ganapathi, M., and Nakul, H. T. (2010). Characterization of sorghum genotypes for physiological parameters and yield under receding soil moisture conditions. Res. J. Agric. Sci. 1, 242–244.
Swigoňová, Z., Lai, J., Ma, J., Ramakrishna, W., Llaca, V., Bennetzen, J. L., et al. (2004). Close split of sorghum and maize genome progenitors. Genome Res. 14, 1916–1923. doi: 10.1101/gr.2332504
Symanczik, S., Lehmann, M. F., Wiemken, A., Boller, T., and Courty, P. E. (2018). Effects of two contrasted arbuscular mycorrhizal fungal isolates on nutrient uptake by Sorghum bicolor under drought. Mycorrhiza 28, 779–785. doi: 10.1007/s00572-018-0853-9
Tadesse, Y., Sagi, L., Swennen, R., and Jacobs, M. (2003). Optimisation of transformation conditions and production of transgenic Sorghum (Sorghum bicolor) via microparticle bombardment. Plant Cell Tissue Organ Culture 75, 1–18. doi: 10.1023/A:1024664817800
Tao, Y., Luo, H., Xu, J., Cruickshank, A., Zhao, X., Teng, F., et al. (2021). Extensive variation within the pan-genome of cultivated and wild sorghum. Nat. Plants 7, 766–773. doi: 10.1038/s41477-021-00925-x
Tardieu, F., Simonneau, T., and Muller, B. (2018). The physiological basis of drought tolerance in crop plants: a scenario-dependent probabilistic approach. Annu. Rev. Plant Biol. 69, 733–759. doi: 10.1146/annurev-arplant-042817-040218
Tari, I., Laskay, G., Takács, Z., and Poór, P. (2013). Response of sorghum to abiotic stresses: A review. J. Agron. Crop Sci. 199, 264–274. doi: 10.1111/jac.12017
Taylor, J. R., Schober, T. J., and Bean, S. R. (2006). Novel food and non-food uses for Sorghum and millets. J. Cereal Sci. 44, 252–271. doi: 10.1016/j.jcs.2006.06.009
Tenorio Berrío, R., Verstaen, K., Vandamme, N., Pevernagie, J., Achon, I., Van Duyse, J., et al. (2022). Single-cell transcriptomics sheds light on the identity and metabolism of developing leaf cells. Plant Physiol. 188, 898–918. doi: 10.1093/plphys/kiab489
Tsuji, W., Ali, M. E. K., Inanaga, S., and Sugimoto, Y. (2003). Growth and gas exchange of three sorghum cultivars under drought stress. Biol. Plantarum 46, 583–587. doi: 10.1023/A:1024875814296
United Nations (2022). Department of Economic and Social Affairs, Population Division, “World population prospects 2022: Summary of Results. Available online at: https://www.un.org/development/desa/pd/sites/www.un.org.development.desa.pd/files/undesa_pd_2022_wpp_key-messages.pdf.
Uttam, G. A., Praveen, M., Rao, Y. V., Tonapi, V. A., and Madhusudhana, R. (2017). Molecular mapping and candidate gene analysis of a new epicuticular wax locus in sorghum (Sorghum bicolor L. Moench). Theor. Appl. Genet. 130, 2109–2125. doi: 10.1007/s00122-017-2945-x
Van Dijk, M., Morley, T., Rau, M. L., and Saghai, Y. (2021). A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2, 494–501. doi: 10.1038/s43016-021-00322-9
Varoquaux, N., Cole, B., Gao, C., Pierroz, G., Baker, C. R., Patel, D., et al. (2019). Transcriptomic analysis of field-droughted sorghum from seedling to maturity reveals biotic and metabolic responses. Proc. Natl. Acad. Sci. 116, 27124–27132. doi: 10.1073/pnas.1907500116
Venkateswaran, K., Elangovan, M., and Sivaraj, N. (2019). “Origin, domestication and diffusion of Sorghum bicolor,” in Breeding Sorghum for diverse end uses (Woodhead Publishing), 15–31.
Visarada, K. B. R. S. and Sai Kishore, N. (2015). Advances in genetic transformation. Sorghum Mol. Breed., 199–215. doi: 10.1007/978-81-322-2422-8_9
Wang, G., Long, Y., Jin, X., Yang, Z., Dai, L., Yang, Y., et al. (2024). SbMYC2 mediates jasmonic acid signaling to improve drought tolerance via directly activating SbGR1 in Sorghum. Theor. Appl. Genet. 137, 72. doi: 10.1007/s00122-024-04578-0
Wang, N., Ryan, L., Sardesai, N., Wu, E., Lenderts, B., Lowe, K., et al. (2023). Leaf transformation for efficient random integration and targeted genome modification in maize and Sorghum. Nat. Plants 9, 255–270. doi: 10.1038/s41477-022-01338-0
Wang, Y., Zhang, L., Zhang, L., Xing, T., Peng, J., Sun, S., et al. (2013). A novel stress-associated protein SbSAP14 from Sorghum bicolor confers tolerance to salt stress in transgenic rice. Mol. Breed. 32, 437–449. doi: 10.1007/s11032-013-9882-4
Watanabe, K., Guo, W., Arai, K., Takanashi, H., Kajiya-Kanegae, H., Kobayashi, M., et al. (2017). High-throughput phenotyping of sorghum plant height using an unmanned aerial vehicle and its application to genomic prediction modeling. Front. Plant Sci. 8, 421. doi: 10.3389/fpls.2017.00421
Wu, E., Lenderts, B., Glassman, K., Berezowska-Kaniewska, M., Christensen, H., Asmus, T., et al. (2014). Optimized Agrobacterium-mediated Sorghum transformation protocol and molecular data of transgenic Sorghum plants. In Vitro Cell. Dev. Biology-Plant 50, 9–18. doi: 10.1007/s11627-013-9583-z
Xin, Z., Li Wang, M., Barkley, N. A., Burow, G., Franks, C., Pederson, G., et al. (2008). Applying genotyping (TILLING) and phenotyping analyses to elucidate gene function in a chemically induced sorghum mutant population. BMC Plant Biol. 8, 1–14. doi: 10.1186/1471-2229-8-103
Xin, Z., Wang, M. L., Burow, G., and Burke, J. (2009). An induced sorghum mutant population suitable for bioenergy research. Bioenergy Res. 2, 10–16. doi: 10.1007/s12155-008-9029-3
Xu, W., Subudhi, P. K., Crasta, O. R., Rosenow, D. T., Mullet, J. E., and Nguyen, H. T. (2000). Molecular mapping of QTLs conferring stay-green in grain sorghum (Sorghum bicolor L. Moench). Genome 43, 461–469. doi: 10.1139/gen-43-3-461
Yahaya, M. A. and Shimelis, H. (2022). Drought stress in sorghum: Mitigation strategies, breeding methods and technologies—A review. J. Agron. Crop Sci. 208, 127–142. doi: 10.1111/jac.12573
Yang, Z., Chi, X., Guo, F., Jin, X., Luo, H., Hawar, A., et al. (2020). SbWRKY30 enhances the drought tolerance of plants and regulates a drought stress-responsive gene, SbRD19, in Sorghum. J. Plant Physiol. 246, 153142. doi: 10.1016/j.jplph.2020.153142
Yue, L., Wang, H., Shan, Q., Kuerban, Z., Mao, H., and Yu, M. (2025). Metabolomic and transcriptomic analyses of drought resistance mechanisms in sorghum varieties. PeerJ 13, e19596. doi: 10.7717/peerj.19596
Zhang, L. M., Leng, C. Y., Luo, H., Wu, X. Y., Liu, Z. Q., Zhang, Y. M., et al. (2018). Sweet Sorghum originated through selection of Dry, a plant-specific NAC transcription factor gene. Plant Cell 30, 2286–2307. doi: 10.1105/tpc.18.00313
Zhang, H., Yu, F., Xie, P., Sun, S., Qiao, X., Tang, S., et al. (2023). A Gγ protein regulates alkaline sensitivity in crops. Science 379, eade8416. doi: 10.1126/science.ade8416
Zhang, D. F., Zeng, T. R., Liu, X. Y., Gao, C. X., Li, Y. X., Li, C. H., et al. (2019). Transcriptomic profiling of Sorghum leaves and roots responsive to drought stress at the seedling stage. J. Integr. Agric. 18, 1980–1995. doi: 10.1016/S2095-3119(18)62119-7
Zhao, Z. Y., Cai, T., Tagliani, L., Miller, M., Wang, N., Pang, H., et al. (2000). Agrobacterium-mediated sorghum transformation. Plant Mol. Biol. 44, 789–798. doi: 10.1023/A:1026507517182
Zhou, C., Yuan, Z., Ma, X., Yang, H., Wang, P., Zheng, L., et al. (2021). Accessible chromatin regions and their functional interrelations with gene transcription and epigenetic modifications in sorghum genome. Plant Commun. 2 (1). doi: 10.1016/j.xplc.2020.100140
Keywords: Sorghum, monocot model, drought, omics, transformation
Citation: Fontanet-Manzaneque JB, Hernández DM, Giordano A and Caño-Delgado AI (2025) Sorghum as a monocot model for drought research. Front. Plant Sci. 16:1665967. doi: 10.3389/fpls.2025.1665967
Received: 14 July 2025; Accepted: 26 August 2025;
Published: 24 September 2025.
Edited by:
Peng Lv, Hebei Academy of Agricultural and Forestry Sciences, ChinaCopyright © 2025 Fontanet-Manzaneque, Hernández, Giordano and Caño-Delgado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Juan B. Fontanet-Manzaneque, anVhbi5mb250YW5ldEBwc2IudmliLXVnZW50LmJl; Ana I. Caño-Delgado, YW5hLmNhbm9AY3JhZ2Vub21pY2EuZXM=
†Present address: Juan B. Fontanet-Manzaneque, VIB-UGent Center for Plant Systems Biology, Ghent, Belgium